
Submitted:
31 July 2023
Posted:
01 August 2023
You are already at the latest version
Abstract
In rice, symbiotic N fixation via nodule bacteroids does not take place naturally. Although N fixation by endophytic and associative diazotrophs has been reported in rice, main organs and seasonal regulation for the N fixation have not been elucidated. In this study, therefore, seasonal changes of nitrogenase (acetylene reduction) activity and carbohydrate contents in culm internode (IN), basal nodes (BN), and crown root (CR) were investigated in manure compost (MC)- and chemical fertilizer-applied rice. Nitrogenase activity increased after rooting, and reached the highest activity in BN of MC-applied rice at panicle initiation. In BN of MC-applied rice, the sucrose content was low whereas the higher glucose content was detected at panicle initiation. Although bacterial and diazotrophic bacterial populations were most abundant in CR, the nitrogenase activity seemed to be restricted by its trace sucrose and glucose contents. In addition, alpha-Proteobacteria and Clostridia phyla were abundant in BN-specific bacterial microbiota at panicle initiation. These results suggest that the increased N fixation in BN at panicle initiation might be sustained both by the non-structural carbohydrate supply from photosynthetic apparatus and BN-specific bacterial microbiota including diazotrophs.
Keywords:
nitrogen fixation
; rice
; basal nodes
; unelongated stem
; carbohydrate
; manure compost
; nitrogenase
; nifH
1. Introduction
Symbiotic diazotrophic bacteria play a crucial role in biological N fixation (BNF) under catalysis of nitrogenases, which can partly supply N compounds to host plants. N fixation in the symbiotic associations between legume plants and diazotrophs inside root nodules are well known in agricultural systems. Dicarboxylates (fumarate, succinate, or L-malate) are supplied from legume plants to nodule bacteroids via the dicarboxylic-acid transport system, and these organic acids fuel the energy for nitrogenase activity [1].
In the non-symbiotic systems, rhizosphere-associative diazotrophic bacteria fix N by using carbon and energy sources supplied from the environment, and release the fixed N probably after lysis of the bacterial cells [2]. In addition, endophytic diazotrophs are often observed and fix N with a wide variety of plant roots including cereals. Dinitrogenase reductase-encoding gene, nifH, and its expression have been detected in agronomically significant non-legume plants and their rhizosphere such as the maize stems and roots, the maize rhizosphere soil, the sorghum roots and rhizosphere, the switch grass shoots and roots, the sugarcane stems and roots, the sugarcane leaf sheath, and the sweet potato stems and storage tubers [3]. It has been reported that the carbohydrate-rich mucilage secreted from the aerial roots of the maize landrace are enriched in many known diazotrophic species and the homologs of genes encoding nitrogenase subunits, and harbor nitrogenase activity [4].
In terms of N fixation in paddy rice fields, the interface of the root-soil system has been reported as the important site of N fixation, and the rhizosphere-associated bacteria responsible for the N fixation were thought to be heterotrophic diazotrophs such as Azotobacter and Clostridia bacteria [5]. BNF by the methane-oxidizing methanotrophs takes place in the root tissues and also in the surface soil. The functional genes for methane oxidation and plant association were abundant in rice roots under low levels of N fertilizer application [6]. It has been shown that methane oxidation and N fixation are simultaneously activated in the root zone of rice in low-N paddy fields, and both processes are likely controlled by OsCCaMK gene encoding Ca2+/calmodulin-dependent protein kinase, which is the central component of common symbiosis pathway [7]. The short filaments and coiled masses of N-fixing cyanobacteria were observed near the epidermis and cortex of roots and shoot tissues [8]. It has been suggested that rice rhizosphere-associative bacteria compete carbon and energy sources supplied from the environment between N fixation by diazotrophs and methane emission by methanogens or sulfate reduction by sulfate-reducing bacteria [9]. However, there is limited information about main organs and seasonal regulation for N fixation of rice cultured in paddy fields.
A rice culm has 13–16 nodes, which are composed of the upper 4 to 5 nodes and the residual basal nodes (BN) [10]. The node is a junction of vasculatures connecting to leaves, stems, panicles, and roots. The long internodes (IN) are separated by the upper nodes and they participate in the culm elongation [11]. On the other hand, BNs form an unelongated stem and they are connected to crown roots (CR). It has been reported that the endophytic bacteria seem to enter the root of rice from the rhizosphere, and colonize the root’s intracellular spaces, aerenchyma, and cortical cells, the vascular tissue, the xylem vessels, the epidermal cells, the intracellular spaces, and the substomatal cavities of leaves and stem [12,13,14]. Therefore, rice roots are exposed to and associated with various kinds of rhizospheric bacteria including diazotrophs, whereas the rice vasculatures may contain abundant carbohydrates produced by photosynthetic apparatus, which could be metabolized by diazotrophs to fuel BNF.
In the previous study, long-term manure compost (MC) application to the paddy soil resulted in the increases of potential nitrogenase activity and bacterial alpha diversity in the soil at the rice panicle initiation stage, compared to those in the long-term chemical fertilizer (CF) application [15]. Therefore, bacterial microbiota in rice cultured in these plots may be affected by the differences in soil bacterial microbiota and chemical compositions in these soils. In this study, seasonal changes of nitrogenase activity, carbohydrate contents, and bacterial microbiota in IN, BN, and CR of rice cultured in MC- and CF-applied plots were investigated from transplantation to ripening.
2. Materials and Methods
2.1. Rice cultivation and sampling
A rice plant (Oryza sativa ssp. japonica) cultivar Udai 21 was cultured in the research farm at Utsunomiya University (Moka, Tochigi, Japan). CF (Japan agricultural cooperatives) and cow MC prepared in the research farm have been separately applied to allophanic andosol fields since 1991. In May and July, CF each equivalent to 3 kg N/1,000 m2 was applied, while in March, MC equivalent to 9.2 kg N/1,000 m2 was applied. Three stocks each of rice plant were collected from 180 m2 (MC) and 270 m2 (CF) plots on 7 June (rooting), 5 July (tillering), 2 August (panicle initiation), and 27 September (ripening). Soil of the rice stocks was shaken off from the root, the aerial part was removed by scissors, and the residual part was washed with a gentle stream of tap water. The residual part was cut and divided into 1-2 cm-length unelongated culm composed of BN, IN that was the 5 cm upper section of the unelongated culm, and CR that had the 5 cm outer section of the unelongated culm by scissors sterilized with 70% ethanol (Figure 1). The rice specimens were stored in plastic bags both at 4°C and −30°C.
2.2. Measurement of nitrogenase activity
Bacterial nitrogenase activity was measured by the acetylene reduction assay. The rice specimens stored at 4°C were used in a day of sampling. Approximately 10 g fresh weight specimen was poured into 100-mL Erlenmeyer flask without addition of external energy and electron sources such as glucose. After covering the flask with a double cap (Kokugo, Tokyo, Japan), the flasks were sealed with plastic tape. After injection of 5 mL acetylene through needle and syringe, the flasks were incubated at 30°C for 24 h.
After incubation, 100 µL of gas phase in the flask was taken by Gastight syringe 81000 (Hamilton, Reno, NV, USA), and ethylene formed was quantified by a gas chromatograph GC-4000 (GL Sciences) equipped with a flame ionization detector and a capillary column Rt-Alumina BOND/KCl (30 m, 0.32 mm ID, 5 µm df, Restek, Tokyo, Japan). N was used as a carrier gas at a flow rate of 3 mL/min. The injector and column temperatures were kept at 200°C and 100°C, respectively.
