
Submitted:
03 May 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
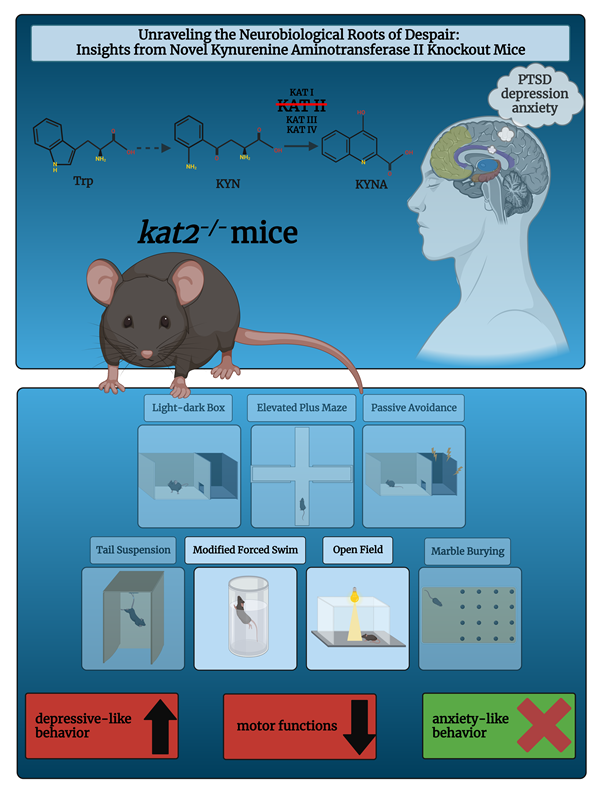
Memory and emotion are highly vulnerable to psychiatric disorders like post-traumatic stress disorder (PTSD), which has been linked to serotonin (5-HT) metabolism disruptions. In fact, over 90% of the 5-HT precursor tryptophan (Trp) is metabolized via the Trp-kynurenine (KYN) meta-bolic pathway, producing a variety of bioactive molecules. The aadat (kat2) gene encodes mito-chondrial kynurenine aminotransferase (KAT) isotype 2, responsible for kynurenic acid (KYNA) production. Little is known about its role in behavior. In CRISPR/Cas9-induced aadat knockout (kat2−/−) mice, we examined the effects on emotion, memory, motor function, Trp and its metabolite levels, enzyme activities in the plasma and the urine of 8-week-old males compared to wild-type mice. Transgenic mice showed more depressive-like behaviors in the forced swim test, but not in the tail suspension, anxiety, or memory tests. They also had fewer center field and corner entries, shorter walking distances, and fewer jumping counts in the open field test. Plasma metabolite levels are generally consistent with those of urine: KYN, antioxidant KYNs, 5-hydroxyindolacetic acid, and indole-3-acetic acid levels are lower; enzyme activities in KATs, kynureninase, and monoamine oxidase/aldehyde dehydrogenase are lower, but kynurenine 3-monooxygenase is higher; and ox-idative stress and excitotoxicity indices are higher. Transgenic mice show depression-like behavior in a learned helplessness model, emotional indifference, and motor deficits, coupled with a decrease in KYNA, a shift of Trp metabolism toward the KYN-3-HK pathway, and a partial decrease in the gut microbial Trp-indole pathway metabolite. This is the first evidence that deleting the aadat gene causes depression-like behaviors that are unique to despair experience, which appears to be linked to excitatory neurotoxic and oxidative stresses. This may lead to the development of a double-hit preclinical model in experience-based depression, better understanding of these complex condi-tions, and more effective therapeutic strategies by elucidating the relationship between Trp me-tabolism and PTSD pathogenesis.
Keywords:
post-traumatic stress disorder (PTSD)
; depression
; tryptophan
; kynurenine
; microbiota
; transgenic mice
1. Introduction
The interaction between memory and emotion involves a complex interplay of neural, cognitive, and physiological processes involving the amygdala, hippocampus, and prefrontal cortex (1-5). Orderly function at multi-layered levels is essential to maintaining sound mental well-being (6-9). The reciprocal interaction between cognitive function and affective states can significantly impact each other. Cognitive impairment can lead to affective disturbances, triggering emotional responses such as frustration, anxiety, and stress, particularly when individuals feel a loss of control over their cognitive abilities (10). Similarly, emotional disturbances such as depression and anxiety can influence memory function, increasing vulnerability to cognitive challenges (11, 12). This intricate bidirectional link between cognition and emotions can lead to changes in brain structure, function, behavior, lifestyle, and neurotransmitter systems (13, 14). Memory impairment and emotional disturbance are associated with a wide range of systematic diseases and neuropsychiatric disorders such as Alzheimer’s disease (AD), Parkinson’s disease, traumatic brain injury, major depressive disorder (MDD), and post-traumatic stress disorder (PTSD) (15-20).
The serotonergic nervous system plays an important role in regulating mood, anxiety, and cognition (21-24). Serotonin (5-hydroxytryptamine, 5-HT) is involved in cognitive processes such as attention, learning, and memory (25-27). Studies indicate that 5-HT enhances long-term memory consolidation and improves cognitive flexibility, which is the ability to switch between different cognitive tasks or mental sets (28-37). 5-HT is implicated in regulating mood and anxiety, influencing cognitive function (38, 39). Mental illnesses like MDD, eating disorders, obsessive-compulsive disorder, schizophrenia (SCZ), and PTSD are associated with dysregulation of 5-HT (40-45). Selective serotonin reuptake inhibitors (SSRIs) are commonly used for these conditions, targeting the serotonergic nervous system (46-48). Furthermore, abnormalities in the serotonergic system also affect norepinephrine and dopamine (49-51).
The complex interplay of tryptophan (Trp)-kynurenine (KYN) and 5-HT metabolism is crucial for comprehending the pathogenesis of mental illnesses (52, 53). The Trp-KYN metabolic system, closely associated with 5-HT metabolism, plays a pivotal role in the production of prooxidants and antioxidants, regulation of the immune system, and the balance between neurotoxicity and neuroprotection (54, 55). Approximately 2% of L-Trp undergoes metabolism through the 5HT metabolic pathway; however, over 90% of Trp is catabolized through the KYN route, which safely to say that it governs Trp metabolism (Figure 1, a,b) (56). Various factors, including stress, inflammation, and the gut microbiome, influence this system (57-59). Dysregulation of the KYN route has been linked to mental health conditions such as MDD, SCZ, and AD (60).
Approximately 5% of dietary Trp undergoes conversion to indole and its derivatives via the gut microbial indole pyruvate pathway, facilitated by gut microbiota like Escherichia coli and Clostridium sporogenes (81-83). These bacteria produce indole and its derivatives, including indole-3-acetic acid (IAA), indole-3-aldehyde, and indoxyl sulfate (ISN), among others (Figure 1, c) (76, 84, 85). These signaling molecules influence a variety of physiological processes in the host, including the intestinal barrier, inflammation, oxidative stress, hormone secretion, and nuclear receptor activation (86-88). Many gastrointestinal and hepatic conditions, such as colorectal cancer, irritable bowel syndrome, non-alcoholic fatty liver disease, and hepatic encephalopathy, are linked to the gut microbial indole pyruvate pathway (80, 86, 89). Disruptions in the gut microbial indole pyruvate pathway can impact the neurotransmitters, hormones, and immune factors in the brain (90-92). This can also impair the communication between the gut and the brain through the vagus nerve (93-95). The increasing attention towards the role of the gut microbial indole pathway in mental health disorders such as depression, anxiety, autism, SCZ, and AD highlights its potential significance in the field of neuropsychiatry (96-100).
However, the understanding of the interplay between Trp-KYN, 5-HT, and indole metabolism in the pathogenesis of mental illnesses remains limited. Kynurenine aminotransferases (KATs) are members of the pyridoxal-5'-phosphate-dependent enzyme family involved in the KYN metabolic pathway. The KYN metabolism is responsible for the conversion of L-KYN to kynurenic acid (KYNA), an antioxidant and neuroprotective metabolite with implications for various central nervous system (CNS) diseases (68, 101, 102). Among the KAT enzymes, kynurenine/alpha-aminoadipate aminotransferase (KAT/AadAT, aka KAT II) is a mitochondrial enzyme encoded in the gene aadat (kat2) (103). KAT II is considered to play the most important role among the four isozymes in the cellular environment due to its highest enzymatic activity close to the physiological pH. Thus, KAT II plays a prominent role in KYNA production in the human brain and is considered a crucial target for managing CNS disorders (104).
Preclinical research has significantly contributed to our understanding of mental illnesses by elucidating the underlying pathomechanisms and identifying potential therapeutic targets (105-113). Researchers have employed preclinical animal models to examine the causes and effects of mental disorders, thereby attaining a comprehensive understanding of their underlying pathology (114-121). In vitro models, such as cell cultures and organoids, have facilitated the investigation of complex molecular pathways linked to mental disorders (122-125). Animal models, along with other in vivo models, have been instrumental in studying the behavioral, cognitive, and physiological dimensions of mental disorders (126-132). These models allow researchers to simulate disease conditions, assess symptomatology, and evaluate the efficacy of potential interventions (132, 133). Transgenic animals are vital in biomedical research, enabling the replication of human conditions through gene deletion or the introduction of altered genes into their genome (134). These animals offer indispensable insights into human diseases, facilitating the exploration of disease mechanisms, experimentation with potential treatments, and assessment of therapeutic effectiveness (135-139). Moreover, they offer crucial insights into changes in structure and imaging techniques in clinical cases (140-158). Preclinical and clinical research collaboratively contribute to innovative therapeutics and personalized medicine (159-164).
This study involved manipulating the gene kat2 in mice to create a knockout (kat2−/−) model, allowing us to observe the behavioral consequences of KAT II deficiency. By focusing on negative valence in emotional domain, memory acquisition, and motor function, we aimed to gain insights into the role of KAT II in these specific behavioral domains in young adult kat2−/− mice. Furthermore, we assess the levels of Trp and its metabolites in three distinct metabolic pathways in both plasma and urine samples, the enzyme activities of Trp metabolism, and the oxidative stress and excitotoxicity indices of KYN metabolites, with the aim of elucidating the Trp metabolic profiles that underlie the behavioral phenotype. This research contributes to our understanding of the genetic factors influencing behaviors related to emotional valence, memory, and motor function and Trp catabolism.
2. Materials and Methods
CRISPR/Cas9 was applied on C57BL/6N and CD1 (ICR) mice to generate knockout kat2-/- mice, and Taqman allelic discrimination was used to prove that the gene had been deleted. The emotional domain, including depression-like and anxiety-like behaviors, was evaluated with the modified forced swim test (FST), tail suspension test (TST), elevated plus maze (EPM), open field (OF) test, and light dark box (LDB) test; the cognitive domain was evaluated with the passive avoidance test (PAT); and the motor domain was evaluated with the OF test. Furthermore, the levels of Trp and its major metabolites, as well as enzyme activities in plasma and urine samples, were determined, and oxidative stress and excitotoxicity indices were calculated.
2.1. Ethical Approval
The Department of Nature Conservation of the Ministry of Agriculture has authorized us to use genetically modified organisms in a closed system of the second security isolation level (TMF/43-20/2015). The import of genetically modified animals has been approved by the Department of Biodiversity and Gene Conservation of the Ministry of Agriculture (BGMF/37-5/2020). In accordance with the guidelines of the 8th Edition of the Guide for the Care and Use of Laboratory Animals, the Use of Animals in Research of the International Association for the Study of Pain, and the directive of the European Economic Community (86/609/ECC), the experiments conducted in this study received ethical approval from two committees. The Scientific Ethics Committee for Animal Research of the Protection of Animals Advisory Board (XI./95/2020) and the Committee of Animal Research at the University of Szeged (I-74-10/2019, I-74-1/2022) both approved the experiments. Furthermore, Directive 2010/63/EU on the protection of animals used for scientific purposes provides guidance for the ethical evaluation of animal use proposals. The directive allows individual institutions to make determinations based on the recommendations of their ethical review committees. These ethical guidelines and regulations ensure that the experiments conducted on animals adhere to the highest standards of animal welfare and scientific integrity. The approval from the Scientific Ethics Committee for Animal Research of the Protection of Animals Advisory Board and the Committee of Animal Research at the University of Szeged demonstrates that the study was conducted in compliance with these ethical principles and regulations.
2.2. Animals
C57BL/6N and CD1 (ICR) mice were purchased from Japan SLC, Inc. (Hamamatsu, Japan) and Charles River Laboratories International, Inc. (Yokohama, Japan), respectively, in order to generate kat2−/− mice utilizing the CRISPR/Cas9 technique. After genetic modifications, breeding, and transport from Japan to Hungary, the animals were housed in groups of 4-5 in polycarbonate cages (530 cm2 floor space) under pathogen-free conditions in the Animal House of the Department of Neurology, University of Szeged, maintained at 24 ± 1°C and 45–55% relative humidity under a 12:12-h light:dark cycle. Throughout the duration of the investigation, mice had unrestricted access to standard rodent food and water. Animal experiments were conducted humanely in accordance with the Regulations for Animal Experiments of Kyushu University and the Fundamental Guidelines for Proper Conduct of Animal Experiments and Related Activities in Academic Research Institutions governed by the Ministry of Education, Culture, Sports, Science, and Technology of Japan, and with the approval of the Institutional Animal Experiment Committees of Kyushu University. After the transport of the animals, the studies complied with the Ethical Codex of Animal Experiments, were approved by the Committee of the Animal Research of the University of Szeged (I-74-10/2019, I-74-1/2022), were authorized by the National Food Chain Safety Office with permission number XI./95/2020, and complied with the guidelines for the Use of Animals in Research of the International Association for the Study of Pain and the directive of the European Parliament (2010/63/EU).
The deletion was introduced into the KATs gene using the CRISPR/Cas9 method. The single guide RNAs (sgRNA) were selected using the CRISPRdirect software. Artificially synthesized the sgRNA were purchased from FASMAC (Atsugi, Japan). The Female C57BL/6N mice were injected with pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG) with a 48-h interval, and mated with male C57BL/6N mice. The fertilized one-cell embryos were collected from the oviducts. Then, 25 ng/µl of the sgRNA and 75 ng/µl Guide-it™ Recombinant Cas9 protein (TaKaRa, Japan) were injected into the cytoplasm of these one-cell-stage embryos. The injected two-cell embryos were then transferred into pseudopregnant ICR mice (Figure 2).
The kat2−/− mouse line expresses a carboxy-terminal truncated polypeptide consisting of the first 47 amino acids of the intact KAT II with a 2-nucleotide deletion (CCDS nucleotide sequence 32-33) in the mRNA.
2.3. DNA Extraction and Sequencing
Genomic DNA of tails collected from mice was extracted using NucleoSpin Tissue (MACHEREY-NAGEL GmbH&Co, KG, Germany). Each targeted fragment around the sgRNA targeting site from the extracted genomic DNA as a part of the KATs genes was amplified with TAKARA Ex Taq (Takara Bio, Japan) and the 1st primers pair and subsequently with 2nd primers pair (Table 1). The polymerase chain reaction (PCR) product was purified with a Fast Gene Gel/PCR Extraction Kit (Nippon Genetics Co., Ltd., Tokyo, Japan), and the PCR products were purified by agarose gel electrophoresis and Monarch Gel Extraction Kit (NEW ENGLAND BioLabs Inc.). Then, the PCR products were sequenced with M-KAT II_2nd_R (Table 1).
2.4. Western Blotting
For Western blotting, tissue extracts from the liver (20 mg) of the knockout and wild-type (WT) mice were prepared by the Total Protein Extraction Kit for Animal Cultured Cells and Tissues (Invent Biotechnologies, Minnesota, USA) according to the manufacturer’s instructions. Subsequently, the tissue extracts were passed through Protein G HP SpinTrapTM (Cytiva, Buckinghamshire, UK) to remove immunoglobulin G. 14 µl of each sample were mixed with 7 µl of 3X SDS Blue Loading Buffer (New England BioLabs) and separated on a 12% SDS-polyacrylamide gel. Subsequently, the protein was transferred to the membranes. The membranes were blocked and incubated with anti-human KAT II rabbit polyclonal antibody (1:500, Invitrogen, Thermo Fisher Scientific, Massachusetts, USA) at room temperature for 2 h, followed by combination with alkaline phosphatase-labeled secondary goat anti-rabbit IgG FC antibody (1:10000, Sigma-Aldrich) at room temperature for 2 h, followed by visualization of dystrophin and utrophin using Western Blue® Stabilized Substrate for Alkaline Phosphatase (Promega, Madison, WI, USA).
2.5. Phenotype Analysis with Modified SHIRPA Test
The RIKEN modified SHIRPA test was conducted to ascertain the comprehensive phenotypic traits of the mutant rodents. The assessment included the evaluation of diverse behaviors and physical attributes such as motion, bowel movements, urination, locomotor activity, startle response, tactile escape, pinna reflex, trunk curling, limb grasping, contact-righting reflex, grip strength, wire maneuver test, corneal reflex, toe pinching, and overall appearance. The animals were also monitored for vocalization, aggression, head bobbing, jumping, circling, retropulsion, grooming, and tail-wagging (165, 166).
2.6. Behavioral Tests
8-week-old male C57BL/6N and kat2-/- mice (n=10-13) were tested. In order to make the results comparable, all behavioral experiments were performed between 8 a.m. and 12 p.m. The animals were transferred to the laboratory, where the measurements were made, one hour before the start of the experiment, thus they had time to acclimatize to the environmental conditions.
2.6.1. Modified Forced Swim Test (FST)
The modified FST was performed as reported previously. The mice were placed individually in a glass cylinder of 12 cm in diameter and 30 cm in height. Water (25±1 °C) was filled to a height of 20 cm. Fresh water was used for each mouse. A 15-min pretest was carried out 24 hours before the 3-min test session. A time-sampling technique was conducted to count the duration of time spent with climbing, swimming, and immobility (167, 168).
2.6.2. Tail Suspension Test (TST)
The mice were placed in a 28x28x23.5 cm wooden box with three side walls and a clip hanging from the top of the box. The animals were suspended by their tails from the base to the middle two-thirds using a clip and allowed to hang for 6 minutes. We measure the duration of immobility. A cotton swab was pre-attached to the clip's interior to prevent the mice's tails from injuring or severely restricting blood circulation. If the animal is able to climb or falls off the clip, it is removed from the experiment and its results are discarded (169, 170).
