
Submitted:
28 July 2023
Posted:
01 August 2023
You are already at the latest version
Abstract
Biofilm formation and lipopolysaccharide (LPS) are implicated in the pathogenesis of gastrointestinal (GI) diseases caused by Gram-negative such as Escherichia coli and Salmonella enterica serovar Typhimurium. Grape seeds, wine industry by-products, have antioxidant, antimicrobial, and anti-inflammatory activities. In the present study, the protective effect of procyanidin-rich grape seed extract (prGSE), from unfermented pomace of Vitis vinifera cv Bellone, on bacterial LPS-induced oxidative stress and epithelial barrier integrity damages has been studied in a model of Caco-2 cells. The in vitro activity of prGSE against formation of biofilm of S. Typhimurium and E. coli, has been investigated. In vivo, prGSE activity using infected Galleria mellonella larvae has been evaluated. The results showed that the prGSE, if administered with LPS, can significantly reduce the LPS-induced permeability alteration. Moreover, the ability of the extract to prevent Reactive Oxygen Species (ROS) production induced by LPS treatment of Caco-2 cells has been demonstrated. prGSE inhibited the biofilm formation of E. coli and S. Typhimurium. In in vivo activity, increased survival of infected G. mellonella larvae after treatment with prGSE has been demonstrated. In conclusion, grape seed extracts could be used in the reduction of GI damage caused by bacterial endotoxin and biofilm of Gram-negative bacteria.
Keywords:
grape seed extract
; Vitis vinifera L.
; anti-virulence factors
; LPS
; bacterial biofilm
; Salmonella Typhimurium
; Escherichia coli
; Galleria mellonella
; Caco-2.
1. Introduction
In recent years, the association between increased intestinal permeability and oxidative stress production has been demonstrated in several chronic gastrointestinal (GI) disorders such as inflammatory bowel disease (IBD), celiac disease, irritable bowel syndrome (IBS) and bacterial infections [1,2,3].
The mechanisms underlying the pathophysiology of these disorders are still unclear, but the integrity of the intestinal barrier could be modulated by several factors - such as mucosal inflammatory events, different dietary patterns and intestinal microbial composition - and it seems to be involved in these processes [4]. Bacterial biofilms, such as those of Escherichia coli and Salmonella, have been implicated in the pathogenesis of many GI diseases like inflammatory bowel disease and irritable bowel syndrome [5]. Moreover, substantial evidence indicates that E. coli is involved in Crohn’s disease and is also a contributing factor in ulcerative colitis pathogenesis [6].
Even if the connection between impaired intestinal permeability and the altered gut microbiota is still not clear, increased intestinal permeability could lead to a translocation of the luminal content (such as bacteria and pathogenic molecules) to the bloodstream, as demonstrated in GI chronic diseases, acute intestinal failure, and Gram-negative sepsis [7,8]. The pathological passage of several substances can enhance the mucosal inflammatory processes on the basis of chronic GI disorders and, through the mesenteric lymph nodes these pathogenic molecules can reach the systemic circulation contaminating sterile organs, including liver, lungs and brain [9].
One of the most known mechanisms of pathological translocation involves lipopolysaccharide (LPS) which is an endotoxin present in the Gram-negative bacteria that is able to induce an immune/inflammatory host response not only locally but also systemic such as during sepsis [10,11,12,13].
In previous studies, it was observed, using an ex vivo experimental model, that acute exposure of human colonic mucosa to pathogenic LPS alters the contractility of human colonic smooth muscle cells, through both LPS mucosal translocation and production of free radicals, suggesting that increased permeability can play a significant role in the onset of GI disease [11,12]. Moreover, numerous studies showed that E. coli infection reduced the mucus thickness, decreased the intestinal barrier integrity and increased the concentrations of LPS [14].
In the last years, in vitro and in vivo studies have demonstrated that several natural products, such as probiotics, prebiotics, and plant-derived extracts, could restore the altered intestinal permeability in conditions of stress, suggesting the possibility of new therapeutic strategies [15,16,17,18]. Moreover, some of them could also modulate the oxidative stress processes and reduce pro-inflammatory cytokines secretion [19]. It has been previously demonstrated that exposure of human colonic mucosa to inulin, a dietary fiber with prebiotic activity, seems to prevent LPS-induced alteration in expression of some key proteins, which promote intestinal motility and inflammation, reducing the radical-mediated oxidative stress [13].
Phenolic compounds from plant matrices and phytochemicals showed different radical scavenging capacity [20]. Seed extracts of Vitis vinifera L., rich in procyanidins (PCs), showed higher antioxidant activity than other plant extracts [21]. Moreover, it has been reported that grape seed extracts (GSE) obtained from different table and wine cultivars have a significant activity against fungi such as Candida species, dermatophytes and Malassezia [22,23]. Kitsiou and colleagues demonstrated a significant inhibitory growth effect of 4% GSE against Salmonella enterica serovar Typhimurium and E. coli [24]. Specifically, grape seeds contain many bioactive molecules, lipids, proteins, carbohydrates and 5-8% polyphenols, depending on the cultivar. Most of the total polyphenols are PCs, which are flavan-3-ols present in monomeric, dimeric, oligomeric, and polymeric forms. The PCs are considered to be primarily responsible for the biological effects of grape seed extract [22,23].
GSE can be obtained from agro-waste of the vine-wine industry, which produces them in thousands of tons per year. Wine by-products are of poor value and their disposal constitutes an ecological and economical management issue for wineries industries. Since it has been recognized as an important natural source of compounds with promising health properties, its use may be part of a circular economy perspective [23].
GSE, containing high levels of PCs rich in phenolic hydrogens scavengers of hydrogen radical donors, could reduce oxidative stress by acting as a regulator of the inflammatory reaction restoring the integrity of the intestinal barrier after LPS exposition [25]. Grape polyphenols have previously been shown to improve gut health and are oxidative damage; however, the mechanism of these beneficial effects is still debated.
In the present study, the activity of procyanidin-rich GSE (prGSE) on bacterial LPS-induced oxidative stress and epithelial barrier integrity damages has been studied in a model of Caco-2 cells. Among two of the major foodborne Gram-negative pathogens, S. Typhimurium and E. coli, the efficacy of prGSE inhibiting the formation of biofilms was evaluated. Moreover, for the first time, the protective effect of prGSE in Galleria mellonella larvae infected, a consolidated in vivo model, has been investigated.
2. Results
2.1. Phytochemical analysis of prGSE
2.1.1. Total Phenolic Content
The results of chemical analysis of V. vinifera (cv Bellone) seed extract from unfermented pomace revealed that it is a very phenol-rich matrix. The analysis was carried out on filtered and unfiltered samples, as described in the methods and materials section. The spectrophotometric analysis and subsequent quantification based on the calibration curve of gallic acid allowed the determination of total phenolic compounds. A difference in phenol content was found between filtered and unfiltered samples before being subjected to the Folin-Ciocalteu assay. The unfiltered samples had a higher concentration of phenols (645 mg/g) than the filtered samples (530 mg/g).
2.1.2. High Performance Liquid Chromatography (HPLC) Analysis
Separation of phenolic compounds in prGSE was achieved within 90 min. The HPLC chromatogram of prGSE was recorded at 278 nm. The components were eluted in the following order: catechin (1), procyanidin B2 (2), epicatechin (3), epicatechin gallate (4) and the polymeric procyanidins (5 and 6) and their concentrations are shown in Table 1. Moreover, two groups of polymeric procyanidins, named Pol 1 (5) and Pol 2 (6), have been separated and quantified. As shown in Table 1, the main components of prGSE are 5 and 6, with a concentration of 365.48 and 78.36 mg/g, respectively. Within the monomeric procyanidins, catechin (1) represents the main one with a concentration of 10.92 mg/g, followed by epicatechin (3) (7.66 mg/g) and epicatechin gallate (4) (2.80 mg/g). Finally, procyanidin B2 (2) shows a concentration of 4.05 mg/g. The total procyanidins content determined by HPLC-DAD is 469.27 mg/g.
2.1.3. Nuclear Magnetic Resonance (NMR) Analysis
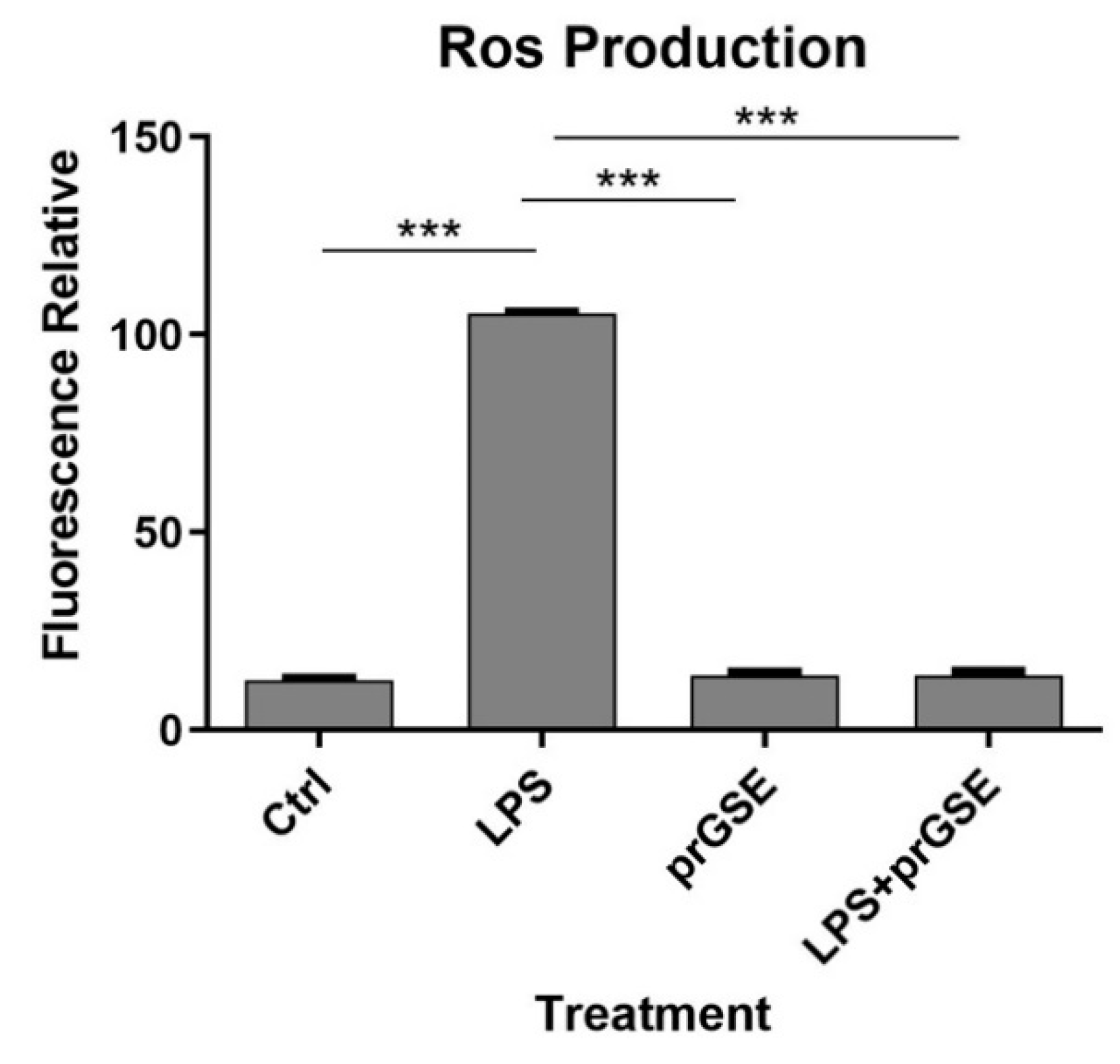
From the 1H NMR spectrum (Figure 1), it was possible to identify and quantify 24 metabolites classified as amino acids, organic acids, carbohydrates, and miscellaneous molecules. Among them, it is interesting the presence of ascorbate, procyanidin B1 and polymeric procyanidins. In particular, the diagnostic resonances of procyanidin B1 are the doublets at 6.05 ppm, 6.13 ppm, 6.15 ppm and two ABX systems at 6.88 ppm (two doublet of doublets), 6.95 ppm (triplet), 6.96 (triplet), 6.98 ppm (doublet) and 7.06 ppm (doublet). For what regards the aromatic moieties of the polymeric procyanidins, they can be assessed by the broad resonances at 6.15 ppm and 6.82 ppm. The carbohydrates, whose resonances are the most abundant in the spectrum, are identified on the basis of their anomeric protons at 4.65 ppm and 5.22 ppm for glucose and at 5.44 ppm for sucrose.