2.3. Measurements of sucrose and glucose concentrations
The rice specimens stored at −30°C were used. Plant extracts containing carbohydrates were prepared by treating approximately 0.04-1.44 g fresh weight rice specimens with 5 mL boiling ethanol/water (4/1, v/v) 1-2 times [16]. Glass tubes for extraction were capped with glass marbles and heated at 80-85°C for 10 min in an aluminum alloy block placed on a hot stirrer. The ethanol/water extracts were put together, and volumes of the ethanol/water solution were measured. Sucrose and glucose concentrations in the ethanol/water solution were measured using Enzytex liquid sucrose/D-glucose (J.K. International, Tokyo, Japan) and Glucose CII-test Wako (Fujifilm Wako Pure Chemical, Osaka, Japan), respectively.
2.4. Evaluation of endophytic bacterial microbiota
The rice specimens stored at −30°C were used. DNA in rice IN, BN, and CR was extracted using DNeasy Plant Mini Kit (Qiagen, Tokyo, Japan). In DNA extraction from rice plant, a 3-mm diameter stainless steel bead (SUS304, Taitec, Koshigaya, Saitama, Japan) was put in NucleoSpin Bead Tubes Type A (Macherey-Nagel, Düren, Germany). Rice plant pieces placed in the beads tubes were subjected to bead beater treatment using a beads crusher µT-12 (Taitec). Construction of the metagenomic library targeting the 16S rRNA gene V3 and V4 region and sequencing by a next-generation sequencer MiSeq (Illumina, San Diego, CA, USA) were performed with an NGS service (Macrogen, Tokyo, Japan) as described previously. The primer sequences were trimmed, pair sequence reads were merged, and the merged reads comprising 400-470 nucleotides were extracted using Geneious Prime ver. 2019.2.3 (Tomy Digital Biology, Tokyo, Japan). Sequence data were analyzed by mothur (v. 1.47.0) [17]. During the data processing, chloroplast sequences were identified and removed. Measurements of alpha diversity, beta diversity, and the relative taxonomic abundance were performed according to the procedure described previously [15]. The sequences were classified with the Greengenes reference database.
2.5. Quantification of bacteria read/chloroplast read and nifH/bacterial 16S rRNA gene
A ratio of bacteria read number to rice chloroplast read number was calculated by an equation:
where a is the total read number and b is the chloroplast read number in the 16S rRNA gene-targeted metagenomic analysis as described above.
Bacterialread/Chloroplasticread = (a − b)/b
Bacterial genes were amplified using quantitative PCR with KOD SYBR qPCR Mix (Toyobo, Osaka, Japan) and primer sets of PolF (TGCGAYCCSAARGCBGACTC)/PolR (ATSGCCATCATYTCRCCGGA) for nifH [18] and Univ-341-F (CCTACGGGAGGCAGCAG)/Univ-907-R (CCCCGTCAATTCCTTTGAGTTT) for 16S rRNA gene [19,20]. The cycle threshold (Ct) of each DNA sample extracted from the rice specimens was determined with ABI7500 real-time PCR system (ThermoFisher Scientific, Tokyo, Japan). Relative concentrations for nifH (CnifH) and 16S rRNA gene (C16S rRNA) were calculated using an equation of 1/2Ct. A ratio of nifH copy number to bacterial 16S rRNA gene copy number was calculated by an equation:
where a is the total read number and b is the chloroplast read number in the 16S rRNA gene-targeted metagenomic analysis as described above.
nifH/bacterial 16S rRNA gene = a·CnifH/((a − b)·C16S rRNA)
3. Results
3.1. Seasonal changes of bacterial nitrogenase activity in the rice parts
In order to investigate the rice parts for endophytic and associative bacterial N fixation, rice stocks were collected from CF and MC plots from June (21 d after transplantation, rooting) to September (133 d, ripening). The bacterial nitrogenase activity was measurable without addition of external energy and electron sources. The nitrogenase activity increased from June to August (77 d, panicle initiation) in BN and IN whereas it increased from June to July (49 d, tillering) and then decreased in August and September in CR (Figure 2). In August, it was significantly higher at BN than IN and CR, and BN collected from MC-applied plot exhibited higher nitrogenase activity than BN collected from CF-applied plot.
3.2. Seasonal changes of sucrose and glucose contents in the rice parts
Seasonal changes of sucrose and glucose contents were observed in IN and BN (Figure 3). These contents were low or not detected in CR from June to September, suggesting restriction of the nitrogenase activity in CR by the low or undetectable concentrations of the carbon sources. In August, regardless of the difference of applied materials, sucrose contents were lower in BN than in IN whereas glucose contents were higher in BN than in IN. The result suggests that consumption of sucrose and its metabolism to glucose might be activated in BN at panicle initiation.
3.3. Bacterial and nifH abundances in the rice parts at panicle initiation
Bacterial DNA was extracted from IN, BN, and CR of rice collected from MC- and CF-applied plots in August. Amplicon sequences of 16S rRNA gene V3 and V4 region revealed that the ratio of bacterial DNA read numbers to chloroplastic read numbers was 0.5-2.3 in IN, 0.6-2.5 in BN, and 5.5-9.7 in CR (Figure 4A). The result suggests that bacterial DNA was more abundant in CR than in IN and BN. Furthermore, the ratio of nifH gene copy number to bacterial 16S rRNA gene copy number was higher in CR than in IN and BN, and it was higher in CR of rice collected from MC-applied plot than in CR of rice collected from CF-applied plot (Figure 4B). The result suggests that population of diazotrophs among the total bacteria might be higher in CR than in IN and BN.
3.4. Evaluation of bacterial microbiota in the rice parts at panicle initiation
The numbers of observed OTUs with given sequence numbers tended to be higher in CR-specific bacterial microbiota in August (Figure 5). The observed OTU number in MC-applied CR steeply rose with the sequence number increase compared to that in CF-applied CR. On the other hand, no significant differences in the rarefaction curves were observed between MC-applied and CF-applied rice plants in IN- and BN-specific bacterial microbiota.
Alpha diversity within the bacterial microbiota of rice parts in August was measured with Shannon and inverse Simpson indices. The higher values for both Shannon and inverse Simpson indices were observed in CR than in IN and BN (Table 1). However, no differences in alpha diversity were observed between the MC- and CF-applied rice parts.
Beta diversity among the bacterial microbiota of rice parts in August were compared with the principal coordinate analysis (PCoA). The dissimilarity of bacterial microbiota was observed between CR and IN or BN (Figure 6). The coordinate points for the IN and BN bacterial microbiota were more closely plotted. The bacterial microbiota in all the plant parts were not distinguished between MC- and CF-applied rice.
Differences in the relative taxonomic abundances were compared within the bacterial microbiota of rice parts in August. The class-level taxonomic profiles in the IN-, BN-, and CR-specific bacterial microbiota were similar between MC- and CF-applied rice parts (Figure 7). In comparison with CR, alpha-Proteobacteria class, among which an unclassified species of Rhizobiales order was included, was abundant, and beta-, gamma-, and delta-Proteobacteria were decreased in IN and BN. Clostridia class, among which two unclassified species of Clostridiales order were included, was specifically higher in BN. An unclassified species of Veillonellaceae family, which belongs to Negativicutes class, was specifically higher in BN. These results indicate higher abundances of total bacterial number and nifH copy number in CR and the dissimilarity between CR- and IN- or BN-specific bacterial microbiota.