2.6.3. Elevated Plus Maze (EPM) Test
The animals were positioned in a plus-shaped apparatus with four arms measuring 35x10 cm. Two of the opposite arms are open, while the other two are closed, forming an angle of 90 degrees. The open arms have no side walls, while the closed arms have walls that are 20 cm tall. The entire apparatus is situated 50 cm off the ground. The device is surrounded by a screen that does not display any visual signals. The mouse was placed in the device's center with its nose facing an open arm and allow it to explore for 5 minutes. We measure the time spent in each part (open arms, closed arms, and central part). The experiment was captured on video using a camera and software (EthoVision XT14). Between each animal, the apparatus was disinfected with 70% ethanol and left exposed to the air for 5 minutes (171, 172).
2.6.4. Light Dark Box (LDB) Test
The LDB apparatus is comprised of larger illuminated (2/3 of the box) and smaller dark (1/3 of the box) compartments that are connected by a 5x5 cm door. The length of time a mouse spent in the lighted compartment during the 5-minute session was determined 5 seconds after a mouse was placed in the bright area. After each session, the box was cleaned with 70% ethanol and allowed to air for 5 minutes (172-174).
2.6.5. Passive Avoidance Test (PAT)
Each mouse was individually placed in a box containing two apparatuses with distinct lighting. The animals began in the bright compartment and had 5 minutes to pass through the 5x5 cm door into the dark, smaller portion of the box. As soon as the animals entered the dark compartment, they received a 0.3 mA electroshock through their paws, and the door shut. After 10 seconds, the animals were removed, and the experiment was repeated 24 hours later. Those animals that did not enter the dark area within 5 minutes during the pre-testing phase were omitted from the measurement. The box was cleaned with 70% ethanol and left to air for 5 minutes between mice (175).
2.6.6. Open Field (OF) Test
A standard table lamp illuminated the center of the 48x40 cm OF box, while the Conducta 1.0 system (Experimetria Ltd.) monitored the mouse's movements. Each mouse was placed individually in the center of the box. Ambulation distance, time spent in the center zone, and number of entries to the center zone were measured for 10 minutes. After each session, the box was wiped down with 70% ethanol and allowed to for 5 minutes (176, 177).
2.7. Ultra-High-Performance Liquid Chromatography with Tandem Mass Spectrometry
Trp and its metabolites were measured in plasma and urine using previously published protocols (178, 179) using ultra-high performance liquid chromatography-tandem mass spectrometry (UHPLC-MS/MS). Picolinic acid multiple reaction monitoring (MRM) showed a change from 124.0 to 106.0 over 1.21 minutes, with 75 V acting as the declustering potential and 13 V acting as the collision energy.
2.8. The Enzyme Activities of Trptophan (Trp) Metabolism
The enzyme activities of each Trp metabolism were determined by dividing the concentration of the product by the concentration of the substrate.
2.9. Oxidative Stress and Excitotoxicity Indices
The oxidative stress index was calculated as the ratios of putative prooxidant metabolite 3-HK concentrations to the sums of putative antioxidant metabolite concentrations (KYNA, AA, and XA) (1) (180-182).
Oxidative stress index = [3-Hydroxykynurenine]/{[Kynurenic acic] + [Anthranilic acid] + [Xanthurenic acid]}
The excitotoxicity index is calculated by dividing the concentration of NMDA receptor agonist QA by that of NMDA receptor antagonist KYNA (2)(183-185).
Excitotoxicity index = [Quinolinic acid]/[Kynurenic acid]
3.0. Statistical Analysis
We used IBM SPSS Statistics 28.0.0.0 for the statistical analysis. The Shapiro–Wilk test was used to determine the distribution of data. In addition, we used a Q-Q plot to find out if two sets of data come from the same distribution. Our data followed a normal distribution. One-way ANOVA test was used to evaluate the results of the TST, OF, FST, and EPM followed by the Tamhane post hoc test. Values p <0.05 were considered statistically significant. Our data are reported as means ± SEM for all parameters and groups.
3. Results
3.1. DNA Sequence Analysis and Western Blot
To generate knockout mice of kat2 gene, 25 ng/µl of sgRNA and 75ng/µl Cas9 protein were injected into the cytoplasm of the one-cell-stage embryos. Sequencing analyses with their founder mice showed that various deletions and/or insertions were introduced in the target sequence. One of the founders was selected and established the homozygous mouse line for further analyses. KAT II knockout mouse line expresses a carboxy-terminal truncated polypeptide consisting of the first 47 amino acids of the intact KAT II with 2 nucleotides deletion (CCDS nucleotide sequence 32-33) in the mRNA. Western blotting with antibodies against KAT II revealed that the band with approximately 50-kDa supposed to be KAT II was not detected in the knockout mice, while it was detected in the WT counterparts (Figure 3).
3.2. Phenotype Analysis with SHIRPA Protocol
We did not detect any significant differences between the knockout mice and their wild-type counterparts.
3.3. Behavioral Tests
3.3.1. Forced Swim Test (FST)
3.3.2. Open Field (OF) Test
The ambulation distance of the kat2-/- mice was significantly shorter in the first 10-minute timeframe than that of their WT counterparts (Figure 4, c; Table 2). The number of jumps was significantly fewer in the kat2-/- mice than that of their WT counterparts (Figure 4, d; Table 2). There were significantly fewer entries into the center and corner zones compared to their WT counterparts (Figure 4, e; Table 2).
3.3.3. Other Behavioral Tests
There were no statistically significant distinctions observed between the transgenic mice and their WT counterparts in TST, PAT, EPM test, and LDB test (Table 2).
3.4. Ultra-High-Performance Liquid Chromatography with Tandem Mass Spectrometry
3.5. Enzyme Activities in Trptophan (Trp) Metabolism
The transgenic mice showed significantly lower KATs, kynureninase, MAO, ALDH, and TMO activities and significantly higher kynurenine 3-monooxygenase activity in plasma samples than wild-type mice. In the urine samples, the transgenic mice showed significantly lower KATs, 3-HA, TPH, and TMO activities, and significantly higher TDO/IDOs (KFA), KMO, KYNU, AADC, MAO, and ALDH activities compared to the wild type (Table 4).
2.9. Oxidative Stress and Excitotoxicity Indices
Transgenic mice had higher levels of oxidative stress and excitotoxicity in both plasma and urine than wild-type mice (Table 5).
3. Discussion
Dysregulation of 5-HT metabolism is a key factor in mental symptom development, with attention focused on its imbalance with neurotransmitters like dopamine, norepinephrine, and biosystems such as substance P (186-189). Alterations in 5-HT precursor Trp metabolism are noted in mental illnesses, but their connection with the Trp-KYN metabolic system remains poorly understood (190-192). Growing evidence suggests that the gut microbial indole pyruvate pathway can influence the microbiome-gut-brain axis, implying that intestinal tryptophan metabolism may play a significant role in psychological health. The microbiome-gut-brain axis is responsible for regulating mood, cognition, stress response, and behavior. As a result, the gut-microbial indole pyruvate pathway can influence the microbiome-gut-brain axis by controlling the production and availability of neurotransmitters, hormones, cytokines, and bioactive metabolites involved in neuropsychiatric conditions.
KATs are cytosolic and mitochondrial aminotransferases that convert KYN to KYNA (69, 193-195). The mitochondrial isoform KAT II exclusively influences cellular bioenergetics due to its exclusive location in the mitochondria (101, 187). CRISPR/Cas9 was employed to knock out the kat2 gene, creating kat2−/− mice. This study aimed to examine the negative emotional aspects and evaluate any behavioral alterations caused by the knockout of the kat2 gene in young adults aged 8 weeks. kat2−/− mice, studied in 8-week-old adults, induces a unique depression-like phenotype marked by increased immobility in FST, likely linked to serotonergic pathways. TST didn't show significant differences, possibly due to FST conditioning. PAT indicated despair-induced depression rather than average-conditioned memory. Anxiety-like behaviors (EPM, LDB, and MB tests) showed no difference, but the OF test revealed shorter ambulation distance, fewer jumping counts, and fewer entries into both center field and corners, suggesting a la belle indifference-like trait. kat2−/− mice exhibited despair-based depression-like behavior without anxiety-like traits, demonstrating motor deficits. The study suggests the kat2 gene deletion leads to a PTSD-like phenotype, including a la belle indifference trait, indicative of complex PTSD with emotional dysregulation (196-198).
The gene knockout significantly alters Trp metabolism in both 5-HT, KYN, and indole pathways in plasma and urine. A major 5-HT metabolite, 5-HIAA, is markedly reduced, possibly explained by scarce mitochondrial enzyme activity. Lower levels of KYNA and antioxidant KYNs indicate decreased production in peripheral tissues of kat2−/− mice. Conversely, 3-HK is significantly elevated. The levels of the gut microbial metabolite IAA, an antioxidant and anti-inflammatory molecule, were reduced. Furthermore, gene knockout affects enzyme activity, puts organisms under oxidative stress, and imposes high excitoneurotoxicity. The study demonstrates that the deletion of the kat2 gene leads to a specific set of characteristics, including behavior similar to depression, impaired motor function, decreased levels of KYNA, and a change in the way tryptophan is metabolized towards the KYN pathway. This phenotype exhibits similarities to PTSD in humans, potentially indicating the presence of complex PTSD due to the observed belle indifference-like trait.
The amygdala encodes and stores fear memory after receiving sensory input from the thalamus, which also consolidates and retrieves memories from the initial stimuli that induce fear (199-201). Fear memory is associated with the release of stress hormones such as adrenaline and cortisol, which stimulate the sympathetic nervous system and the hypothalamic-pituitary-adrenal axis (39, 202-205). This study does not show evidence of fear memory acquisition. In contrast, the encoding and storage of memories associated with despair occur in the prefrontal cortex, which plays a crucial role in the cognitive and emotional processing of negative experiences (206). Recalling distressing memories, triggered by cues linked to the initial negative encounter, results in the disruption of serotonin, norepinephrine, and dopamine regulation. Although fear and despair memories have similarities in terms of encoding and retrieval processes, they are associated with different brain regions, neurotransmitters, and neural circuits (207, 208).
Furthermore, despair memory and despair experience differ. The latter pertains to an instantaneous, personal feeling of despair or hopelessness, prompted by present circumstances, as opposed to a remembrance of past experiences (209). Despair memory involves the consolidation and retrieval of long-term memories, influenced by stress and emotion (210). In contrast, a despair experience entails immediate emotional responses influenced by factors like cognitive assessments, environmental cues, and physiological states (211). Additionally, la belle indifference arises from a discrepancy between cognitive and emotional symptom processing, including altered emotional processing in the amygdala and insula, changed self-awareness in the medial prefrontal cortex, and adjusted activity in the somatosensory cortex influenced by dopamine and serotonin (212). Thus, kat2−/− mice show more despair-based depression-like behavior involving a change in 5-HT metabolism.
Approximately 60% of individuals on antidepressants, including SSRIs, for two months experience a 50% reduction in depression symptoms (213). The observation aligns with the monoamine hypothesis, suggesting depression's pathogenesis is linked to low 5-HT levels. Transgenic models are used to study 5-HT dysmetabolism behaviors, with a focus on the Tph gene, which encodes tryptophan hydroxylase, a key enzyme in 5-HT synthesis (214). Preclinical studies found normal 5-HT levels with no behavioral changes in Tph1−/− mice, while Tph2−/− mice's behaviors are inconclusive (215, 216). The knock-in mice of the TPH2 variant (R439H) showed depression-like behavior in TST (217). Intriguingly, Tph1/Tph2−/− mice exhibited contrasting behaviors: antidepressant-like in FST, depressive in TST, and anxious in the MB test, accompanied by low 5-HT levels in the brain and periphery (218). 5-HT1A receptor knockout (5-HT1AR−/−) mice display heightened fear memory to contextual cues, suggesting a role for 5-HT receptors in PTSD-like phenotype (219). 5-HT 2C receptor knockout 5-HT2CR−/− mice attenuates fear responses in contextual or cued but not compound context-cue fear conditioning (220). Knockout of the 5-HTT gene in mice (5-HTT−/−) leads to impaired stress response, fear extinction, and abnormal corticolimbic structure (221).
Over 90% of 5-HT precursor Trp undergoes catabolism in the Trp-KYN metabolic system, generating a variety of bioactive molecules including prooxidants, antioxidants, inflammation suppressants, neurotoxins, neuroprotectants, and/or immunomodulators (222). Growing evidence indicates disrupted KYN metabolism in MDD, bipolar disorder, and SCZ (223-225). Earlier, KYN metabolites were suggested to be either neuroprotective or neurotoxic (226). However, increasing evidence suggests that KYN metabolites exhibit versatile actions, potentially influenced by concentrations and the microenvironment (227). Previously, cognitive and motor functions of 129/SvEv kat2−/− mice were reported. These transgenic mice exhibited transient hyperlocomotive activity and motor coordination issues at postnatal day 21. However, from postnatal day 17 to 26, they demonstrated notable improvements in cognitive functions, particularly in object exploration and recognition tasks in PAT and T-maze tests (228, 229).
Other biosystems play an important role in the pathogenesis of PTSD include dopaminergic and gamma-aminobutyric acid (GABA)ergic system and cannabinoids. Catechol-O-methyltransferase (COMT) degrades dopamine. COMT−/− mice exhibited an increased response to repeated stress exposures (230). Glutamic acid decarboxylase (GAD) synthesizes GABA (231). GAD6−/− mice shows increased generalized fear and impaired extinction of cued fear (232). GABA receptor subunit B1a knockout GABAB1a−/− mice showed a generalization of conditioned fear to nonconditioned stimuli (233). Cannabinoid 1 receptor (CBIR) knockout CB1R−/− mice showed an increased response to repeated stress exposures (234).
The potential of this study is to characterize the negative valence of emotional domain in context with aversive-conditioned memory and despair experience in the young adult (8 week) of kat2−/− mice. The findings complement the previous studies of kat2−/− mice in the early adolescence (2 and 1/2 to 4 week) to reveal that, toward the adulthood, there is a dynamic change in emotional susceptibility and motor function derived from despair experience in adjunct to Trp metabolism. Furthermore, urinary Trp metabolite levels are generally consistent with plasma levels, suggesting that urinary samples may serve as non-invasive biomarkers for Trp metabolism status. This study may shed new light on the deletion of the kat2 gene as a new avenue toward understanding a KYN metabolite as an oxidative stressor, a potential barrier between aversive-conditioned memory and despair experience, a distinction between memory and experience, their mechanism for the formation of intrusive memories, and the pathogenesis of PTSD. The ultimate goal is to probe a potential interventionable stage in age where the progression of PTSD is preventable and to identify targets which drugs or psychotherapy can relieve symptoms of PTSD. The greatest challenge lies in preclinical animal models that are difficult to simulate and interpolate to mental illnesses to achieve high model validity.
This study suggests that behavioral sampling in rodents can distinguish between fear-, memory-, and despair-based depression-like behavior associated with Trp metabolism gene deletions. Further research incorporating neurochemical, neurogenetic, and electrophysiological biomarkers may reinforce this finding. Additionally, using inhibitory RNA or antisense RNA on neurotransmitters in specific brain regions could elucidate the precise mechanisms underlying emotional behaviors. Preclinical research drives advances in clinical applications like precision medicine and drug discovery. The study acknowledges weaknesses, noting distinctions in interpreting animal behaviors and drug responses compared to humans. Recent perspectives consider depression-like behavior in FST as related to different stages of stress-coping behaviors (235). Consequently, Translational research has limitations that necessitate careful interpretation. This study employed animal models with standard protocols, focusing on the negative valence of the emotional domain and motor function in kat2−/− mice. Further exploration with diverse models such as sucrose preference tests, fear condition tests, and those using non-standard protocols is crucial for a more accurate characterization of kat2−/− mouse behavior. Notably, the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition, emphasizes four symptom clusters in PTSD diagnosis (236-238). The transgenic mice in this study did not exhibit signs related to negative cognitions and mood, and arousal state and reactivity were not investigated.
5. Conclusions
Psychiatric disorders, including PTSD, have a significant impact on memory and emotion, and disruptions in 5-HT metabolism have been associated with these disorders. The Trp-KYN metabolic pathway plays a crucial role in metabolizing over 95% of the serotonin precursor Trp. To investigate the effects of gene deletion on negative valence in emotion, memory, and motor function, transgenic kat2−/− mice were created and compared to WT mice. The kat2−/− mice exhibited depression-like behavior characterized by despair experiences, diminished motor functions, and la belle indifference-like characteristics without anxiety-like behavior. This study provides insights into the negative valence of the emotional domain in the context of aversive-conditioned memory and despair experiences in 8-week-old kat2−/− mice. Understanding the complex interplay between memory, emotion, and genetic factors is crucial for advancing our knowledge of psychiatric disorders (239, 240). By elucidating the specific effects of gene deletion on negative valence and related behaviors, this research contributes to our understanding of the underlying mechanisms and potential interventions.
