
Submitted:
01 August 2023
Posted:
02 August 2023
You are already at the latest version
Abstract
The growth of plant and process of yield formation in crops are moderated by surrounding conditions as well as interaction of genetic background of plants and environment. In the last two decades significant changes in climate can be observed generating unfavorable and harmful impact on plant development. Drought stress can be considered as one of the most dangerous environmental factor influencing plant life cycle and decreasing production of biomass and finally the yield. Plants can react to water deficit in a wide degree what is dependent on the species, the genetic variability within the species, the phase of the plant ontogenesis, the intensity of stress, and other potential stress factors. In plants there can be observed hybrids between distinct taxa which exhibit certain traits adopted to tolerate stress conditions better than parental plants. Oat × maize addition (OMA) plants are good example of fertile hybrids with retained maize chromosomes as addition to complete oat genome. Hybrids generated via wide crossing can exhibit morphological, physiological and biochemical variations implemented by the occurrence of extra chromosomes of maize, as well as interaction of maize and oat chromatin. The molecular analysis indicated 12.5% OMA lines among all tested descendants of wide oat-maize crossing. On the fourteenth day of the drought, a significant effect of stress on the content of soluble sugars and phenolic compounds was observed, soil drought significantly reduced plant biomass above ground and several yield components. Less water lost revealed by results of the excised leaf water loss test was correlated with high yield of OMA lines. Phenolic compounds content might be used as biochemical indicator of plant drought tolerance since there were significant correlation with high kernel yields of plants subjected to drought stress.
Keywords:
agronomic traits
; drought stress
; Grande I
; maize
; oat
; OMA
; phenolic compounds
; soluble sugars
1. Introduction
Drought is acknowledged as one of the most important factors lowering agricultural yield, including crops like wheat, rice, and maize [1]. Chlorophyll concentration in leaves may decrease due to extreme drought conditions as well as a breakdown of the photosynthetic system, particularly PSI and PSII [2]. However, it is still unknown how two photosystems interact under stress, as well as how drought stress affects how the photosynthetic linear electron transport mechanism reacts [3]. Chlorophyll is unquestionably a necessary component of photosynthesis in vegetation, therefore it might be viewed as a vital indication of plant production and function [4]. Through adjustments to multiple systems that function at various levels of the physiological, biochemical, and genetic processes, plants have developed exceptional capacities to govern growth and development in harsh environmental conditions. Depending on the severity of the stress, the kind of crop, genetic characteristics, the presence of other stressors, and the stage of plant development, a plant's response to drought stress may differ [5]. In one of the way of plant defense system are involved soluble sugars.
Carbohydrates are made of carbon, oxygen and hydrogen atoms. Single carbohydrate molecules are simple sugars, such as glucose or fructose. When one carbohydrate molecule is made up of several smaller ones, we are talking about oligosaccharides, such as sucrose. Polysaccharides such as starch, glycogen or cellulose are polysaccharides composed of many sugar molecules. Each type of carbohydrate plays important roles in plants, e.g. as an energy reserve, performing signal or regulatory functions, affecting the maintenance of turgor or causing the opening and closing of stomata [6]. In conditions of increasing water scarcity, the efficiency of all processes in the plant is at risk. Plants that are more tolerant of drought stress do better when intracellular pressure drops due to water loss. It depends to a large extent on the concentration of cell sap in the cytosol. Its key components are soluble sugars, the accumulation of which is correlated with resistance to drought stress [7]. Accumulation of sugar reserves as a preparation of plants for the stress of water shortage has been observed in many very different species, such as grasses or potatoes [8,9]. Storing large amounts of carbohydrates can inhibit photosynthesis while stimulating respiration. In addition, their stock can be used after the drought subsides as a source of readily available carbon [10]. The concentration of soluble sugars in the cell sap is a direct response to a given stressor. The functions of some sugars may be responsible for an indirect reaction, such as inducing the expression of some genes responsible for plant resistance or being precursors of many defense substances synthesized in the plant as a result of stress. Hexokinase signal transduction, for which hexoses are responsible, activates many genes, which increases the production of peroxidase or pathogen-related PR proteins [11]. The role of soluble sugars in the plant is also extremely important in the case of biotic stresses. By influencing changes in the water potential in cells infected with the pathogen, they limit its development. For example, higher resistance of grasses to snow mold was demonstrated at low water content and high concentration of soluble sugars in cells [12,13].
Another big group of molecules generated by plants in order to fight with stress factor are phenolic compounds. As a result of the stress factor, the synthesis of various types of compounds is induced in plants, which participate in the proper reaction aimed at limiting its harmful effects. The synthesized compounds may be specific to a given stressor protein. Heat shock proteins (HSP) or dehydrins, i.e. LEA (late embryogenesis abundant) proteins, occur during drought stress. In response to biotic stresses, for example, pathogenesis related (PR) proteins are synthesized, which are responsible, among others, for the hypersensitivity reaction, which cuts off the infection site from healthy tissue [14]. Phenolic compounds are important chemical compounds, the amount of which present in plants varies during the growing season and depends to a large extent on stress factors. This is a very diverse group of secondary metabolites. Phenolic compounds are divided into simple and complex phenols which are their derivatives. Simple phenols commonly found in plants are e.g. coumarins, vanillin, caffeic acid, salicylic acid or phenylpropanoids, i.e. cinnamic acid and its derivatives. Coumarins are seed germination inhibitors, they also inhibit cell division. They also have a protective function against herbivorous animals, affecting the taste and smell of plants. Phenolic compounds in root secretions have a significant impact on the phenomenon of allelopathy. Secreted by oat plants, scopoletin interrupts the reproduction cycles of many pests and diseases of cereals. For this reason, oat is considered a phytosanitary plant, improving its position in cereal crop rotations [15]. In turn, polymers of phenylpropanoids, such as lignin and suberin, saturate the cell walls of the cork - the secondary covering tissue. A large group of complex phenols are tannins, also called tannins, and flavonoids. They mainly perform protective functions, being poisons and repellents protecting against pathogens and pests. The toxic properties of phenols include: on protein denaturation. Anthocyanins, flavonols and flavones are individual groups of flavonoids that make up the majority of known groups of phenolic compounds. Absorption of different spectra of visible light causes different colors of aboveground parts of plants, mainly leaves. In addition to the ability to attract pollinating insects, resulting from the properties of flavonoids, their advantage is also protection against ultraviolet radiation. Increased content of phenolic compounds as a plant response to stress also contribute to the neutralization of reactive oxygen species (ROS), which are often the fastest reaction and effect of the stressor [16].
In crop species, there are several instances of sexual hybridization where stable viable hybrids have been produced, such as the oat (Avena sativa L. 2n = 6x = 42) and maize (Zea mays L. 2n = 2x = 20), which are particularly intriguing since they are clearly related plant species that are capable of sexual reproduction [17]. The discovery of keeping corn chromosomes in pollinated oat plants, led to the development of the oat × maize addition (OMA) lines [18] and their use in streamlining the study of the maize genome [17]. Numerous applications of OMA lines in research might be enumerated as follows: investigations on the expression of maize genes in the oat genetic background, including looking at elements of gene regulation [19], the potential acquisition of novel traits [20], and disease resistance [21], but also abiotic stress like drought [22]. The OMA lines are crosses between plants with C3 and C4 photosynthesis, allowing researchers to better understand C4 photosynthesis [20]. In order to follow the genetics of the maize C4 pathway and identify chromosomes/chromosome regions significant in this process, OMA lines may also be used. The C4 photosynthesis is thought to be more efficient, and plants that perform that type of CO2 assimilation are more resistant to photooxidation [20,23,24]. Other uses of OMA lines have been noted in the field of molecular genetics, including investigations into the structure of the maize centromere and knob [18], the behavior of chromosomes during meiosis [25], the use of in situ hybridization (FISH) to physically map single-copy sequences on maize chromosomes [26], and the procedure of flow-cytometry to separate individual maize chromosomes [27]. Additionally, the presence of maize chromatin frequently indicates morphological (such as thickened shoots, straight leaf blades, and bent panicles) and physiological (such as abnormal panicle growth and chlorophyll production) anomalies, but their characteristics vary depending on the specific addition of maize chromosomes as well as oat genetic background [23]. The greater tolerance of the OMA lines to different stressors, including resistance to environmental variables, such as Puccinia coronata f. sp. avenae or Puccinia graminis f. sp. avenae, is therefore assumed, as well as their effect on the behavior of the photosynthetic apparatus [28,29,30]. Numerous environmental conditions can impair the photosynthetic process and, as a result, alter the photosynthetic apparatus through structural and functional abnormalities [31], which in turn limits plant growth and biomass production [32,33].
Oat are a very significant cereal crop because of their great nutritional value, unique biological characteristics, and use in the cosmetics sector [34]. With a seeded area of 9.77 million hectares and a total harvest of 25.18 million tons in 2020, an average yield of 25.77 dt/ha was produced. Oat are mostly farmed in Australia, Canada, and Russia. In 2020, these three nations cultivated 45% of all agricultural land worldwide. Canada topped Russia in terms of harvest volume (4.57 million t vs 4.13 million t, respectively) thanks to higher oat grain yields [35]. Drought can diminish cereal yields by lowering the number of viable ears, the number of kernels per ear, and disrupted grain filling (lower thousand grain weight) [34]. However, oat exhibit increased vulnerability to drought stress and require more water during the vegetative phase than other cereal crops [36]. When being sown in early spring, spring cereals require longer time than winter cereals to reach the crucial stage of shooting at the stem, which increases the risk of drought-related losses due to a shortage of rainfall when the water stores in the soil run out [37]. The pouring of the grain is another crucial growth stage because a shortage of water at this stage causes the grain to fill poorly, resulting in a reduction in the weight of a thousand grains. Both winter and spring cereals fall under this [38]. In regions where late spring droughts are common, plant breeding produces early crop types that begin the most important water-demanding stages of the cycle as soon as feasible to get through them before protracted drought conditions arise [39]. Studies on the impact of dryness during grain filling on winter barley reveal a production decline of even more than 80% in both greenhouse and field circumstances [40]. Drought may spread to even more locations as a result of climate change, and its impacts may be more severe [41]. The first aim of the study were to identify OMA lines among plants generated by wide crossing with maize as pollinator, then to investigate possible differences in OMA lines according to Excised Leaf Water Loss (ELWL) test, and determining selected biochemical changes and agronomic characteristics of OMA plants developed from the cross between oat and maize under water availability and simulated soil drought conditions. Also detection if the possible changes are stable in subsequent generations (F2 and F3). The extra goal was to choose the lines best suited to development and cultivation under water limitation based on the aforementioned results.
2. Results
2.1. Molecular identification of OMA lines
Based on the results of electrophoretic separations, introgression of maize genetic material was found in 15 descendants resulting from wide crossing of oat with maize out of 120 tested genotypes the rest were oat DH (doubled haploids) lines. Therefore it was detected that 12.5 % of plants generated as a result of oat pollination with maize were OMA lines (oat maize addition). The OMA lines with the Grande 1 retrotransposon fragment (500 bp) presence are annotated as follows: 1b, 9, 12, 18, 23, 26, 35, 42, 43, 55, 78b, 83, 97, 114 and 119. The example result of DNA separation is presented in Figure 1.
2.2. Biochemical analysis
All the traits had a normal distribution. The results of MANOVA indicated that treatment, generation, genotype as well as all interactions were statistically significant (p<0.001) when examined in all ten quantitative traits jointly. Analysis of variance revealed that the main effects of all factors and all interaction were significant for all the traits of study, except for differences between control and stress for phenolic compounds content (p=0.818).
The mean water loss of all genotypes after 6 hours averaged 28.60%, and ELWL0-6 h values ranged from 20.81% to 42.08%. Genotypes 18, 119 and 35 lost the most water, and 23, 9 and 12 the least. Detailed results of each tested genotype are presented in Table 1.
On the first day of drought, the plants subjected to water restriction possessed in average 34% higher content of soluble sugars compared to the objects normally watered. A statistically insignificant decrease occurred only in line no. 18 and in the Bingo variety. The remaining lines showed an increase, and in 9 genotypes it was statistically significant. The highest increases of 70% were observed in lines no. 43, 12, 9 and 1b. The highest contents of soluble sugars in the control objects were recorded in lines no. 23 and 35 and in the Bingo variety, while the lowest contents were found in lines no. 43 and 42. Among the OMA lines in drought stress, the highest contents of soluble sugars were found in lines no. No. 18 and 42 (Table 2).
