
Submitted:
31 July 2023
Posted:
02 August 2023
You are already at the latest version
Abstract
Background: One of the main diseases of worldwide concern is non–insulin dependent Diabetes mellitus. 90-95% of patients with diabetes suffers from Type II diabetes, a metabolic diabetes whose prevalence is increasing in the world.Given the current prevalence of diabetes type II in society, research of natural compounds able to regulate glycaemia and insulin–resistance is nowadays covering importance, as alternative or co–adjuvant treatment to existing hypoglycaemic medicines. In pre–clinical steps of research, C. elegans is considered a promising in vivo model for the study of the molecular mechanism of glucose as part of anti–diabetic bioactivity.Most of the studies of insulin resistance with this in vivo model are carried out by using mutant strains such as daf-2 and the effects of the compounds are attributed a posteriori, after a physiological response of the worms rather than directly measuring preservation of activity inside the worm.
Methods: The in vitro α-glucosidase inhibition of two oral formulations with Origanum vulgare extract before and after a simulated process of gastrointestinal digestion was evaluated. After confirming the in vitro activity, the α-glucosidase inhibition was tested in vivo through the new method proposed.
Results: The crude extract showed an IC50 value similar to acarbose (positive control), before and after the gastrointestinal digestion. The encapsulated extract (111.86±1.25 µg/mL) did not present statistical differences with the positive control (110.10±1.15 µg/mL) after digestion. However, the powder formulation showed the highest hypoglycaemic activity (59.55±0.85 µg/mL).In vivo, the IC50 values showed similar profile than in vitro.
Conclusion: A new simple and reliable method has been developed to determine pharmacological activities inside in vivo model C. elegans.The method has been evaluated with a hydroalcoholic extract of O. vulgare. The method is accurate and valid for the quantification of the inhibition of in vivo, allowing the direct attribution of this in vivo hypoglycaemic activity to the treatment. In addition, it opens the door to further research into in vitro pharmacological activities that require an in vivo counterpart.
Keywords:
C. elegans
; hyperglycemia
; O. vulgare
; in vitro digestion
1. Introduction
One of the main diseases of worldwide concern is non–insulin dependent Diabetes mellitus (NIDDM), caused by an insulin resistance or a decreased secretion by Langerhans β cells of this hypoglycemic hormone.
After ingestion, sugars and carbohydrates are digested and reduced to glucose, which is absorbed in the intestine. This increase of glycaemia (glucose levels in blood) promotes the release of hormone insulin in the pancreas that lowers glucose levels. The β–cells in pancreas secrete insulin so that glucose can enter into cells as a cell nutrient. In addition, insulin stimulates the formation of glycogen in the liver and thus blood glucose to normal values. The antagonistic hormone produced by the endocrine side of the pancreas, glucagon, stimulates glycogen degradation when there is a lack of glucose intake and glucose is needed. This is a simplification of endocrine feedback to control glycaemia and body energy balance. However, as occurs with the antioxidant reactionsin the body, these can also be altered giving rise a worldwide–extended disease: Diabetes. There are three main types of diabetes, depending on the cause of the problem [1].
The first type of diabetes (Type I diabetes), whose cause is autoimmune since the organism itself destroys the β–cells of the pancreas,concerns5–10% of all diabetic patients.[2]. For being a non-curable disease, patients need to inject insulin daily as treatment because this function of the pancreas is partially or totally disabled. Although engineering and pharmaceuticals have played such a helpful role with diabetics (insulin pumps, different types of insulin depending on the speed of action, mobile applications with glycaemia predictions…), science continues to search for a cure in theories such as the microencapsulation of β–cells, transplantation or genetic theories.
On the other side, 90-95% of patients with diabetes suffers from Type II diabetes, a metabolic diabetes whose prevalence is increasing in the world [2]. Generally, these patients are adults whose risk factors (overweight, sedentary lifestyle, non–healthy diet or family history) lead their bodies to insulin resistance.This disease can be sufferedsilently and asymptomatically without medical diagnosis.Therefore, glycaemic monitoring is important to keep resistance under control with hygienic–dietary changes in lifestyle. As prevention, glycaemic can be periodically monitored in pharmacies or MD’s and takes theFindrisk test (questioning the risk of suffering type II diabetes in 10 years). A low score in this test does not mean a lack of pre–diabetes or diabetes. Treatment begins with daily oral medicines that affect different pathways (DPP–4, GLUT–1, α–glycosidase…). If glycaemia and HbA1c (glycosylated haemoglobin that reflects mean glycaemia within last 2–3 months) values remain under control, monotherapy is sufficient. However, when the patient presents glycaemic alterations –reflected by a high HbA1c value (>7%) – more than one type of hypoglycaemic treatment is needed and sometimes insulin is part of this treatment.
Last but not least, there is a third type of diabetes (Gestational diabetes) that can appear during pregnancy. Both mother and child may develop diabetes if the glycaemia is not controlled during pregnancy.
For all the three types of diabetes, patient education and selection of the optimal treatment for them are important. When the disease is not treatedwell, blood glucose is so high that it can cause tissue damage due to overproduction of superoxide in cells, leading to potentially fatal complications such as retinopathies, limb amputations, neuropathies, renal and cardiovascular (CVD) pathologies or even premature death.
Most of the current preclinical research with in vivo model focus on the first type of diabetes, since the profile is a young normal-weighted patient and the cause is autoimmune and cannot be prevented. Nevertheless, could there be an effective natural alternative to prevent and treat Type II diabetes whose prevalence is 530 million adults worldwide[3] and now also occurring increasingly frequently in children[2]?
Among antidiabetic treatments, inhibition of the α–glucosidase pathway has been shown to be effective in delaying intestinal absorption of polysaccharides and disaccharides. This enzyme, located in the membrane–bound epithelium of the small intestine, is responsible for the cleavage of glucose from ingested disaccharides.
Despite the antidiabetic efficacy of acarbose (lowering HbA1c), digestive problems such as flatulence (probably affecting more than 1 in 10 people), stomach pain and diarrhoea (probably affecting fewer than 1 in 10 people) are the main side–effects that limit it success in prescribing[4]. In addition, acarbose causes rare side effects (likely to affect less than 1 in 100 people): nausea, vomiting, increasing liver enzymes and indigestion [5].
Given the current prevalence of diabetes type II in society [1], research of natural compounds able to regulate glycaemia and insulin–resistance is nowadays covering importance in industry, as alternative or co–adjuvant treatment to existing hypoglycaemic medicines.Many plants with α–glucosidaseinhibitory activity have been shown to generate less side effects [6,7,8,9,10].