Author Contributions
Conceptualization, Á.S., E.O., L.V., and M.T.; methodology, Á.S., Z.G., E.S., M.S., D.M., K.T., K.O., H.I., S.Y., and M.T.; software, Á.S., E.S., M.S., K.O.; validation, Á.S., Z.G., E.S., M.S., K.T., K.O., and M.T.; formal analysis, Á.S., Z.G., E.S., M.S., K.O., and M.T.; investigation, Á.S., Z.G., E.S., M.S., D.M., K.T., K.O., and M.T.; resources, Á.S. and E.O.; data curation, Á.S., Z.G., E.S., M.S., K.O.; writing—original draft preparation, Á.S., Z.G., M.T.; writing—review and editing, Á.S., Z.G., E.S., M.S., D.M., J.T., E.O., L.V., and M.T.; visualization, Á.S., K.O.; supervision, J.T., E.O., L.V., and M.T.; project administration, J.T., E.O., and L.V; funding acquisition, J.T., E.O., L.V., and M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research, Development, and Innovation Office—NKFIH K138125, SZTE SZAOK-KKA No:2022/5S729, the HUN-REN Hungarian Research Network, JSPS Joint Research Projects under Bilateral Programs Grant Number JPJSBP120203803, and University of Szeged Open Access Fund, Grant ID: 6846.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of the Scientific Ethics Committee for Animal Research of the Protection of Animals Advisory Board (XI./95/2020) and the Committee of Animal Research at the University of Szeged (I-74-1/2022).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors express their gratitude to Dr. Nikolett Nánási for her chemical analysis. The figures are created with biorender.com.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 5-HT: serotonin |
| AD: Alzheimer’s disease |
| CBIR: cannabinoid 1 receptor |
| CNS: central nervous system |
| COMT: catechol-O-methyltransferase |
| EPM: elevated plus maze |
| FST: forced swim test |
| GABA: gamma-aminobutyric acid |
| GAD: glutamic acid decarboxylase |
| hCG: human chorionic gonadotropin |
| IAA: indole-3-acetic acid |
| ISN: indoxyl sulfate |
| KAT II: α-aminoadipate aminotransferase/kynurenine aminotransferase II |
| KATs: kynurenine aminotransferases |
| KYN: kynurenine |
| KYNA: kynurenic acid |
| LDB: light dark box |
| MDD: major depressive disorder |
| MRM: multiple reaction monitoring |
| OF: open field |
| PAT: passive avoidance test |
| PCR: polymerase chain reaction |
| PMSG: pregnant mare serum gonadotropin |
| PTSD: posttraumatic stress disorder |
| SCZ: schizophrenia |
| sgRNA: single guide RNA |
| SSRI: selective serotonin reuptake inhibitors |
| TNA tryptophanase |
| Trp: tryptophan |
| TST: tail suspension test |
| UHPLC-MS: ultra-high performance liquid chromatography-tandem mass spectrometry |
| WT: wild-type |
References
- Tyng CM, Amin HU, Saad MN, Malik AS (2017): The influences of emotion on learning and memory. Frontiers in psychology.1454.
- Battaglia S, Garofalo S, di Pellegrino G, Starita F (2020): Revaluing the role of vmPFC in the acquisition of Pavlovian threat conditioning in humans. Journal of Neuroscience. 40:8491-8500.
- Battaglia S, Harrison BJ, Fullana MA (2022): Does the human ventromedial prefrontal cortex support fear learning, fear extinction or both? A commentary on subregional contributions. Molecular Psychiatry. 27:784-786.
- Sumsuzzman DM, Choi J, Jin Y, Hong Y (2021): Neurocognitive effects of melatonin treatment in healthy adults and individuals with Alzheimer’s disease and insomnia: a systematic review and meta-analysis of randomized controlled trials. Neuroscience & Biobehavioral Reviews. 127:459-473.
- Borgomaneri S, Battaglia S, Sciamanna G, Tortora F, Laricchiuta D (2021): Memories are not written in stone: Re-writing fear memories by means of non-invasive brain stimulation and optogenetic manipulations. Neuroscience & Biobehavioral Reviews. 127:334-352.
- Hasson U, Chen J, Honey CJ (2015): Hierarchical process memory: memory as an integral component of information processing. Trends in cognitive sciences. 19:304-313.
- Clewett D, Sakaki M, Nielsen S, Petzinger G, Mather M (2017): Noradrenergic mechanisms of arousal’s bidirectional effects on episodic memory. Neurobiology of learning and memory. 137:1-14.
- Battaglia S, Nazzi C, Thayer J (2023): Fear-induced bradycardia in mental disorders: Foundations, current advances, future perspectives. Neuroscience & Biobehavioral Reviews.105163.
- Battaglia S, Di Fazio C, Vicario CM, Avenanti A (2023): Neuropharmacological modulation of N-methyl-D-aspartate, noradrenaline and endocannabinoid receptors in fear extinction learning: Synaptic transmission and plasticity. International Journal of Molecular Sciences. 24:5926. [CrossRef]
- Hayes JP, VanElzakker MB, Shin LM (2012): Emotion and cognition interactions in PTSD: a review of neurocognitive and neuroimaging studies. Frontiers in integrative neuroscience. 6:89.
- Dillon DG, Pizzagalli DA (2018): Mechanisms of memory disruption in depression. Trends in neurosciences. 41:137-149.
- Mathews A, MacLeod C (2005): Cognitive vulnerability to emotional disorders. Annu Rev Clin Psychol. 1:167-195.
- Hasebe K, Kendig MD, Morris MJ (2021): Mechanisms underlying the cognitive and behavioural effects of maternal obesity. Nutrients. 13:240.
- Junges VM, Closs VE, Nogueira GM, Gottlieb MGV (2018): Crosstalk between Gut Microbiota and Central Nervous System: A Focus on Alzheimer's Disease. Curr Alzheimer Res. 15:1179-1190.
- Guzmán-Vélez E, Feinstein JS, Tranel D (2014): Feelings without memory in Alzheimer disease. Cognitive and behavioral neurology. 27:117.
- Mack J, Marsh L (2017): Parkinson’s disease: cognitive impairment. Focus. 15:42-54.
- Mckee AC, Daneshvar DH (2015): The neuropathology of traumatic brain injury. Handbook of clinical neurology. 127:45-66.
- Li M, Feng L, Liu X, Zhang M, Fu B, Wang G, et al. (2018): Emotional working memory in patients with major depressive disorder. Journal of International Medical Research. 46:1734-1746. [CrossRef]
- Samuelson KW (2011): Post-traumatic stress disorder and declarative memory functioning: a review. Dialogues in clinical neuroscience. 13:346-351.
- Dere E, Pause BM, Pietrowsky R (2010): Emotion and episodic memory in neuropsychiatric disorders. Behavioural brain research. 215:162-171.
- Berger M, Gray JA, Roth BL (2009): The expanded biology of serotonin. Annual review of medicine. 60:355-366.
- Ressler KJ, Nemeroff CB (2000): Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depress Anxiety. 12 Suppl 1:2-19.
- Bacqué-Cazenave J, Bharatiya R, Barrière G, Delbecque JP, Bouguiyoud N, Di Giovanni G, et al. (2020): Serotonin in Animal Cognition and Behavior. Int J Mol Sci. 21. [CrossRef]
- Meneses A, Liy-Salmeron G (2012): Serotonin and emotion, learning and memory. Rev Neurosci. 23:543-553.
- Švob Štrac D, Pivac N, Mück-Šeler D (2016): The serotonergic system and cognitive function. Translational neuroscience. 7:35-49.
- Schmitt JA, Wingen M, Ramaekers JG, Evers EA, Riedel WJ (2006): Serotonin and human cognitive performance. Curr Pharm Des. 12:2473-2486.
- Buhot MC, Martin S, Segu L (2000): Role of serotonin in memory impairment. Ann Med. 32:210-221.
- Battaglia S, Cardellicchio P, Di Fazio C, Nazzi C, Fracasso A, Borgomaneri S (2022): Stopping in (e) motion: Reactive action inhibition when facing valence-independent emotional stimuli. Frontiers in Behavioral Neuroscience. 16:998714.
- Cowen P, Sherwood AC (2013): The role of serotonin in cognitive function: evidence from recent studies and implications for understanding depression. Journal of psychopharmacology. 27:575-583.
- Battaglia S, Cardellicchio P, Di Fazio C, Nazzi C, Fracasso A, Borgomaneri S (2022): The influence of vicarious fear-learning in “infecting” reactive action inhibition. Frontiers in Behavioral Neuroscience. 16:946263.
- Battaglia S, Thayer JF (2022): Functional interplay between central and autonomic nervous systems in human fear conditioning. Trends in Neurosciences.
- Battaglia S, Orsolini S, Borgomaneri S, Barbieri R, Diciotti S, di Pellegrino G (2022): Characterizing cardiac autonomic dynamics of fear learning in humans. Psychophysiology. 59:e14122.
- Di Gregorio F, La Porta F, Petrone V, Battaglia S, Orlandi S, Ippolito G, et al. (2022): Accuracy of EEG biomarkers in the detection of clinical outcome in disorders of consciousness after severe acquired brain injury: preliminary results of a pilot study using a machine learning approach. Biomedicines. 10:1897. [CrossRef]
- Borgomaneri S, Battaglia S, Avenanti A, di Pellegrino G (2021): Don't hurt me no more: State-dependent transcranial magnetic stimulation for the treatment of specific phobia. Journal of affective disorders. 286:78-79.
- Khalil R, Godde B, Karim AA (2019): The link between creativity, cognition, and creative drives and underlying neural mechanisms. Frontiers in neural circuits. 13:18. [CrossRef]
- Borgomaneri S, Battaglia S, Garofalo S, Tortora F, Avenanti A, di Pellegrino G (2020): State-dependent TMS over prefrontal cortex disrupts fear-memory reconsolidation and prevents the return of fear. Current Biology. 30:3672-3679. e3674.
- Battaglia S, Garofalo S, di Pellegrino G (2018): Context-dependent extinction of threat memories: Influences of healthy aging. Scientific reports. 8:12592.
- Albert PR, Vahid-Ansari F, Luckhart C (2014): Serotonin-prefrontal cortical circuitry in anxiety and depression phenotypes: pivotal role of pre-and post-synaptic 5-HT1A receptor expression. Frontiers in behavioral neuroscience. 8:199.
- Tortora F, Hadipour AL, Battaglia S, Falzone A, Avenanti A, Vicario CM (2023): The role of Serotonin in fear learning and memory: a systematic review of human studies. Brain Sciences. 13:1197.
- Brewerton TD (1995): Toward a unified theory of serotonin dysregulation in eating and related disorders. Psychoneuroendocrinology. 20:561-590.
- Savitz J, Lucki I, Drevets WC (2009): 5-HT(1A) receptor function in major depressive disorder. Prog Neurobiol. 88:17-31. [CrossRef]
- Nutt DJ (2001): Neurobiological mechanisms in generalized anxiety disorder. J Clin Psychiatry. 62 Suppl 11:22-27; discussion 28.
- Steiger H (2004): Eating disorders and the serotonin connection: state, trait and developmental effects. J Psychiatry Neurosci. 29:20-29.
- Meltzer HY, Li Z, Kaneda Y, Ichikawa J (2003): Serotonin receptors: their key role in drugs to treat schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 27:1159-1172.
- Kelmendi B, Adams TG, Yarnell S, Southwick S, Abdallah CG, Krystal JH (2016): PTSD: from neurobiology to pharmacological treatments. Eur J Psychotraumatol. 7:31858.
- Chu A, Wadhwa R (2022): Selective Serotonin Reuptake Inhibitors.[Updated 2022 May 8]. StatPearls [Internet] Treasure Island (FL): StatPearls Publishing.
- Alvares GA, Quintana DS, Hickie IB, Guastella AJ (2016): Autonomic nervous system dysfunction in psychiatric disorders and the impact of psychotropic medications: a systematic review and meta-analysis. J Psychiatry Neurosci. 41:89-104.
- Stahl SM, Lee-Zimmerman C, Cartwright S, Morrissette DA (2013): Serotonergic drugs for depression and beyond. Curr Drug Targets. 14:578-585.
- Teleanu RI, Niculescu A-G, Roza E, Vladâcenco O, Grumezescu AM, Teleanu DM (2022): Neurotransmitters—Key Factors in Neurological and Neurodegenerative Disorders of the Central Nervous System. International Journal of Molecular Sciences. 23:5954.
- Liu Y, Zhao J, Guo W (2018): Emotional Roles of Mono-Aminergic Neurotransmitters in Major Depressive Disorder and Anxiety Disorders. Front Psychol. 9:2201.
- Nutt DJ (2006): The role of dopamine and norepinephrine in depression and antidepressant treatment. J Clin Psychiatry. 67 Suppl 6:3-8.
- Muneer A (2020): Kynurenine pathway of tryptophan metabolism in neuropsychiatric disorders: pathophysiologic and therapeutic considerations. Clinical Psychopharmacology and Neuroscience. 18:507.
- Tanaka M, Török N, Tóth F, Szabó Á, Vécsei L (2021): Co-players in chronic pain: neuroinflammation and the tryptophan-kynurenine metabolic pathway. Biomedicines. 9:897.
- Huang Y, Zhao M, Chen X, Zhang R, Le A, Hong M, et al. (2023): Tryptophan Metabolism in Central Nervous System Diseases: Pathophysiology and Potential Therapeutic Strategies. Aging and Disease. 14:858. [CrossRef]
- Polyák H, Galla Z, Nánási N, Cseh EK, Rajda C, Veres G, et al. (2023): The tryptophan-kynurenine metabolic system is suppressed in cuprizone-induced model of demyelination simulating progressive multiple sclerosis. Biomedicines. 11:945. [CrossRef]
- Hubková B, Valko-Rokytovská M, Čižmárová B, Zábavníková M, Mareková M, Birková A (2022): Tryptophan: Its metabolism along the kynurenine, serotonin, and indole pathway in malignant melanoma. International Journal of Molecular Sciences. 23:9160.
- Mor A, Tankiewicz-Kwedlo A, Krupa A, Pawlak D (2021): Role of kynurenine pathway in oxidative stress during neurodegenerative disorders. Cells. 10:1603.
- Bosi A, Banfi D, Bistoletti M, Giaroni C, Baj A (2020): Tryptophan metabolites along the microbiota-gut-brain axis: an interkingdom communication system influencing the gut in health and disease. International Journal of Tryptophan Research. 13:1178646920928984.
- Spekker E, Tanaka M, Szabó Á, Vécsei L (2021): Neurogenic inflammation: The participant in migraine and recent advancements in translational research. Biomedicines. 10:76.
- Réus GZ, Jansen K, Titus S, Carvalho AF, Gabbay V, Quevedo J (2015): Kynurenine pathway dysfunction in the pathophysiology and treatment of depression: Evidences from animal and human studies. Journal of psychiatric research. 68:316-328.
- Jones AW (2023): Brief history of the alcohol biomarkers CDT, EtG, EtS, 5-HTOL, and PEth. Drug Testing and Analysis.
- Höglund E, Øverli Ø, Winberg S (2019): Tryptophan metabolic pathways and brain serotonergic activity: a comparative review. Frontiers in endocrinology.158.
- Jayamohananan H, Kumar MKM, Aneesh T (2019): 5-HIAA as a potential biological marker for neurological and psychiatric disorders. Advanced pharmaceutical bulletin. 9:374.
- Hardeland R (2010): Melatonin metabolism in the central nervous system. Current neuropharmacology. 8:168-181.
- Ishidoh K, Kamemura N, Imagawa T, Oda M, Sakurai J, Katunuma N (2010): Quinolinate phosphoribosyl transferase, a key enzyme in de novo NAD+ synthesis, suppresses spontaneous cell death by inhibiting overproduction of active-caspase-3. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research. 1803:527-533. [CrossRef]
- Raffaelli N, Sorci L, Amici A, Emanuelli M, Mazzola F, Magni G (2002): Identification of a novel human nicotinamide mononucleotide adenylyltransferase. Biochemical and biophysical research communications. 297:835-840.
- Jauch R, Humm A, Huber R, Wahl MC (2005): Structures of Escherichia coli NAD synthetase with substrates and products reveal mechanistic rearrangements. Journal of Biological Chemistry. 280:15131-15140.
- Tanaka M, Tóth F, Polyák H, Szabó Á, Mándi Y, Vécsei L (2021): Immune influencers in action: metabolites and enzymes of the tryptophan-kynurenine metabolic pathway. Biomedicines. 9:734.
- Tanaka M, Szabó Á, Spekker E, Polyák H, Tóth F, Vécsei L (2022): Mitochondrial impairment: A common motif in neuropsychiatric presentation? The link to the tryptophan–kynurenine metabolic system. Cells. 11:2607.
- Fila M, Chojnacki J, Pawlowska E, Szczepanska J, Chojnacki C, Blasiak J (2021): Kynurenine pathway of tryptophan metabolism in migraine and functional gastrointestinal disorders. International Journal of Molecular Sciences. 22:10134.
- Wirthgen E, Hoeflich A, Rebl A, Günther J (2018): Kynurenic acid: the Janus-faced role of an immunomodulatory tryptophan metabolite and its link to pathological conditions. Frontiers in immunology. 8:1957.
- Takahashi H, Price J (1958): Dehydroxylation of xanthurenic acid to 8-hydroxyquinaldic acid. Journal of Biological Chemistry. 233:150-153.
- Walczak K, Langner E, Szalast K, Makuch-Kocka A, Pożarowski P, Plech T (2020): A tryptophan metabolite, 8-hydroxyquinaldic acid, exerts antiproliferative and anti-migratory effects on colorectal cancer cells. Molecules. 25:1655. [CrossRef]
- Espi M, Koppe L, Fouque D, Thaunat O (2020): Chronic kidney disease-associated immune dysfunctions: impact of protein-bound uremic retention solutes on immune cells. Toxins. 12:300.
- Mishra P, Kaur S, Sharma AN, Jolly RS (2016): Characterization of an indole-3-acetamide hydrolase from Alcaligenes faecalis subsp. parafaecalis and its application in efficient preparation of both enantiomers of chiral building block 2, 3-dihydro-1, 4-benzodioxin-2-carboxylic acid. PloS one. 11:e0159009.
- Ye X, Li H, Anjum K, Zhong X, Miao S, Zheng G, et al. (2022): Dual role of indoles derived from intestinal microbiota on human health. Frontiers in Immunology. 13:903526. [CrossRef]
- Mousseau DD (1993): Tryptamine: a metabolite of tryptophan implicated in various neuropsychiatric disorders. Metabolic brain disease. 8:1-44.
- Brydges CR, Fiehn O, Mayberg HS, Schreiber H, Dehkordi SM, Bhattacharyya S, et al. (2021): Indoxyl sulfate, a gut microbiome-derived uremic toxin, is associated with psychic anxiety and its functional magnetic resonance imaging-based neurologic signature. Scientific reports. 11:21011. [CrossRef]
- Hou Y, Li J, Ying S (2023): Tryptophan Metabolism and Gut Microbiota: A Novel Regulatory Axis Integrating the Microbiome, Immunity, and Cancer. Metabolites. 13:1166.
