
Submitted:
02 August 2023
Posted:
03 August 2023
You are already at the latest version
Abstract
The basilar arterial endothelium mediates blood-vessel relaxation, partly through release of nitric oxide (NO). Apoptosis of cerebrovascular endothelial cells is linked to high mortality rate in chickens infected with highly pathogenic avian influenza virus, but interestingly ducks exhibit greater resistance to this virus. In this study, we examined the responsiveness of duck basilar arteries (BAs) to various vasoactive substances, including 5-hydroxytryptamine (5-HT), histamine (His), angiotensin (Ang) II, noradrenaline (NA), acetylcholine (ACh), and avian bradykinin ornithokinin (OK), aiming to characterize the receptor subtypes involved and the role of endothelial NO in vitro. Our findings suggest that arterial contraction is mediated by 5-HT1 and H1 receptors, while relaxation is induced by β3-adrenergic and M3 receptors. Additionally, OK elicited a biphasic response in duck BAs, and Ang II had no effect. Endothelial NO appears to be crucial in relaxation mediated by M3 and OK receptors, but not β3-adrenergic receptors, in the duck BA. The reduced endothelial NO involvement in the receptor-mediated relaxation response in duck BAs represents a clear difference from the corresponding response reported in chicken BAs. This physiological difference may explain differences in lethality between ducks and chickens when vascular endothelial cells are infected by the virus.
Keywords:
cerebral artery
; duck
; endothelium
; highly pathogenic avian influenza
; nitric oxide
1. Introduction
The duck’s vascular endothelium may hold the key to answering an intriguing question for poultry scientists: why are ducks markedly more resilient to highly pathogenic avian influenza virus (HPAIV) than chickens? HPAIV is frequently fatal to gallinaceous poultry including chickens, but this seems not to be the case for ducks [5]. Interestingly, according to one report, no wild duck from the genus Anas experimentally infected with HPAIV showed symptoms or succumbed to the disease [15]. By contrast, the vascular endothelium has been implicated in fatal HPAIV outcomes in chickens. This lethality has been linked to apoptosis of vascular endothelial cells in the liver, kidney, and brain [13,36]. This acute apoptosis is reportedly associated with high levels of endothelial nitric oxide (NO) release during HPAIV infection, as NO acts with other damaging oxidants to promote excessive inflammation [2]. From a physiological perspective, HPAIV infection and the associated vascular endothelial apoptosis in chickens may be exacerbated by the high levels of NO released locally. Considering human physiology, NO can promote proinflammatory effects, including increased vascular permeability, cytotoxicity, and inflammatory cell infiltration, when present at high concentrations [3,28], and any related loss of endothelial function in vital arteries could quickly prove fatal.
As a potential site of HPAIV-related vascular endothelial injury, the basilar artery (BA) may provide a fruitful line of research. It is a vital artery that constantly supplies blood to the hindbrain, and a common cerebrovascular feature across vertebrates, contributing to maintenance of cerebral circulatory volume [26]. Its role in birds appears to be the same as that in mammals, although chickens are the only avian species in which BA reactivity has been studied [16,25,37].
In reports on the vascular response in the BA in chickens, the cerebrovascular endothelium has been identified as a powerful source of spontaneous NO release, based on strong contractions noted after application of Nω-nitro-L-arginine (L-NNA), a NO synthase (NOS) inhibitor [25]. Furthermore, previous investigations of endothelial cell receptors have indicated that beta-3 adrenergic receptors (β3-ARs), histamine (His) H1 receptors, and muscarinic acetylcholine (ACh) M3 receptors are involved in the relaxation of the chicken BA via the NO pathway, suggesting this arterial relaxation is strongly dependent on endothelial release of NO [16,25,37].
Until now, studies on the BA in poultry species have only targeted chickens, but differences in this artery and the local vascular response between chickens and ducks can be hypothesized. Evolutionarily speaking, ducks and chickens are separated by a distance of 90 million years and avian genomes tend to be more strongly conserved than mammalian genomes [27,32]. We thus considered that ducks may have a less complex vascular endothelium than their younger avian relatives, chickens. Interestingly, HPAIV reportedly rarely shows endothelial tropism in either wild or domestic ducks [31,33]. However, there is a paucity of evidence on the cerebrovascular endothelium in ducks.
Against this background, we aimed to characterize the BA in ducks, by investigating its capacity for endothelial NO release, and its response to a range of vasoactive substances we had previously used in our studies of chicken BA.
2. Materials and Methods
2.1. Tissue preparation
BAs were obtained from freshly slaughtered Aigamo ducks (Anas platyrhynchos / Anas platyrhynchos var. domesticus hybrid; n =71, both sexes, weight: 2.97 ± 0.04 kg). The ducks had been raised for meat and allowed to forage on insects in rice paddies, in farms in Kagoshima prefecture, Japan, and were slaughtered in accordance with the relevant Japanese law on agricultural animals. The sampled arteries were transferred to our laboratory in ice-cold physiological saline (119 mM NaCl, 4.7 mM KCl, 1.6 mM CaCl2, 1.2 mM MgCl2, 25 mM NaHCO3, 1.2 mM KH2PO4, and 10 mM glucose, pH 7.4) aerated with carbogen [95% (vol/vol) O2, 5% (vol/vol) CO2]. Each artery was immediately dissected free of adherent tissues under a stereomicroscope. All experiments were performed in accordance with the Guidelines for Animal Experiments of Kagoshima University.
2.2. Reagents
The following reagents were used at the described concentrations: His hydrochloride (10-6–10-3 M, Sigma-Aldrich, St. Louis, MO, USA), isoproterenol (10-9–10-5 M, Sigma-Aldrich), ketanserin tartrate (10-6 M, Sigma-Aldrich), methiothepin maleate (10-8–10-6 M, Sigma-Aldrich), diphenhydramine hydrochloride (10-7–10-5 M, Sigma-Aldrich), cimetidine (10-5 M, Sigma-Aldrich), angiotensin (Ang) II acetate salt (10-8–10-4 M, Sigma-Aldrich), butoxamine hydrochloride (10-6 M, Sigma-Aldrich), SR 59230A (10-7–10-6 M, Sigma-Aldrich), L-NNA (10-4 M, Sigma-Aldrich), noradrenaline (NA, 10-9–10-4 M, Tokyo Chemical Industry, Tokyo, Japan), phentolamine mesylate (10-5 M, Tokyo Chemical Industry), 5-hydroxytryptamine (5-HT)-creatinine sulfate (10-8–10-4 M; Merck, Darmstadt, Germany), adrenaline (10-9–10-5 M, Daiichi Sankyo, Tokyo, Japan), ACh chloride (10-9–10-5 M, Daiichi Sankyo), procaterol (10-9 –10-5 M, Fujifilm-Wako, Tokyo, Japan), atenolol (10-6–10-5 M, LKT Laboratories, Tokyo, Japan), hexahydro-sila-difenidol hydrochloride, p-fluoroanalog (pFHHSiD, 10-7, 10-6 M; Research Biochemical, Natick, MA, USA), avian bradykinin ornithokinin (OK, 10-9–10-5 M, BEX Co. Ltd., Tokyo, Japan), and sodium nitroprusside (SNP, 10−4 M; Nacalai Tesque, Kyoto, Japan). All reagents were used in accordance with the relevant manufacturer’s instructions.
2.3. Functional study
Three or four rings, approximately 2 mm wide, were cut from each duck BA. Each ring was mounted horizontally between two L-shaped stainless-steel holders (outer diameter 0.03 mm), with one part fixed to an isometric force transducer. The mounted ring was then immersed in a 4-mL water-jacketed micro-tissue organ bath (UMTB-1, Unique Medical Co. Ltd., Tokyo, Japan), containing oxygenated physiological saline at 41°C (pH 7.4). Each suspended ring was left to equilibrate for at least 30 min under a resting tension of 0.5 mN. This tension was chosen to facilitate induction of maximal contractions in the artery without deflection of the steel. KCl (60 mM) was applied every 30 min until the amplitude of the contraction reached a constant value.
Changes in the KCl concentration of the physiological saline were compensated for by equimolar adjustment of the NaCl concentration. The isometric tension was recorded with an amplifier (AP-621G, Nihon Kohden Kogyo, Tokyo, Japan), digitized with an analog-digital converter (PowerLab/8SP, AD Instruments Co., Castle Hill, NSW, Australia), and stored on the hard disk of a personal computer. In a preliminary experiment, 5-HT induced sufficient contraction and retained the tonic phase, whereas other vasoconstrictors, such as Ang II and NA, failed to induce contractions. Therefore, we used 5-HT to induce the precontracted condition before applying vasodilators. The cumulative concentration-response curve of agonists was obtained by adding a solution of each agonist directly to the fluid in the bath. Antagonists or inhibitors were added to the bathing media 30 min before each agonist. The antagonists had no effect on the resting vascular tone. The log concentration-ratio of EC50 values (i.e., the concentration producing a half-maximal response) in the absence or presence of an antagonist was calculated and plotted against the logarithm of the antagonist concentration to obtain the pA2 values [1].
At the end of the relaxant response, SNP (10−4 M) was applied to produce maximal relaxation, which was taken as 100%.