On the fourteenth day of drought, a decrease in the content of soluble sugars was noted compared to the first day of drought. In 3 genotypes and in the variety Bingo, the level of soluble sugars in combination with drought was lower than in the control objects, but significant decreases occurred only in line no. 78b and in the variety Bingo, where they amounted to 34% and 39%, respectively. In 11 OMA lines, an increase in the content of soluble sugars was observed as the effect of drought, but significant only in 6 lines, and the highest, over 100%, was observed in lines no. 18 and 1b. Among the control plants, the highest content of soluble sugars was recorded in lines no. 23 and 119 and in the Bingo variety, while the lowest in lines no. 42 and 18. In the group of plants subjected to drought stress, the highest contents were found in lines no. 83 and 1b and the lowest in no. 42 and 78b (Table 2).
On the first day of drought, there were no statistically significant differences in the average content of phenolic compounds between control plants and plants in drought conditions. But significant differences were observed between individual genotypes, such as in line no. 119, in which an increase in the content of phenolic compounds due to drought stress was observed, and in line no. 1b a decrease compared to control objects. Statistically significant differences as a result of drought occurred only in these two genotypes. Among the control objects, the highest content of phenolic compounds was observed in lines no. 43 and 42, and the lowest in lines no. 23 and 119 and in the Bingo variety. In the objects subjected to drought, the highest content of phenolic compounds was recorded in lines no. 114 and 43, and the lowest in lines no. 1b and 55 and in the Bingo variety (Table 3). On the fourteenth day of drought, significant differences in the content of phenolic compounds between control plants and plants growing in drought conditions were observed. On average, 6% more phenolic compounds were found in plants subjected to drought compared to control plant. As a result of drought, the highest increase in the content of phenolic compounds (57%) was recorded in line no. 9, and the only significant decrease occurred in line no. 114 and amounted to 21%. In the control conditions, the highest concentrations of phenolic compounds were recorded in lines no. 42 and 114, and the lowest in lines no. 18 and 26. In drought conditions, the highest concentrations of phenolic compounds were recorded in lines no. lines no. 18 and 35 (Table 3).
2.3. Analysis of primary and secondary shoots biomass and selected yield elements
Significant differences in the total mass of shoots were found between control and drought objects. On average, as a result of drought, 43% less biomass of aboveground parts was collected. No increase in shoot weight due to drought was found in any of the genotypes. Only for line no. 114, the decrease in the total weight collected was not statistically significant. In the remaining OMA lines and in the Bingo cultivar, the decreases in the harvested biomass of above-ground parts due to drought were statistically significant. The highest decreases were found in lines no. 18 and 78b and in the Bingo variety, where they amounted to 58%, 56% and 75%, respectively. Among the control plants the highest shoot weights were recorded on lines no. 78b and 1b, and the lowest on lines no. 114 and 83. On the other hand, when considering combination with drought, the highest masses of shoots were found in lines no. 1b and 119, and the lowest in lines no. 83 and 18 and in the Bingo cultivar (Table 4).
The number of developed grains differed in particular genotypes. As a result of drought, there was a statistically significant decrease in the number of kernels in drought combination by an average of 31% compared to control objects. Statistically significant decreases in the number of kernels occurred in lines no. 23, 78b, 83 and 9 and in the Bingo variety, where they amounted to 52%, 44%, 38%, 25% and 62%, respectively. Among the control plants, the most kernels were found in lines no. 78b and 9, and the least in lines no. 114 and 18. Whereas in drought conditions, the most kernels were found in lines no. 9 and 78b, and the least in lines no. 18 and 114 (Table 4).
The mass of kernels developed by single plant decreased in combination with drought by an average of 34% compared to control objects. Only in three lines statistically insignificant increases in kernel mass were found. This was the case with an overall very low number of developed grains. In the remaining genotypes, decreases were noted. In lines no. 1b, 83, 78b and 9 and in the Bingo variety, they were statistically significant. Among the objects of the control combination, the highest weights of kernels were recorded in lines no. 9 and 78b and in the Bingo variety, and the lowest in lines no. 18 and 114. Among the plants subjected to drought, the highest weights were found in lines No. 9 and 78b, and the lowest on lines 18 and 114 (Table 4).
2.4. Correlations among biochemical parameters, biomass and yield elements
Soluble sugars content in first day of drought (20% of soil field capacity) was significantly negatively correlated with phenolic compounds content in first day of drought in both control (–0.70) and drought stress (–0.39) (Figure 1 and Figure 2, respectively). Similarly, for both treatments, control and stress, significant positive correlations were observed between the number of grains and the mass of grains plant–1 (0.98 and 0.99, respectively) (Figure 1 and Figure 2). In addition, under control conditions, correlations were observed between ELWL after 0-3 hours and ELWL after 4-6 hours, ELWL after 0-3 hours and ELWL after 0-6 hours, ELWL after 4-6 hours and ELWL after 0-6 hours, soluble sugars content in first day of drought and soluble sugars content after two weeks of drought, phenolic compounds content in first day of drought and phenolic compounds content after two weeks of drought, Moreover, a positive correlation between the mass of stems plant–1 and the number of grains as well as negative between phenolic compounds content in first day of drought and soluble sugars content after two weeks of drought were observed (Figure 1).
In addition, under stress conditions, a positive correlation was observed between five pairs of traits: soluble sugars content in first day of drought and the mass of stems plant–1, soluble sugars content in first day of drought and the number of grains, soluble sugars content in first day of drought and the mass of grains plant–1, phenolic compounds content after two weeks of drought and the number of grains, and phenolic compounds content after two weeks of drought and the mass of grains plant–1 (Figure 2).
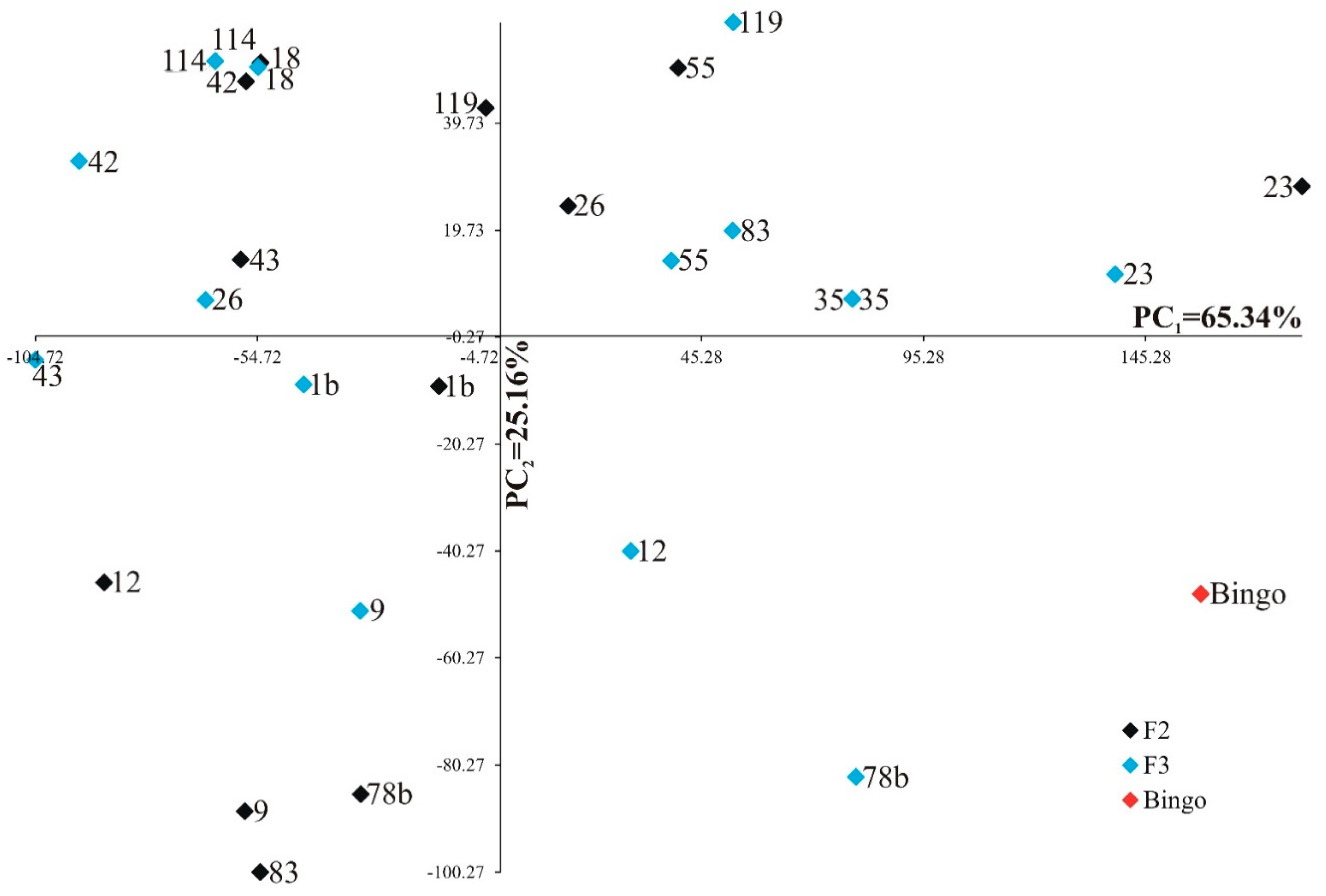
Each trait was of varying significance and had different shares in the joint multivariate variation in the examined genotypes. Analysis of the first two principal components for studied genotypes is shown in Figure 3 (for control) and Figure 4 (for drought stress). In the graphs the coordinates of the point for a studied lines and cultivar Bingo are the values for the first and second principal component. For control condition, the first two principal components accounted for 90.50% of the total variability between the individual genotypes (Figure 3). The most significant positive linear relationship with the first principal component was found for soluble sugars content in first day of drought and soluble sugars content after two weeks of drought, whereas the most significant negative linear relationship was found for phenolic compounds content in first day of drought. The second principal component was significantly negatively correlated with the number of grains and the mass of grains plant–1.
For drought stress, the first two principal components accounted for 94.04% of the total variability between the individual OMA lines and cultivar Bingo (Figure 4). The most significant positive linear relationship with the first principal component was found for soluble sugars content in first day of drought and soluble sugars content after two weeks of drought, while the most significant negative linear relationship was found for phenolic compounds content in first day of drought. The second principal component was significantly positively correlated with the number of grains and the mass of grains plant–1, whereas it was significantly negatively correlated with soluble sugars content after two weeks of drought.
Figure 5.
Distribution of fourteen OMA lines in F2 and F3 generations and cultivar Bingo in the space of the first two principal components for drought stress.
Figure 5.
Distribution of fourteen OMA lines in F2 and F3 generations and cultivar Bingo in the space of the first two principal components for drought stress.
Excised-leaf water loss after 0-3 hours (observed in control only) determined the mass of stems plant–1, the number of grains and the mass of grains plant–1 observed in drought stress (Table 5).
In all three cases, the impact was inversely proportional, and the coefficient of determination was not large, ranging from 2.6 (for the mass of grains plant–1) to 3.2 (for the number of grains). The mass of stems plant–1 was determined by ELWL after 0-6 hours, with values explained at 3.0% (Table 5).
3. Discussion
When certain related species are crossed, the pollinator's chromosomes might be totally excluded, resulting in haploid offspring generation. This method widely known as interspecies or intergeneric crossing (wide crossing) is utilized in plant breeding to speed breeding activities by getting a totally homozygous generation [42]. When the mentioned above technique is applied in crossing of oat with maize, certain maize chromosomes are retained in embryogenesis, during mitotic cell divisions and they act like oat chromosomes and are permanently integrated into the genome of newly produced hybrids. OMA lines (oat x maize addition lines) can be detected with molecular methods application, e.g. PCR with specific retrotransposon Grande I primer application. The method was used by many authors to detect OMA lines [22,43], the percentage of OMA lines could be high even 47% [43] but in our studied in the group of 120 offspring plants resulted from wide crossing oat with maize, 12.5% OMA lines were detected. OMA lines could be useful genotypes in plant breeding, but also they aid in the mapping of the maize genome [44].
The water loss test (ELWL) performed in our experiment revealed significant differences between the tested genotypes. Significant differences in the values of ELWL parameters between different genotypes were found in many cereal species [45,46]. Different crop plants, and within them different genotypes, for which low values of ELWL parameters were found, during drought have a greater ability to maintain water balance in the leaves, which is largely due to soluble sugars in the plant. The combination of low water loss and high content of soluble sugars allows to maintain water balance during stresses and provides such genotypes with tolerance to drought stress, in effect higher yield stabilization [46]. In our experiment, line no. 23 was characterized by the lowest water loss among all genotypes and was also characterized by the highest content of soluble sugars, especially compared to other drought-stressed genotypes. Negative correlations between the percentage of water loss and the analyzed yield components indicate genotypes that cope better with soil drought stress. If water loss is lower, yield under drought stress conditions is higher. On this basis, potentially valuable genotypes can be selected for recombinant breeding aimed at creating varieties tolerant to drought stress. Drought stress might possess strong impact on various physiological parameters of plant, generally stress including also biotic stress might reduce plant performance e.g. fluorescence parameters like Fv/Fm and PI indices decreased in barley DH lines that had been infected with Fusarium culmorum spores [47], and other PS II (photosystem II) characteristics, such RC/CSo decreased [48].