- Li X, Zhang B, Hu Y, Zhao Y (2021): New insights into gut-bacteria-derived indole and its derivatives in intestinal and liver diseases. Frontiers in Pharmacology. 12:769501.
- Gao J, Xu K, Liu H, Liu G, Bai M, Peng C, et al. (2018): Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Frontiers in cellular and infection microbiology. 8:13. [CrossRef]
- Ranhotra HS (2023): Discrete interplay of gut microbiota L-tryptophan metabolites in host biology and disease. Molecular and Cellular Biochemistry.1-18.
- Hyland NP, Cavanaugh CR, Hornby PJ (2022): Emerging effects of tryptophan pathway metabolites and intestinal microbiota on metabolism and intestinal function. Amino Acids. 54:57-70.
- Kumar P, Lee JH, Lee J (2021): Diverse roles of microbial indole compounds in eukaryotic systems. Biological Reviews. 96:2522-2545.
- Su X, Gao Y, Yang R (2022): Gut microbiota-derived tryptophan metabolites maintain gut and systemic homeostasis. Cells. 11:2296.
- Gasaly N, De Vos P, Hermoso MA (2021): Impact of bacterial metabolites on gut barrier function and host immunity: a focus on bacterial metabolism and its relevance for intestinal inflammation. Frontiers in immunology. 12:658354.
- Hubbard TD, Murray IA, Perdew GH (2015): Indole and tryptophan metabolism: endogenous and dietary routes to Ah receptor activation. Drug Metabolism and Disposition. 43:1522-1535.
- Fu Y, Lyu J, Wang S (2023): The role of intestinal microbes on intestinal barrier function and host immunity from a metabolite perspective. Frontiers in Immunology. 14.
- Madella AM, Van Bergenhenegouwen J, Garssen J, Masereeuw R, Overbeek SA (2022): Microbial-derived tryptophan catabolites, kidney disease and gut inflammation. Toxins. 14:645.
- Sun L-J, Li J-N, Nie Y-Z (2020): Gut hormones in microbiota-gut-brain cross-talk. Chinese medical journal. 133:826-833.
- Mittal R, Debs LH, Patel AP, Nguyen D, Patel K, O'Connor G, et al. (2017): Neurotransmitters: The critical modulators regulating gut–brain axis. Journal of cellular physiology. 232:2359-2372.
- Tran SM-S, Mohajeri MH (2021): The role of gut bacterial metabolites in brain development, aging and disease. Nutrients. 13:732.
- Han Y, Wang B, Gao H, He C, Hua R, Liang C, et al. (2022): Vagus nerve and underlying impact on the gut microbiota-brain Axis in behavior and neurodegenerative diseases. Journal of inflammation research.6213-6230. [CrossRef]
- Gershon MD, Margolis KG (2021): The gut, its microbiome, and the brain: connections and communications. The Journal of clinical investigation. 131. [CrossRef]
- Caspani G, Swann J (2019): Small talk: microbial metabolites involved in the signaling from microbiota to brain. Current opinion in pharmacology. 48:99-106.
- Zhou Y, Chen Y, He H, Peng M, Zeng M, Sun H (2023): The role of the indoles in microbiota-gut-brain axis and potential therapeutic targets: A focus on human neurological and neuropsychiatric diseases. Neuropharmacology.109690.
- Caspani G, Kennedy S, Foster JA, Swann J (2019): Gut microbial metabolites in depression: understanding the biochemical mechanisms. Microbial Cell. 6:454.
- Chernikova MA, Flores GD, Kilroy E, Labus JS, Mayer EA, Aziz-Zadeh L (2021): The brain-gut-microbiome system: pathways and implications for autism spectrum disorder. Nutrients. 13:4497.
- Ju S, Shin Y, Han S, Kwon J, Choi TG, Kang I, Kim SS (2023): The Gut–Brain Axis in Schizophrenia: The Implications of the Gut Microbiome and SCFA Production. Nutrients. 15:4391.
- Pappolla MA, Perry G, Fang X, Zagorski M, Sambamurti K, Poeggeler B (2021): Indoles as essential mediators in the gut-brain axis. Their role in Alzheimer's disease. Neurobiology of disease. 156:105403.
- Tanaka M, Bohár Z, Martos D, Telegdy G, Vécsei L (2020): Antidepressant-like effects of kynurenic acid in a modified forced swim test. Pharmacological Reports. 72:449-455.
- Martos D, Tuka B, Tanaka M, Vécsei L, Telegdy G (2022): Memory enhancement with kynurenic acid and its mechanisms in neurotransmission. Biomedicines. 10:849.
- Goh DL, Patel A, Thomas GH, Salomons GS, Schor DS, Jakobs C, Geraghty MT (2002): Characterization of the human gene encoding α-aminoadipate aminotransferase (AADAT). Molecular genetics and metabolism. 76:172-180.
- Modoux M, Rolhion N, Mani S, Sokol H (2021): Tryptophan metabolism as a pharmacological target. Trends in Pharmacological Sciences. 42:60-73.
- Palotai M, Telegdy G, Tanaka M, Bagosi Z, Jászberényi M (2014): Neuropeptide AF induces anxiety-like and antidepressant-like behavior in mice. Behavioural brain research. 274:264-269.
- Tanaka M, Telegdy G (2008): Involvement of adrenergic and serotonergic receptors in antidepressant-like effect of urocortin 3 in a modified forced swimming test in mice. Brain research bulletin. 77:301-305.
- Tanaka M, Schally A, Telegdy G (2012): Neurotransmission of the antidepressant-like effects of the growth hormone-releasing hormone antagonist MZ-4-71. Behavioural brain research. 228:388-391.
- Telegdy G, Tanaka M, Schally AV (2011): Effects of the growth hormone-releasing hormone (GH-RH) antagonist on brain functions in mice. Behavioural brain research. 224:155-158.
- Tanaka M, Csabafi K, Telegdy G (2013): Neurotransmissions of antidepressant-like effects of kisspeptin-13. Regulatory peptides. 180:1-4.
- Rákosi K, Masaru T, Zarándi M, Telegdy G, Tóth GK (2014): Short analogs and mimetics of human urocortin 3 display antidepressant effects in vivo. Peptides. 62:59-66.
- Tanaka M, Vécsei L (2022): Editorial of special issue ‘dissecting neurological and neuropsychiatric diseases: Neurodegeneration and neuroprotection’. MDPI, pp 6991.
- Tran KN, Nguyen NPK, Nguyen LTH, Shin H-M, Yang I-J (2023): Screening for Neuroprotective and Rapid Antidepressant-like Effects of 20 Essential Oils. Biomedicines. 11:1248.
- Tanaka M, Szabó Á, Vécsei L (2022): Integrating armchair, bench, and bedside research for behavioral neurology and neuropsychiatry. MDPI, pp 2999. [CrossRef]
- Tanaka M, Kádár K, Tóth G, Telegdy G (2011): Antidepressant-like effects of urocortin 3 fragments. Brain Research Bulletin. 84:414-418.
- Telegdy G, Adamik A, Tanaka M, Schally A (2010): Effects of the LHRH antagonist Cetrorelix on affective and cognitive functions in rats. Regulatory peptides. 159:142-147.
- Baliellas DE, Barros MP, Vardaris CV, Guariroba M, Poppe SC, Martins MF, et al. (2023): Propentofylline Improves Thiol-Based Antioxidant Defenses and Limits Lipid Peroxidation following Gliotoxic Injury in the Rat Brainstem. Biomedicines. 11:1652. [CrossRef]
- Montanari M, Imbriani P, Bonsi P, Martella G, Peppe A (2023): Beyond the Microbiota: Understanding the Role of the Enteric Nervous System in Parkinson’s Disease from Mice to Human. Biomedicines. 11:1560.
- Garifulin R, Davleeva M, Izmailov A, Fadeev F, Markosyan V, Shevchenko R, et al. (2023): Evaluation of the Autologous Genetically Enriched Leucoconcentrate on the Lumbar Spinal Cord Morpho-Functional Recovery in a Mini Pig with Thoracic Spine Contusion Injury. Biomedicines. 11:1331. [CrossRef]
- Bueno CRdS, Tonin MCC, Buchaim DV, Barraviera B, Junior RSF, Santos PSdS, et al. (2023): Morphofunctional Improvement of the Facial Nerve and Muscles with Repair Using Heterologous Fibrin Biopolymer and Photobiomodulation. Pharmaceuticals. 16:653.
- Scalise S, Zannino C, Lucchino V, Lo Conte M, Scaramuzzino L, Cifelli P, et al. (2022): Human iPSC modeling of genetic febrile seizure reveals aberrant molecular and physiological features underlying an impaired neuronal activity. Biomedicines. 10:1075. [CrossRef]
- Tanaka M, Szabó Á, Vécsei L, Giménez-Llort L (2023): Emerging translational research in neurological and psychiatric diseases: from in vitro to in vivo models. MDPI, pp 15739.
- Datki Z, Sinka R (2022): Translational biomedicine-oriented exploratory research on bioactive rotifer-specific biopolymers. Advances in Clinical and Experimental Medicine.1-5.
- Kwon K-M, Lee M-J, Chung H-S, Pak J-H, Jeon C-J (2022): The Organization of Somatostatin-Immunoreactive Cells in the Visual Cortex of the Gerbil. Biomedicines. 10:92.
- Chen B, Hasan MM, Zhang H, Zhai Q, Waliullah A, Ping Y, et al. (2023): UBL3 Interacts with Alpha-synuclein in Cells and the Interaction is Downregulated by the EGFR Pathway Inhibitor Osimertinib. Biomedicines. 11:1685. [CrossRef]
- Song A, Cho G-W, Vijayakumar KA, Moon C, Ang MJ, Kim J, et al. (2021): Neuroprotective effect of valproic acid on salicylate-induced tinnitus. International Journal of Molecular Sciences. 23:23. [CrossRef]
- Ibos KE, Bodnár É, Bagosi Z, Bozsó Z, Tóth G, Szabó G, Csabafi K (2021): Kisspeptin-8 induces anxiety-like behavior and hypolocomotion by activating the HPA axis and increasing GABA release in the nucleus accumbens in rats. Biomedicines. 9:112.
- Puri S, Kenyon BM, Hamrah P (2022): Immunomodulatory Role of Neuropeptides in the Cornea. Biomedicines. 10.
- Mirchandani-Duque M, Barbancho MA, López-Salas A, Alvarez-Contino JE, García-Casares N, Fuxe K, et al. (2022): Galanin and Neuropeptide Y Interaction Enhances Proliferation of Granule Precursor Cells and Expression of Neuroprotective Factors in the Rat Hippocampus with Consequent Augmented Spatial Memory. Biomedicines. 10. [CrossRef]
- Taschereau-Dumouchel V, Michel M, Lau H, Hofmann SG, LeDoux JE (2022): Putting the "mental" back in "mental disorders": a perspective from research on fear and anxiety. Mol Psychiatry. 27:1322-1330.
- Li J, Li C, Subedi P, Tian X, Lu X, Miriyala S, et al. (2023): Light Alcohol Consumption Promotes Early Neurogenesis Following Ischemic Stroke in Adult C57BL/6J Mice. Biomedicines. 11. [CrossRef]
- Petković A, Chaudhury D (2022): Encore: Behavioural animal models of stress, depression and mood disorders. Front Behav Neurosci. 16:931964.
- Bahor Z, Nunes-Fonseca C, Thomson LD, Sena ES, Macleod MR (2016): Improving our understanding of the in vivo modelling of psychotic disorders: A protocol for a systematic review and meta-analysis. Evid Based Preclin Med. 3:e00022.
- Tanaka M, Spekker E, Szabó Á, Polyák H, Vécsei L (2022): Modelling the neurodevelopmental pathogenesis in neuropsychiatric disorders. Bioactive kynurenines and their analogues as neuroprotective agents-in celebration of 80th birthday of Professor Peter Riederer. J Neural Transm (Vienna). 129:627-642.
- Sobolewska-Nowak J, Wachowska K, Nowak A, Orzechowska A, Szulc A, Płaza O, Gałecki P (2023): Exploring the Heart-Mind Connection: Unraveling the Shared Pathways between Depression and Cardiovascular Diseases. Biomedicines. 11.
- Tug E, Fidan I, Bozdayi G, Yildirim F, Tunccan OG, Lale Z, Akdogan D (2024): The relationship between the clinical course of SARS-CoV-2 infections and ACE2 and TMPRSS2 expression and polymorphisms. Adv Clin Exp Med. 33:39-51.
- Fan P, Miranda O, Qi X, Kofler J, Sweet RA, Wang L (2023): Unveiling the Enigma: Exploring Risk Factors and Mechanisms for Psychotic Symptoms in Alzheimer's Disease through Electronic Medical Records with Deep Learning Models. Pharmaceuticals (Basel). 16.
- Festa F, Medori S, Macrì M (2023): Move Your Body, Boost Your Brain: The Positive Impact of Physical Activity on Cognition across All Age Groups. Biomedicines. 11. [CrossRef]
- Alhaddad A, Radwan A, Mohamed NA, Mehanna ET, Mostafa YM, El-Sayed NM, Fattah SA (2023): Rosiglitazone Mitigates Dexamethasone-Induced Depression in Mice via Modulating Brain Glucose Metabolism and AMPK/mTOR Signaling Pathway. Biomedicines. 11.
- Statsenko Y, Habuza T, Smetanina D, Simiyu GL, Meribout S, King FC, et al. (2023): Unraveling Lifelong Brain Morphometric Dynamics: A Protocol for Systematic Review and Meta-Analysis in Healthy Neurodevelopment and Ageing. Biomedicines. 11. [CrossRef]
- Dang J, Tao Q, Niu X, Zhang M, Gao X, Yang Z, et al. (2022): Meta-Analysis of Structural and Functional Brain Abnormalities in Cocaine Addiction. Front Psychiatry. 13:927075. [CrossRef]
- Okanda Nyatega C, Qiang L, Jajere Adamu M, Bello Kawuwa H (2022): Altered striatal functional connectivity and structural dysconnectivity in individuals with bipolar disorder: A resting state magnetic resonance imaging study. Front Psychiatry. 13:1054380.
- Du H, Yang B, Wang H, Zeng Y, Xin J, Li X (2023): The non-linear correlation between the volume of cerebral white matter lesions and incidence of bipolar disorder: A secondary analysis of data from a cross-sectional study. Front Psychiatry. 14:1149663.
- Chen Y, Yu R, DeSouza JFX, Shen Y, Zhang H, Zhu C, et al. (2023): Differential responses from the left postcentral gyrus, right middle frontal gyrus, and precuneus to meal ingestion in patients with functional dyspepsia. Front Psychiatry. 14:1184797. [CrossRef]
- Adamu MJ, Qiang L, Nyatega CO, Younis A, Kawuwa HB, Jabire AH, Saminu S (2023): Unraveling the pathophysiology of schizophrenia: insights from structural magnetic resonance imaging studies. Front Psychiatry. 14:1188603.
- Chang CH, Wang WL, Shieh YH, Peng HY, Ho CS, Tsai HC (2022): Case Report: Low-Frequency Repetitive Transcranial Magnetic Stimulation to Dorsolateral Prefrontal Cortex and Auditory Cortex in a Patient With Tinnitus and Depression. Front Psychiatry. 13:847618.
- Zakia H, Iskandar S (2022): Case report: Depressive disorder with peripartum onset camouflages suspected intracranial tuberculoma. Front Psychiatry. 13:932635.
- Nyatega CO, Qiang L, Adamu MJ, Kawuwa HB (2022): Gray matter, white matter and cerebrospinal fluid abnormalities in Parkinson's disease: A voxel-based morphometry study. Front Psychiatry. 13:1027907. [CrossRef]
- Rymaszewska J, Wieczorek T, Fila-Witecka K, Smarzewska K, Weiser A, Piotrowski P, Tabakow P (2022): Various neuromodulation methods including Deep Brain Stimulation of the medial forebrain bundle combined with psychopharmacotherapy of treatment-resistant depression-Case report. Front Psychiatry. 13:1068054.
- Kim BH, Kim SH, Han C, Jeong HG, Lee MS, Kim J (2023): Antidepressant-induced mania in panic disorder: a single-case study of clinical and functional connectivity characteristics. Front Psychiatry. 14:1205126.
- Zhou J, Cao Y, Deng G, Fang J, Qiu C (2023): Transient splenial lesion syndrome in bipolar-II disorder: a case report highlighting reversible brain changes during hypomanic episodes. Front Psychiatry. 14:1219592.
- Veldema J (2023): Non-Invasive Brain Stimulation and Sex/Polypeptide Hormones in Reciprocal Interactions: A Systematic Review. Biomedicines. 11.
- Manuello J, Costa T, Cauda F, Liloia D (2022): Six actions to improve detection of critical features for neuroimaging coordinate-based meta-analysis preparation. Neurosci Biobehav Rev. 137:104659.
- Nani A, Manuello J, Mancuso L, Liloia D, Costa T, Vercelli A, et al. (2021): The pathoconnectivity network analysis of the insular cortex: A morphometric fingerprinting. Neuroimage. 225:117481. [CrossRef]
- Liloia D, Crocetta A, Cauda F, Duca S, Costa T, Manuello J (2022): Seeking Overlapping Neuroanatomical Alterations between Dyslexia and Attention-Deficit/Hyperactivity Disorder: A Meta-Analytic Replication Study. Brain Sci. 12.
- Liloia D, Cauda F, Uddin LQ, Manuello J, Mancuso L, Keller R, et al. (2023): Revealing the Selectivity of Neuroanatomical Alteration in Autism Spectrum Disorder via Reverse Inference. Biol Psychiatry Cogn Neurosci Neuroimaging. 8:1075-1083. [CrossRef]
- Tanaka M, Chen C (2023): Editorial: Towards a mechanistic understanding of depression, anxiety, and their comorbidity: perspectives from cognitive neuroscience. Front Behav Neurosci. 17:1268156.
- Battaglia S, Schmidt A, Hassel S, Tanaka M (2023): Case reports in neuroimaging and stimulation. Frontiers in Psychiatry. 14:1264669.
