
Submitted:
03 August 2023
Posted:
04 August 2023
You are already at the latest version
Abstract
Phytoestrogens are plant-derived bioactive compounds with estrogen-like properties. Their potential health benefits, especially in cancer prevention and treatment, have been a subject of considerable research in the past decade. Phytoestrogens exert their effects, at least in part, through interactions with estrogen receptors (ERs), mimicking or inhibiting the actions of natural estrogens. Recently, there has been growing interest in exploring the impact of phytoestrogens on osteosarcoma (OS), a type of bone malignancy that primarily affects children and young adults, and currently presenting limited treatment options. Considering the critical role of estrogen/ERs axis in bone development and growth, the modulation of ERs has emerged as a highly promising approach in the treatment of OS. This review provides an extensive overview of current literature on the effects of phytoestrogens on human OS models. It delves into the multiple mechanisms through which these molecules regulate the cell cycle, apoptosis and key pathways implicated in the growth and progression of OS, including ERs signaling. Moreover, potential interactions between phytoestrogens and conventional chemotherapy agents commonly used in OS treatment will be examined. Understanding the impact of these compounds in OS holds great promise for developing novel therapeutic approaches that can augment current OS treatment modalities.
Keywords:
osteosarcoma
; phytoestrogens
; anticancer effects
; estrogen receptors
1. Introduction
Osteosarcoma (OS) is the most primary bone tumor, and a major cause of cancer death during the second and third decades of life with a worldwide incidence of 3.4 cases per million people per year, for all races and both genders [1,2]. Current disease management strategies include the surgical resection of all clinically visible tumors and systemic front-line chemotherapy which uses high doses of methotrexate (MTX), cisplatin (CDDP), and doxorubicin (DOX). This determines an overall survival level of 65–70% at 5 years [3,4]. However, today, several patients continue to develop metastases with an elective site in the lung, which causes a high mortality rate. This means that 20–30% of patients are refractory to these conventional treatments [5]. This discomforting scenario is frequently attributed to the ineffectiveness of chemotherapy, which can be influenced by chemo-resistance phenomena [6]. In addition, chemotherapeutic agents often produce various side effects, including cardiotoxicity, hepatotoxicity, and renal toxicity, which contribute to the increased likelihood of OS recurrence and progression [7].
While there are an increasing number of targeted therapies being developed and an improvement in the survival rate of other cancers, OS still stands where it was decades ago. These unsatisfactory results mean that complementary and alternative treatment options merit more attention. In this regard, dietary supplements and phytotherapeutic agents, with high anticancer efficacy and nominal toxicity to normal tissues, have emerged as a promising avenue that is worth investigating [8].
Over the past decade, significant attention has been devoted to studying phytoestrogens through mechanistic in vitro research which, together with epidemiological observations, have provided evidence that supports their chemopreventive and chemotherapeutic effects in different types of malignancies, including breast, prostate, and colon cancer [8]. Phytoestrogens are found in various plants, especially soy and soy-based foods, and herbal medicines [9]. They possess a structural similarity to the estrogen hormone, which enables them to interact with ERs in the body, exerting both estrogenic and anti-estrogenic effects [10,11,12]. This characteristic has sparked interest in gaining a better understanding of the potential influence of this class of plant substances on OS, considering that estrogen signaling can be implicated in the growth and progression of this malignancy [12,13,14]. In recent years, studies have shed light on the multifaceted effects of phytoestrogens on human OS cells. These compounds have been reported to exhibit both pro-apoptotic and anti-proliferative properties, suggesting their potential as therapeutic agents for OS [8,15]. Several underlying mechanisms have been proposed, including modulation of ERs, inhibition of angiogenesis, regulation of apoptosis-related proteins and interference with cell signaling pathways involved in OS development, including the phosphatidylinositol 3-kinase (PI3K)/Akt pathway, and the mitogen-activated protein kinase (MAPK) signaling and Wnt/β-catenin pathway [15]. Phytoestrogens also possess antioxidant and anti-inflammatory properties, which contribute to their anticancer effects by attenuating oxidative stress and inflammation-mediated processes [8,15].
In this work, we aim to provide a comprehensive synthesis of the existing body of literature surrounding the impact of phytoestrogens on human OS models, with particular attention being paid to the potential of these molecules as ER modulators. By examining preclinical studies and molecular investigations, we seek to gain insights into the mechanisms through which phytoestrogens exert their effects on OS cells and provide support for the future development of phytoestrogens as effective and safe agents for the therapy of OS.
2. Phytoestrogens: Chemical Classification and General Aspects
Phytoestrogens are produced by plants (more than 300 various species) as secondary metabolites which play crucial roles in various plant functions such as defense against pathogens, pigmentation and protection from UV radiation, photosynthetic stress and reactive oxygen species [16]. The quantity of phytoestrogens produced by a plant increases significantly during extreme growing conditions [17]. The human diet is rich in plants-containing phytoestrogens (i.e., vegetables, legumes, cereals, fruits, nuts etc.), and beverages such as wine, cider, beer, tea, and many more. Many edible plants contain multiple classes of phytoestrogens, adding to their diversity and potential health benefits [9]. Regarding their structural features, phytoestrogens represent a large and heterogenic class of non-steroidal substances characterized by a close structural similarity to the principal mammalian estrogen 17β-estradiol (E2) [11,12].
Shared structures include a phenolic ring and a pair of hydroxyl groups in opposite positions on the molecule (as in the case of E2 molecule), which are responsible for the interaction of phytoestrogens with the ligand binding domain of ERα and ERβ.
The exact position and number of these hydroxyl substituents is crucial in determining the binding affinity for the ERs and the activation of hormonal signaling [18,19]. The estrogenic or antiestrogenic properties of phytoestrogens in the target cells depend on their phenolic ring [20].
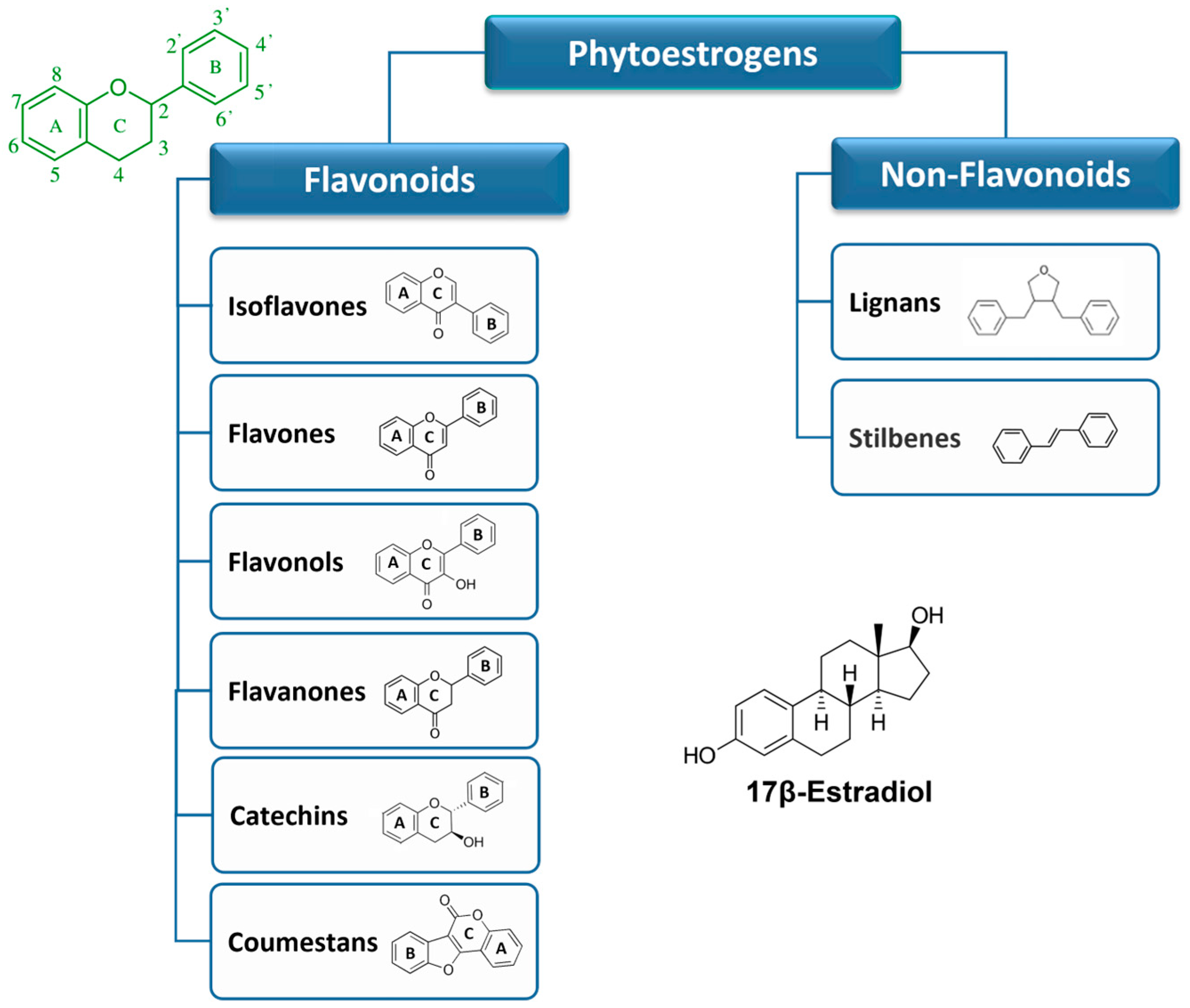
The phytoestrogens have been categorized into two main groups: flavonoids and non-flavonoids based on their chemical structure and properties. The classification and the basic structures of the most representative dietary phytoestrogens are illustrated in Figure 1. For further details see section 6 and 7. Isoflavones, flavones, flavonols, flavanones, catechins and coumestans belong to flavonoids. Non-flavonoids include, stilbenes, and lignans. The most commonly occurring phytoestrogens are the flavonoids (of which the coumestans and isoflavones have the greatest estrogenic effects) and the lignans [8,23].
Figure 1.
Basic chemical structures of the major classes of phytoestrogens. The different types of phytoestrogens share a structural similarity with 17β-Estradiol. In green, the flavonoid skeleton showing rings A, B and C and the numbering.
Figure 1.
Basic chemical structures of the major classes of phytoestrogens. The different types of phytoestrogens share a structural similarity with 17β-Estradiol. In green, the flavonoid skeleton showing rings A, B and C and the numbering.
2.1. Flavonoids
Flavonoids are a large group of substituted phenolic compounds [24]. They share a common structure consisting of a fifteen-carbon skeleton composed of two benzene rings (A and B) connected by a heterocyclic pyran structure (C), in a C6–C3–C6 arrangement, as illustrated in Figure 1 [25]. The basic flavonoid skeleton can have numerous substituents, including hydroxyl groups typically found at positions 4′, 5′, and 7. Flavonoids can be further classified into different subclasses. Specifically, isoflavones are flavonoids where the B ring is connected to the heterocyclic ring at the C3 position.
On the other hand, flavonoids in which the B ring is linked at position 2 are divided into several subgroups, namely flavones, flavonols, flavanones and catechins, depending on the degree of saturation and oxidation of the C ring (Figure 1). Coumestans are a distinct flavonoid which are characterized by a 1-benzoxolo (3,2-c)chromen-6-one structure formed by a benzoxole fused with a chromen-2-one [8]. The presence and position of hydroxyl groups and/or additional substituents contribute to the diversity of flavonoids and their biological activities [25,26,27].
Moreover, the addition of lipophilic prenyl side-chains can occur at different positions of the flavonoid skeleton, resulting in various prenylated derivatives with improved bioactivities and higher affinity to biological membranes. Prenylated flavonoids are much less common than flavonoids in nature [28].
Flavonoids are some of the most prevalent compounds found in fruits, vegetables, legumes, and tea and are generally concentrated in the fruit skin, bark, and flowers of plants [23]. Certain flavonoids, such as the flavonol quercetin are found in all plant products (i.e., fruit, vegetables, cereals, leguminous plants, tea, and wine), but others are specific components of particular foods (i.e., flavanones in citrus fruit and isoflavones in soya). In most cases, food contains complex mixtures of flavonoids; for many food products, the composition is less known (for review, see [29]). In plants, most flavonoids are conjugated with one or more sugar residues linked to hydroxyl groups or aromatic carbons, so they mainly exist as glycosides [30].
2.2. Non-flavonoids
Non-flavonoids encompass a broad range of plant compounds that do not possess the characteristic flavonoid structure. Their structure consists of phenolic acids in either C6–C1 (benzoic acid) or C6–C3 (cinnamic acid) conformations and are mainly represented by lignans and stilbenes (Figure 1). Non-flavonoids may occur in the form of aglycones and glycosides [30].
Lignans are dimers of phenylpropanoid units connected via two specific carbons (C-2–C-2’) and are typically found in plant cell walls [31]. They are widespread and their content is high in common foods, including grains, nuts, seeds, vegetables, and drinks such as tea, coffee or wine. Plant lignans are the principal source of dietary phytoestrogens of Western diet [32]. Compounds, such as pinoresinol, lariciresinol, sesamin, enterolactone, and enterodiol can be found in this group [31].
Stilbenes are among the most relevant non-flavonoid phytoestrogens which consist of a 1,2- diphenylethylene nucleus that generates two isomers (cis and trans), with trans-isomer being the most stable and biologically active [33,34]. More than 400 stilbene compounds have been identified in plants, with various structures ranging from monomers to octamers with different substituents, such as glycosyl, hydroxyl, methyl or isopropyl radicals [33]. Monomeric stilbenes have been studied the most. These include resveratrol and polydatin (section 7). They are naturally occurring in fruits, mostly in grapes, berries and peanuts [35]. In general, the occurrence of stilbenes in human diet is limited, but represents an important part of phytoestrogen intake by people who follow a Mediterranean diet or who regularly drink wines.
2.3. Metabolism of Dietary Phytoestrogens
Each class of dietary phytoestrogens has its own structural particularities, and studies regarding their bioavailability and metabolism are still far from being completed. There is no relation between the quantity of phytoestrogens in food and their bioavailability in the human body. Indeed, the rate and extent of absorption of dietary phytoestrogens in the intestine is determined primarily by their chemical structure and by factors such as molecular size and solubility, extent of glycosylation, hydroxylation, acylation and degree of polymerization [12,36]. Most ingested phytoestrogens (e.g., isoflavones, lignans and stilbenes), are predominantly present as estrogenically inactive glycosides in plant material [37]. After ingestion, phytoestrogens undergo extensive metabolization mediated both by tissue enzymes and gut microbiota, either prior to absorption or during enterohepatic circulation. The intestinal flora is capable of transforming aglycones into bioactive metabolites that are more similar to estrogens, being able to interfere with the endogenous estrogen signaling and associated cellular processes. In some cases, these metabolites have greater biological activities and different impacts on targeted tissues than their parent precursors [38,39]. For a detailed background on the absorption and metabolism of different phytoestrogens see ref. [40]. Thus, individual variability in gut microbiota can influence the metabolism of these estrogenic molecules, contributing to their intake and beneficial effects [41]. Consequently, the identification, quantification, and individual differences among phytoestrogens metabolites are important issues when researching the health effects of phytoestrogens in humans.
3. Phytoestrogen Mechanisms of Action - Anticancer Related Effects
In recent years, significant efforts have been made to elucidate the molecular mechanisms underlying the biological effects of phytoestrogens in both physiological and pathological conditions. The bioavailability and metabolism of phytoestrogens, as well as their effects on enzymes, nuclear receptors, and intracellular transduction mechanisms, play a crucial role in determining the overall impact of these compounds on cancer risk and progression [8]. However, the debate surrounding these effects persists, and further clarification is still needed. Phytoestrogens show a complex mode of action via interaction with the ER subtypes (i.e., ERα and ERβ), acting as either estrogen, triggering receptor pathways, or anti-estrogens, blocking normal estrogenic activity [10,21,42].
The dichotomy of ER modulating action induced by phytoestrogens led to the insertion of these compounds in the class of selective ER modulators (SERMs) [43,44,45,46] and probably provides an explanation regarding the conflicting evidence about the risks and benefits of these molecules on human health [47]. The activation of ER signaling pathways plays a vital role in the malignant progression of multiple cancers by comprehensively regulating downstream genes. The two ER subtypes have been described with different tissue distribution and ligand-binding affinities. ERα is mainly found in breast and uterine tissues and has been associated with pro-oncogenic responses while ERβ is the predominant isoform in the brain, bones, and blood vessels and is related to tumor-suppressive responses [19,48]. The alteration of the ERα/ERβ ratio in the affected tissues is one of the main reasons of the variability of estrogen-dependent cancer biology [49], and correlates with the response to the treatments and prognosis [50,51].
Phytoestrogens are known to bind ERs with much lower affinities than that of E2 (from 1/100 to 1/10,000), suggesting their weak estrogenic activities [19,48,52].