In our experiment the content of soluble sugars in oat plants increased as a result of soil drought. However, there were significant differences among tested genotypes, which was proved by significant interaction of two experimental factors T×G (Treatment and Genotype) both in the first day and 14-th day of drought. During prolonged drought conditions, the content of soluble sugars decreased (comparison of the first and fourteenth day of drought), but it was still higher in drought-treated plants than in control objects. The accumulation of soluble sugars in plants has been found in many species as a reaction to drought stress, e.g. in maize, oats, rapeseed or rice, in which the reaction to salinity stress was additionally studied [48,49,50]. The beneficial effect of soluble sugars during persistent water shortages was basically manifested by regulating the osmotic pressure of plants, which prevents significant decreases in turgor. Sinay and Karuwal (2014) in a greenhouse experiment with nine corn varieties, investigated the effect of watering intervals on the content of soluble sugars [51]. In the control objects, all objects were watered every other day, while the conditions of water deficiency were induced on two levels: watering only every 8 or 12 days. As a result of the induced drought conditions, significant increases in the content of soluble sugars in maize leaves were found, depending on the genotype, several times to several hundred times compared to the values in control objects [52]. In addition, a significant positive correlation was found between the content of soluble sugars and the analyzed yield components.
The content of phenolic compounds is another possible parameter applied in our studies to indicate genotypes tolerant to soil drought stress. During the experiments, it increased in both control and drought objects when compare the first and the fourteenth day of drought. In most cases, the values due to stress were higher. In both dates (first and fourteenth day of drought), the genotype had a significant impact on the content of phenolic compounds, which indicates a different reaction of the tested OMA lines to water deficiency. In experiments on drought stress in spring triticale, an increase in the content of phenolic compounds was found, and this parameter was considered useful in selecting genotypes less susceptible to drought stress [53]. In potato, increased content of phenolic compounds was also observed as a result of drought, during which the analyzed secondary metabolites were responsible for the good general condition of plants [54]. The positive effect of increased phenolic compounds content as antioxidants supporting plants in the fight against ROS (reactive oxygen species) induces by stress, has also been demonstrated in other species, such as rapeseed or potato [51,54].
The last analyzed parameters were aboveground biomass and selected yield elements. The occurrence of a two-week period of drought during the growing season of oat-maize hybrids affected the total dry mass of shoots, as well as the number and mass of kernels. Genotype, treatment and their interaction had a significant impact on the differences. The average decrease in the total aboveground mass was 43%. In the most susceptible genotypes, the decrease reached 75%, and in the tolerant ones the differences were not statistically significant. The experiments also showed a significant reduction in the number and weight of kernels. Among all analyzed lines, the highest values of yield components were achieved by lines no. 9 and 78b, especially in the case of the number and weight of kernels. The total biomass of above-ground parts of these lines decreased as a result of drought, but it was one of the smallest percentage decreases in these values compare to the rest of tested genotype. These lines were the only ones among the 14 analyzed OMA lines that achieved higher or similar values to the Bingo variety in the control, and even exceeded the standard object (cv. Bingo) in drought conditions. None of the genotypes reached a higher number and weight of kernels under drought stress when compare to lines no. 9 and 78b. Bingo variety reached similar values only for some yield elements. Studies conducted on wheat showed lower biomass values in plants growing in drought conditions compared to control combination with sufficient access to water. The decrease in yield was 59% for fresh weight and 51% for dry weight [55]. Other studies on drought stress in barley revealed a significant decrease in the number of kernels as well as their weight due to drought in both mild and severe drought stress. As a result of drought, the grain filling period was shortened by as much as ⅓ in the case of severe drought stress. The pouring of grain ended earlier, which caused a shortening of the vegetation period and led to a situation where, 17 days after the beginning of this phase, the average weight of one kernel was higher in drought-stressed plants than in control objects. Grain pouring in the control objects continued after 17 days from the beginning of this phase, so much higher grain weights than those in drought conditions were reached only after the end of drought vegetation [56]. In our experiment, faster maturation and dying of plants subjected to drought stress were also observed. In the experiment interesting and significant correlation was found, a higher production of biomass and a higher number and weight of kernels were associated with higher contents of soluble sugars on the first day of drought or higher contents of phenolic compounds on the fourteenth day of drought.
4. Materials and Methods
4.1. Molecular identification of OMA lines
The plant material for the study were obtained by wide crossing of oat with maize, at The Franciszek Górski Institute of Plant Physiology, Polish Academy of Sciences in Krakow. Intraspecific oat hybrids (cross combinations of cultivars or breeding lines) pollinated with maize allowed to generate a population of DH lines and OMA lines equivalent to the classic F2 generation, but with the difference that only homozygous forms constituted this population [57]. A total of 120 descendants of wide crosses were tested to detect oat-maize hybrids. The control objects were: maize variety Waza (positive control) and oat variety Stoper (negative control). Identification of hybrids was carried out using the PCR method, with the application of the marker retrotransposon fragment sequence Grande 1 present in multiple copies on each maize chromosome [23]. The leaves of plants grown in controlled greenhouse conditions were used for the study.
4.2. Grande I amplification
The harvested oat leaves were lyophilized under reduced pressure (40 μbar) at a heat exchanger temperature of -52°C (FreeZone 6L lyophilizer, Labconco, USA). The plant material was then homogenized using a cryogenic homogenization mill (MM400, Retsch, Germany) for 5 minutes at a frequency of 25 Hz in a 2000 μl round-bottom tubes together with 5 grinding balls made of zirconium oxide (Ø5 mm). Genomic DNA isolation was performed using the Genomic Mini AX Plant enhanced performance kit from A&A Biotechnology, the DNA was then dried in a concentrator (Concentrator Plus, Eppendorf, Germany) for 10 minutes at 20 mbar. Then dried DNA samples were diluted in TE buffer (Tris, EDTA) for 48 hours. The concentration of the DNA solution was measured using the NanoDrop 2000c spectrophotometer (Thermo Scientific, USA). Finally the DNA concentration 50 ng/μl was used in the PCR reaction. Apart from the OMA lines samples, the maize DNA isolated from cv. Waza (positive control) and oat DNA from variety cv. Stoper (negative control) was used in the reaction. The reaction mixture contained: 15.5 μl of water, 3 μl of MgCl2 solution, 2 μl of free nucleotides (dNTP) solution, 2.5 μl of Taq 10x buffer, 1 μl of Grande 1R primer solution, 1 μl of Grande 1F primer solution, 0,42 μl of Taq polymerase (Thermo Scientific, USA). The primers used bound to highly conserved elements, such as the Grande 1 retrotransposons, which are common on each maize chromosome [18] (GenBank accession number X976040. The Grande 1 primer sequence was as follows:
GRANDE 1F: 5' - AAAGACCTCACGAAAGGCCCAAGG - 3'
GRANDE 1R: 5' - AAATGGTTCATGCCGATTGCACG - 3'
The PCR reaction was carried out under the following conditions: pre-denaturation - 94°C for 5 minutes; 25 cycles:denaturation - 94°C for 30 seconds, primer annealing - 58°C for 30 seconds, polymerization - 72°C for 30 seconds; final polymerization at 72°C for 5 minutes and cooling the samples to 4°C. Separation of the amplification products according to the length of the newly formed DNA fragments was carried out by electrophoresis in 1.5% agarose gel in TBE buffer (Tris, boric acid, EDTA – Sigma Aldrich, USA) at a voltage of 110 V for 60 minutes. Archiving of electrophoretic separations was made using a gel archive system (Amersham - Pharmacia Biotech, USA) and the Liscap Capture Application ver. 1.0. The analysis of the obtained PCR products was performed using the GelScan ver. 1.45 (Kucharczyk - Electrophoretic Techniques, Poland). Computer analysis of the gels allowed the detection of oat DH lines and OMA lines.
4.3. Greenhouse experiment
OMA lines that were chosen based on molecular analyses were replicated to create F3 offspring, which were then evaluated in a greenhouse experiment in the spring of 2020. The Bingo oat variety was used as a reference genotype and two generations (F2 and F3) of fourteen OMA lines (No. 1b, 9, 12, 18, 23, 26, 35, 42, 43, and 55) were evaluated. The plants were given a soil drought treatment, while the control group consisted of the same plants that were consistently watered during the trial. In Krakow's Department of Plant Breeding, Physiology and Seed Science, the experiment was conducted in the greenhouse. Four replications of the experimental factor—i.e., drought conditions and optimal soil water content —were prepared for each OMA line.
A total of 240 pots were used in the experiment (15 genotypes, including 14 OMA lines and the cv. Bingo; four replicates; two generations; and two treatments). In February 2020, the genotypes under study had their seeds planted. The pots held a total of 2500 g of soil and sand combined in equal parts. In order to simulate a drought, watering was restricted after the soil humidity level reached 20%. The control pots received regular watering to 70% of soil humidity throughout the simulation of drought conditions, whereas the pots under the stress of the drought received only water to a moisture level of 20%. The following experiments and activities were conducted during the greenhouse experiment:
1. Performing ELWL (Excised Leave Water Loss)
2. Collection of leaves for soluble sugars and phenolic compound content in the first day of reaching 20% humidity simultaneously with control combination (70% soil humidity).
3. Collection of leaves for soluble sugars and phenolic compound content in the fourteenth day of drought (maintaining 20% humidity) simultaneously with control combination (70% soil humidity).
4. Harvesting mature shoots, weighing the biomass of aboveground parts and total mass of kernels from all shoots.
4.4. Water loss test
A physiological indicator widely used as a selection criterion for drought tolerance is the rate of Excised Leaf Water Loss (ELWL), which in soil drought conditions is negatively correlated with grain yield [58]. The water loss test was performed on OMA lines grown under controlled greenhouse conditions. The cut leaves of oat plants at the BBCH 27 development stage (end of tillering) were transferred to the growth chamber, where the following measurement conditions were maintained: temperature 20°C, air humidity 50% and lighting 250 µmol m2 s-1 (HPS "Agro" lamps, Osram). Water loss was monitored by weighing the leaves immediately after cutting (0 hours), after 3 and 6 hours, and after drying at 70 °C for 48 hours. The water loss in the tested plants was calculated on the basis of the formula given by Clarke and McCaig (1982) [59]:
where: FW0 - fresh mass, FW3 – fresh mass after 3 hours, FW6 – fresh mass after 6 hours, DW – dry weight after drying for 48 hours at 70°C.
• ELWL0-3 h = (FW0 – FW3) / (FW0 – DW),
• ELWL4-6 h = (FW3 – FW6) / (FW3 – DW),
• ELWL0-6 h = (FW0 – FW6) / (FW0 – DW),
4.5. Analysis of biochemical parameters
The leaves for biochemical analysis were collected twice during the experiment: on the first day of drought (when the soil reach the 20% of capacity) and on the fourteenth day of drought (the maintenance of soil 20% capacity) both from control plants and plants subjected to soil drought. One leaf was collected from the main shoots of each 3 pots of the F2 generation and 3 pots of the F3 generation. The harvested oat leaves were lyophilized under reduced pressure (40 μbar) at a heat exchanger temperature of –52°C (FreeZone 6L lyophilizer, Labconco, USA). The plant material was then homogenized using a cryogenic homogenization mill (MM400, Retsch, Germany) for 5 minutes at a frequency of 25 Hz in a 2000 μl round-bottom tubes together with 5 grinding balls made of zirconium oxide (Ø5 mm). For biochemical measurements, 5 mg of lyophilized and homogenized leaves were utilized with addition of 2 ml of 80% ethanol, then the tubes were centrifuged at 2800 rpm for 20 minutes (Eppendorf Centrifuge 5702 R, Germany). The total amount of sugars was determined by the phenol method by Dubois et al. (1956) [60]. The reaction mixture contained: 0.2 ml of distilled water, 20 μl of supernatant, 0.2 ml of 5% phenol and 1 ml of concentrated H2SO4. After 10 minutes from mixing the reaction components, the absorbance was measured at a wavelength of 490 nm (Synergy 2 spectrophotometer, BioTek, USA). On its basis, the content of soluble sugars in the samples was calculated, which was then presented as the content in the dry matter of plant tissue [mg g–1 DM]. The content of phenolic compounds was determined by the method of Folin and Ciocalteu [61]. The reaction mixture contained: 1 ml of water, 20 µl of supernatant, 0.5 ml of 25% Na2CO3 and 0.125 ml of Folin and Ciocalteu reagent (diluted 1:1 with distilled water shortly before use). After 30 minutes, the absorbance was measured on a spectrophotometer (Synergy 2, BioTek, USA) at a wavelength of 760 nm. The content of phenolic compounds was determined in mg of chlorogenic acid per 1 g of dry matter of plant tissue [mg g–1 DM].