2.4. Statistical analysis
Results are expressed as the mean ± SEM, and statistical analyses were performed using Student’s t-test or the Bonferroni test after one-way ANOVA (Stat View J-4.5, Abacus Concepts Inc., Berkeley, CA, USA). P-values < 0.05 were considered statistically significant.
3. Results
3.1. Maintenance of arterial resting tone by spontaneous NO and PGs
To elucidate the resting tone balance of the BA, the effects of L-NNA, an NOS inhibitor, and indomethacin, a cyclo-oxygenase inhibitor, on resting tone were investigated. The real-time vasomotive tension changes induced by L-NNA (10−4 M)- and indomethacin (10−5 M) are illustrated in Figure 1. L-NNA induced contraction (30.5 ± 5.4% to 60 mM KCl, n = 4 ducks) under resting tension, and indomethacin induced relaxation (-36.1 ± 2.1% to 10−4 M SNP, n = 4 ducks) under contraction induced by L-NNA. The contraction induced by 60 mM KCl was 1.84 ± 0.04 mN.
3.2. Responsiveness to vasoactive substances
To evaluate which of receptor subtypes is predominant in duck BAs, we investigated the effect of a range of substances with a known vasoactive effect in chickens and other species. Specifically, the agents used to generate effects on the vascular endothelium were 5-HT, His, Ang II, ACh, NA [a beta-1 (β1) and non-selective alpha (α) AR agonist], and OK (an avian bradykinin receptor agonist). Contractile response was measured under a resting tension condition. Under this condition, 5-HT and His induced contraction in concentration-dependent manners with respective half-maximal effective concentrations (logarithmically adjusted; hereafter, pEC50 values) of 5.84 ± 0.06 and 4.37 ± 0.09, whereas no effect was noted for NA in the absence or presence of propranolol (a non-selective β-AR antagonist, 10-5 M), or Ang II (Figure 2A and Table 1). Relaxation response was measured under precontraction with 5-HT. Following precontraction, ACh, NA, and OK induced relaxation in a concentration-dependent manner, yielding respective pEC50 values of 6.18 ± 0.17, 6.28 ± 0.05, and 6.20 ± 0.25. Phentolamine (10−5 M), a non-selective α-AR antagonist, had no significant effect on the response to NA (Figure 2B and Table 1).
3.3. Involvement of 5-HT receptor subtype
To ascertain the predominant 5-HT receptor subtype in the duck BA, we investigated the effects of methiothepin (a non-selective 5-HT receptor antagonist) and ketanserin (a 5-HT2 receptor selective antagonist) on 5-HT-induced contraction. The Schild plot for methiothepin showed a slope of 1.16 ± 0.19 (Figure 3C), which did not significantly diverge from unity. Its calculated pA2 value was 8.52 ± 0.17, whereas ketanserin (10-6 M) had no significant effect for the 5-HT-induced contraction in the duck BAs (Figure 3B).
3.4. Involvement of His receptor subtype
To ascertain the predominate His receptor subtype in the duck BA, we investigated the effect of diphenhydramine (a H1 receptor antagonist) and cimetidine (a H2 receptor selective antagonist) on His-induced contraction. The Schild plot for diphenhydramine (10-7–10-5 M) showed a slope of 1.09 ± 0.15 (Figure 4B), which did not significantly diverge from unity. Its calculated pA2 value was 6.89 ± 0.13, whereas cimetidine (10-5 M) had no significant effect (Figure 4).
3.5. Responsiveness to β-ARs agonists
To elucidate the roles of β-ARs in the duck BA, we attempted to generate the relaxation effects of different β-AR agonists, and monitored the vascular response. The agents applied were isoproterenol (a non-selective β-AR agonist), NA (a β1-AR and non-selective α-AR agonist), adrenaline (a non-selective β-AR and non-selective α-AR agonist), and procaterol (a β2-AR agonist). Three of the four β-AR agonists induced concentration-dependent relaxation of the duck BAs; the exception was procaterol (Figure 5). Their rank order of potency as inducers of relaxation was isoproterenol > NA > adrenaline (Table 1). None of these agents induced contraction.
3.6. Involvement of β-AR subtype
To ascertain the predominant β-AR subtype in the duck BA, we investigated the effects of atenolol (a β1-AR antagonist), butoxamine (a β2-AR antagonist), and SR 59230A (a β3-AR antagonist) on isoproterenol-induced relaxation. In this experiment, atenolol (a β1-AR antagonist, 10-6 M) and butoxamine (a β2-AR antagonist, 10-6 M) had no significant effect on isoproterenol-induced relaxation (Figure 6A). In contrast, only SR 59230A effectively antagonized the isoproterenol-induced relaxation and its slope of Schild plot was 1.18 ± 0.11, which did not significantly diverge from unity (Figure 6C). Its calculated pA2 value was 7.03 ± 0.08. Furthermore, L-NNA (10-4 M) did not affect the isoproterenol-induced relaxation in duck BAs (data not shown).
3.7. Involvement of muscarinic ACh receptor subtype
To ascertain the predominant muscarinic ACh receptor subtype in the duck BA, we investigated the effects of atropine (a non-selective muscarinic ACh receptor antagonist), pirenzepine (a muscarinic ACh M1 receptor selective antagonist), methoctramine (a muscarinic ACh M2 receptor selective antagonist), and pFHHSiD (a muscarinic ACh M3 receptor selective antagonist) on ACh-induced contraction. Atropine shifted the concentration-response curve for ACh to the right at 10-8 M, and largely abolished ACh-induced relaxation at 10-7 M (Figure 7A). For selective muscarinic antagonists, the respective slopes of Schild plots for pirenzepine and pFHHSiD were 1.06 ± 0.10 and 0.97 ± 0.08, neither of which significantly diverged from unity. The respective pA2 values yielded by pirenzepine and pFHHSiD were 6.52 ± 0.12 and 8.06 ± 0.13 (Figure 7E,F), whereas methoctramine did not affect the ACh-induced relaxation in duck BAs (data not shown). L-NNA mostly abolished ACh-induced relaxation of duck BAs (Figure 7D).
3.8. Effects of L-NNA and indomethacin on OK-induced relaxation
To investigate OK-induced relaxation in the duck BA, we applied L-NNA and L-NNA plus indomethacin. In the presence of L-NNA (10-4 M), OK-induced relaxation was attenuated significantly (p < 0.01) and even transferred to contraction at 10-5 M, whereas indomethacin (10-5 M) abolished this contraction (Figure 8).
4. Discussion
The present study is the first to demonstrate the responsiveness of BAs to vasoactive substances in ducks. We showed that NO is a diminished endothelial mediator in ducks, relative to its reported role in chickens. This information will prove especially useful for comparisons between ducks, which are not prone to disease and death after HPAIV infection, and chickens, which are.
We demonstrated the involvement of spontaneously released endothelial NO, based on the contraction induced by the NOS inhibitor L-NNA, followed by the relaxation induced by indomethacin, which inhibits cyclooxygenase, under resting tension (Figure 1). Our findings in ducks are similar to results obtained in some mammalian species including dogs [30], pigs and cattle [17], horses [34], and bats [9]. In porcine BAs, further studies demonstrated that spontaneous NO is released from endothelial cells [18] and spontaneous thromboxane A2, which is formed from PGH2 and inhibited by indomethacin, is released from endothelial and smooth muscle cells [18,21]. The mechanism of maintenance of cerebrovascular tone in the duck partially resembles that in the chicken. However, a key difference was that the L-NNA-induced contraction in ducks (30.5%) was markedly weaker than that in chicken (122.1%) [25], suggesting that duck shows less involvement of spontaneous endothelial NO than chicken.
We also investigated the receptor subtypes involved in contraction and relaxation, and their location (smooth muscle or endothelial cells) in the duck BA.
5-HT receptors appear to play a similar role in ducks and chickens, as the receptor agonist induced a concentration-dependent contraction in isolated duck BAs, in both the avian species we evaluated, and we found that the 5-HT1 receptor may be the dominant receptor of this subtype (Figure 3). Based on an evaluation of half-maximal-effect concentrations, we found that the 5-HT receptor antagonist, methiothepin, produced a similar effect in the duck BA to that observed in the chicken BA and the rabbit saphenous vein, where vascular response (vasocontraction) is mediated via activation of 5-HT1 receptor 16, 35]. Furthermore, the effects of methiothepin and the 5-HT2 antagonist ketanserin in the duck BA (respective rightward shifts in the concentration-response curve of 35.5-fold and 2.7-fold) were similar to the corresponding shifts (30-fold and 3-fold, respectively) we previously observed in the chicken BA [16], suggesting ducks and chickens share a similar characterization for this receptor type.
We also profiled the involvement of His receptors. The His-induced concentration-dependent contraction in isolated duck BAs in this study resembled those observed in porcine, bovine, equine, and dolphin BAs, with a similar half-maximal effect to that in the bovine report, but a smaller half-maximal effect than those in the porcine, equine, and cetacean reports [11,22,23]. The H1-receptor antagonist diphenhydramine (10-7–10-5 M) produced an effect of similar extent in duck and dolphin BAs, but its effect in ducks was smaller than in the bovine and porcine reports [22,23]. In line with the known vascular response in these mammalian species, we consider that H1 receptor activation induces the contraction of the duck BA. Our His receptor findings present a contrast to those previously reported in chickens. Okuno et al. (2008) reported that both H1 and H2 receptors, which are located on respective endothelial cells and smooth muscle cells, are involved in the relaxation of the chicken BA. However, in ducks, H1 receptors play a dominant role in His-induced contraction, while H2 receptor were not involved.