The identified molecules were integrated and normalized for the number of protons originating that resonance, then by the internal standard and then for the extract dry weight. The measured amounts are reported in Table 2.
2.2. Caco-2 experiments
2.2.1. Effect of the Different Treatments with LPS, prGSE, and LPS-prGSE on the Caco-2 cell Monolayer Viability/Cytotoxicity
The effects of prGSE on cell viability using the MTT assay has been preliminarily assessed. To this scope, Caco-2 cells were exposed to increasing concentrations (3.125 – 6.25 – 12.5 – 25 and 50 µg/mL) of prGSE for 24, 48 and 72 hours. The MTT assay results suggest that prGSE applied at 3.125 – 6.25 – 12.5 – 25 and 50 µg/mL had no significant cytotoxic effects (p>0.05) on the Caco-2 cells after treatment. According to these results, in the following experiments, a prGSE concentration of 6.25 µg/ml was used.
2.2.2. FD-4 Permeability Analysis after LPS, prGSE, and LPS-prGSE on the Caco-2 cells
To evaluate the alteration of the Caco-2 cell monolayer integrity, the paracellular penetration amount of FD-4 across Caco-2 monolayers was investigated. FD-4 is a large molecule with a molecular weight of 4 kDa. The cumulative increase of FD-4 fluorescence in the receiver compartment was measured using a multiplate reader (Tecan), calculated and plotted as a function of time, every 20 min over 6 h (from 20 min up to 360 min in Figure 2). FD-4 permeability across the epithelial barrier is significantly increased following LPS treatment (10 μg/mL for 24 h) compared to the control (4.647±0.306 vs 0.479±0.178 pmols; p<0.0313), which is in agreement with the previous literature [26,27,28,29]. Co-treatment with prGSE was able to prevent the LPS induced alteration (4.647±0.306 vs 0.457±0.147 pmols; p<0.0313.) showing a value similar to the control. Interestingly, prGSE alone did not alter the permeability values (0.397±0.103 vs 0.479±0.178 pmols, p=ns) excluding a possible harmful effect of the extract.
2.2.3. Analysis of Reactive Oxygen Species (ROS) production after LPS, prGSE, and LPS-prGSE treatment on the Caco-2 cells
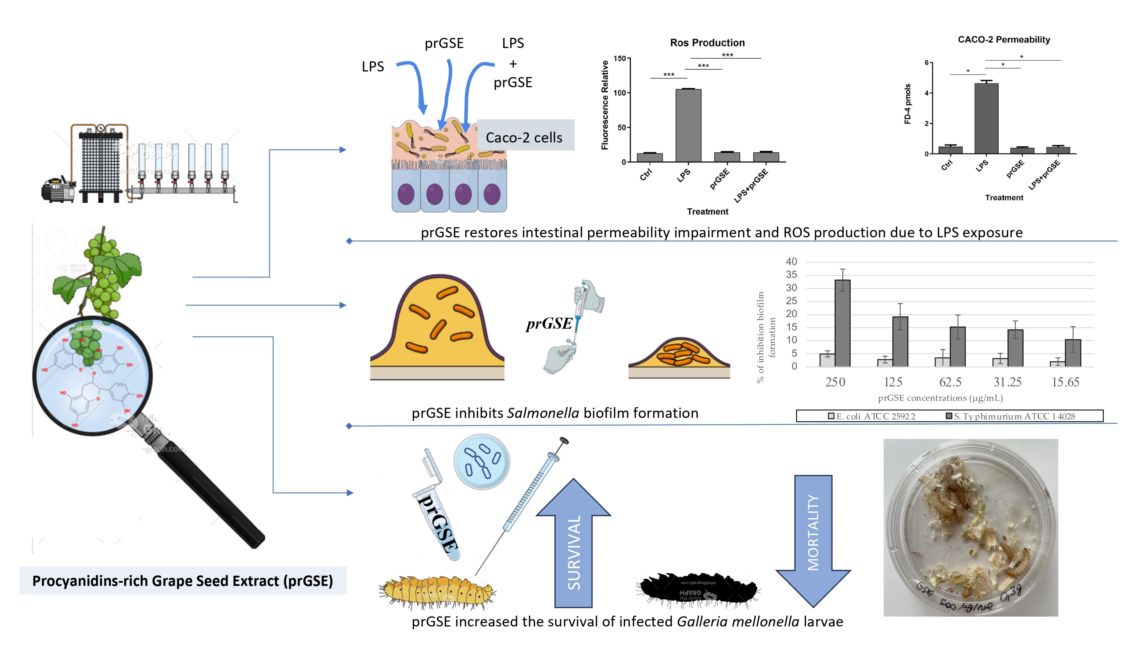
In order to explore the presence of oxidative stress following exposure to the different treatment analyzed, intracellular ROS levels were measured in LPS-, prGSE-, LPS+prGSE-treated Caco2 monolayers via the carboxy-H2 DCFDA fluorescent probe, as described previously [30,31]. LPS treatment significantly increased the ROS production compared to the control (104±1.90 vs 12.9±1.08; p < 0.0001) while the co-treatment prGSE-LPS, with the prGSE concentration of 6.25 µg/mL, is able to prevent this production (19.0±2.00 vs 104±1.90; p < 0.0001). prGSE alone didn't determine a significant production of ROS compared to the control (11.3±1.30 vs 19.2±2.0; p > 0.0001).
Figure 3.
Reactive Oxygen Species (ROS) levels were analyzed by using carboxy-H2DCFDA staining by fluorescence plate reader. The experiments were executed in triplicate. Data are reported as fluorescence relatives. The experiments were executed in triplicate. Data are reported as mean ± SD. Statistical analysis was performed using a two-way ANOVA, followed by Bonferroni’s post-hoc correction test ***p<0.0001. Ctrl (Control); LPS (Lipopolysaccharide); prGSE (procyanidins-rich Grape Seed Extract); prGSE+LPS (Mix).
Figure 3.
Reactive Oxygen Species (ROS) levels were analyzed by using carboxy-H2DCFDA staining by fluorescence plate reader. The experiments were executed in triplicate. Data are reported as fluorescence relatives. The experiments were executed in triplicate. Data are reported as mean ± SD. Statistical analysis was performed using a two-way ANOVA, followed by Bonferroni’s post-hoc correction test ***p<0.0001. Ctrl (Control); LPS (Lipopolysaccharide); prGSE (procyanidins-rich Grape Seed Extract); prGSE+LPS (Mix).
2.3. In Vitro and In Vivo Activity of prGSE Against Salmonella Typhimurium and Escherichia coli Cells and Virulence Factors
2.3.1. In Vitro Antibacterial Activity Evaluation
PrGSE antibacterial activity was investigated against S. Typhimurium ATCC 14028 and E. coli ATCC 25922, using the standard microdilution method [32]. PrGSE showed the capacity to inhibit Gram-negative bacteria growth in a dose-dependent manner, being more active against S. Typhimurium. MIC values for S. Typhimurium are lower than those obtained with E. coli. PrGSE displayed a Geometric Mean (GM) MIC50 value of 44.17 µg/mL against S. Typhimurium, while for E. coli GM MIC50 value is 55.58 µg/mL.

Table 3.
Antibacterial activity of procyanidins-rich grape seed extract against Salmonella Typhimurium ATCC 14028 and Escherichia coli ATCC 25922 planktonic cells.
Table 3.
Antibacterial activity of procyanidins-rich grape seed extract against Salmonella Typhimurium ATCC 14028 and Escherichia coli ATCC 25922 planktonic cells.
| GM MIC50 (µg/mL) | GM MIC90 (µg/mL) | GM MIC100 (µg/mL) | |
|---|---|---|---|
| Salmonella Typhimurium ATCC 14028 | 44.17 | 88.39 | 222.73 |
| Escherichia coli ATCC 25922 | 55.68 | 99.21 | 396.85 |
3 MIC50=Minimum Inhibitory Concentration of 50% bacterial growth; MIC90=Minimum Inhibitory Concentration of 90% bacterial growth; MIC100=Minimum Inhibitory Concentration of 100% bacterial growth; GM=Geometric mean.
2.3.2. In Vitro Antibiofilm Activity
The anti-biofilm activity was evaluated after 24 h of incubation, using CV assay. prGSE displayed its major anti-biofilm activity against S. Typhimurium, with a percentage of inhibition of 33% at 250 µg/mL. At the same concentration, prGSE inhibits E. coli biofilm formation of nearly 5%. Thus, it can be stated that prGSE has no activity against E. coli biofilm. On the other hand, prGSE showed a dose-dependent inhibition of S. Typhimurium biofilm formation. As shown in Figure 4, the percent rate of inhibition decreases with the concentration of prGSE used.
2.3.3. In Vivo Antibacterial Activity
In the past decades, G. mellonella larvae have been largely used to test the antimicrobial activity of both chemical and natural compounds because of the immune system remarkably similar with mammals. This animal model meets the bioethics principle of the 3Rs (Replacement, Reduction, and Refinement) in animal experimentation and is characterized by simple handling and experimental procedures [33]. G. mellonella larvae were infected with S. Typhimurium ATCC 14028 and E. coli ATCC 25922. After they were infected, they were treated with 500 µg/mL, 250 µg/mL, or 125 µg/mL of prGSE. The survival of the larvae was reported daily for 5 days. Survival rate was 100% for both untreated larvae and larvae injected with PBS. Conversely, the mortality rate in the group infected with S. Typhimurium and E. coli was 60% and 40%, respectively. The in vivo antibacterial assay showed significant results at the concentration of 500 µg/mL and 250 µg/mL (P value <0.01 and 0.05, respectively) against S. Typhimurium (Figure 5). When treated with 5 µg of prGSE, G. mellonella larvae infected with S. Typhimurium showed a survival percent rate of 100%. After 5 days, only one larva treated with 0.5 µg of prGSE died. The survival rate decreased to 50% after 5 days for the group of larvae treated with 0.05 µg of prGSE. E. coli infected larvae, treated with 5 µg of prGSE, survived with a percent rate of 100%. 40% of larvae died for both the group treated with 0.5 µg and 0.05 µg of prGSE, after 5 days (Figure 6).