Moreover, unlike E2, which binds both ERα and ERβ with similar affinity, several phytoestrogens, including genistein, daidzein and naringenin display a substantially higher affinity for ERβ [19,53,54,55] (Table 1). Hence, considering the contrasting pro- and anti-cancerous effects exerted by ERα and ERβ respectively, along with the unique expression patterns of these receptors in various cell types and tissues, the specific impact of phytoestrogens on each ER subtype becomes crucial in shaping the effects of these compounds on cancer progression [56].

Table 1.
Relative binding affinities of various dietary phytoestrogens for ERα and ERβ, with values for E2 arbitrarily set as 100. The phytoestrogens reported are those discussed in the sections 6 and 7.
Table 1.
Relative binding affinities of various dietary phytoestrogens for ERα and ERβ, with values for E2 arbitrarily set as 100. The phytoestrogens reported are those discussed in the sections 6 and 7.
| Phytoestrogen | Relative binding affinity | References | |
|---|---|---|---|
| ERα | ERβ | ||
|
Isoflavones: Genistein Daidzein |
|||
| 4 | 87 | ||
| 0.1 | 0.5 | [19] | |
| Biochanin A | < 0.01 | < 0.01 | |
| Formononetin | < 0.01 | < 0.01 | |
| Flavonols: | |||
| Quercetin | 0.01 | 0.04 | [19] |
| Galangin | ND | ND | |
| Flavones: | |||
| Apigenin | 0.3 | 0.6 | [19] |
| Flavanones: | |||
| Naringenin | 0.01 | 0.11 | [19] |
| Stilbenes: | |||
| Resveratrol | 6.11–11.2a | 4.7–15.66a | [53] |
| Polydatin | ND | ND | |
a values reported by Bowers et al. [53], were obtained using a different methodology.
The actions of phytoestrogens via ERs can be mediated by genomic and/or non-genomic mechanisms, in a dose and tissue-specific manner [53,57]. The ER-mediated genomic effects of phytoestrogens result in the regulation of target genes, which include anti-inflammatory, anti-apoptotic, metabolic and mytocondrial genes, as well as an improvement in mitochondrial biogenesis and function, which leads to increased resistance to stress [58,59] (Figure 2).
Figure 2.
Intracellular mediators of the effects of phytoestrogens on cancer cells. Phytoestrogen can interact with the two types of ER, the intracellular ERα and ERβ and membrane-associated mERs and GPER, activating downstream genomic and non-genomic effects which ultimately affect cell cancer phenotypes. The genomic pathway can involve ER interactions with other transcription factors (TFs) such as, CREB, AP-1, Sp1 and NF-κB. Phytoestrogens can also act through ER-independent mechanisms which induce oxidative stress-mediated signaling by generating ROS, as well as interacting with other nuclear receptors, such as PPARs and ERRα/γ.
Figure 2.
Intracellular mediators of the effects of phytoestrogens on cancer cells. Phytoestrogen can interact with the two types of ER, the intracellular ERα and ERβ and membrane-associated mERs and GPER, activating downstream genomic and non-genomic effects which ultimately affect cell cancer phenotypes. The genomic pathway can involve ER interactions with other transcription factors (TFs) such as, CREB, AP-1, Sp1 and NF-κB. Phytoestrogens can also act through ER-independent mechanisms which induce oxidative stress-mediated signaling by generating ROS, as well as interacting with other nuclear receptors, such as PPARs and ERRα/γ.
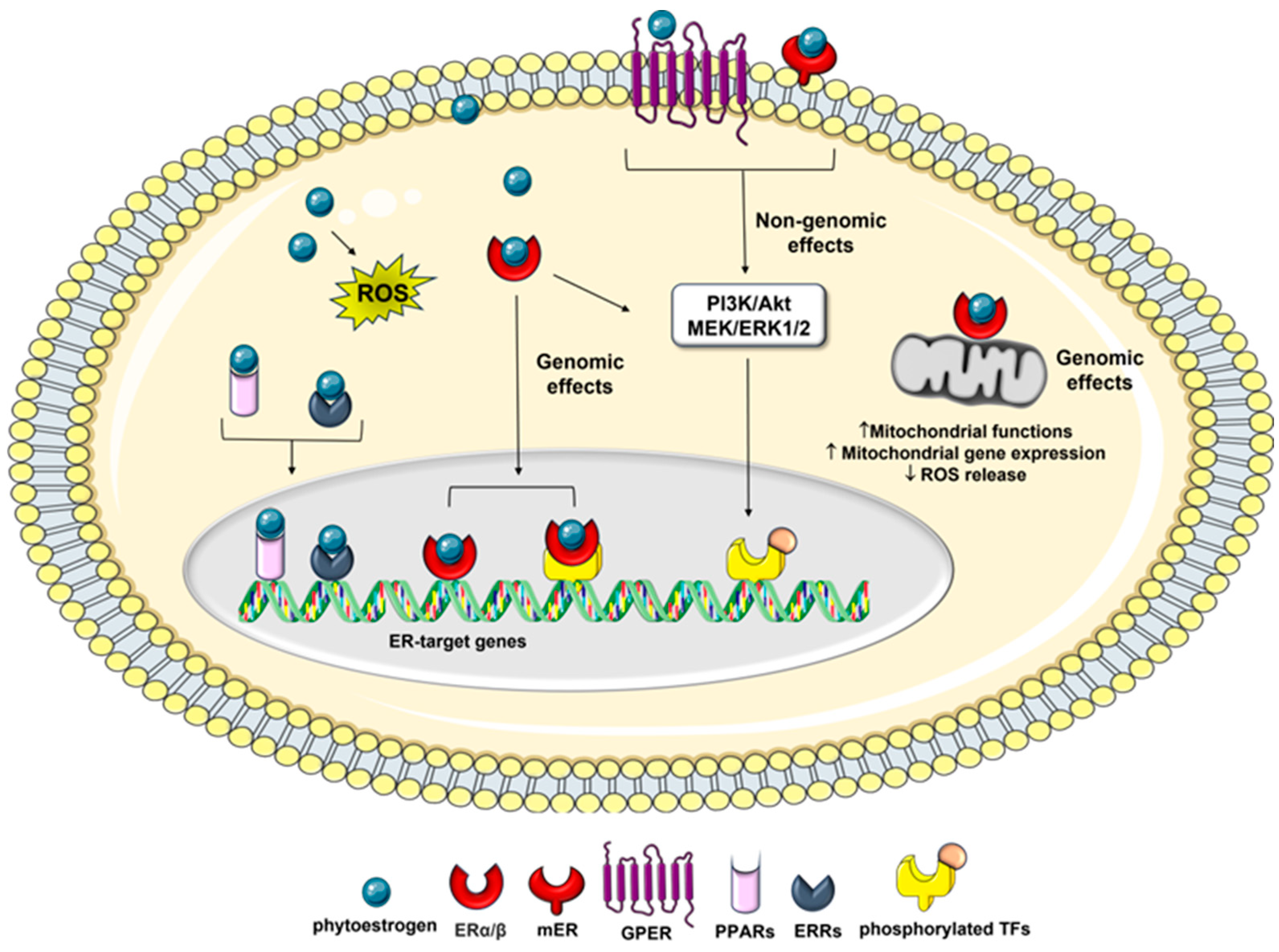
Phytoestrogens also modulate several therapeutically important oncogenic signaling pathways including epithelial-mesenchymal transition (EMT), and MAPK associated pathways [47,60], and recruit transcription factors such as, response element binding protein (CREB), the activator protein 1 (AP-1), the stimulating protein 1 (Sp1), and the nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB), which are correlated with cell cycle regulation, angiogenesis, metastasis, and apoptosis [61].
Besides nuclear events mediated by intracellular ER binding, many of these compounds also exert non-genomic effects through the activation of the membrane-associated ERs (mERs) and/or G-protein-coupled estrogen receptor 1 (GPER1/GPR30), which are involved in a diverse array of disorders including cancer [62,63,64]. Among the membrane-initiated non-genomic effects is the activation of signaling cascades, such as the mitogen-activated protein extracellular kinase/extracellular signal-regulated kinase (MEK/ERK) and PI3K pathways, which affect cancer cell apoptosis and proliferation [10,65].
In addition, they can exert estrogenic activity by cross-talk with many other pathways, including those related to membrane-associated growth factor receptors, such as human epidermal growth factor receptor (EGFR/HER) and insulin-like growth factor 1-receptor (IGF1R) [66], as well as nuclear receptors, including [67], peroxisome proliferator-activated receptors (PPARs) [68], and estrogen-related receptor alpha/gamma (ERRα/γ) (Figure 2) [69]. Moreover, phytoestrogens can promote apoptosis and prevent the reproduction of malignant cells by blocking neo-angiogenesis, tyrosine-kinase, and topoisomerase proteins [70]. Several studies have also reported that in addition to the classical estrogen receptor signaling, and the genomic and non-genomic effects mentioned above, some phytoestrogens, including genistein and resveratrol, exert their anticancer effects by epigenetic mechanism, such as the modulation of chromatin structure [43,71], and the regulation of different cancer associated miRNAs [72,73], suggesting new therapeutic strategies for cancer.
Phytoestrogens are mostly known for their potent antioxidant activity, i.e., another biological activity that indirectly reduces the risk of various degenerative diseases linked to oxidative stress, including cancer [74]. The chemical structures of these compounds consist in a long-conjugated system that includes phenolic groups. This structural arrangement confers them significant antioxidant properties [8], which have been linked to their chemoprevention potential, particularly in Asian populations. It is worth noting that in these populations, there is a correlation between soy consumption and a reduced occurrence of estrogen-related cancers [8,26].
On the other hand, at high concentrations, phytoestrogens may have pro-oxidant effects and induce cell death. Flavonoids autoxidize in aqueous medium and may form highly reactive radicals in the presence of transition metals. This effect has been described for several compounds, including genistein [75,76] and resveratrol [77], suggesting that a combination of phytoestrogens with anti-cancer treatments may render cancer cells more sensitive to treatment, in part by increasing reactive oxygen species (ROS) production. However, given the high concentration of these compounds required for these activities, their impact on cancer onset and progression appears to be related to other cellular effects besides the modulation of oxidative stress [78,79].
Some phytoestrogens have also been shown to possess anti-inflammatory properties and modulate immune responses. They can inhibit the production of inflammatory mediators and reduce the expression of pro-inflammatory genes, contributing to their potential as anticancer agents [8,15].
As a whole, phytoestrogens exert a plethora of effects through multiple synergistic signaling pathways, which contribute to the outcome of phytoestrogen exposure on health and/or cancer cells. The specific effects of phytoestrogen exposure on the cancer initiation, progression, and development may differ depending on the ERα/ERβ ratio in the affected tissue and the different selectivity and concentration of phytoestrogens [49]. In this regard, the majority of reported findings indicate distinct effects at low and high concentrations of phytoestrogens, which may be attributed to the capacity of these molecules to interact with and modulate ERs, thereby influencing endocrine functions [80]. Indeed, some studies have raised concerns about the potential adverse effects of soy products, particularly in high doses or when consumed by individuals with hormone-sensitive cancers [10,43]. Hence, it is crucial to gain a comprehensive understanding of how phytoestrogens interact with the ERs to fully evaluate their toxicologic and pharmacologic properties.
4. Molecular Basis of Osteosarcoma Pathogeneses
The difficulty in establishing efficacious OS therapy is linked to the unclear specific markers for diagnosis and treatment. It is also due to the complexity of the OS genome, low incidence of this tumor, and significant biologic differences between OS subtypes. Nevertheless, the heterogeneity in the genotype of OS has translated into several expression profiles of macromolecular biomarkers, which are helpful in the clinic [2,81,82,83]. There are many genetic mutations observed in OS patients. The p53 and retinoblastoma (Rb) genes are well-known tumour-suppressor genes. Both germline and somatic mutations of p53 and Rb gene have been proven to be involved in OS pathogenesis [83,84,85]. Inherited cancer predisposition syndromes, such as Li–Fraumeni, hereditary retinoblastoma, Rothmund–Thomson, Bloom or Werner syndrome, may also influence the high appearance of this kind of tumor in young patients [84,86,87,88,89]. Among other genes mutated in more than 10% of OS cases, c-Myc plays a role in OS development and promotes cell invasion by activating MEK–ERK pathways. High expression of c-Myc in OS tumors correlates with the formation of metastasis and poor prognosis [90].
Several studies have consistently demonstrated that OS cells have the capacity to develop and secrete a range of growth factors that exert autocrine and paracrine effects. Vascular endothelial growth factor (VEGF), transforming growth factor (TGF), IGF-I and IGF-II, and connective tissue growth factor (CTGF) are deregulated in OS, which leads to tumor progression and growth in target cells [83,91,92,93]. Parathyroid hormone-related peptide (PTHrP) and its receptor have also been implicated in OS progression and metastasis development, with PTHrP conferring OS chemoresistance by blocking signaling via p53 [94]. Epigenetic events have emerged as significant risk factors for OS, since the DNA methylation pattern of specific genes or gene regions and histone modifications may be involved in tumor development [95]. In addition, a variety of studies have found abnormally-expressed levels of micro-RNAs (miRs), which have the potential to become prognostic biomarkers of OS. Overexpression of this molecule results in proliferation, migration and invasion of tumor cells [127,128]. Among the miRNAs deregulated in osteosarcoma are miR-421, miR-16, miR-200b and miR-101 [82,98,99].
OS is a highly metastatic tumour, and pulmonary metatases are the most common cause of death [83,100]. The ability of OS cells to metastasise has been found to be correlated with multiple processes and various cytophysiological changes, including changing the adhesion capabilities between cells and the extracellular matrix (ECM) and disrupting intercellular interactions [101,102]. Degradation of the ECM and components of the basement membrane caused by the concerted action of proteinases, such as matrix metalloproteinases (MMPs), cathepsins, and the plasminogen activator (PA), can play a critical role in OS invasion and metastasis [102]. Moreover, in metastatic forms of OS, some specific genetic changes have been observed, which include upregulation of the Wnt/β-catenin and Src pathway, neurogenic locus notch homolog protein 1 and 2 (Notch1/Notch2) receptors [103,104] together with the downregulation of the Fas/Fas ligand pathway (a cell death pathway), which increases the metastatic potential of human OS [105,106].
In both primary bone cancer and bone metastases, the bone remodeling process creates a favorable environment for tumor establishment and progression. Osteoblasts and osteoclasts are the primary regulators of bone metabolism [107]. Specifically, osteoblasts secrete multiple components of ECM and MMPs in OS niche, which are rich promoters of OS development. Moreover, osteoclasts play a pivotal role as bone-resorbing cells, and significant osteolysis exhibited in some OS cases can be directly attributed to the heightened activity of osteoclasts [102].
It has been demonstrated that OS is a condition characterized by deregulation in the signaling triad, receptor activator of nuclear factor kB Ligand (RANKL), its receptors RANK and osteoprotegerin (OPG) [108,109]. In its canonical function, RANKL, which is secreted by osteoblasts, induces bone destruction by mature osteoclasts. In response, osteoblasts secrete OPG—RANKL decoy receptor—and in this way inhibit osteoclast differentiation and resultant bone resorption [108,110]. The RANKL/OPG ratio in the blood is increased in high-grade OS, leading to the establishment of a vicious cycle between pathological bone remodeling and OS growth [110]. RANKL/RANK-signaling regulates OS cell migration and tissue-specific metastatic behavior in the lungs, but has no direct impact on OS-associated bone destruction and does not impact OS cell proliferation [108,111]. Thus, osteoclast pathways of differentiation, maturation, and activation constitute another compelling therapeutic target since the inhibition of bone resorption at the tumor-bone interface may lead to reduced local OS invasion [108].
Among the possible mechanisms that contribute to OS development in bone microenvironment are alterations in the osteogenic pathway, which lead to the differentiation of mesenchymal stem cells (MSCs) into mature osteoblasts [82,112]. Defects in osteogenic differentiation or exposure to new non-native stimuli such as pro-inflammatory cytokines and pro-tumor agents, may cause an imbalance between cell differentiation and proliferation, so contributing to a malignant phenotype. OS cells share more characteristics with undifferentiated osteo-progenitors than with differentiated osteoblasts, including a high proliferative capacity and resistance to apoptosis. Indeed, osteogenic regulators associated with mature osteoblast phenotypes, such as CTGF, RUNX2, alkaline phosphatase (ALP), osteopontin (OPN), osteocalcin (OCN), are very lowly expressed in both primary OS tumors and OS cell lines [113].
Although not well understood, some of the potential defects in the MSC differentiation cascade may include genetic and/or epigenetic changes in Wnt signaling, Rb and p53. These alterations may lead to uncontrolled cell proliferation and disrupted differentiation, so producing a tumorigenic phenotype [82,112]. Interestingly, treatments of human OS cells with therapeutic agents such as peroxisome proliferator-activated receptor (PPAR) agonists [113], growth factors (e.g., PTHrP) [114], and SERMs [115] enable terminal differentiation and subsequent tumor inhibition. Hence, a better understanding of the relationship between defects in osteogenic differentiation and tumor development is of fundamental importance for the treatment of OS and promoting differentiation offers a potential for disease control.