4. Discussion
Sucrose originated from carbon assimilation in leaves is transported and translocated from photosynthetic apparatus to non-photosynthetic sinks such as fruits, seeds, and developing tubers [21]. The synergy between abscisic acid and sucrose increased grain yield and quality by improving the source-sink relationship. Thus, sucrose is an energy source of plants and a signaling molecule that regulates plant development. This study showed that the sucrose content increased after rooting in IN of rice culm. It has been reported that the upper leaf sheath of rice serves as a temporary starch sink before heading and it subsequently becomes a carbon source tissue to the growing panicle at the post-heading stage. The time of sink–source transition in upper leaf sheaths is highly correlated to the panicle exsertion [22]. Although sucrose translocation might be activated at panicle initiation in rice culm, the sucrose contents in BN were lower than those in IN. Considering the significantly higher nitrogenase activity in BN compared to those in IN at panicle initiation, the translocated sucrose may be consumed by the endophytic bacteria in BN to fuel the nitrogenase activity. In addition, there is another possibility that the translocated sucrose might be hydrolyzed into glucose and fructose by various isoforms of invertase [23]. Grasses including rice have the ability to buffer the sink–source interaction by transiently storing carbohydrates in stem tissue when production from the source is greater than whole-plant demand [24]. Cells surrounding rice stem small vascular bundles accumulated more non-structural carbohydrates than those surrounding large vascular bundles before heading via enhancing sucrose hydrolysis and starch accumulation in small vascular bundles [23]. In this study, the high glucose contents were observed after tillering, suggesting that the glucose contents detected in BN might originate from the translocated sucrose and the glucose contents might sustain the nitrogenase activity in BN after tillering. It has been shown through cultivation of the rice genotypes for 2 years that the stem N-fixing (acetylene reduction) activity was correlated with the stem levels of soluble sugars [25]. The rice basal nodes as a junction of the vascular systems are important in distribution of iron and zinc, which are taken up by root, to leaf sheath, developing tiller bud, and panicle [26,27]. Considering possibility that rhizosphere bacteria enter root and might be distributed to the rice parts through the vascular systems as shown in the cases of iron and zinc, the basal nodes could be the first part for the endophytic bacteria to contact to high concentrations of non-structural carbohydrates. The results in this study together with the knowledge from preceded studies suggest that basal nodes of rice unelongated culm might be a main part for N fixation by endophytic diazotrophs at panicle initiation and thereafter, which was affected by the contents of translocated non-structural carbohydrates such as sucrose and glucose.
This study revealed that the ratio of bacterial read number to rice chloroplastic read number and the ratio of nifH gene copy number to bacterial 16S rRNA gene copy number were higher in CR DNA than in IN and BN DNA. It was also shown that the number of OTUs with given sequence numbers and the indices of alpha diversity were higher in CR-specific bacterial microbiota than those in IN and BN. These results suggest that total bacterial and diazotrophic bacterial populations might be more abundant and diverse in CR than in IN and BN. It has been reported that more diverse endophytic bacterial OTUs are detected in roots than in stems of rice samples [28]. In addition, nifH gene copy numbers are not significantly correlated with N-fixing activities in soil even in the presence of external glucose and malate solution [29,30,31]. Considering that no or trace amounts of sucrose and glucose were detected in CR throughout the rice cultivation period, it could be concluded that the lower nitrogenase activity in CR might be due to the restriction of carbon source supply even in the presence of abundant and diverse populations of diazotroph.
PCoA and comparison of relative taxonomic abundances in rice bacterial microbiota at panicle initiation indicated the dissimilarity of bacterial community structures between CR and IN or BN. The class of alpha-Proteobacteria and an unclassified species of Rhizobiales order were abundant in IN and BN, whereas beta-, gamma-, and delta-Proteobacteria were abundant in CR. The class of Clostridia and two unclassified species of Clostridiales order were specifically higher in BN. The phylum abundances of rice root and stem have been demonstrated in the preceded studies. Endophytic bacterial diversity in rice root consists of alpha, beta, gamma, delta, and epsilon subclasses of Proteobacteria, and Cytophaga/Flexibacter/Bacteroides (CFB) phylum [32], and the dominant group was beta-Proteobacteria [33,34]. In the stems of four rice genotypes, the classification by class levels have revealed that alpha-Proteobacteria is the most prevalent at the heading stage [25]. The previously shown dominant phyla, beta-Proteobacteria in root and alpha-Proteobacteria in stem, are consistent with the dominant phyla shown in this study. BNF by some diazotrophic bacteria like Azotobacter, Clostridium, Azospirillum, Herbaspirillum, Burkholderia, and Rhizobium can substitute for urea-N in CF [35]. In addition, several Rhizobium species have been isolated from rice root and rhizosphere [36,37,38,39,40]. In the class of Negativicutes, which was not a dominant class in the rice parts, an unclassified species of Veillonellaceae family was specifically higher in BN-specific bacterial microbiota. The high abundance of Veillonellaceae family has been shown first on rice straw and only later in the rice straw-amended soil [41]. Metagenomic analysis targeting 16S rRNA gene have reported the increase of Veillonellaceae family in the rice base including 10 cm stem and 1 cm root caused by Azospirillum sp. B510 inoculation [42]. Uncultivated diazotrophs such as Veillonellaceae and Costridiaceae dominated the communities of responsive phylotypes in anaerobic soil microcosms amended with glucose and glucose plus urea [43]. Therefore, unclassified species of Rhizobiales and Clostridiales orders and Veillonellaceae family may be responsible for N fixation in BN with the translocated and metabolized carbohydrates at panicle initiation.
In this study, factors that enhanced the nitrogenase activities in BN of MC-applied rice at panicle initiation could not be clarified. It has been suggested that low N application leads to the increased accumulation and remobilization of stem starch and non-structural carbohydrates in the rice cultivar SY63 via increases of the activities of adenosine diphosphate-glucose pyrophosphorylase, starch synthase, starch branching enzyme, α-amylase, β-amylase, and sucrose phosphate synthase [44]. In addition, this study showed that the sucrose contents in MC-applied rice BN were maintained at lower levels than those in CF-applied rice BN. Therefore, soil nutritional conditions such as the lower ammonia contents caused by MC application [15] may result in changes of non-structural carbohydrate flux and consumption of carbon sources by endophytic bacteria in BN of rice unelongated culm.
5. Conclusions
BN of unelongated culm exhibited the highest nitrogenase activity and the decreased sucrose content at panicle initiation of rice cultured in MC-applied plot. Although bacterial and diazotrophic bacterial populations were higher in CR than in BN, the trace nitrogenase activity was detected in CR, suggesting restriction of the activity by the limited carbohydrate supply from photosynthetic apparatus to CR. In IN and BN at panicle initiation, alpha-Proteobacteria including an unclassified species of Rhizobiales order was abundant in BN and IN, and Clostridia phylum and an unclassified species of Veillonellaceae family were specifically higher in BN. It is likely that the BN-specific bacterial microbiota together with the higher contents of glucose in BN might participate in the increased nitrogenase activity at panicle initiation and ripening.