3. Discussion
The use of plants as a source of medical remedies has a long history and, despite the intensive production of synthetic drugs by pharmaceutical companies, natural products still attract the attention of the scientific community. Recently, there has been an increased demand for natural treatments that can limit the damaging effects given by common antifungal, antibiotic and anti-inflammatory drugs. In addition to whole plants or their organs as sources of bioactive natural products, agro-industrial by-products and waste represent value-added matrices.
The recovery of V. vinifera seeds, a by-product of the winemaking process, comes within the concept of the circular economy; this model of production and consumption can exploit existing products and natural resources with the aim of extending their life cycle and reducing by-products and waste derived from them. This study shows how V. vinifera seeds from unfermented pomace, can be reintroduced into production cycles due to their content of several bioactive compounds. Phytochemical characterization of prGSE obtained from the cultivar Bellone showed the presence of several phenolic compounds. From a qualitative point of view, the prGSE of the present investigation shows the same phytochemical profile of other cultivars, having as main components monomeric, dimeric, and polymeric procyanidins [22]. However, the content of each compound can vary depending on the matrix. In fact, the concentrations of the metabolites in GSE are dependent on several factors, both endogenous and exogenous, such as cultivar, climatic conditions, soil nutrients, altitude, and soil type, consequently it is of crucial importance to use a titrated extract [34].
PrGSE has a high concentration of polymeric procyanidins (443.84 mg/g over 469.27 mg/g of total procyanidins) and these results could explain the high activity of this extract. In fact, in a previous study a significant correlation between the content of polymeric flavan-3-ols with a polymerization degree ≥4 in GSEs and antimicrobial activity was demonstrated [22].
Enterobacteriaceae is a large family of Gram-negative bacteria, which comprises many symbiont and pathogenic bacteria, such as Salmonella and Escherichia coli. Pathogenesis in Salmonella and E. coli infections is an outcome of multiple virulence factors, including LPS (bacterial endotoxin) and biofilm production. LPS is an outer component of the cell wall and plays a role in the endotoxic shock mechanism and in the pathophysiology of the infections [35]. Biofilm is a community of bacteria enclosed in an extracellular substance that renders bacteria resistant to stresses, immune system clearance, and antibiotics. Biofilm additionally mediates pathogen-host interactions and is an essential factor in chronic infections [36]. It is known that Gram-negative bacteria are significantly represented in the intestinal microbiota of patients affected by Chronic Intestinal Disorders such as IBD or IBS [37], and endotoxemia has been detected in 48% of patients with Crohn’s disease and 28% of patients with ulcerative colitis [38].
The effect of LPS on intestinal mucosa is not completely understood; it’s already known that it is able to alter the intestinal permeability [39], possibly facilitating a pathological translocation of several substances. Moreover, it has been demonstrated that it binds the Toll Like Receptor 4 on the surface of the intestinal epithelial cells, which is able to activate several local inflammatory processes, such as ROS production and synthesis of inflammatory cytokines [40,41].
In the present study, the effect of prGSE against LPS-induced damage on Caco-2 cells was investigated. Caco-2 was chosen for this test since it is a colonic adenocarcinoma cell line which could considered as intestinal epithelium model because it forms monolayers with characteristics of intestinal epithelial cells such as the formation of microvillus and expresses brush-border proteins [10,42]. Previous studies on the Caco-2 cell monolayer model have shown an inverse relationship between intestinal epithelial resistance and paracellular permeability, following exposure to different harmful stimuli [10,29,43,44,45].
The results of the present investigation showed that the prGSE if administered with LPS was able to significantly reduce the permeability alteration due to this endotoxin. It has been previously demonstrated, in an animal model of intestinal inflammation and in cell culture, that PCs are able to prevent the impaired intestinal permeability even if it seems that this protective effect is mediated by the polymeric PCs but not by oligomers [46,47]. In accordance with this evidence, the prGSE was rich in polymeric procyanidins. The positive effect of PCs on the altered permeability induced by LPS seems to be induced by the overexpression of the tight junction protein such as occludin and zona occludens (ZO)-1 [48,49]. Claudin-2 is a pore-forming claudin that forms high conductance, paracellular cation-selective pores [50] and it has been demonstrated that in IBD patients, its expression is altered determining a changing in tight-junction structure [51,52].
Another important result of the present investigation was the ability of the extract to prevent ROS production induced by LPS treatment of Caco-2 cells. It is well known that oxidative stress is one of the most important mechanisms on the basis of the inflammatory processes due to Gram-negative infection on the intestinal mucosa [53,54,55].
For several years the antioxidant and anti-inflammatory capacities of various natural extracts have been studied with a view to being able to identify new potential therapeutic approaches for acute and chronic inflammatory states involving the intestinal mucosa [56]. For this reason, in this study the protective role of prGSE was explored in order to identify new natural substances deriving from waste products capable of being reused for preventive and curative purposes. It was also interesting to observe that a treatment of Caco-2 with the extract alone didn’t alter the mucosal permeability nor did it determine the production of ROS, confirming that this extract does not cause damage to the intestinal epithelial cells.
Among Enterobacteriaceae, E. coli and S. Typhimurium, the main zoonotic bacteria that can be transmitted from animals to humans via the food chain, affect millions of people every year [57]. Salmonella can cause persistent intestinal infection, gut microbiota imbalance and chronic inflammation [58]. Most bacteria live not planktonically, but as residents of sessile biofilm communities. Biofilm formation in the GI tract has an adverse effect on the immune response of the host. The formation of biofilm allows bacteria to persist in the inflammation environment, accelerate mucosa damage, and create an immune cell-related chronic infection reservoir [59]. Protection against intestinal infections, including those of bacterial origin, and enhancement of the epithelial gut barrier function represent an objective to improve human health. In the present study it has been demonstrated that prGSE inhibits S. Typhimurium biofilm, which is responsible for drug resistance and intestinal damage.
Moreover, the activity of prGSE in G. mellonella larvae infected with E. coli and S. Typhimurium has been investigated. In vitro tests provide partial information on the antagonistic effects against bacterial pathogens, they essentially determine the inhibition of growth, bactericidal activity, and inhibition of biofilm formation. The results from the in vivo test can efficiently complement in vitro results. G. mellonella larvae represent a cost-effective and simple in vivo model.
G. mellonella in vivo model can be considered as an alternative to vertebrates to investigate enteric bacteria pathogens [60], since similarities have been described between mammalian digestive apparatus and intestinal epithelial cells from larvae [61]. The basic tissue architecture of the midgut is similar to those found in the human intestine, such as epithelial arrangements of columnar cells and smooth septate junctions that control permeability—analogous to tight junctions [62]. The insect peritrophic matrix is the functional equivalent to the mammalian mucus layer, which acts as a barrier for the epithelial cells and impedes pathogen movement into the body cavity (i.e., the haemocoel) [63,64]. Moreover, some microbial communities characterized in the midgut of G. mellonella are similar to those found in crypts of the human intestine [65,66].
Lately, reduction of Listeria monocytogenes virulence and Staphylococcus aureus infections were successfully demonstrated using G. mellonella larvae [67,68,69]. Additionally, G. mellonella has been used to evaluate the infectivity of gut pathogens such as L. monocytogenes [70], Campylobacter jejunis [71], Vibrio spp. [72], Shigella spp. [73], and S. enterica[74]. For the first time in this study the activity of prGSE has been demonstrated in an in vivo model of G. mellonella. The results obtained by the G. mellonella protection assay showed activity of prGSE by reducing the mortality of both the larvae infected with S. Typhimurium and the larvae infected with E. coli. The protective effects conferred in the G. mellonella larvae could have been caused by different mechanisms, including enhancement of the larval immune system.
In the present study, the results obtained demonstrated that prGSE is able to modulate some important virulence factors of Gram-negative bacteria, suggesting a possible use of this extract as an
alternative treatment in maintaining gastrointestinal health. Further studies are needed to confirm its protective and anti-inflammatory properties even if these preliminary results are in line with the current Literature and are promising towards possible uses in clinical practice.
4. Materials and Methods
4.1. Extraction and phytochemical analysis of prGSE
4.1.1. Plant Material
PrGSE were obtained from unfermented pomace of Vitis vinifera L. cv. Bellone. Plant material was harvested in September 2021 (Pietra Pinta winery, Latium region). All samples were stored at -20°C until use.
4.1.2. Sample Preparation
The seeds were separated from the unfermented pomace by sieving, washed and frozen to facilitate the freeze-drying process. The freeze-dried seeds were weighed and ground into a fine powder. They were extracted with an EtOH/H2O mixture (7:3 v/v) acidified with formic acid pH 3, using the matrix/solvent ratio of 1g/10 mL. Extraction was conducted at 40°C for 3h. Removal of the solid residue was achieved by vacuum filtration; the filtrate was then concentrated using the rotary evaporator (BUCHI R Pro 220). Finally, the extract was frozen and lyophilized.
4.1.3. Total Phenolic Content
For the total phenolic compounds’ quantification, the Folin–Ciocalteu method was performed. A stock solution of gallic acid with a concentration of 2.4 mg/mL and different dilutions were prepared for the calibration curve. A 50% EtOH/H2O solution of the lyophilized extract with a concentration of 1 mg/mL and a sodium carbonate solution at 10.75% were prepared. To test whether filtration affected the content of total polyphenols, the samples were subjected to filtration with 45 µm PTFE syringe filters or not. At room temperature, 1 mL of sample solution was placed in a 20 mL volumetric flask, and 2 mL of distilled water and 1 mL of Folin-Ciocalteu reagent were then added to the flask. Following the addition of Folin-Ciocalteu reagent, a waiting time of 3 min was observed. Afterward, 4 mL of sodium carbonate solution and 12 mL of distilled water were added to the flask. A blank with all the components except the extract was also prepared. The samples were then shaken and incubated at room temperature for 90 min. Subsequently, the samples were placed in a quartz cuvette cell and then read on a spectrophotometer (Shimadzu UV-1280) at an absorbance of 750 nm [75]. All total phenolic compound concentrations have been expressed in gallic acid equivalents.