In pre–clinical steps of research, C. elegans is considered a promising in vivo model for the study of the molecular mechanism of glucose as part of anti–diabetic bioactivity [11]. The strains are cheap to breed and can be frozen, allowing for long–term storage. Moreover, thanks to be the first multicellular organism with a complete genome sequence [12], mutations and molecular identification of many key genes can be studied to identify behavioural defects. Indeed, at least 38 % of the protein–coding genes in C. elegans have predicted orthologs in the human genome [13], 60 – 80 % of human genes have predicted orthologs in the C. elegans genome [14], and 40 % of genes known to be associated with human diseases have clear orthologs in the C. elegans genome [15].
Most of the studies of insulin resistance with this in vivo model are carried out by using mutant strains such as daf-2, an insulin-like receptor gene, to understand the antidiabetic behaviour of a compound. However, the effects of the compounds are attributed a posteriori [11,16,17,18], after a physiological response of the worms rather than directly measuring preservation of activity inside the worm. In this context, there is a need on finding a simple and reliable method with this in vivo model in order to quantify directly pharmacological activity of the compound given as treatment for Type II diabetes.
In previous study, hydroalcoholic extract of Origanumvulgareshowed significant antioxidant activity both in vitro and in vivo [19]. According to literature, this medicinal plant count with many different pharmacological activities that might be significant in metabolic age-related pathologies of current interest in society such as Diabetes type II [20,21,22]. Therefore, themain aim of this work isto find a new method able to easily determine hypoglycaemic activity in vivo model and so, certainly attribute the response to the effect of the treatment.
In order to achieve that, the study starts evaluating the in vitro hypoglycemic activity of two oral formulations with Origanumvulgare extract before and after a simulated process of gastrointestinal digestion. After confirming the in vitro activity, the hypoglycemic activity is testedin vivo through the new method proposed. Results would determine the robustness of the method and if there is an in vitro-in vivo co-relation.
2. Results and Discussion
Diabetes mellitus is a chronic disease that, if left untreated, leads to serious and disabling complications that reduce the quality of life of those affected and increase the cost of care. It is the most common endocrine disease and its prevalence is steadily increasing worldwide. Of the different types of diabetes, type II is the most common. It is characterised by a gradual course in which changes in the mass and function of pancreatic beta cells precede a decrease in the responsiveness of peripheral tissues to insulin. These patients do not initially require insulin therapy, and glycaemic control can be achieved through weight loss, physical activity, or oral hypoglycemic agents. Oral hypoglycemic agents such as biguanides, thiazolidinediones, and sulfonylureas can be effective in controlling hyperglycemia. However, they have significant side effects, including hypoglycemia and gastrointestinal problems [23].
There is an urgent need for effective substitutes to reduce diabetes complications with fewer side effects. In recent years, the search for alternative medicines for the treatment of diabetes has attracted considerable attention. Natural products play a key role in this effort. There are more than 400 traditional medicinal plants known to have antidiabetic properties from their bioactive compounds. However, only a few of them have been sufficiently studied scientifically and medically to prove their efficacy [24].
Naturally, occurring polysaccharides are primary metabolites found in all plants and are therefore essential for their development. These compounds, usually derived from medicinal plants, grains, fruits, vegetables, edible fungi and foods, have been extensively studied in recent years due to their low toxicity and numerous pharmacological activities, including antidiabetic activity [25,26]. Several studies have shown that purified polysaccharides from pumpkin, sea cucumber, goji berry, mushrooms, green beans, tea, and oats have beneficial effects on glucose homeostasis, reduce diabetic complications through the defence mechanism against oxidative stress damage, and ultimately improve insulin sensitivity [27,28,29]. Carbohydrates are ingested in the form of polysaccharides and are converted in the gastrointestinal tract by the digestive enzymes α-amylase (salivary or pancreatic) and α-glucosidase (small intestinal) into monosaccharides, which are readily absorbed by the small intestine and enter the bloodstream. Thus, inhibition of these enzymes prevents this conversion and is an effective step in controlling blood glucose levels in diabetics. Carbohydrates that are not digested in the small intestine are metabolised in the large intestine by intestinal bacteria, which explains the frequent side effects of these drugs (meteorism, flatulence, diarrhoea, etc.).
Therefore, the search for extracts/compounds that inhibit these enzymes could be an effective strategy to combat this pathology. In this work, the inhibitory activity of the enzyme α-glucosidase was tested two formulations proposed for a hydroalcoholic extract of O. vulgare. Matsui et al. [30] describe a method to spectrophotometrically determine the inhibition of this enzyme by a substrate that decomposes into a yellow compound and glucose after exposure to α-glucosidase. Acarbose, one of the drugs marketed for oral treatment of noninsulin-dependent diabetes (type II diabetes), was used as a positive control in the analysis because it impairs intestinal absorption of carbohydrates and reduces postprandial hyperglycemia by competitively inhibiting the action of intestinal α-glucosidase, which hydrolyses oligosaccharides and polysaccharides. Rodriguez-Solana et al. [31] also used this compound (1 mg/ml) as a positive control and showed 78.34% inhibition of the enzyme α-glucosidase and 78.33% inhibition for α-amylase. As shown in Figure 1, in this current study, the same positive control at 0.5 mg/mL showed a percentage of inhibition of 76.03±2.08% before digestion and 78.42±2.69% after the gastrointestinal digestion process. In vitro was OV-E as effective as acarbose, a marketed oral anti-diabetic drug used as a positive control.
In general, the inhibition of the enzyme was enhanced with the intestinal fraction, what suggests that not only might the activity be maintained in the intestine – where the enzyme is physiological located -, but also improved. The crude extract (OV-E) before the gastrointestinal digestion process showed an IC50 value similar to acarbose (positive control), 149.40±3.36 and 141.47±3.57 µg/mL, respectively with p>0.05 (Figure 1).According to the results, two formulations after the digestion step presented higher bioactivity than the extract before digestion (OV-E). Neither the activity of the excipients nor the digestion enzymes can be related to this enhance of activity since the Blank did not present any inhibitory activity (being highest 0.16±0.03%, with p<0.05 – Figure 1). In the case of the capsules, even if the hard gelatine capsules were completely disaggregated, the intestinal fraction of the empty capsules, used as control, did not present any potentially inhibitory activity, being IC50=201.51±1.12 µg/mL, with p<0.001 for the rest of results.
Once dismissed possible secondary pharmacological activities from the pharmaceutical forms, the encapsulated extract (111.86±1.25 µg/mL) did not present statistical differences with the intestinal fraction of the positive control (110.10±1.15 µg/mL) with p=0.282. Thus, this dosage form did not show the same improvement in the pharmacological activity, but the efficacy was similar to acarbose after digestion. However, the extract in powder showed the highest hypoglycaemic because the IC50 value was the lowest (59.55±0.85 µg/mL) with p<0.05.
