
Submitted:
31 July 2023
Posted:
01 August 2023
You are already at the latest version
Abstract
When “hijacked” by compulsive behaviors that affect the reward and stress centers of the brain, functional changes in dopamine circuitry occur as the consequence of pathological brain adaptation. As a brain explanans for mental health, with a central functional role in behavioral regulation from healthy reward seeking to pathological adaptation to stress in response to adversity, we may use dopamine regulation in interaction with other brain mechanisms, as discussed herein, to gather a deeper insight into environmentally triggered mental health changes to understand, for example, specific behavioral changes observed during and after the COVID-19 pandemic. Pandemic-related adversities, the stresses they engendered, and the long lockdown periods where people had to rely on digital tools to get feed-back rewards via the internet can be seen as the major triggers of changes in motivation and reward seeking behavior worldwide. The pathological adaptation of dopamine-mediated reward circuitry in the brain offers a highly plausible explanation why, when pushed by fate and circumstance into a physiological brain state of anti-reward, human behavior and mental health changes almost inevitably depending on individual vulnerability to stress. A unified conceptual account that places dopamine function at the centre of the current global mental health context is proposed.
Keywords:
Dopamine
; Brain
; Reward
; Stress
; Pathological Adaptation
; Behavior
; Covid19 pandemic
; Digital Addiction
; Global Mental Health
1. Introduction
The neurotransmitter dopamine fulfills a critical function in regulating the responses of the mesolimbic system, also known as the reward system, in the neural circuits of the mammalian brain [1]. The reward system governs and regulates responses ranging from pleasure and craving to disgust and anhedonia, triggered by chemical and other stimuli and guiding a larger proportion of our behavior than we may be aware of, or are ready to admit [2]. In ancient times, the function of the reward system made the difference between life and death because it made us and other species deploy most of our or their attention and behavioral effort towards things that were important for survival of the species, such as food, sleep, and sex. Our ancestors did not have a supermarket around the corner but had to hunt for food, and often had to deploy a considerable amount of time and energy to find it. An individual who found sweet fruit in the environment, for example, had better eat it as quickly and in as large quantities as possible before another did. Ripe fruits with higher sugar content providing more energy than other foods, the human preference for sweet food stuffs may well be due to the evolutionary advantage that craving and eating such high-calorie food has brought. Responding selectively to survival-relevant stimuli thus, in the course of evolution, has become hardwired into the brain's reward system [3]. Food, sleep, physical contact, and sex are primary stimuli that reinforce the neural connections of the reward system, and a craving for these primary stimuli is inherent in humans [4], as well as most other mammals [5]. The reward system is composed of brain structures that are responsible for mediating the physiological and cognitive processing of reward in a natural, neurobiological process during which the brain associates diverse stimuli such as substances, situations, events, or activities with a positive or desirable outcome. This results in adjustments of an individual’s behavior, ultimately leading them to search for that particular positive stimulus. Reward requires the coordinated release of heterogenous neurotransmitters where dopamine plays the central role by mediating the reward value of food, drink, sex, social interaction, or specific substances or stimuli, as in abuse and addiction [6]. Moreover, the influence of the dopaminergic reward pathways on circadian rhythms [1], translated by the fact that reward-related activities such as feeding, exercise, sex, substance use, and social interactions lead to an elevated level of dopamine and alter the circadian rhythms of the central nervous system [1,7], has hitherto unsuspected implications for human health, especially in the post-COVID-19 digital age.
The discovery of the reward system harks back to experiments from 1954 [8]. Scientistsimplanted electrodes in the brains of rats in order to stimulate specific areas of the brain with light electrical currents. The electrodes were activated by a lever that the rats could pull themselves. The researchers were able to observe how the rats pressed the lever up to 2,000 times within an hour to receive the internal electric stimulation, and for several hours until they were completely exhausted. Primary reinforcements such as food and sleep became less attractive than the stimulation that followed the lever pressure. Animal studies have also shown that there are reward circuits in the brain that originate in the evolutionarily ancient midbrain and connect with other brain areas [9]. One of the most important messengers in these regulatory circuits is the neurotransmitter dopamine. Dopamine is primarily released when a reward comes as a surprise or when stimuli appear that indicate a reward [10,11], but is not as such responsible for the positive feelings during consumption of the reward, which are essentially mediated by serotonin and endorphins. Interactions with the endocannabinoid system, which is uniquely placed to influence dopaminergic and serotonergic neurotransmission, generate neuromodulatory effects at cellular and circuit levels [12]. Technological advances that facilitate the precise identification and control of genetically targeted neuronal populations may soon achieve a better understanding of the complex functional links between these systems and their potential relevance for motivated behavior and reward response. The dopaminergic and opiodergic reward pathways of the brain are, as explained above, critical for survival since they provide the drive for eating, love and reproduction, the so-called ‘natural rewards’. This involves the release of dopamine in the nucleus accumbens and frontal lobes, however, dopamine is not the only reward transmitter, and dopaminergic neurons are not the final common path for all rewards. Dopamine antagonists and lesions of the dopamine systems appear to spare the rewarding effects of nucleus accumbens and frontal cortex brain stimulation and the rewarding effects of apomorphine [13]. Reward circuitry is multisynaptic and dopamine is the critical link in this circuitry.
Release of dopamine can likewise be produced by ‘unnatural rewards’ such as alcohol, cocaine, methamphetamine, heroin, nicotine, marijuana, and other drugs, of by compulsive activities such as gambling, eating, and sex, and by risk taking behaviors [14]. Drugs can trigger a particularly powerful release of dopamine compared to primary reinforcers or ‘natural rewards’. While primary reinforcers can increase dopamine levels by as much as 100 percent, drugs like cocaine can spike dopamine levels by as much as 1000 percent. The body perceives the strong increase in dopamine release as a particularly high reward that is “better than expected”. While a saturation effect sets in with the primary reinforcers at some point because no more dopamine is released, this is not the case with drugs, at least in the initial phase of the development of dependence [14]. Drugs interfere with the reward system by directly stimulating dopamine release. The typical course of an addiction development is characterized by the person resorting to the addictive substance again and again [15]. Like compulsory and excessive sugar and alcohol consumption, repeated drug use (abuse) leads to addiction by causing changes the activity of the brain’s reward system, which preferentially responds to the target substance and to stimuli (context) directly associated with the habit. This can be specific places, moments in time, or social cues and people (consuming with friends). As the person's attention becomes increasingly focused on a substance of abuse, other primary reinforcers lose their appeal [16]. The person is then interested in little else but the drug craved, in other words, the reward system is « hijacked » by the drug. Eventually, in later stages of addiction, a strong association develops between drug-associated stimuli, such as specific locations where the drug is used, and associated behavioral responses that lead to use. As addictive behavior becomes more and more of a habit that cannot be controlled by willpower, the reward system becomes increasingly networked with other brain areas that control habitual behavior [16,17]. This connectivity also explains the compulsive consumption behavior in drug addicts when they relapse without any conscious decision to use again, as addictive behavior is triggered more or less automatically by certain stimuli beyond individual conscious awareness [18,19,20].