Author Contributions
Conceptualization, I.M., H.H., C.H. and Y.T.; methodology, I.M., H.H., C.H. and Y.T.; software, I.M.; formal analysis, Z.A. and M.T.; resources, H.H., C.H. and Y.T.; data curation, Z.A. and I.M.; writing—original draft preparation, I.M.; writing—review and editing, I.M.; funding acquisition, I.M. and H.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a Grant-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science (17K07709)
Data Availability Statement
The raw data associated with this study were deposited into the DNA Data Bank of Japan (DDBJ), BioProject database (accession: PRJDB14253).
Acknowledgments
This work was carried out based on management in paddy rice production by stuffs of Utsunomiya University Farm.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lodwig, E.M.; Hosie, A.H.; Bourdes, A.; Findlay, K.; Allaway, D.; Karunakaran, R.; Downie, J.; Poole, P.S. Amino-acid cycling drives nitrogen fixation in the legume–Rhizobium symbiosis. Nature 2003, 422, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, K.; Makaju, S.; Ibrahim, R.; Missaoui, A. Current progress in nitrogen fixing plants and microbiome research. Plants 2020, 9, 97. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, T.; Terakado-Tonooka, J.; Bao, Z.; Minamisawa, K. Molecular analyses of the distribution and function of diazotrophic rhizobia and methanotrophs in the tissues and rhizosphere of non-leguminous plants. Plants 2019, 8, 408. [Google Scholar] [CrossRef]
- Van Deynze, A.; Zamora, P.; Delaux, P.-M.; Heitmann, C.; Jayaraman, D.; Rajasekar, S.; Graham, D.; Maeda, J.; Gibson, D.; Schwartz, K.D. Nitrogen fixation in a landrace of maize is supported by a mucilage-associated diazotrophic microbiota. PLoS biology 2018, 16, e2006352. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, T.; Terakado-Tonooka, J.; Minamisawa, K. Exploration of bacterial N2-fixation systems in association with soil-grown sugarcane, sweet potato, and paddy rice: a review and synthesis. Soil Science and Plant Nutrition 2017, 63, 578–590. [Google Scholar] [CrossRef]
- Ikeda, S.; Sasaki, K.; Okubo, T.; Yamashita, A.; Terasawa, K.; Bao, Z.; Liu, D.; Watanabe, T.; Murase, J.; Asakawa, S. Low nitrogen fertilization adapts rice root microbiome to low nutrient environment by changing biogeochemical functions. Microbes and environments 2014, 29, 50–59. [Google Scholar] [CrossRef]
- Bao, Z.; Watanabe, A.; Sasaki, K.; Okubo, T.; Tokida, T.; Liu, D.; Ikeda, S.; Imaizumi-Anraku, H.; Asakawa, S.; Sato, T. A rice gene for microbial symbiosis, Oryza sativa CCaMK, reduces CH4 flux in a paddy field with low nitrogen input. Applied and environmental microbiology 2014, 80, 1995–2003. [Google Scholar] [CrossRef]
- Bidyarani, N.; Prasanna, R.; Chawla, G.; Babu, S.; Singh, R. Deciphering the factors associated with the colonization of rice plants by cyanobacteria. Journal of Basic Microbiology 2015, 55, 407–419. [Google Scholar] [CrossRef]
- Maeda, I. Potential of phototrophic purple nonsulfur bacteria to fix nitrogen in rice fields. Microorganisms 2021, 10, 28. [Google Scholar] [CrossRef]
- Yamaji, N.; Sasaki, A.; Xia, J.X.; Yokosho, K.; Ma, J.F. A node-based switch for preferential distribution of manganese in rice. Nature Communications 2013, 4, 2442. [Google Scholar] [CrossRef]
- Yamamuro, C.; Ihara, Y.; Wu, X.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. The Plant Cell 2000, 12, 1591–1605. [Google Scholar] [CrossRef] [PubMed]
- Gyaneshwar, P.; James, E.K.; Mathan, N.; Reddy, P.M.; Reinhold-Hurek, B.; Ladha, J.K. Endophytic colonization of rice by a diazotrophic strain of Serratia marcescens. Journal of bacteriology 2001, 183, 2634–2645. [Google Scholar] [CrossRef] [PubMed]
- James, E.K.; Gyaneshwar, P.; Mathan, N.; Barraquio, W.L.; Reddy, P.M.; Iannetta, P.P.; Olivares, F.L.; Ladha, J.K. Infection and colonization of rice seedlings by the plant growth-promoting bacterium Herbaspirillum seropedicae Z67. Molecular Plant-Microbe Interactions 2002, 15, 894–906. [Google Scholar] [CrossRef] [PubMed]
- Mano, H.; Morisaki, H. Endophytic bacteria in the rice plant. Microbes and environments 2008, 23, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Ao, Z.; Xia, J.; Seino, H.; Inaba, K.; Takahashi, Y.; Hayakawa, C.; Hirai, H.; Maeda, I. Adaptations of Potential Nitrogenase Activity and Microbiota with Long-Term Application of Manure Compost to Paddy Soil. Environments 2023, 10, 103. [Google Scholar] [CrossRef]
- Chow, P.S.; Landhäusser, S.M. A method for routine measurements of total sugar and starch content in woody plant tissues. Tree physiology 2004, 24, 1129–1136. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Applied and environmental microbiology 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Poly, F.; Monrozier, L.J.; Bally, R. Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Research in microbiology 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Applied and environmental microbiology 1993, 59, 695–700. [Google Scholar] [CrossRef]
- Teske, A.; Wawer, C.; Muyzer, G.; Ramsing, N.B. Distribution of sulfate-reducing bacteria in a stratified fjord (Mariager Fjord, Denmark) as evaluated by most-probable-number counts and denaturing gradient gel electrophoresis of PCR-amplified ribosomal DNA fragments. Applied and Environmental Microbiology 1996, 62, 1405–1415. [Google Scholar] [CrossRef]
- Chen, T.; Li, G.; Islam, M.R.; Fu, W.; Feng, B.; Tao, L.; Fu, G. Abscisic acid synergizes with sucrose to enhance grain yield and quality of rice by improving the source-sink relationship. BMC plant biology 2019, 19, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-J.; Wang, S.-J. Molecular regulation of sink–source transition in rice leaf sheaths during the heading period. Acta Physiologiae Plantarum 2008, 30, 639–649. [Google Scholar] [CrossRef]
- Zhang, G.; Cui, K.; Li, G.; Pan, J.; Huang, J.; Peng, S. Stem small vascular bundles have greater accumulation and translocation of non-structural carbohydrates than large vascular bundles in rice. Physiologia Plantarum 2022, 174, e13695. [Google Scholar] [CrossRef] [PubMed]