5. Estrogen Receptors as a Potential Target for the Treatment of Osteosarcoma
A number of studies have established a correlation between the rapid bone growth experienced during puberty, when endogenous sex hormones, growth hormones, and IGF1 levels are at their highest, and OS development [116]. During the growth phase, there is a greater bone turnover, and so the possibility for defects to occur in the differentiation process and in the signaling pathways is amplified [1].
The estrogen/ERs axis is widely recognized for its essential role in bone development and growth from puberty through adulthood, acting via a variety of mechanisms and cell types [14,117,118]. Both ER isoforms, α and β, have been identified in bone, although their levels are lower than in reproductive tissues, and can be affected by many parameters including the cell differentiation state [118,119,120]. ERα and ERβ have been observed to antagonize each other (activation vs. repression of transcription) in bone cells [118]. Therefore, ER isoform-specific activation could potentially lead to large differences in the patterns of gene expression induced by the ER-isoform homodimers [121,122]. In this regard, further work still needs to be done to identify different genes that are regulated by estrogens and SERMs, including phytoestrogens, in each bone cell type during different stages of differentiation.
Several lines of evidence have hinted that the estrogens, which play prominent and well-described roles in osteoclast biology [14], might have a therapeutic differentiating effect on OS cells [123]. Accordingly, the loss of estrogens or the impaired functionality of the ERs appears to be involved in bone-forming cancers such as OS [14,124,125].
The expression of ERs is regulated by the methylation state in the promoter region. Their lack of expression, caused by epigenetic silencing (methylation) in the promoter region, is associated with the development of bone cancer and the metastasis of tumors in bone [126,127]. In line with this evidence, ERs have attracted widespread attention as promising targets for treating OS.
ERα is essential in osteogenesis and regulates cell growth in various tumors, including OS [128,129,130]. The activation of the ERs, especially ERα, triggers the downstream Wnt/beta-catenin signaling cascade, which promotes osteogenesis [128]. Moreover, targeting ERα-sensitive OS cells treated with methotrexate enhances the cytotoxic effects on OS when combined with DOX treatment [131].
OS typically originates from osteoblasts and/or MSCs, both of which normally express ERα [118,132]. However, the majority of OS tumors do not exhibit ERα expression, primarily due to the hypermethylation occurring in the ERα promoter region [124,126,130]. Notably, 143B cells demonstrate complete methylation of the ERα promoter, while U2OS and MG63 cells display partial methylation [124]. Interestingly, the use of demethylating agents to reverse the epigenetic silencing had a noteworthy effect on OS cell lines in vitro. It promoted differentiation and suppressed proliferation, and these effects were partially replicated in vivo, demonstrating potential disease control [124]. This suggests that the presence of ERα could serve as a therapeutic target and a prognostic factor for predicting the response to chemotherapy [124,130].
Regarding ERβ, while its specific functions have not been as extensively studied as those of ERα, evidence has emphasized its role as a tumor-inhibiting factor in estrogen-sensitive malignant tumors [133]. The anti-tumor effects of ERβ have also been reported in OS [13,115,134,135]. Specifically, it was found that ERβ mediates the proliferation, migration and invasion of U2OS cells by regulating the integrin, IAP, NF-kB/Bcl-2, and PI3K/Akt signaling pathways. Moreover, silencing of Erβ promoted the metastatic phenotype in U2OS cells and OS tumors of mice, through activating the Wnt signaling pathway [136]. ERβ also plays an important role in inducing autophagy in U2OS cells by downregulating of the expression of P62 and phosphorylated mammalian target of rapamycin (p-mTOR) [13,137]. These findings suggest a role of ERβ agonists as potential candidates for therapy in OS. This is a noteworthy aspect considering that many phytoestrogens exhibit a higher binding affinity for ERβ compared to Erα [19].
The challenge in OS therapy lies in the lack of clearly defined markers for diagnosis and treatment. Because of the critical role of ERs in bone formation, whether or not the control of ERs by SERMs can modulate new bone formation and affect the prognosis or chemosensitivity of bone tumors is an interesting subject of study.
6. Anti-Osteosarcoma Effects of Flavonoids
6.1. Genistein and Related Isoflavones
Genistein is a 15-carbon skeleton compound, chemically known as 5,7-dihydroxy-3-(4-hydroxyphenyl)chromen-4-one, which was first isolated from Genista tinctoria. It is the most prominent isoflavone from soy, and soy-based food products [138,139].
In addition to genistein, soy foods are rich in daidzein, which differs from genistein due to the lack of the hydroxyl group at position 5 [140,141]. Genistein and daidzein derive from the O-methylated precursors biochanin A and formononetin, respectively [141,142] (Figure 3), which are generally less prevalent in soy and are found mostly in clover and alfalfa sprouts [141,143]. They are present as either aglycons or as glycosides [143,144]. The presence of hydroxyl groups and sugars increases their solubility in water, whilst methyl groups confer them lipophilicity [23].
Figure 3.
Chemical structure of the most common isoflavones and some of their possible targets in OS cells. Isoflavones, especially genistein, bind ERβ with a significant higher affinity than ERα. Downward arrows represent downregulation or reduction. Upward arrows represent upregulation or increase.
Figure 3.
Chemical structure of the most common isoflavones and some of their possible targets in OS cells. Isoflavones, especially genistein, bind ERβ with a significant higher affinity than ERα. Downward arrows represent downregulation or reduction. Upward arrows represent upregulation or increase.
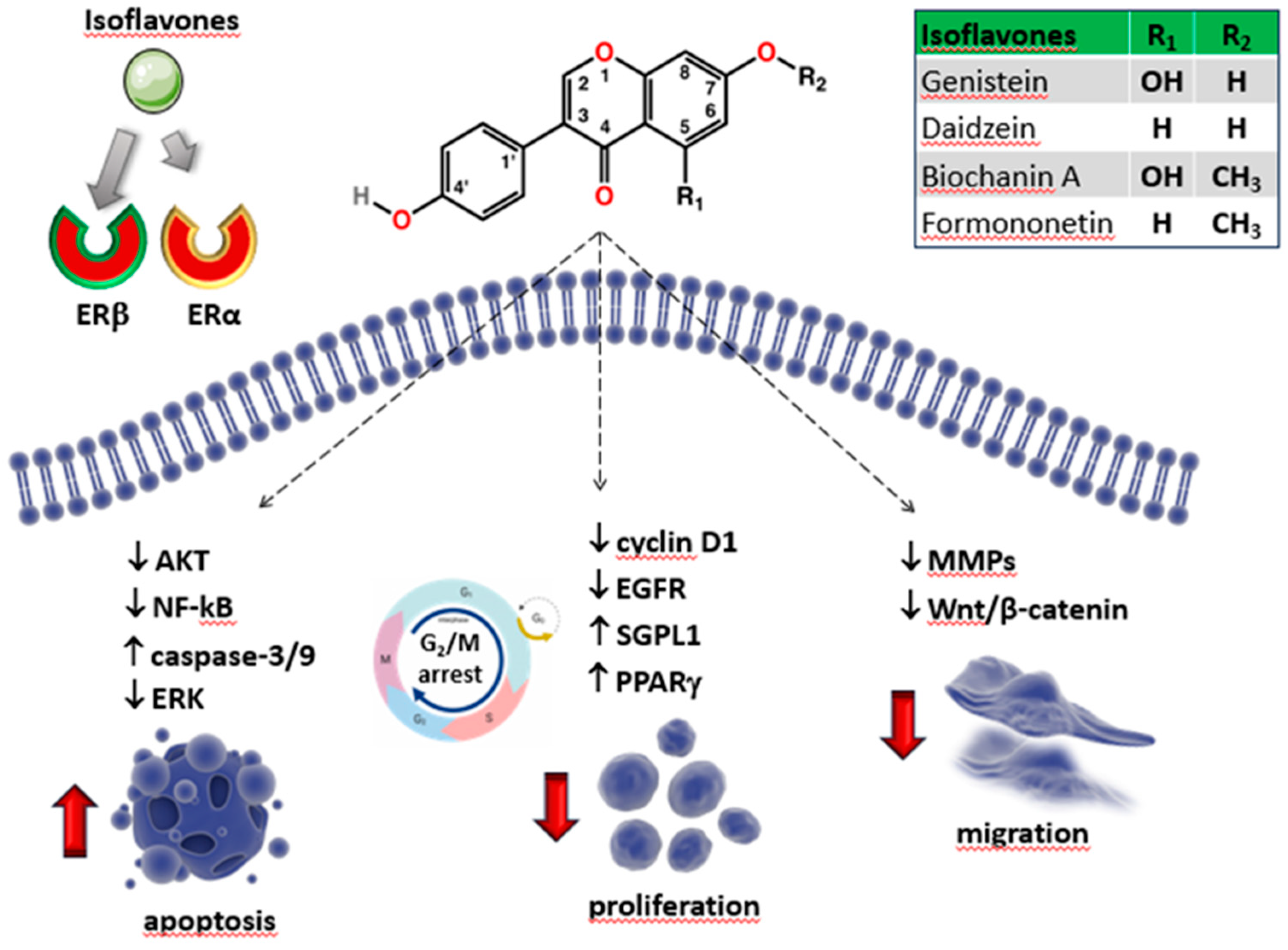
Genistein and its related compounds are well known for their weak estrogen-like activity in mammals, including the prevention of bone loss and other estrogen decrease-related conditions [145]. Indeed, data have revealed a reduced occurrence of breast and prostate tumors, as well as osteoporosis, in human populations that consume a soybean-rich diet [146]. Isoflavones interfere with ERα and ERβ isoforms with higher binding affinity for the latter, particularly genistein (20- to 30-fold higher) and daidzein (five-fold) [19] (Table 1), producing distinct clinical effects from estrogens and contributing to the treatment of various cancers, such as gastric, breast, prostate and non-small cell lung cancer prostate cancer [139,147,148,149].
Consistent with the above-reported Erβ anti-proliferative effects [10], it is believed that the chemotherapeutic potential of isoflavones is mainly due to their interactions with ERβ and vary in different cancer types, based upon their specific selectivity for target cells and their concentration, highlighting the complexity of dietary isoflavone functions [150,151,152]. ER-independent signaling mechanisms have also been described for isoflavones, including protein kinase regulation, enzymatic inhibition, growth factor modulation, antioxidant activity, or epigenetic changes [43].
Among the soybean isoflavones, genistein has shown to be one of the most potent inhibitors of cell proliferation in vitro, by affecting the protein tyrosine kinase (PTK) activity [153], suppressing the mammalian DNA topoisomerase II [139], and interacting with ERs on the nuclear envelope to promote G2/M phase arrest in various types of cancer cell lines [170,180]. The anticancer efficiency of genistein has been proven in numerous preclinical investigations [139]. Genistein from soy extracts, its free form, and its glycoside genistin are readily bioavailable and well tolerated with minimal toxicity [139,142]. Studies on OS cells suggested that genistein and its naturally occurring prenylated derivatives exert an inhibitory activity by binding ER isoforms with a statistically significant effect from 1 nmol/l [155]. The specific activation of ER signaling by genistein has been reported to be involved in the down-regulation of the expression of the EGFR gene. This influences events strictly controlled by its signaling pathway, such as the differentiation of U2OS human OS cells that stably express ERs [156]. Indeed, previous studies have demonstrated that genistein and daidzein can induce osteoblast differentiation through the enhanced expression of ALP, bone morphogenetic protein 2 (BMP-2), and OPG, [157,158], which suggests that soybeans are able to prevent osteoporosis. In addition, the treatment of osteoblasts with genistein, and other soybean isoflavones, induces calcified bone noduli [159]. The osteogenic activity of genistein was also observed in human MG-63 OS cell line [160], and murine cell line LM8 [161], using a variety of approaches. Treatment with genistein switched the osteoblasts towards a more differentiated phenotype that was able to synthesize many of the essential factors required for the production of new mineralized extracellular matrix, such as collagen type I (Col I) and ALP, even in a nonosteogenic growth medium [160].
As mentioned earlier, extensive vascularization and rapid growth are considered to be associated with the high metastatic potential and recurrence rate of OS [83,100].
The modulation of ECM components is a key element in cancer progression and invasion [101,102]. Glycosaminoglycans (GAGs)/proteoglycans (PGs) are the major macromolecules composing ECM [162]. Their localization can be both extracellular and cellular (cell membrane and intracellular granules) and they participate in the regulation of various cellular events, such as cell adhesion, migration, differentiation and proliferation, critically affecting broader aspects in cancer initiation and progression [162]. In vitro studies have shown that pathogenesis of OS implies qualitative and quantitative changes in PGs, which have important consequences on cell proliferation and/or differentiation [163]. Nikitovic and coauthors [164] investigated the activity of genistein on the synthesis and distribution of GAGs/PGs in MG-63 and SaOS-2 cell lines, which differ in the density of the ERs they express. They observed a dose-dependent inhibitory effect of genistein on OS cell growth, which was accompanied by a reduction of both secreted and cell-associated GAGs/PGs by both cell lines, so suggesting a PTK mechanism. On the other hand, MG-63 cell line showed a complex pattern of PG synthesis, indicating a dual action of genistein via the PTK mechanism and through the ER receptors, which are present in much higher density in MG-63 cells compared to SaOS-2 cells [164].
Other than ERs, genistein can also regulate other nuclear malignancy-related receptors such as peroxisome proliferator-activated receptor γ (PPARγ), an isotype of PPARs, which may further explain its activity against a range of cancers, including OS [165,166]. In OS, PPARγ plays a key role in suppressing cell proliferation and promoting osteoblastic terminal differentiation, suggesting a potential use of PPARγ agonists as chemotherapeutic and/or chemopreventive agents for human OS [113]. It has been reported that genistein treatment triggers growth inhibition and G2/M cell cycle arrest in human MG-63 cells, by acting as a non-toxic activator of PPARγ expression [166]. In addition, the genistein-induced PPARγ signaling resulted in the downstream activation of the tumor suppressor phosphatase and tensin homolog (PTEN), which in turn potently regulated the PI3K/Akt pathway, involved in cell proliferation and survival [166]. Additionally, the anti-OS activity of genistein was examined in SaOS-2 and MG-63 OS cells in combination with Calcitriol [167], the most active component of the vitamin D family [168]. The synergistic action of both these substances had already proven effective in preventing osteoporosis and reducing hip fracture risk in postmenopausal women [169]. In OS cells, combined treatment significantly increased the expression of the enzyme sphingosine-1-phosphate lyase (SGPL1) [167], which irreversibly degrades sphingosine-1-phosphate (S1P), whose production and secretion is associated with an increased capability of migration and invasion of cancer cells [170]. Besides the synergistic effects on the proliferation behavior, co-treatment increased the expression of vitamin D receptor (VDR) and ERβ and influenced the cellular metabolism by decreasing extracellular acidification (which is a measure of glycolytic flux in cancer cells) as well as cell respiration rates (a measure of mitochondrial respiration) [167].
Gemcitabine is one of the commonly used anti-metabolite drugs. It is an analog of cytosine arabinoside which shows pronounced anti-tumor activity, in vitro and in vivo, in a variety of solid tumors, including OS [171]. Data reported by Zhang et al., showed that the combination of gemcitabine and genistein enhance the antitumor efficacy of gemcitabine in MG-63 and U2OS cell lines and help to overcome resistance to gemcitabine treatment [172]. The induction of NF-kB activation and up-regulation of its target genes by many anticancer agents, including gemcitabine, may mediate the cellular resistance to anticancer drugs [173]. In contrast, the activation of the Akt and NF-kB signaling pathways can be inhibited by genistein [147]. Combined treatment of genistein with gemcitabine reversed the cancer’s resistance to gemcitabine by abrogating the Akt/NF-κB pathway, which led to a significant reduction in cell viability and induction of apoptosis in OS cell lines [172,174]. The structure of genistein and some of its potential effects on OS cells are reported in Figure 3 and Table 2.
6.1.1. Daidzein
The analog of genistein, daidzein (7,4-dihydroxyisoflavone) (Figure 3), is a well-studied isoflavone with a high nutritive value. It is predominantly found in soy and many unfermented foods not only in the form of daidzein, a glycoside conjugate, but also as acetylglycoside and aglycone [175]. The complex daidzein pharmacokinetic features, along with its insolubility in water and oil, have blocked their use as a highly common compound in medicine or as a nutraceutical [175].
Although daidzein has been proven to have in vitro antitumor effects on a variety of cancers [8,176,177,178], there are few reports concerning its inhibitory activity towards OS [156,179] (Table 2). Daidzein inhibited proliferation and cell cycle progression and promoted apoptosis in U2OS cell lines that stably express ERs [156]. In addition, similarly to genistein, daidzein is able to modulate the level of EGFR through the specific activation of both ER isoforms, thus affecting multiple intracellular events that are tightly controlled by the EGFR transduction pathway, such as the maturation of osteoblasts [156].