4.6. Analysis of primary and secondary shoots biomass and selected yield elements
At the stage of kernels’ full maturity all plant of control and subjected to drought stress condition were harvested. The primary and secondary shoots biomass generated by plants was assessed, as well as the percentage of grains in the overall biomass yield. It was noted the quantity of shoots, their weight, and the weight of the kernels. Each shoot was measured independently, and the total amount of branching on each individual plant was then calculated and the results were gathered for each individual plant as described by Warzecha et al. 2023 [22].
4.7. Statistical analysis
The normality of distribution of the ten traits was tested using Shapiro–Wilk’s normality test [62] to verify whether the analysis of variance (ANOVA) met the assumption that the ANOVA model residuals followed a normal distribution. Three-way multivariate analysis of variance (MANOVA) was performed. Three-way analyses of variance (ANOVA) were carried out to determine the main effects of treatment, generation, and genotype as well as their interactions on the variability of the particular traits. The mean values and standard deviations of traits were calculated for treatments, generation, genotypes, and their combinations. Additionally, Fisher’s least significant differences (LSDs) were calculated for individual traits at the 0.05 level, and on this basis homogeneous groups were generated. The relationships between observed traits were estimated using Pearson’s linear correlation coefficients based on the means of the genotypes, independent for control and drought stress. The relationships of the observed traits are presented in heatmaps. The results were also analyzed using multivariate methods. Principal component analysis (PCA), independent for control and drought stress, was utilized to show a multi-trait assessment of the similarity of the tested genotypes in a lower number of dimensions with the least possible loss of information [63]. The effect of excised-leaf water loss values, independent: after 0-3 hours, after 4-6 hours and after 0-6 hours, on the mass of stems plant–1, the number of grains, the mass of grains plant–1 was assessed by regression analysis. All these analyses were conducted using the GenStat v. 22 statistical software package [64].
5. Conclusions
Among the 120 tested plants generated by wide crossing oat with maize, the presence of 105 DH (doubled haploids) lines and 15 OMA (oat × maize addition) lines was confirmed, which is 12.5% of all tested lines. The analysis of variance showed a significant effect of soil drought on the content of soluble sugars on the first day of soil drought, and on the fourteenth day of soil drought, a significant effect of stress on the content of soluble sugars and phenolic compounds was noted. Moreover, drought caused a significant decrease in the above-ground biomass of plants and selected yield elements. The tested OMA lines had a significant impact on the values of the excised leaf water loss (ELWL) test as well as biochemical parameters and selected yield components in response to drought stress. In the entire experiment, lines no. 9 and 78b were the most distinctive, as they were the only ones among the analyzed genotypes to achieve higher values of the tested yield components than the Bingo cultivar, both in the control and in the soil drought conditions. It is worth to note that for the OMA line no. 9 significant correlation between high yield values and low water loss from the leaves (ELWL test) were found. The ELWL values classified line no. 9 among the 3 genotypes with the lowest water loss, therefore it can be concluded that this genotype is more tolerant to drought stress. OMA lines no. 9 and 78b should be considered as the best candidates for further investigation and possible application in breeding program on oat resistance to soil drought stress. Moreover, in both lines, high content of phenolic compounds was found, correlated with a high yield of kernels in drought conditions. This parameter can be utilized as a biochemical indicator of oat tolerance to soil drought in oat.
Author Contributions
Conceptualization, T.W. and E.S.; methodology, T.W.; software, E.S.; validation, T.W., E.S. and J.B.; formal analysis, J.B.; investigation, T.W., E.S., R.B. and M.W.; resources, T.W.; data curation, T.W., E.S., R.B. and M.W.; writing—original draft preparation, T.W., J.B. and R.B.; writing—review and editing, T.W, J.B. and A.S.; visualization, J.B.; supervision, T.W.; project administration, T.W.; funding acquisition, J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data in this manuscript are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.L.D.; Condon, A.G.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 2010, 33, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.E.; Chaitanya, K.V. Photosynthesis and antioxidative defense mechanisms in deciphering drought stress tolerance of crop plants. Biol. Plant. 2016, 60, 201–218. [Google Scholar] [CrossRef]
- Peng, Y.; Nguy-Robertson, A.; Arkebauer, T.; Gitelson, A.A. Assessment of canopy chlorophyll content retrieval in maize and soybean: implications of hysteresis on the development of generic algorithms. Remote Sens. 2017, 9, 226. [Google Scholar] [CrossRef]
- Demirevska, K.; Zasheva, D.; Dimitrov, R.; Simova-Stoilova, L.; Stamenova, M.; Feller, U. Drought stress effects on Rubisco in wheat: changes in the Rubisco large subunit. Acta Physiol. Plant. 2009, 31, 1129–1138. [Google Scholar] [CrossRef]
- Okagaki, R.J.; Kynast, R.G.; Livingston, S.M.; Russell, C.D.; Rines, H.W.; Phillips, R.L. Mapping maize sequences to chromosomes using oat-maize chromosome addition materials. Plant Physiol. 2001, 125, 1228–1235. [Google Scholar] [CrossRef]
- Thomas, H. Drought resistance in plants. [W:] Mechanisms of environmental stress resistance in plants. Red. A. S. Basra, R. K. Basra. Harwood Academic Publishers, Netherlands, ss. 1-42, 1997.
- Hoekstra, F.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 8, 431–438. [Google Scholar] [CrossRef]
- Stitt, M.; Hurry, V. A plant for all seasons: alterations in photosynthetic carbon metabolism during cold acclimation in Arabidopsis. Curr. Opin. Plant Biol. 2002, 5, 199–206. [Google Scholar] [CrossRef]
- Nasir, M.W.; Toth, Z. Effect of Drought Stress on Potato Production: A Review. Agronomy 2022, 12, 635. [Google Scholar] [CrossRef]
- Paul, M.; Pellny, T. Carbon metabolite feedback regulation of leaf photosynthesis and development. J. Exp. Bot. 2003, 54(382), 539–547. [Google Scholar] [CrossRef]
- Herbers, K.; Meuwly, P.; Frommer, W.B.; Metraux, J.P.; Sonnewald, U. Systemic acquired resistance mediated by the ectopic expression of invertase: possible hexose sensing in the secretory pathway. The Plant Cell 1996, 8(5), 793–803. [Google Scholar] [CrossRef]
- Farrar, J.F. Fluxes and turnover of sucrose and fructans in healthy and diseased plants. J. Plant Physiol. 1989, 134(2), 137–140. [Google Scholar] [CrossRef]
- Yoshida, M.; Abe, J.; Moriyama, M.; Shimokawa, S.; Nakamura, Y. Seasonal changes in the physical state od crown water associated with freezing tolerance in winter wheat. Physiol. Plant. 1997, 99, 396–370. [Google Scholar] [CrossRef]
- Woloshuk, C.P.; Meulenhof, J.S.; Sela-Buurlage, M.; van den Elzen, P.J.M.; Cornelissen, B.J.C. Pathogen-induced proteins with inhibitor activity toward Phytophtora infestans. Plant Cell 1991, 3, 619–628. [Google Scholar] [PubMed]
- Tejeda-Sartorius, O.; Soto-Hernández, R.M.; San Miguel-Chávez, R.; Trejo-Téllez, L.I.; Caamal-Velázquez, H. Endogenous Hormone Profile and Sugars Display Differential Distribution in Leaves and Pseudobulbs of Laelia anceps Plants Induced and Non-Induced to Flowering by Exogenous Gibberellic Acid. Plants 2022, 11, 845. [Google Scholar] [CrossRef] [PubMed]
- Basharat, S.; Huang, Z.; Gong, M.; Lv, X.; Ahmed, A.; Hussain, I.; Li, J.; Du, G.; Liu, L. A review on current conventional and biotechnical approaches to enhance biosynthesis od steviol glycoside in Stevia rebaudiana. Chin. J. Chem. Eng. 2021, 30, 92–104. [Google Scholar] [CrossRef]
- Rines, H.W.; Phillips, R.L.; Kynast, R.G.; Okagaki, R.J.; Galatowitsch, M.W.; Huettl, P.A.; Stec, A.O.; Jacobs, M.S.; Suresh, J.; Porter, H.L.; Walch, M.D.; Cabral, C.B. Addition of individual chromosomes of maize inbreds B73 and Mo17 to oat cultivars Starter and Sun II: Maize chromosome retention, transmission, and plant phenotype. Theor. Appl. Genet. 2009, 119, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Ananiev, E.V.; Phillips, R.L.; Rines, H.W. Chromosome-specific molecular organization of maize (Zea mays L.) centromeric regions. Proc. Natl. Acad. Sci. U.S.A. 1998, 95, 13073–13078. [Google Scholar] [CrossRef]
- Muehlbauer, G.J.; Riera-Lizarazu, O.; Kynast, R.G.; Martin, D.; Phillips, R.L.; Rines, H.W. A maize chromosome 3 addition line of oat exhibits expression of the maize homeobox gene liguleless3 and alteration of cell fates. Genome 2000, 43, 1055–1064. [Google Scholar] [CrossRef]
- Kowles, R.V.; Walch, M.D.; Minnerath, J.M.; Bernacchi, C.J.; Stec, A.O.; Rines, H.W.; Phillips, R.L. Expression of C4 photosynthetic enzymes in oat-maize chromosome addition lines. Maydica 2008, 53, 69–78. [Google Scholar]
- Walch, M.D. Expression of maize pathogenesis-related and photosynthetic genes in oat × maize addition lines. University of Minnesota, Minneapolis, Saint Paul, 2007.
- Warzecha, T.; Bathelt, R.; Skrzypek, E.; Warchoł, M.; Bocianowski, J.; Sutkowska, A. Studies of Oat-Maize Hybrids Tolerance to Soil Drought Stress. Agriculture 2023, 13, 243. [Google Scholar] [CrossRef]
- Kynast, R.G.; Riera-Lizarazu, O.; Vales, M.I.; Okagaki, R.J.; Maquieira, S.B.; Chen, G.; Ananiev, E.V.; Odland, W.E.; Russell, C.D.; Stec, A.O. , et al. A complete set of maize individual chromosome additions to the oat genome. Plant Physiol. 2001, 125, 1216–1227. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Yu, J.; Li, H.; Huang, W.; Xu, L.; Zhao, Y.; Zhang, T.; Xu, W.; Jiang, J.; Su, Z.; Jin, W. Transcriptional and epigenetic adaptation of maize chromosomes in Oat-Maize addition lines. Nucleic Acids Res. 2018, 46, 5012–5028. [Google Scholar] [CrossRef] [PubMed]
- Bass, H.W.; Riera-Lizarazu, O.; Ananiev, E.V.; Bordoli, S.J.; Rines, H.W.; Phillips, R.L.; Sedat, J.W.; Agard, D.A.; Cande, W.Z. Evidence for the coincident initiation of homolog pairing and synapsis during the telomere-clustering (bouquet) stage of meiotic prophase. J. Cell Sci. 2000, 113, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Amarillo, F.I.E.; Bass, H.W. A transgenomic cytogenetic sorghum (Sorghum propinquum) bacterial artificial chromosome fluorescence in situ hybridization map of maize (Zea mays L.) pachytene chromosome 9, evidence for regions of genome hyperexpansion. Genetics 2007, 177, 1509–1526. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Arumuganathan, K.; Rines, H.W.; Phillips, R.L.; Riera-Lizarazu, O.; Sandhu, D.; Zhou, Y.; Gill, K.S. Flow cytometric sorting of maize chromosome 9 from an oat-maize chromosome addition line. Theor. Appl. Genet. 2001, 102, 658–663. [Google Scholar] [CrossRef]
- Kynast, R.; Okagaki, R.; Rines, H.; Phillips, R. Maize individualized chromosome and derived radiation hybrid lines and their use in functional genomics. Funct. Integr. Genomic 2002, 2, 60–69. [Google Scholar] [CrossRef]
- Ohm, H.; Shaner, G. Breeding oat for resistance to diseases. [W:] Oat science and technology. Red. H. Marshall, M. Sorrells. American Society of Agronomy, Madison, Wisconsin 1992, ss. 657–698.