4.1.4. High Performance Liquid Chromatography (HPLC) Analysis
HPLC analyses were performed with an HPLC system consisting of a 1260 Infinity II flexible pump, a 1260 Infinity II autosampler, and an HS 1260 Infinity II diode array detector set at lambda of 278 nm. An InfinityLab Poroshell 120 EC-C18 column (3.0 mm x 150 mm, 2.7 µm) was used as a stationary phase at a temperature of 27°C. The gradient elution method was performed starting with 95% H2O for 5 min, then with 84% H2O for 2 min, 80% H2O for 4 min, 70% H2O for 3 min, 20% H2O for 4 min, until reaching 100 % ACN for 5 min, followed by an equilibration time of 10 min with a flow rate of 1 mL/min. A stock solution of the standards (catechin, procyanidin B2, epicatechin and epicatechin gallate) was prepared each at the concentration of 1 mg/mL except for procyanidin B2 the concentration of which was 0.5 mg/mL, and this was solubilized in a 70% (v/v) hydroalcoholic mixture. Five dilutions were performed from the stock solution and a volume of 2 µL was injected to construct calibration curves for each standard, and elution times and peak subtended areas were identified. The seed extract at the concentration of 10 mg/mL was solubilized in the same 70% hydroalcoholic mixture and 5 µL was injected for a total of three replicates. Polymeric procyanidins with a degree of polymerization ≥4 were quantified at 278 nm according to the method described by Simonetti and colleagues [22], using procyanidin B2 as an external standard in a concentration range of 0.001-0.2 μg and a five-point calibration curve with R2 = 0.9998.
4.1.5. Nuclear Magnetic Resonance (NMR) Analysis
A known amount of dried extract of V. vinifera seeds was resuspended in 700 μL of D2O containing 3-(trimethylsilyl)-propionic-2,2,3,3-d4 acid sodium salt (TSP) as an internal chemical shift and concentration internal standard at a final concentration of 2mM. The NMR experiments were carried out on at 298 K on a JNM-ECZ 600R spectrometer operating at the proton frequency of 600 MHz and equipped with a multinuclear z-gradient inverse probe head, and the monodimensional 1H spectra were acquired employing a presat pulse sequence, 64k data points, a spectral width of 15 ppm (9.03 kHz), a recycle delay of 5.72s, a presat time of 2s and an acquisition time of 5.81s. The spectra were processed with an exponential window function of 0.1 Hz, phase corrected and then the baseline was optimized with Akima algorithm. Resonance assignment was carried out on the basis of resonance chemical shift, multiplicity, software database (Chenomix) and literature data [76,77].
4.2. Caco-2 Culture experiments
4.2.1. Cell culture
Human epithelial colorectal adenocarcinoma (Caco-2) cells purchased from the American Type Culture Collection (ATCC, VA, USA), were maintained under standard cell culture conditions, in a humidified 37°C incubator, with 5% CO2, in Dulbecco’s modified of Eagle’s Medium with 4,5 g/L glucose & L-glutamine without sodium pyruvate (DMEM high glucose, Corning, Sigma-Aldrich, Milan, Italy) containing 10% (vol/vol) fetal bovine serum (FBS, Euroclone, South-America), 5% (vol/vol) L-glutamine (Aurogene, Rome, Italy), 5% (vol/vol) Penicillin/Streptomycin (Merck, Sigma-Aldrich, Milan, Italy) and 0,5% (vol/vol) HEPES (Dominique Dutscher, France). L-glutamine has been added after 21 days. Caco-2 were periodically screened for contamination. Cells seeded at a density of 5.0 x 104 per insert were grown on transwell chambers (12 mm with 0.4 μm pore polyester membrane inserts; Corning, Sigma-Aldrich, Milan, Italy) placed in a 12-well plate, 500 μL of media were placed in the apical compartments and 1600 μL of media were placed in the basolateral compartments. Experiments were performed 21 days after seeding when the cells reached confluence and differentiation. Fresh media was changed every other day in the apical and basolateral compartments of the well until the day of experimentation [10].
4.2.2. Caco-2 cell Monolayer Viability/Cytotoxicity
To determine the effect of extracts on Caco-2 cells and to choose the correct concentration to use for experiments, viability was assessed using an MTT assay after 24, 48 and 72 h, to exposure of prGSE extract at different concentrations (from 50 to 3.125 mg/mL, w/v). Then, the medium was removed, and each well was washed with 200 ml of PBS. The metabolic activity of the cells was evaluated via their capacity to reduce the yellow MTT (0.5 mg/mL in serum-free DMEM) to a blue formazan product using 3 h of incubation at 37°C. Then, the medium containing MTT was removed from each well of the 96-multiwell, washed in PBS and the formazan crystals were dissolved in DMSO. The absorbance was read at 570 nm using a microplate reader Tecan Infinite M200-Pro. Cell proliferation values were expressed as percentages from the relative absorbance measured in the treated wells versus control wells (untreated cells).
4.2.3. Treatment of Caco-2 cells with LPS and prGSE
Cells were challenged apically with LPS, prGSE and LPS + prGSE for 24 h at 37°C. LPS (bacterial lipopolysaccharide from a pathogenic strain of E. coli 0111:B4 w/v, Sigma-Aldrich, Milan, Italy) at the concentration of 10 mg/ml and prGSE (6.25 mg/mL, w/v) were dissolved in plain DMEM.
4.2.4. Cell permeability assay
Epithelial barrier function was assessed by measuring the unidirectional paracellular flux of fluorescein isothiocyanate-dextran (FD-4, Sigma-Aldrich, Milan, Italy) from the apical to basolateral compartments. In brief, prior to transport studies, the culture medium on both sides of the Caco-2 cell monolayer was removed by aspiration and replaced with prewarmed Krebs-Hensleit Buffer (KHB, pH 7.4) for 1 h at 37°C. The cells were then washed in PBS, and FD-4 was applied to the apical side of the Caco-2 monolayer, at 1 mg/mL final concentration in prewarmed KHB [10], and its paracellular permeability was calculated from the apical to basolateral direction by collecting undernatants. The concentration of FD-4 in the solution was measured every 20 min over 6 h, by removing an aliquot from the receiver compartment (undernatants) and replacing it with an equal volume of fresh KHB. Fluorescence readings were carried out at ex/em 490/520 nm by using a multiplate reader (Tecan). Fluorescence values were converted into a concentration of fluorescein (pmol), using a standard curve. Each experiment was performed in triplicates with independent controls among the three conditions.
4.2.5. Analysis of Oxidative Stress
Intracellular ROS production was assessed by adding 2′,7′-dichlorodihydrofluorescein diacetate (H2-DCF-DA, Sigma Aldrich), according to Dinicola and colleagues [30,31,78], with slight modifications in Caco-2 cells exposed to LPS, prGSE and LPS + prGSE for 24 h in complete medium. The day of the experimentation, cells were incubated for 3 h. Incubation with 400 μM H2O2 (Sigma-Aldrich, Milan, Italy) was used as positive control for ROS. After incubation the culture medium was replaced with PBS and cells were loaded with 10 µM H2-DCF-DA for 30 min. After incubation, PBS and H2-DCF-DA were removed and the cells were gently washed twice in PBS. Then, red-free medium was added. The increase in cell fluorescence was measured at excitation and emission wavelengths of 485 and 530 nm, respectively, using an Infinite F200 auto microplate reader (Tecan), at 25 °C. Each experiment was performed in triplicates with independent controls among the three conditions.
4.3. In Vitro and In Vivo Activity of prGSE Against Salmonella enterica serovar Typhimurium and Escherichia coli Cells and Virulence Factors
4.3.1. Bacterial strains and growth conditions
Bacterial strains (Salmonella enterica serovar Typhimurium ATCC 14028 and Escherichia coli ATCC 25922) were obtained from the American Culture Type Collection (ATCC, Rockville, MD, USA). Bacteria were grown on Mueller-Hinton agar (Sigma Aldrich, St. Louis, MO, USA) at 37°C for 24 h.
4.3.2. Antimicrobial susceptibility tests
The antifungal activity of prGSE against two Gram-negative bacteria, S. Typhimurium and E. coli, was determined using the standard microdilution method [32]. Bacterial suspensions were obtained adjusting bacterial concentration to obtain a turbidity visually comparable to that of the 0.5 McFarland turbidity standard (≈108 CFU/mL). Suspensions were 100-fold diluted with Mueller-Hinton broth (Sigma Aldrich, St. Louis, MO, USA) to obtain a final concentration of 106 CFU/mL and 100 μl of these suspensions added to the wells. Serial two-fold dilutions of prGSE in concentrations ranging from 500 μg/ml to 0.97 μg/ml were used to determine Minimum Inhibitory Concentration (MIC). Plates were incubated for 24 h at 37° C. The MIC endpoint is the lowest concentration of prGSE where no visible growth is seen in the wells, compared with the growth-control wells. The tests were conducted in double, in three different replicates.
4.3.3. Anti-biofilm Activity
Anti-biofilm activity of prGSE was investigated following the method previously described with some modification [79]. 107 CFU/mL bacterial suspensions were added to flat-bottomed, 48-wells microtiter plates, together with prGSE in concentrations ranging from 250 μg/ml to 15.65 μg/ml and incubated for 24h at 37°C. After that time, supernatant was removed, and bacterial cells and extracellular biofilm matrix were fixed with 200 μL of methanol and incubated at room temperature for 15 minutes. After methanol was removed, the biofilm total biomass was quantified with Crystal Violet (CV; Sigma-Aldrich) solution. After the incubation, the biofilm was washed with phosphate buffered saline (PBS) four times and left to dry. The bound dye to biofilms was solubilized in ethanol 96% for 30 minutes. The absorbance measurement was recorded at 595 nm using a microplate reader and the total biomass calculated. The experiments were conducted in triple, in three different replicates.
4.3.4. In vivo G. mellonella survival assay
In vivo studies were conducted using G. mellonella larvae, obtained from Bigserpens (Frosinone, Italy), as previously described [80,81]. Larvae of G. mellonella of 0.3 ± 0.03 g were selected. Thirteen groups of larvae (10 larvae for each group, 1 group for each treatment) were inoculated in the last left proleg with 105 cells of E. coli or 105 cells of S. Typhimurium together with 10 μL of prGSE at the concentrations of 500 µg/mL, 250 µg/mL,125 µg/mL. Three groups with respectively 500 µg/mL, 250 µg/mL,125 µg/mL of prGSE were defined. Three control groups were used: one group of larvae with no treatment applied, one group were treated with sterile phosphate buffered saline (PBS) and two groups were treated with E. coli and S. Typhimurium, respectively. The larvae were then incubated at 37°C and monitored for 120 h. They were considered dead when they did not respond to physical stimulation (a slight pressure with forceps). Each experiment was repeated at least three times and reported as a percent survival rate.
4.4. Statistical Analysis
Data are presented as mean ± standard deviation of at least two independent experiments. Data were analyzed using GraphPad Prism v.9, (La Jolla, San Diego, CA, USA) software tool. Normally distributed data were analyzed for significance by unpaired t-test and two-way ANOVA, followed by post-hoc test (Bonferroni’s). The Mantel-Cox log-rank test was used to judge the statistical significance relative to the control in in vivo tests. Significance was at the 0.05 level.