- Cauda F, Nani A, Liloia D, Manuello J, Premi E, Duca S, et al. (2020): Finding specificity in structural brain alterations through Bayesian reverse inference. Human brain mapping. 41:4155-4172. [CrossRef]
- Balogh L, Tanaka M, Török N, Vécsei L, Taguchi S (2021): Crosstalk between existential phenomenological psychotherapy and neurological sciences in mood and anxiety disorders. Biomedicines. 9:340.
- Gregorio F, Battaglia S (2023): Advances in EEG-based functional connectivity approaches to the study of the central nervous system in health and disease. Advances in Clinical and Experimental Medicine: Official Organ Wroclaw Medical University.
- Hakamata Y, Hori H, Mizukami S, Izawa S, Yoshida F, Moriguchi Y, et al. (2023): Blunted diurnal interleukin-6 rhythm is associated with amygdala emotional hyporeactivity and depression: a modulating role of gene-stressor interactions. Frontiers in Psychiatry. 14:1196235. [CrossRef]
- Rassler B, Blinowska K, Kaminski M, Pfurtscheller G (2023): Analysis of Respiratory Sinus Arrhythmia and Directed Information Flow between Brain and Body Indicate Different Management Strategies of fMRI-Related Anxiety. Biomedicines. 11:1028.
- Vasiliu O (2023): Efficacy, Tolerability, and Safety of Toludesvenlafaxine for the Treatment of Major Depressive Disorder—A Narrative Review. Pharmaceuticals. 16:411.
- Tanaka M, Szabó Á, Körtési T, Szok D, Tajti J, Vécsei L (2023): From CGRP to PACAP, VIP, and Beyond: Unraveling the Next Chapters in Migraine Treatment. Cells. 12:2649. [CrossRef]
- Masuya H, Inoue M, Wada Y, Shimizu A, Nagano J, Kawai A, et al. (2005): Implementation of the modified-SHIRPA protocol for screening of dominant phenotypes in a large-scale ENU mutagenesis program. Mamm Genome. 16:829-837. [CrossRef]
- Mandillo S, Tucci V, Hölter SM, Meziane H, Banchaabouchi MA, Kallnik M, et al. (2008): Reliability, robustness, and reproducibility in mouse behavioral phenotyping: a cross-laboratory study. Physiol Genomics. 34:243-255. [CrossRef]
- Detke MJ, Rickels M, Lucki I (1995): Active behaviors in the rat forced swimming test differentially produced by serotonergic and noradrenergic antidepressants. Psychopharmacology (Berl). 121:66-72.
- Khisti RT, Chopde CT, Jain SP (2000): Antidepressant-like effect of the neurosteroid 3alpha-hydroxy-5alpha-pregnan-20-one in mice forced swim test. Pharmacol Biochem Behav. 67:137-143.
- Steru L, Chermat R, Thierry B, Simon P (1985): The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology (Berl). 85:367-370.
- Cryan JF, Mombereau C, Vassout A (2005): The tail suspension test as a model for assessing antidepressant activity: review of pharmacological and genetic studies in mice. Neurosci Biobehav Rev. 29:571-625.
- Lister RG (1987): The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology (Berl). 92:180-185.
- Pellow S, Chopin P, File SE, Briley M (1985): Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J Neurosci Methods. 14:149-167.
- Costall B, Coughlan J, Horovitz ZP, Kelly ME, Naylor RJ, Tomkins DM (1989): The effects of ACE inhibitors captopril and SQ29,852 in rodent tests of cognition. Pharmacol Biochem Behav. 33:573-579.
- Onaivi ES, Martin BR (1989): Neuropharmacological and physiological validation of a computer-controlled two-compartment black and white box for the assessment of anxiety. Prog Neuropsychopharmacol Biol Psychiatry. 13:963-976.
- van der Poel AM (1967): Ethological study of the behaviour of the albino rat in a passive-avoidance test. Acta Physiol Pharmacol Neerl. 14:503-505.
- Stanford SC (2007): The Open Field Test: reinventing the wheel. J Psychopharmacol. 21:134-135.
- Walsh RN, Cummins RA (1976): The Open-Field Test: a critical review. Psychol Bull. 83:482-504.
- Galla Z, Rajda C, Rácz G, Grecsó N, Baráth Á, Vécsei L, et al. (2021): Simultaneous determination of 30 neurologically and metabolically important molecules: A sensitive and selective way to measure tyrosine and tryptophan pathway metabolites and other biomarkers in human serum and cerebrospinal fluid. J Chromatogr A. 1635:461775. [CrossRef]
- Galla Z, Rácz G, Grecsó N, Baráth Á, Kósa M, Bereczki C, Monostori P (2021): Improved LC-MS/MS method for the determination of 42 neurologically and metabolically important molecules in urine. J Chromatogr B Analyt Technol Biomed Life Sci. 1179:122846.
- Sahin EK, Caykoylu A, Senat A, Erel O (2019): A comprehensive study of oxidative stress in patients with somatic symptom disorder. Acta Neuropsychiatrica. 31:100-105.
- Polat N, Beyaztas H, Aktas S, Maden O, Guler EM (2023): Comparison of oxidative stress parameters, thiol-disulfide homeostasis, and pro-inflammatory cytokines levels in patients with bipolar disorder and their first-degree relatives. Journal of Psychiatric Research. 162:103-112. [CrossRef]
- Juchnowicz D, Dzikowski M, Rog J, Waszkiewicz N, Zalewska A, Maciejczyk M, Karakuła-Juchnowicz H (2021): Oxidative stress biomarkers as a predictor of stage illness and clinical course of schizophrenia. Frontiers in Psychiatry. 12:728986.
- Savitz J, Drevets WC, Smith CM, Victor TA, Wurfel BE, Bellgowan PS, et al. (2015): Putative neuroprotective and neurotoxic kynurenine pathway metabolites are associated with hippocampal and amygdalar volumes in subjects with major depressive disorder. Neuropsychopharmacology. 40:463-471. [CrossRef]
- Barone P (2019): The ‘Yin’and the ‘Yang’of the kynurenine pathway: excitotoxicity and neuroprotection imbalance in stress-induced disorders. Behavioural pharmacology. 30:163-186.
- Globus MY-T, Ginsberg MD, Busto R (1991): Excitotoxic index—a biochemical marker of selective vulnerability. Neuroscience letters. 127:39-42.
- Schwarz MJ, Ackenheil M (2002): The role of substance P in depression: therapeutic implications. Dialogues Clin Neurosci. 4:21-29.
- Tanaka M, Török N, Vécsei L (2021): Are 5-HT(1) receptor agonists effective anti-migraine drugs? Expert Opin Pharmacother. 22:1221-1225.
- Kindler J, Lim CK, Weickert CS, Boerrigter D, Galletly C, Liu D, et al. (2020): Dysregulation of kynurenine metabolism is related to proinflammatory cytokines, attention, and prefrontal cortex volume in schizophrenia. Mol Psychiatry. 25:2860-2872.
- Van Praag HM, Kahn RS, Asnis GM, Wetzler S, Brown SL, Bleich A, Korn ML (1987): Denosologization of biological psychiatry or the specificity of 5-HT disturbances in psychiatric disorders. Journal of Affective Disorders. 13:1-8.
- Li D, Yu S, Long Y, Shi A, Deng J, Ma Y, et al. (2022): Tryptophan metabolism: Mechanism-oriented therapy for neurological and psychiatric disorders. Front Immunol. 13:985378. [CrossRef]
- Miura H, Ozaki N, Sawada M, Isobe K, Ohta T, Nagatsu T (2008): A link between stress and depression: shifts in the balance between the kynurenine and serotonin pathways of tryptophan metabolism and the etiology and pathophysiology of depression. Stress. 11:198-209.
- Correia AS, Vale N (2022): Tryptophan metabolism in depression: A narrative review with a focus on serotonin and kynurenine pathways. International journal of molecular sciences. 23:8493.
- Han Q, Cai T, Tagle DA, Li J (2010): Structure, expression, and function of kynurenine aminotransferases in human and rodent brains. Cell Mol Life Sci. 67:353-368. [CrossRef]
- Okada K, Angkawidjaja C, Koga Y, Kanaya S (2014): Structural and mechanistic insights into the kynurenine aminotransferase-mediated excretion of kynurenic acid. J Struct Biol. 185:257-266.
- Kucukkarapinar M, Yay-Pence A, Yildiz Y, Buyukkoruk M, Yaz-Aydin G, Deveci-Bulut TS, et al. (2022): Psychological outcomes of COVID-19 survivors at sixth months after diagnose: the role of kynurenine pathway metabolites in depression, anxiety, and stress. J Neural Transm (Vienna). 129:1077-1089. [CrossRef]
- Ford JD, Courtois CA (2021): Complex PTSD and borderline personality disorder. Borderline Personal Disord Emot Dysregul. 8:16.
- Horowitz MJ (2011): Stress response syndromes: PTSD, grief, adjustment, and dissociative disorders. Jason Aronson, Incorporated.
- Dorfman WI, Walker LE (2007): The Anxiety, Somatoform and Dissociative Disorders. First Responder’s Guide to Abnormal Psychology: Applications for Police, Firefighters and Rescue Personnel.87-107.
- Ehrlich I, Humeau Y, Grenier F, Ciocchi S, Herry C, Lüthi A (2009): Amygdala inhibitory circuits and the control of fear memory. Neuron. 62:757-771.
- Hartley CA, Phelps EA (2010): Changing fear: the neurocircuitry of emotion regulation. Neuropsychopharmacology. 35:136-146.
- Cowansage KK, Shuman T, Dillingham BC, Chang A, Golshani P, Mayford M (2014): Direct reactivation of a coherent neocortical memory of context. Neuron. 84:432-441.
- Barsy B, Kocsis K, Magyar A, Babiczky Á, Szabó M, Veres JM, et al. (2020): Associative and plastic thalamic signaling to the lateral amygdala controls fear behavior. Nat Neurosci. 23:625-637. [CrossRef]
- Tsigos C, Chrousos GP (2002): Hypothalamic-pituitary-adrenal axis, neuroendocrine factors and stress. J Psychosom Res. 53:865-871.
- Rodrigues SM, LeDoux JE, Sapolsky RM (2009): The influence of stress hormones on fear circuitry. Annu Rev Neurosci. 32:289-313.
- Battaglia S, Di Fazio C, Mazzà M, Tamietto M, Avenanti A (2024): Targeting Human Glucocorticoid Receptors in Fear Learning: A Multiscale Integrated Approach to Study Functional Connectivity. International Journal of Molecular Sciences. 25:864.
- Spielberg JM, Stewart JL, Levin RL, Miller GA, Heller W (2008): Prefrontal Cortex, Emotion, and Approach/Withdrawal Motivation. Soc Personal Psychol Compass. 2:135-153.
- Frankland PW, Josselyn SA, Köhler S (2019): The neurobiological foundation of memory retrieval. Nat Neurosci. 22:1576-1585.
- Grace AA (2016): Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat Rev Neurosci. 17:524-532.
- Westerhof GJ, Bohlmeijer ET, McAdams DP (2017): The Relation of Ego Integrity and Despair to Personality Traits and Mental Health. J Gerontol B Psychol Sci Soc Sci. 72:400-407.
- Jing L, Duan TT, Tian M, Yuan Q, Tan JW, Zhu YY, et al. (2015): Despair-associated memory requires a slow-onset CA1 long-term potentiation with unique underlying mechanisms. Sci Rep. 5:15000. [CrossRef]
- van der Kolk BA (1994): The body keeps the score: memory and the evolving psychobiology of posttraumatic stress. Harv Rev Psychiatry. 1:253-265.
- van Meerkerk-Aanen PJ, de Vroege L, Khasho D, Foruz A, van Asseldonk JT, van der Feltz-Cornelis CM (2017): La belle indifférence revisited: a case report on progressive supranuclear palsy misdiagnosed as conversion disorder. Neuropsychiatr Dis Treat. 13:2057-2067.
- Reid S, Barbui C (2010): Long term treatment of depression with selective serotonin reuptake inhibitors and newer antidepressants. Bmj. 340:c1468.
- Raison S, Weissmann D, Rousset C, Pujol JF, Descarries L (1995): Changes in steady-state levels of tryptophan hydroxylase protein in adult rat brain after neonatal 6-hydroxydopamine lesion. Neuroscience. 67:463-475.
- Jacobsen JP, Medvedev IO, Caron MG (2012): The 5-HT deficiency theory of depression: perspectives from a naturalistic 5-HT deficiency model, the tryptophan hydroxylase 2Arg439His knockin mouse. Philos Trans R Soc Lond B Biol Sci. 367:2444-2459.
- Xu CJ, Wang JL, Jing P, Min L (2019): Tph2 Genetic Ablation Contributes to Senile Plaque Load and Astrogliosis in APP/PS1 Mice. Curr Alzheimer Res. 16:219-232.
- Angoa-Pérez M, Kane MJ, Briggs DI, Herrera-Mundo N, Sykes CE, Francescutti DM, Kuhn DM (2014): Mice genetically depleted of brain serotonin do not display a depression-like behavioral phenotype. ACS Chem Neurosci. 5:908-919.
- Sbrini G, Hanswijk SI, Brivio P, Middelman A, Bader M, Fumagalli F, et al. (2022): Peripheral Serotonin Deficiency Affects Anxiety-like Behavior and the Molecular Response to an Acute Challenge in Rats. Int J Mol Sci. 23. [CrossRef]
- Klemenhagen KC, Gordon JA, David DJ, Hen R, Gross CT (2006): Increased fear response to contextual cues in mice lacking the 5-HT1A receptor. Neuropsychopharmacology. 31:101-111.
- Bouchekioua Y, Nebuka M, Sasamori H, Nishitani N, Sugiura C, Sato M, et al. (2022): Serotonin 5-HT(2C) receptor knockout in mice attenuates fear responses in contextual or cued but not compound context-cue fear conditioning. Transl Psychiatry. 12:58. [CrossRef]
- Wellman CL, Izquierdo A, Garrett JE, Martin KP, Carroll J, Millstein R, et al. (2007): Impaired stress-coping and fear extinction and abnormal corticolimbic morphology in serotonin transporter knock-out mice. J Neurosci. 27:684-691. [CrossRef]
- Sorgdrager FJH, Naudé PJW, Kema IP, Nollen EA, Deyn PP (2019): Tryptophan Metabolism in Inflammaging: From Biomarker to Therapeutic Target. Front Immunol. 10:2565.
- Myint AM (2012): Kynurenines: from the perspective of major psychiatric disorders. Febs j. 279:1375-1385.
- Battaglia MR, Di Fazio C, Battaglia S (2023): Activated Tryptophan-Kynurenine metabolic system in the human brain is associated with learned fear. Front Mol Neurosci. 16:1217090.
- Skorobogatov K, Autier V, Foiselle M, Richard JR, Boukouaci W, Wu CL, et al. (2023): Kynurenine pathway abnormalities are state-specific but not diagnosis-specific in schizophrenia and bipolar disorder. Brain Behav Immun Health. 27:100584. [CrossRef]
- Liang Y, Xie S, He Y, Xu M, Qiao X, Zhu Y, Wu W (2022): Kynurenine Pathway Metabolites as Biomarkers in Alzheimer's Disease. Dis Markers. 2022:9484217.
- Marszalek-Grabska M, Walczak K, Gawel K, Wicha-Komsta K, Wnorowska S, Wnorowski A, Turski WA (2021): Kynurenine emerges from the shadows - Current knowledge on its fate and function. Pharmacol Ther. 225:107845.
- Yu P, Di Prospero NA, Sapko MT, Cai T, Chen A, Melendez-Ferro M, et al. (2004): Biochemical and phenotypic abnormalities in kynurenine aminotransferase II-deficient mice. Mol Cell Biol. 24:6919-6930. [CrossRef]
- Potter MC, Elmer GI, Bergeron R, Albuquerque EX, Guidetti P, Wu HQ, Schwarcz R (2010): Reduction of endogenous kynurenic acid formation enhances extracellular glutamate, hippocampal plasticity, and cognitive behavior. Neuropsychopharmacology. 35:1734-1742.
- Desbonnet L, Tighe O, Karayiorgou M, Gogos JA, Waddington JL, O'Tuathaigh CM (2012): Physiological and behavioural responsivity to stress and anxiogenic stimuli in COMT-deficient mice. Behav Brain Res. 228:351-358.
- Kash SF, Tecott LH, Hodge C, Baekkeskov S (1999): Increased anxiety and altered responses to anxiolytics in mice deficient in the 65-kDa isoform of glutamic acid decarboxylase. Proc Natl Acad Sci U S A. 96:1698-1703.
- Sangha S, Narayanan RT, Bergado-Acosta JR, Stork O, Seidenbecher T, Pape HC (2009): Deficiency of the 65 kDa isoform of glutamic acid decarboxylase impairs extinction of cued but not contextual fear memory. J Neurosci. 29:15713-15720.
- Zhang WH, Zhou J, Pan HQ, Wang XY, Liu WZ, Zhang JY, et al. (2017): δ Subunit-containing GABA(A) receptor prevents overgeneralization of fear in adult mice. Learn Mem. 24:381-384.
- Sideris A, Piskoun B, Russo L, Norcini M, Blanck T, Recio-Pinto E (2016): Cannabinoid 1 receptor knockout mice display cold allodynia, but enhanced recovery from spared-nerve injury-induced mechanical hypersensitivity. Mol Pain. 12. [CrossRef]
- de Kloet ER, Molendijk ML (2016): Coping with the Forced Swim Stressor: Towards Understanding an Adaptive Mechanism. Neural Plast. 2016:6503162.
- Lee DJ, Bovin MJ, Weathers FW, Palmieri PA, Schnurr PP, Sloan DM, et al. (2019): Latent factor structure of DSM–5 posttraumatic stress disorder: Evaluation of method variance and construct validity of novel symptom clusters. Psychological Assessment. 31:46. [CrossRef]
- McSweeney LB, Koch EI, Saules KK, Jefferson S (2016): Exploratory factor analysis of Diagnostic and Statistical Manual, criteria for posttraumatic stress disorder. The Journal of Nervous and Mental Disease. 204:9-14.
- Seligowski AV, Rogers AP, Orcutt HK (2016): Relations among emotion regulation and DSM-5 symptom clusters of PTSD. Personality and Individual Differences. 92:104-108.