- Rines, H.W.; Phillips, R.L.; Kynast, R.G.; Okagaki, R.; Odland, W.E.; Stec, A.O.; Jacobs, M.S.; Granath, S.R. Maize chromosome additions and radiation hybrids in oat and their use in dissecting the maize genome. In Proceedings of International congress “In the wake of the double helix: from the green revolution to the gene revolution". Bologna, Italy, 27–31 May 2003; pp. 427–441. [Google Scholar]
- Jastrzebska, M.; Kostrzewska, M.; Wanic, M. Wpływ deficytu wody i interakcji międzygatunkowych na wybrane parametry fizjologiczne roślin jęczmienia jarego i koniczyny czerwonej. Fragm. Agron. 2016, 33, 44–59, [In polish]. [Google Scholar]
- Lawlor, D.W.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a critical evaluation of mechanisms and integration of processes. Ann. Bot. 2009, 103, 561–579. [Google Scholar] [CrossRef]
- Akıncı, Ş.; Lösel, D.M. Plant water-stress response mechanisms. In Water stress, Rahman, I.M.M., Hasegawa, H., Eds. InTech: Rijeka, Croatia, 2012; pp. 15–42.
- Sadras, V.O.; Calderini, D.F. (Editors) Crop Physiology Case Histories for Major Crops 2021. Copyright © 2021 Elsevier Inc. [CrossRef]
- Food and Agriculture Organization of the United Nations (FAOSTAT). 2022. https://www.fao.org/faostat/en/%3F%23data/ (access 22.03.2022).
- Pisulewska, E.; Tobiasz-Salach, R.; Witkowicz, R.; Cieślik, E.; Bobrecka-Jamro, D. Effect of habitat conditions on content and quality of lipids in selected oat forms. Żywność. Nauka. Technologia. Jakość 2011, 3(76), 66–77. [Google Scholar] [CrossRef]
- Oleksiak, T.; Spyroglou, I.; Pacoń, D.; Matysik, P.; Pernisová, M.; Rybka, K. Effect of drought on wheat production in Poland between 1961 and 2019. Crop Sci. 2021, 62, 728–743. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Shakhatreh, Y.; Kafawin, O.; Ceccarelli, S.; Saoub, H. Selection of barley lines for drought tolerance in low rainfall areas. J. Agron. Sci. 2001, 186, 119–127. [Google Scholar] [CrossRef]
- Samarah, N.; Alqudah, A.; Amayreh, J.; McAndrews, G. The Effect of Late-terminal Drought Stress on Yield Components of Four Barley Cultivars. J. Agron. Crop Sci. 2009, 195, 427–441. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P. Global Synthesis of Drought Effects on Maize and Wheat Production. Public Library of Science ONE 2016, 11(5), e0156362. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Tanaka, H.; Eltayeb, A.; Tsujimoto, H. Wide hybridization between oat and pearl millet belonging to different subfamilies of Poaceae. Plant Reprod. 2013, 26, 25–32. [Google Scholar] [CrossRef]
- Skrzypek, E.; Warzecha, T.; Noga, A.; Warchol, M.; Czyczylo-Mysza, I.; Dziurka, K.; Marcinska, I.; Kaploniak, K.; Sutkowska, A.; Nita, Z.; Werwinska, K.; Idziak-Helmcke, D.; Rojek, M.; Hosiawa-Baranska, M. Complex characterization of oat (Avena sativa L.) lines obtained by wide crossing with maize (Zea mays L.). PeerJ 2018, 6, e5107. [Google Scholar] [CrossRef]
- Okagaki, R.; Kynast, R.; Livingston, S.; Russell, C.; Rines, H.; Phillips, R. Mapping maize sequences to chromosomes using oat-maize chromosome addition materials. Plant Physiol. 2001, 125, 1228–1235. [Google Scholar] [CrossRef]
- Munjal, R.; Dhanda, S. Physiological evaluation of wheat (Triticum aestivum L.) genotypes for drough tresistance. Indian J. Genet. Plant Breed. 2005, 65(4), 307–308. [Google Scholar]
- Sallam, A.; Alqudah, A.; Dawood, M.; Baenziger, S.; Börner, A. Drought stress tolerance in wheat and barley: advances in physiology, breeding and genetics research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef]
- Warzecha, T.; Skrzypek, E.; Sutkowska, A. Effect of Fusarium culmorum infection on the selected physiological and biochemical parameters of barley (Hordeum vulgare L.) DH lines. Physiol. Mol. Plant Pathol. 2015, 89, 62–69. [Google Scholar] [CrossRef]
- Warzecha, T.; Skrzypek, E.; Adamski, T.; Surma, M.; Kaczmarek, Z.; Sutkowska, A. Chlorophyll a fluorescence parameters of hulled and hull-less barley (Hordeum vulgare L.) DH lines inoculated with Fusarium culmorum. Plant Pathol. J. 2019, 35(2), 112–124. [Google Scholar] [CrossRef] [PubMed]
- Mohammadkhani, N.; Heidari, R. Drought-induced accumulation of soluble sugars and proline in two maize varieties. World Appl. Sci. J. 2008, 3(3), 448–453. [Google Scholar]
- Redillas, M.; Park, S.H.; Lee, J.; Kim, Y.; Jeong, J.; Jung, H.; Bang, S.; Hahn, T.R.; Kim, J.K. Accumulation of trehalose increases soluble sugar contents in rice plants conferring tolerance to drought and salt stress. Plant Biotechnol. Rep. 2011, 6, 89–96. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Differential responses of phenolic compounds of Brassica napus under drought stress. Iran. J. Plant Physiol. 2018, 8(3), 2417–2425. [Google Scholar]
- Sinay, H.; Karuwal, R. Proline and total soluble sugar content at the vegetative phase of six corn cultivars from Kisar Island Maluku, grown under drought stress conditions. Int. J. Advance Agricultural Res. 2014, 2, 77–82. [Google Scholar]
- Hura, T.; Hura, K.; Grzesiak, S. Leaf dehydration induces different content of phenolics and ferulic acid in drought-resistant and -sensitive genotypes of spring triticale. Verlag der Zeitschrift für Naturforschung 2014, 64(1–2), 85–95. [Google Scholar] [CrossRef]
- André, C.; Schafleitner, R.; Guignard, C.; Oufir, M.; Aliaga, C.; Nomberto, G.; Hoffmann, L.; Hausman, J.F.; Evers, D.; Larondelle, Y. Modification of the health-promoting value of potato tubers field grown under drought stress: emphasis on dietary antioxidant and glycoalkaloid contents in five native Andean cultivars (Solanum tuberosum L.). J. Agric. Food Chem. 2009, 57, 599–609. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.; Etminan, A.; Moghaddam, M.; Siddique, K. Physiological responses to drought stress in wild relatives of wheat: implications for wheat improvement. Acta Physiol. Plant. 2017, 39, 106. [Google Scholar] [CrossRef]
- Samarah, N. Effects of drought stress on growth and yield of barley. Agron. Sustainable Dev. 2005, 25, 145–149. [Google Scholar] [CrossRef]
- Marcińska, I.; Nowakowska, A.; Skrzypek, E.; Czyczyło-Mysza, I. Production of double haploids in oat (Avena sativa L.) by pollination with maize (Zea mays L.). Cent. Eur. J. Biol. 2013, 8, 306–313. [Google Scholar] [CrossRef]
- Czyczyło-Mysza, I.; Marcińska, I.; Skrzypek, E.; Bocianowski, J.; Dziurka, K.; Rančić, D.; Radošević, R.; Pekić-Quarrie, S.; Dodig, D.; Quarrie, S. Genetic analysis of water loss of excised leaves associated with drought tolerance in wheat. PeerJ 2018, 6, e5063. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.M.; McCaig, T.N. Evaluation of techniques for screening for drought resistance in wheat. Crop Sci. 1982, 22(3), 503–506. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Roberts, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagent. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Skomra, U.; Bocianowski, J.; Agacka, M. Agro-morphological differentiation between European hop (Humulus lupulus L.) cultivars in relation to their origin. J. Food Agric. Environ. 2013, 11(3&4), 1123–1128. [Google Scholar]
- VSN International Genstat for Windows 23rd Edition. VSN International, Hemel Hempstead, UK, 2023.
Figure 1.
Electrophoretic separation of PCR products generated with Grande 1 primers application (M – 100 bp marker, W – maize cv. Waza, S – oat cv. Stoper, 35 - OMA line, 36-41 - oat DH lines, 42-43 - OMA lines, 44-51 – oat DH lines).
Figure 1.
Electrophoretic separation of PCR products generated with Grande 1 primers application (M – 100 bp marker, W – maize cv. Waza, S – oat cv. Stoper, 35 - OMA line, 36-41 - oat DH lines, 42-43 - OMA lines, 44-51 – oat DH lines).
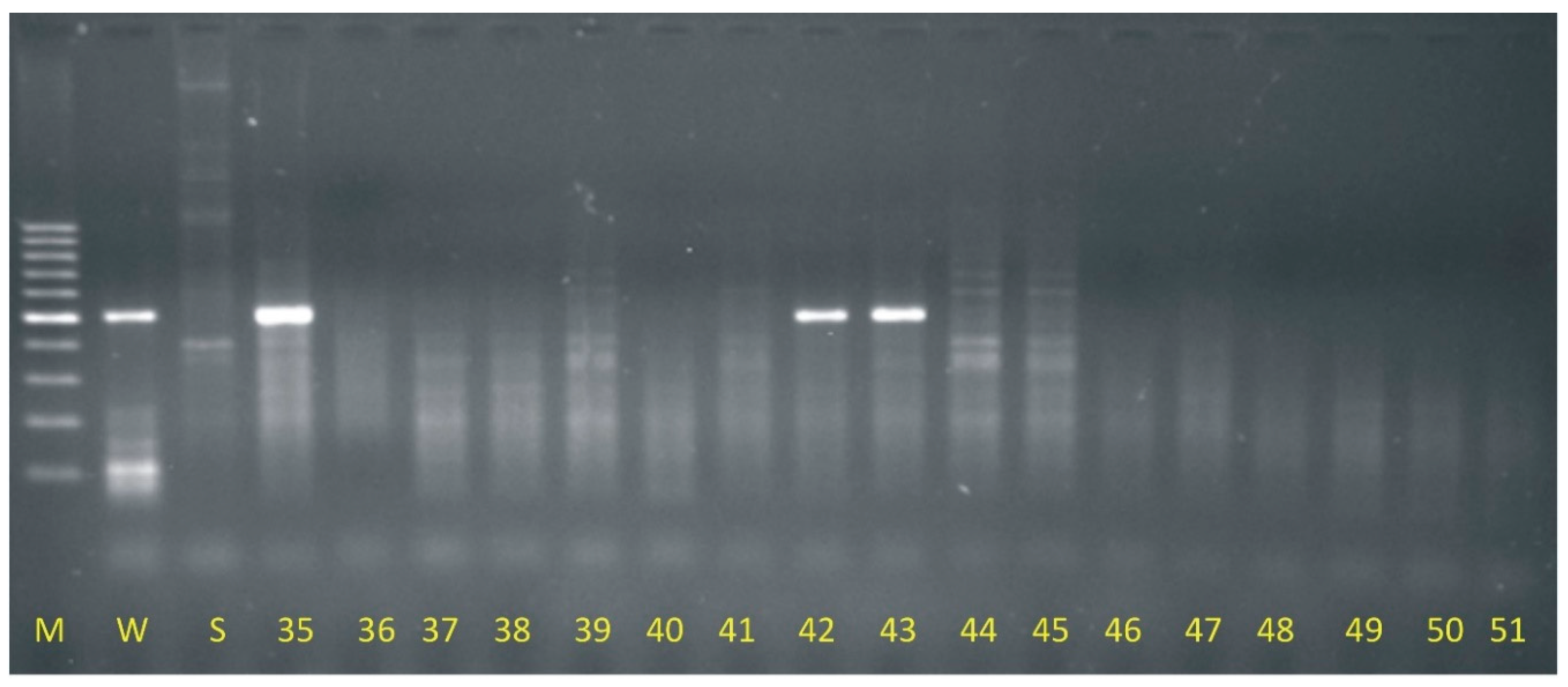
Figure 2.