Author Contributions
Conceptualization: G.P., G.S., D.DEV., F.S., A.A.; methodology: R.M.N., G.B., E.I., A.A., L.D.; C.B.; validation R.M.N., G.B., E.I., L.D., C.B., F.S.; formal analysis: R.M.N., G.B., E.I., A.A., C.B., F.S.; writing—original draft preparation: R.M.N., G.B., E.I., A.A., G.S., F.S.; writing—review and editing: G.S., A.A., G.B., M.P.L.G.; supervision: G.P., G.S., A.A., M.P.L.G., G.B., L.D. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the POR FESR Lazio 2014-2020 - Azione 1.2.1 PROT. A0375-2020-36590- “Gruppi di ricerca 2020”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- C, P.; F, P.; D, C.; J, T.; Sh, I. Inflammatory Bowel Disease and Primary Sclerosing Cholangitis: A Review of the Phenotype and Associated Specific Features. Gut Liver 2018, 12. [Google Scholar] [CrossRef]
- Schultz, B.M.; Paduro, C.A.; Salazar, G.A.; Salazar-Echegarai, F.J.; Sebastián, V.P.; Riedel, C.A.; Kalergis, A.M.; Alvarez-Lobos, M.; Bueno, S.M. A Potential Role of Salmonella Infection in the Onset of Inflammatory Bowel Diseases. Front. Immunol. 2017, 8, 236225. [Google Scholar] [CrossRef]
- T, V.; J, T.; R, F. The Role of Intestinal Permeability in Gastrointestinal Disorders and Current Methods of Evaluation. Front. Nutr. 2021, 8. [Google Scholar] [CrossRef]
- Ij, M.; M, M.; J, W.; N, M.; D, W.; R, A.; J, B.-W.; E, M. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10. [Google Scholar] [CrossRef]
- Nayak, S.K. Biofilm-Mediated Gastrointestinal Diseases. In Biofilms in Human Diseases: Treatment and Control; Springer International Publishing AG, 2019; pp. 167–176. [Google Scholar]
- Martinez-Medina, M.; Garcia-Gil, L.J. Escherichia Coli in Chronic Inflammatory Bowel Diseases: An Update on Adherent Invasive Escherichia Coli Pathogenicity. World J. Gastrointest. Pathophysiol. 2014, 5, 213. [Google Scholar] [CrossRef] [PubMed]
- A, T.; A, M.; A, B.; J, F.; M, S. Preventing Bacterial Translocation in Patients with Leaky Gut Syndrome: Nutrition and Pharmacological Treatment Options. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- A, A.; A, de G; M, R. The Gut-Liver Axis in Liver Disease: Pathophysiological Basis for Therapy. J. Hepatol. 2020, 72. [Google Scholar] [CrossRef]
- R, N.; H, Y. Bacterial Translocation from the Gut to the Distant Organs: An Overview. Ann. Nutr. Metab 2017, 71 Suppl 1. [Google Scholar] [CrossRef]
- Gori, M.; Altomare, A.; Cocca, S.; Solida, E.; Ribolsi, M.; Carotti, S.; Rainer, A.; Francesconi, M.; Morini, S.; Cicala, M.; et al. Palmitic Acid Affects Intestinal Epithelial Barrier Integrity and Permeability In Vitro. Antioxidants 2020, 9. [Google Scholar] [CrossRef]
- Mp, G.; R, S.; A, A.; S, C.; M, D.P.; S, C.; G, S.; R, A.; S, E.; S, M.; et al. Human Colonic Myogenic Dysfunction Induced by Mucosal Lipopolysaccharide Translocation and Oxidative Stress. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2013, 45. [Google Scholar] [CrossRef]
- V, P.; A, A.; Mp, G.; V, L.; S, C.; S, C.; R, P.; R, A.; L, D.G.; M, C. Antioxidant Activity of Inulin and Its Role in the Prevention of Human Colonic Muscle Cell Impairment Induced by Lipopolysaccharide Mucosal Exposure. PloS One 2014, 9. [Google Scholar] [CrossRef]
- Guarino, M.P.L.; Altomare, A.; Barera, S.; Locato, V.; Cocca, S.; Franchin, C.; Arrigoni, G.; Vannini, C.; Grossi, S.; Campomenosi, P.; et al. Effect of Inulin on Proteome Changes Induced by Pathogenic Lipopolysaccharide in Human Colon. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Wang, D.; He, Y.; Liu, K.; Deng, S.; Fan, Y.; Liu, Y. Sodium Humate Alleviates Enterotoxigenic Escherichia Coli-Induced Intestinal Dysfunction via Alteration of Intestinal Microbiota and Metabolites in Mice. Front. Microbiol. 2022, 13, 809086. [Google Scholar] [CrossRef]
- M, F.; M, A.-B.; M, A.-G.; Jp, G.M.; X, S.-R.; A, H.-P.; E, E.; Am, G.-C.; D, G.; B, L.; et al. Present and Future Therapeutic Approaches to Barrier Dysfunction. Front. Nutr. 2021, 8. [Google Scholar] [CrossRef]
- Mp, G.; A, A.; E, S.; M, M.; C, S.; R, A.; G, D.; L, M.; R, C.; M, C. Effect of Acute Mucosal Exposure to Lactobacillus Rhamnosus GG on Human Colonic Smooth Muscle Cells. J. Clin. Gastroenterol 2008, 42 Pt 2. [Google Scholar] [CrossRef]
- F, A.; A, S.; A, A.; P, M.; C, P.; B, A.; R, C.; M, G.; M, M.; M, C.; et al. Lactobacillus Rhamnosus Protects Human Colonic Muscle from Pathogen Lipopolysaccharide-Induced Damage. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2013, 25. [Google Scholar] [CrossRef]
- P, L.; S, B.; J, G.; Hj, W.; G, F.; J, F.-G.; S, V. Beyond Heat Stress: Intestinal Integrity Disruption and Mechanism-Based Intervention Strategies. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- I, H.; W, H.; L, T.; A, M.; S, J.; X, D.; C, Y.; W, H.; G, Z.; Z, K.; et al. Phytochemicals and Inflammatory Bowel Disease: A Review. Crit. Rev. Food Sci. Nutr. 2020, 60. [Google Scholar] [CrossRef]
- Pasqualetti, V.; Locato, V.; Fanali, C.; Mulinacci, N.; Cimini, S.; Morgia, A.M.; Pasqua, G.; De Gara, L. Comparison between In Vitro Chemical and Ex Vivo Biological Assays to Evaluate Antioxidant Capacity of Botanical Extracts. Antioxidants 2021, 10, 1136. [Google Scholar] [CrossRef]
- N, M.; A, V.; V, P.; M, I.; C, G.; M, B.; G, D.A.; A, C.; V, L.; C, D.V.; et al. Effects of Ionizing Radiation on Bio-Active Plant Extracts Useful for Preventing Oxidative Damages. Nat. Prod. Res. 2019, 33. [Google Scholar] [CrossRef]
- G, S.; Ar, S.; Fd, D.; N, M.; M, I.; F, C.; E, P.; E, G.; S, P.; D, A.; et al. Evaluation of Anti-Candida Activity of Vitis Vinifera L. Seed Extracts Obtained from Wine and Table Cultivars. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Simonetti, G.; Brasili, E.; Pasqua, G. Antifungal Activity of Phenolic and Polyphenolic Compounds from Different Matrices of Vitis Vinifera L. against Human Pathogens. Molecules 2020, 25, 3748. [Google Scholar] [CrossRef]
- Kitsiou, M.; Purk, L.; Gutierrez-Merino, J.; Karatzas, K.A.; Klymenko, O.V.; Velliou, E. A Systematic Quantitative Determination of the Antimicrobial Efficacy of Grape Seed Extract against Foodborne Bacterial Pathogens. Foods 2023, 12, 929. [Google Scholar] [CrossRef]
- Gupta, M.; Dey, S.; Marbaniang, D.; Pal, P.; Ray, S.; Mazumder, B. Grape Seed Extract: Having a Potential Health Benefits. J. Food Sci. Technol. 2020, 57, 1205. [Google Scholar] [CrossRef]
- Guo, H.; Xu, Y.; Huang, W.; Zhou, H.; Zheng, Z.; Zhao, Y.; He, B.; Zhu, T.; Tang, S.; Zhu, Q. Kuwanon G Preserves LPS-Induced Disruption of Gut Epithelial Barrier In Vitro. Molecules 2016, 21. [Google Scholar] [CrossRef]
- Y, H.; K, I.; R, K.; M, M.; T, U.; Y, I.; K, T. Protective Effects of Lactoferrin against Intestinal Mucosal Damage Induced by Lipopolysaccharide in Human Intestinal Caco-2 Cells. Yakugaku Zasshi 2008, 128. [Google Scholar] [CrossRef]
- A, B.; A, Z.; M, G.; D, D.; B, S. LPS Induces Hyper-Permeability of Intestinal Epithelial Cells. J. Cell. Physiol. 2017, 232. [Google Scholar] [CrossRef]
- Y, W.; J, T.; B, C.; B, W.; D, Z.; B, W. Effects of Alcohol on Intestinal Epithelial Barrier Permeability and Expression of Tight Junction-Associated Proteins. Mol. Med. Rep. 2014, 9. [Google Scholar] [CrossRef]
- Calabriso, N.; Massaro, M.; Scoditti, E.; Verri, T.; Barca, A.; Gerardi, C.; Giovinazzo, G.; Carluccio, M.A. Grape Pomace Extract Attenuates Inflammatory Response in Intestinal Epithelial and Endothelial Cells: Potential Health-Promoting Properties in Bowel Inflammation. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Atzeri, A.; Lucas, R.; Incani, A.; Peñalver, P.; Zafra-Gómez, A.; Melis, M.P.; Pizzala, R.; Morales, J.C.; Deiana, M. Hydroxytyrosol and Tyrosol Sulfate Metabolites Protect against the Oxidized Cholesterol Pro-Oxidant Effect in Caco-2 Human Enterocyte-like Cells. Food Funct. 2016, 7, 337–346. [Google Scholar] [CrossRef]
- M07: Dilution AST for Aerobically Grown Bacteria - CLSI . Available online: https://clsi.org/standards/products/microbiology/documents/m07/ (accessed on 17 July 2023).