- Slewinski, T.L. Non-structural carbohydrate partitioning in grass stems: a target to increase yield stability, stress tolerance, and biofuel production. Journal of Experimental Botany 2012, 63, 4647–4670. [Google Scholar] [CrossRef]
- Okamoto, T.; Shinjo, R.; Nishihara, A.; Uesaka, K.; Tanaka, A.; Sugiura, D.; Kondo, M. Genotypic variation of endophytic nitrogen-fixing activity and bacterial flora in rice stem based on sugar content. Frontiers in Plant Science 2021, 1610. [Google Scholar] [CrossRef]
- Che, J.; Yamaji, N.; Ma, J.F. Role of a vacuolar iron transporter OsVIT2 in the distribution of iron to rice grains. New Phytologist 2021, 230, 1049–1062. [Google Scholar] [CrossRef]
- Mu, S.; Yamaji, N.; Sasaki, A.; Luo, L.; Du, B.; Che, J.; Shi, H.; Zhao, H.; Huang, S.; Deng, F. A transporter for delivering zinc to the developing tiller bud and panicle in rice. The Plant Journal 2021, 105, 786–799. [Google Scholar] [CrossRef]
- Wang, W.; Zhai, Y.; Cao, L.; Tan, H.; Zhang, R. Endophytic bacterial and fungal microbiota in sprouts, roots and stems of rice (Oryza sativa L.). Microbiological Research 2016, 188, 1–8. [Google Scholar] [CrossRef]
- Knelman, J.E.; Legg, T.M.; O’Neill, S.P.; Washenberger, C.L.; González, A.; Cleveland, C.C.; Nemergut, D.R. Bacterial community structure and function change in association with colonizer plants during early primary succession in a glacier forefield. Soil Biology and Biochemistry 2012, 46, 172–180. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, M.; Chen, A.; Zhang, W.; Wei, W.; Sheng, R. Impact of fertilization regimes on diazotroph community compositions and N2-fixation activity in paddy soil. Agriculture, Ecosystems & Environment 2017, 247, 1–8. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, J.; Li, Y.; Chen, D.; Ao, J.; Zhou, W.; Shen, D.; Li, Q.; Huang, Z.; Jiang, Y. Influence of nitrogen and phosphorus additions on N2-fixation activity, abundance, and composition of diazotrophic communities in a Chinese fir plantation. Science of the Total Environment 2018, 619, 1530–1537. [Google Scholar] [CrossRef]
- Ramšak, A.; Peterka, M.; Tajima, K.; Martin, J.C.; Wood, J.; Johnston, M.E.; Aminov, R.I.; Flint, H.J.; Avguštin, G. Unravelling the genetic diversity of ruminal bacteria belonging to the CFB phylum. FEMS Microbiology Ecology 2000, 33, 69–79. [Google Scholar] [CrossRef]
- Sun, L.; Qiu, F.; Zhang, X.; Dai, X.; Dong, X.; Song, W. Endophytic bacterial diversity in rice (Oryza sativa L.) roots estimated by 16S rDNA sequence analysis. Microbial ecology 2008, 55, 415–424. [Google Scholar] [CrossRef]
- Yasuda, M.; Dastogeer, K.M.; Sarkodee-Addo, E.; Tokiwa, C.; Isawa, T.; Shinozaki, S.; Okazaki, S. Impact of Azospirillum sp. B510 on the rhizosphere microbiome of rice under field conditions. Agronomy 2022, 12, 1367. [Google Scholar] [CrossRef]
- Choudhury, A.; Kennedy, I. Prospects and potentials for systems of biological nitrogen fixation in sustainable rice production. Biology and Fertility of Soils 2004, 39, 219–227. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, J.; Cao, Y.; Sheirdil, R.; Wang, X.; Zhang, L. Isolation and proposal novel rice promoting endophytic bacteria, Rhizobium oryzicola sp. nov. Int J Syst Evol Microbiol 2015, 65, 2931–2936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, L.; Ma, X.; Sui, X.H.; Jiang, R. Rhizobium pseudoryzae sp. nov., isolated from the rhizosphere of rice. International journal of systematic and evolutionary microbiology 2011, 61, 2425–2429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-X.; Tang, X.; Sheirdil, R.A.; Sun, L.; Ma, X.-T. Rhizobium rhizoryzae sp. nov., isolated from rice roots. International Journal of Systematic and Evolutionary Microbiology 2014, 64, 1373–1377. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, X.; Wang, J.; Gong, Q.; Zhang, X.; Zhang, G. Isolation, identification and characterization of endophytic bacterium Rhizobium oryzihabitans sp. nov., from rice root with biotechnological potential in agriculture. Microorganisms 2020, 8, 608. [Google Scholar] [CrossRef]
- Zhao, J.-J.; Zhang, J.; Sun, L.; Zhang, R.-J.; Zhang, C.-W.; Yin, H.-Q.; Zhang, X.-X. Rhizobium oryziradicis sp. nov., isolated from rice roots. International journal of systematic and evolutionary microbiology 2017, 67, 963–968. [Google Scholar] [CrossRef]
- Wegner, C.E.; Liesack, W. Microbial community dynamics during the early stages of plant polymer breakdown in paddy soil. Environmental microbiology 2016, 18, 2825–2842. [Google Scholar] [CrossRef]
- Bao, Z.; Sasaki, K.; Okubo, T.; Ikeda, S.; Anda, M.; Hanzawa, E.; Kakizaki, K.; Sato, T.; Mitsui, H.; Minamisawa, K. Impact of Azospirillum sp. B510 inoculation on rice-associated bacterial communities in a paddy field. Microbes and environments 2013, 28, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Lianfeng, W.; Zhongjun, J. Heterotrophy-coordinated diazotrophy is associated with significant changes of rare taxa in soil microbiome. Pedosphere 2022, 32, 402–413. [Google Scholar] [CrossRef]
- Li, G.; Hu, Q.; Shi, Y.; Cui, K.; Nie, L.; Huang, J.; Peng, S. Low nitrogen application enhances starch-metabolizing enzyme activity and improves accumulation and translocation of non-structural carbohydrates in rice stems. Frontiers in plant science 2018, 9, 1128. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The rice parts collected at panicle initiation. Pictures of IN (A), BN (B), and CR (C) were shown.
Figure 1.
The rice parts collected at panicle initiation. Pictures of IN (A), BN (B), and CR (C) were shown.

Figure 2.
Changes of nitrogenase activity in the rice parts after seedling transplantation. IN (triangle), BN (circle), and CR (square) of rice cultured in MC-applied (red) and CF-applied (blue) plots were sampled. A plot with an error bar indicates of a mean of measurements using 3 different stocks ± SD.
Figure 2.
Changes of nitrogenase activity in the rice parts after seedling transplantation. IN (triangle), BN (circle), and CR (square) of rice cultured in MC-applied (red) and CF-applied (blue) plots were sampled. A plot with an error bar indicates of a mean of measurements using 3 different stocks ± SD.