In a previous study with flavonoid fisetin from leaves of Rhus succedanea L. [32], which was more active than acarbose, encapsulation in nanoparticles did not improved this inhibitory activity against α–glucosidase. Using the computational molecular docking approach, the authors explained their results in relation to the structural determinants involved on the interaction between flavonoids and the enzyme, considering that the 3’,4’–dihydroxyl groups of B ring in flavonoids are crucial in engaging direct binding with the active–site residues. Since the flavonoid content was higher in the intestinal fraction of the capsule than in OV–P, our hypothesis relays on the fact that in OV–P, the binding site of flavonoids might be uncluttered and the chemical compounds are already completely released, comparing to the physical protection of the capsule as a formulation container. Thus, the extract given as powder clearly showed to enhance hypoglycaemic activity in vitro.
Once the intestinal absorbable fractions have been shown to inhibit the enzyme in the site of action in vitro, there is the doubt whether they will be maintaining the activity inside a living organism. Thus, the inhibitory activity of the pharmaceutical forms – and the positive control – after digestion was determined inside C. elegans. The new method proposed was designed in order to quantify directly the activity of the samples inside this in vivo model, instead of attributing physiological effects a posteriori. Figure 2 recompiles the inhibitory percentages of OV-C, OV-P and Acarbose-Int in vivo as well as the calculated value of IC50.
Results for in vivo pharmacological activity were again expressed as percentage of inhibition.At first glance, the extract encapsulated (OV-C) presented a similar behaviour as positive control (Acarbose-Int), as in vitro. Nevertheless, OVE given as powder showed to inhibit significantly higher the enzyme than the positive control at any of the concentrations, being at 50 mg/mL 52.98±0.99% against 31.66±0.63%, respectively with p<0.05. In this sense, the samples showed similar patron of activity among them than in the experiment performed in vitro.
Whereas in vitro tests are used to demonstrate the intrinsic activity of compounds, in vivo tests focus on the physiological effects and provide a second and essential line of evidence for antioxidant activity. Although sometimes is there no direct correlation between in vitro and in vivo results, our results showed a scale of 1.30 times on the values (Figure 3).
This mathematical relation can still only be attributed to this case and should not be settled as universal until more data from different plants is recompiled. This new method set up provides a fast track to test treatments for diabetes in vivo without involving genes or any other costly material. Results confirm the robustness of the method. The assay was replicated three times from the start using independent worm stocks to validate the method. The reproducibility was tested by carrying out a minimum of 12 determinations for each compound concentration per repetition of the experiment under the same conditions. In contrast, our detectable results (more than 1% inhibition) had a C.V. of 1.87-6.72%, which is a good indication of the repeatability of the method, since it is less than 20%. For this reason, our method is accurate and valid for the quantification of the inhibition of in vivo, allowing the direct attribution of this in vivo hypoglycaemic activity to the treatment. In addition, it opens the door to further research into in vitro pharmacological activities that require an in vivo counterpart.
3. Material and Methods
3.1. Preparation of the extract and formulations
The two pharmaceutical forms used for this study were prepared as previously on de Torre et al. [33]. Briefly, the hydroalcoholic extract (OV-E) was prepared from O. vulgare flowered aerial-dried parts by cold maceration with ethanol-water 50% v/v. Then, two simple solid oral pharmaceutical forms for this extract (500 mg with 30% of rosmarinic acid) were designed, divided powder and hard gelatine capsules.
3.2. In vitro gastrointestinal digestion process
Thein vitrogastrointestinal digestion process was performed according to Gayoso et al. [34] with some modifications previously described by de Torre et al. [33]. Each pharmaceutical form (equivalent a 500 mg of extract) weresubmitted into the gastrointestinal digestion process obtaining the lyophilised intestinal absorbable fraction, called for each sample: intestinal absorbable fraction of the powder (OV-P), intestinal absorbable fraction of the capsules (OV-C) and intestinal absorbable fraction of the positive control (Acarbose-Int). Finally, thefractions were lyophilised (Cryodos50, Telstar, Barcelona, Spain). A control with two empty capsules and a blank with the excipients and without plant extract were treated under the same conditions.
3.3. In vitro α–glucosidase activity assay
According to Matsui[30]and Shuyuan et al. [10], the inhibitory activity of α–glucosidase can be measured thanks to an analogous disaccharide (4–Nitrophenyl–α–D–glucopyranoside –p–NPG–, used as substrate). The enzyme breaks down the α-chemical bond of the substrate, so that carbohydrate D–Glucopyranoside and the phenolic compound 4–Nitrophenylare released. 4–Nitrophenyl turns yellow at pH 6.8, making it quantifiable at 405 nm (Figure 4).
The more inhibitory activity the extract has, the less yellow the solution will have (measurable, lower absorbance value at 405 nm). To enhance colour, a basic solution must be added (optimal efficient medium pH 6.8) [35] so that there are enough electrons for the p–nitrophenolate anion to make resonance and stabilize [36]. This protocol was established in the laboratory.
The enzyme α–glucosidase needs to be stored at –20 ºC, and kept cold during handling. The enzyme was obtained from Saccharomyces cerevisae (#G0660, Sigma–Aldrich Co., St. Louis, MO) and the optimal pH is 6.8, the same as the intestinal one [37]. The enzyme was resuspended in 0.1 M PBS buffer, pH 6.8, at a concentration of 0.5 U/L –within the range suggested 0.2 U/L–1U/L [38].
As the enzyme, substrate (4–Nitrophenyl–α–D–glucopyranoside– #N1377, Sigma–Aldrich Co., St. Louis, MO) must be stored at –20 ºC and kept on ice while handling. After several titration tests performed, the optimal substrate concentration was 1 mM in 0.1 M PBS buffer and pH 6.8.
It is necessary a solution to stop the reaction and measure the inhibitory activity avoiding biodegradation. 80 µL of 0.15 M Na2CO3 was added toeach well after the reaction occurred.
The different lyophilised sampleswere diluted in 0.1 M PBS buffer, pH 6.8 at six serial concentrations (1,000–0.03125 µg/mL). Acarbose (#A8980,Sigma–Aldrich Co., St. Louis, MO) was used as the positive control at the same concentrations and the sample solvent (PBS) was used for the negative control. Finally, the intestinal digestion fractions of OV1 were also analysed, as well as the intestinal fractions of the corresponding positive control (Acarbose–Int at the same concentrations).
Every plate needs a positive control (120 µL Acarbose) and a negative control (120 µL PBS) with the corresponding blanks. 20 µL PBS was used for blanks instead of enzyme (20 µL 0.5 U/L in PBS) and the volume of substrate was constant for each condition (20 µL).Sample, acarbose, PBS and enzyme were added to the 96–well plate and incubated for 15 min at 37 ºC with shaking.Then, substrate was added according to the design. It was incubated under the same circumstances for an additional 15 min. After the incubation time, 80 µL of the stop solution were added to each sample.