For adrenergic receptors, noradrenaline had no effect on BA at resting tension and phentolamine (a non-selective α-ARs antagonist) had no effect on noradrenaline-induced relaxation under precontraction (Figure 2), suggesting that α-ARs are not involved in duck BA reactivity. This presents a contrast with the chicken BA, where α-ARs are present and involved in contraction, and phentolamine inhibits this contraction [37]. For β-ARs, the duck BA appears to possess β1-ARs, but with a smaller population for this receptor subtype than in chickens, based on the similar potency ranking but smaller effect size for isoproterenol and NA [37]. β2-ARs appear not to play a major role in the duck BA, although they are present as a non-dominant subtype in chickens, based on findings for the effect of procaterol [37]. Interestingly, the effect of the β-AR agonist isoproterenol-induced relaxation was not inhibited by the NOS inhibitor L-NNA (data not shown); therefore, we suggest that NO plays no role in β-AR mediation in ducks, which represents a physiological difference from chickens.
We consider that the β3-AR is the predominant β-AR subtype in the duck BA, based on results obtained with a range of β-AR antagonists (Figure 6). Most pertinently, only the β3-AR antagonist SR 59230A demonstrated an effective antagonistic effect (pA2 value: 7.03) in duck BAs comparable to that in rat urinary bladder tissue (7.27) and guinea pig gastric fundus (7.35), where β3-ARs are known to predominate [8,24]. Accordingly, we suggest that the β3-AR is predominantly involved in isoproterenol-induced relaxation in the duck BA, a similar phenomenon to that we reported for chickens. However, this relaxation is endothelium dependent in chickens, but not in ducks.
Our findings for muscarinic ACh receptors (Figure 7) resembled those observed in mouse, chicken, and bat BAs [9,12,16], suggesting their activation is involved in the response to ACh. We further suggest that the M3 receptor is the predominant subtype involved in ACh-induced relaxation, and M1 and M2 receptors may not be involved in duck BAs. We found no effect for the M2 antagonist methoctramine and only a weak effect for the M1 antagonist pirenzepine. The latter effect was smaller than that reported in cat cerebral arteries (8.08), where muscarinic M1 receptor predominate [4]. However, the M3 antagonist pFHHSiD yielded a calculated pA2 value (8.06) similar to those reported in bovine coronary arteries (7.64), human uterine artery (8.17), and chicken BAs (7.55), where the muscarinic M3 receptor predominate [6,14,16]. In contrast to our findings on β-ARs, we consider that NO is involved in muscarinic M3 receptor-mediated relaxation in ducks, as it is in other species including mice, bats, and chickens [9,12,37], based on the abolition of the ACh-induced relaxation by the NO inhibitor L-NNA seen in this study (Figure 7D).
We revealed that avian bradykinin OK induced relaxation and contraction in duck BAs, by regulation of respective NO and cyclooxygenase pathways. In porcine BAs, bradykinin produced NO and prostaglandin F2α then induced relaxation followed by contraction [10,19]. OK receptors are the avian homolog to mammalian bradykinin receptors, and are known to be involved in blood vessel dilatation, smooth muscle contraction, increased vascular permeability, and inflammation [29]. OK receptors reportedly potentiate pro-inflammatory responses in chicken macrophages [7]. Pharmacological research on these receptors is hampered by a lack of established antagonists; accordingly, it was not possible to investigate the relevant receptor subtypes in this study. However, we anticipate this line of research will be enabled by advances in cell culture in the future.
BA responsiveness to vasoactive substances is known to differ widely between mammalian species, according to differences in receptor subtypes and their distribution on smooth muscle or endothelial cells. To our knowledge, BA responses have never shown fully identical characteristics in any two mammalian species, and we speculate that similar diversity will be observable across avian species, based on the present study, which is the second characterization of BA response in an avian species. However, one feature common to ducks in this study and to chickens in our previous report was β3-AR-mediated vasodilation, which has never been reported in mammalian cerebrovascular systems [20]. We speculate that β3-ARs may play an important role in the cardiovascular system of birds, and this represents an interesting line of future research.
5. Conclusions
In conclusion, we suggest that the BA of the duck is characterized by a smaller endothelial release of NO, and a smaller degree of endothelial involvement in its reactivity, than the BA of the chicken. 5-HT1 and H1 receptors may be involved in arterial contraction, and β3 and M3 receptors may be involved in relaxation. NO plays a role with M3, but not β3-adrenergic receptors. This physiological difference may help to explain why severe effects (including death) of HPAIV infection are seen in chickens, but not ducks.
Author Contributions
Conceptualization, S.W. and A.M.; methodology, M.Z.I. and H.T.T.N.; formal analysis, S.W. and A.M.; investigation, S.W., T.O. and R.S.; resources, A.M., Y.U. and M.S.; data curation, S.W.; writing—original draft preparation, S.W. and A.M.; writing—review and editing, H.S., Y.U., M.S., A.M. and T.O.; supervision, A.M.; project administration, A.M.; funding acquisition, A.M. and H.T.T.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by JPSP KAKENHI Grant Number 16580242 and Vietnam National Foundation for Science and Technology Development (NAFOSTED) under the grant number 09/2019/ TN.
Institutional Review Board Statement
All applicable international, national, and/or university guidelines were strictly followed for this study. This study does not involve human participant and was performed in accordance with the Guidelines for Animal Experiments of Kagoshima University. Individual ethical and animal experimental approval was not required from Kagoshima University. Our animal experimental facilities and programs have been fully accredited by AAALAC International since 2017.
Acknowledgments
We would like to thank Nobuyoshi Kawasaki, Toshinori Otani, Yoshikazu Yamada and Aiko Noguchi of Nihon Yuuki Co.,Ltd (Soo, Kagoshima, Japan) for supplying us with duck BAs, and Rieko Uchida of Kagoshima University for supporting our research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Arunlakshana, O.; Schild, H.O. Some quantitative uses of drug antagonists. Br. J. Pharmacol. Chemother. 1959, 14, 48–58. [CrossRef]
- Burggraaf, S.; Bingham, J.; Payne, J.; Kimpton, W.G.; Lowenthal, J.W.; Bean, A.G.D. Increased inducible nitric oxide synthase expression in organs is associated with a higher severity of H5N1 influenza virus infection. PLOS ONE 2011, 6, e14561. [CrossRef]
- Cirino, G.; Fiorucci S.; Sessa W. C. Endothelial nitric oxide synthase: the Cinderella of inflammation? Trends Pharmacol. Sci. 2003 24, 91–95. [CrossRef]
- Dauphin, F.; Ting, V.; Payette, P.; Dennis, M.; Hamel, E. Vasocontractile muscarinic M1 receptors in cat cerebral arteries: pharmacological identification and detection of mRNA. Eur. J. Pharmacol. Mol. Pharmacol. 1991, 207, 319–327. [CrossRef]
- Davis, R.L.; Choi, G.; Kuiken, T.; Quéré, P.; Trapp, S.; Short, K.R.; Richard, M. The culture of primary duck endothelial cells for the study of avian influenza. BMC Microbiol. 2018, 18, 138. [CrossRef]
- Friedrich, B.; Elisabeth K.; Jürgen S.; Walter R. K. Characterization of muscarinic receptors of bovine coronary artery by functional and radioligand binding studies. Eur. J. Pharmacol. 1991, 196, 247–255. [CrossRef]
- Guabiraba, R.; Garrido, D.; Bailleul, G.; Trotereau, A.; Pinaud, M.; Lalmanach, A.-C.; Chanteloup, N.K.; Schouler, C. Unveiling the participation of avian kinin ornithokinin and its receptors in the chicken inflammatory response. Veter- Immunol. Immunopathol. 2017, 188, 34–47. [CrossRef]
- Horinouchi, T.; Yamamoto, Y.; Koike, K. Effect of SR59230A on atypical β-adrenoceptor mediating relaxation in the guinea pig gastric fundus. Pharmacology 2001, 62, 98–102. [CrossRef]
- Islam, Z.; Kojima, S.; Sameshima, M.; Obi, T.; Yamazaki-Himeno, E.; Shiraishi, M.; Miyamoto, A. Vasomotor effects of noradrenaline, 5-hydroxytryptamine, angiotensin II, bradykinin, histamine, and acetylcholine on the bat (Rhinolophus ferrumequinum) basilar artery. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2021, 250, 109190. [CrossRef]
- Islam, Z.; Miyagi, K.; Matsumoto, T.; Nguyen, H.T.T.; Yamazaki-Himeno, E.; Shiraishi, M.; Miyamoto, A. Bradykinin induces NO and PGF2α production via B2 receptor activation from cultured porcine basilar arterial endothelial cells. Naunyn-Schmiedeberg's Arch. Pharmacol. 2014, 387, 697–702. [CrossRef]
- Islam, Z.; Sawatari, Y.; Kojima, S.; Kiyama, Y.; Nakamura, M.; Sasaki, K.; Otsuka, M.; Obi, T.; Shiraishi, M.; Miyamoto, A. Vasomotor effects of 5-hydroxytryptamine, histamine, angiotensin II, acetylcholine, noradrenaline, and bradykinin on the cerebral artery of bottlenose dolphin (Tursiops truncatus). J. Veter- Med Sci. 2020, 82, 1456–1463. [CrossRef]
- Islam, M. Z.; Watanabe Y.; Nguyen H. T. T.; Yamazaki-Himeno E.; Obi T.; Shiraishi M.; Miyamoto A. 2014a. Vasomotor effects of acetylcholine, bradykinin, noradrenaline, 5-hydroxytryptamine, histamine and angiotensin II on the mouse basilar artery. J. Vet. Med. Sci. 2014a, 76, 1339–1345.