A heatmap showing correlation coefficients between all pairs of observed traits in control. [t[1]—ELWL after 0-3 hours, t[2]—ELWL after 4-6 hours, t[3]—ELWL after 0-6 hours, t[4]—soluble sugars content in first day of drought (20% of soil field capacity), t[5]—phenolic compounds content in first day of drought (20% of soil field capacity), after two weeks of drought (maintaining 20% of soil field capacity), t[6]—soluble sugars content after two weeks of drought (maintaining 20% of soil field capacity), t[7]—phenolic compounds content after two weeks of drought (maintaining 20% of soil field capacity), t[8]—the mass of stems plant–1, t[9]—the number of grains, t[10]—the mass of grains plant–1].
Figure 2.
A heatmap showing correlation coefficients between all pairs of observed traits in control. [t[1]—ELWL after 0-3 hours, t[2]—ELWL after 4-6 hours, t[3]—ELWL after 0-6 hours, t[4]—soluble sugars content in first day of drought (20% of soil field capacity), t[5]—phenolic compounds content in first day of drought (20% of soil field capacity), after two weeks of drought (maintaining 20% of soil field capacity), t[6]—soluble sugars content after two weeks of drought (maintaining 20% of soil field capacity), t[7]—phenolic compounds content after two weeks of drought (maintaining 20% of soil field capacity), t[8]—the mass of stems plant–1, t[9]—the number of grains, t[10]—the mass of grains plant–1].
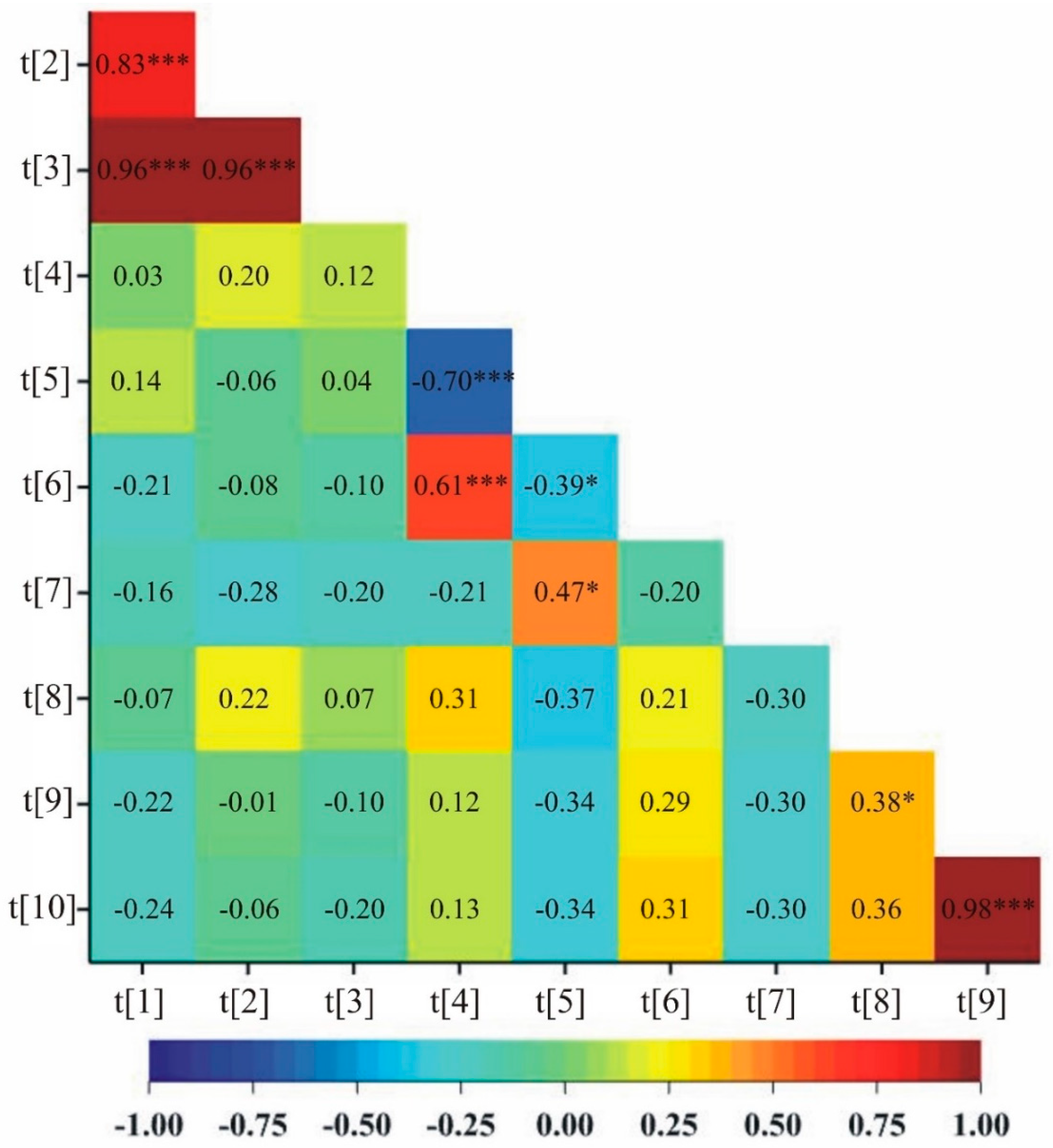
Figure 3.
A heatmap showing correlation coefficients between all pairs of observed traits in drought stress. [t[4]—soluble sugars content in first day of drought (20% of soil field capacity), t[5]—phenolic compounds content in first day of drought (20% of soil field capacity), after two weeks of drought (maintaining 20% of soil field capacity), t[6]—soluble sugars content after two weeks of drought (maintaining 20% of soil field capacity), t[7]—phenolic compounds content after two weeks of drought (maintaining 20% of soil field capacity), t[8]—the mass of stems plant–1, t[9]—the number of grains, t[10]—the mass of grains plant–1].
Figure 3.
A heatmap showing correlation coefficients between all pairs of observed traits in drought stress. [t[4]—soluble sugars content in first day of drought (20% of soil field capacity), t[5]—phenolic compounds content in first day of drought (20% of soil field capacity), after two weeks of drought (maintaining 20% of soil field capacity), t[6]—soluble sugars content after two weeks of drought (maintaining 20% of soil field capacity), t[7]—phenolic compounds content after two weeks of drought (maintaining 20% of soil field capacity), t[8]—the mass of stems plant–1, t[9]—the number of grains, t[10]—the mass of grains plant–1].
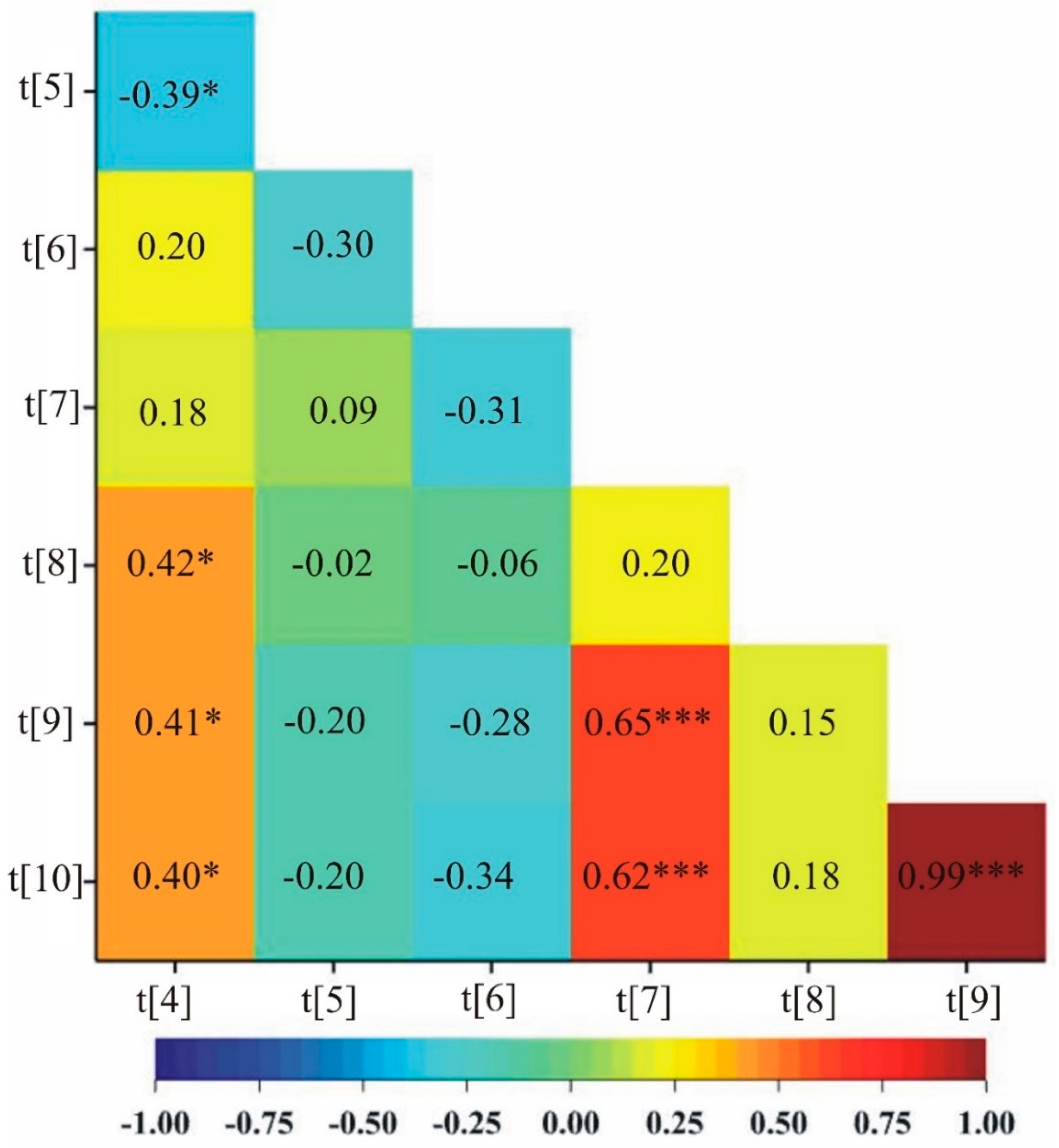
Figure 4.
Distribution of fourteen OMA lines in F2 and F3 generations and cultivar Bingo in the space of the first two principal components for control.
Figure 4.
Distribution of fourteen OMA lines in F2 and F3 generations and cultivar Bingo in the space of the first two principal components for control.

Table 1.
Mean values, standard deviations and homogeneous group for excised-leaf water loss.
| Trait | Excised-leaf water loss (ELWL) | ||||||
|---|---|---|---|---|---|---|---|
| after 0-3 hours | after 4-6 hours | after 0-6 hours | |||||
| Genotype | Mean | s.d. | Mean | s.d. | Mean | s.d. | |
| Bingo | 19.71c-g | 3.677 | 13.96c-e | 3.318 | 30.82b-e | 5.665 | |
| F2 | 114 | 22.80bc | 5.225 | 12.97c-f | 4.272 | 32.64b-d | 7.846 |
| 119 | 17.39e-h | 1.493 | 12.88c-f | 3.044 | 28.01b-f | 3.551 | |
| 12 | 16.55gh | 2.033 | 13.66c-e | 11.022 | 27.98b-f | 9.052 | |
| 18 | 28.32a | 5.774 | 19.33b | 2.403 | 42.08a | 6.264 | |
| 1b | 22.27b-d | 7.371 | 14.06c-e | 6.472 | 32.85b-d | 11.078 | |
| 23 | 15.26gh | 3.659 | 10.25e-h | 4.050 | 23.84e-g | 6.563 | |
| 26 | 15.43gh | 1.828 | 9.98e-h | 1.210 | 23.86e-g | 2.622 | |
| 35 | 21.34b-f | 3.189 | 16.41b-d | 2.724 | 34.21bc | 4.467 | |
| 42 | 16.26gh | 2.910 | 10.83e-h | 3.337 | 25.26e-g | 5.302 | |
| 43 | 18.88c-h | 6.296 | 11.07e-h | 6.225 | 27.57c-g | 10.275 | |
| 55 | 21.69b-e | 3.930 | 16.91bc | 4.978 | 34.80b | 7.022 | |
| 78b | 15.72gh | 0.820 | 10.05e-h | 0.751 | 24.20e-g | 0.110 | |
| 83 | 18.24d-h | 3.523 | 11.39e-h | 4.057 | 27.47c-g | 5.886 | |
| 9 | 15.58gh | 2.293 | 7.58gh | 1.295 | 21.96fg | 3.203 | |
| F3 | 114 | 22.80bc | 5.225 | 12.97c-f | 4.272 | 32.64b-d | 7.846 |
| 119 | 16.9f-h | 3.878 | 11.72d-h | 1.959 | 26.67d-g | 3.323 | |
| 12 | 17.05f-h | 3.994 | 9.46e-h | 2.458 | 24.82e-g | 5.661 | |
| 18 | 28.32a | 5.774 | 19.33b | 2.403 | 42.08a | 6.264 | |
| 1b | 17.24e-h | 2.189 | 8.51f-h | 3.023 | 24.25e-g | 4.328 | |
| 23 | 15.29gh | 1.644 | 8.43f-h | 0.910 | 22.42fg | 2.120 | |
| 26 | 16.87f-h | 1.483 | 12.28c-g | 2.539 | 27.05d-g | 3.351 | |
| 35 | 21.34b-f | 3.189 | 16.41b-d | 2.724 | 34.21bc | 4.467 | |
| 42 | 16.3gh | 4.258 | 10.10e-h | 3.367 | 24.65e-g | 6.52 | |
| 43 | 15.96gh | 2.685 | 11.32e-h | 3.181 | 25.39eg | 5.052 | |
| 55 | 16.74gh | 1.091 | 12.80c-f | 1.808 | 27.4c-g | 1.867 | |
| 78b | 25.62ab | 4.516 | 26.76a | 6.265 | 45.33a | 7.675 | |
| 83 | 18.22d-h | 0.497 | 11.61e-h | 0.751 | 27.72c-g | 0.173 | |
| 9 | 14.81h | 0.997 | 7.05h | 1.681 | 20.81g | 2.279 | |
| LSD0.05 | 4.533 | 4.768 | 7.07 | ||||
Table 2.