The absorbance measurement was made with the Power WaVe XS de BioTek® spectrophotometer at 405nm.The results were processed with KC Junior BioTek data analysis software. Inhibition (%) of α-glucosidase was calculated by using the following equation:
where, Inhibitory (%, percentage of inhibition) is obtained from the absorbance of the sample (AbsS) without its blank (AbsSB: Sample Blank Absorbance) against the absorbance of the negative control (AbsC) without its blank (AbsCB: Negative Control Blank Absorbance).
The inhibitory concentration (IC50) was calculated by GraphPad Prism v 4.00 analysis.
3.4. In vivo α–glucosidase activity assay
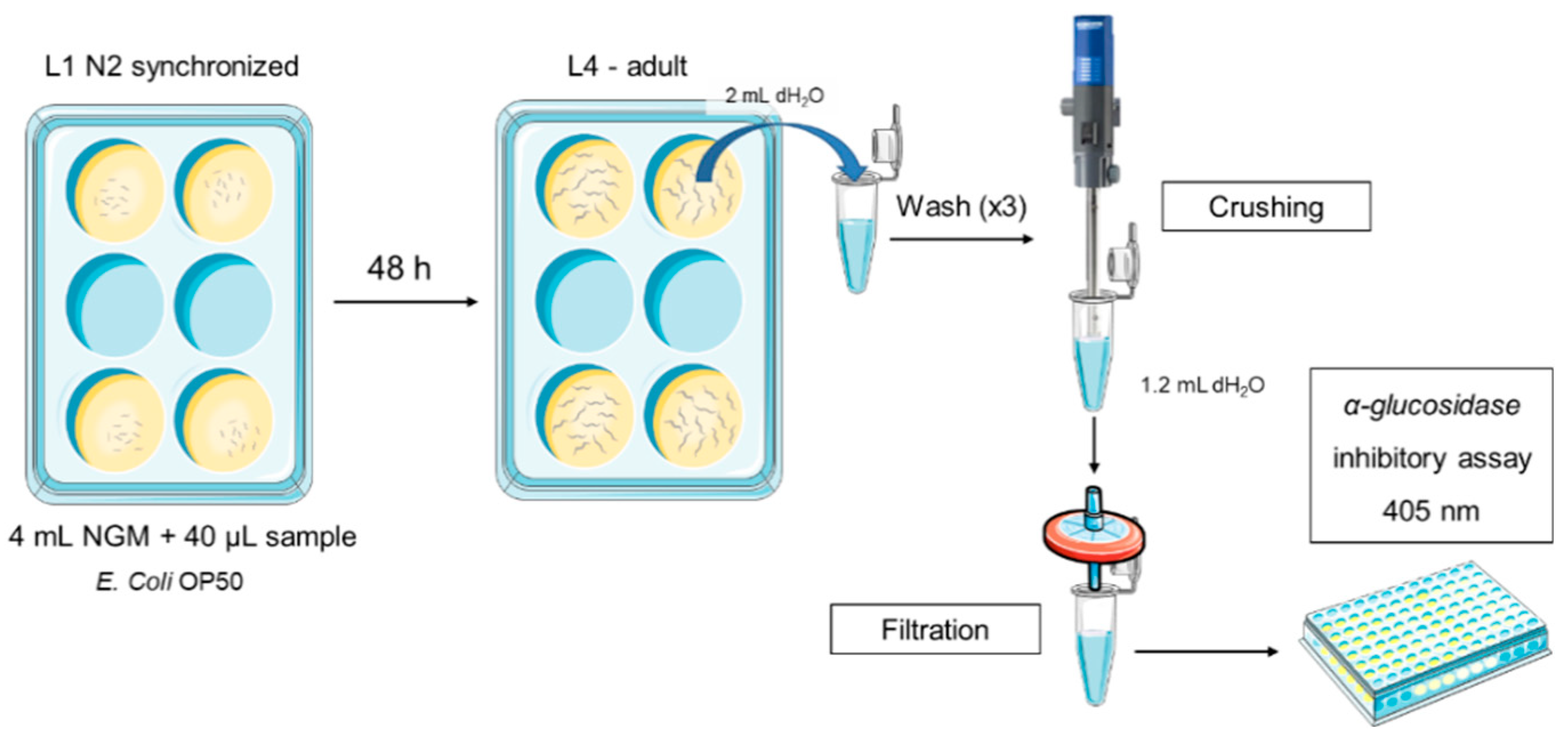
C. elegans was cultured as described previously and the strain used was N2 Bristol. All assays were performed in 6–well cell culture plates with 4 mL of Nematode Growth Medium (NGM) per well at 20 °C.
Intestinal samples of each formulation and acarbose as positive control were resuspended in distilled water at different concentrations: 0, 0.1, 1, 10, 20, 50 and 100 mg/mL.40 µL of each condition were added per well.
100 µL of E. coli OP50 were seeded on each well as the worm food source and 2,000 L1 synchronized worms were placed onto each well. After 48 h, they were collected with 2 mL of sterile water per plate, washed three times and pelleted by centrifugation (314 g / 4 min / 20 ºC). Then, worms were suspended in 1.75 mL of sterile water and crushed for 20 s at maximum power using Ultraturrax T25. Final solution was filtered through a 0.45 µm filter (Figure 5). Finally, hypoglycaemic in vivo activity assay was performed in the same manner as previously described in vitro.
3.5. Statistical analysis
All experiments were performed in triplicate. Means, standard deviations and graphs were obtained with Microsoft Excel 2013 (Microsoft Corp., Redmond, WA). Statistical analysis was performed using Stata v.12 (StataCorp LLC, College Station, TX). Normality was checked by Shapiro–Wilkinson test. Differences were estimated by ANOVA followed by pairwise comparison post hoc test using Tukey’s method (95 % CL) or post–estimation margins to check interaction among groups.
4. Conclusions
A new simple and reliable method has been developed to determine pharmacological activities inside in vivo model C. elegans. Thanks to this method, functional effects can be certainly attributed to the intake of a non–fluorimetrically traceable compound.
Two oral pharmaceutical forms were designed with OV-E extract, hard gelatine capsules and lyophilised powder to suspend in water. After digestion process, encapsulation better preserves chemical composition and α-glucosidase activity, both in vitro and in vivo. By contrast, lyophilised powder shows better hypoglycaemic activity.