- Serrano, I.; Verdial, C.; Tavares, L.; Oliveira, M. The Virtuous Galleria Mellonella Model for Scientific Experimentation. Antibiotics 2023, 12, 505. [Google Scholar] [CrossRef]
- Giannini, B.; Mulinacci, N.; Pasqua, G.; Innocenti, M.; Valletta, A.; Cecchini, F. Phenolics and Antioxidant Activity in Different Cultivars/Clones of Vitis Vinifera L. Seeds over Two Years. Plant Biosyst. - Int. J. Deal. Asp. Plant Biol. 2016. [Google Scholar]
- Abdel-Rhman, S.H. Role of Pseudomonas Aeruginosa Lipopolysaccharides in Modulation of Biofilm and Virulence Factors of Enterobacteriaceae. Ann. Microbiol. 2019, 69, 299–305. [Google Scholar] [CrossRef]
- Harrell, J.E.; Hahn, M.M.; D’Souza, S.J.; Vasicek, E.M.; Sandala, J.L.; Gunn, J.S.; McLachlan, J.B. Salmonella Biofilm Formation, Chronic Infection, and Immunity Within the Intestine and Hepatobiliary Tract. Front. Cell. Infect. Microbiol. 2021, 10, 624622. [Google Scholar] [CrossRef]
- A, D.; P, N.; M, D.; G, M.K.; J, N.; S, A.; G, L. Increased Serum Levels of Lipopolysaccharide and Antiflagellin Antibodies in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2015, 27. [Google Scholar] [CrossRef]
- O, P.R.; A, L.S.R.; E, A.A.; A, de la H, M.; E, R.S.; A, A.M. Serum Lipopolysaccharide-Binding Protein in Endotoxemic Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2007, 13. [Google Scholar] [CrossRef]
- S, G.; R, A.-S.; Hm, S.; Ty, M. Lipopolysaccharide Causes an Increase in Intestinal Tight Junction Permeability in Vitro and in Vivo by Inducing Enterocyte Membrane Expression and Localization of TLR-4 and CD14. Am. J. Pathol. 2013, 182. [Google Scholar] [CrossRef]
- S, B.; Pk, D.; Jk, T. Lipopolysaccharide Activates NF-KappaB by TLR4-Bcl10-Dependent and Independent Pathways in Colonic Epithelial Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295. [Google Scholar] [CrossRef]
- Mt, A. Toll-like Receptor Signalling in the Intestinal Epithelium: How Bacterial Recognition Shapes Intestinal Function. Nat. Rev. Immunol. 2010, 10. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Rodrigues, M.; Santos, A.O.; Alves, G.; Silva, L.R. Antioxidant Status, Antidiabetic Properties and Effects on Caco-2 Cells of Colored and Non-Colored Enriched Extracts of Sweet Cherry Fruits. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Ty, M.; D, N.; V, B.; H, N.; N, H. Ethanol Modulation of Intestinal Epithelial Tight Junction Barrier. Am. J. Physiol. 1999, 276. [Google Scholar] [CrossRef]
- Rm, C.; C, F.; A, T.; M, A.; Ca, M.; S, G. Differential Effect of Ethanol and Hydrogen Peroxide on Barrier Function and Prostaglandin E2 Release in Differentiated Caco-2 Cells: Selective Prevention by Growth Factors. J. Pharm. Sci. 2009, 98. [Google Scholar] [CrossRef]
- Ty, M.; D, H.; Lt, T.; D, N.; N, H.; D, B. Cytoskeletal Regulation of Caco-2 Intestinal Monolayer Paracellular Permeability. J. Cell. Physiol. 1995, 164. [Google Scholar] [CrossRef]
- Zt, B.; Sl, G.; Mr, D.; Km, G.; L, Y.; Sf, O.; Jd, L.; Ap, N. Cocoa Procyanidins with Different Degrees of Polymerization Possess Distinct Activities in Models of Colonic Inflammation. J. Nutr. Biochem. 2015, 26. [Google Scholar] [CrossRef]
- Bianchi, M.G.; Chiu, M.; Taurino, G.; Brighenti, F.; Rio, D.D.; Mena, P.; Bussolati, O. Catechin and Procyanidin B2 Modulate the Expression of Tight Junction Proteins but Do Not Protect from Inflammation-Induced Changes in Permeability in Human Intestinal Cell Monolayers. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- K, G.-C.; R, C.; I, G.; A, A.; M, P.; F, V.; X, T.; M, B. Protective Effect of Proanthocyanidins in a Rat Model of Mild Intestinal Inflammation and Impaired Intestinal Permeability Induced by LPS. Mol. Nutr. Food Res. 2019, 63. [Google Scholar] [CrossRef]
- H, W.; T, L.; Ym, L.; Zp, G.; Kq, Z.; Jy, S.; Js, X.; Yp, C. Granny Smith Apple Procyanidin Extract Upregulates Tight Junction Protein Expression and Modulates Oxidative Stress and Inflammation in Lipopolysaccharide-Induced Caco-2 Cells. Food Funct. 2018, 9. [Google Scholar] [CrossRef]
- R, R.; S, M.; Sm, K.; B, O.; Jd, S.; S, A.; D, G.; M, F. Claudin-2, a Component of the Tight Junction, Forms a Paracellular Water Channel. J. Cell Sci. 2010, 123. [Google Scholar] [CrossRef]
- T, O.; H, M.; T, J. Changes in the Expression of Claudins in Active Ulcerative Colitis. J. Gastroenterol. Hepatol. 2008, 23 Suppl 2. [Google Scholar] [CrossRef]
- Das, P.; Goswami, P.; Das, T.; Nag, T.; Sreenivas, V.; Ahuja, V.; Panda, S.; Gupta, S.D.; Makharia, G. Comparative Tight Junction Protein Expressions in Colonic Crohn’s Disease, Ulcerative Colitis, and Tuberculosis: A New Perspective. Virchows Arch. 2012. [CrossRef]
- Talavera, M.M.; Nuthakki, S.; Cui, H.; Jin, Y.; Liu, Y.; Nelin, L.D. Immunostimulated Arginase II Expression in Intestinal Epithelial Cells Reduces Nitric Oxide Production and Apoptosis. Front. Cell Dev. Biol. 2017, 5, 227133. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, S.; Zhao, R.C. The Roles of Mesenchymal Stem Cells in Tumor Inflammatory Microenvironment. J. Hematol. Oncol.J Hematol Oncol 2014, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, B.; Wang, X.; Yu, Q.; Fang, R. Epidermal Growth Factor, through Alleviating Oxidative Stress, Protect IPEC-J2 Cells from Lipopolysaccharides-Induced Apoptosis. Int. J. Mol. Sci. 2018, 19, 848. [Google Scholar] [CrossRef] [PubMed]
- C, G.-Q.; E, R.-G.; R, B.-D.; M, P.; A, A.; Mt, B.; X, T. Health-Promoting Properties of Proanthocyanidins for Intestinal Dysfunction. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2017/2018 | EFSA. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/6007 (accessed on 26 July 2023).
- Zha, L.; Garrett, S.; Sun, J. Salmonella Infection in Chronic Inflammation and Gastrointestinal Cancer. Diseases 2019, 7. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, S.; He, Q.; Sun, K.; Wang, X.; Zhang, X.; Li, Y.; Zeng, J. Crosstalk between Microbial Biofilms in the Gastrointestinal Tract and Chronic Mucosa Diseases. Front. Microbiol. 2023, 14, 1151552. [Google Scholar] [CrossRef]
- G, M.; A, R.; V, C.; Py, D. Galleria Mellonella as a Suitable Model of Bacterial Infection: Past, Present and Future. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- N, R.; C, N.-L.; D, L. The Insect Galleria Mellonella as a Powerful Infection Model to Investigate Bacterial Pathogenesis. J. Vis. Exp. JoVE 2012. [Google Scholar] [CrossRef]
- Lf, G.; Pr, B.; S, B. The Structure and Function of the Smooth Septate Junction in a Transporting Epithelium: The Malpighian Tubules of the New Zealand Glow-Worm Arachnocampa Luminosa. Tissue Cell 1980, 12. [Google Scholar] [CrossRef]
- Pm, C.; At, C.; Er, H.; Pd, E.; Kh, G. Proteomic Analysis of the Peritrophic Matrix from the Gut of the Caterpillar, Helicoverpa Armigera. Insect Biochem. Mol. Biol. 2008, 38. [Google Scholar] [CrossRef]
- T, K.; O, B.; O, O.; N, B.; B, L. Genetic Evidence for a Protective Role of the Peritrophic Matrix against Intestinal Bacterial Infection in Drosophila Melanogaster. Proc. Natl. Acad. Sci. U. S. A. 2011, 108. [Google Scholar] [CrossRef]
- Mukherjee, K.; Hain, T.; Fischer, R.; Chakraborty, T.; Vilcinskas, A. Brain Infection and Activation of Neuronal Repair Mechanisms by the Human Pathogen Listeria Monocytogenes in the Lepidopteran Model Host Galleria Mellonella. Virulence 2013, 4, 324. [Google Scholar] [CrossRef] [PubMed]
- Emery, H.; Johnston, R.; Rowley, A.F.; Coates, C.J. Indomethacin-Induced Gut Damage in a Surrogate Insect Model, Galleria Mellonella. Arch. Toxicol. 2019, 93, 2347–2360. [Google Scholar] [CrossRef] [PubMed]
- Grounta, A.; Harizanis, P.; Mylonakis, E.; Nychas, G.-J.E.; Panagou, E.Z. Investigating the Effect of Different Treatments with Lactic Acid Bacteria on the Fate of Listeria Monocytogenes and Staphylococcus Aureus Infection in Galleria Mellonella Larvae. PLOS ONE 2016, 11, e0161263. [Google Scholar] [CrossRef] [PubMed]
- A, U.; I, U.; S, M.; K, V. Eugenol in Combination with Lactic Acid Bacteria Attenuates Listeria Monocytogenes Virulence in Vitro and in Invertebrate Model Galleria Mellonella. J. Med. Microbiol. 2016, 65. [Google Scholar] [CrossRef]
- Scalfaro, C.; Iacobino, A.; Nardis, C.; Franciosa, G. Galleria Mellonella as an in Vivo Model for Assessing the Protective Activity of Probiotics against Gastrointestinal Bacterial Pathogens. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- K, M.; R, R.; R, F.; A, V. Galleria Mellonella as a Model Host to Study Gut Microbe Homeostasis and Brain Infection by the Human Pathogen Listeria Monocytogenes. Adv. Biochem. Eng. Biotechnol. 2013, 135. [Google Scholar] [CrossRef]
- Nj, S.; Mc, B.; Ol, C.; Se, R.; Rm, L.R.; Mj, W.; Fj, S.; Rw, T. Galleria Mellonella as an Infection Model for Campylobacter Jejuni Virulence. J. Med. Microbiol. 2011, 60. [Google Scholar] [CrossRef]
- Wagley, S.; Borne, R.; Harrison, J.; Baker-Austin, C.; Ottaviani, D.; Leoni, F.; Vuddhakul, V.; Titball, R.W. Galleria Mellonella as an Infection Model to Investigate Virulence of Vibrio Parahaemolyticus. Virulence 2018, 9, 197. [Google Scholar] [CrossRef]
- S, B.; H, G.; Y, Z.; Cl, H.; Dv, Z.; Mm, V. The Galleria Mellonella Larvae as an in Vivo Model for Evaluation of Shigella Virulence. Gut Microbes 2017, 8. [Google Scholar] [CrossRef]
- Card, R.; Vaughan, K.; Bagnall, M.; Spiropoulos, J.; Cooley, W.; Strickland, T.; Davies, R.; Anjum, M.F. Virulence Characterisation of Salmonella Enterica Isolates of Differing Antimicrobial Resistance Recovered from UK Livestock and Imported Meat Samples. Front. Microbiol. 2016, 7, 186885. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- V, S.; E, B.; F, S.; A, C.; O, G.; A, M.; G, P.; Am, P. Biostimulant Effects of Chaetomium Globosum and Minimedusa Polyspora Culture Filtrates on Cichorium Intybus Plant: Growth Performance and Metabolomic Traits. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- G, C.; F, S.; P, G.; S, M.; L, F.; G, G.; Ar, C.; G, C.; Me, D.C.; F, A. Mantonico and Pecorello Grape Seed Extracts: Chemical Characterization and Evaluation of In Vitro Wound-Healing and Anti-Inflammatory Activities. Pharm. Basel Switz. 2020, 13. [Google Scholar] [CrossRef]
- Dinicola, S.; Cucina, A.; Pasqualato, A.; D’Anselmi, F.; Proietti, S.; Lisi, E.; Pasqua, G.; Antonacci, D.; Bizzarri, M. Antiproliferative and Apoptotic Effects Triggered by Grape Seed Extract (GSE) versus Epigallocatechin and Procyanidins on Colon Cancer Cell Lines. Int. J. Mol. Sci. 2012, 13, 651. [Google Scholar] [CrossRef]
- Provenzani, R.; San-Martin-Galindo, P.; Hassan, G.; Legehar, A.; Kallio, A.; Xhaard, H.; Fallarero, A.; Yli-Kauhaluoma, J. Multisubstituted Pyrimidines Effectively Inhibit Bacterial Growth and Biofilm Formation of Staphylococcus Aureus. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Cui, Z.-H.; He, H.-L.; Zheng, Z.-J.; Yuan, Z.-Q.; Chen, Y.; Huang, X.-Y.; Ren, H.; Zhou, Y.-F.; Zhao, D.-H.; Fang, L.-X.; et al. Phentolamine Significantly Enhances Macrolide Antibiotic Antibacterial Activity against MDR Gram- Bacteria. Antibiotics 2023, 12, 760. [Google Scholar] [CrossRef]
- Efficacy and Safety of Phage Therapy against Salmonella Enterica Serovars Typhimurium and Enteritidis Estimated by Using a Battery of in Vitro Tests and the Galleria Mellonella Animal Model. Microbiol. Res. 2022, 261, 127052. [CrossRef]
Figure 1.