Recently, Zhu and coworkers investigated the underlying mechanism of daidzein against OS by means of an in-depth systematic pharmacology analysis, which identified the Src-MAPK pathway as the highest-ranked target of daidzein in 143B and U2OS cells and in in vivo OS models [179].
6.1.2. Biochanin A
Biochanin A (5,7-dihydroxy-4’-methoxy-isoflavone) (Figure 3) is a phytoestrogenic isoflavone found in legumes, particularly red clover (Trifolium pratense), but it is also present in cabbage, alfalfa, and many other herbal products [180]. The biological effects of biochanin A observed in vitro and in vivo are different from those observed for its derivative genistein. It plays complex roles in the regulation of multiple biological functions by binding DNA and some specific proteins or acting as a competitive substrate for some enzymes. Biochanin A extract from plants is already commercially available because of its potential benefits to human health despite its limited bioavailability [181,182]. It is used for the reduction of oxidative stress, treatment of osteoporosis, anti-inflammatory effects, regulation of blood glucose levels, and for treatment of allergies [183,184]. Moreover, like other isoflavones, biochanin A exhibits chemopreventive activity against various cancers [182].
Several studies have suggested that biochanin A has the potential to prevent and treat OS [185,186] (Table 2). The antiproliferative potential of biochanin A and its underlying mechanisms in human OS have been investigated by Hsu and et al. They reported that biochanin A dose-dependently inhibits cell growth and induces apoptosis in MG-63 and U2OS OS cells by triggering the activation of the intrinsic mitochondrial pathway and caspase-9 and -3 and increasing the Bax: Bcl-2/Bcl-XL ratio.
DOX is probably the most studied drug in combination with phytoestrogens. It is an antitumor antibiotic which acts by interfering with the enzymes involved in DNA replication or by causing strand breakage [187]. Hsu et al. also demonstrated that a combination of the chemotherapeutic agent DOX with biochanin A had synergistic cytotoxicity [185]. Supporting evidence for the anti-OS activity of biochanin A has been recently reported by Zhao et al. [186]. Biochanin A treatment clearly increased apoptotic rates and decreased migration and invasion ability in MG-63 and U2OS cells, in a time and dose dependent manner. Relevant genes involved in cell proliferation, apoptosis, invation and migration (i.e. proliferating cell nuclear antigen (PCNA), caspase-3, cyclinD1, Bcl-2, MMP-9, N-cadherin and E-cadherin) showed altered expressions in both OS cell lines [186].
E-cadherin is an important factor of cell-cell adhesion. It is classified as a cancer depressor because the loss of E-cadherin may lead to destruction of cytoskeleton promoting cells invasion and migration [188]. The expression of N-cadherin in epithelial tumour characterizes tumorigenesis as it may induce EMT, reinforce tumour cell activeity and promote the interaction between tumour and neighbouring cells [189]. OS cells treated with biochanin A exhibited decreased expression of MMP-9 and N-cadherin, while showing increased expression of E-cadherin. These findings strongly indicated that biochanin A may possess a suppressive function in cancer invasion and migration by mediating the process of tumor EMT [186].
6.1.3. Formononetin
Formononetin (7-hydroxy, 4′-methoxy isoflavone) (Figure 3) is the active ingredient of the traditional Chinese medicine astragalus, angelica, and Pueraria lobate. Formononetin exhibits anticancer effects against ovarian cancer, colorectal cancer, and gastric cancer, by suppressing cell viability and inducing apoptosis through the regulation of the estrogen-dependent signalling pathway [190,191].
Reports have also documented the anti-OS activity of formononetin (Table 2). It promoted the apoptosis of human bone cancer in vitro and in vivo in nude mice that had undergone orthotopic tumor implants, by modulating the expression levels of the apoptosis-related factors ERK, Akt [192], Bcl-2, Bax, caspase-3 and decreasing the level of miR-375, an ER signaling related miRNA, in ER-positive U2OS cells [193]. Further evidence showed that in MG-63 cells, the anti-proliferative function of formononetin is related to the up-regulation of the expression of the tumor suppressor PTEN gene, via miR-214-3p [194], which is one of the miRs with oncogene properties that is considerably increased in OS [195]. Recently a bioinformatic-based network pharmacology has been used to disclose other therapeutic targets and bio-mechanisms of anti-OS formononetin activity [196]. The biological processes of formononetin against OS were principally linked to the regulation of cell motility, cell proliferation as well as the regulation of gene expression. The main core targets of formononetin were determined as estrogen receptor 1 (ESR1), TP53 and receptor tyrosine-protein kinase erbB-2 (ERBB2). Interestingly, they were representatively validated following formononetin treatment in vivo, in tumor-bearing mice and clinical cases, suggesting that these predictive targets might be potential biomarkers for the screening and treatment of OS [196].
6.2. Flavonols
6.2.1. Quercetin
An intake of flavonols is found to be associated with a wide range of health benefits which includes antioxidant potential and anti-inflammatory effects. Flavonols may also play a role in reducing the risk of chronic diseases such as cardiovascular disease, cancer, and neurodegenerative disorders [197].
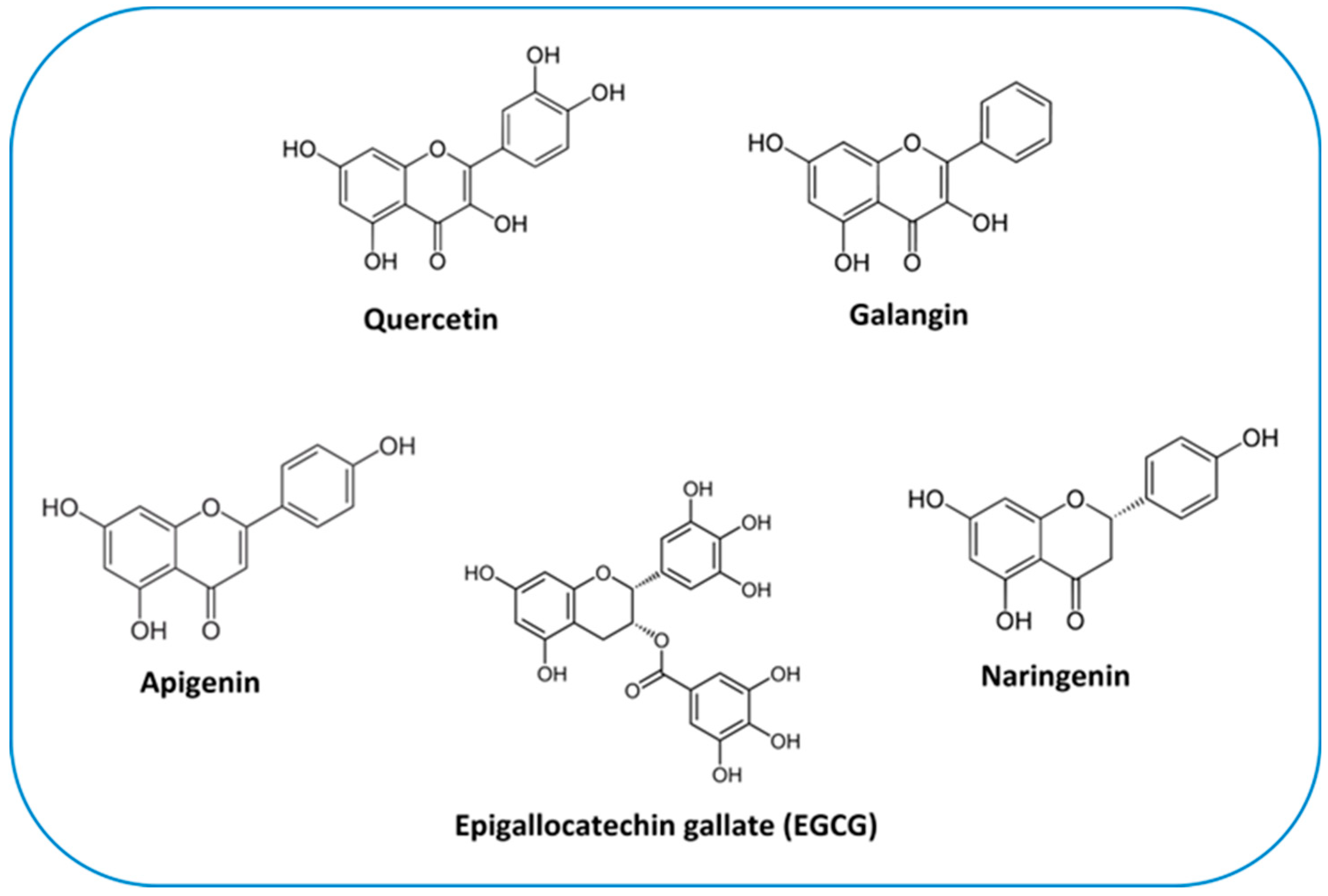
Quercetin (3,5,7,3′,4′-pentahydroxyflavone) (Figure 4) is the most abundant plant pigment in the extensive class of flavonols. It can be found in numerous plant species and in daily foods such as vegetables (caper contains the greatest quantity in relation to its weight), fruit, nuts and teas. The average daily dietary intake of quercetin is estimated to be 16 mg [198]. Due to its potent anti-inflammatory and antioxidant effects, quercetin is considered to be an advantageous agent for therapeutic purposes for a number of diseases including cardiovascular diseases, arthritis, allergy and diabetes [199]. Indeed, it is commercially accessible as a supplementary agent. Oral administration of 1 g quercetin per day is safe and is absorbed up to 60% [200].
Figure 4.
Chemical structure of flavonoids discussed throughout the text.
Extensive studies have demonstrated the efficacy of quercetin for the prevention and treatment of several types of cancer, in in vivo and in vitro models [201]. Quercetin can significantly prevent the cell cycle, promote cell apoptosis, and suppress tumor invasive behaviour via a variety of mechanisms [202]. There is evidence that several of the effects of quercetin on the survival of cancer cells might rely on ER-dependent mechanisms.
In particular, it has been shown that quercetin can impair the ERα-mediated rapid signaling, preventing the anti-apoptotic and proliferative ERK/MAPK and PI3K/AKT pathway activation [203]. Thus, quercetin can influence cancer cell proliferation and survival by acting as partial antagonists of ERα-activated rapid signals [204,205]. On the contrary, quercetin can behave as an E2-mimetic agent in the presence of ERβ, by activating the p38/MAPK and the downstream pro-apoptotic pathway, indicating a differential agonist or partial antagonistic effect of this compound in the definition of their anti-carcinogenic potential [206].
A strong cytotoxic activity of quercetin against OS has been largely demonstrated (Table 2). Although the related studies are mainly limited to in vivo and in vitro investigations, findings are promising. Quercetin treatment results in the suppression of proliferation, cell cycle arrest, induction of apoptosis, and reduced potential for adhesion and migration in several human OS cells lines, including 143B [207,208], HOS, MG-63 [209,210], U2OS and SaOS-2 cells [211]. Regarding the effect on OS cell growth, a number of key elements of the cellular apoptotic signaling pathway seems to be involved in the quercetin-dependent regulation of apoptosis. In an early study, Liang and coworkers [209] evaluated the effects of quercetin on the viability of human OS MG-63 cells. Quercetin treatment resulted in a decreased expression of the anti-apoptotic protein Bcl-2, which was paralleled with the increase of pro-apoptotic proteins BAX and cytochrome C, activation of caspase-3 and -9, and loss of mitochondrial membrane potential, indicating that quercetin was able to promote apoptosis via activation of the mitochondrial-dependent pathway [209]. Additional evidence demonstrated that the administration of quercetin to HOS and ATCC 1543 human OS cell lines leads to the cell cycle arrest at the G(1)/S phase accompanied by downregulation of cyclin D1, one of the cyclins required for G(1) to S progression. Subsequent apoptosis was induced by gradual activation of caspase-3 and subsequent poly (ADP-ribose) polymerase (PARP) cleavage [212].
The ability of quercetin to induce apoptotic cell death has also been associated with overcoming drug-resistance in OS cells. High-dose methotrexate (HDMTX) is considered to be a key agent in determining the chemotherapeutic outcome of OS patients [213]. However, drug resistance often develops in the late stage of treatment. Data reported by Xie et al., [214] revealed an apoptosis-inducing activity of quercetinin in an MTX-resistant OS model (U2OS/MTX300 cells). Exploring the mechanism underlying these effects, the authors found that cell death was accompanied by mitochondrial dysfunction and dephosphorylation of Akt, suggesting that quercetin-induced apoptosis might be associated with the apoptosis pathway of mitochondria and Akt activity [214]. Similar evidence was provided by Yin et al. [215], further sustaining the role of quercetin as a potential chemotherapeutic agent for MTX-resistant osteosarcoma.
OS is known to be a disease with a high propensity for metastasis to the lung [5]. In comparison to other cell lines, only 143B cells are able to generate a reliable and reproducible in vivo mouse model that develops primary tumors after intratibial injection within three to five weeks [216]. This mouse model also more closely reflects human disease because it develops metastases in the lung, which is the main location of metastasis in humans [217]. An early study by Berndt et al., [207] evaluated the anticancer properties of quercetin in human a 143B OS cell line. Quercetin treatment caused an arrest of 143B cells in the G2/M transition of the cell cycle. The cell cycle arrest was followed by cell death via activation of the apoptotic signaling pathway, as shown by a dose-dependent caspase-3, caspase-7 and PARP cleavage, after 36 h of incubation with quercetin. The pan-caspase inhibitor Z-VAD prevented PARP cleavage-dependent apoptosis. Quercetin also effectively blocked some hallmarks of metastatic behavior such as adhesion and migration. Overall, these findings suggested a role of quercetin as a potential drug that can target cells of the primary tumor and metastasizing foci [207].
Quercetin’s ability to induce cell death in OS cell lines is not limited to apoptosis. A recent work by Wu et al., [218] showed for the first time a role of quercetin in promoting autophagic cell death in human OS cells. It is well accepted that autophagy plays a two-faced role in cancer as a tumor suppressor or as a pro-oncogenic mechanism. Indeed, there is a dynamic relationship between the rate of protein degradation through autophagy, i.e. autophagic flux, and the susceptibility of tumors to undergo apoptosis which is critical in the autophagy/cancer relationship [219]. Therefore, an accurate modulation of this pathway is the challenge faced when targeting autophagy in the clinical setting [220].
The quercetin dependent increase of autophagic flux was shown by a dose-dependent up-regulation of LC3B-II/LC3B-I ratio, a hallmark of autophagy, in quercetin-treated MG-63 cells. Moreover, pharmacological inhibition of autophagy or genetic blocking autophagy by autophagy-related gene 5 (ATG5) knockdown efficiently protected against quercetin-induced cell death, supporting the role of quercetin in triggering autophagic cell death in MG-63 cells [218]. The authors also investigated the effect of quercetin exposure on the transcription factor nuclear protein 1 (NUPR1) activity [218]. NUPR1 is a master regulator of autophagy flux typically expressed in response to stress signals induced by genotoxic signals and agents [221]. Under stressful conditions such as chemotherapy treatment, an interplay between the homeostatic NUPR1 and autophagy pathways may occur in cancer cells, ultimately dictating their fate between cell death or survival [221]. Data revealed that quercetin increased NUPR1 expression and activated NUPR1 reporter activity, which contributed to the expression of autophagy-related genes by disturbing ROS homeostasis, indicating an important role for NUPR1 signalling in quercetin-induced autophagy in MG-63 cells [218].
Activating local cell invasion and distant metastasis represents another important hallmark of cancer that mainly reflects the progression of carcinomas to a higher grade of malignancy. Cell migration is a critical process for cancer cell spread, invasion, and distant metastasis [222]. Hypoxia-inducible factor (HIF)-1α was shown to be correlated with tumor grade, metastasis, and poor outcomes in various cancers including OS.
Its upregulation results in increased cell proliferation and migration, as well as the development of chemotherapeutic resistance [223]. Lan et al., [210] reported a dose- and time-dependent reduction of cell migration and invasion in human HOS and MG-63 cell lines, which was paralleled with a significant quercetin-induced downregulation of the expression of HIF-1α and downstream genes such as, VEGF, MMP-2, and MMP-9, which play essential roles in promoting cancer invasion and metastasis. In addition, quercetin treatment suppressed the formation and proliferation of metastatic lung tumors in vivo, in an OS nude mouse model [210]. In U2OS and SaOS-2 cells, the quercetin-mediated inhibition of the metastatic phenotype was associated with a significant downregulation of parathyroid hormone receptor 1 (PTHR1) [211], an important G-protein coupled receptor involved in OS pathophysiology [224].
Many lines of compelling data indicated the role of quercetin in sensitizing cancer cells to the action of several anti-cancer drugs [225,226]. In this respect, synergistic anti-tumor activities of quercetin and CDDP, a widely used chemotherapeutic agent, have been reported in cancer treatment in several in vivo and in vitro models [225,226,227].