Author Contributions
M.P.d.T, M.I.C., and J.L.V. conceived the experimental design and wrote the paper. M.P.d.T. performed in vitro and in vivo experiments. M.P.d.T. designed and performed statistical analyses. M.P.d.T, R.Y.C., M.I.C., and J.L.V. critically reviewed the data contributing to the draft of the manuscript.
Funding
We are grateful to the PIUNA (Plan Investigador de la Universidad de Navarra, Ref. PIUNA/2018) for the financial support of this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850. [Google Scholar] [CrossRef] [PubMed]
- Diabetes. Available online: https://www.who.int/health-topics/diabetes#tab=tab_1 (accessed on Sep 4, 2021).
- Type 2 diabetes mellitus: Prevalence and risk factors - UpToDate Available online:. Available online: https://www.uptodate.com/contents/type-2-diabetes-mellitus-prevalence-and-risk-factors (accessed on Jul 23, 2023).
- Public Assessment Report Decentralised Procedure ACARBOSE 50MG TABLETS ACARBOSE 100MG TABLETS (Acarbose) TEVA UK LIMITED;
- Höpfner, M.; Durani, B.; Spengler, M.; Fölsch, U.R. Effect of acarbose and simultaneous antacid therapy on blood glucose. Arzneimittelforschung. 1997, 47, 1108–11. [Google Scholar] [PubMed]
- Srinivasan, S.; Muthukumaran, J.; Muruganathan, U.; Venkatesan, R.S.; Jalaludeen, A.M. Antihyperglycemic effect of syringic acid on attenuating the key enzymes of carbohydrate metabolism in experimental diabetic rats. Biomed. Prev. Nutr. 2014, 4, 595–602. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Berkay Yılmaz, Y.; Antika, G.; Salehi, B.; Tumer, T.B.; Kulandaisamy Venil, C.; Das, G.; Patra, J.K.; Karazhan, N.; Akram, M.; Iqbal, M.; Imran, M.; Sen, S.; Acharya, K.; Dey, A.; Sharifi-Rad, J. Phytochemical constituents, biological activities, and health-promoting effects of the genus Origanum. Phyther. Res. 2021, 35, 95–121. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.; Moreira, E.; Grosso, C.; Andrade, P.B.; Valentão, P.; Romano, A. Phenolic profile, antioxidant activity and enzyme inhibitory activities of extracts from aromatic plants used in Mediterranean diet. J. Food Sci. Technol. 2017, 54, 219–227. [Google Scholar] [CrossRef]
- Raafat, K. Identification of phytochemicals from North African plants for treating Alzheimer’s diseases and of their molecular targets by in silico network pharmacology approach. J. Tradit. Complement. Med. 2020, 11, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yu, Z.; Zhu, H.; Zhang, W.; Chen, Y. In vitro α-glucosidase inhibitory activity of isolated fractions from water extract of Qingzhuan dark tea. BMC Complement. Altern. Med. 2016, 16, 378. [Google Scholar] [CrossRef]
- Liao, V.H. Use of Caenorhabditis elegans To Study the Potential Bioactivity of Natural Compounds. J. Agric. Food Chem. 2018, 66, 1737–1742. [Google Scholar] [CrossRef]
- C. elegans Sequencing Consortium. Genome sequence of the nematode C. elegans: a platform for investigating biology. Science 1998, 282, 2012-2018.
- Shaye, D.D.; Greenwald, I. OrthoList: A Compendium of C. elegans Genes with Human Orthologs. PLoS One 2011, 6, e20085. [Google Scholar] [CrossRef]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discov. 2006, 5, 387–398. [Google Scholar] [CrossRef]
- Culetto, E.; Sattelle, D.B. A role for Caenorhabditis elegans in understanding the function and interactions of human disease genes. Hum. Mol. Genet. 2000, 9, 869–877. [Google Scholar] [CrossRef]
- Schlotterer, A.; Kukudov, G.; Bozorgmehr, F.; Hutter, H.; Du, X.; Oikonomou, D.; Ibrahim, Y.; Pfisterer, F.; Rabbani, N.; Thornalley, P.; Sayed, A.; Fleming, T.; Humpert, P.; Schwenger, V.; Zeier, M.; Hamann, A.; Stern, D.; Brownlee, M.; Bierhaus, A.; Nawroth, P.; Morcos, M. C. elegans as Model for the Study of High Glucose– Mediated Life Span Reduction. Diabetes 2009, 58, 2450–2456. [Google Scholar] [CrossRef] [PubMed]
- Morcos, M.; Hutter, H. The Model Caenorhabditis elegans in Diabetes Mellitus and Alzheimer’s Disease. J. Alzheimer’s Dis. 2009, 16, 897–908. [Google Scholar] [CrossRef]
- Forsythe, M.E.; Love, D.; Lazarus, B.D.; Kim, E.J.; Prinz, W.; Ashwell, G.; Krause, M.K.; Hanover, J. Caenorhabditis elegans ortholog of a diabetes susceptibility locus: oga-1 (O-GlcNAcase) knockout impacts O-GlcNAc cycling, metabolism, and dauer. Proc. Natl. Acad. Sci. 2006, 103, 11952–11957. [Google Scholar] [CrossRef] [PubMed]
- de Torre, M.P.; Cavero, R.Y.; Calvo, M.I.; Vizmanos, J.L. A simple and a reliable method to quantify antioxidant activity in vivo. Antioxidants 2019, 8, 142. [Google Scholar] [CrossRef] [PubMed]
- Lemhadri, A.; Zeggwagh, N.A.; Maghrani, M.; Jouad, H.; Eddouks, M. Anti-hyperglycaemic activity of the aqueous extract of Origanum vulgare growing wild in Tafilalet region. J. Ethnopharmacol. 2004, 92, 251–256. [Google Scholar] [CrossRef]
- Vujicic, M.; Nikolic, I.; Kontogianni, V.G.; Saksida, T.; Charisiadis, P.; Vasic, B.; Stosic-Grujicic, S.; Gerothanassis, I.P.; Tzakos, A.G.; Stojanovic, I. Ethyl Acetate Extract of Origanum vulgare L. ssp. hirtum Prevents Streptozotocin-Induced Diabetes in C57BL/6 Mice. J. Food Sci. 2016, 81, H1846–H1853. [Google Scholar] [CrossRef]
- Pezzani, R.; Vitalini, S.; Iriti, M. Bioactivities of Origanum vulgare L.: an update. Phytochem. Rev. 2017, 16, 1253–1268. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides. Molecules 2019, 24, 2556. [Google Scholar] [CrossRef]