- Ito, T.; Kobayashi, Y.; Morita, T.; Horimoto, T.; Kawaoka, Y. Virulent influenza A viruses induce apoptosis in chickens. Virus Res. 2002, 84, 27–35. [CrossRef]
- Jovanović, A.; Grbović, L.; Tulić, I. Endothelium-dependent relaxation in response to acetylcholine in the human uterine artery. Eur. J. Pharmacol. 1994, 256, 131–139. [CrossRef]
- Keawcharoen, J.; van Riel, D.; van Amerongen, G.; Bestebroer, T.; Beyer, W.E.; van Lavieren, R.; Osterhaus, A.D.; Fouchier, R.A.; Kuiken, T. Wild Ducks as Long-Distance Vectors of Highly Pathogenic Avian Influenza Virus (H5N1). Emerg. Infect. Dis. 2008, 14, 600–607. [CrossRef]
- Matsumoto, F.; Watanabe, Y.; Obi, T.; Islam, M.Z.; Yamazaki-Himeno, E.; Shiraishi, M.; Miyamoto, A. Characterization of 5-hydroxytryptamine-induced contraction and acetylcholine-induced relaxation in isolated chicken basilar artery. Poult. Sci. 2012, 91, 1158–1164. [CrossRef]
- Miyamoto, A.; Matsumoto, M.; Nishio, A. Endothelial Modulation of Vascular Tone in Isolated Porcine and Bovine Basilar Arteries.. J. Veter- Med Sci. 1994, 56, 947–950. [CrossRef]
- Miyamoto, A.; Hashiguchi, Y.; Obi, T.; Ishiguro, S.; Nishio, A. Ibuprofen or ozagrel increases NO release and l-nitro arginine induces TXA2 release from cultured porcine basilar arterial endothelial cells. Vasc. Pharmacol. 2007, 46, 85–90. [CrossRef]
- Miyamoto, A.; Ishiguro, S.; Nishio, A. Stimulation of bradykinin B2 -receptors on endothelial cells induces relaxation and contraction in porcine basilar artery in vitro. Br. J. Pharmacol. 1999, 128, 241–247. [CrossRef]
- Miyamoto, A.; Ito, K.; Nishio, A. Characterization of β-Adrenoceptors in Pig Basilar Artery from Functional and Radioligand Binding Studies. Jpn. J. Pharmacol. 1993, 61, 93–99. [CrossRef]
- Miyamoto, A.; Nakamoto, T.; Matsuoka, Y.; Ishiguro, S.; Nishio, A. The role of thromboxane A2 in regulating porcine basilar arterial tone. J. Veter- Pharmacol. Ther. 1998, 21, 223–227. [CrossRef]
- Miyamoto, A.; Nishio, A. Characterization of histamine receptors in isolated pig basilar artery by functional and radioligand binding studies. Life Sci. 1993, 53, 1259–1266. [CrossRef]
- Miyamoto, A.; Nishio A. Vasomotor effects of histamine on bovine and equine basilar arteries in vitro. Vet. Res. Commun. 1994, 18, 447–456. [CrossRef]
- Obara, K.; Suzuki, S.; Shibata, H.; Yoneyama, N.; Hamamatsu, S.; Yamaki, F.; Higai, K.; Tanaka, Y. Noradrenaline-induced relaxation of urinary bladder smooth muscle is primarily triggered through the β3-adrenoceptor in rats. Biol. Pharm. Bull. 2019, 42, 736–743. [CrossRef]
- Okuno, T.; Yabuki, A.; Shiraishi, M.; Obi, T.; Miyamoto, A. Histamine-induced modulation of vascular tone in the isolated chicken basilar artery: A possible involvement of endothelium. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2008, 147, 339–344. [CrossRef]
- Rahamt, S.; Gilland E. Comparative anatomy of the carotid-basilar arterial trunk and hindbrain penetrating arteries in vertebrates. Open Anat. J. 2014, 6, 1–26. [CrossRef]
- Ramirez, V.; Savoie, P.; Morais, R. Molecular characterization and evolution of a duck mitochondrial genome. J. Mol. Evol. 1993, 37, 296–310. [CrossRef]
- Redington, A.E. Modulation of nitric oxide pathways: Therapeutic potential in asthma and chronic obstructive pulmonary disease. Eur. J. Pharmacol. 2006, 533, 263–276. [CrossRef]
- Schmaier, A.H. The contact activation and kallikrein/kinin systems: pathophysiologic and physiologic activities. J. Thromb. Haemost. 2016, 14, 28–39. [CrossRef]
- Shirahase, H.; Usui, H.; Kurahashi, K.; Fujiwara, M.; Fukui, K. Possible Role of Endothelial Thromboxane A2 in the Resting Tone and Contractile Responses to Acetylcholine and Arachidonic Acid in Canine Cerebral Arteries. J. Cardiovasc. Pharmacol. 1987, 10, 517–522. [CrossRef]
- Short, K.R.; Veldhuis Kroeze, E.J.B.; Reperant, L.A.; Richard, M.; Kuiken, T. Influenza virus and endothelial cells: a species specific relationship. Front. Microbiol. 2014, 5, 653. [CrossRef]
- Skinner, B.M.; Robertson, L.B.; Tempest, H.G.; Langley, E.J.; Ioannou, D.; E Fowler, K.; Crooijmans, R.P.; Hall, A.D.; Griffin, D.K.; Völker, M. Comparative genomics in chicken and Pekin duck using FISH mapping and microarray analysis. BMC Genom. 2009, 10, 1–11. [CrossRef]
- Tong, Z.W.M.; Karawita, A.C.; Kern, C.; Zhou, H.; Sinclair, J.E.; Yan, L.; Chew, K.Y.; Lowther, S.; Trinidad, L.; Challagulla, A.; et al. Primary Chicken and Duck Endothelial Cells Display a Differential Response to Infection with Highly Pathogenic Avian Influenza Virus. Genes 2021, 12, 901. [CrossRef]
- Ueno, D.; Yabuki, A.; Obi, T.; Shiraishi, M.; Nishio, A.; Miyamoto, A. Characterization of bradykinin-induced endothelium-independent contraction in equine basilar artery. J. Veter- Pharmacol. Ther. 2009, 32, 264–270. [CrossRef]
- van Heuven-Nolsen D.; Tysse Klasen T. H.; Luo Q. F.; Saxena P. R. 5-HT1-like receptors mediate contractions of the rabbit saphenous vein. Eur. J. Pharmacol. 1990, 191, 375–382. [CrossRef]
- Vreman, S.; Bergervoet, S.A.; Zwart, R.; Stockhofe-Zurwieden, N.; Beerens, N. Tissue tropism and pathology of highly pathogenic avian influenza H5N6 virus in chickens and Pekin ducks. Res. Veter- Sci. 2022, 146, 1–4. [CrossRef]
- Wu, S.; Ootawa, T.; Sekio, R.; Smith, H.; Islam, Z.; Uno, Y.; Shiraishi, M.; Miyamoto, A. Involvement of beta3-adrenergic receptors in relaxation mediated by nitric oxide in chicken basilar artery. Poult. Sci. 2023, 102, 102633. [CrossRef]
Figure 1.
Typical contraction induced by Nω-nitro-L-arginine (L-NNA, 10-4 M) and relaxation induced by indomethacin (10-5 M) under precontracted condition induced by L-NNA. Relaxation induced by sodium nitroprusside was taken as 100%, in Figures 2, 5 and 8.
Figure 1.
Typical contraction induced by Nω-nitro-L-arginine (L-NNA, 10-4 M) and relaxation induced by indomethacin (10-5 M) under precontracted condition induced by L-NNA. Relaxation induced by sodium nitroprusside was taken as 100%, in Figures 2, 5 and 8.