Regarding OS, the efficacy of quercetin in enhancing CDDP sensitivity has been demonstrated by Zhang et al. in 143B cells [208]. miR-217 is a tumor suppressor which inhibits cell proliferation and metastasis in OS. Its over-expression could reverse CDDP chemoresistance in lung cancer cells [228]. Expression of miR-217 was upregulated after quercetin and/or CDDP treatment, while its target Kirsten rat sarcoma virus (KRAS) was downregulated both at mRNA and protein levels, thus suggesting an involvement of the miR-217-KRAS axis in the QUE-improved sensitivity of CDDP [208]. Quercetin has also been shown to be effective in boosting the anticancer activity of MTX on SaOS-2 cancer cells. A decline in MTX IC50 value was observed from 13.7 ng/ml to 8.45 ng/ml in the presence of quercetin. Moreover, the mRNA expression outcomes indicated that the combination therapy significantly up-regulates the tumor suppressor genes, such as p53, CBX7, and CYLD, and declines anti-apoptotic genes BCL-2 and miR-223, which can lead to proliferation, inhibition and apoptosis inducement [229].
Taken together, these findings provide experimental evidence of the effectiveness of quercetin in treating OS in human cell lines. On the other hand, further research including clinical trials are needed to support the future development of quercetin as an effective and safe candidate agent for the prevention and/or therapy of OS.
6.2.2. Galangin
Galangin (3,5,7-trihydroxyflavone) (Figure 4) is another plant flavonol from the flavonoid group of polyphenols. It is primarily extracted from the rhizome of Alpinia officinarum, which has been used as a herbal medicine in Asia for decades. Studies have shown that galangin has anti-inflammatory [230], antibacterial [231] and antiviral [232] activities in vitro. Currently, its antitumor properties are subject of attention. Studies have shown that galangin suppresses the proliferation and functions of various tumor cells [233,234,235]. However, research on galangin’s effects on OS specifically is limited (Table 2). In a study conducted by Yang et al. [236], exposure to galangin significantly suppressed MG-63 and U2OS cells by inhibiting their proliferation and invasion and triggering their apoptosis, in a concentration-dependent manner. The underlying mechanism was associated with the suppression of PI3K and its downstream regulators, cyclin D1 and MMP-2/9, and upregulation of p27Kip1, caspase-3, and caspase-8 [236]. Other evidence indicated that, in addition to effectively attenuating cell proliferation, incubation of MG-63 and U2OS cells with galangin increased dose dependently the expression levels of several markers for osteogenic differentiation such as, Col I, ALP, OPN, OCN and the transcription factor Runx2. Galangin could also attenuate OS growth in vivo, in the xenograft mouse model [237]. There are several cytokines that control bone formation, among which TGF-β1 has been proven to be fundamental in osteoblastic differentiation and bone matrix synthesis, through Smads-dependent signaling [238]. TGF-β1 secretion and the phosphorylation of Smad2 and Smad3 were triggered in a dose-dependent manner after galangin treatment, clearly indicating that galangin-mediated cell differentiation was dependent on its selective activation of the TGF-β1/Smad2/3 signaling pathway [237].
6.3. Apigenin
Apigenin (4′,5,7-trihydroxyflavone), is the main compound in the flavone group (Figure 4). It is widely contained in many fruits and vegetables such as oranges, tea, chamomile, onions and wheat sprouts [239]. Apigenin is thought to protect cells against oxidative damage by enhancing mitochondrial function [240]. Furthermore, it possess anticancer properties in vitro and in vivo, inducing cell cycle arrest and DNA damage in different types of cancer cells [239,241,242].
Results reported by Lin et al. showed that apigenin triggers apoptosis in U2OS cells and inhibits the xenograft tumor growth [243]. Liu et al., [244] confirmed the cytotoxic role of apigenin on both U2OS and MG-63 cells and investigated the underlying molecular mechanisms of its anti-OS effect.
The canonical Wnt-β-catenin signaling pathway is widely expressed in bone tissue and cells, and its deregulation is closely associated with the progression of OS [103,104,245]. It has been reported that a decreased β-catenin expression can downregulate MMP-14 expression, thereby resulting in suppression of the invasion and motility of MG-63 cells [246]. Apigenin was able to decrease the expression of β-catenin. Moreover, overexpression of β-catenin reversed the inhibitory effect of apigenin on OS cells, and knockdown of β-catenin enhanced apigenin-inhibited proliferation and invasion in OS cells. These results supported the hypothesis that Wnt/β-catenin pathway is involved in OS cell proliferation and invasion in response to apigenin [244].
6.4. Naringenin
Citrus fruits (grapefruit and oranges) and tomatoes are rich sources of flavanones, which are known for their numerous health benefits due to their ability to scavenge free radicals in various basal metabolic conditions [26].
Naringenin (4′,5,7-trihydroxyflavonone), especially abundant in the Mediterranean diet, is the most extensively studied flavanone (Figure 4). Despite its limited bioavailability [247], it shows great promise in various therapeutic applications. Its notable benefits in in vivo and in vitro models, which have been comprehensively reviewed by Arafah et al. [248], include anti-inflammatory, antioxidant, and anticancer properties. However, a scarce number of clinical studies have been conducted to date, compromising its commercial exploitation [247]. The generation of ROS is one of the key mechanisms responsible for promoting different stages of cancer [249]. Naringenin, as an antioxidant, does efficiently counter such effects [248]. In this regard, it has been reported that a long-term administration of naringenin (20 mg daily) inhibits OS progression and local recurrence in the patients (n = 47) who underwent OS surgery by improving the antioxidant and anti-inflammatory capacities of OS patients [250].
The cytotoxicity of naringenin has been demonstrated in numerous in vitro studies using various kinds of cancer cells e.g., breast, colon and liver cancer cell lines [251,252]. Antioxidant activities, kinase and glucose uptake inhibition have been proposed as molecular mechanisms for these effects. In addition, naringenin stimulation is believed to inhibit unregulated growth and induces apoptotic cascade in different cancer cell types by ERα or ERβ signalling [252]. Notably, naringenin shows an anti-estrogenic effect only in ERα containing cells; whereas in ERβ containing cells, naringenin mimics the E2 effects [252]. A recent work conducted by Lee et al., [253] has shown for the first time the cytotoxic and antiproliferative effects of naringenin on human OS cells. Although a high naringenin concentration was used during the study, it selectively inhibited the growth of OS cells (IC50 values for HOS and U2OS cells were 276 and 389 μM, respectively), with less cytotoxicity in normal human bone cells. The suppression of cell growth was accompanied by a significant increase in intracellular ROS generation and mitochondrial dysfunction, resulting in the activation of endoplasmic reticulum stress-mediated autophagy and apoptosis. This evidence suggested that this flavanone exerts its mechanism of action in cancer OS cells through ROS-mediated endoplasmic reticulum stress signaling pathways [253].
6.5. Catechins
Catechins, which are considered as readily applicable and safe phytochemicals [254], are the major bioactive constituent in green tea polyphenols. Their basic structure consists of a flavan-3-ol unit with a catechol (1,2-dihydroxybenzene) moiety. There are different types of green tea catechins (GTC), including epicatechin (EC), epicatechin gallate (ECG), epigallocatechin (EGC), and epigallocatechin gallate (EGCG), among others. These variations occur due to the number and position of hydroxyl groups attached to the flavan-3-ol structure. The arrangement of hydroxyl groups determines the distinct properties and potential health benefits of each catechin [255]. EGCG is the most abundant and biologically active catechin [256]. Its chemical structure is reported in Figure 4. Based on decades of research, EGCG has received considerable attention due to its inhibitory activities against carcinogenesis at all stages, i.e. initiation, promotion and progression [257,258]. Other catechins, such as ECG and EGC, have been shown to have similar, albeit lower, activities in numerous studies [259,260]. However, in humans, plasma bioavailability of GTCs is very low, which has been in part attributed to their oxidation, metabolism and efflux [261]. The low bioavailability and absorption of GTCs are considered to be the major reason behind the differing effects between in vitro and in vivo studies. In fact, extensive studies on the improvement of GTC bioavailability should help in this regard [261].
The bone strength mainly relies on selenium (Se), calcium, and vitamin (K and D) contents. Se deficiency is associated with the risk of developing multiple cancers including OS [262]. To minimize the risk associated with Se deficiency, its doping with hydroxyapatite (HAp) can be an effective approach which may potentially reduce the growth of OS cells. Currently, HAp has received considerable attention in reconstructive surgeries, orthodontic, and three-dimensional printing of scaffolds, owing to its high bioactive and osteoconductive properties [263,264]. Owing to the known antitumor properties of catechins, a recent study by Khan et al. aimed to develop catechin-modified Se-doped HAp nanocomposites (CC/Se-HAp) for potential application in OS therapy [265]. Cell toxicity analysis showed that CC/Se-HAp were rapidly internalized into the MNNG/HOS cells and improved anticancer activity compared to Se-doped HAp nanocomposite, by inducing ROS-mediated apoptosis through the activation of caspase-3 pathway [265]. This suggests that CC/Se-HAp nanoparticles possess the potential for the targeted treatment of OS. Further studies on the anti-OS properties of catechins have mainly focused on EGCG (Table 2). This green tea polyphenol can effectively inhibit the tumor characteristics of OS cells (i.e., 143B, MG-63, U2OS and SaOS-2), including proliferation, migration, and apoptosis [266,267,268]. Moreover, it protects local bone tissue from destruction and prevents lung metastasis of tumor cells [267,268]. These effects may occur by regulating the activity of the Wnt/β-catenin pathway, as demonstrated by cell and animal experiments [268]. Additional EGCG targets have been identified. In MG-63 and U2OS, the inhibitory activity of EGCG treatment has been partially associated with the upregulation of miR-1, one of the miRNAs critically involved in the pathogenesis and progression of human OS [267]. Moreover, combined administration of EGCG and IL-1 receptor antagonist (IL-1Ra) efficiently downregulated IL-1-induced IL-6 and IL-8 release from U2OS cells. This treatment approach also reduced the secretion of invasiveness-promoting MMP-2 and pro-angiogenic VEGF, without affecting the metabolic response and caspase-3 activity [266]. EGCG has also been shown to act as a chemosensitizer for DOX in OS models. The synergistic interaction between EGCG and DOX has been shown to be mediated by SOX2 overlapping transcript (SOX2OT), which contributes to suppress OS via autophagy and stemness inhibition ([269], p. 7).
Table 2.
Effects of flavonoids on osteosarcoma. Downward arrows represent downregulation or reduction. Upward arrows represent upregulation or increase.
Table 2.
Effects of flavonoids on osteosarcoma. Downward arrows represent downregulation or reduction. Upward arrows represent upregulation or increase.
| Phytoestrogen | Cell Line/ in vivo model |
Concentrations | Combined treatment |
Molecular Mechanism | Observed Effects | References |
|---|---|---|---|---|---|---|
| Genistein | U2OS | 1µM | ER-mediated down-regulation of EGFR |
↑ differentiation ↑ apoptosis ↑ cell cycle arrest |
[156] | |
| MG-63 | 2.5 – 30 μmol/L | ↑ matrix vesicles, ↑ ALP activity |
↑differentiation ↑mineralized bone noduli ↓ proliferation |
[160] | ||
| MG-63, SaOS-2 | 10 – 30 µM | ↓ PTK, ↑ ER | ↓ synthesis and secretion of GAGs/PGs | [164] | ||
| MG-63 | 40 – 80 µM | ↑ PPARγ pathway | ↑cell cycle arrest ↓ proliferation |
[166] | ||
| SaOS-2, MG-63 | 1 – 100 µM | calcitriol 10 nM for 48 h |
↑ SGPL1, VDR, ERβ | ↑ calcitriol sensitivity ↓ proliferation ↓ extracellular acidification ↓ mitochondrial respiration |
[167] | |
| MG-63, U2OS | 10 - 100 μmol/L | gemcitabine 0.5 µmol/l for 72 h |
↓ Akt/NF-κB pathway | ↑ gemcitabine sensitivity ↑ apoptosis |
[270] | |
| Daidzein | U2OS | 1 µM | ER-mediated down-regulation of EGFR |
↑ differentiation ↑ apoptosis ↑cell cycle arrest |
[156] | |
| 143B, U2OS xenograft mouse model |
10 - 500 μmol/L 20 mg/kg every 2 days |
↓ Src-ERK pathway | ↑ apoptosis ↑ cell cycle arrest ↓ migration ↓ tumour weights |
[179] | ||
| Biochanin A | MG-63, U2OS | 5 - 30 μg/mL | DOX 1μg/mL for 24 h |
↑ caspase-3/9, ↑ Bax: Bcl-2/Bcl-XL ratio |
↑ DOX sensitivity ↓ proliferation ↑ apoptosis |
[185] |
| MG-63, U2OS | 5 - 80 μM | ↓ PCNA, cyclin D1, Bcl-2, ↑ Bax, caspase-3; ↓ MMP-9, N-cadherin, ↑ E-cadherin |
↑ cell cycle arrest ↑ apoptosis ↓ migration, invation |
[186] | ||
| Formononetin | U2OS, tumor-bearing nude mice |
20 - 80 μM, 80 mg/kg/d |
↓ Bcl-2, miR-375, ↑ Bax | ↑ apoptosis ↓ tumour weights |
[193] | |
| U2OS | 5 - 100 μM |
↑ caspase-3 and Bax, ↓ Bcl-2 ↓ ERK and PI3K/AKT pathway |
↑ apoptosis |
[192] | ||
| MG-63 | 15 - 45 μM |
↑ miR-214-3p/PTEN pathway | ↓ proliferation ↑ apoptosis |
[194] | ||
| tumor-bearing nude mice |
25-100mg/kg/d | ↓ ESR1, p53, ERBB2 | ↓ tumour weights | [196] | ||
|
Quercetin |
MG-63 | 20 - 320 μM |
↑ cytochrome C, caspase-3/9, Bax, ↓ Bcl-2 |
↑ apoptosis |
[209] | |
| HOS, ATCC 1543 |
10 - 1000 µM | ↓ cyclin D1, ↑ caspase-3, cleaved PARP |
↑ cell cycle arrest ↓ proliferation ↑ apoptosis |
[212] | ||
| U2OS/MTX300 | 10 - 50 µM | ↑ cytochrome C, caspase-3, Bax, cleaved PARP; ↓Bcl-2, Akt | ↑ apoptosis ↓ proliferation |
[214] | ||
| 143B | 10 - 100 µM | ↑ caspase-3, cleaved PARP | ↑ cell cycle arrest ↑ apoptosis ↓ adhesion ↓ migration |
[207] | ||
| MG-63 | 50 - 200 µM | ↑ LC3B-II/LC3B-I ratio, ↓ ROS - NUPR1 pathway |
↑ autophagy | [218] | ||
| HOS, MG-63, tumor-bearing nude mice |
20 - 100 µM 25-100 mg/kg/d |
↓ HIF-1α, VEGF, MMP-2/9 | ↓ migration, invasion ↓ tumor growth |
[210] | ||
| U2OS, SaOS-2 | ↓PTHR1 | ↓ proliferation ↓ adhesion ↓ migration, invasion |
[211] | |||
| 143B | 5 μM | CDDP 5 μM for 24 h |
↑ miR-217- KRAS axis | ↑ CDDP sensitivity ↓ proliferation ↓ migration, invasion |
[208] | |
| SaOS-2 | 10 - 200 μM |
MTX 10-200 μM for 48 h |
↑ p53, CBX7, CYLD, ↓ Bcl-2, miR-223 |
↑ MTX sensitivity ↓ proliferation ↑ apoptosis |
[229] | |
| Galangin | MG-63, U2OS | 5 - 300 µM | ↓ PI3K/Akt pathway ↓ cyclin D1, MMP-2/9, ↑ p27Kip1, caspase-3/8 |
↓ proliferation ↑ apoptosis ↓ invasion |
[236] | |
| MG-63, U2OS tumor xenograft mouse |
25 - 100 μM 50, 100 mg/kg/d |
↑ TGF-β1/Smad2/3 pathway ↑Col I, ALP, OPN, OCN |
↑ differentiation ↓ tumor growth |
[237] | ||
| Apigenin | U2OS tumor xenograft mouse |
50 - 200 μM 2 mg/kg every 3 days |
↑ caspase-3/8/9, BAX, AIF |
↑ apoptosis ↓ tumor growth |
[243] | |
| U2OS, MG-63 |
20 - 100 μg/ml |
↓ Wnt/β-catenin pathway | ↓ proliferation ↓ invasion |
[244] | ||
| Naringenin | HOS, U2OS | 100 - 500 μM | ↑ ROS-Mediated ER Stress | ↑ autophagy ↑ apoptosis |
[253] | |
|
EGCG |
MG-63, 143B, SaOS-2 tumor-bearing nude mice |
10 - 50 μM 10 - 40 mg/kg every 2 days |
↓ Wnt/β-catenin pathway | ↓ proliferation ↓ migration, invasion ↓ tumor growth |
[268] | |
| MG-63, U2OS tumor-bearing nude mice |
0.0125 - 0.1 g/L 30 mg/kg/d |
↑ miR-1 | ↓ proliferation ↓ tumor growth |
[267] | ||
| U2OS | 5 - 50 μM | IL-1Ra1 ng/ml | ↓ MMP-2, VEGF ↓ IL-6/8 |
↓ proliferation ↓ invasion |
[266] | |
| U2OS, SaOS-2 | 20 μg/ml | DOX 1 - 2.5 μM | ↓ SOX2OT | ↑ autophagy | [269], p. 7 |
7. Anti-Osteosarcoma Effects of Non-Flavonoids
7.1. Stilbenes
7.1.1. Resveratrol
Resveratrol (3,5,4′-trihydroxy-trans-stilbene) is a powerful nutritional polyphenol found in more than 70 plant species and is especially abundant in food products such as red grapes (up to 14 mg/L) and derived products, peanuts, mulberries and soy [35,271]. Resveratrol is a phytoalexin. These chemicals are characterized by their low molecular weight and their ability to inhibit the progress of infections and other adverse stressful conditions for the plants [272]. Resveratrol is characterized by a stilbene structure which consists of two phenolic rings bonded together by a double styrene bond, which is responsible for the isometric cis- and trans-forms of resveratrol (Figure 5). The trans-isomer appears to be the more predominant and stable natural form [273]. There are many synthetic and natural analogues of resveratrol, as well as adducts, derivatives, and conjugates, including glucosides [274]. In red and white grape juice, resveratrol exists mostly as polydatin, its glycosidic form (see next section). Red grape juices contain significant amounts of trans-polydatin, followed by cis-polydatin and trans-resveratrol. These compounds are considered to be the primary compounds responsible for the health benefits associated with wine consumption [271].