The following parts of this article discuss how functional changes in dopamine circuitry, when hijacked by certain compulsive and addicitve behaviors affecting the whole reward system of the brain, may provide global insight into mental health changes in individuals and groups, and provide an understanding of specific health behavior changes observed during and after the COVID-19 pandemic. Pandemic-related adversities, the stresses they engendered, and the long lockdown periods where people had to rely on digital tools to get feed-back rewards via the internet can be seen as the major triggers of changes in motivation and reward seeking behavior worldwide. The pathological adaptation of dopamine-mediated reward circuitry in the brain offers a highly plausible explanation why individuals of all nations, during and after the pandemic, were pushed by fate and circumstance into a physiological brain state of anti-reward the clinical term for which is anhedonia. A unified conceptual model that places dopamine function at the centre of the global mental health context is discussed prior to the conclusions.
2. Dopamine, Reward, and Behavioral Regulation
The important role of dopamine in the neurocircuitry of the reward system provides a functional brain-based interpretation of change in health-related behavior. It needs to be examined in the conceptual framework of the neurobiology of motivation in both healthy behavior and in addiction. The resources of modern neuroscience offer an opportunity to conceptualize the dopaminergic neurocircuitry and its multiple interactions with other pathways, functions, and mechanisms in healthy reward and in addictive processes. They provide the keys to understanding vulnerability and behavioral changes in individuals and groups under conditions of stress and environmental challenges, as will be discussed in greater detail in chapter 3. There are essentially three specific types of behavioral change that have been functionally linked to dopamine interactions in the brain.
2.1. Impulsive Behavior
To attempt to clarify the concept of vulnerability to impulsivity, individual or population vulnerability is to be defined as an outcome of other concurrent variables in a potentially preventable, reversible process [21]. A vulnerable population may refer to individuals with disabilities (physical or mental impairment that substantially limits major life activities), or individuals who represent socially or economically disadvantaged demographic populations, such as homeless, rural, adopted, elderly, racial/ethnic minorities, gender minorities, religious minorities, children with parents who are active-duty members of the Armed Forces, veterans, and others. Vulnerability is to be placed on a continuum of severity, and various characteristics and circumstances, or combinations of variables. The concept of vulnerability is important in public health. Vulnerable individuals often resort to alternative means of coping with stressors compared with more resilient individuals, and may therefore be at higher risk of developing extreme reward seeking behaviors in the form of substance or behavioral addictions. There is an initial relationship between vulnerability (predisposition) to impulsivity and subsequently expressed compulsive behavior, in both health and addiction [22]. This relationship can be explained by the neuroplasticity of the circuits involved, where plasticity-dependent deregulation of dopamine interactions in the brain accounts for how circuits may gradually become dysregulated, stay dysregulated, and then provide further vulnerability to further dysregulation [23]. The neural systems basis of compulsive behavior involves dopaminergic as well as serotoninergic mechanisms within the corticostriatal circuitry [24,25,26]. Also, increasing evidence supports a role of the noradrenergic system in compulsive behavior [27,28], with an involvement of noradrenergic mechanisms in the regulation of glucocorticoid levels [29,30] linked to stress-induced repetitive behaviors [31]. The proven role of noradrenergic interactions in anxiety, coping and impulse control [32,33,34,35] contributes to both the emergence and to the increase in severity of compulsions [36,37,38].
2.2. Craving and Bingeing
Preoccupation, a motivation-related mental state triggered by aversive internal or external stimuli may produce excessive reward-seeking in healthy and in addictive behavior [39,40,41]. Human neuroimaging studies show that the prefrontal cortex (orbitofrontal, medial prefrontal, prelimbic/cingulate) and the basolateral amygdala are critical in cue-induced craving and bingeing behavior. Whether the underlying mechanisms reflect a disruption of frontal brain regions secondary to changes in striatal dopamine activity, or alternatively reflect a primary disruption of frontal regions that regulate dopamine cell activity, remains to be determined. Dopamine neurons exist in multiple types, connected with distinct brain networks, ensuring distinct roles in motivational control. Some dopamine neurons encode motivational value, supporting brain networks for seeking, evaluation, and value learning. Others encode motivational salience, supporting brain networks for orienting, cognition, and general motivation [41]. In extreme reward-seeking behavior, increase in severity of the compulsion leads to craving and bingeing, which are often, but not always, relating to a specific stage of the addiction cycle [41,42], where neuroplasticity intervenes with a change in the firing activities of mesolimbic dopamine neurons during initial stimulus (substance or other drug) exposure, and is then translated to engagement of the dorsal striatum, disruption of frontal system function, and recruitment of the brain’s stress systems [43]. Similar to what happens in the case of drug abuse, chronic, repeated stimulations of dopaminergic system in the NAc by palatable food and associated cues shifts signalling to dorso-striatal dopaminergic pathways, which results in habit formation for compulsive eating, and reflects a maladaptive, stimulus-driven habit that overrides voluntary goal-directed behavior [43,44]. Through a process of consolidation, this produces a powerful drive for food- or drug-seeking behavior even months after dieting or withdrawal, and provides a brain explanation for relapse in addiction. Relapse following withdrawal is the result of healthy neuroadaptive processes gone wrong [45], or pathological adaptation, in the central nervous system, opposing the acute reinforcing actions of the drug via a deregulation of the mechanisms that mediate positive reinforcement. Such deregulation explains the emergence of anxiety, anhedonia, and depression during withdrawal.
2.3. Negative Affect
Motivational changes related to the sudden withdrawal of reward [46,47], in both health and addiction, produce negative affect and changes in mood that can lead to anhedonia [46], depression and, in the extreme, suicide. Negative affect is closely linked to the brain’s stress systems [4] involving the corticotropin releasing factor (CRF), which interacts with dopaminergic brain circuitry. The primary physiological stress response is partly mediated by the hypothalamic–pituitary–adrenocortical (HPA) axis in mammals, with stressors activating the central nervous system and the hypothalamus and thereby the CRF to the pituitary gland. The pituitary gland will in its turn release adrenocorticotropic hormone (ACTH), which induces glucocorticoid synthesis and release from the adrenal tissue into the blood. The glucocorticoids will then affect target tissues throughout the body as each step of the HPA axis is self-regulated by an array of feedback loops. This explains why numerous other neurotransmitter systems, such as dynorphin, NPY, substance P, nociceptin, and orexin systems interact with the brain’s stress systems. Stressful situations are unavoidable element in everyday life. Stressors activate a number of complex mental and physiological reactions in the organism [49], thus affecting the state of health of an individual. Stress is the main risk factor in the development of sleep disorders, depression, addiction and other health issues that result from the addiction [50]. Reward-related behavior in stressed individuals affects dopamine levels and dopaminergic neuronal activity in the mesolimbic dopamine system. Changes in mesolimbic dopaminergic neurotransmission are important for coping with the stress, as they allow adaption to behavioral responses to various environmental stimuli. Modulation of the dopaminergic reward system is necessary for monitoring and selecting the optimal process for coping with stressful situations, and aversive stressful events may negatively regulate the dopaminergic reward system, perturbing reward sensitivity, which is closely associated with chronic stress-induced depression [51]. The mesolimbic dopamine system is excited not only by reward but also by aversive stressful stimuli, which adds further intriguing complexity to the relationship between stress and the reward system [52]. Internal factors altered by stress may produce drug addiction vulnerability [53] by neuro-inflammatory, neurotrophic, and neurotransmitter factors impacting mechanisms of craving and relapse susceptibility, and linked to sleep disorders [49,53] and depression [54].