1H spectrum of procyanidins-rich grape seed extract.
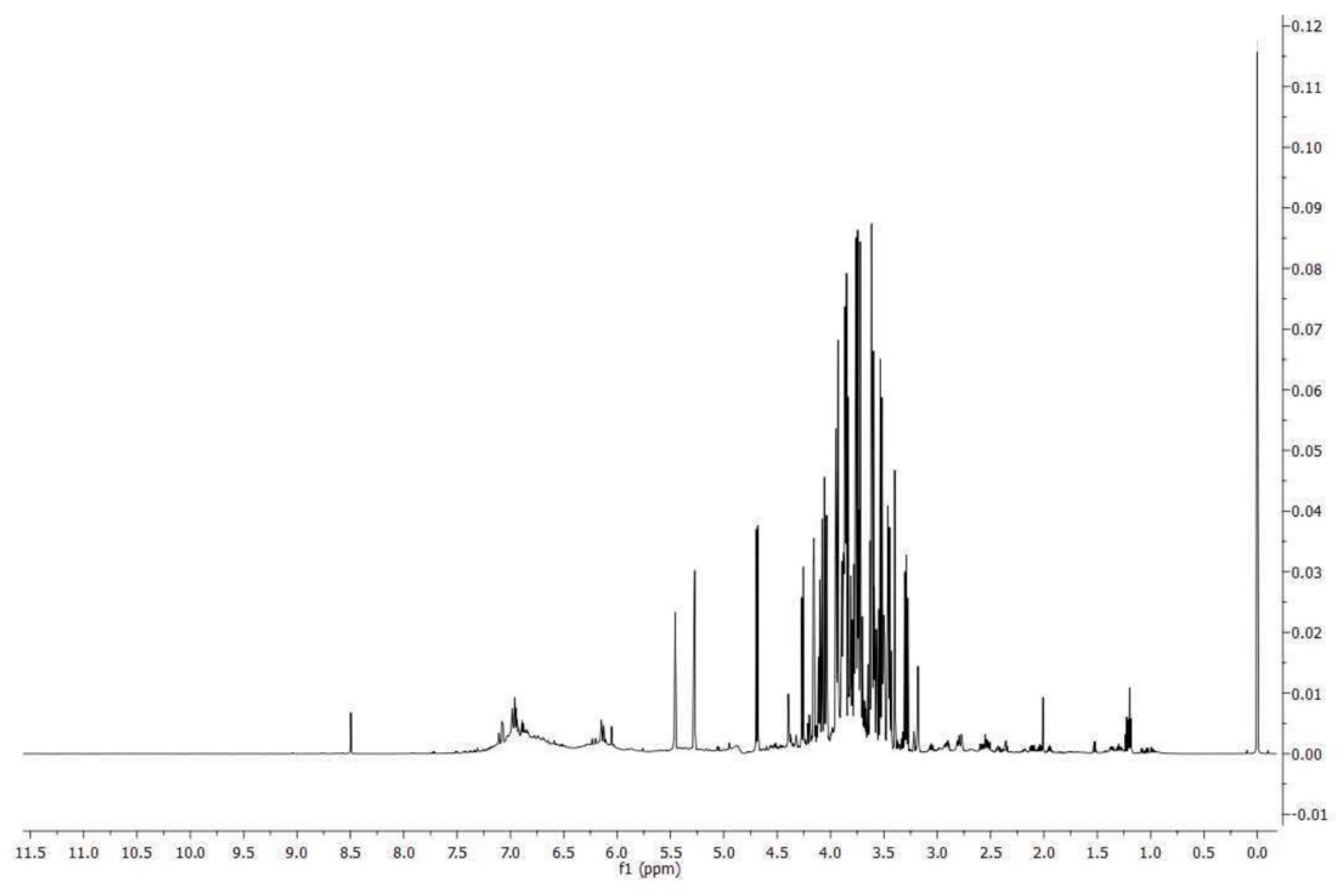
Figure 2.
Barrier function of the Caco-2 cell monolayers was analyzed through the FD-4 permeability assay. LPS treatment (10 ug/mL) for 24 h, prGSE (6.25 ug/mL) for 24 h and LPS with prGSE treatment for 24 h. The amount of FD-4 (expressed in pmol) accumulated in the receiver compartment, was plotted for each treatment. The experiments were executed in triplicate. Data are reported as mean ± SD. Statistical analysis was performed using a two-way ANOVA, followed by Bonferroni’s post-hoc correction test. *p<0.05. Ctrl (Control); LPS (Lipopolysaccharide); prGSE (procyanidins-rich Grape Seed Extract); prGSE+LPS (Mix).
Figure 2.
Barrier function of the Caco-2 cell monolayers was analyzed through the FD-4 permeability assay. LPS treatment (10 ug/mL) for 24 h, prGSE (6.25 ug/mL) for 24 h and LPS with prGSE treatment for 24 h. The amount of FD-4 (expressed in pmol) accumulated in the receiver compartment, was plotted for each treatment. The experiments were executed in triplicate. Data are reported as mean ± SD. Statistical analysis was performed using a two-way ANOVA, followed by Bonferroni’s post-hoc correction test. *p<0.05. Ctrl (Control); LPS (Lipopolysaccharide); prGSE (procyanidins-rich Grape Seed Extract); prGSE+LPS (Mix).
Figure 4.
Percentage of inhibition biofilm formation of Escherichia coli ATCC 25922 and Salmonella enterica serovar Typhimurium ATCC 14028 after 24 h of incubation.
Figure 4.
Percentage of inhibition biofilm formation of Escherichia coli ATCC 25922 and Salmonella enterica serovar Typhimurium ATCC 14028 after 24 h of incubation.
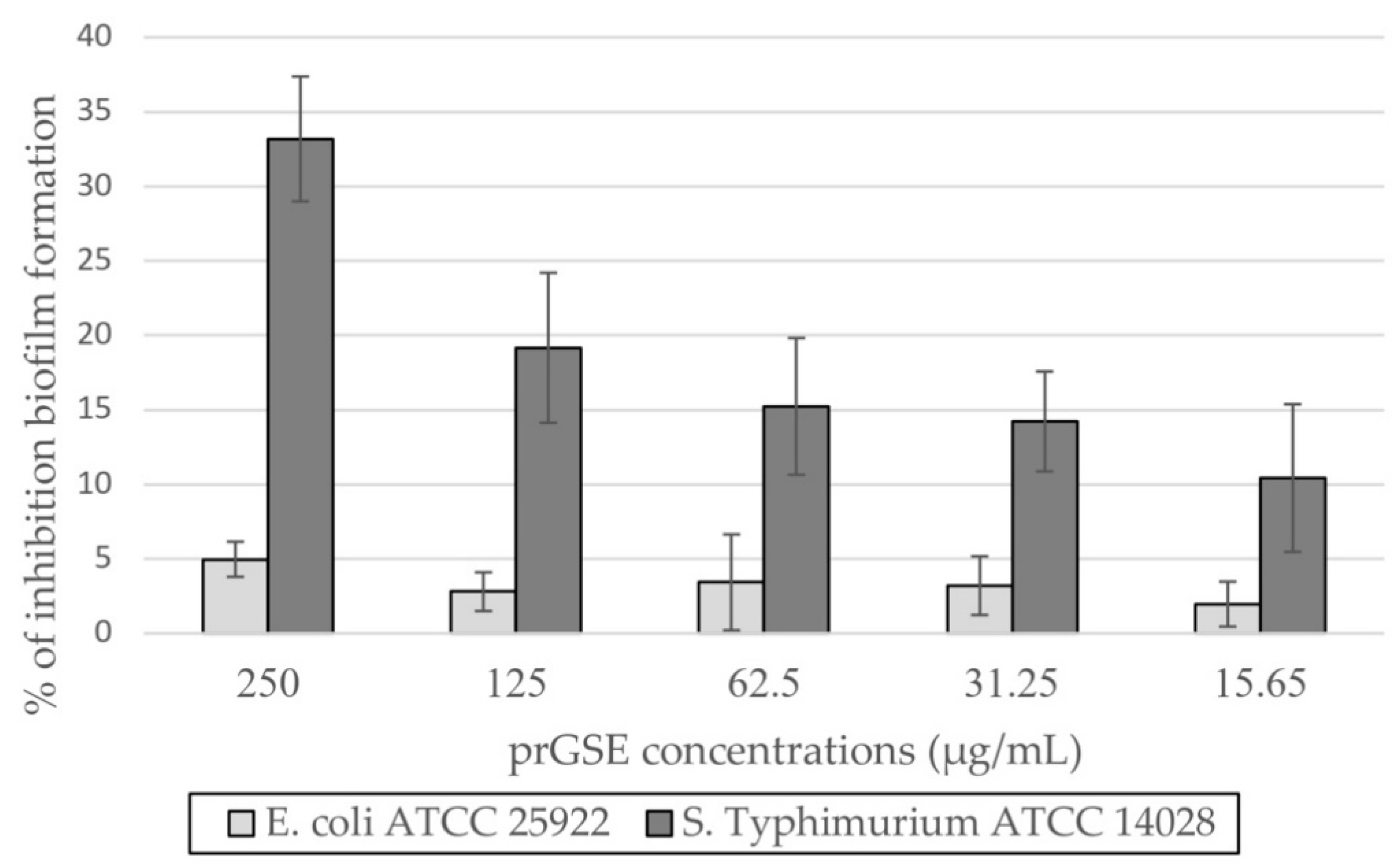
Figure 5.
Survival curves of Galleria mellonella larvae infected with Salmonella enterica serovar Typhimurium ATCC 14028 and treated with 500 µg/mL (a), 250 µg/mL (b), 125 µg/mL (c) of procyanidins-rich grape seed extract (prGSE). The Mantel-Cox log-rank test was used to judge the statistical significance relative to the control. * p < 0.05 compared to the control; ** p < 0.01 compared to the control; ns=not significant.
Figure 5.