- Di Gregorio F, Steinhauser M, Maier ME, Thayer JF, Battaglia S (2024): Error-related cardiac deceleration: functional interplay between error-related brain activity and autonomic nervous system in performance monitoring. Neuroscience & Biobehavioral Reviews.105542.
- Battaglia S, Nazzi C, Thayer JF (2024): Genetic differences associated with dopamine and serotonin release mediate fear-induced bradycardia in the human brain. Translational Psychiatry. 14:24.
Figure 1.
Tryptophan (Trp) metabolism. (a) The serotonin (5-HT) pathway: A fraction exceeding 2% of L-Trp is metabolized within the 5-HT pathway. The rate-limiting enzyme tryptophan hydroxylase 1 and 2 (TPH1, TPH2) converts Trp to 5-hydroxytryptophan (5-HTP), which is then decarboxylated by aromatic L-amino acid decarboxylase (AADC) to 5-HT. 5-HT is oxidized by monoamine oxidase A and B (MAOA, MAOB) in different tissues to 5-hydroxyindole acetaldehyde (5-HIAL), which is subsequently further oxidized to 5-hydroxyindoleacetic acid (5-HIAA) by aldehyde dehydrogenase (ALDH) or reduced to 5-hydroxytryptophol (5-HTOL) by alcohol dehydrogenase (ADH). 5-HIAA is the main metabolite and a marker of serotonergic activity, whereas 5-HTOL is a minor pathway of 5-HT degradation (61). On the other hand, 5-HT synthetize melatonin (N-acetyl-5-methoxytryptamine). First, 5-HT is converted into N-acetylserotonin (NAS) by arylalkylamine N-acetyltransferase (AANAT), then hydroxyindole-O-methyltransferase (HIOMT) transform melatonin (62-64). (b) The kynurenine (KYN) pathway: More than 90% of Trp enters the KYN pathway, which produces a variety of biomolecules. The primary metabolites include N-formyl-L-kynurenine (NFK), KYN, kynurenic acid (KYNA), anthranilic acid (AA), 3-hydroxykynurenine (3-HK), xanthurenic acid (XA), 3-hydroxyanthranilic acid (3-HAA), quinolinic acid (QA), picolinic acid (PA), and nicotinamide adenine dinucleotide (NAD+). These metabolites are produced through the catalytic actions of various enzymes, namely tryptophan 2,3-dioxygenase (TDO), indoleamine 2,3-dioxygenases (IDOs), kynurenine formamidase (KFA), kynurenine 3-monooxygenase (KMO), kynurenine aminotransferases (KATs), kynureninase (KYNU), 3-hydroxyanthranilate oxidase (3-HAO), quinolinate phosphoribosyl transferase (QPRT) (65), nicotinamide mononucleotide adenylyltransferase (NMNAT)(66), NAD synthetase (67), amino-β-carboxymuconate-semialdehyde-decarboxylase (ACMSD) and 2-aminomuconic-6-semialdehyde dehydrogenase (AMSD) (68-70). KYNA is subsequently metabolized by the gut microbiome to quinaldic acid (QAA) and 8-hydroxyquinaldic acid (71). 8-hydroxyquinaldic acid can be dehydroxylated from xanthurenic acid (72, 73). (c) The gut microbial indole pyruvate pathway: The metabolism of Trp is accomplished through four distinct pathways, which include the indoxyl sulfate pathway, the indole-3-acetamide (IAM) pathway, the tryptamine pathway, and the indole-3-propionic acid (IPA) pathway. The pyridoxal phosphate-dependent tryptophanase (TNA) enzyme serves as the rate-limiting component of the indoxyl sulfate pathway. Its primary function is to facilitate the transformation of Trp into indole, which is passing through the gut epithelium, then hydroxylated into 3-hydroxyindole (indoxyl) and ultimately transformed into indoxyl sulfate (INS) by p450 cytochrome and sulfanate in the liver (74). In the IAM pathway, tryptophan-2-monooxygenase (TMO) catalyzes the conversion of Trp to IAM. This is followed by the conversion of IAM to indole-3-acetic acid (IAA) by indole-3-acetamide hydrolase, which can then be further metabolized into indole-3-aldehyde (IAld) or decarboxylized into 3-methylindole (skatole) (75, 76). Tryptophan decarboxylase (TrD) catalyzes the conversion of Trp to tryptamine by amino acid decarboxylase (AAD), which subsequently undergoes conversion into indole-3-acetaldehyde (IAAld) (77). IAAld can be further converted into IAA by indole-3-acetaldehyde dehydrogenase. It is also worth noting that IAAld can be reversibly converted into indole-3-ethanol (tryptophol) by IAD reductase and tryptophol dehydrogenase (78). The transformation of Trp into indole-3-pyruvic acid (IPyA) is catalyzed by aromatic amino acid aminotransferase (ArAT), resulting in the formation of either tryptamine, or indole-3-lactic acid (ILA), indole acrylic acid (IA), and ultimately IPA (78-80). Black arrows: the host pathways, Yellow arrows: the gut microbiome pathways. AA: anthranilic acid, acdA: acyl-coenzim A dehydrogenase, AAD: amino acid decarboxylase, AADC: aromatic L-amino acid decarboxylase, AANAT: arylalkylamine N-acetyltransferase, ACMSD: amino-β-carboxymuconate-semialdehyde-decarboxylase, ADH: alcohol dehydrogenase, AldA: indole-3-acetaldehyde dehydrogenase, ALDH: aldehyde dehydrogenase, ArAT: aromatic amino acid aminotransferase, flBC: phenyllactate dehydratase, fldH: phenyllactate dehydrogenase, 3-HAA: 3-hydroxyanthranilic acid, 3-HAO: 3-hydroxyanthranilate oxidase, 5-HIAA: 5-hydroxyindoleacetic acid, 5-HIAL: 5-hydroxyindole-3-acetaldehyde, HIOMT: hydroxyindole-O-methyltransferase , 3-HK: 3-hydroxykynurenine, 5-HT: serotonin/5-hydroxytryptamine, 5-HTOL: 5-hydroxytryptophol, 5-HTP: 5-hydroxytryptophan, IA: indole acrylic acid, IAA: indole-3-acetic acid, IaaH: indole-3-acetamide hydrolase, IAAld: indole -3-acetaldehyde, IAld: indole-3-aldehyde, IAA: indole-3-acetic acid, IAM: indole-3-acetamide, IDO1/2: indoleamine 2,3-dioxygenases 1 and 2, ILA: indole-3-lactic acid, INS: indoxyl sulfate, IPA: indole-3-propionate, IPyA: indole-3-pyruvic acid, KAT III: kynurenine aminotransferase III/cysteine conjugate beta-lyase 2, KATs: kynurenine aminotransferases, KFA: kynurenine formamidase, KMO: kynurenine 3-monooxygenase, KYN: kynurenine, KYNA: kynurenic acid, KYNU: kynureninase, MAOA/B: monoamine oxidase A and B, NAAD: nicotinic acid adenine dinucleotide, NAD+: nicotinamide adenine dinucleotide, NAS: N-acetylserotonin, NFK: N-formyl-L-kynurenine, NMN: nicotinic acid mononucleotide, NMNAT: nicotinamide mononucleotide adenylyltransferase, PA: picolinic acid, QA: quinolinic acid, QAA: quinaldic acid, QPRT: quinolinate phosphoribosyl transferase, TDO: tryptophan-2,3-dioxygenase, TMO: tryptophan-2-monooxygenase, TNA: tryptophanase, TrD: tryptophan decarboxylase, Trp: tryptophan, TPH1/2: tryptophan hydroxylase 1 and 2, XA: xanthurenic acid, ?: unknown.
Figure 1.
Tryptophan (Trp) metabolism. (a) The serotonin (5-HT) pathway: A fraction exceeding 2% of L-Trp is metabolized within the 5-HT pathway. The rate-limiting enzyme tryptophan hydroxylase 1 and 2 (TPH1, TPH2) converts Trp to 5-hydroxytryptophan (5-HTP), which is then decarboxylated by aromatic L-amino acid decarboxylase (AADC) to 5-HT. 5-HT is oxidized by monoamine oxidase A and B (MAOA, MAOB) in different tissues to 5-hydroxyindole acetaldehyde (5-HIAL), which is subsequently further oxidized to 5-hydroxyindoleacetic acid (5-HIAA) by aldehyde dehydrogenase (ALDH) or reduced to 5-hydroxytryptophol (5-HTOL) by alcohol dehydrogenase (ADH). 5-HIAA is the main metabolite and a marker of serotonergic activity, whereas 5-HTOL is a minor pathway of 5-HT degradation (61). On the other hand, 5-HT synthetize melatonin (N-acetyl-5-methoxytryptamine). First, 5-HT is converted into N-acetylserotonin (NAS) by arylalkylamine N-acetyltransferase (AANAT), then hydroxyindole-O-methyltransferase (HIOMT) transform melatonin (62-64). (b) The kynurenine (KYN) pathway: More than 90% of Trp enters the KYN pathway, which produces a variety of biomolecules. The primary metabolites include N-formyl-L-kynurenine (NFK), KYN, kynurenic acid (KYNA), anthranilic acid (AA), 3-hydroxykynurenine (3-HK), xanthurenic acid (XA), 3-hydroxyanthranilic acid (3-HAA), quinolinic acid (QA), picolinic acid (PA), and nicotinamide adenine dinucleotide (NAD+). These metabolites are produced through the catalytic actions of various enzymes, namely tryptophan 2,3-dioxygenase (TDO), indoleamine 2,3-dioxygenases (IDOs), kynurenine formamidase (KFA), kynurenine 3-monooxygenase (KMO), kynurenine aminotransferases (KATs), kynureninase (KYNU), 3-hydroxyanthranilate oxidase (3-HAO), quinolinate phosphoribosyl transferase (QPRT) (65), nicotinamide mononucleotide adenylyltransferase (NMNAT)(66), NAD synthetase (67), amino-β-carboxymuconate-semialdehyde-decarboxylase (ACMSD) and 2-aminomuconic-6-semialdehyde dehydrogenase (AMSD) (68-70). KYNA is subsequently metabolized by the gut microbiome to quinaldic acid (QAA) and 8-hydroxyquinaldic acid (71). 8-hydroxyquinaldic acid can be dehydroxylated from xanthurenic acid (72, 73). (c) The gut microbial indole pyruvate pathway: The metabolism of Trp is accomplished through four distinct pathways, which include the indoxyl sulfate pathway, the indole-3-acetamide (IAM) pathway, the tryptamine pathway, and the indole-3-propionic acid (IPA) pathway. The pyridoxal phosphate-dependent tryptophanase (TNA) enzyme serves as the rate-limiting component of the indoxyl sulfate pathway. Its primary function is to facilitate the transformation of Trp into indole, which is passing through the gut epithelium, then hydroxylated into 3-hydroxyindole (indoxyl) and ultimately transformed into indoxyl sulfate (INS) by p450 cytochrome and sulfanate in the liver (74). In the IAM pathway, tryptophan-2-monooxygenase (TMO) catalyzes the conversion of Trp to IAM. This is followed by the conversion of IAM to indole-3-acetic acid (IAA) by indole-3-acetamide hydrolase, which can then be further metabolized into indole-3-aldehyde (IAld) or decarboxylized into 3-methylindole (skatole) (75, 76). Tryptophan decarboxylase (TrD) catalyzes the conversion of Trp to tryptamine by amino acid decarboxylase (AAD), which subsequently undergoes conversion into indole-3-acetaldehyde (IAAld) (77). IAAld can be further converted into IAA by indole-3-acetaldehyde dehydrogenase. It is also worth noting that IAAld can be reversibly converted into indole-3-ethanol (tryptophol) by IAD reductase and tryptophol dehydrogenase (78). The transformation of Trp into indole-3-pyruvic acid (IPyA) is catalyzed by aromatic amino acid aminotransferase (ArAT), resulting in the formation of either tryptamine, or indole-3-lactic acid (ILA), indole acrylic acid (IA), and ultimately IPA (78-80). Black arrows: the host pathways, Yellow arrows: the gut microbiome pathways. AA: anthranilic acid, acdA: acyl-coenzim A dehydrogenase, AAD: amino acid decarboxylase, AADC: aromatic L-amino acid decarboxylase, AANAT: arylalkylamine N-acetyltransferase, ACMSD: amino-β-carboxymuconate-semialdehyde-decarboxylase, ADH: alcohol dehydrogenase, AldA: indole-3-acetaldehyde dehydrogenase, ALDH: aldehyde dehydrogenase, ArAT: aromatic amino acid aminotransferase, flBC: phenyllactate dehydratase, fldH: phenyllactate dehydrogenase, 3-HAA: 3-hydroxyanthranilic acid, 3-HAO: 3-hydroxyanthranilate oxidase, 5-HIAA: 5-hydroxyindoleacetic acid, 5-HIAL: 5-hydroxyindole-3-acetaldehyde, HIOMT: hydroxyindole-O-methyltransferase , 3-HK: 3-hydroxykynurenine, 5-HT: serotonin/5-hydroxytryptamine, 5-HTOL: 5-hydroxytryptophol, 5-HTP: 5-hydroxytryptophan, IA: indole acrylic acid, IAA: indole-3-acetic acid, IaaH: indole-3-acetamide hydrolase, IAAld: indole -3-acetaldehyde, IAld: indole-3-aldehyde, IAA: indole-3-acetic acid, IAM: indole-3-acetamide, IDO1/2: indoleamine 2,3-dioxygenases 1 and 2, ILA: indole-3-lactic acid, INS: indoxyl sulfate, IPA: indole-3-propionate, IPyA: indole-3-pyruvic acid, KAT III: kynurenine aminotransferase III/cysteine conjugate beta-lyase 2, KATs: kynurenine aminotransferases, KFA: kynurenine formamidase, KMO: kynurenine 3-monooxygenase, KYN: kynurenine, KYNA: kynurenic acid, KYNU: kynureninase, MAOA/B: monoamine oxidase A and B, NAAD: nicotinic acid adenine dinucleotide, NAD+: nicotinamide adenine dinucleotide, NAS: N-acetylserotonin, NFK: N-formyl-L-kynurenine, NMN: nicotinic acid mononucleotide, NMNAT: nicotinamide mononucleotide adenylyltransferase, PA: picolinic acid, QA: quinolinic acid, QAA: quinaldic acid, QPRT: quinolinate phosphoribosyl transferase, TDO: tryptophan-2,3-dioxygenase, TMO: tryptophan-2-monooxygenase, TNA: tryptophanase, TrD: tryptophan decarboxylase, Trp: tryptophan, TPH1/2: tryptophan hydroxylase 1 and 2, XA: xanthurenic acid, ?: unknown.
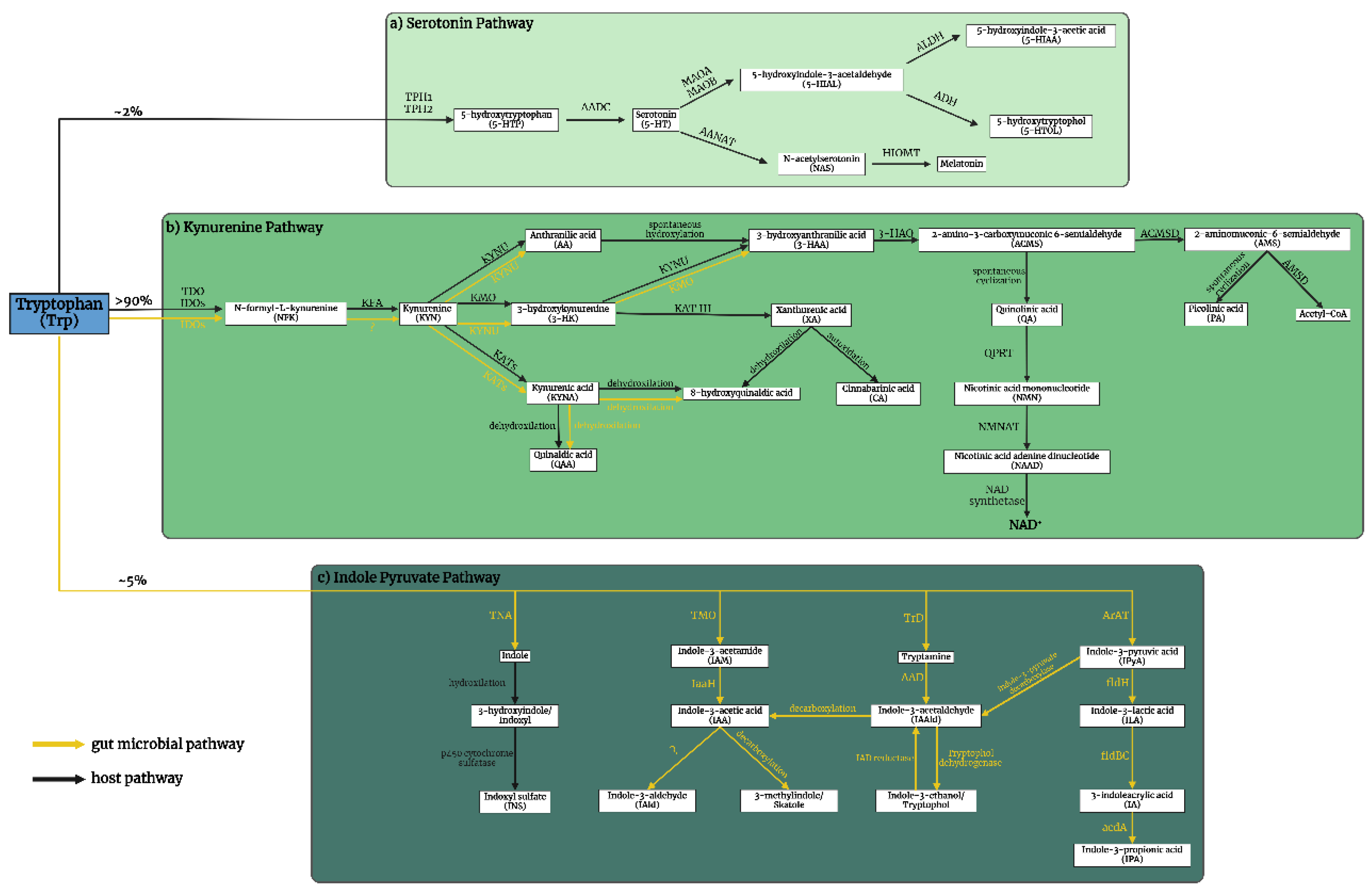
Figure 2.