Figure 5.
Downregulation of various pathways involved in the anti-OS activity of the stilbenes, resveratrol and its glycosidic form polydatin.
Figure 5.
Downregulation of various pathways involved in the anti-OS activity of the stilbenes, resveratrol and its glycosidic form polydatin.
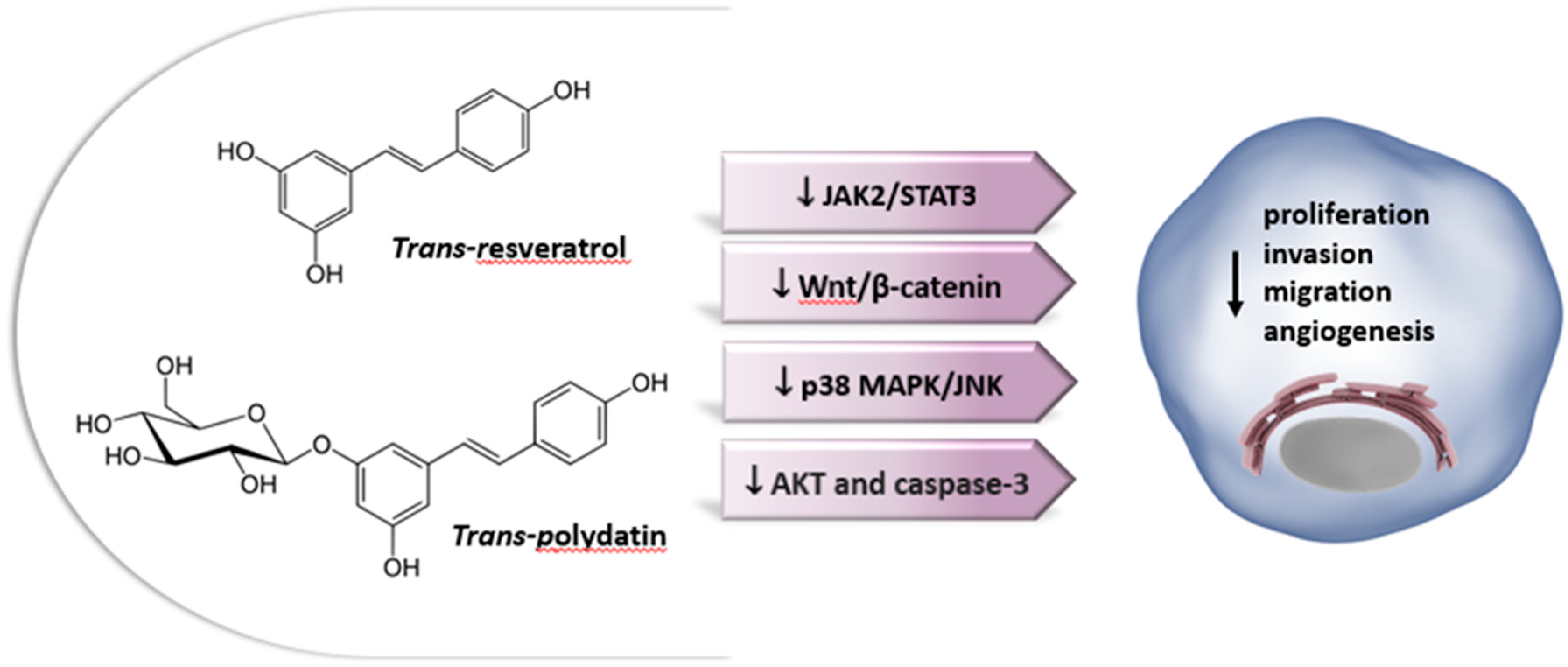
Resveratrol is commonly used as a nutraceutical in the management of high cholesterol, cancer, heart disease, as well as many other pathological conditions [275,276,277]. Its chemopreventive and anticancer effects, have been documented by in vivo and clinical studies in a wide variety of tumor cell types, highlighting its role in diverse cellular events associated with all stages of carcinogenesis, i.e., tumor initiation, promotion, and progression [8,278,279,280,281]. Unlike other phytoestrogens which bind ERβ with higher affinity, resveratrol binds ERα and ERβ with comparable affinity, but with 7,000-fold lower affinity than estradiol, acting as mixed agonist/antagonist [53]. Many lines of compelling data indicate that the effects of resveratrol on the survival of estrogen-related cancer cells might rely on ER-dependent mechanisms [282,283].
Regarding the bone microenvironment, resveratrol has been shown to have multiple bioactivities including antioxidative, anti-inflammatory, estrogen-like and proliferative properties that can influence bone metabolism [107,284]. In particular, in normal osteoblasts and osteoclasts, it regulates cell proliferation, cellular senescence and apoptosis, and inflammation processes, reducing the activity of NF-B and MAPK, and also acts through an epigenetic control, modulating the expression and activity of sirtuin-1 (SIRT-1), which is capable of increasing osteoblast survival and differentiation [285].
A body of studies on the effect of resveratrol on OS demonstrated a strong suppression of the cell viability, self-renewal ability and tumorigenesis of OS cells, whereas no significant inhibition effect on normal osteoblasts was observed [286,287,288,289,290,291]. However, the underlying mechanisms of action of resveratrol on OS cells have only been partially defined (Table 3). The Janus kinase 2/signal transducer and activator of transcription 3 (JAK2/STAT3) pathway is involved in different biological processes such as immunity, cell division, cell death, and tumor formation [292]. Aberrantly activated JAK2/STAT3 is frequently detected in many cancer diseases that are usually refractory to standard chemotherapy [292]. Studies have demonstrated a critical role of STAT3 signaling in the persistence of cancer stem cells (CSCs) [293], which is a primary cause of tumor relapse and metastasis [294]. Peng et al. [289], investigated the underlying molecular mechanism of resveratrol activity against OS CSCs, reporting that resveratrol treatment inhibited tumorigenesis ability by decreasing the synthesis of cytokines such as IL-6, IFN-γ, TNF-α and Oncostatin M, and preventing JAK2/STAT3 signaling pathway, which was consistent with the decline of the CSC marker CD133. On the other hand, exogenous STAT3 activation had the opposite effect and could abrogate the effect of resveratrol on CSCs, suggesting that resveratrol may be a promising therapeutic agent for OS management [289].
Additional studies have reported a correlation between resveratrol activity and the canonical Wnt/β-catenin cascade [288,295]. Aberrant activation of the Wnt-β-catenin signaling pathway plays a critical role in OS pathogenesis, which makes the Wnt/β-catenin pathway a hot topic in OS research [103,104,245]. Nevertheless, the development of therapeutic agents specifically targeting the aberrant Wnt activation in OS cells is still in its infancy. Exploring the effects of the resveratrol treatment against U2OS cells, Xie and coworkers found that the antitumor effects of resveratrol occurred through suppressing the activity of the Wnt/β-catenin pathway and the expression of related genes such as, c-myc, cyclin D1, MMP-2 and MMP-9 [295].
Moreover, resveratrol was able to up-regulate the expression level of Connexin 43 (Cx43) and E-cadherin [295], two important mediators of cell-cell adhesion which are critical in tumor progression [188,296]. The association between the anti-OS effect of resveratrol and the Wnt signaling was further demonstrated by Zou et al. [288].
Using β-catenin as a drug target, they performed a high content screening of botanical extracts to identify potential drugs against the human MG-63 OS cell line. β-catenin is a pivotal member of the canonical Wnt signaling pathway with the dual functions of regulating the coordination of cell-cell adhesion and gene transcription [297]. Aberrant expression of β-catenin activates numerous downstream target genes of the Wnt signaling pathway, a number of which are associated with cancer progression [298,299]. In a total of 14 botanical extracts assessed, resveratrol markedly downregulated the expression of β-catenin and significantly inhibited MG-63 cell proliferation [288].
OS is characterized by a high metastatic potential which is associated with a high death rate [83,100]. Proteinase enzymes, such as cathepsins, MMPs, and PA, are involved in many steps of tumor metastasis, including tumor invasion, migration, host immune escape, angiogenesis, and tumor growth [300]. MMPs, especially MMP-2 and MMP-9, are usually over-expressed in a wide range of human cancer types, including OS, providing a potential therapeutic target [102]. Based on in vitro, in vivo, and clinical evidence, Yang and coauthors showed that resveratrol can suppress the metastatic potential of human OS cells (i.e., HOS, MG-63, U2OS, Saos-2, and 143B) through transcriptional and epigenetic regulation of MMP-2, by respectively inhibiting cAMP CREB-DNA-binding activity and upregulating miR-328, which was initiated by the inhibition of the p38 MAPK/JNK pathways. Consistently, suppression of miR-328 significantly relieved MMP-2 and motility inhibition imposed by resveratrol treatment [301]. The proliferation, invasion and metastasis of tumor cells, as well as tumor relapse, are strongly correlated with the interactions of several factors, in which angiogenesis is a prerequisite. During this process, VEGF functions as the most significant vascular endothelial stimulating factor [302,303]. According to Liu et al. [304], resveratrol exerts a time and dose-dependent inhibition of OS cell invasion capabilities and proliferation, which is mediated by the downregulation of VEGF expression [304].
Recently, De Luca and colleagues [291] made a series of important observations on different human OS cell lines (i.e. MG-63, SaOS-2, KHOS, U2OS). They found that resveratrol was involved in the pAKT and caspase-3 pathway, causing cell growth inhibition and increase in apoptosis. Moreover, a significant increase in osteoblastic differentiation genes, such as osterix (Osx), OPN, ALP, Col I alpha 1, and OCN, was observed, suggesting that resveratrol may act as an inducer of differentiation, which is known to make OS cells more vulnerable to the action of chemotherapeutic agents [305]. Furthermore, they highlighted an epigenetic action of resveratrol on the promoters of interleukins IL-6 and IL-8, whose role in tumor progression is a well-described process in several cancer models [306,307]. The epigenetic change induced the reduction of the secretion of interleukins IL-6 and IL-8, which further explained the inhibitory effects of resveratrol on OS cellular growth and motility. In line with data previously reported [286], the OS cell lines examined, which represent the various typical characteristics of OS, responded in a variable manner to resveratrol treatment. This explained why by calculating the IC50 of resveratrol, different values were obtained, i.e. around 120 µM for MG-63 and SaOS-2 and around 60 µM for KHOS and U2OS treatment [291].
Strong evidence from breast, gastric, and prostate cancer cells subjected to combined treatment with resveratrol–DOX or resveratrol–CDDP has shown a synergistic behavior of resveratrol towards chemotherapeutic agents [308,309,310]. According to De Luca et al., the cotreatment of resveratrol with DOX and CDDP increased their cytotoxic effect on OS cells, suggesting that resveratrol might be a promising therapeutic adjuvant agent for OS cell treatment [291].
7.1.2. Polydatin
Polydatin (3, 4′, 5-trihydroxystibene-3-β-mono-D-glucoside), also known as piceid, is a naturally occurring glucoside derivative of resveratrol, in which the glucoside group linked to position C-3 replaces the hydroxyl group [311,312] (Figure 5).
Polydatin was first extracted from the roots of Polygonum cuspidatum (Polygonaceae), which have a long history of use in traditional Chinese and Japanese medicines, but it also exists in a variety of other sources including dietary plants such as grape, peanut, berries and chocolate [312]. The trans form of polydatin is well known for its high therapeutic potential in a variety of medical domains, for example infection, inflammation, cardiovascular disorders and aging-related diseases such as osteoporosis [313,314,315,316]. The Chinese FDA has approved polydatin for multiple phase II clinical trials, mainly for anti-shock applications [317].
Glucose substitution gives polydatin a more hydrophilic character than resveratrol, resulting in a significantly increased bioavailability and higher health-promoting/disease-modifying activities [318,319]. Furthermore, comparative studies of polydatin and resveratrol regarding antioxidative effects in vivo have revealed a better antioxidant activity of polydatin than resveratrol [311,320]. Polydatin has been recognized as a potent anticancer agent, with the ability to regulate various signaling pathways involved in the progression of several kinds of cancers [321,322]. It is mainly involved in cell cycle regulation, apoptosis, autophagy, signaling pathways, EMT, inhibition of inflammation and metastasis, and regulation of enzymes related to oxidative stress [323,324].
Moreover, recent studies by Mele et al. demonstrated that polydatin exerts a significant cytotoxic effect on cancer cells by Glucose-6-phosphate dehydrogenase inhibition, a rate-limiting enzyme in the pentose phosphate pathway which is altered in different malignant tumors [325]. A beneficial role of polydatin was also documented in prevention and treatment of OS. In an early study, Xu et al. evaluated the anti-OS activity of polydatin in human OS cell lines (i.e., 143B and MG-63). Polydatin dose-dependently inhibited proliferation by suppressing the β-catenin signaling and promoted apoptosis via upregulated expressions of Bax/Bcl-2 and caspase-3 in OS cells [326].
Polydatin also induced apoptosis via different mechanisms such as reducing the expression/phosphorylation of STAT3 and increasing the expression of autophagy-related genes (Atg12, Atg14, BECN1, PIC3K3), thereby triggering autophagic cell death in MG-63 cells [327]. Further studies reported the efficacy of polydatin in drug-resistant models of OS [328,329]. The therapeutic effect of polydatin against DOX-resistant OS, in vitro and in a MG-63/DOX xenograft model, occurs via the oncogene Taurine upregulated gene 1 (TUG1) mediated suppression of Akt signaling, which promotes apoptosis and prevents cell proliferation [329]. Additionally, polydatin enhances the chemosensitivity to the antineoplastic agent paclitaxel of U2OS and MG-63 cells and their paclitaxel-resistant variants, suppressing cell growth and migration and inducing cell-cycle arrest in the S phase [328]. Interestingly, a recent work investigated the role accomplished by polydatin, alone or after radiation therapy, in the osteogenic differentiation of SaOS-2 and MG-63 [330].
As mentioned above, the osteogenic differentiation has a role in the pathogenesis of OS considering that OS tumors deregulate the signaling pathways associated with osteogenic differentiation by arresting the cells as undifferentiated precursors [112].
In combination with radiotherapy, the pretreatment with polydatin promoted a radiosensitizing effect on OS cancer cells as demonstrated by the increased mineralization and osteogenic markers levels. The differentiation process was paralleled by the activation of the Wnt-β-catenin pathway and cell cycle arrest in S-phase. Additionally, the secretion of sphingolipid, ceramides, and their metabolites were analyzed by mass spectrometry in OS treated cells. In past years, evidence has accumulated demonstrating that 2′-hydroxy ceramide/sphingolipids have distinct biological functions to regulate various cellular processes and cell differentiation by binding to specific target proteins [331].
Moreover, Sphingosine-1-phosphate has been reported to inhibit osteoclast formation and mineralization [332]. MS analysis demonstrated that polydatin-induced osteogenic differentiation was mediated by an increased expression and secretion of ceramides and sphingolipids and pretreatment with polydatin sensitized OS cells to ionizing radiation, suggesting that polydatin, in combination with radiotherapy, can consolidate the response to therapy of OS cells [330].