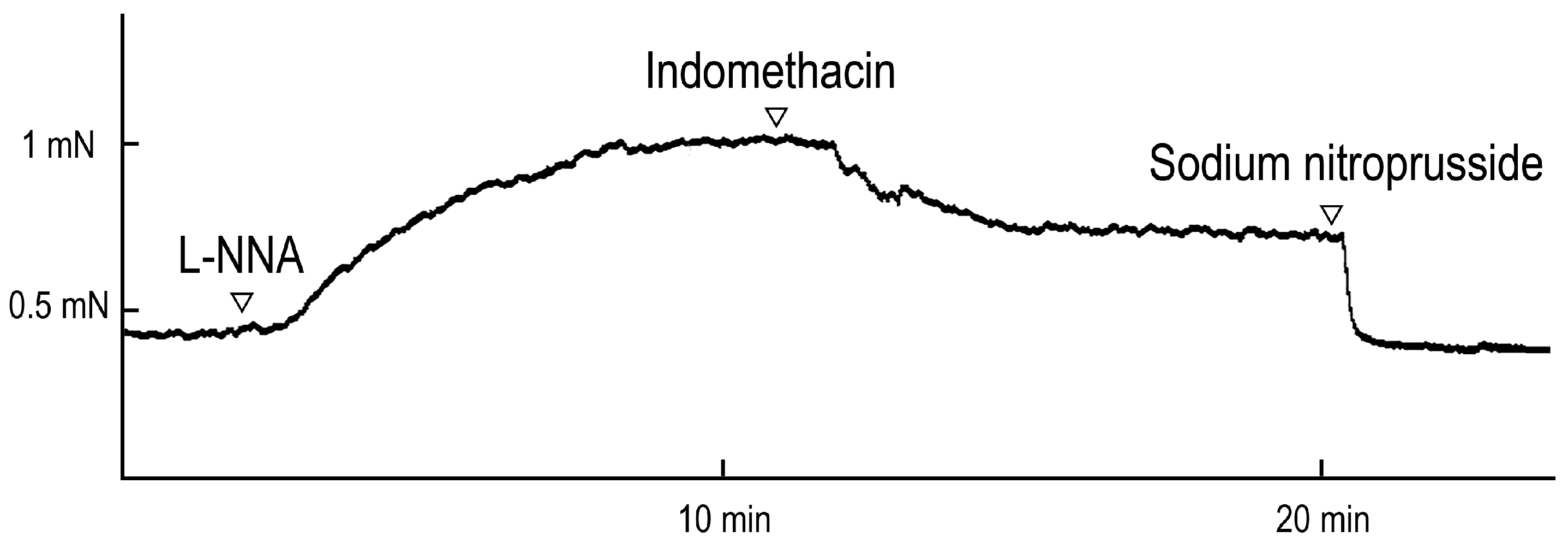
Figure 2.
[A] Responsiveness of duck basilar arteries to 5-hydroxytrypatamine (5-HT), histamine (His), noradrenaline (NA), NA in the presence of propranolol, and angiotensin (Ang) II under resting tension. [B]: Responsiveness of isolated duck basilar arteries to NA, NA in the presence of phentolamine (10-5 M) acetylcholine (ACh), and ornithokinin (OK) under precontracted condition induced with 5-HT. The contraction induced by 60 mM KCl [A] or the relaxation induced by 10-4 M sodium nitroprusside [B] were taken as 100% contraction and relaxation, respectively. Each point represents the mean ± SEM of 4–6 ducks.
Figure 2.
[A] Responsiveness of duck basilar arteries to 5-hydroxytrypatamine (5-HT), histamine (His), noradrenaline (NA), NA in the presence of propranolol, and angiotensin (Ang) II under resting tension. [B]: Responsiveness of isolated duck basilar arteries to NA, NA in the presence of phentolamine (10-5 M) acetylcholine (ACh), and ornithokinin (OK) under precontracted condition induced with 5-HT. The contraction induced by 60 mM KCl [A] or the relaxation induced by 10-4 M sodium nitroprusside [B] were taken as 100% contraction and relaxation, respectively. Each point represents the mean ± SEM of 4–6 ducks.
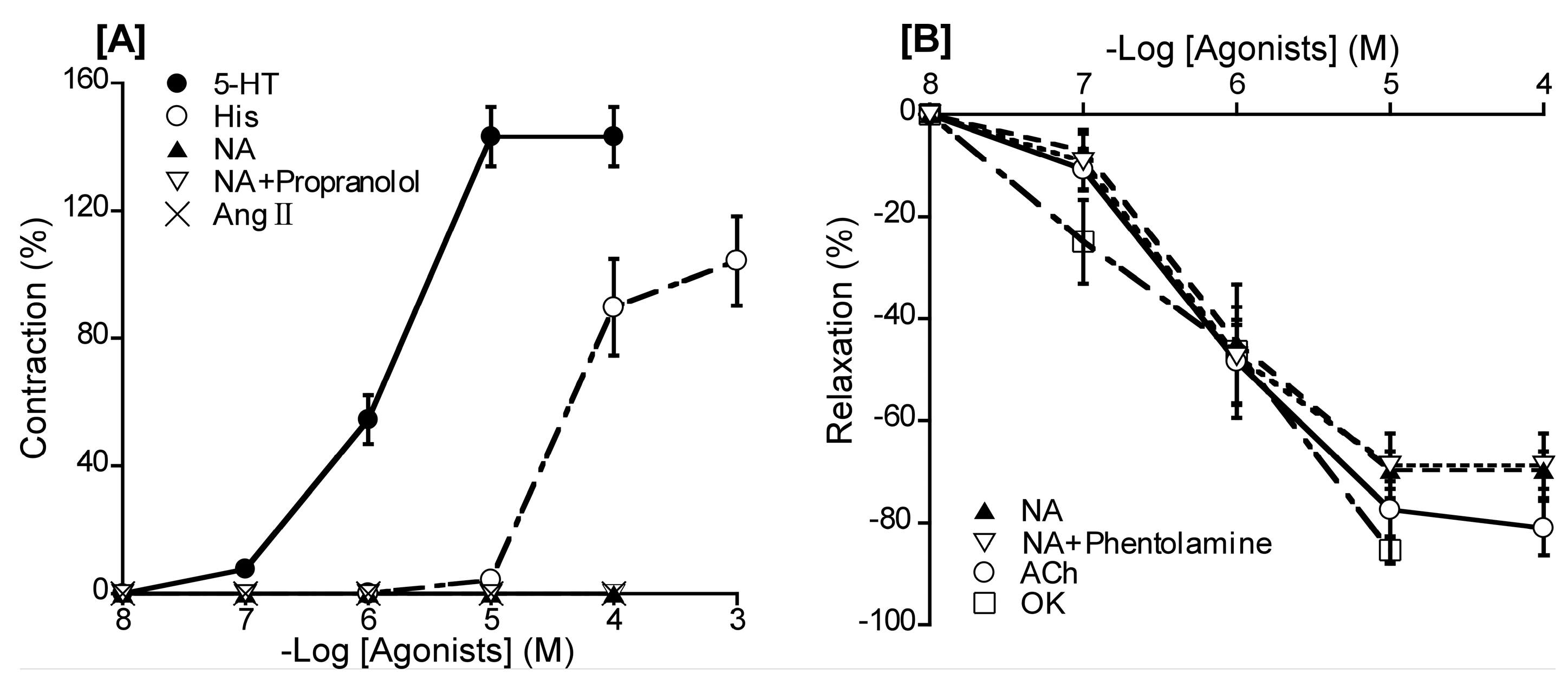
Figure 3.
Effects of methiothepin (Δ: 10-8 M, ▽: 10-7 M, ○: 10-6 M) [A], and ketanserin (○: 10-6 M) [B] on 5-hydroxytrypatamine (5-HT)-induced contraction (●) in duck basilar arteries, with Schild plot for methiothepin [C]. The maximal contraction induced by 5-HT was taken as 100%. Each point represents the mean ± SEM of 6 ducks.
Figure 3.
Effects of methiothepin (Δ: 10-8 M, ▽: 10-7 M, ○: 10-6 M) [A], and ketanserin (○: 10-6 M) [B] on 5-hydroxytrypatamine (5-HT)-induced contraction (●) in duck basilar arteries, with Schild plot for methiothepin [C]. The maximal contraction induced by 5-HT was taken as 100%. Each point represents the mean ± SEM of 6 ducks.