2.4. Sleep
While healthy positive stimulation of the reward system promotes healthy sleep, compulsive reward seeking on the other hand leads to the asynchronization of circadian rhythms [55,56,57,58], centrally controlled by serotonin and responsible for dysfunctional sleep patterns. Pathological changes in sleep patterns and behavior are a product of pathological brain adaptation involving antagonistic processes (allostasis) that lead to irritability, dysphoria, anxiety and anhedonia in what is sometimes called the brain’s anti-reward system [55,56,57,58,59,60,61]. Because compulsive reward seeking litterally usurps everyday behavior, i.e. eating, sexuality, exercise, and other, it « hijacks » the natural effect of substances or activities that produce pleasure through the release of dopamine [1,12,13]. Natural reward at first decreases the reward threshold, while chronic addiction to a substance or a behavior increases this threshold, hence the need to consume more of a given drug to reach it, thus, compulsive behavior ultimately leads to addiction by increasing the concentration of extracellular dopamine through mechanisms including (i) decrease in the inhibitory tone exerted by GABAergic neurons on dopaminergic neurons, (ii) release of opioids and endogenous cannabinoids, and (iii) a direct action on dopaminergic neurons by increasing their frequency of discharge [61]. Dopamine not only underlies the pleasurable effects but is also involved in much more complex phenomena of attribution of the "added value" ("incentive value") associated with a behavior or a substance. A contextual cue associated with a drug, after a period of conditioning, replaces the value of the drug itself and is able to precipitate relapse by an overwhelming, urgent and irrepressible desire to consume [10,11,13,61]. This process is related to changes in the amygdala [62] resulting in negative emotional states and sleep disruption [63,64].
3. Dopamine and Post-Pandemic Mental Health
In 2022 the World Health Organisation (WHO) issued a brief [65] with facts and figures showing that the COVID-19 pandemic has had a severe impact on the mental health and wellbeing of people around the world. While many individuals have adapted, others have experienced mental health problems that only in some cases are a consequence of COVID-19 infection. The brief states further that the pandemic also continues to impede access to mental health services and has raised concerns about increases in suicidal behavior, stressing the causal role of psychological and environmental factors producing ontological insecurity individuals and populations. In the wake of the COVID-19 pandemic, experts were witnessing a
- ➢
- significant rise in addictions and related mental illnesses
- ➢
- significant rise in corresponding anti-depressant prescription uptake
- ➢
- increased risk of suicidal ideation or suicide
Any potential doubts about the impact of COVID-19 on mental health have been largely put to rest by the most recent papers on the topic [66,67,68,69,70]. Loneliness, physical exhaustion, sleep disturbances, and anhedonia linked to compulsive or addictive behaviors such as binge-eating [71,72], substance abuse [73], and digital reward craving [74] were identified as linked to suicidal ideation [71,75,76], in particular in younger individuals, during and after the pandemic. These reflect some of the most negative effects of pandemic-related adversity on mental health world-wide.
3.1. Adversity and Vulnerability
Mental health issues arise from a specific context. The effects of the pandemic show beyond all reasonable doubt that they can arise when adversity begins to affect people’s capacity of coping with the stresses adversity produces. Conditions of extreme adversity challenge the stress and immune system responses needed for coping and, beyond a certain threshold, such conditions can trigger behavioral changes as listed and discussed here above ; all of these have been functionally linked to the complex brain pathways involving dopamine release, reward mechanisms, and their interaction or regulation thereof [52,54]. Having to cope with adversity during childhood increases the risk of mental health problems in adulthood, and it has been suggested that this effect may be mediated by increased striatal dopamine neurotransmission [77]. Also, while resilience may be viewed as an individual’s positive adaptation to experiences of adversity [78],it is also known that conditions of adversity reinforce already exisiting vulnerability to mental illness, and may create new vulnerabilty in specific adverse contexts such as a pandemic [68], or armed conflict [79]. It has been confirmed that stress responses can cause functional changes in the amygdala and the dopaminergic circuits of the brain in relation to stress-induced cortisol release mechanisms [80,81,82]. Studies showing cortisol-induced increase in serotonin in subjects with major depressive disorder offer insight into functional links between stress, sleep disorder, and depression, where increase in one brain correlate affects another, thereby creating or increasing physiological vulnerability to mental illness in individuals under prolonged stress.
3.2. From Reward to Anhedonia
The COVID-19 pandemic brought negative health consequences such as consuming high fat and sugar foods and increased body weight [83], anxiety and depression [84], and anhedonia [83,85]. Brain dopamine not only plays a critical role in the subjective pleasure associated with positive rewards, but also in the objective reinforcement and incentive motivation associated with food and water, brain stimulation, psychomotor stimulants, and opiate or other drug rewards [61,62]. Brain dopamine plays a very important role in reinforcement of response habits, conditioned preferences, and synaptic plasticity in the brain circuitry that governs learning and memory. Therein, the process that leads from the initial subjective pleasure associated with a stimulus to craving followed by a negative reward response in the brain and producing, ultimately, what is called anhedonia is a fundamental aspect of the neurobiology of motivation [86,87]. Dopamine is intimately linked to the brain genesis of anhedonia through its interaction with the glucocorticoid system that responds to chronic stress. Prolonged stress leads to inflammatory processes that negatively impact on functional connectivity in the corticostriatal (dopaminergic) reward circuits and symptoms, in close interaction with the endocannabinoid and orexin systems [88] thereby affecting the synthesis and release of dopamine [89]. Through this process, stimuli and activities we once enjoyed doing and that gave us pleasure become by and by aversive and, ultimately, detrimental to our well-being. Anhedonia, i.e. the inability to experience pleasure from natural rewards, is thus a direct consequence of our brain’s response to chronic stress [89,90].