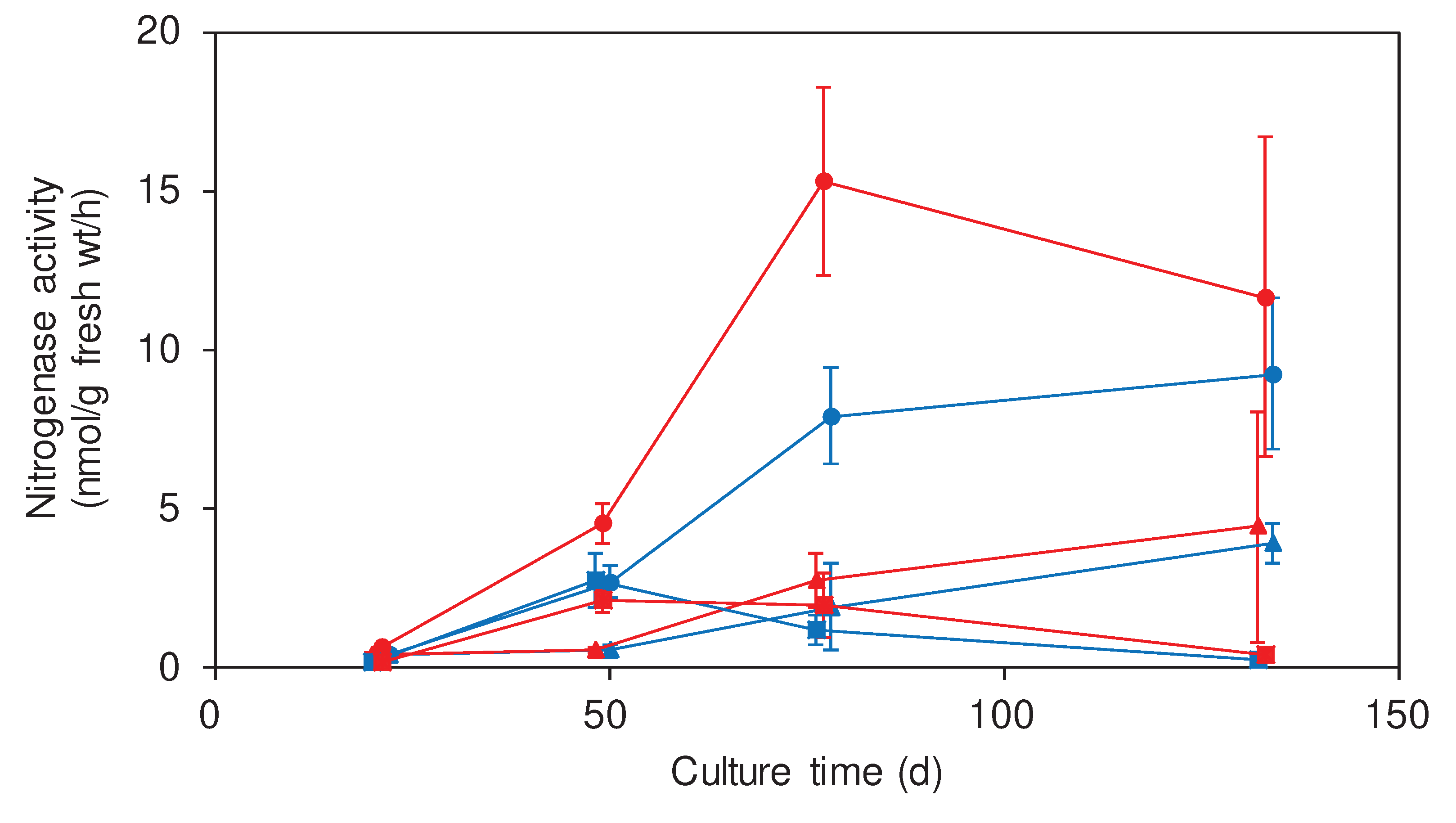
Figure 3.
Seasonal changes of non-structural carbohydrate contents in the rice parts after seedling transplantation. Sucrose (A) and glucose (B) contents in IN (triangle), BN (circle), and CR (square) were measured in rice cultured in MC-applied (red) and CF-applied (blue) plots and sampled after the days of seedling transplantation. A plot with an error bar indicates a mean of measurements using 3 different stocks ± SD.
Figure 3.
Seasonal changes of non-structural carbohydrate contents in the rice parts after seedling transplantation. Sucrose (A) and glucose (B) contents in IN (triangle), BN (circle), and CR (square) were measured in rice cultured in MC-applied (red) and CF-applied (blue) plots and sampled after the days of seedling transplantation. A plot with an error bar indicates a mean of measurements using 3 different stocks ± SD.
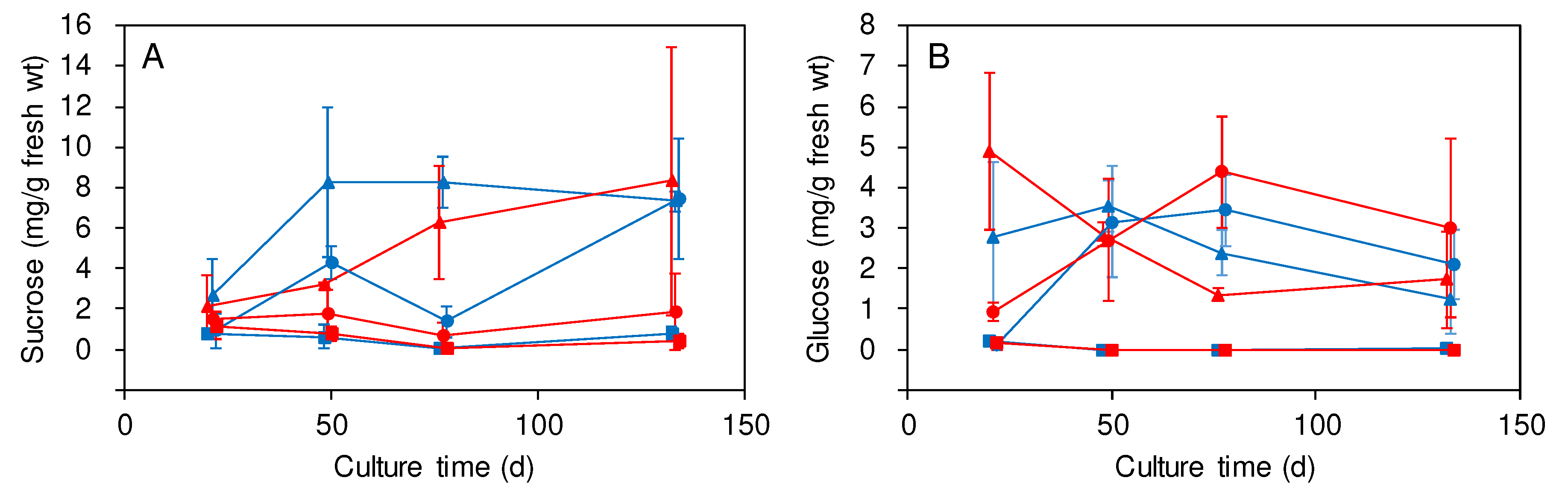
Figure 4.
Abundances of bacteria and nifH genes in the rice parts at panicle initiation. Bacterial read/chloroplastic read (A) was calculated based on the 16S rRNA gene amplicon sequence. nifH/bacterial 16S rRNA gene (B) was calculated based on Ct values of the quantitative PCR. A column with an error bar indicates a mean of measurements using 3 different stocks ± SD.
Figure 4.
Abundances of bacteria and nifH genes in the rice parts at panicle initiation. Bacterial read/chloroplastic read (A) was calculated based on the 16S rRNA gene amplicon sequence. nifH/bacterial 16S rRNA gene (B) was calculated based on Ct values of the quantitative PCR. A column with an error bar indicates a mean of measurements using 3 different stocks ± SD.