Generation of the knockout kat2-/- mice. Female C57BL/6N mice were treated with pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG) with a 48-hour interval between administrations, then mated with male C57BL/6N mice. From the oviducts, fertilized one-cell embryos were collected and injected with single guide RNA (sgRNA) and Guide-itTM Recombinant Cas9 protein. At the two-cell stage, the embryos were transferred into pseudopregnant ICR mice. PMSG: pregnant mare serum gonadotropin; hCG: human chorionic gonadotropin; sgRNA: single guide RNA.
Figure 2.
Generation of the knockout kat2-/- mice. Female C57BL/6N mice were treated with pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG) with a 48-hour interval between administrations, then mated with male C57BL/6N mice. From the oviducts, fertilized one-cell embryos were collected and injected with single guide RNA (sgRNA) and Guide-itTM Recombinant Cas9 protein. At the two-cell stage, the embryos were transferred into pseudopregnant ICR mice. PMSG: pregnant mare serum gonadotropin; hCG: human chorionic gonadotropin; sgRNA: single guide RNA.
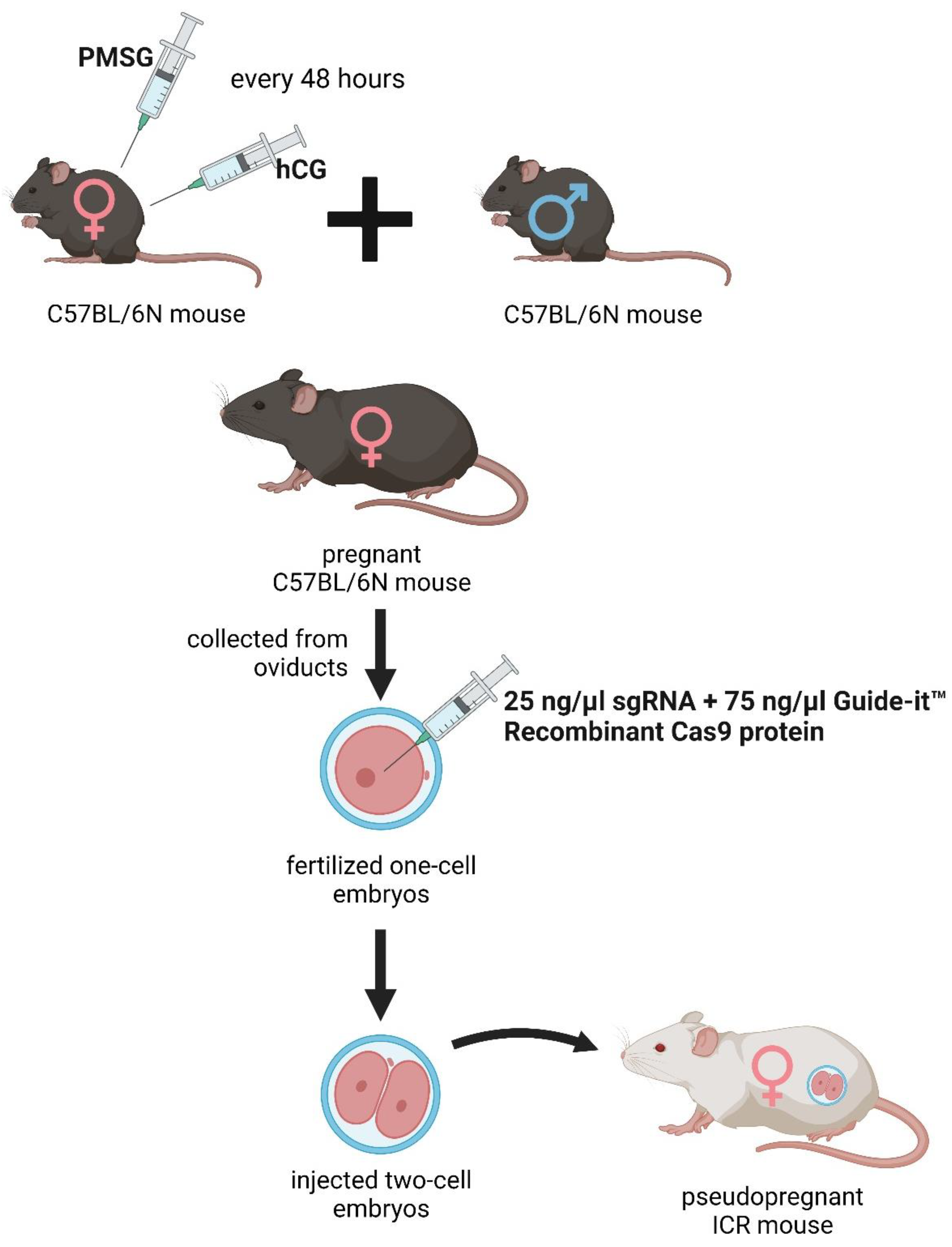
Figure 3.
DNA sequence analysis of knockout kat2-/- mouse line. a) Genomic sequences around the mutation site of knockout kat2-/- mouse strain. b) Western blot analysis of knockout kat2-/- mouse line.
Figure 3.
DNA sequence analysis of knockout kat2-/- mouse line. a) Genomic sequences around the mutation site of knockout kat2-/- mouse strain. b) Western blot analysis of knockout kat2-/- mouse line.
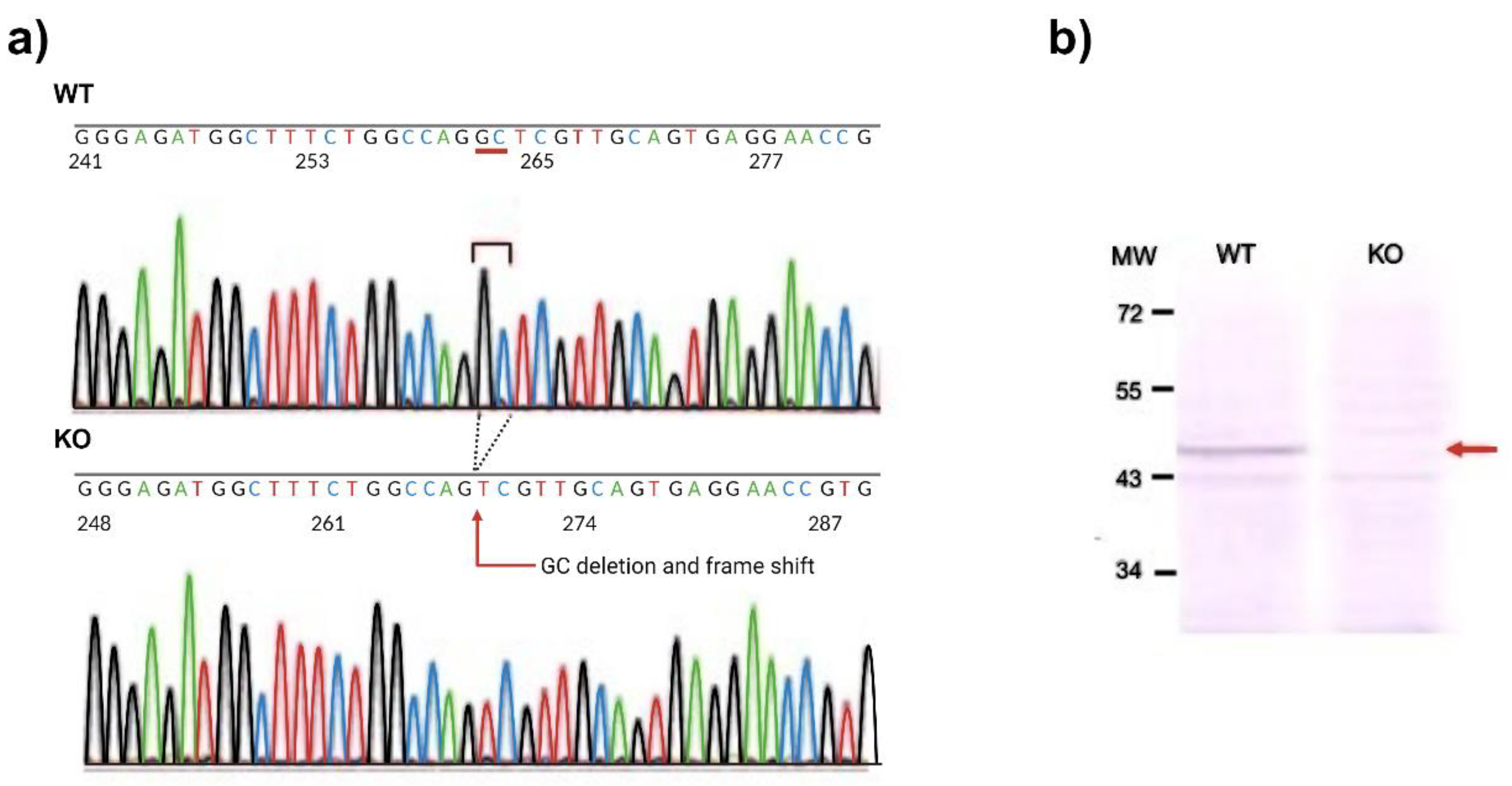
Figure 4.
Behavioral tests. a) Time spent immobile in the modified forced swim test (FST); b) Time spent swimming in the modified FST; c) Ambulation distance in the open field (OF) test; d) Number of jumps in the OF test; and e) Number of entries into the center and corner zones in the OF test. Wild-type mice (light green); kat2-/- mice (dark green). WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout mice; FST: forced swim test; OF: open field test; •: outliner. Mean ± SEM. *: p ≤ 0.05; **: p ≤ 0.01; ***: p ≤ 0.001.
Figure 4.
Behavioral tests. a) Time spent immobile in the modified forced swim test (FST); b) Time spent swimming in the modified FST; c) Ambulation distance in the open field (OF) test; d) Number of jumps in the OF test; and e) Number of entries into the center and corner zones in the OF test. Wild-type mice (light green); kat2-/- mice (dark green). WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout mice; FST: forced swim test; OF: open field test; •: outliner. Mean ± SEM. *: p ≤ 0.05; **: p ≤ 0.01; ***: p ≤ 0.001.
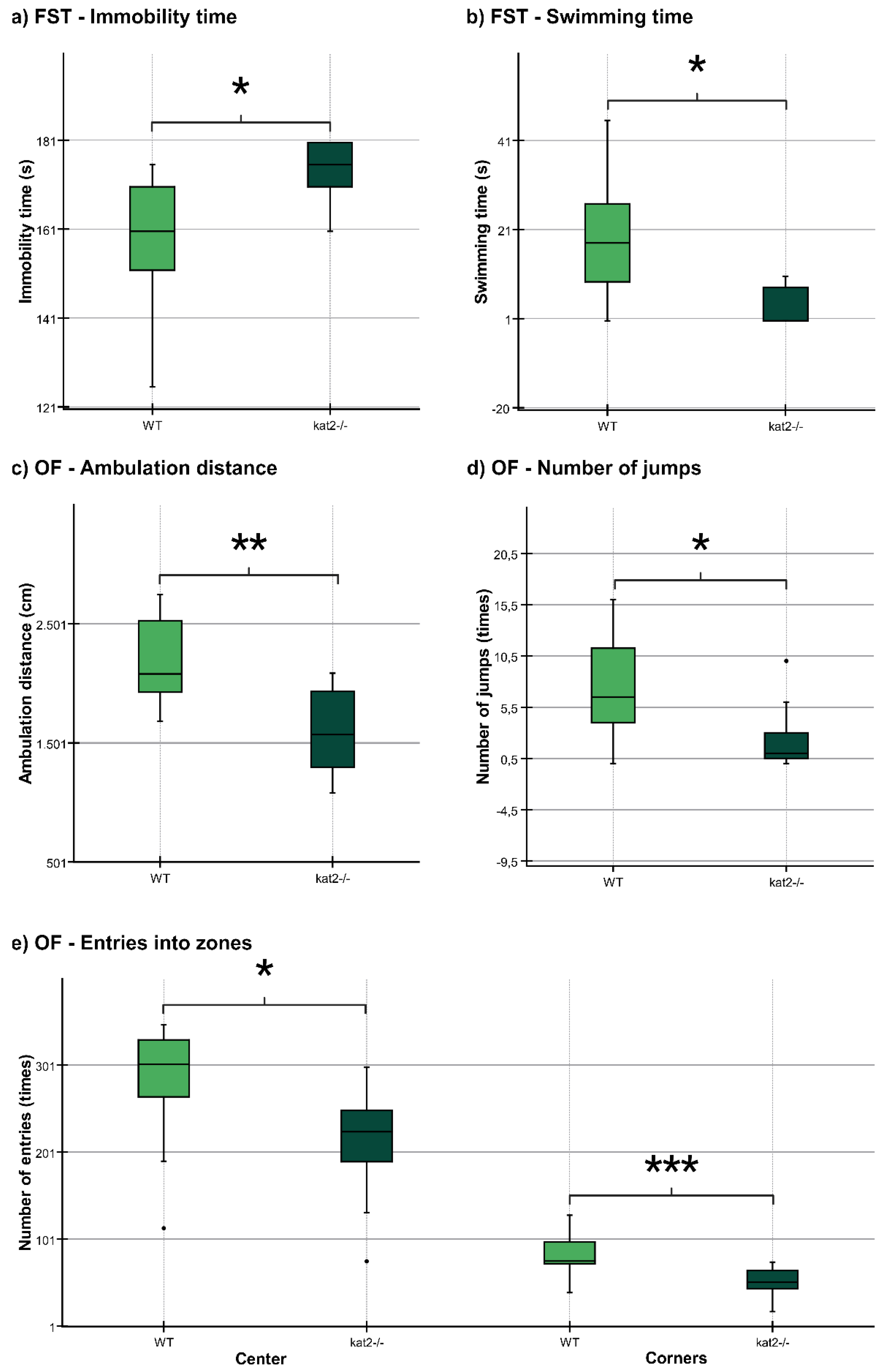
Figure 5.
Concentration of tryptophan and its metabolites in plasma. We marked wild-type with light, and kat2-/- mice results with dark green boxes. WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout; Trp: tryptophan; 5-HTP: 5-hydroxytryptophan; 5-HT: serotonin; 5-HIAA: 5-hydroxyindoleacetic acid; IAA: indole-3-acetic acid; INS: indoxyl-sulphate; KYN: kynurenine; KYNA: kynurenic acid; QAA: quinaldic acid; AA: anthranilic acid; 3-HK: 3-hydroxykynurenine; 3-HAA: 3-hydroxyanthranilic acid; QA: quinolinic acid; PA: picolinic acid; XA: xanthurenic acid; •: outliner; : far out. Mean+SEM; * ≤ 0.05; ** ≤ 0.01; *** ≤ 0.001.
Figure 5.
Concentration of tryptophan and its metabolites in plasma. We marked wild-type with light, and kat2-/- mice results with dark green boxes. WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout; Trp: tryptophan; 5-HTP: 5-hydroxytryptophan; 5-HT: serotonin; 5-HIAA: 5-hydroxyindoleacetic acid; IAA: indole-3-acetic acid; INS: indoxyl-sulphate; KYN: kynurenine; KYNA: kynurenic acid; QAA: quinaldic acid; AA: anthranilic acid; 3-HK: 3-hydroxykynurenine; 3-HAA: 3-hydroxyanthranilic acid; QA: quinolinic acid; PA: picolinic acid; XA: xanthurenic acid; •: outliner; : far out. Mean+SEM; * ≤ 0.05; ** ≤ 0.01; *** ≤ 0.001.
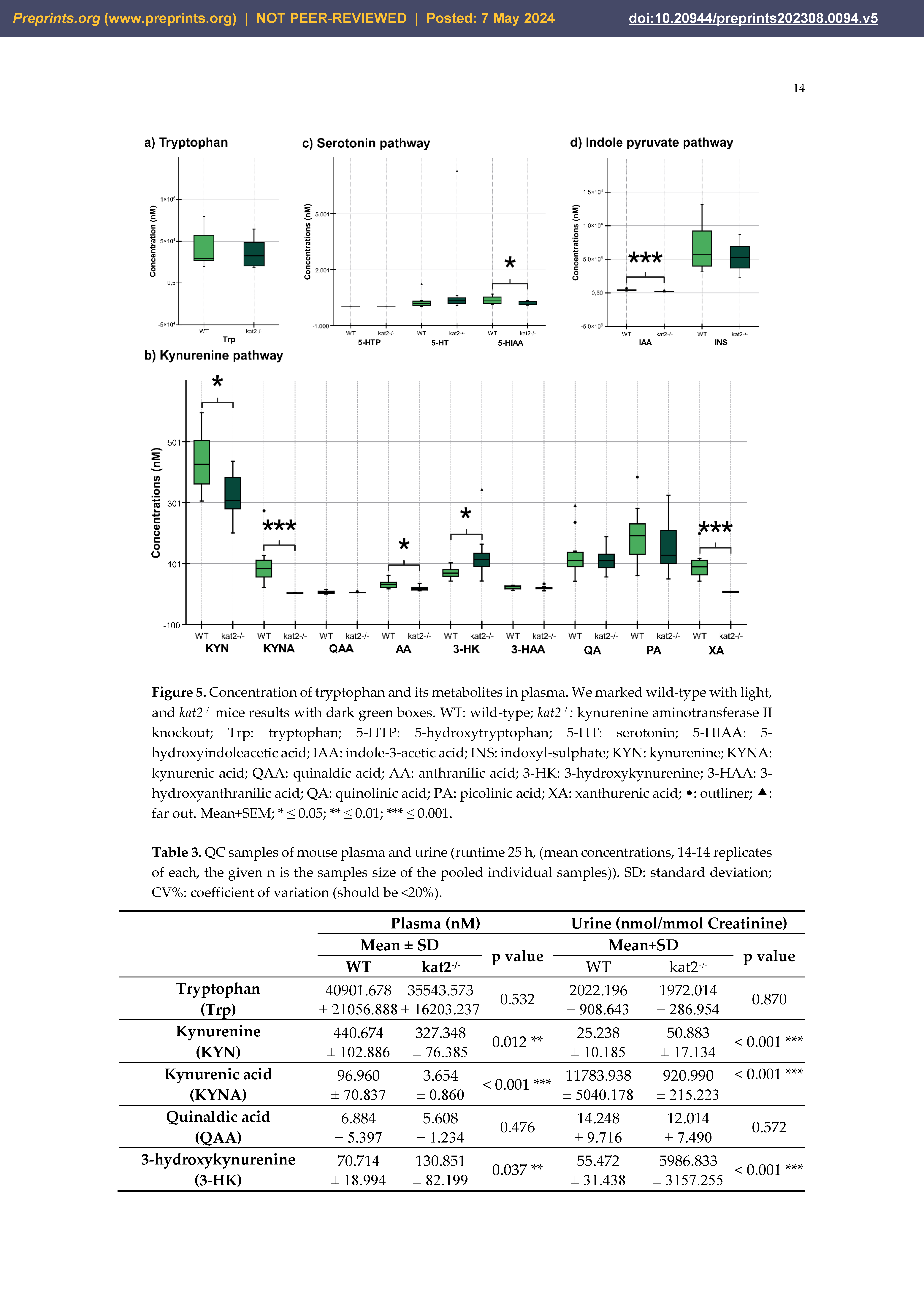
Figure 6.