3.3. The New «Digital Drug»
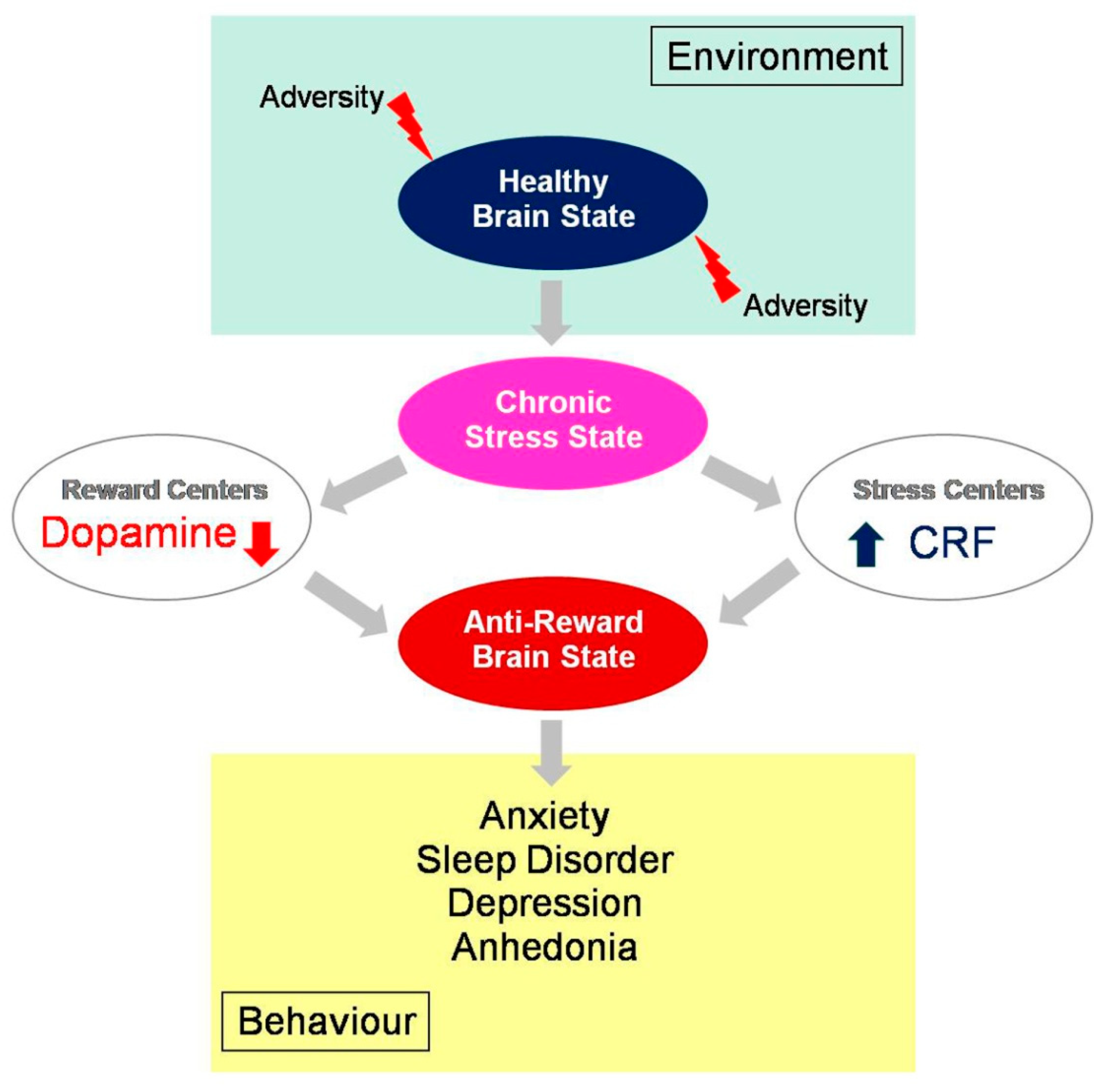
COVID-19 spread across the world in a rapid pace, and to further limit the spread of infection, lockdown was declared in most parts of the world. People were forced to stay indoors and the internet was the only source of entertainment. This context engendered addictive behavior with measurable negative effects on anxiety and sleep quality, especially among younger individuals [91,92]. In digital addiction, the internet is a channel [94,95] through which individuals may access whatever content they want (games, social media, shopping, sex and so forth), wherever they want, and whenever they want it. The development of the addictive response is thereby digitally facilitated and instantly available to anyone - adults, adolescents, and young children [93,94,95,96]. At an advanced stage, digital addiction [95] is associated with significant and permanent symptomatic psychological, cognitive, and physiological states, with measurable dopamine deficiency and impaired mental health [97]. Psychological stress [98,99,100,101], anxiety and depression [102,103,104], eating disorders [105,106,107], sleeplessness [108,109], and mood changes with suicidal ideation [110,111] are the most frequently reported. A compilation of cross-national studies on more than 89,000 participants from 31 nations, performed almost ten years ago well before the COVID-19 pandemic, already suggested a global prevalence estimate for digital addiction of 6% worldwide [96]. Interdependent variables such as socio-cultural factors, biological vulnerabilities (genetic predisposition, pre-existing metabolic disorders), and psychological factors (personality characteristics, negative affect) play a critical role here [112,113,114,115,116,117]. Excessive seeking of the new digital drug and addiction to the internet have been identified as part of the consequences of the COVID-19 pandemic with a major impact on mental health of predominantly young people and teenagers [118,119]. This involves, like all compulsive behavior loops, the brain’s reward circuitry with all the complex interactions between environmental, metabolic, and neurobiological changes in the brain discussed here above. Dopamine, and the modulation thereof under conditions of adversity and stress, therein (Figure 1) is to be conceived as the common functional denominator [93,120,121,122,123].
4. Conclusions
Dopamine regulates reward-related behaviors as discussed here above through the mesolimbic dopaminergic pathway. Chronic stress affects dopamine levels and dopaminergic neuronal activity in that same brain system. By regulating behavioral responses environmental stimuli, dopaminergic neuromodulation is a unifying brain explanation for healthy and pathological processes of coping with stress. Adversity as a consequence of the prolonged stressful events engendered by the COVID-19 pandemic, correlated with increasing digital reward seeking worldwide stressful, has perturbed reward sensitivity, produced chronic stress-induced depression, and mesolimbic dopamine regulation in individuals across populations and nations. As a central neurotransmitter at the heart of an intriguingly complex relationship between stress and the brain’s reward system, it ensures a critical function in the slow process from healthy reward response to bran state of anti-reward that has become a global challenge for mental health worldwide. Future research, from molecular to integrative brain circuitry levels and behavior, has acquired a novel global significance in this new mental health context [124], prompting towards further investigations into the hitherto still poorly explored links between dopamine and other neurotransmitter (serotonine, noradrenaline) and hormonal (cortisol, oxytocine) functions.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data can be found in the literature cited in this communication.
Conflicts of Interest
The author declares no conflict of interest.
References
- Lewis RG, Florio E, Punzo D, Borrelli E. The Brain's Reward System in Health and Disease. Adv Exp Med Biol 2021, 1344, 57–69. [Google Scholar] [CrossRef]
- Kringelbach ML, Berridge KC. The Functional Neuroanatomy of Pleasure and Happiness. Discov Med 2010, 9, 579–587. [Google Scholar]
- Robbins TW, Everitt BJ. Neurobehavioural mechanisms of reward and motivation. Curr Opin Neurobiol 1996, 6, 228–36. [Google Scholar] [CrossRef] [PubMed]
- De Decker A, Verbeken S, Sioen I, Van Lippevelde W, Braet C, Eiben G, Pala V, Reisch LA, De Henauw S; I. Family Consortium. Palatable food consumption in children: interplay between (food) reward motivation and the home food environment. Eur J Pediatr. 2017, 176, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Everitt, BJ. Sexual motivation: a neural and behavioural analysis of the mechanisms underlying appetitive and copulatory responses of male rats. Neurosci Biobehav Rev. 1990, 14, 217–32. [Google Scholar] [CrossRef]
- Comings DE, Blum K. Reward deficiency syndrome: genetic aspects of behavioral disorders. Prog Brain Res 2000, 126, 325–41. [Google Scholar] [CrossRef]
- Johnston, JD. Physiological responses to food intake throughout the day. Nutr Res Rev 2014, 27, 107–18. [Google Scholar] [CrossRef]
- Olds J, Milner P. Positive reinforcement produced by electrical stimulation of septal area and other regions of rat brain. J Comp Physiological Psychology 1954, 47, 419–27. [Google Scholar] [CrossRef]
- Kapsimali M, Dumond H, Le Crom S, Coudouel S, Vincent JD, Vernier P. Evolution et développement des systèmes neuromodulateurs dopaminergiques chez les Vertébrés [Evolution and development of dopaminergic neurotransmitter systems in vertebrates]. J Soc Biol 2000, 194, 87–93. [Google Scholar] [CrossRef]
- Haber SN, Knutson B. The reward circuit: linking primate anatomy and human imaging. Neuropsychopharmacology 2010, 35, 4–26. [Google Scholar] [CrossRef]
- Schultz, W. Neuronal Reward and Decision Signals: From Theories to Data. Physiol Rev 2015, 95, 853–951. [Google Scholar] [CrossRef] [PubMed]
- Peters KZ, Cheer JF, Tonini R. Modulating the Neuromodulators: Dopamine, Serotonin, and the Endocannabinoid System. Trends Neurosci 2021, 44, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Wise RA, Rompre PP. Brain dopamine and reward. Annu Rev Psychol 1989, 40, 191–225. [Google Scholar] [CrossRef] [PubMed]
- Substance Abuse and Mental Health Services Administration US Office of the Surgeon General (US). Facing Addiction in America: The Surgeon General's Report on Alcohol, Drugs, and Health Washington (DC) US Department of Health and Human Services, 2016; CHAPTER 2, THE NEUROBIOLOGY OF SUBSTANCE USE, MISUSE, AND ADDICTION. Available online at: https://www.ncbi.nlm.nih.gov/books/NBK424849/. 4248.