Mean values, standard deviations and homogeneous group for soluble sugars content (mg g–1 of d.w.).
Table 2.
Mean values, standard deviations and homogeneous group for soluble sugars content (mg g–1 of d.w.).
| Trait | First day of drought (20% of soil field capacity) | After two weeks of drought (maintaining 20% of soil field capacity) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | C | S | Average | C | S | Average | |||||||
| Genotype | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | |
| Bingo | 374.9 | 19.98 | 340.1 | 42.86 | 357.5bc | 36.96 | 263.9 | 77.13 | 162.1 | 8.13 | 213.0b-f | 74.64 | |
| F2 | 114 | 204.6 | 28.81 | 295.8 | 26.54 | 250.2g-k | 55.09 | 139.6 | 12.26 | 218.6 | 10.10 | 179.1f-j | 43.47 |
| 119 | 244.0 | 48.65 | 305.7 | 10.88 | 274.8e-i | 46.38 | 202.2 | 32.22 | 272.6 | 14.41 | 237.4a-c | 44.14 | |
| 12 | 160.1 | 19.47 | 352.3 | 114.43 | 256.2g-k | 127.8 | 142.5 | 22.68 | 206.6 | 3.88 | 174.5f-j | 37.45 | |
| 18 | 221.2 | 38.25 | 206.7 | 70.09 | 214.0jk | 52.84 | 122.0 | 16.74 | 264.8 | 37.10 | 193.4d-i | 80.89 | |
| 1b | 250.3 | 24.60 | 415.6 | 26.73 | 332.9b-e | 91.51 | 130.7 | 18.47 | 314.2 | 64.12 | 222.5b-e | 107.39 | |
| 23 | 419.7 | 29.45 | 548.0 | 45.13 | 483.8a | 77.09 | 243.4 | 20.49 | 219.9 | 34.43 | 231.6b-d | 29.09 | |
| 26 | 279.1 | 75.43 | 383.6 | 62.29 | 331.3bc-e | 84.98 | 150.4 | 32.41 | 183 | 38.38 | 166.7h-l | 37.22 | |
| 35 | 330.2 | 36.01 | 402.3 | 15.70 | 366.3b | 46.36 | 186.7 | 25.69 | 195.6 | 22.16 | 191.1e-i | 22.72 | |
| 42 | 225.1 | 41.08 | 241.1 | 31.87 | 233.1i-k | 35.09 | 101.9 | 22.33 | 79.9 | 4.82 | 90.9n | 19.02 | |
| 43 | 190.8 | 26.63 | 303.5 | 20.56 | 247.1h-k | 64.17 | 177.4 | 40.47 | 207.7 | 35.99 | 192.6d-i | 38.97 | |
| 55 | 310.6 | 30.05 | 379.5 | 39.92 | 345.1b-d | 49.27 | 147.3 | 4.55 | 204.7 | 21.07 | 176.0f-j | 33.73 | |
| 78b | 195.8 | 33.04 | 324.3 | 26.41 | 260g-k | 74.04 | 193.8 | 54.08 | 120.8 | 30.89 | 157.3i-l | 56.46 | |
| 83 | 167.4 | 44.50 | 437.7 | 48.72 | 302.5c-h | 150.79 | 199.1 | 33.07 | 279.7 | 38.16 | 239.4ab | 54.31 | |
| 9 | 186.5 | 42.29 | 356.6 | 57.23 | 271.5f-j | 102.13 | 142.2 | 35.16 | 204.7 | 13.83 | 173.4f-j | 41.58 | |
| F3 | 114 | 204.6 | 28.81 | 295.8 | 26.54 | 250.2g-k | 55.09 | 139.6 | 12.26 | 218.6 | 10.10 | 179.1f-j | 43.47 |
| 119 | 309.3 | 19.76 | 291.3 | 34.64 | 300.3c-h | 27.82 | 190.5 | 26.76 | 208.1 | 46.86 | 199.3c-h | 36.56 | |
| 12 | 269.3 | 66.51 | 383.4 | 29.55 | 326.4b-f | 77.43 | 188.5 | 42.90 | 153.8 | 33.26 | 171.2g-k | 40.08 | |
| 18 | 219.5 | 48.8 | 206.7 | 70.09 | 213.1k | 56.33 | 124.6 | 17.23 | 264.8 | 37.10 | 194.7d-i | 79.62 | |
| 1b | 210.6 | 29.86 | 360.2 | 38.35 | 285.4e-i | 86.03 | 151.8 | 30.76 | 261.4 | 61.75 | 206.6b-g | 73.99 | |
| 23 | 389.3 | 26.23 | 500.1 | 30.32 | 444.7a | 64.76 | 197.1 | 51.67 | 247.7 | 15.50 | 222.4b-e | 44.48 | |
| 26 | 201.2 | 25.52 | 349.7 | 45.73 | 275.5e-i | 86.46 | 120.7 | 6.20 | 141.9 | 23.68 | 131.3lm | 19.64 | |
| 35 | 330.2 | 36.01 | 402.3 | 15.70 | 366.3b | 46.36 | 186.7 | 25.69 | 195.6 | 22.16 | 191.1e-i | 22.72 | |
| 42 | 178.9 | 47.00 | 294.0 | 1.54 | 236.5i-k | 68.81 | 113.1 | 16.64 | 75.9 | 2.94 | 94.5mn | 22.75 | |
| 43 | 147.8 | 12.36 | 279.5 | 33.99 | 213.6jk | 74.25 | 152.7 | 32.65 | 103.3 | 0.00 | 128.0l-n | 33.97 | |
| 55 | 292.6 | 31.49 | 421.2 | 138.04 | 356.9bc | 115.38 | 175.9 | 8.81 | 243.0 | 7.91 | 209.4b-g | 36.70 | |
| 78b | 327.1 | 108.05 | 289.1 | 28.59 | 308.1b-g | 75.94 | 154.7 | 18.25 | 110.7 | 16.95 | 132.7k-m | 28.62 | |
| 83 | 301.9 | 113.68 | 300.4 | 75.66 | 301.1c-h | 89.40 | 193.3 | 43.89 | 354.7 | 0.00 | 274.0a | 90.95 | |
| 9 | 231.9 | 1.51 | 358.0 | 28.28 | 294.9d-h | 69.88 | 109.1 | 10.60 | 172.3 | 70.26 | 140.7j-l | 57.50 | |
| LSD0.05 | 58.341 | 39.71 | |||||||||||
Table 3.
Mean values, standard deviations and homogeneous group for phenolic compounds content (mg g–1 of d.w.).
Table 3.
Mean values, standard deviations and homogeneous group for phenolic compounds content (mg g–1 of d.w.).
| Trait | First day of drought (20% of soil field capacity) | After two weeks of drought (maintaining 20% of soil field capacity) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | C | S | Average | C | S | Average | |||||||
| Genotype | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | |
| Bingo | 19.41 | 3.568 | 23.22 | 1.856 | 21.31m | 3.380 | 36.60 | 2.218 | 41.76 | 2.873 | 39.18c-e | 3.639 | |
| F2 | 114 | 32.84 | 2.407 | 37.93 | 7.56 | 35.39ab | 5.862 | 41.35 | 6.317 | 32.29 | 1.910 | 36.82d-h | 6.489 |
| 119 | 26.96 | 3.464 | 32.62 | 3.352 | 29.79d-h | 4.371 | 35.87 | 2.412 | 30.07 | 0.640 | 32.97g-k | 3.504 | |
| 12 | 27.67 | 3.062 | 34.32 | 7.745 | 30.99c-f | 6.509 | 27.66 | 0.856 | 38.12 | 5.552 | 32.89g-k | 6.690 | |
| 18 | 30.13 | 2.423 | 24.74 | 4.503 | 27.43e-k | 4.417 | 27.45 | 3.816 | 25.26 | 3.168 | 26.35l | 3.451 | |
| 1b | 33.41 | 0.412 | 20.86 | 1.874 | 27.14e-k | 6.821 | 38.30 | 2.271 | 38.32 | 4.649 | 38.31c-f | 3.387 | |
| 23 | 24.79 | 1.780 | 25.67 | 2.016 | 25.23i-m | 1.823 | 31.24 | 2.349 | 33.48 | 5.414 | 32.36i-k | 4.043 | |
| 26 | 22.85 | 0.774 | 25.55 | 3.639 | 24.2k-m | 2.831 | 31.15 | 3.630 | 34.08 | 1.871 | 32.61h-k | 3.098 | |
| 35 | 26.74 | 3.360 | 24.59 | 1.854 | 25.67h-l | 2.762 | 31.56 | 2.153 | 31.36 | 4.996 | 31.46k | 3.563 | |
| 42 | 33.36 | 3.572 | 31.00 | 0.549 | 32.18b-d | 2.681 | 46.34 | 0.870 | 42.62 | 1.840 | 44.48ab | 2.395 | |
| 43 | 38.26 | 0.833 | 34.62 | 9.858 | 36.44a | 6.762 | 40.58 | 1.873 | 39.58 | 8.265 | 40.08b-d | 5.573 | |
| 55 | 23.25 | 1.679 | 24.79 | 2.318 | 24.02k-m | 2.049 | 36.02 | 0.280 | 35.26 | 5.003 | 35.64d-k | 3.305 | |
| 78b | 31.79 | 4.030 | 28.08 | 3.141 | 29.94d-g | 3.891 | 33.40 | 2.428 | 32.23 | 8.697 | 32.81h-k | 5.944 | |
| 83 | 28.56 | 3.085 | 25.30 | 1.191 | 26.93f-k | 2.778 | 32.51 | 2.456 | 40.88 | 2.626 | 36.70d-i | 5.053 | |
| 9 | 28.67 | 0.898 | 29.37 | 4.215 | 29.02d-j | 2.846 | 34.75 | 1.965 | 48.36 | 2.962 | 41.56a-c | 7.634 | |
| F3 | 114 | 32.83 | 2.711 | 37.93 | 7.56 | 35.38ab | 5.922 | 40.64 | 4.586 | 32.29 | 1.910 | 36.46d-j | 5.522 |
| 119 | 22.08 | 2.454 | 30.91 | 2.233 | 26.5g-l | 5.196 | 35.33 | 2.95 | 36.39 | 0.490 | 35.86d-k | 2.038 | |
| 12 | 27.83 | 4.926 | 29.38 | 2.532 | 28.61d-j | 3.719 | 34.73 | 1.461 | 39.92 | 3.809 | 37.33c-g | 3.849 | |
| 18 | 30.13 | 2.423 | 24.74 | 4.503 | 27.43e-k | 4.417 | 27.45 | 3.816 | 25.26 | 3.168 | 26.35l | 3.451 | |
| 1b | 32.02 | 4.865 | 26.52 | 5.512 | 29.27d-i | 5.639 | 32.92 | 1.200 | 40.03 | 1.269 | 36.48d-j | 3.971 | |
| 23 | 19.80 | 2.061 | 25.69 | 3.102 | 22.74lm | 3.984 | 32.60 | 2.509 | 32.92 | 0.895 | 32.76h-k | 1.753 | |
| 26 | 27.20 | 1.263 | 24.00 | 3.431 | 25.60i-l | 2.943 | 28.64 | 2.813 | 35.45 | 2.738 | 32.04jk | 4.456 | |
| 35 | 26.74 | 3.360 | 24.59 | 1.854 | 25.67h-l | 2.762 | 31.56 | 2.153 | 31.36 | 4.996 | 31.46k | 3.563 | |
| 42 | 34.16 | 2.533 | 28.27 | 1.407 | 31.21b-e | 3.678 | 42.74 | 4.226 | 36.74 | 3.238 | 39.74cd | 4.735 | |
| 43 | 37.11 | 1.394 | 31.57 | 1.703 | 34.34a-c | 3.291 | 38.20 | 5.292 | 31.88 | 0.000 | 35.04e-k | 4.841 | |
| 55 | 26.93 | 0.452 | 21.88 | 2.420 | 24.40k-m | 3.142 | 30.51 | 1.918 | 38.92 | 3.628 | 34.71f-k | 5.238 | |
| 78b | 24.50 | 2.914 | 30.05 | 1.902 | 27.27e-k | 3.741 | 31.90 | 2.957 | 44.79 | 3.590 | 38.35c-f | 7.536 | |
| 83 | 29.26 | 0.232 | 25.13 | 1.888 | 27.20e-k | 2.532 | 33.67 | 4.322 | 29.66 | 7.933 | 31.67k | 6.291 | |
| 9 | 20.48 | 3.051 | 29.52 | 4.478 | 25.00j-m | 5.993 | 32.78 | 0.823 | 57.93 | 8.004 | 45.35a | 14.44 | |
| LSD0.05 | 4.181 | 4.453 | |||||||||||
Table 4.