Various novel drug delivery systems, including nanoparticles [333], liposomes [334], micelles [335], quantum dots [336] and polymeric nanocapsules [337] have been designed to enhance polydatin pharmacodynamics and pharmacokinetics [338]. Polycaprolactone (PCL) is a biodegradable hydrophobic polyester used to obtain clinically applicable implantable nanostructures [339]. In their study, Lama and coworkers demonstrated that PCL nanofibers complexed to polydatin supported the adhesion and promoted the osteogenic differentiation in both SaOS-2 cells and bone MSCs, providing evidence of the osteogenic capacity of polydatin to create a biomimetic, innovative and patented scaffold for both anticancer and regenerative purposes [340].
7.2. Lignans
In contrast to other types of phytoestrogens, lignan-type phytoestrogens have rarely been studied, although they are widely distributed in the plant kingdom. Cereals, grains, berries, and garlic are good dietary sources of lignans [32]. The plant lignans may occur in the form of aglycones and glycosides. After consummation, they can be converted by gut bacteria to form enterolignans (enterodiol and enterolactone), also known as “mammalian lignans”, which have a variety of biologic activities, including tissue-specific ER activation, and anti-inflammatory and apoptotic effects, that may influence disease risk in humans [341].
The effects of enterodiol and enterolactone on the viability and differentiation of MG-63 cells have been examined. Data suggested that both enterolactone and enterodiol have biphasic effects on cell proliferation, ALP activity and transcriptional levels of osteonectin (ON) and Col I, showing induction at low doses and inhibition at high doses.
The dose-dependent effects have been linked to the estrogenic and antiestrogenic properties of estrogen-like molecules as well as their ability to induce multiple signaling transduction [342].
Table 3.
Effects of non-flavonoids on osteosarcoma. Downward arrows represent downregulation or reduction. Upward arrows represent upregulation or increase.
Table 3.
Effects of non-flavonoids on osteosarcoma. Downward arrows represent downregulation or reduction. Upward arrows represent upregulation or increase.
| Phytoestrogen | Cell Line/ in vivo model |
Concentrations | Combined treatment | Molecular Mechanism | Observed Effects | References |
|---|---|---|---|---|---|---|
| Resveratrol | MNNG/HOS, MG-63 tumor xenograft mouse |
10 - 40 μM 100 mg/kg/d |
↑caspase-3, Bax, cleaved PARP, ↓ Bcl-2, Bcl-xL; ↓ cytokines ↓ JAK2/STAT3 pathway |
↑ apoptosis ↓ CSCs survival ↓ tumor growth |
[289] | |
| U2OS | 6 - 24 μg/ml | ↓ Wnt/β-catenin pathway ↓ β-catenin, c-myc, cyclin D1, ↓MMP-2/9 ↑ Cx43, E-cadherin |
↓ proliferation ↑ apoptosis ↓ migration, invasion |
[295] | ||
| MG-63 | 10 - 40 µg/ml | ↓ β-catenin signaling | ↓ proliferation | [288] | ||
| HOS, MG-63, U2OS, SaOS-2, 143B HOS orthotopic graft model |
25 - 200 μM 40, 100-mg/kg/d |
↓ p38 MAPK/JNK pathways ↓ CREB, ↓ MMP-2, ↑ miR-328 |
↓ migration, invasion, adhesion ↓ tumor growth, metastasis |
[301] | ||
| U2OS | 10 - 40 μM | ↓ VEGF | [304] | |||
| MG-63, SaOS-2, KHOS, U2OS | 50 -100 µM | DOX 0.1-10 µM or CDDP 0.2-2 µg/mL for 24 h |
↓ pAKT, ↑ caspase-3 ↓ IL-6/8 ↑ Osx, OPN, ALP, Col I, OCN |
↓ proliferation ↑ apoptosis ↑ differentiation ↑ DOX/CDDP sensitivity |
[291] | |
| Polydatin | 143B, MG-63 | 1 - 100 µM | ↓ β-catenin signaling ↑ Bax/Bcl-2, caspase-3 |
↓ proliferation ↑ apoptosis |
[326] | |
| MG-63 | 10 -160 µM | ↓ STAT3 signaling | ↑ apoptosis, ↑ autophagy | [327,328] | ||
| Polydatin | SaOS-2/ DOX, MG-63/DO xMG-63/ DOX xenograft model |
50 - 250 μM 150 mg/kg/d |
↓ TUG1/Akt signaling | ↓ proliferation ↑ apoptosis ↓ tumor growth |
[329] | |
| U2OS, MG-63 | paclitaxel | ↓ proliferation ↓ migration ↑cell-cycle arrest |
[328] | |||
| SaOS-2 | 1 - 150 μM |
ionizing radiation | ↓ Wnt/β-catenin pathway ↑ lipid metabolite secretion |
↑ differentiation ↑cell-cycle arrest ↑radiation sensitivity |
[330] | |
| Enterodiol, Enterolactone | MG-63 | 0.1–10 mg/ml | ↑ALP activity ↑ ON, Col I |
↓ proliferation ↑ differentiation |
[342] |
8. Conclusions and Perspectives
OS is a disease of multifactorial origin which involves a complex interaction between a wide variety of factors and mechanisms that when acting together promote the deregulation of cellular signaling pathways, causing disturbances in bone tissue homeostasis [82,83]. Though rare, OS is the prevalent form of bone cancer among children and young adults. Despite advancements in treatment modalities, the prognosis for OS patients remains unfavorable, particularly in cases of metastasis and resistance to conventional therapies [5,6]. With the aim of improving patient outcomes, researchers have turned their attention to exploring new second-line treatment options (i.e., complementary and integrative medicine).
Evidence collected from literature indicates that phytoestrogens hold promise as a complementary therapeutic approach for OS, making them attractive candidates for further exploration. Mechanistically, phytoestrogens can interfere with genomic and non-genomic signaling pathways, such as NF-kB, PI3K/Akt or MAPK/ERK, control cell cycle progression, initiate apoptosis events, inhibit angiogenesis and metastasis. Several reports have also highlighted their ability to enhance the efficacy of chemotherapy agents and overcome drug resistance. Among them, genistein, resveratrol, and quercetin have the most evidence of effectiveness [172,174,208,214,229,291,329].
Despite the abundance of in vitro studies, the role and involvement of ERα and/or ERβ in the phytoestrogens-dependent modulation of OS cells has been only partially investigated to date, although the estrogen signaling may be involved in the growth and progression of this malignancy [14,124,126].
To fully assess the impact of these compounds on OS initiation, progression, and development, the Erα/ERβ ratio in specific OS models should also be considered. This is important due to the considerable variation in ER-mediated cellular response based on cell type, along with the selectivity and concentration of phytoestrogens. Taken together, these factors play a crucial role in determining the multifaced effects exerted by these plant estrogenic molecules on cancer cells [49,50,51].
Although in vitro and preclinical studies have shown promising results, it is crucial to evaluate the true potential of phytoestrogens as a viable option for OS therapy through the rigorous assessment of their efficacy, safety, optimal dosage, and treatment duration in clinical settings. In this regard, preliminary clinical studies have been conducted to investigate the pharmacokinetics of compounds derived from plant sources, including phytoestrogens, in individuals with premalignancies or patients with different types of cancer [80,175,343]. Among the various molecules studied, only clinical investigations on green tea extracts have consistently yielded results, supporting their potential for chemoprevention in premalignant cervical disease, prostate cancer and leukoplakia. Moreover, existing data generally supports the safety of small doses of purified phytoestrogen consumption as a medication for breast cancer [201]. Conversely, further clinical trials are needed to sustain the role of soy isoflavones in preventing cancer development and progression [201,343]. Overall, the protective effect of phytoestrogens against cancer development remains a topic of ongoing debate [80]. This controversy stems primarily from a notable discrepancy between the outcomes of mechanistic studies conducted in vitro and the observations made in clinical settings. The main reason for these inconsistencies is the frequent use of non-physiological concentrations of phytoestrogens in mechanistic studies, which are difficult to achieve through normal dietary intake [80,344].
Interestingly, some ongoing research is focusing on the development of highly specific and long-acting analogues or drug delivery strategies to improve the pharmacodynamics and pharmacokinetics of phytoestrogensin in human OS models [265,281,333,337,340]. This may encourage the exploitation of the therapeutic potential of phytoestrogens in managing OS and other types of cancers.
Author Contributions
Writing—original draft preparation, A.C., C.C., G.F.F.; Table and Figure preparation, A.C., C.C., Writing–Review & Editing, A.C., C.C., M.G.; Supervision, C.C., M.G. and S.M. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The financial support from the Italian Ministery of University and Research (MUR Prin 2022PFFPBL) is gratefully acknowledged.
Conflicts of Interest
The authors declare no conflict of interest.
References
- G. Ottaviani e N. Jaffe, «The Epidemiology of Osteosarcoma», in Pediatric and Adolescent Osteosarcoma, N. Jaffe, O. S. Bruland, e S. Bielack, A c. di, in Cancer Treatment and Research, vol. 152. Boston, MA: Springer US, 2009, pp. 3–13. doi: 10.1007/978-1-4419-0284-9_1. [CrossRef]
- S. K. Denduluri et al., «Molecular pathogenesis and therapeutic strategies of human osteosarcoma», J Biomed Res, vol. 30, ago. 2015, doi: 10.7555/JBR.29.20150075. [CrossRef]
- B. Zhang, Y. Zhang, R. Li, J. Li, X. Lu, e Y. Zhang, «The efficacy and safety comparison of first-line chemotherapeutic agents (high-dose methotrexate, doxorubicin, cisplatin, and ifosfamide) for osteosarcoma: a network meta-analysis», J Orthop Surg Res, vol. 15, fasc. 1, p. 51, dic. 2020, doi: 10.1186/s13018-020-1576-0. [CrossRef]
- L. Marchandet, M. Lallier, C. Charrier, M. Baud’huin, B. Ory, e F. Lamoureux, «Mechanisms of Resistance to Conventional Therapies for Osteosarcoma», Cancers, vol. 13, fasc. 4, p. 683, feb. 2021, doi: 10.3390/cancers13040683. [CrossRef]
- Y. Arima, H. Nobusue, e H. Saya, «Targeting of cancer stem cells by differentiation therapy», Cancer Sci, vol. 111, fasc. 8, pp. 2689–2695, ago. 2020, doi: 10.1111/cas.14504. [CrossRef]
- S. Shukla, S. Ohnuma, e S. V. Ambudkar, «Improving Cancer Chemotherapy with Modulators of ABC Drug Transporters», CDT, vol. 12, fasc. 5, pp. 621–630, mag. 2011, doi: 10.2174/138945011795378540. [CrossRef]
- J.-J. Monsuez, J.-C. Charniot, N. Vignat, e J.-Y. Artigou, «Cardiac side-effects of cancer chemotherapy», International Journal of Cardiology, vol. 144, fasc. 1, pp. 3–15, set. 2010, doi: 10.1016/j.ijcard.2010.03.003. [CrossRef]
- M. Torrens-Mas e P. Roca, «Phytoestrogens for Cancer Prevention and Treatment», Biology (Basel), vol. 9, fasc. 12, p. 427, nov. 2020, doi: 10.3390/biology9120427. [CrossRef]
- X. J. Hu, W. R. Song, L. Y. Gao, S. P. Nie, G. Eisenbrand, e M. Y. Xie, «Assessment of dietary phytoestrogen intake via plant-derived foods in China», Food Addit Contam Part A Chem Anal Control Expo Risk Assess, vol. 31, fasc. 8, pp. 1325–1335, 2014, doi: 10.1080/19440049.2014.930562. [CrossRef]
- M. Cipolletti, V. Solar Fernandez, E. Montalesi, M. Marino, e M. Fiocchetti, «Beyond the Antioxidant Activity of Dietary Polyphenols in Cancer: the Modulation of Estrogen Receptors (ERs) Signaling», Int J Mol Sci, vol. 19, fasc. 9, p. 2624, set. 2018, doi: 10.3390/ijms19092624. [CrossRef]
- C. R. Sirtori, A. Arnoldi, e S. K. Johnson, «Phytoestrogens: end of a tale?», Ann Med, vol. 37, fasc. 6, pp. 423–438, 2005, doi: 10.1080/07853890510044586. [CrossRef]
- V. S. Ionescu, A. Popa, A. Alexandru, E. Manole, M. Neagu, e S. Pop, «Dietary Phytoestrogens and Their Metabolites as Epigenetic Modulators with Impact on Human Health», Antioxidants (Basel), vol. 10, fasc. 12, p. 1893, nov. 2021, doi: 10.3390/antiox10121893. [CrossRef]
- Z. Yang, M. Yang, W. Yu, e H. Tao, «Molecular mechanisms of estrogen receptor β -induced apoptosis and autophagy in tumors: implication for treating osteosarcoma», J Int Med Res, vol. 47, fasc. 10, pp. 4644–4655, ott. 2019, doi: 10.1177/0300060519871373. [CrossRef]
- S. C. Manolagas, C. A. O’Brien, e M. Almeida, «The role of estrogen and androgen receptors in bone health and disease», Nat Rev Endocrinol, vol. 9, fasc. 12, pp. 699–712, dic. 2013, doi: 10.1038/nrendo.2013.179. [CrossRef]
- M. Tobeiha et al., «Potential of natural products in osteosarcoma treatment: Focus on molecular mechanisms», Biomedicine & Pharmacotherapy, vol. 144, p. 112257, dic. 2021, doi: 10.1016/j.biopha.2021.112257. [CrossRef]
- T. P. Kondratyuk e J. M. Pezzuto, «Natural Product Polyphenols of Relevance to Human Health», Pharmaceutical Biology, vol. 42, fasc. sup1, pp. 46–63, gen. 2004, doi: 10.3109/13880200490893519. [CrossRef]
- P. Vuorela et al., «Natural Products in the Process of Finding New Drug Candidates», CMC, vol. 11, fasc. 11, pp. 1375–1389, giu. 2004, doi: 10.2174/0929867043365116. [CrossRef]
- R. J. Miksicek, «Estrogenic Flavonoids: Structural Requirements for Biological Activity», Experimental Biology and Medicine, vol. 208, fasc. 1, pp. 44–50, gen. 1995, doi: 10.3181/00379727-208-43830. [CrossRef]
- G. G. Kuiper et al., «Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta», Endocrinology, vol. 139, fasc. 10, pp. 4252–4263, ott. 1998, doi: 10.1210/endo.139.10.6216. [CrossRef]
- J. C. Le Bail, Y. Champavier, A. J. Chulia, e G. Habrioux, «Effects of phytoestrogens on aromatase, 3beta and 17beta-hydroxysteroid dehydrogenase activities and human breast cancer cells», Life Sci, vol. 66, fasc. 14, pp. 1281–1291, feb. 2000, doi: 10.1016/s0024-3205(00)00435-5. [CrossRef]
- I. M. C. M. Rietjens, A. M. Sotoca, J. Vervoort, e J. Louisse, «Mechanisms underlying the dualistic mode of action of major soy isoflavones in relation to cell proliferation and cancer risks», Mol. Nutr. Food Res., vol. 57, fasc. 1, pp. 100–113, gen. 2013, doi: 10.1002/mnfr.201200439. [CrossRef]
- P. Roca, J. Sastre-Serra, M. Nadal-Serrano, D. G. Pons, M. del M. Blanquer-Rosselló, e J. Oliver, «Phytoestrogens and mitochondrial biogenesis in breast cancer. Influence of estrogen receptors ratio», Curr Pharm Des, vol. 20, fasc. 35, pp. 5594–5618, 2014, doi: 10.2174/1381612820666140306100709. [CrossRef]