- 15. Jahan AR, Burgess DM. Substance Use Disorder. [Updated 2023 Apr 29]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing, 2023; Available online at: https://www.ncbi.nlm.nih.gov/books/NBK570642/. /.
- Volkow ND, Michaelides M, Baler R. The Neuroscience of Drug Reward and Addiction. Physiol Rev 2019, 99, 2115–2140. [Google Scholar] [CrossRef]
- Kato A, Shimomura K, Ognibene D, Parvaz MA, Berner LA, Morita K, Fiore VG. Computational models of behavioral addictions: State of the art and future directions. Addict Behav 2023, 140, 107595. [Google Scholar] [CrossRef]
- Michaelsen MM, Esch T. Understanding health behavior change by motivation and reward mechanisms: a review of the literature. Front Behav Neurosci. 2023, 17, 1151918. [Google Scholar] [CrossRef]
- Koob GF, & Volkow ND. Neurobiology of addiction: a neurocircuitry analysis. Lancet Psychiatry, 2016, 3, 760–773.
- Keramati M, Gutkin B. Imbalanced decision hierarchy in addicts emerging from drug-hijacked dopamine spiraling circuit. PLoS One. 2013, 8, e61489. [Google Scholar] [CrossRef]
- Sussman S, Sinclair DL. Substance and Behavioral Addictions, and Their Consequences among Vulnerable Populations. Int J Environ Res Public Health 2022, 19, 6163. [Google Scholar] [CrossRef]
- Maldonado R, Calvé P, García-Blanco A, Domingo-Rodriguez L, Senabre E, Martín-García E. Vulnerability to addiction. Neuropharmacology. 2021, 186, 108466. [Google Scholar] [CrossRef]
- Velazquez-Sanchez C, Muresan L, Marti-Prats L, Belin D. The development of compulsive coping behaviour is associated with a downregulation of Arc in a Locus Coeruleus neuronal ensemble. Neuropsychopharmacology 2023, 48, 653–663. [Google Scholar] [CrossRef]
- Moreno M, Flores P. Schedule-induced polydipsia as a model of compulsive behavior: neuropharmacological and neuroendocrine bases. Psychopharmacology 2012, 219, 647–59. [Google Scholar] [CrossRef] [PubMed]
- Robbins TW, Koob GF. Selective disruption of displacement behaviour by lesions of the mesolimbic dopamine system. Nature 1980, 285, 409–12. [Google Scholar] [CrossRef] [PubMed]
- Mora S, Merchan A, Vilchez O, Aznar S, Klein AB, Ultved L, et al. Reduced cortical serotonin 5-HT2A receptor binding and glutamate activity in high compulsive drinker rats. Neuropharmacology 2018, 143, 10–9. [Google Scholar] [CrossRef] [PubMed]
- Ansquer S, Belin-Rauscent A, Dugast E, Duran T, Benatru I, Mar AC, et al. Atomoxetine decreases vulnerability to develop compulsivity in high impulsive rats. Biol Psychiatry. 2014, 75, 825–32. [Google Scholar] [CrossRef] [PubMed]
- Higgins GA, Brown M, St John J, MacMillan C, Silenieks LB, Thevarkunnel S. Effects of 5-HT2C receptor modulation and the NA reuptake inhibitor atomoxetine in tests of compulsive and impulsive behaviour. Neuropharmacology 2020, 170, 108064. [Google Scholar] [CrossRef]
- Ziegler DR, Cass WA, Herman JP. Excitatory influence of the locus coeruleus in hypothalamic-pituitary-adrenocortical axis responses to stress. J Neuroendocrinol 1999, 11, 361–9. [Google Scholar] [CrossRef]
- Thrivikraman KV, Kinkead B, Owens MJ, Rapaport MH, Plotsky PM. Locus coeruleus noradrenergic modulation of diurnal corticosterone, stress reactivity and cardiovascular homeostasis in male rats. Neuroendocrinology. 2021, 112, 763–76. [Google Scholar]
- Lustberg D, Iannitelli AF, Tillage RP, Pruitt M, Liles LC, Weinshenker D. Central norepinephrine transmission is required for stress-induced repetitive behavior in two rodent models of obsessive-compulsive disorder. Psychopharmacology 2020, 237, 1973–87. [Google Scholar] [CrossRef]
- Bari A, Robbins TW. Noradrenergic versus dopaminergic modulation of impulsivity, attention and monitoring behaviour in rats performing the stop-signal task: possible relevance to ADHD. Psychopharmacology 2013, 230, 89–111. [Google Scholar] [CrossRef]
- Pattij T, Vanderschuren LJ. The neuropharmacology of impulsive behaviour. Trends Pharmacol Sci 2008, 29, 192–9. [Google Scholar] [CrossRef]
- Daviu N, Bruchas MR, Moghaddam B, Sandi C, Beyeler A. Neurobiological links between stress and anxiety. Neurobiol Stress 2019, 11, 100191. [Google Scholar] [CrossRef] [PubMed]
- Tanaka M, Yoshida M, Emoto H, Ishii H. Noradrenaline systems in the hypothalamus, amygdala and locus coeruleus are involved in the provocation of anxiety: basic studies. Eur J Pharmacol 2000, 405, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Coban DA, Tan O. The relationship between childhood trauma and obsessive-compulsive disorder, comorbid attention deficit hyperactivity disorder, and impulsivity. Arch Neuropsychiatry 2019, 57, 37–43. [Google Scholar]
- Saxena S, Maidment KM, Vapnik T, Golden G, Rishwain T, Rosen RM, et al. Obsessive-compulsive hoarding: symptom severity and response to multimodal treatment. J Clin Psychiatry 2002, 63, 21–7. [Google Scholar] [CrossRef]
- Rosa-Alcázar Á, García-Hernández MD, Parada-Navas JL, Olivares-Olivares PJ, Martínez-Murillo S, Rosa-Alcázar AI. Coping strategies in obsessive-compulsive patients during Covid-19 lockdown. Int J Clin Health Psychol 2021, 21, 100223. [Google Scholar] [CrossRef]
- Mathis V, Kenny PJ. From controlled to compulsive drug-taking: The role of the habenula in addiction. Neurosci Biobehav Rev 2019, 106, 102–111. [Google Scholar] [CrossRef]
- Asensio S, Hernández-Rabaza V, Orón Semper JV. What Is the "Trigger" of Addiction? Front Behav Neurosci 2020, 14, 54. [Google Scholar] [CrossRef]