- Sukalingam, K.; Ganesan, K.; Ponnusamy, K. Evaluation of Antidiabetic Activity of Polyherbal Formulations on Type 2 Diabetic Patients: A Single Blinded Randomized Study Quick Response code. Int. J. Integr. Med. Sci. 2015, 2, 90–98. [Google Scholar]
- Wang, D.; Li, C.; Fan, W.; Yi, T.; Wei, A.; Ma, Y. Hypoglycemic and hypolipidemic effects of a polysaccharide from Fructus Corni in streptozotocin-induced diabetic rats. Int. J. Biol. Macromol. 2019, 133, 420–427. [Google Scholar] [CrossRef]
- Zhang, X.L.; Guo, Y.S.; Wang, C.H.; Li, G.Q.; Xu, J.J.; Chung, H.Y.; Ye, W.C.; Li, Y.L.; Wang, G.C. Phenolic compounds from Origanum vulgare and their antioxidant and antiviral activities. Food Chem. 2014, 152, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Mao, G. hua; Zhang, M.; Li, F.; Zou, Y.; Zhou, Y.; Zheng, W.; Zheng, D. heng; Yang, L. qing; Wu, X. yang Anti-diabetic effects of polysaccharides from ethanol-insoluble residue of Schisandra chinensis (Turcz.) Baill on alloxan-induced diabetic mice. Chem. Res. Chinese Univ. 2012, 29, 99–102. [Google Scholar] [CrossRef]
- Liu, C.; Song, J.; Teng, M.; Zheng, X.; Li, X.; Tian, Y.; Pan, M.; Li, Y.; Lee, R.J.; Wang, D. Antidiabetic and Antinephritic Activities of Aqueous Extract of Cordyceps militaris Fruit Body in Diet-Streptozotocin-Induced Diabetic Sprague Dawley Rats. Oxid. Med. Cell. Longev. 2016, 2016, 9685257. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Y.; Sarker, M.M.R.; Yan, X.; Yang, C.; Zhao, L.; Lv, X.; Liu, B.; Zhao, C. Structural characterization and antidiabetic potential of a novel heteropolysaccharide from Grifola frondosa via IRS1/PI3K-JNK signaling pathways. Carbohydr. Polym. 2018, 198, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Yoshimoto, C.; Osajima, K.; Oki, T.; Osajima, Y. In Vitro Survey of α -Glucosidase Inhibitory Food Components. Biosci. Biotechnol. Biochem. 1996, 60, 2019–2022. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Solana, R.; Coelho, N.; Santos-Rufo, A.; Gonçalves, S.; Pérez-Santín, E.; Romano, A. The influence of in vitro gastrointestinal digestion on the chemical composition and antioxidant and enzyme inhibitory capacities of carob liqueurs obtained with different elaboration techniques. Antioxidants 2019, 8, 563. [Google Scholar] [CrossRef]
- Sechi, M.; Syed, D.N.; Pala, N.; Mariani, A.; Marceddu, S.; Brunetti, A.; Mukhtar, H.; Sanna, V. Nanoencapsulation of dietary flavonoid fisetin: Formulation and in vitro antioxidant and α-glucosidase inhibition activities. Mater. Sci. Eng. C 2016, 68, 594–602. [Google Scholar] [CrossRef]
- de Torre, M.P.; Vizmanos, J.L.; Cavero, R.Y.; Calvo, M.I. Improvement of antioxidant activity of oregano (Origanum vulgare L.) with an oral pharmaceutical form. Biomed. Pharmacother. 2020, 129, 110424. [Google Scholar] [CrossRef]
- Gayoso, L.; Claerbout, A.S.; Calvo, M.I.; Cavero, R.Y.; Astiasarán, I.; Ansorena, D. Bioaccessibility of rutin, caffeic acid and rosmarinic acid: Influence of the in vitro gastrointestinal digestion models. J. Funct. Foods 2016, 26, 428–438. [Google Scholar] [CrossRef]
- Anton, S.D.; Martin, C.K.; Han, H.; Coulon, S.; Cefalu, W.T.; Geiselman, P.; Williamson, D.A. Effects of stevia, aspartame, and sucrose on food intake, satiety, and postprandial glucose and insulin levels. Appetite 2010, 55, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Langley, F.M. Nitrophenyl glucoside hydrolysis as a potential time-temperature integrator reaction. Food Chem. 1998, 62, 65–68. [Google Scholar] [CrossRef]
- Johnston, M.; Kim, J.-H. Glucose as a hormone: receptor-mediated glucose sensing in the yeast Saccharomyces cerevisiae. Biochem. Soc. Trans. 2005, 33, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.T.; Sousa, J.L.C.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: an in vitro and in silico structure-activity relationship study. J. Enzyme Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
α–glucosidase inhibitory activity in vitro expressed as percentage of inhibition (mean±SD µg/mL). Orange line represents positive control before (Acarbose – orange line) and after (Acarbose Int. – orange dashed line) digestion. OV–E (grey line): non–digested hidroalcoholic extract (OV crude extract); OV–P (red dashed line): dry powder after gastrointestinal digestion; OV–C (blue dashed line): capsules after gastrointestinal digestion; Empty caps (dashed grey line): blank control (black dashed line) of hard gelatine capsules after gastrointestinal digestion. Table below shows IC50 values (mean±SD µg/mL). Values with different letter present significant differences (p< 0.05) and same letter indicates no significant differences (p>0.05), within the same concentration. n.d.: non-determined.
Figure 1.
α–glucosidase inhibitory activity in vitro expressed as percentage of inhibition (mean±SD µg/mL). Orange line represents positive control before (Acarbose – orange line) and after (Acarbose Int. – orange dashed line) digestion. OV–E (grey line): non–digested hidroalcoholic extract (OV crude extract); OV–P (red dashed line): dry powder after gastrointestinal digestion; OV–C (blue dashed line): capsules after gastrointestinal digestion; Empty caps (dashed grey line): blank control (black dashed line) of hard gelatine capsules after gastrointestinal digestion. Table below shows IC50 values (mean±SD µg/mL). Values with different letter present significant differences (p< 0.05) and same letter indicates no significant differences (p>0.05), within the same concentration. n.d.: non-determined.