Mean values, standard deviations and homogeneous group for the mass of stems plant–1 (g), the number of grains and the mass of grains plant–1 (g) in control combination (C) and combination with soil drought (D).
Table 4.
Mean values, standard deviations and homogeneous group for the mass of stems plant–1 (g), the number of grains and the mass of grains plant–1 (g) in control combination (C) and combination with soil drought (D).
| Trait | The mass of stems/plant (g) | The number of grains | The mass of grains/plant (g) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | C | D | Average | C | D | Average | C | D | Average | ||||||||||
| Genotype | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | |
| Bingo | 17.16 | 3.157 | 4.29 | 3.089 | 10.72d-f | 7.300 | 124.25 | 17.34 | 47.75 | 31.13 | 86bc | 46.40 | 6.242 | 0.8845 | 1.833 | 1.5724 | 4.037b-d | 2.5885 | |
| F2 | 114 | 9.04 | 4.200 | 6.21 | 1.499 | 7.62f | 3.288 | 1.00 | 1.15 | 2.67 | 1.70 | 1.83i | 1.61 | 0.047 | 0.0543 | 0.091 | 0.0781 | 0.069k | 0.0665 |
| 119 | 14.87 | 2.860 | 10.56 | 1.717 | 12.72a-e | 3.172 | 8.50 | 3.00 | 7.75 | 2.87 | 8.12i | 2.75 | 0.420 | 0.0671 | 0.285 | 0.1125 | 0.353k | 0.1121 | |
| 12 | 16.57 | 3.515 | 9.13 | 1.212 | 12.85a-e | 4.66 | 90.75 | 39.51 | 76.25 | 21.39 | 83.50b-d | 30.42 | 3.558 | 1.4544 | 3.021 | 0.8043 | 3.290c-f | 1.1252 | |
| 18 | 13.57 | 2.224 | 5.72 | 1.501 | 9.65ef | 4.551 | 8.25 | 2.50 | 0.25 | 0.50 | 4.25i | 4.59 | 0.311 | 0.093 | 0.006 | 0.0115 | 0.159k | 0.1746 | |
| 1b | 17.29 | 2.151 | 12.99 | 2.872 | 15.14a | 3.286 | 76.75 | 12.69 | 35.50 | 21.11 | 56.12d-f | 27.32 | 3.464 | 0.4248 | 1.485 | 0.8583 | 2.475e-h | 1.2295 | |
| 23 | 14.95 | 2.448 | 11.34 | 0.772 | 13.15a-d | 2.557 | 58.75 | 17.21 | 37.00 | 4.69 | 47.88e-g | 16.48 | 2.423 | 0.7491 | 1.459 | 0.2433 | 1.941g-j | 0.7288 | |
| 26 | 13.71 | 3.182 | 9.19 | 1.199 | 11.45de | 3.282 | 44.50 | 17.18 | 43.25 | 10.63 | 43.88e-g | 13.24 | 1.998 | 0.6854 | 1.991 | 0.4541 | 1.994g-j | 0.5383 | |
| 35 | 14.83 | 1.180 | 7.84 | 1.514 | 11.33de | 3.944 | 69.25 | 23.91 | 58.25 | 10.40 | 63.75c-e | 18.05 | 2.962 | 0.8024 | 2.577 | 0.3947 | 2.769d-g | 0.6204 | |
| 42 | 14.06 | 1.962 | 7.92 | 2.904 | 10.99de | 4.005 | 16.00 | 14.21 | 25.75 | 15.13 | 20.88g-i | 14.55 | 0.782 | 0.8016 | 1.346 | 0.8691 | 1.064jk | 0.8306 | |
| 43 | 12.91 | 1.657 | 9.34 | 1.993 | 11.13de | 2.555 | 29.75 | 14.45 | 17.75 | 21.61 | 23.75g-i | 18.19 | 1.395 | 0.6518 | 0.89 | 1.1859 | 1.143i-k | 0.9262 | |
| 55 | 14.15 | 1.972 | 9.65 | 1.065 | 11.90b-e | 2.82 | 26.25 | 7.41 | 51.5 | 17.62 | 38.88e-h | 18.40 | 1.194 | 0.2872 | 2.19 | 0.7574 | 1.692g-j | 0.7514 | |
| 78b | 16.76 | 3.276 | 8.85 | 1.047 | 12.80a-e | 4.786 | 132.00 | 73.14 | 82.00 | 9.27 | 107.00ab | 55.17 | 5.089 | 2.3964 | 3.343 | 0.3876 | 4.216bc | 1.8430 | |
| 83 | 12.67 | 1.022 | 6.86 | 1.252 | 9.77ef | 3.28 | 139.50 | 36.57 | 104.00 | 22.38 | 121.75a | 33.88 | 5.442 | 1.0742 | 4.016 | 0.7586 | 4.729ab | 1.1498 | |
| 9 | 12.86 | 2.528 | 9.19 | 5.206 | 11.03de | 4.266 | 141.75 | 21.61 | 104.00 | 70.15 | 122.88a | 52.12 | 7.041 | 2.7310 | 3.978 | 2.709 | 5.510a | 3.0038 | |
| F3 | 114 | 9.04 | 4.200 | 6.21 | 1.499 | 7.62f | 3.288 | 1.00 | 1.15 | 2.67 | 1.70 | 1.83i | 1.61 | 0.047 | 0.0543 | 0.091 | 0.0781 | 0.069k | 0.0665 |
| 119 | 18.89 | 6.158 | 11.14 | 1.247 | 15.01ab | 5.837 | 9.25 | 3.20 | 9.75 | 1.50 | 9.50hi | 2.33 | 0.382 | 0.0910 | 0.364 | 0.0409 | 0.373k | 0.0661 | |
| 12 | 13.12 | 2.378 | 9.06 | 0.344 | 11.09de | 2.678 | 103.25 | 25.22 | 74.75 | 15.22 | 89.00bc | 24.58 | 3.975 | 1.0943 | 2.945 | 0.5318 | 3.460c-e | 0.9683 | |
| 18 | 13.57 | 2.224 | 5.72 | 1.501 | 9.65ef | 4.551 | 8.25 | 2.50 | 0.25 | 0.50 | 4.25i | 4.59 | 0.311 | 0.093 | 0.006 | 0.0115 | 0.159k | 0.1746 | |
| 1b | 17.26 | 4.163 | 12.42 | 0.823 | 14.84a-c | 3.798 | 62.75 | 31.61 | 47.50 | 13.38 | 55.12d-f | 23.90 | 2.692 | 1.3115 | 2.034 | 0.6319 | 2.363e-i | 1.0159 | |
| 23 | 18.02 | 1.880 | 9.57 | 1.597 | 13.79a-d | 4.798 | 76.25 | 18.08 | 27.50 | 8.35 | 51.88e-g | 29.14 | 3.044 | 0.6402 | 1.067 | 0.3224 | 2.055f-j | 1.1560 | |
| 26 | 13.38 | 1.545 | 9.35 | 0.906 | 11.36de | 2.453 | 49.25 | 14.48 | 39.75 | 5.56 | 44.50e-g | 11.35 | 2.411 | 0.7548 | 1.817 | 0.2941 | 2.114f-j | 0.6180 | |
| 35 | 14.83 | 1.180 | 7.84 | 1.514 | 11.33de | 3.944 | 69.25 | 23.91 | 58.25 | 10.4 | 63.75c-e | 18.05 | 2.962 | 0.8024 | 2.577 | 0.3947 | 2.769d-g | 0.6204 | |
| 42 | 11.77 | 1.970 | 7.99 | 6.697 | 9.88ef | 4.997 | 18.50 | 13.58 | 27.75 | 32.95 | 23.12g-i | 23.85 | 0.726 | 0.5186 | 1.305 | 1.6926 | 1.016jk | 1.1996 | |
| 43 | 15.06 | 2.137 | 8.41 | 0.238 | 11.74c-e | 3.819 | 43.00 | 42.58 | 48.00 | 16.17 | 45.50e-g | 29.93 | 1.909 | 1.8288 | 2.281 | 0.4809 | 2.095f-j | 1.2539 | |
| 55 | 13.85 | 2.33 | 7.48 | 0.970 | 10.66d-f | 3.788 | 54.25 | 24.30 | 57.00 | 11.34 | 55.62d-f | 17.61 | 2.498 | 1.0145 | 2.317 | 0.6464 | 2.407e-h | 0.7934 | |
| 78b | 19.80 | 2.438 | 7.36 | 0.687 | 13.58a-d | 6.851 | 167.50 | 56.97 | 84.75 | 18.39 | 126.12a | 59.10 | 6.179 | 1.1685 | 3.314 | 0.6160 | 4.747ab | 1.7583 | |
| 83 | 6.25 | 6.306 | 2.13 | 2.933 | 4.19g | 5.056 | 48.00 | 55.43 | 11.50 | 22.34 | 29.75f-i | 43.72 | 2.151 | 2.4832 | 0.418 | 0.8081 | 1.284h-k | 1.9442 | |
| 9 | 13.64 | 1.701 | 8.38 | 1.909 | 11.01de | 3.273 | 119.00 | 26.14 | 91.25 | 23.41 | 105.12ab | 27.35 | 4.724 | 0.8791 | 3.448 | 1.1002 | 4.086bc | 1.1468 | |
| LSD0.05 | 3.211 | 29.80 | 1.241 | ||||||||||||||||
Table 5.
The effect of excised-leaf water loss (ELWL) values, independent: after 0-3 hours, after 4-6 hours and after 0-6 hours (independent variable, x), on the mass of stems plant–1, the number of grains, the mass of grains plant–1 (dependent variable, y) estimated by regression model y=a x + b.
Table 5.
The effect of excised-leaf water loss (ELWL) values, independent: after 0-3 hours, after 4-6 hours and after 0-6 hours (independent variable, x), on the mass of stems plant–1, the number of grains, the mass of grains plant–1 (dependent variable, y) estimated by regression model y=a x + b.
| Independent variable | Dependent variable | Model | Percentage variance accounted | Standard error of observations |
|---|---|---|---|---|
| ELWL after 0-3 hours | The mass of stems plant–1 | y=21.53*** –0.314* x | 3.0 | 4.87 |
| The number of grains | y=20.184*** –0.028* x | 3.2 | 4.86 | |
| The mass of grains plant–1 | y=20.109*** –0.637* x | 2.6 | 4.88 | |
| ELWL after 4-6 hours | The mass of stems plant–1 | y=15.14*** –0.285 x | 2.0 | 5.23 |
| The number of grains | y=13.051*** –0.0057 x | - | 5.3 | |
| The mass of grains plant–1 | y=12.935*** –0.074 x | - | 5.3 | |
| ELWL after 0-6 hours | The mass of stems plant–1 | y=33.23*** –0.500* x | 3.0 | 7.77 |
| The number of grains | y=30.39*** –0.0286 x | 0.8 | 7.86 | |
| The mass of grains plant–1 | y=30.23*** –0.611 x | 0.4 | 7.88 |
* p<0.05; *** p<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