- Bromberg-Martin ES, Matsumoto M, Hikosaka O. Dopamine in motivational control: rewarding, aversive, and alerting. Neuron. 2010, 68, 815–34. [Google Scholar] [CrossRef]
- 42. Schaefer LM, Forester G, Burr EK, Laam L, Crosby RD, Peterson CB, Crow SJ, Engel SG, Dvorak RD, Wonderlich SA. Examining the role of craving in affect regulation models of binge eating: Evidence from an ecological momentary assessment study. J Psychopathol Clin Sci 2023. [CrossRef]
- 43. Moore CF, Panciera JI, Sabino V, Cottone P. Neuropharmacology of compulsive eating. Philos Trans R Soc Lond B Biol Sci. 2018, 373, 20170024. [CrossRef]
- Parylak SL, Koob GF, Zorrilla EP. The dark side of food addiction. Physiol. Behav 2011, 104, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Weiss F, Ciccocioppo R, Parsons LH, Katner S, Liu X, Zorrilla EP, Valdez GR, Ben-Shahar O, Angeletti S, Richter RR. Compulsive drug-seeking behavior and relapse. Neuroadaptation, stress, and conditioning factors. Ann N Y Acad Sci. 2001, 937, 1–26. [Google Scholar]
- Koob, GF. Anhedonia, Hyperkatifeia, and Negative Reinforcement in Substance Use Disorders. Curr Top Behav Neurosci 2022, 58, 147–165. [Google Scholar] [CrossRef]
- Piper, ME. Withdrawal: Expanding a Key Addiction Construct. Nicotine Tob Res 2015, 17, 1405–15. [Google Scholar] [CrossRef] [PubMed]
- Backström T, Winberg S. Central corticotropin releasing factor and social stress. Front. Neurosci, 2013, 7, 117. [Google Scholar] [CrossRef]
- Han KS, Kim L, Shim I. Stress and sleep disorder. Exp Neurobiol 2012, 21, 141–50. [Google Scholar] [CrossRef]
- Grabowska K, Ziemichód W, Biała G. Recent Studies on the Development of Nicotine Abuse and Behavioral Changes Induced by Chronic Stress Depending on Gender. Brain Sci 2023, 13, 121. [Google Scholar] [CrossRef]
- Hales CA, Stuart SA, Griffiths J, Bartlett J, Arban R, Hengerer B, Robinson ES. Investigating neuropsychological and reward-related deficits in a chronic corticosterone-induced model of depression. Psychoneuroendocrinology, 2023, 147, 105953. [CrossRef]
- Baik, JH. Stress and the dopaminergic reward system. Exp Mol Med 2020, 52, 1879–1890. [Google Scholar] [CrossRef]
- Sochal M, Ditmer M, Gabryelska A, Białasiewicz P. The Role of Brain-Derived Neurotrophic Factor in Immune-Related Diseases: A Narrative Review. Journal of Clinical Medicine 2022, 11, 6023. [Google Scholar] [CrossRef]
- Correia AS, Cardoso A, Vale N. Oxidative Stress in Depression: The Link with the Stress Response, Neuroinflammation, Serotonin, Neurogenesis and Synaptic Plasticity. Antioxidants (Basel) 2023, 12, 470. [Google Scholar] [CrossRef]
- Smevik H, Habli S, Saksvik SB, Kliem E, Evensmoen HR, Conde V, Petroni A, Asarnow RF, Dennis EL, Eikenes L, Kallestad H, Sand T, Thompson PM, Saksvik-Lehouillier I, Håberg AK, Olsen A. Poorer sleep health is associated with altered brain activation during cognitive control processing in healthy adults. Cereb Cortex 2023, 33, 7100–7119. [Google Scholar] [CrossRef]
- Uccella S, Cordani R, Salfi F, Gorgoni M, Scarpelli S, Gemignani A, Geoffroy PA, De Gennaro L, Palagini L, Ferrara M, Nobili L. Sleep Deprivation and Insomnia in Adolescence: Implications for Mental Health. Brain Sci, 2023, 13, 569. [CrossRef]
- Sollenberger NA, Sequeira S, Forbes EE, Siegle GJ, Silk JS, Ladouceur CD, Ryan ND, Dahl RE, Mattfeld AT, McMakin DL. More time awake after sleep onset is linked to reduced ventral striatum response to rewards in youth with anxiety. J Child Psychol Psychiatry. 2023, 64, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Alloy LB, Chat IK, Grehl MM, Stephenson AR, Adogli ZV, Olino TM, Ellman LM, Miller GE, Nusslock R. Reward and Immune Systems in Emotion (RISE) prospective longitudinal study: Protocol overview of an integrative reward-inflammation model of first onset of major depression in adolescence. Brain Behav Immun Health, 2023, 30, 100643. [CrossRef]
- Scaplen KM, Kaun KR. Reward from bugs to bipeds: a comparative approach to understanding how reward circuits function. J Neurogenet. 2016, 30, 133–48. [Google Scholar] [CrossRef]
- Blum K, Gondré-Lewis M, Steinberg B, et al. Our evolved unique pleasure circuit makes humans different from apes: Reconsideration of data derived from animal studies. J Syst Integr Neurosci. 2018, 4, 10. [Google Scholar]
- Wise, RA. Dopamine and Reward: The Anhedonia Hypothesis 30 years on. Neurotox Res. 2008, 14, 169–183. [Google Scholar] [CrossRef]
- Volkow ND, Michaelides M, Baler R. The Neuroscience of Drug Reward and Addiction. Physiol Rev, 2019, 99, 2115–2140. [CrossRef]
- Gong L, Liao T, Liu D, Luo Q, Xu R, Huang Q, Zhang B, Feng F, Zhang C. Amygdala Changes in Chronic Insomnia and Their Association with Sleep and Anxiety Symptoms: Insight from Shape Analysis. Neural Plast, 2019, 2019, 8549237. [CrossRef]
- Cirrincione L, Plescia F, Malta G, Campagna M, Lecca LI, Skerjanc A, Carena E, Baylon V, Theodoridou K, Fruscione S, Cannizzaro E. Evaluation of Correlation between Sleep and Psychiatric Disorders in a Population of Night Shift Workers: A Pilot Study. Int J Environ Res Public Health. 2023, 20, 3756. [Google Scholar] [CrossRef]
- World Health Organization. Mental health and COVID-19: Early evidence of the pandemics impact. Geneva: World Health Organization, https://apps.who.int/iris/bitstream/handle/10665/352189/WHO-2019-nCoV-Sci-Brief-Mental-health-2022.1-eng.pdf. 2022.