Concentration of tryptophan and its metabolites in urine. We marked wild-type with light, and kat2-/- mice results with dark green boxes. WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout; Trp: tryptophan; 5-HTP: 5-hydroxytryptophan; 5-HT: serotonin; 5-HIAA: 5-hydroxyindoleacetic acid; IAA: indole-3-acetic acid; INS: indoxyl-sulphate; KYN: kynurenine; KYNA: kynurenic acid; QAA: quinaldic acid; AA: anthranilic acid; 3-HK: 3-hydroxykynurenine; 3-HAA: 3-hydroxyanthranilic acid; QA: quinolinic acid; PA: picolinic acid; XA: xanthurenic acid; •: outliner; : far out. Mean+SEM; * ≤ 0.05; ** ≤ 0.01; *** ≤ 0.001.
Figure 6.
Concentration of tryptophan and its metabolites in urine. We marked wild-type with light, and kat2-/- mice results with dark green boxes. WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout; Trp: tryptophan; 5-HTP: 5-hydroxytryptophan; 5-HT: serotonin; 5-HIAA: 5-hydroxyindoleacetic acid; IAA: indole-3-acetic acid; INS: indoxyl-sulphate; KYN: kynurenine; KYNA: kynurenic acid; QAA: quinaldic acid; AA: anthranilic acid; 3-HK: 3-hydroxykynurenine; 3-HAA: 3-hydroxyanthranilic acid; QA: quinolinic acid; PA: picolinic acid; XA: xanthurenic acid; •: outliner; : far out. Mean+SEM; * ≤ 0.05; ** ≤ 0.01; *** ≤ 0.001.
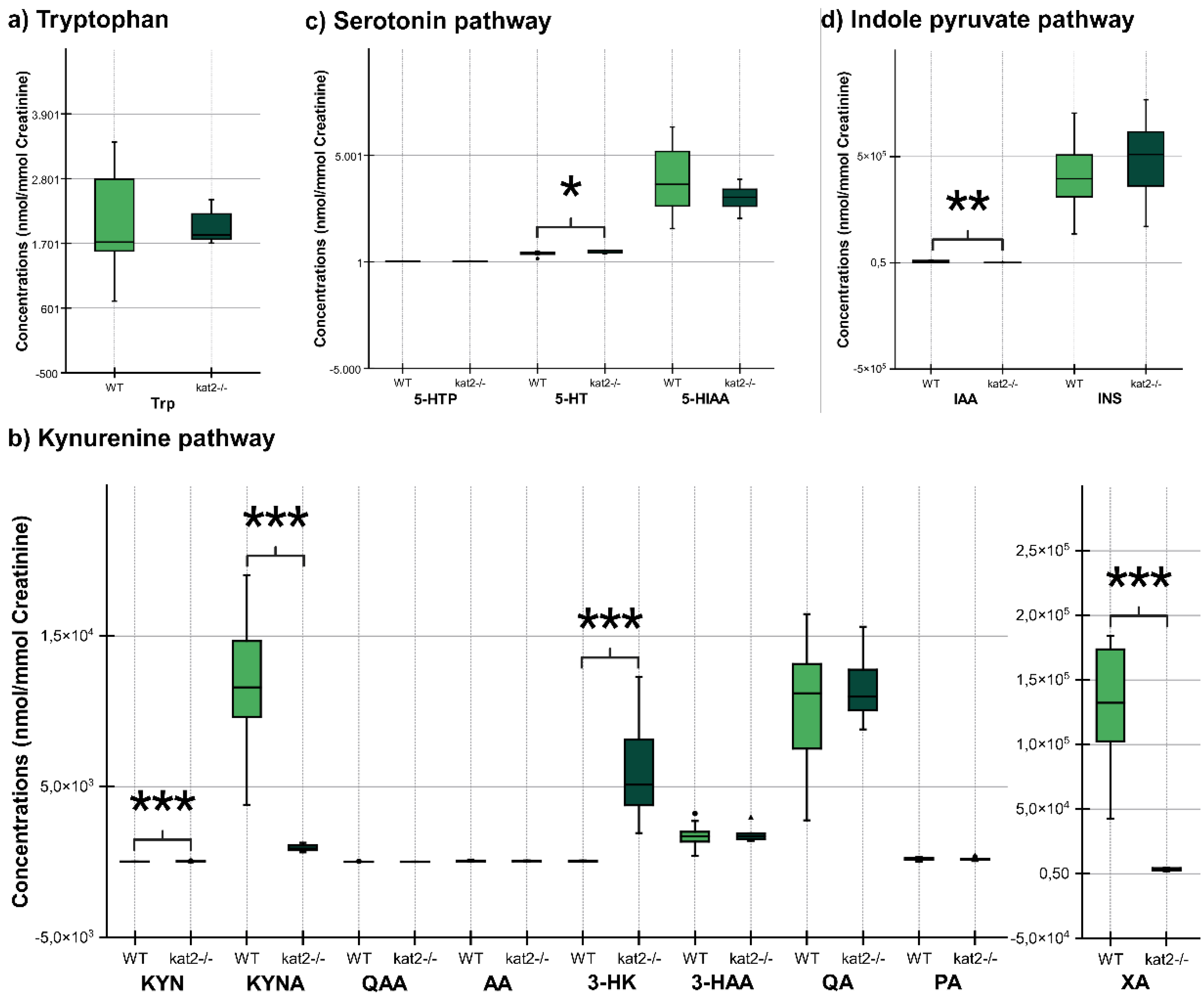

Table 1.
Properties of sgRNA, primers and KAT gene. sgRNA: single guide RNA; KAT: kynurenine aminotransferase; KAT II: aminoadipate aminotransferase; CCDS: Consensus Coding Sequence.
Table 1.
Properties of sgRNA, primers and KAT gene. sgRNA: single guide RNA; KAT: kynurenine aminotransferase; KAT II: aminoadipate aminotransferase; CCDS: Consensus Coding Sequence.
| Name of sgRNA | Sequence | ||
|---|---|---|---|
| M-KAT II-2 | GTTCCTCACTGCAACGAGCCguuuuagagcuagaaauagcaaguuaaaaaaggcuaguccguuaucaacuugaaaaaguggcacggacucggugcuuuu |
||
| Name of primer | Sequence | ||
| M-KAT II_1st_F | CCCTCTGTGGATGGACTTTG | ||
| M-KAT II_1st _R | TTGAAAGATGTGCCTCATGC | ||
| M-KAT II_2nd_F | GGATGGACTTTGTCCCTTCT | ||
| M-KAT II_2nd_R | ATGTGCCTCATGCTTGGCCC | ||
| Name of KAT gene | Transcript ID | CCDS | CCDS Nucleotide Sequence |
| Aadat-201 | ENSMUST00000079472.4 | CCDS22320 | 32-33 (2 nucleotide deletion) |
Table 2.
Behaviors of kat2−/− mice and the wild-type counterparts.
| Test type | Number of animals | Perspectives | Mean ± SEM of wild-type |
Mean ± SEM of kat2-/- |
p-value |
| Modified forced swim test (FST) | WT: n = 12 kat2-/-: n = 11 |
Immobility time (s) | 157,73 ± 4,97 | 174,09 ± 2,00 | 0,022 * |
| Swimming time (s) | 18,18 ± 4,44 | 3,18 ± 1,39 | 0,014 * | ||
| Climbing time (s) | 4,09 ± 1,26 | 1,82 ± 1,82 | 0,681 | ||
| Tail suspension test (TST) | WT: n = 10 kat2-/-: n = 13 |
Immobility time (s) | 194,50 ± 21,11 | 209,58 ± 18,65 | 0,625 |
| Passive avoidance (PA) test | WT: n = 12 kat2-/-: n = 12 |
Time spent in the lit box on the training day (s) | 48,33 ± 8,44 | 64,67 ± 16,10 | 0,979 |
| Time spent in the lit box on the test day (s) | 256,0 ± 22,2 | 283,8 ± 11,4 | 0,822 | ||
| Elevated plus maze (EPM) test | WT: n = 10 kat2-/-: n = 11 |
Time spent in the open arms (s) | 42,90 ± 19,48 | 30,64 ± 13,17 | 0,500 |
| Light-dark box (LDB) test | WT: n = 12 kat2-/-: n = 11 |
Time spent in the lit box (s) | 119,00 ± 9,06 | 113,91 ± 7,36 | 0,957 |
| Open-field test (OFT) | WT: n = 12 kat2-/-: n = 11 |
Number of entries to the center zones (times) | 281,67 ± 19,96 | 210,73 ± 19,66 | 0,011 * |
| Number of entries to the corner zones (times) | 83,08 ± 7,78 | 51,27 ± 5,39 | 0,001 *** | ||
| Ambulation distance (cm) | 2191,75 ± 105,21 | 1609,27 ± 115,17 | 0,002 ** | ||
| Number of jumps (times) | 7,33 ± 1,43 | 2,45 ± 0,93 | 0,034 * |
Table 3.
QC samples of mouse plasma and urine (runtime 25 h, (mean concentrations, 14-14 replicates of each, the given n is the samples size of the pooled individual samples)). SD: standard deviation; CV%: coefficient of variation (should be <20%).
Table 3.
QC samples of mouse plasma and urine (runtime 25 h, (mean concentrations, 14-14 replicates of each, the given n is the samples size of the pooled individual samples)). SD: standard deviation; CV%: coefficient of variation (should be <20%).
| Plasma (nM) | Urine (nmol/mmol Creatinine) | |||||
| Mean ± SD | p value | Mean+SD | p value | |||
| WT | kat2-/- | WT | kat2-/- | |||
|
Tryptophan (Trp) |
40901.678 ± 21056.888 |
35543.573 ± 16203.237 |
0.532 | 2022.196 ± 908.643 |
1972.014 ± 286.954 |
0.870 |
|
Kynurenine (KYN) |
440.674 ± 102.886 |
327.348 ± 76.385 |
0.012 ** | 25.238 ± 10.185 |
50.883 ± 17.134 |
< 0.001 *** |
|
Kynurenic acid (KYNA) |
96.960 ± 70.837 |
3.654 ± 0.860 |
< 0.001 *** | 11783.938 ± 5040.178 |
920.990 ± 215.223 |
< 0.001 *** |
|
Quinaldic acid (QAA) |
6.884 ± 5.397 |
5.608 ± 1.234 |
0.476 | 14.248 ± 9.716 |
12.014 ± 7.490 |
0.572 |
|
3-hydroxykynurenine (3-HK) |
70.714 ± 18.994 |
130.851 ± 82.199 |
0.037 ** | 55.472 ± 31.438 |
5986.833 ± 3157.255 |
< 0.001 *** |
|
Xanthurenic acid (XA) |
93.624 ± 45.637 |
7.406 ± 1.452 |
< 0.001 *** | 127228.662 ± 52582.223 |
3273.334 ±1021.511 |
< 0.001 *** |
|
Anthranilic acid (AA) |
32.655 ± 13.114 |
19.335 ± 7.280 |
0.012 ** | 69.112 ± 45.347 |
60.862 ± 22.368 |
0.612 |
|
3-Hydroxyanthranilic acid (3-HAA) |
22.992 ± 6.140 |
20.920 ± 5.921 |
0.452 | 1741.538 ± 824.887 |
1789.475 ± 454.422 |
0.874 |
|
Quinolinic acid (QA) |
132.185 ± 75.409 |
112.000 ± 41.600 |
0.468 | 10059.485 ± 4601.597 |
11718.491 ± 2401.051 |
0.326 |
|
Picolinic acid (PA) |
193.797 ± 93.230 |
154.895 ± 88.753 |
0.352 | 190.435 ± 91.394 |
193.898 ± 113.072 |
0.941 |
|
5-Hydroxytryptophan (5-HTP) |
2.790 ± 1.577 |
2.708 ± 1.297 |
0.901 | 21.742 ± 8.520 |
19.297 ± 3.833 |
0.419 |
|
Serotonin (5-HT) |
277.309 ± 353.179 |
1010.379 ± 2219.355 |
0.316 | 371.974 ± 125.489 |
479.383 ± 63.304 |
0.027 * |
|
5-hydroxyindoleacetic acid (5-HIAA) |
362.241 ± 199.450 |
201.217 ± 99.184 |
0.035 ** | 3774.968 ± 1666.005 |
2969.725 ± 598.373 |
0.167 |
|
Indole-3-acetic acid (IAA) |
457.329 ± 153.046 |
229.142 ± 68.266 |
< 0.001 *** | 6030.306 ± 4737.901 |
1513.400 ± 1097.122 |
0.009 ** |
|
Indoxyl-sulphate (INS) |
6738.111 ± 3559.896 |
5404.257 ± 2292.535 |
0.332 | 400636.750 ± 185880.105 |
497063.585 ± 190235.646 |
0.267 |
Table 4.
Enzymes activities in plasma and urine.
| Enzyme | Product/Substrate | Plasma | Urine | ||||
|---|---|---|---|---|---|---|---|
| Mean ± SD | p value | Mean ± SD | p value | ||||
| WT | kat2-/- | WT | kat2-/- | ||||
| TDO/IDOs (KFA) | KYN/Trp | 0.013 ± 0.007 |
0.011 ± 0.006 |
0.532 | 0.013 ± 0.002 |
0.026 ± 0.006 |
< 0.001 *** |
| KATs | KYNA/KYN | 0.205 ± 0.107 |
0.011 ± 0.002 |
< 0.001 *** | 476.464 ± 164.156 |
18.937 ± 5.057 |
< 0.001 *** |
| KMO | 3-HK/KYN | 0.168 ± 0.062 |
0.386 ± 0.180 |
0.002 ** | 2.219 ± 0.827 |
122.983 ± 75.543 |
< 0.001 *** |
| KYNU | AA/KYN | 0.075 ± 0.028 |
0.059 ± 0.016 |
0.120 | 2.593 ± 0.862 |
1.253 ± 0.529 |
< 0.001 *** |
| KYNU | 3-HAA/3-HK | 0.330 ± 0.070 |
0.194 ± 0.080 |
< 0.001 *** | 35.177 ± 16.776 |
0.372 ± 0.182 |
< 0.001 *** |
| KAT III | XA/3-HK | 1.374 ± 0.714 |
0.070 ± 0.033 |
< 0.001 *** | 2702.990 ± 1524.430 |
0.629 ± 0.229 |
< 0.001 *** |
| 3-HAO | QA/3-HAA | 5.l771 ± 2.978 |
5.486 ± 1.994 |
0.804 | 6.240 ± 2.487 |
6.856 ± 1.779 |
0.532 |
| 3-HA | PA/3-HAA | 8.797 ± 4.263 |
7.681 ± 4.872 |
0.592 | 1.286 ± 0.594 |
0.119 ± 0.085 |
< 0.001 *** |
| TPH | 5-HTP/Trp | < 0.001 ± <0.001 |
< 0.001 ± < 0.001 |
0.128 | 15.653 ± 6.024 |
0.010 ± 0.003 |
< 0.001 *** |
| AADC | 5-HT/5-HTP | 97.585 ± 87.384 |
307.233 ± 509.276 |
0.216 | 10.209 ± 2.530 |
25.997 ± 7.185 |
< 0.001 *** |
| MAO &ALDH | 5-HIAA/5-HT | 4.217 ± 4.818 |
0.905 ± 0.712 |
0.045 * | < 0.001 ± < 0.001 |
6.181 ± 0.859 |
< 0.001 *** |
| TMO(TrD, ArAT) | IAA/Trp | 0.013 ± 0.005 |
0.007 ± 0.002 |
0.005 ** | 5.243 ± 1.794 |
0.786 ± 0.636 |
< 0.001 *** |
| TNA | INS/Trp | 0.208 ± 0.178 |
0.170 ± 0.089 |
0.555 | 203.069 ± 108.000 |
248.916 ± 81.413 |
0.298 |
Table 5.
The oxidative stress and excitotoxicity indices in the plasma and urine.
| Oxidative stress index | ||||||
|---|---|---|---|---|---|---|
| Oxidant/antioxidant metabolites |
Plasma (nM) | Urine (nmol/mmol Creatinine) | ||||
| Mean ± SD | p value | Mean ± SD | p value | |||
| WT | kat2-/- | WT | kat2-/- | |||
| 3-HK/KYNA+AA+XA | 0.378 ± 0.163 |
4.090 ± 1.478 |
< 0.001 *** | 0.085 ± 0.011 |
1.352 ± 0.473 |
< 0.001 *** |
| Excitotoxicity index | ||||||
|
NMDA agonist/antagonist metabolites |
Plasma (nM) |
Urine (nmol/mmol Creatinine) |
||||
| QA/KYNA | 1.648 ± 0.810 |
30.514 ± 8.618 |
< 0.001 *** | 0.884 ± 0.320 |
13.092 ± 2.833 |
< 0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