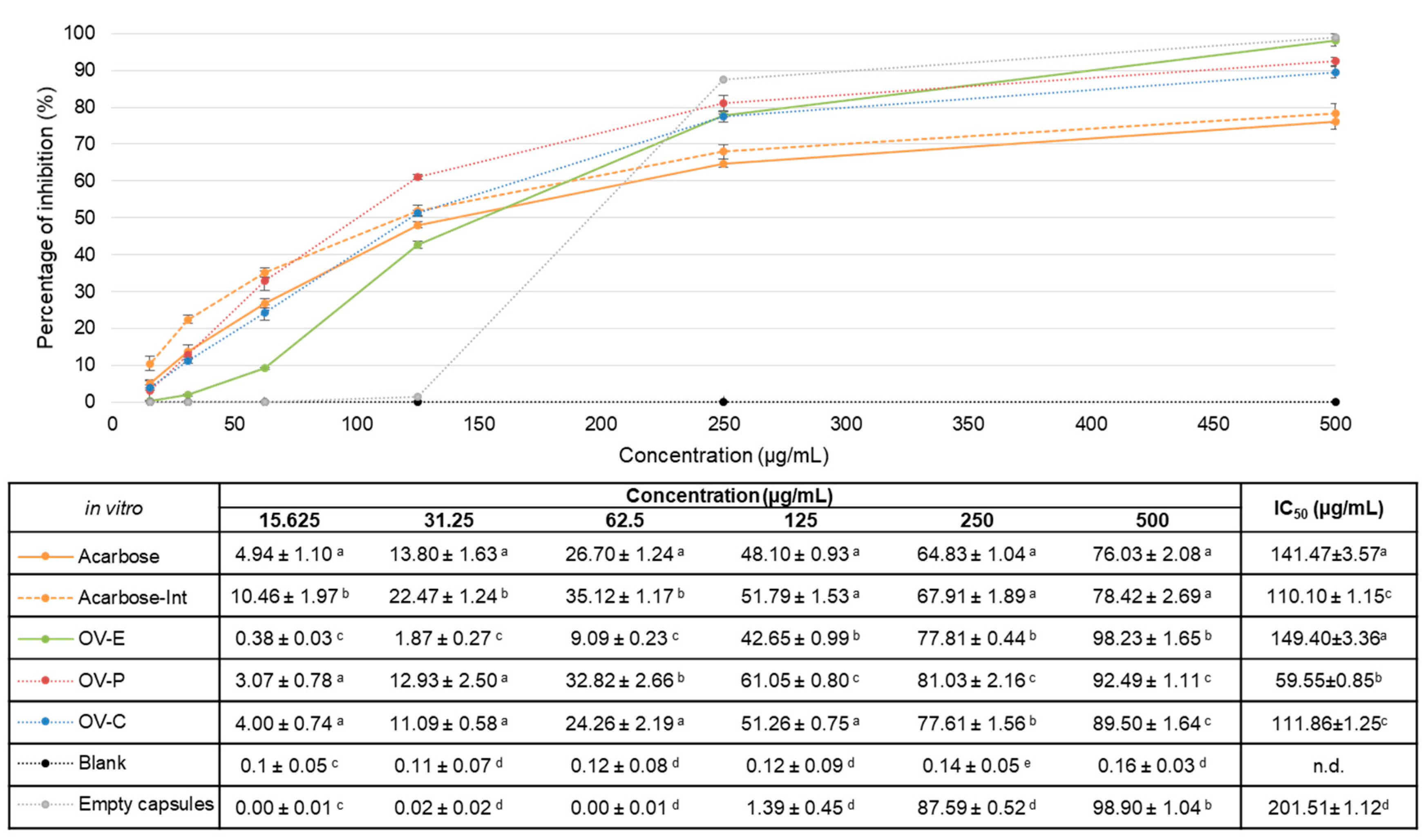
Figure 2.
α–glucosidase inhibitory activityin vivo expressed as percentage of inhibition (mean±SD %). OV–P (red line): dry powder after gastrointestinal digestion; OV–C (blue line): capsules after gastrointestinal digestion; Acarbose-Int (grey line): intestinal fraction of positive control. Table below shows percentage of inhibition values (mean±SD %). Values with different letter present significant differences (p< 0.05) and same letter indicates no significant differences (p>0.05), within same concentration.
Figure 2.
α–glucosidase inhibitory activityin vivo expressed as percentage of inhibition (mean±SD %). OV–P (red line): dry powder after gastrointestinal digestion; OV–C (blue line): capsules after gastrointestinal digestion; Acarbose-Int (grey line): intestinal fraction of positive control. Table below shows percentage of inhibition values (mean±SD %). Values with different letter present significant differences (p< 0.05) and same letter indicates no significant differences (p>0.05), within same concentration.
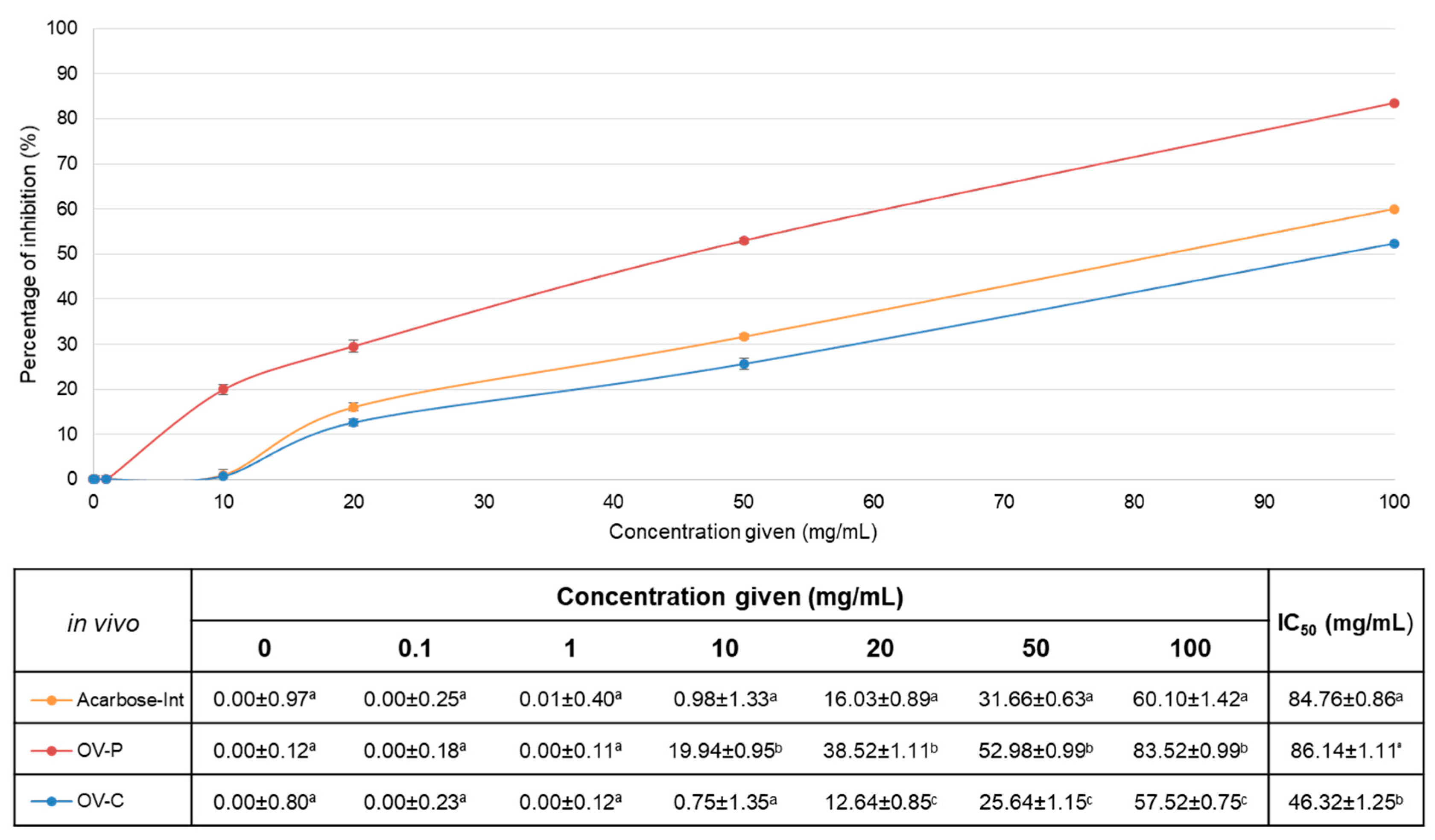
Figure 3.