- Oakes, MB. Ontological insecurity in the post-covid-19 fallout: using existentialism as a method to develop a psychosocial understanding to a mental health crisis. Med Health Care Philos. 2023 Jun 30. [CrossRef]
- Tomlinson M, Marlow M. COVID-19 and mental health: Building back better or reimagining a new way forward? PLoS Med 2023, 20, e1004216. [Google Scholar] [CrossRef]
- Winkler P, Mohrova Z, Mlada K, Kuklova M, Kagstrom A, Mohr P, et al. Prevalence of current mental disorders before and during the second wave of COVID-19 pandemic: An analysis of repeated nationwide cross-sectional surveys. J Psychiatr Res 2021, 139, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Pierce M, McManus S, Hope H, Hotopf M, Ford T, Hatch SL, et al. Mental health responses to the COVID-19 pandemic: a latent class trajectory analysis using longitudinal UK data. Lancet Psychiatry 2021, 8, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Knudsen AKS, Stene-Larsen K, Gustavson K, Hotopf M, Kessler RC, Krokstad S, et al. Prevalence of mental disorders, suicidal ideation and suicides in the general population before and during the COVID-19 pandemic in Norway: A population-based repeated cross-sectional analysis. Lancet Reg Health Eur 2021, 4, 100071. [Google Scholar] [CrossRef]
- Davies HL, Hübel C, Herle M, Kakar S, Mundy J, Peel AJ, Ter Kuile AR, Zvrskovec J, Monssen D, Lim KX, Davies MR, Palmos AB, Lin Y, Kalsi G, Rogers HC, Bristow S, Glen K, Malouf CM, Kelly EJ, Purves KL, Young KS, Hotopf M, Armour C, McIntosh AM, Eley TC, Treasure J, Breen G. Risk and protective factors for new-onset binge eating, low weight, and self-harm symptoms in >35,000 individuals in the UK during the COVID-19 pandemic. Int J Eat Disord 2023, 56, 91–107. [Google Scholar] [CrossRef]
- Güzel Â, Mutlu NL, Molendijk M. COVID-19-related changes in eating disorder pathology, emotional and binge eating and need for care: a systematic review with frequentist and Bayesian meta-analyses. Eat Weight Disord 2023, 28, 19. [Google Scholar] [CrossRef]
- Martinelli TF, Nagelhout GE, Best D, Vanderplasschen W, van de Mheen D. Factors associated with problematic substance use before and during the COVID-19 pandemic among a drug addiction recovery cohort: A prospective study in the Netherlands, Belgium, and UK. J Subst Use Addict Treat. 2023, 148, 209025. [Google Scholar] [CrossRef]
- Zhao L, Li X, Yang Q, Peng Y, Jiang L, Jia P, Shi W. The longitudinal association between internet addiction and depressive and anxiety symptoms among Chinese adolescents before and during the COVID-19 pandemic. Front Public Health. 2023, 10, 1096660. [Google Scholar] [CrossRef]
- Stanley IH, Flarity KM, April MD. Suicide Ideation, Plans, and Attempts Attributed to the COVID-19 Pandemic Among US Veterans. JAMA Netw Open. 2023, 6, e2320193. [Google Scholar] [CrossRef] [PubMed]
- Tardeh S, Adibi A, Mozafari AA. Prevalence of Suicide Ideation and Attempt during COVID-19 Pandemic: A Systematic Review and Meta-Analysis. Int J Prev Med. 2023, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Egerton A, Valmaggia LR, Howes OD, Day F, Chaddock CA, Allen P, Winton-Brown TT, Bloomfield MAP, Bhattacharyya S, Chilcott J, Lappin JM, Murray RM, McGuire P. Adversity in childhood linked to elevated striatal dopamine function in adulthood. Schizophr Res 2016, 176, 171–176. [Google Scholar] [CrossRef]
- Chuang SP, Wu JYW, Wang CS. Resilience and Quality of Life in People with Mental Illness: A Systematic Review and Meta-Analysis. Neuropsychiatr Dis Treat 2023, 19, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Østergaard MLD, Aponte-Canencio DM, Barajas Ortiz Y, Velez Botero HJ, Simon Modvig J, Brasholt M. Vulnerability factors in conflict-related mental health. Med Confl Surviv 2023, 39, 63–80. [Google Scholar] [CrossRef]
- Oswald LM, Wong DF, McCaul M, Zhou Y, Kuwabara H, Choi L, Brasic J, Wand GS. Relationships among ventral striatal dopamine release, cortisol secretion, and subjective responses to amphetamine. Neuropsychopharmacology 2005, 30, 821–32. [Google Scholar] [CrossRef]
- Wand GS, Oswald LM, McCaul ME, Wong DF, Johnson E, Zhou Y, Kuwabara H, Kumar A. Association of amphetamine-induced striatal dopamine release and cortisol responses to psychological stress. Neuropsychopharmacology 2007, 32, 2310–20. [Google Scholar] [CrossRef]
- Yaribeygi H, Panahi Y, Sahraei H, Johnston TP, Sahebkar A. The impact of stress on body function: A review. EXCLI J 2017, 16, 1057–1072. [Google Scholar] [CrossRef]
- Fleischer E, Landaeta-Díaz L, González-Medina G, Horovitz O. Anxiety, anhedonia, and related food consumption in Israelis populations: An online cross-sectional study two years since the outbreak of COVID-19. Heliyon 2023, 9, e17211. [Google Scholar] [CrossRef]
- Vindegaard N, Benros ME. COVID-19 pandemic and mental health consequences: Systematic review of the current evidence. Brain Behav Immun. 2020, 89, 531–542. [Google Scholar] [CrossRef]
- Costa R, Pinto TM, Conde A, Mesquita A, Motrico E, Figueiredo B. Women's perinatal depression: Anhedonia-related symptoms have increased in the COVID-19 pandemic. Gen Hosp Psychiatry 2023, 84, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne SJ, Melendez SI, Olmstead MC. Investigating dopamine and glucocorticoid systems as underlying mechanisms of anhedonia. Psychopharmacology (Berl). 2018, 235, 3103–3113. [Google Scholar] [CrossRef]
- Kim HJJ, Zagzoog A, Ceni C, Ferrisi R, Janz N, Laprairie RB. Dual Cannabinoid and Orexin Regulation of Anhedonic Behaviour Caused by Prolonged Restraint Stress. Brain Sci 2023, 13, 314. [Google Scholar] [CrossRef]
- Gold MS, Blum K, Febo M, Baron D, Modestino EJ, Elman I, Badgaiyan RD. Molecular role of dopamine in anhedonia linked to reward deficiency syndrome (RDS) and anti- reward systems. Front Biosci (Schol Ed) 2018, 10, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Phillips R, Walsh E, Jensen T, Nagy G, Kinard J, Cernasov P, Smoski M, Dichter G. Longitudinal associations between perceived stress and anhedonia during psychotherapy. J Affect Disord 2023, 330, 206–213. [Google Scholar] [CrossRef]
- Zhang C, Liu B, Pawluski J, Steinbusch HWM, Kirthana Kunikullaya U, Song C. The effect of chronic stress on behaviors, inflammation and lymphocyte subtypes in male and female rats. Behav Brain Res 2023, 439, 114220. [Google Scholar] [CrossRef] [PubMed]
- Huang Y, Zhao N. Generalized anxiety disorder, depressive symptoms and sleep quality during COVID-19 outbreak in China: A web-based cross-sectional survey. Psychiatry Res 2020, 288, 112954. [Google Scholar] [CrossRef]
- Lin M-P. Prevalence of Internet Addiction during the COVID-19 Outbreak and Its Risk Factors among Junior High School Students in Taiwan. International Journal of Environmental Research and Public Health 2020, 17, 8547. [Google Scholar] [CrossRef]
- Dresp-Langley B, Hutt A. Digital Addiction and Sleep. Int J Environ Res Public Health. 2022, 19, 6910. [Google Scholar] [CrossRef]