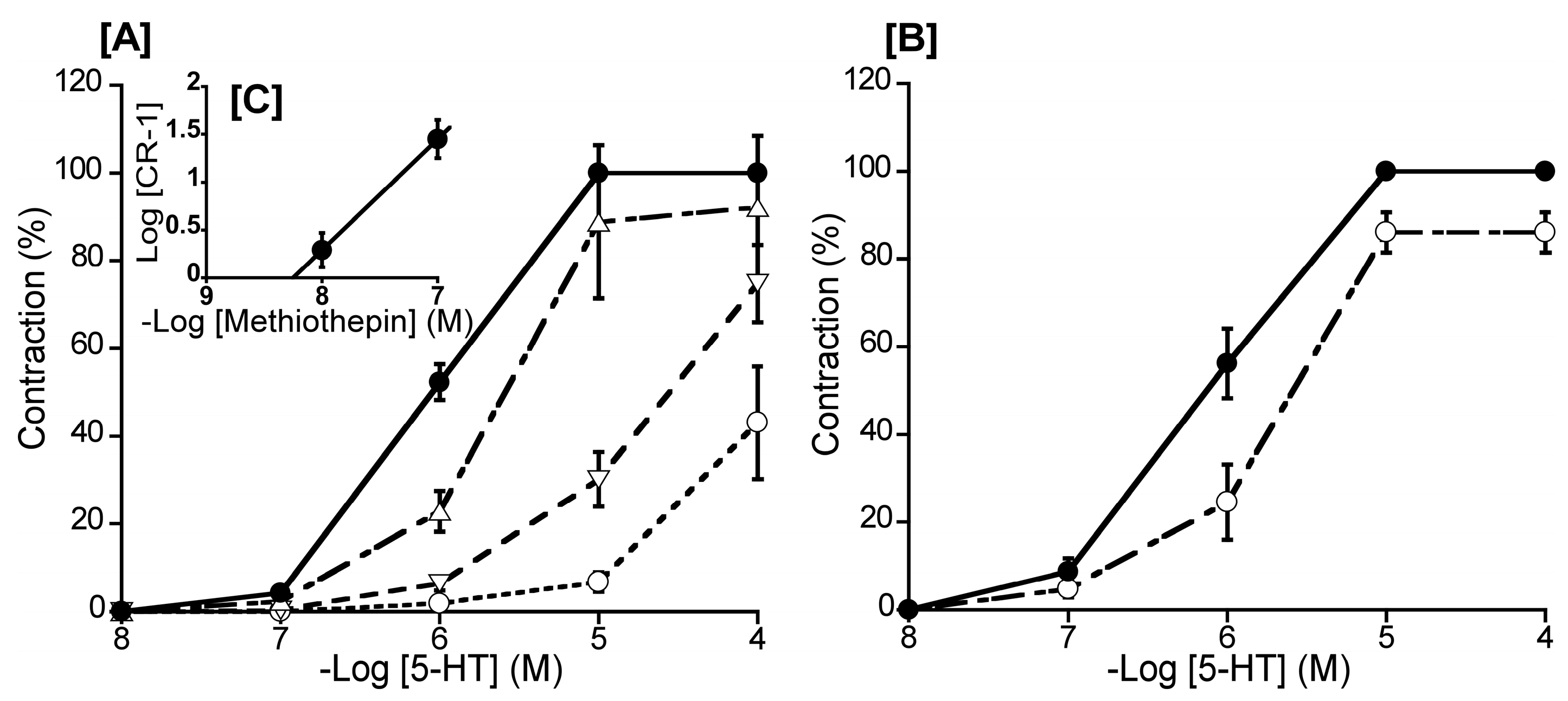
Figure 4.
Effects of diphenhydramine (Δ: 10-7 M, ▽: 10-6 M, ○: 10-5 M) and cimetidine (○: 10-5 M) [A] on histamine (His)-induced contraction (●) in duck basilar arteries, with Schild plot for diphenhydramine [B]. The maximal contraction induced by His was taken as 100%. Each point represents the mean ± SEM of 5 ducks.
Figure 4.
Effects of diphenhydramine (Δ: 10-7 M, ▽: 10-6 M, ○: 10-5 M) and cimetidine (○: 10-5 M) [A] on histamine (His)-induced contraction (●) in duck basilar arteries, with Schild plot for diphenhydramine [B]. The maximal contraction induced by His was taken as 100%. Each point represents the mean ± SEM of 5 ducks.
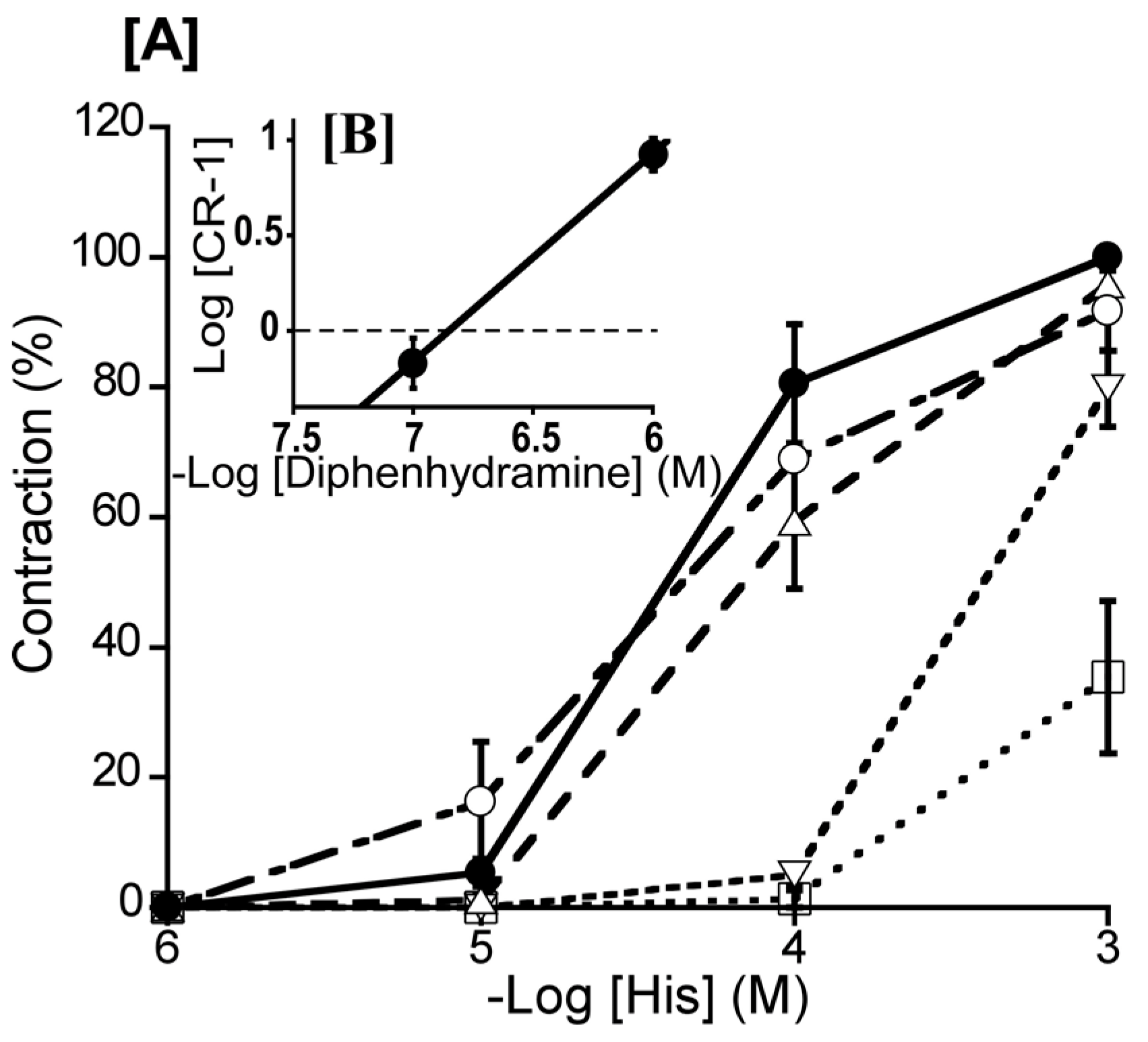
Figure 5.
Responsiveness of duck basilar arteries to isoproterenol (●), noradrenaline (▲), adrenaline (▼), and procaterol (×) under precontracted condition. The relaxation induced by 10-4 M sodium nitroprusside were taken as 100%. Each point represents the mean ± SEM of 4–6 ducks.
Figure 5.
Responsiveness of duck basilar arteries to isoproterenol (●), noradrenaline (▲), adrenaline (▼), and procaterol (×) under precontracted condition. The relaxation induced by 10-4 M sodium nitroprusside were taken as 100%. Each point represents the mean ± SEM of 4–6 ducks.