IC50 values of intestinal absorbable fractions of acarbose (grey, Acarbose-Int), capsules (blue, OV-C) and powder (red, OV-P). On the left, values of the in vitro assay (mean±SD µg/mL). On the right, values for in vivo assay (mean±SD mg/mL). ***: non-statistical differences, p < 0.05.
Figure 3.
IC50 values of intestinal absorbable fractions of acarbose (grey, Acarbose-Int), capsules (blue, OV-C) and powder (red, OV-P). On the left, values of the in vitro assay (mean±SD µg/mL). On the right, values for in vivo assay (mean±SD mg/mL). ***: non-statistical differences, p < 0.05.
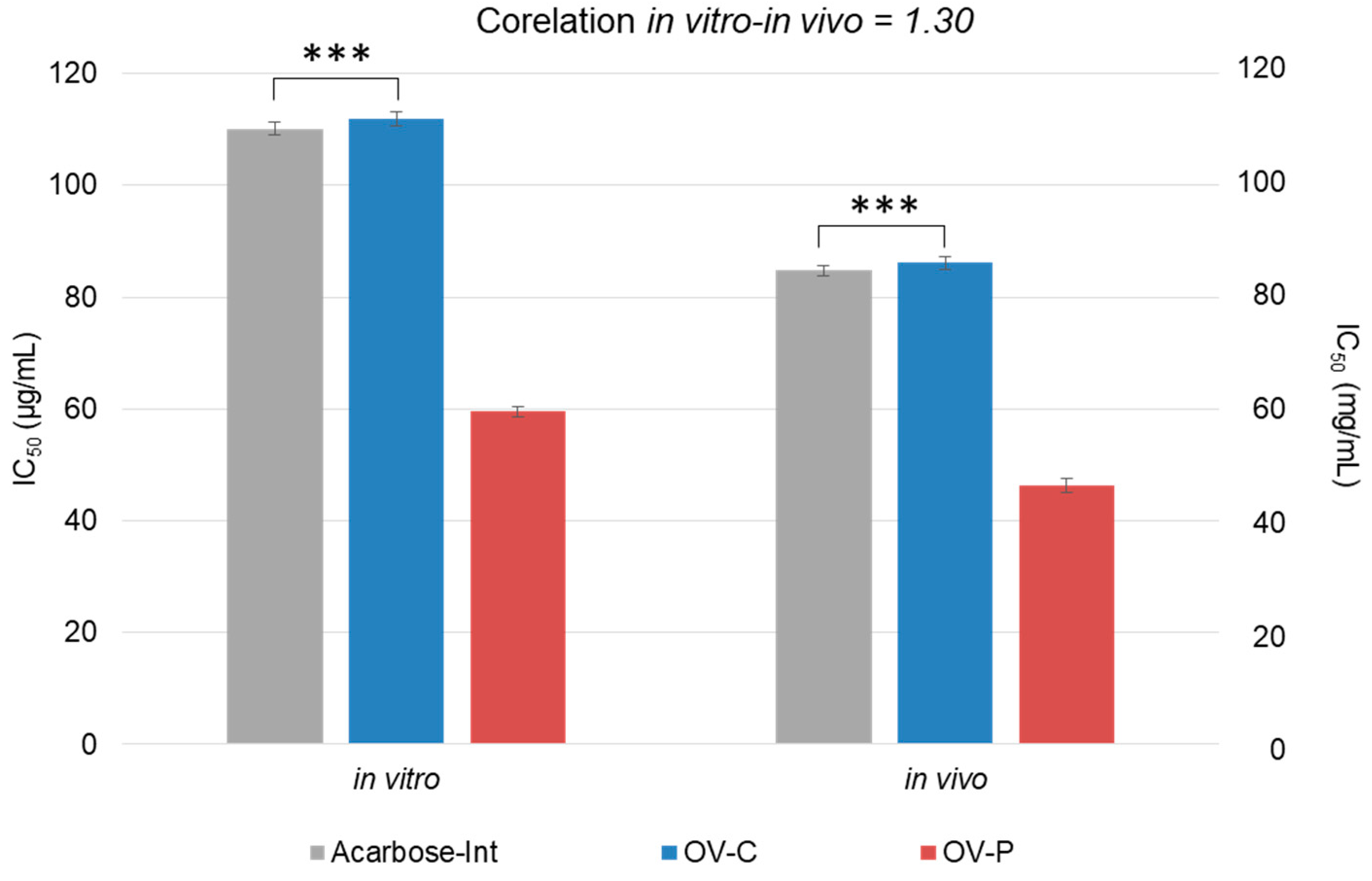
Figure 4.
Scheme of the inhibition and activity of α–glucosidase. Reactive p–NPG (4–Nitrophenyl–α–D–glucopyranoside) is an analogous disaccharide that is broken down by the enzyme into P–nitrophenol (a compound with a yellow colour measurable at 405 nm) and glucose. In the presence of any α–glucosidase inhibitor (e.g. acarbose), a yellow coloration would not result as the enzyme is removed.
Figure 4.
Scheme of the inhibition and activity of α–glucosidase. Reactive p–NPG (4–Nitrophenyl–α–D–glucopyranoside) is an analogous disaccharide that is broken down by the enzyme into P–nitrophenol (a compound with a yellow colour measurable at 405 nm) and glucose. In the presence of any α–glucosidase inhibitor (e.g. acarbose), a yellow coloration would not result as the enzyme is removed.
Figure 5.
Scheme of the adapted method for hypoglycaemic activity in vivo.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