- Kuss DJ, Griffiths MD, Karila L, Billieux, J. Internet addiction: a systematic review of epidemiological research for the last decade. Curr Pharm Des 2014, 20, 4026–52. [Google Scholar] [CrossRef]
- Griffiths MD, Pontes HM. Internet Addiction Disorder and Internet Gaming Disorder are not the same. J Addict Res Ther, 2014, 5, e1. [Google Scholar]
- Cheng C, Li AY. Internet addiction prevalence and quality of (real) life: a meta-analysis of 31 nations across seven world regions. Cyberpsychol Behav Soc Netw. 2014, 17, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Liu M, Luo J. Relationship between peripheral blood dopamine level and internet addiction disorder in adolescents: A pilot study. Int J Clin Exp Med, 2015, 8, 9943–9948. [Google Scholar]
- Nalwa K, Anand AP. Internet addiction in students: a cause of concern. Cyberpsychol Behav, 2003, 6, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Cao F, Su L. Internet addiction among Chinese adolescents: prevalence and psychological features. Child Care and Health Development, 2006, 33, 275–281. [Google Scholar]
- Fu KW, Chan WS, Wong PW, Yip PS. Internet addiction: prevalence, discriminant validity and correlates among adolescents in Hong Kong. Br J Psychiatry, 2010, 196, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Saikia AM, Das J, Barman P, Bharali MD. Internet Addiction and its Relationships with Depression, Anxiety, and Stress in Urban Adolescents of Kamrup District, Assam. J Family Community Med, 2019, 26, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Dalbudak E, et al. Relationship of internet addiction severity with depression, anxiety and alexithymia, temperament, and character in university students. Cyberpsychol Behav Soc Netw, 2013, 16, 272–278. [Google Scholar] [CrossRef]
- Shakya HB, Christakis NA. Association of ‘Facebook’ use with compromised well-being: A longitudinal study. American Journal of Epidemiology 2017, 185, 203–211. [Google Scholar]
- Lau JTF, Walden DL, Wu AMS, Cheng KM, Lau MCM, Mo PKH. Bidirectional predictions between Internet addiction and probable depression among Chinese adolescents. J Behav Addict, 2018, 7, 633–643. [Google Scholar] [CrossRef]
- Hinojo-Lucena FJ, Aznar-Díaz I, Cáceres-Reche MP, Trujillo-Torres JM, Romero-Rodríguez JM. Problematic Internet Use as a Predictor of Eating Disorders in Students: A Systematic Review and Meta-Analysis Study. Nutrients, 2019, 11, e2151. [Google Scholar] [CrossRef] [PubMed]
- Moreno MA, Eickhoff J, Zhao Q, Suris, JC. College Students and Problematic Internet Use: A Pilot Study Assessing Self-Appraisal and Independent Behavior Change. J Adolesc Health, 2019, 64, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy S, Chetlapalli SK. Internet addiction: Prevalence and risk factors: A cross-sectional study among college students in Bengaluru, the Silicon Valley of India. Indian J Public Health, 2015, 59, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Li JB, Lau JTF, Mo PKH, Su XF, Tang J, Qin ZG, Gross DL. Insomnia partially mediates the association between problematic Internet use and depression among secondary school students in China. J Behav Addict 2017, 6, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Chen YL, Gau SS. Sleep problems and internet addiction among children and adolescents: a longitudinal study. J Sleep Res 2016, 25, 458–65. [Google Scholar] [CrossRef]
- Kim K, et al. Internet addiction in Korean adolescents and its relation to depression and suicidal ideation: a questionnaire survey. Int J Nurs Stud 2006, 43, 185–192. [Google Scholar] [CrossRef]
- Keles B, McCrae N, Grealish A. A systematic review: the influence of social media on depression, anxiety and psychological distress in adolescents, International Journal of Adolescence and Youth. 2020, 25, 79–93.
- Yau YH, Potenza MN. Gambling disorder and other behavioral addictions: recognition and treatment. Harv Rev Psychiatry 2015, 23, 134–146. [Google Scholar] [CrossRef]
- Greenfield, D. Treatment Considerations in Internet and Video Game Addiction: A Qualitative Discussion. Child and Adolescent Clinics of North America Reviews, Youth Internet Habits and Mental Health 2018. [Google Scholar]
- Twenge, JM. Have Smartphones Destroyed a Generation? The Atlantic, 2017. [Google Scholar]
- Shaw M, Black DW. Internet addiction: definition, assessment, epidemiology and clinical management. CNS Drugs 2008, 22, 353–365. [Google Scholar] [CrossRef]
- Kuss DJ, Griffiths MD, Karila L, Billieux, J. Internet addiction: a systematic review of epidemiological research for the last decade. Curr Pharm Des 2014, 20, 4026–52. [Google Scholar] [CrossRef] [PubMed]
- Rumpf H, et al. Including gaming disorder in the ICD-11: The need to do so from a clinical and public health perspective. Journal of Behavioral Addictions 2018, 7, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Onukwuli VO, Onyinye EN, Udigwe IB, Umeh UM, Enebe JT, Umerah AT. Internet addiction during the COVID-19 pandemic among adolescents in southeast Nigeria and implications for adolescent care in the post-pandemic era: A cross-sectional study. SAGE Open Med 2023, 11, 20503121231152763. [Google Scholar] [CrossRef]
- Kumar G, Dash P, Jnaneswar A, Suresan V, Jha K, Ghosal S. Impact of internet addiction during COVID-19 on anxiety and sleep quality among college students of Bhubaneswar city. J Educ Health Promot 2022, 11, 156. [Google Scholar] [CrossRef]
- Melca IA, Teixeira EK, Nardi AE, Spear AL. Association of Internet Addiction and Mental Disorders in Medical Students: A Systematic Review. Prim Care Companion CNS Disord 2023, 25, 22r03384. [Google Scholar] [CrossRef]
- Dresp-Langley, B. Children's Health in the Digital Age. Int J Environ Res Public Health 2020, 17, 3240. [Google Scholar] [CrossRef]
- Dresp-Langley, B. Dresp-Langley B. Consciousness Beyond Neural Fields: Expanding the Possibilities of What Has Not Yet Happened. Front Psychol 2022, 12, 762349. [Google Scholar] [CrossRef]
- Oster, H. The interplay between stress, circadian clocks, and energy metabolism. J Endocrinol 2020, 247, R13–R25. [Google Scholar] [CrossRef]
- Arias D, Saxena S, Verguet, S. Quantifying the global burden of mental disorders and their economic value. eClinicalMedicine 2022, 54, 101675. [Google Scholar] [CrossRef]
Figure 1.
The behavioral consequences of an anti-reward brain state, with depleted dopamine and increased cortisol levels in the centers that regulate the brain’s response to reward in a process of pathological adaptation to chronic stress under adverse environmental conditions.
Figure 1.
The behavioral consequences of an anti-reward brain state, with depleted dopamine and increased cortisol levels in the centers that regulate the brain’s response to reward in a process of pathological adaptation to chronic stress under adverse environmental conditions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