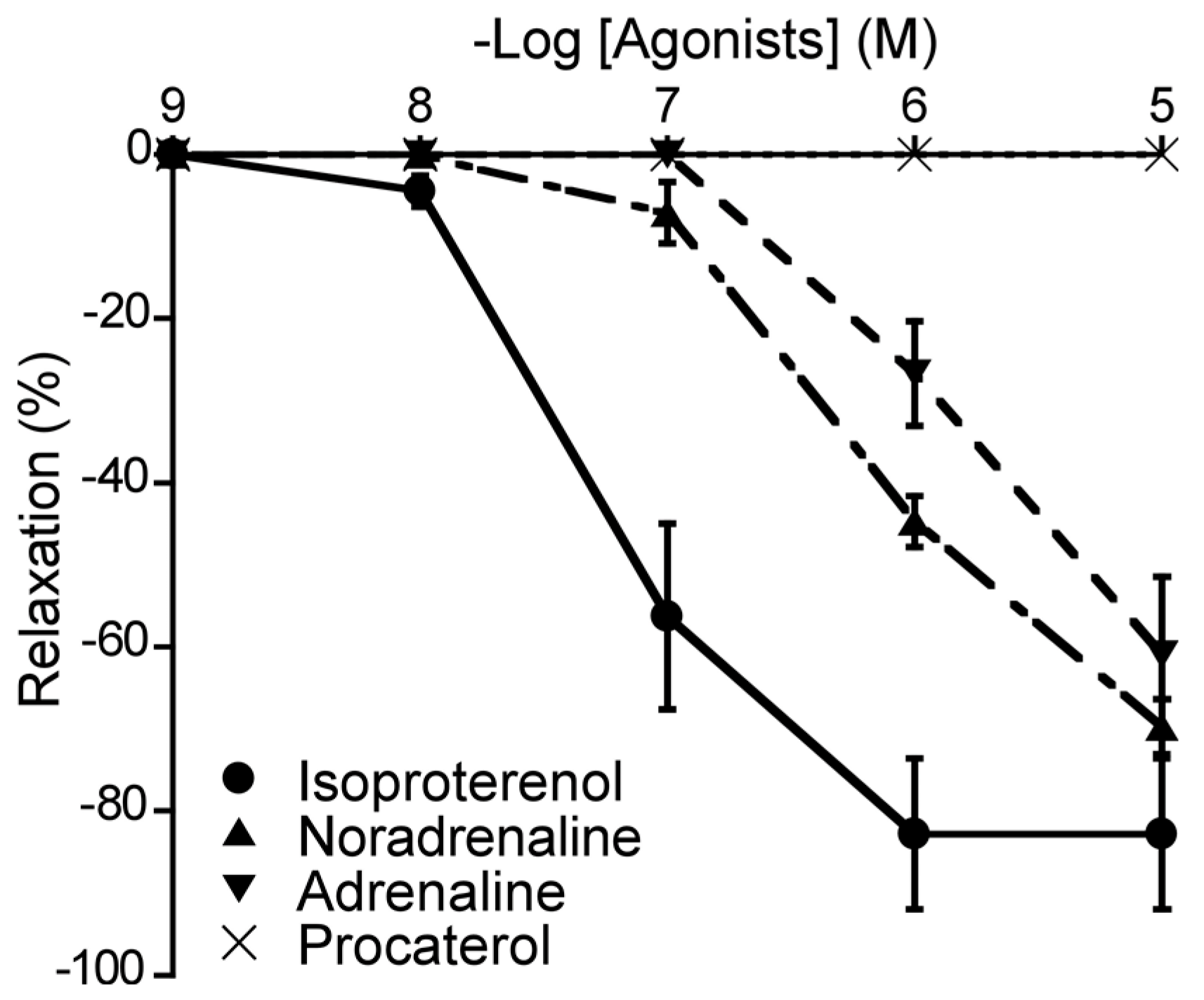
Figure 6.
Effects of atenolol (▲: 10-6 M) and butoxamine (▼: 10-6 M) [A], SR 59230A (○: 10-7 M, ▲: 10-6 M) [B] on isoproterenol-induced relaxation (●) in duck basilar arteries, with Schild plot for SR 59230A [C]. The maximal relaxation induced by isoproterenol was taken as 100%. Each point represents the mean ± SEM of 4 or 5 ducks.
Figure 6.
Effects of atenolol (▲: 10-6 M) and butoxamine (▼: 10-6 M) [A], SR 59230A (○: 10-7 M, ▲: 10-6 M) [B] on isoproterenol-induced relaxation (●) in duck basilar arteries, with Schild plot for SR 59230A [C]. The maximal relaxation induced by isoproterenol was taken as 100%. Each point represents the mean ± SEM of 4 or 5 ducks.
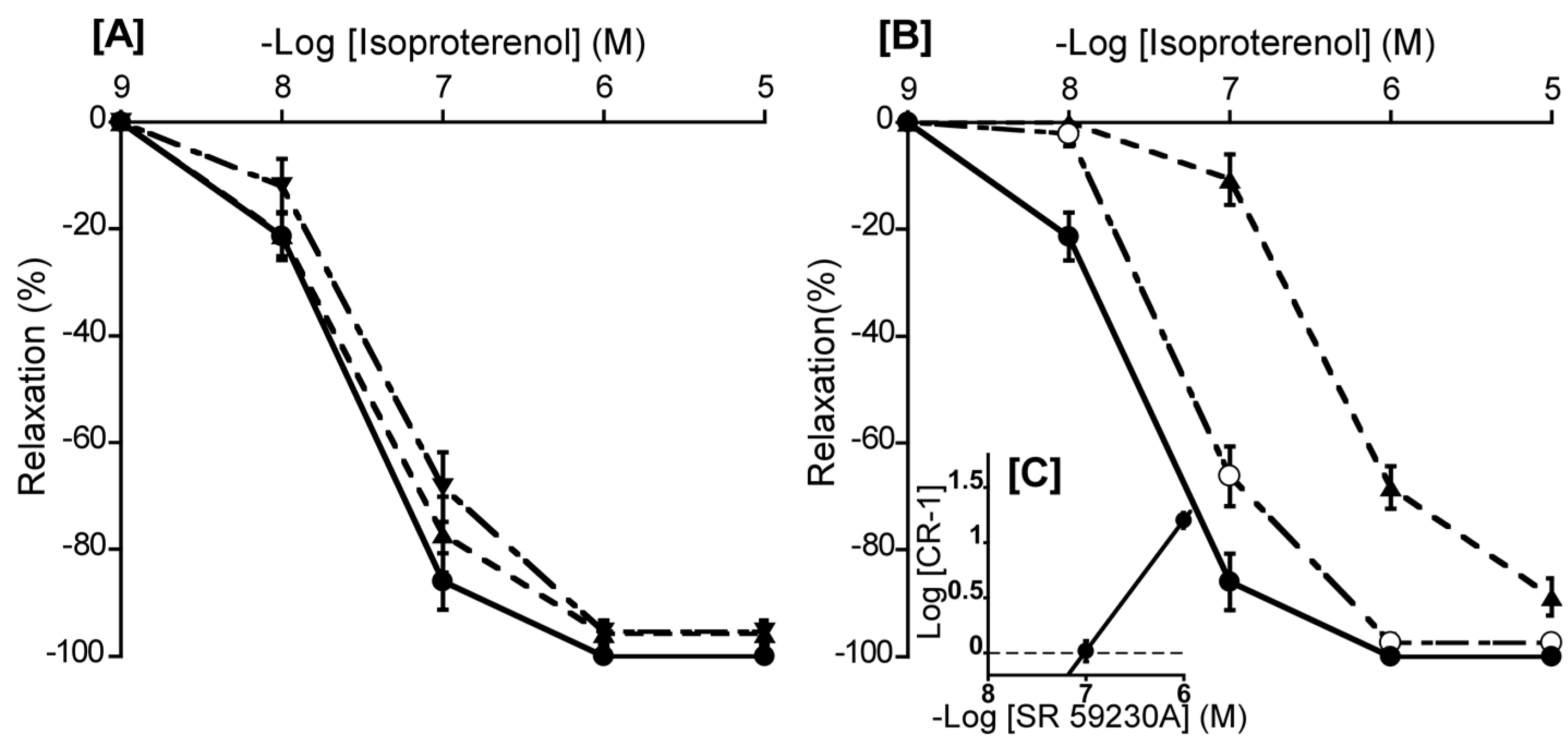
Figure 7.
Effects of atropine (○: 10-8 M, Δ: 10-7 M) [A], pirenzepine (■: 10-6.5 M, □: 10-6 M) [B], hexahydro-sila-difenidol hydrochloride, p-fluoroanalog (pFHHSiD; Δ: 10-7 M, □: 10-6 M) [C], and Nω-nitro-L-arginine (◇:10-4 M) [D] on acetylcholine (ACh)-induced relaxation (●) in duck basilar arteries, with Schild plots for pirenzepine [E], and pFHHSiD [F]. The maximal relaxation induced by ACh was taken as 100%. Each point represents the mean ± SEM of 5 ducks. (**p < 0.01 vs. control).
Figure 7.
Effects of atropine (○: 10-8 M, Δ: 10-7 M) [A], pirenzepine (■: 10-6.5 M, □: 10-6 M) [B], hexahydro-sila-difenidol hydrochloride, p-fluoroanalog (pFHHSiD; Δ: 10-7 M, □: 10-6 M) [C], and Nω-nitro-L-arginine (◇:10-4 M) [D] on acetylcholine (ACh)-induced relaxation (●) in duck basilar arteries, with Schild plots for pirenzepine [E], and pFHHSiD [F]. The maximal relaxation induced by ACh was taken as 100%. Each point represents the mean ± SEM of 5 ducks. (**p < 0.01 vs. control).