Survival curves of Galleria mellonella larvae infected with Salmonella enterica serovar Typhimurium ATCC 14028 and treated with 500 µg/mL (a), 250 µg/mL (b), 125 µg/mL (c) of procyanidins-rich grape seed extract (prGSE). The Mantel-Cox log-rank test was used to judge the statistical significance relative to the control. * p < 0.05 compared to the control; ** p < 0.01 compared to the control; ns=not significant.
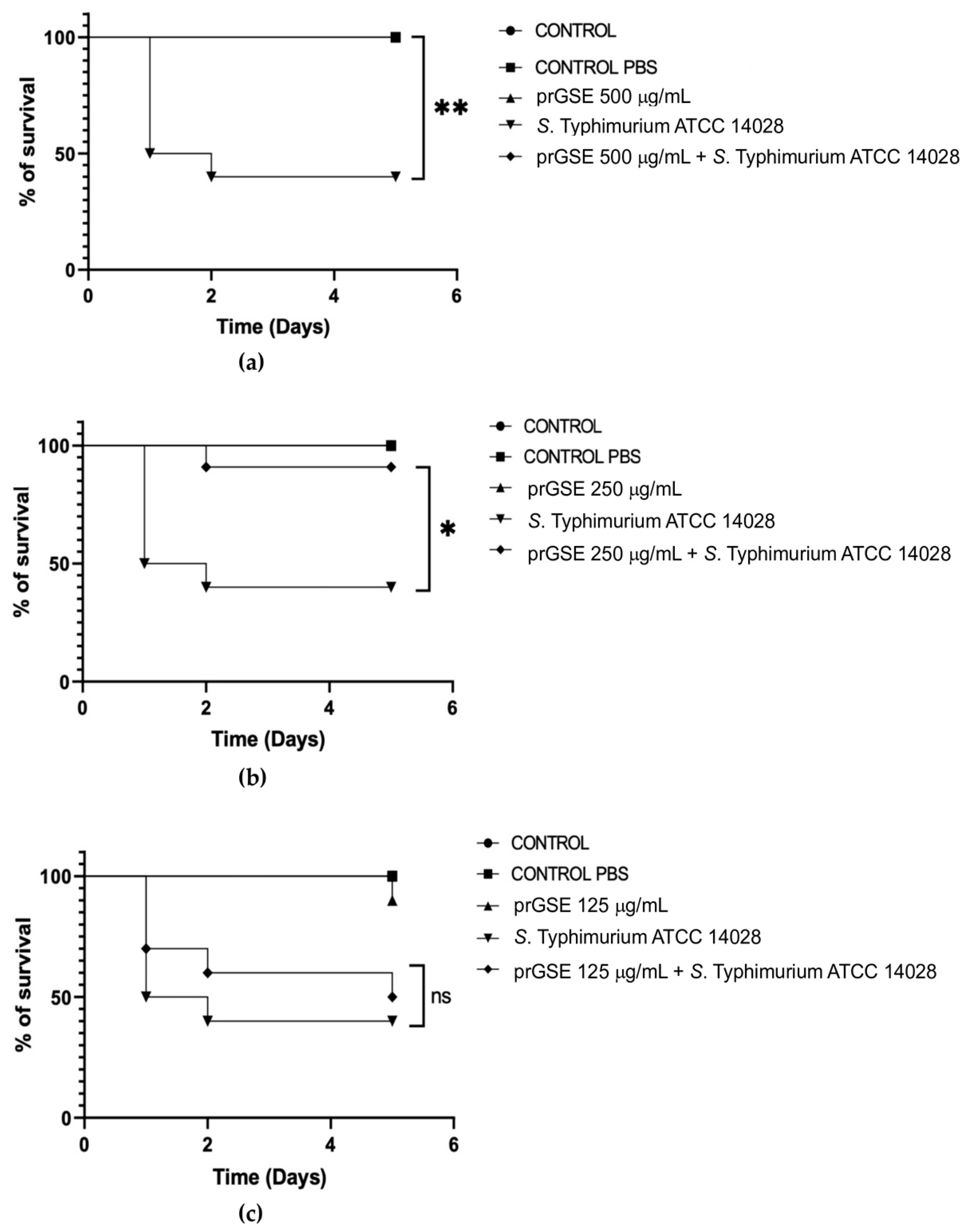
Figure 6.
Survival curves of Galleria mellonella larvae infected with Eschericia coli ATCC 25922 and treated with 500 µg/mL (a), 250 µg/mL (b), 125 µg/mL (c) of procyanidins rich grape seed extract (prGSE). The Mantel-Cox log-rank test was used to judge the statistical significance relative to the control. * p < 0.05 compared to the control; ** p < 0.01 compared to the control; ns=not significant.
Figure 6.
Survival curves of Galleria mellonella larvae infected with Eschericia coli ATCC 25922 and treated with 500 µg/mL (a), 250 µg/mL (b), 125 µg/mL (c) of procyanidins rich grape seed extract (prGSE). The Mantel-Cox log-rank test was used to judge the statistical significance relative to the control. * p < 0.05 compared to the control; ** p < 0.01 compared to the control; ns=not significant.
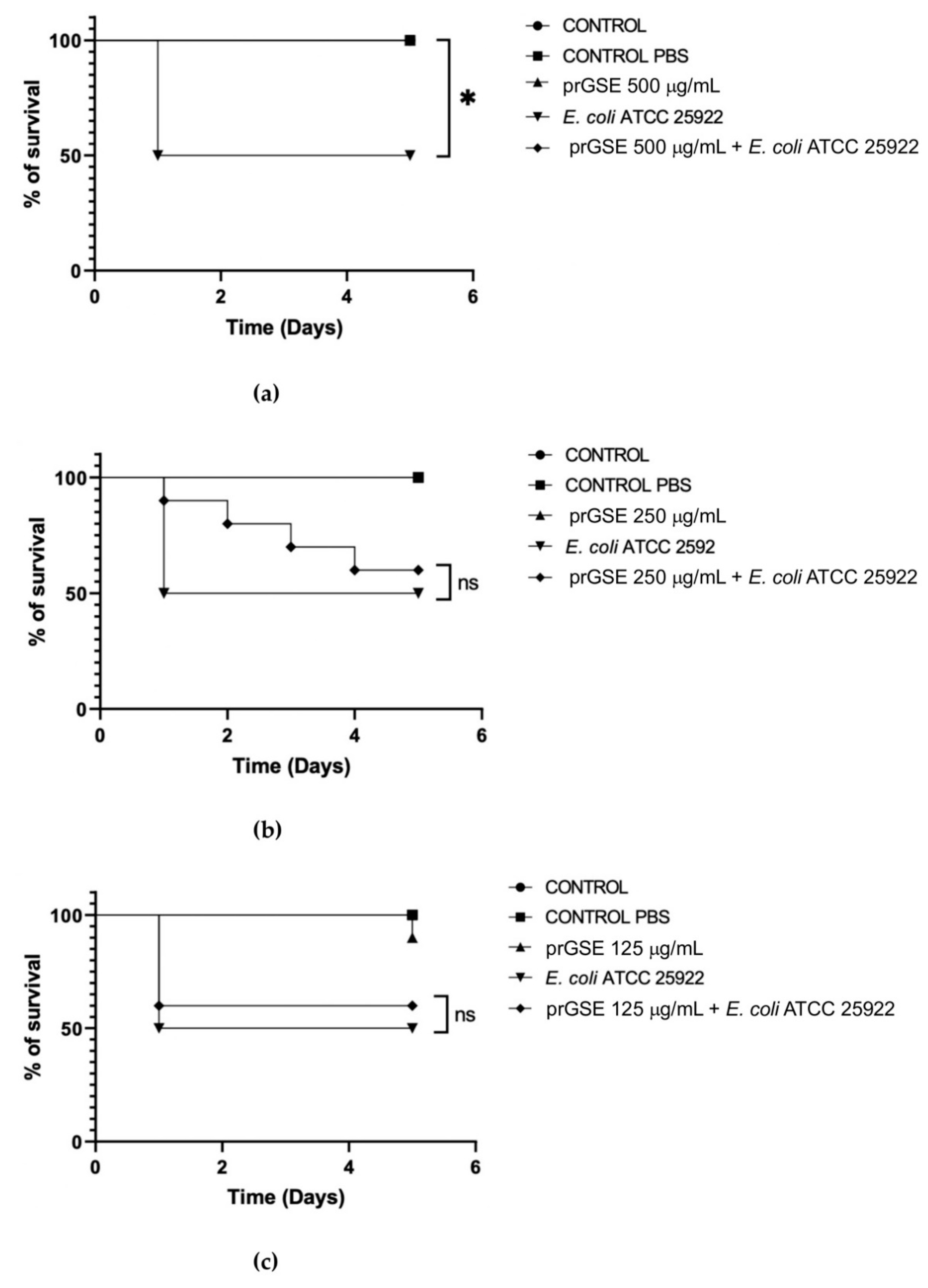
Table 1.
Compounds determined by HPLC-DAD, their elution times and concentrations.
| Compound | Retention Time (Min) | Area | mg/g |
|---|---|---|---|
| Catechin (1) | 11.65 | 408.94 | 10.92 |
| Procyanidin B2 (2) | 16.86 | 106.99 | 4.05 |
| Epicatechin (3) | 19.03 | 372 | 7.66 |
| Epicatechin gallate (4) | 30.54 | 261.12 | 2.80 |
| Procyanidin Pol 1 (5) | 48.59 | 9694.75 | 365.48 |
| Procyanidin Pol 2 (6) | 52.00 | 2078.38 | 78.36 |
1 Compounds determined by HPLC-DAD, their elution times, and concentrations. SD% values are lower than 5%.
Table 2.
Molecule amount measured by 1H NMR in procyanidins-rich grape seed extract.
| Molecule | Amount (mg/100 mg dry extract) | |
|---|---|---|
| Amino acids | Leucine | 0.01461 ± 0.00073 |
| Isoleucine | 0.01417 ± 0.00071 | |
| Valine | 0.0228 ± 0.0011 | |
| Threonine | 0.00199 ± 0.00010 | |
| Alanine | 0.0403 ± 0.0021 | |
| GABA | 0.1199 ± 0.0062 | |
| Glutamine | 0.1536 ± 0.0077 | |
| Aspartate | 0.372 ± 0.019 | |
| Asparagine | 0.027 ± 0.0014 | |
| Phenylalanine | 0.034 ± 0.0017 | |
| Tryptophan | 0.0838 ± 0.0042 | |
| Organic acids | Lactate | 0.00633 ± 0.00032 |
| Acetate | 0.056 ± 0.0028 | |
| Citrate | 0.1027 ± 0.0051 | |
| Malate | 0.445 ± 0.022 | |
| Ascorbate | 0.1343 ± 0.0067 | |
| Formate | 0.1587 ± 0.0079 | |
| Carbohydrates | Glucose | 13.55 ± 0.68 |
| Sucrose | 8.17 ± 0.41 | |
| Miscellaneous molecules | Ethanol | 0.1031 ± 0.0052 |
| Choline | 0.1146 ± 0.0057 | |
| Procyanidin B1 | 2.66 ± 0.13 | |
| Polymeric Procyanidin (eq. Procyanidin B1) | 25.3 ± 1.27 | |
| Trigonelline | 0.025 ± 0.0012 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