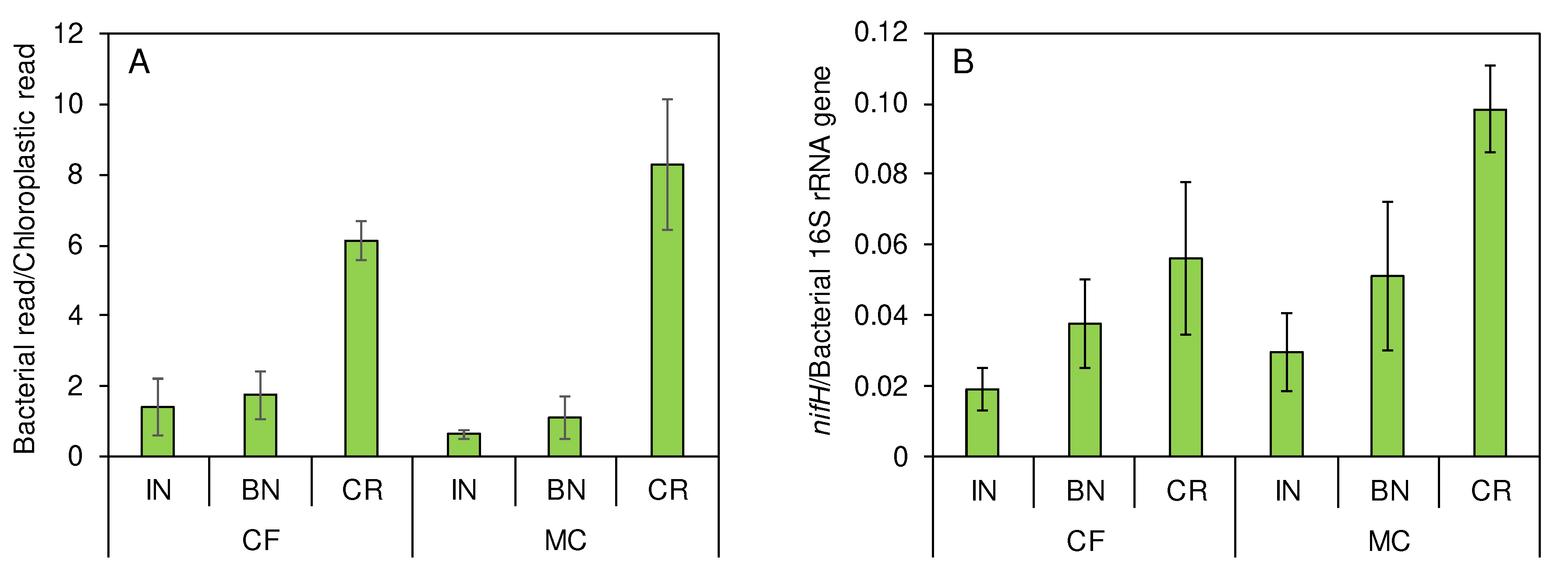
Figure 5.
Rarefaction curves of OTUs observed in the bacterial microbiota of the rice parts at panicle initiation. Curves for IN (green), BN (blue), and CR (red) of rice cultured in MC-applied (A) and CF-applied (B) plots.
Figure 5.
Rarefaction curves of OTUs observed in the bacterial microbiota of the rice parts at panicle initiation. Curves for IN (green), BN (blue), and CR (red) of rice cultured in MC-applied (A) and CF-applied (B) plots.
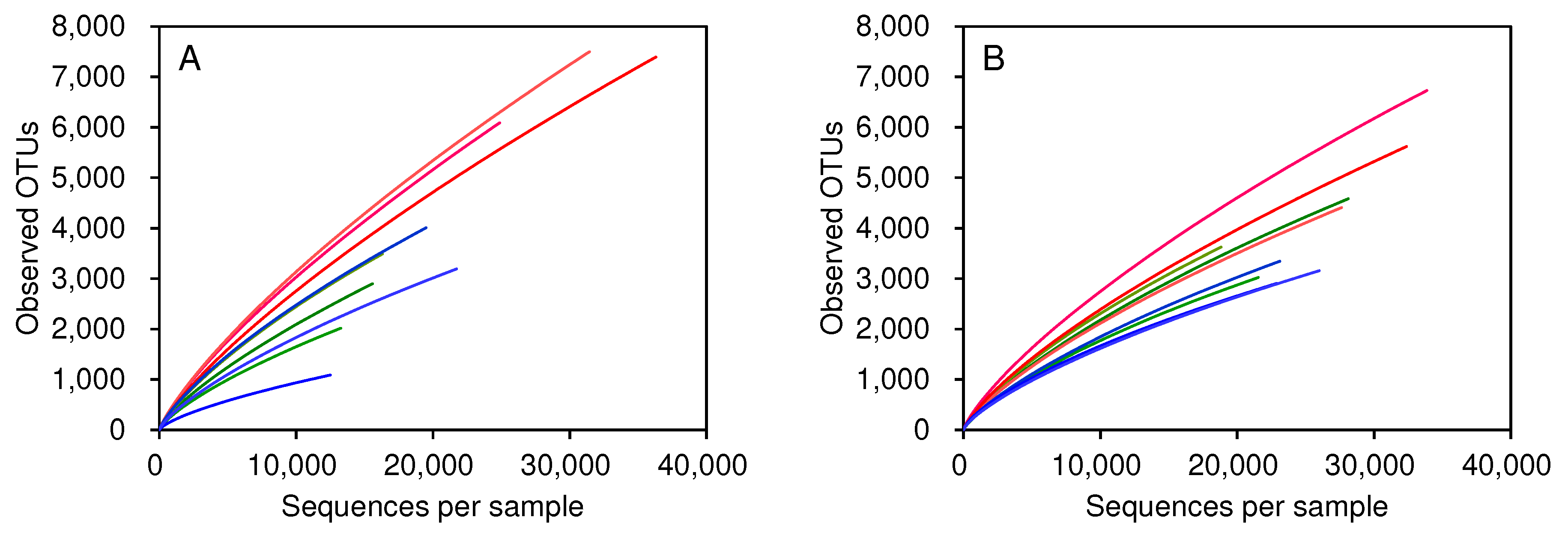
Figure 6.
PCoA of the bacterial microbiota in the rice parts at panicle initiation. Coordinate points for IN (green), BN (blue), and CR (red) of rice cultured in MC-applied (closed) and CF-applied (open) plots. The dotted drawings show 95% confidence ellipses for IN and BN (blue) and CR (red).
Figure 6.
PCoA of the bacterial microbiota in the rice parts at panicle initiation. Coordinate points for IN (green), BN (blue), and CR (red) of rice cultured in MC-applied (closed) and CF-applied (open) plots. The dotted drawings show 95% confidence ellipses for IN and BN (blue) and CR (red).