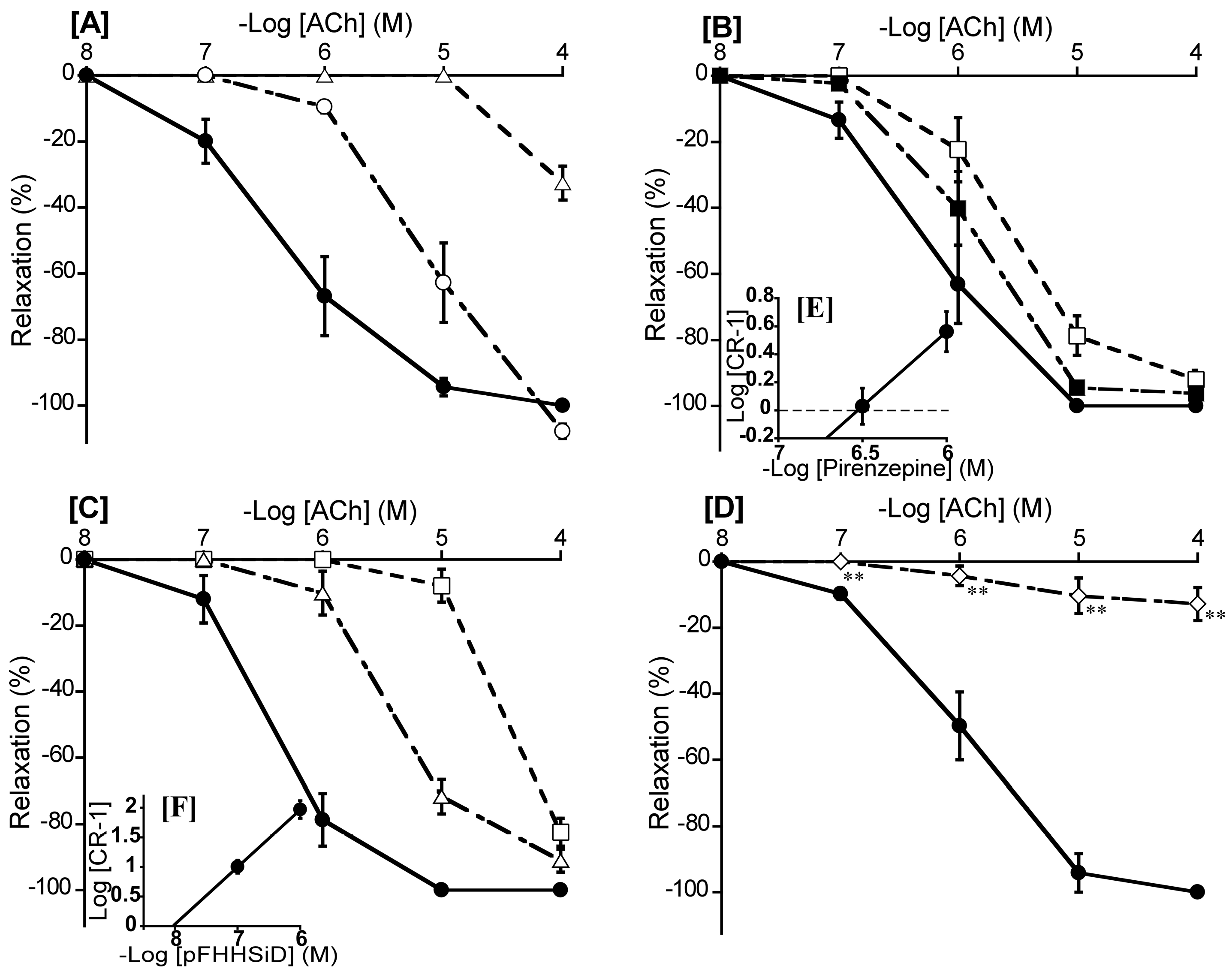
Figure 8.
Effects of Nω-nitro-L-arginine (○: 10-4 M, L-NNA) and L-NNA plus indomethacin (10-5 M) (▽) on ornithokinin (OK)-induced relaxation (●) in duck basilar arteries. The contraction induced by 60 mM KCl and the relaxation induced by 10-4 M sodium nitroprusside were taken as 100% contraction and relaxation, respectively. Each point represents the mean ± SEM of 4 ducks. (*p < 0.05, **p < 0.01 vs. control).
Figure 8.
Effects of Nω-nitro-L-arginine (○: 10-4 M, L-NNA) and L-NNA plus indomethacin (10-5 M) (▽) on ornithokinin (OK)-induced relaxation (●) in duck basilar arteries. The contraction induced by 60 mM KCl and the relaxation induced by 10-4 M sodium nitroprusside were taken as 100% contraction and relaxation, respectively. Each point represents the mean ± SEM of 4 ducks. (*p < 0.05, **p < 0.01 vs. control).
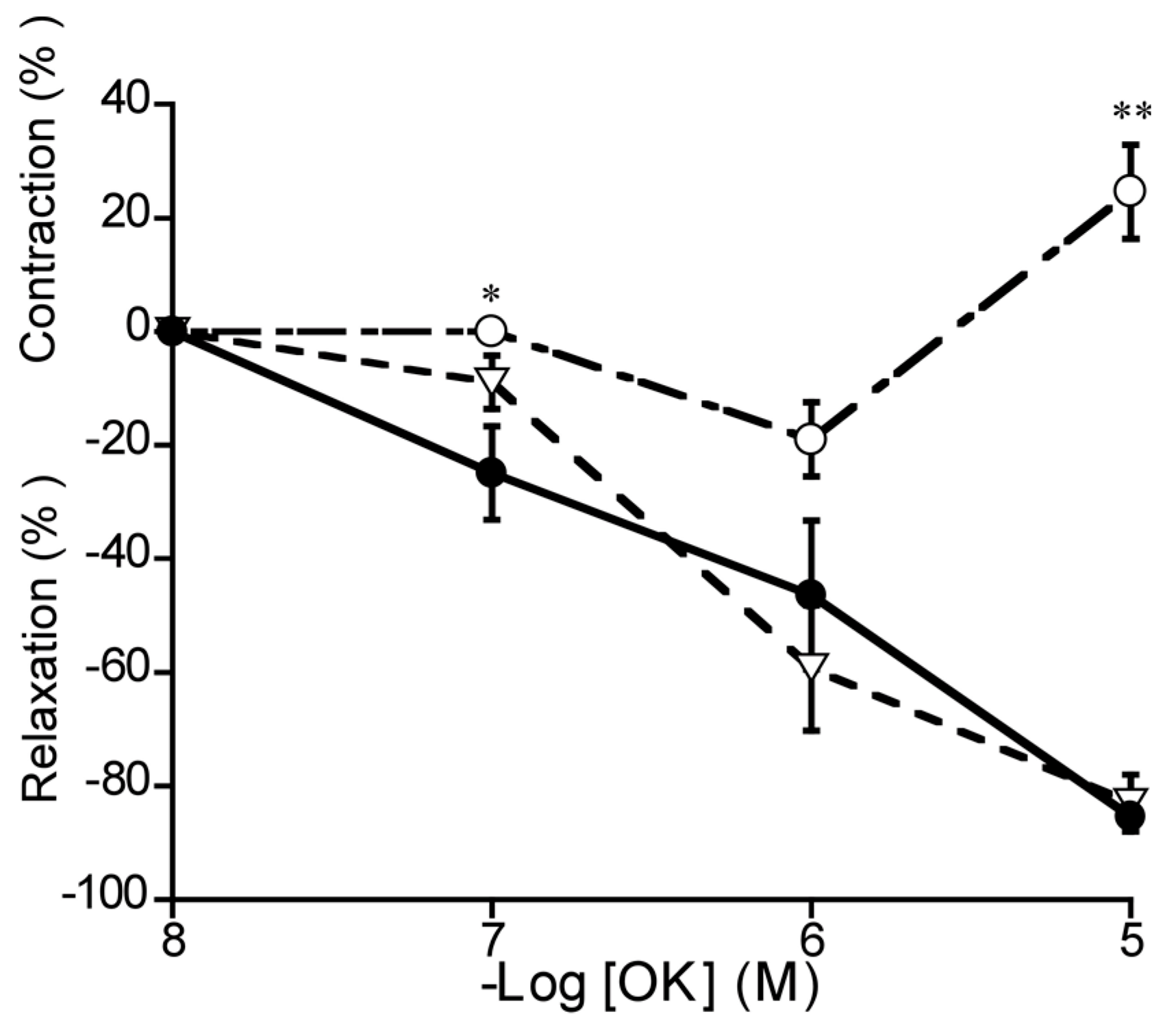

Table 1.
The pEC50 values and maximal response (Emax) for agonists.
| Agonists | pEC50 | Emax (%) |
|---|---|---|
| Resting tension | ||
| 5-Hydroxytryptamine | 5.84 ± 0.06 | 143.2 ± 9.3a |
| Histamine | 4.37 ± 0.09 | 104.3 ± 14.0a |
| Noradrenaline | - | No response |
| Noradrenaline + Propranolol | - | No response |
| Angiotesin II | - | No response |
| Precontracted condition | ||
| Acetylcholine | 6.18 ± 0.17 | -81.0 ± 5.4b |
| Noradrenaline | 6.28 ± 0.05 | -69.7 ± 3.7b |
| Noradrenaline + Phentolamine | 6.30 ± 0.11 | -68.8 ± 6.3b |
| Ornithokinin | 6.20 ± 0.25 | -85.4 ± 2.7b |
| Isoproterenol | 7.22 ± 0.11 | -82.8 ± 9.2b |
| Adrenaline | 5.89 ± 0.10 | -60.9 ± 9.5b |
aThe contraction induced by 60 mM KCl was taken as 100%; bThe relaxation induced by 10-4 M sodium nitroprusside was taken as 100%. Each value represents the mean ± SEM of 4–6 ducks. The concentration of propranolol and phentolamine is 10-5 M.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