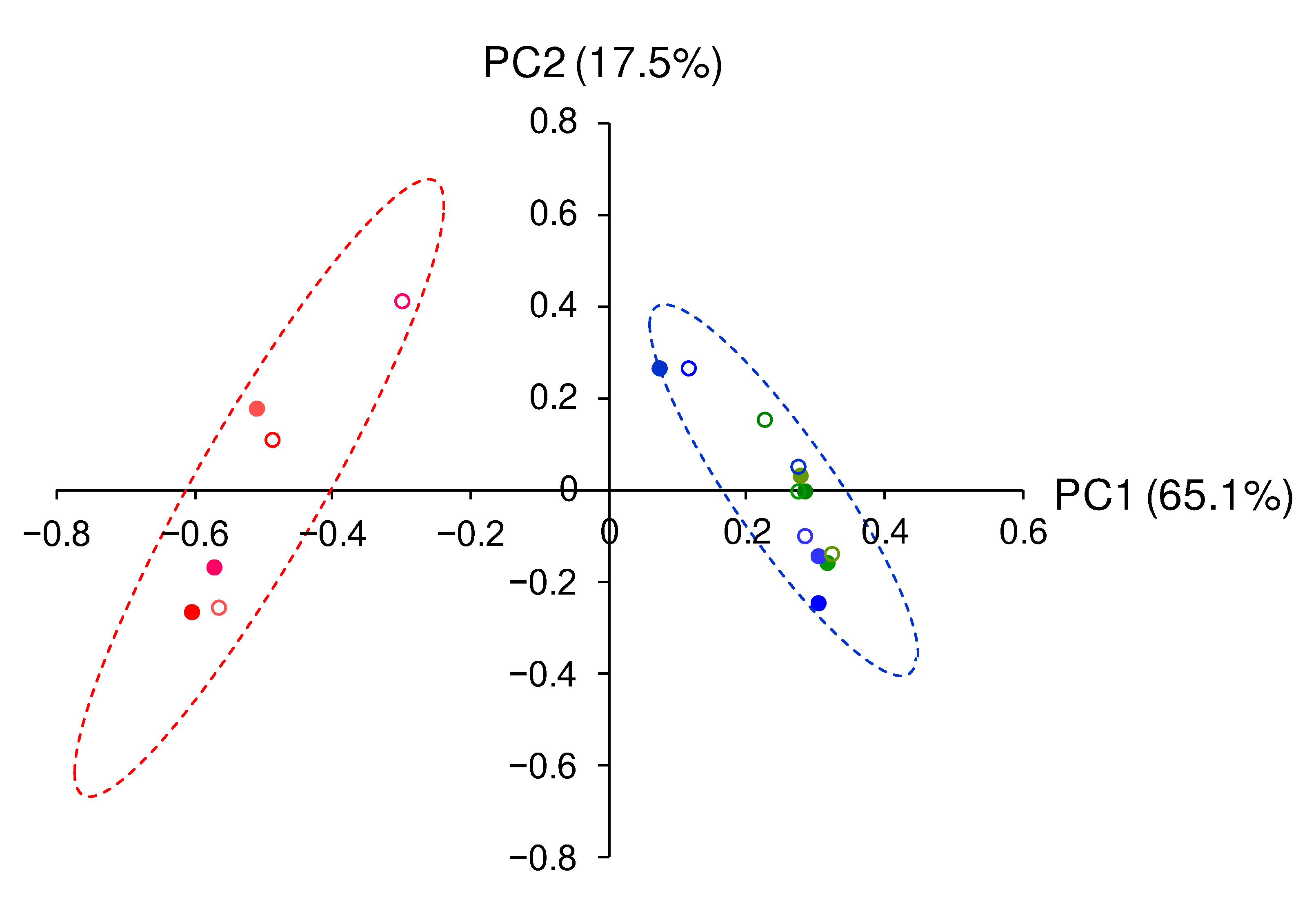
Figure 7.
Class-level taxonomic profiles in the rice parts at panicle initiation. A column with an error bar indicates a mean of measurements using data obtained from 3 different stocks + SD.
Figure 7.
Class-level taxonomic profiles in the rice parts at panicle initiation. A column with an error bar indicates a mean of measurements using data obtained from 3 different stocks + SD.
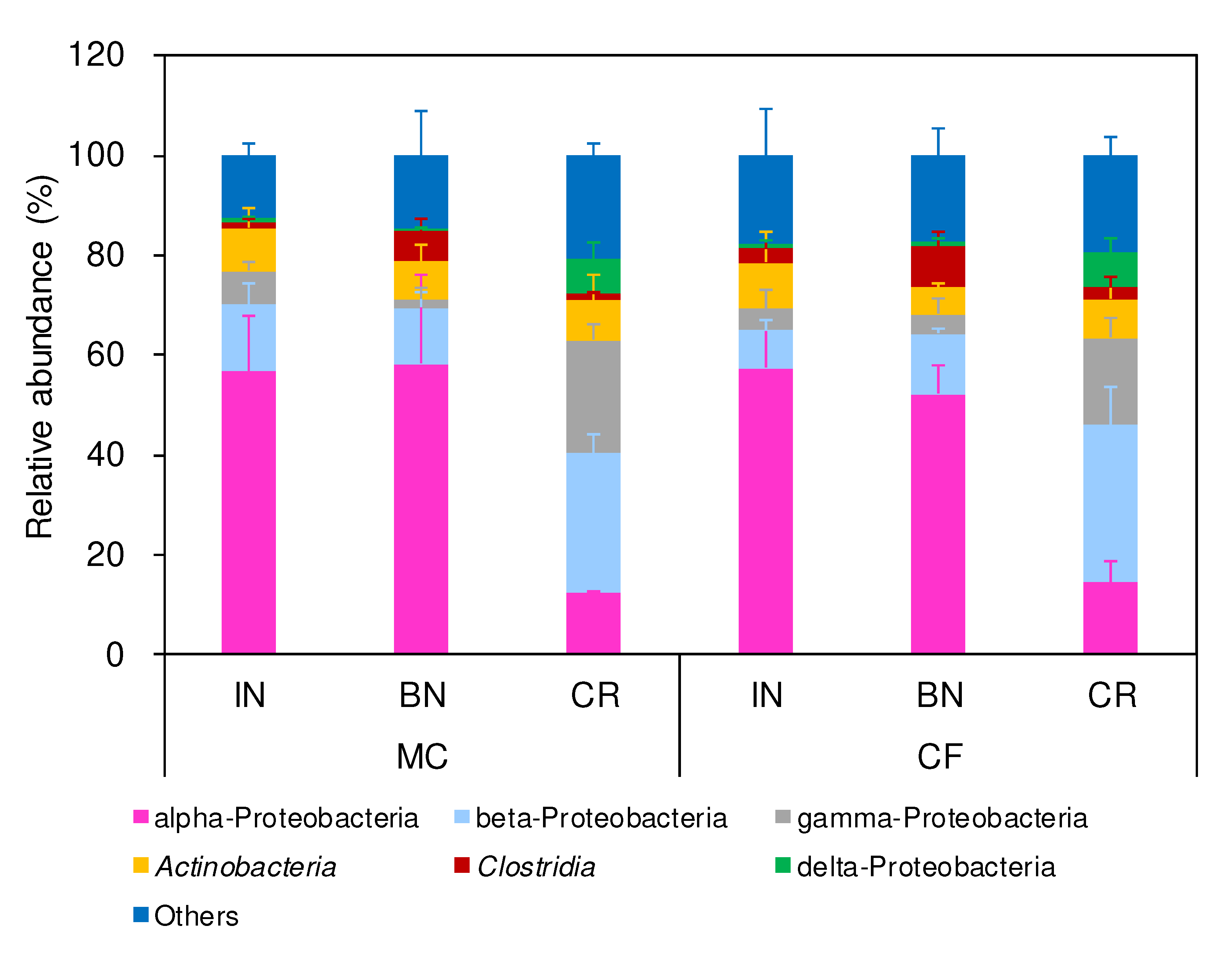

Table 1.
Shannon and inverse Simpson indices of the bacterial microbiota in the rice parts at panicle initiation. A mean of measurements using data obtained from 3 different stocks ± SD was shown.
Table 1.
Shannon and inverse Simpson indices of the bacterial microbiota in the rice parts at panicle initiation. A mean of measurements using data obtained from 3 different stocks ± SD was shown.
| Material applied | Rice part | Shannon | Inverse Simpson |
|---|---|---|---|
| MC | IN BN CR |
5.39 ± 0.63 4.87 ± 1.34 6.46 ± 0.28 |
14.5 ± 6.2 15.6 ± 15.6 33.1 ± 12.1 |
| CF | IN BN CR |
5.45 ± 0.42 5.35 ± 0.33 6.07 ± 0.55 |
18.1 ± 10.9 23.7 ± 14.0 43.4 ± 23.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