- I. Nikolić, I. Savić-Gajić, A. Tačić, e I. Savić, «Classification and biological activity of phytoestrogens: A review», Adv Techol, vol. 6, fasc. 2, pp. 96–106, 2017, doi: 10.5937/savteh1702096N. [CrossRef]
- R. E. Mutha, A. U. Tatiya, e S. J. Surana, «Flavonoids as natural phenolic compounds and their role in therapeutics: an overview», Futur J Pharm Sci, vol. 7, fasc. 1, p. 25, 2021, doi: 10.1186/s43094-020-00161-8. [CrossRef]
- T.-Y. Wang, Q. Li, e K.-S. Bi, «Bioactive flavonoids in medicinal plants: Structure, activity and biological fate», Asian J Pharm Sci, vol. 13, fasc. 1, pp. 12–23, gen. 2018, doi: 10.1016/j.ajps.2017.08.004. [CrossRef]
- B. A. Graf, P. E. Milbury, e J. B. Blumberg, «Flavonols, flavones, flavanones, and human health: epidemiological evidence», J Med Food, vol. 8, fasc. 3, pp. 281–290, 2005, doi: 10.1089/jmf.2005.8.281. [CrossRef]
- A. N. Panche, A. D. Diwan, e S. R. Chandra, «Flavonoids: an overview», J Nutr Sci, vol. 5, p. e47, 2016, doi: 10.1017/jns.2016.41. [CrossRef]
- M.-J. Xu et al., «Simultaneous characterization of prenylated flavonoids and isoflavonoids in Psoralea corylifolia L. by liquid chromatography with diode-array detection and quadrupole time-of-flight mass spectrometry», Rapid Commun Mass Spectrom, vol. 26, fasc. 19, pp. 2343–2358, ott. 2012, doi: 10.1002/rcm.6361. [CrossRef]
- C. Manach, A. Scalbert, C. Morand, C. Rémésy, e L. Jiménez, «Polyphenols: food sources and bioavailability», The American Journal of Clinical Nutrition, vol. 79, fasc. 5, pp. 727–747, mag. 2004, doi: 10.1093/ajcn/79.5.727. [CrossRef]
- A. Crozier, I. B. Jaganath, e M. N. Clifford, «Dietary phenolics: chemistry, bioavailability and effects on health», Nat Prod Rep, vol. 26, fasc. 8, pp. 1001–1043, ago. 2009, doi: 10.1039/b802662a. [CrossRef]
- A. Durazzo et al., «Dietary Lignans: Definition, Description and Research Trends in Databases Development», Molecules, vol. 23, fasc. 12, p. 3251, dic. 2018, doi: 10.3390/molecules23123251. [CrossRef]
- C. Rodríguez-García, C. Sánchez-Quesada, E. Toledo, M. Delgado-Rodríguez, e J. J. Gaforio, «Naturally Lignan-Rich Foods: A Dietary Tool for Health Promotion?», Molecules, vol. 24, fasc. 5, p. 917, mar. 2019, doi: 10.3390/molecules24050917. [CrossRef]
- C. Rivière, A. D. Pawlus, e J.-M. Mérillon, «Natural stilbenoids: distribution in the plant kingdom and chemotaxonomic interest in Vitaceae», Nat Prod Rep, vol. 29, fasc. 11, pp. 1317–1333, nov. 2012, doi: 10.1039/c2np20049j. [CrossRef]
- J. A. Sirerol, M. L. Rodríguez, S. Mena, M. A. Asensi, J. M. Estrela, e A. L. Ortega, «Role of Natural Stilbenes in the Prevention of Cancer», Oxid Med Cell Longev, vol. 2016, p. 3128951, 2016, doi: 10.1155/2016/3128951. [CrossRef]
- J. Burns, T. Yokota, H. Ashihara, M. E. J. Lean, e A. Crozier, «Plant foods and herbal sources of resveratrol», J Agric Food Chem, vol. 50, fasc. 11, pp. 3337–3340, mag. 2002, doi: 10.1021/jf0112973. [CrossRef]
- T. Clavel e J. O. Mapesa, «Phenolics in Human Nutrition: Importance of the Intestinal Microbiome for Isoflavone and Lignan Bioavailability», in Natural Products, K. G. Ramawat e J.-M. Mérillon, A c. di, Berlin, Heidelberg: Springer Berlin Heidelberg, 2013, pp. 2433–2463. doi: 10.1007/978-3-642-22144-6_94. [CrossRef]
- M. T. Viggiani, L. Polimeno, A. Di Leo, e M. Barone, «Phytoestrogens: Dietary Intake, Bioavailability, and Protective Mechanisms against Colorectal Neoproliferative Lesions», Nutrients, vol. 11, fasc. 8, p. 1709, lug. 2019, doi: 10.3390/nu11081709. [CrossRef]
- S. V. Luca et al., «Bioactivity of dietary polyphenols: The role of metabolites», Critical Reviews in Food Science and Nutrition, vol. 60, fasc. 4, pp. 626–659, feb. 2020, doi: 10.1080/10408398.2018.1546669. [CrossRef]
- A. Sarfraz et al., «Biochanin A: A novel bioactive multifunctional compound from nature», Sci Total Environ, vol. 722, p. 137907, giu. 2020, doi: 10.1016/j.scitotenv.2020.137907. [CrossRef]
- E. Bowey, H. Adlercreutz, e I. Rowland, «Metabolism of isoflavones and lignans by the gut microflora: a study in germ-free and human flora associated rats», Food Chem Toxicol, vol. 41, fasc. 5, pp. 631–636, mag. 2003, doi: 10.1016/s0278-6915(02)00324-1. [CrossRef]
- A. V. Sirotkin e A. H. Harrath, «Phytoestrogens and their effects», Eur J Pharmacol, vol. 741, pp. 230–236, ott. 2014, doi: 10.1016/j.ejphar.2014.07.057. [CrossRef]
- I. Paterni, C. Granchi, J. A. Katzenellenbogen, e F. Minutolo, «Estrogen receptors alpha (ERα) and beta (ERβ): Subtype-selective ligands and clinical potential», Steroids, vol. 90, pp. 13–29, nov. 2014, doi: 10.1016/j.steroids.2014.06.012. [CrossRef]
- I. M. C. M. Rietjens, J. Louisse, e K. Beekmann, «The potential health effects of dietary phytoestrogens», Br J Pharmacol, vol. 174, fasc. 11, pp. 1263–1280, giu. 2017, doi: 10.1111/bph.13622. [CrossRef]
- V. C. Jordan, «SERMs: meeting the promise of multifunctional medicines», J Natl Cancer Inst, vol. 99, fasc. 5, pp. 350–356, mar. 2007, doi: 10.1093/jnci/djk062. [CrossRef]
- S. Bedell, M. Nachtigall, e F. Naftolin, «The pros and cons of plant estrogens for menopause», J Steroid Biochem Mol Biol, vol. 139, pp. 225–236, gen. 2014, doi: 10.1016/j.jsbmb.2012.12.004. [CrossRef]
- M. G. M. van de Schans, J.-P. Vincken, P. de Waard, A. R. M. Hamers, T. F. H. Bovee, e H. Gruppen, «Glyceollins and dehydroglyceollins isolated from soybean act as SERMs and ER subtype-selective phytoestrogens», J Steroid Biochem Mol Biol, vol. 156, pp. 53–63, feb. 2016, doi: 10.1016/j.jsbmb.2015.11.020. [CrossRef]
- G.-A. Lee, K.-A. Hwang, e K.-C. Choi, «Roles of Dietary Phytoestrogens on the Regulation of Epithelial-Mesenchymal Transition in Diverse Cancer Metastasis», Toxins (Basel), vol. 8, fasc. 6, p. 162, mag. 2016, doi: 10.3390/toxins8060162. [CrossRef]
- M. F. McCarty, «Isoflavones made simple - genistein’s agonist activity for the beta-type estrogen receptor mediates their health benefits», Med Hypotheses, vol. 66, fasc. 6, pp. 1093–1114, 2006, doi: 10.1016/j.mehy.2004.11.046. [CrossRef]
- C. Thomas e J.-Å. Gustafsson, «The different roles of ER subtypes in cancer biology and therapy», Nat Rev Cancer, vol. 11, fasc. 8, pp. 597–608, ago. 2011, doi: 10.1038/nrc3093. [CrossRef]
- P. Jonsson, A. Katchy, e C. Williams, «Support of a bi-faceted role of estrogen receptor β (ERβ) in ERα-positive breast cancer cells», Endocrine-Related Cancer, vol. 21, fasc. 2, pp. 143–160, apr. 2014, doi: 10.1530/ERC-13-0444. [CrossRef]
- B. Huang, M. Warner, e J.-Å. Gustafsson, «Estrogen receptors in breast carcinogenesis and endocrine therapy», Molecular and Cellular Endocrinology, vol. 418, pp. 240–244, dic. 2015, doi: 10.1016/j.mce.2014.11.015. [CrossRef]
- H. van der Woude, M. G. R. Ter Veld, N. Jacobs, P. T. van der Saag, A. J. Murk, e I. M. C. M. Rietjens, «The stimulation of cell proliferation by quercetin is mediated by the estrogen receptor», Mol Nutr Food Res, vol. 49, fasc. 8, pp. 763–771, ago. 2005, doi: 10.1002/mnfr.200500036. [CrossRef]
- J. L. Bowers, V. V. Tyulmenkov, S. C. Jernigan, e C. M. Klinge, «Resveratrol acts as a mixed agonist/antagonist for estrogen receptors alpha and beta», Endocrinology, vol. 141, fasc. 10, pp. 3657–3667, ott. 2000, doi: 10.1210/endo.141.10.7721. [CrossRef]
- S. O. Mueller, S. Simon, K. Chae, M. Metzler, e K. S. Korach, «Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor alpha (ERalpha) and ERbeta in human cells», Toxicol Sci, vol. 80, fasc. 1, pp. 14–25, lug. 2004, doi: 10.1093/toxsci/kfh147. [CrossRef]
- J. An, C. Tzagarakis-Foster, T. C. Scharschmidt, N. Lomri, e D. C. Leitman, «Estrogen receptor beta-selective transcriptional activity and recruitment of coregulators by phytoestrogens», J Biol Chem, vol. 276, fasc. 21, pp. 17808–17814, mag. 2001, doi: 10.1074/jbc.M100953200. [CrossRef]
- D. P. McDonnell, «The molecular determinants of estrogen receptor pharmacology», Maturitas, vol. 48 Suppl 1, pp. S7-12, ago. 2004, doi: 10.1016/j.maturitas.2004.03.006. [CrossRef]
- G. G. Kuiper et al., «Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta», Endocrinology, vol. 138, fasc. 3, pp. 863–870, mar. 1997, doi: 10.1210/endo.138.3.4979. [CrossRef]
- E. L. Robb e J. A. Stuart, «Resveratrol interacts with estrogen receptor-β to inhibit cell replicative growth and enhance stress resistance by upregulating mitochondrial superoxide dismutase», Free Radic Biol Med, vol. 50, fasc. 7, pp. 821–831, apr. 2011, doi: 10.1016/j.freeradbiomed.2010.12.038. [CrossRef]
- D. Surico et al., «Modulation of Oxidative Stress by 17 β-Estradiol and Genistein in Human Hepatic Cell Lines In Vitro», Cell Physiol Biochem, vol. 42, fasc. 3, pp. 1051–1062, 2017, doi: 10.1159/000478752. [CrossRef]
- A. K. Tanwar, N. Dhiman, A. Kumar, e V. Jaitak, «Engagement of phytoestrogens in breast cancer suppression: Structural classification and mechanistic approach», Eur J Med Chem, vol. 213, p. 113037, mar. 2021, doi: 10.1016/j.ejmech.2020.113037. [CrossRef]
- Y. Yamaguchi, «Microenvironmental regulation of estrogen signals in breast cancer», Breast Cancer, vol. 14, fasc. 2, pp. 175–181, 2007, doi: 10.2325/jbcs.975. [CrossRef]
- E. R. Prossnitz e M. Barton, «The G-protein-coupled estrogen receptor GPER in health and disease», Nat Rev Endocrinol, vol. 7, fasc. 12, pp. 715–726, ago. 2011, doi: 10.1038/nrendo.2011.122. [CrossRef]
- R. Viñas, Y.-J. Jeng, e C. S. Watson, «Non-Genomic Effects of Xenoestrogen Mixtures», IJERPH, vol. 9, fasc. 8, pp. 2694–2714, lug. 2012, doi: 10.3390/ijerph9082694. [CrossRef]
- L. Molina, F. A. Bustamante, K. D. Bhoola, C. D. Figueroa, e P. Ehrenfeld, «Possible role of phytoestrogens in breast cancer via GPER-1/GPR30 signaling», Clin Sci (Lond), vol. 132, fasc. 24, pp. 2583–2598, dic. 2018, doi: 10.1042/CS20180885. [CrossRef]
- M. Razandi, A. Pedram, I. Merchenthaler, G. L. Greene, e E. R. Levin, «Plasma membrane estrogen receptors exist and functions as dimers», Mol Endocrinol, vol. 18, fasc. 12, pp. 2854–2865, dic. 2004, doi: 10.1210/me.2004-0115. [CrossRef]
- D. C. Márquez, J. Lee, T. Lin, e R. J. Pietras, «Epidermal growth factor receptor and tyrosine phosphorylation of estrogen receptor», Endocrine, vol. 16, fasc. 2, pp. 73–81, nov. 2001, doi: 10.1385/ENDO:16:2:073. [CrossRef]
- P. Gong, Z. Madak-Erdogan, J. A. Flaws, D. J. Shapiro, J. A. Katzenellenbogen, e B. S. Katzenellenbogen, «Estrogen receptor-α and aryl hydrocarbon receptor involvement in the actions of botanical estrogens in target cells», Mol Cell Endocrinol, vol. 437, pp. 190–200, dic. 2016, doi: 10.1016/j.mce.2016.08.025. [CrossRef]
- Q. Huang e Q. Chen, «Mediating Roles of PPARs in the Effects of Environmental Chemicals on Sex Steroids», PPAR Res, vol. 2017, p. 3203161, 2017, doi: 10.1155/2017/3203161. [CrossRef]
- K. Kumari, A. K. Adhya, A. K. Rath, P. B. Reddy, e S. K. Mishra, «Estrogen-related receptors alpha, beta and gamma expression and function is associated with transcriptional repressor EZH2 in breast carcinoma», BMC Cancer, vol. 18, fasc. 1, p. 690, dic. 2018, doi: 10.1186/s12885-018-4586-0. [CrossRef]
- S. Barnes, «The biochemistry, chemistry and physiology of the isoflavones in soybeans and their food products», Lymphat Res Biol, vol. 8, fasc. 1, pp. 89–98, mar. 2010, doi: 10.1089/lrb.2009.0030. [CrossRef]
- S. Lecomte, F. Demay, F. Ferrière, e F. Pakdel, «Phytochemicals Targeting Estrogen Receptors: Beneficial Rather Than Adverse Effects?», Int J Mol Sci, vol. 18, fasc. 7, p. 1381, giu. 2017, doi: 10.3390/ijms18071381. [CrossRef]
- K. Pandima Devi, T. Rajavel, M. Daglia, S. F. Nabavi, A. Bishayee, e S. M. Nabavi, «Targeting miRNAs by polyphenols: Novel therapeutic strategy for cancer», Semin Cancer Biol, vol. 46, pp. 146–157, ott. 2017, doi: 10.1016/j.semcancer.2017.02.001. [CrossRef]
- C.-J. Hsieh, Y.-L. Hsu, Y.-F. Huang, e E.-M. Tsai, «Molecular Mechanisms of Anticancer Effects of Phytoestrogens in Breast Cancer», Curr Protein Pept Sci, vol. 19, fasc. 3, pp. 323–332, 2018, doi: 10.2174/1389203718666170111121255. [CrossRef]
- G. L. Russo, I. Tedesco, C. Spagnuolo, e M. Russo, «Antioxidant polyphenols in cancer treatment: Friend, foe or foil?», Seminars in Cancer Biology, vol. 46, pp. 1–13, ott. 2017, doi: 10.1016/j.semcancer.2017.05.005. [CrossRef]
- C. Park et al., «Induction of G2/M Cell Cycle Arrest and Apoptosis by Genistein in Human Bladder Cancer T24 Cells through Inhibition of the ROS-Dependent PI3k/Akt Signal Transduction Pathway», Antioxidants (Basel), vol. 8, fasc. 9, p. 327, ago. 2019, doi: 10.3390/antiox8090327. [CrossRef]
- Y.-C. Hsiao et al., «Genistein induces apoptosis in vitro and has antitumor activity against human leukemia HL-60 cancer cell xenograft growth in vivo», Environmental Toxicology, vol. 34, fasc. 4, pp. 443–456, apr. 2019, doi: 10.1002/tox.22698. [CrossRef]
- S. Rodríguez-Enríquez et al., «Resveratrol inhibits cancer cell proliferation by impairing oxidative phosphorylation and inducing oxidative stress», Toxicol Appl Pharmacol, vol. 370, pp. 65–77, mag. 2019, doi: 10.1016/j.taap.2019.03.008. [CrossRef]
- A. Scalbert, C. Manach, C. Morand, C. Rémésy, e L. Jiménez, «Dietary Polyphenols and the Prevention of Diseases», Critical Reviews in Food Science and Nutrition, vol. 45, fasc. 4, pp. 287–306, giu. 2005, doi: 10.1080/1040869059096. [CrossRef]
- F. Virgili e M. Marino, «Regulation of cellular signals from nutritional molecules: a specific role for phytochemicals, beyond antioxidant activity», Free Radical Biology and Medicine, vol. 45, fasc. 9, pp. 1205–1216, nov. 2008, doi: 10.1016/j.freeradbiomed.2008.08.001. [CrossRef]
- M. Asensi, A. Ortega, S. Mena, F. Feddi, e J. M. Estrela, «Natural polyphenols in cancer therapy», Critical Reviews in Clinical Laboratory Sciences, vol. 48, fasc. 5–6, pp. 197–216, dic. 2011, doi: 10.3109/10408363.2011.631268. [CrossRef]
- J. W. Martin, J. A. Squire, e M. Zielenska, «The genetics of osteosarcoma», Sarcoma, vol. 2012, p. 627254, 2012, doi: 10.1155/2012/627254. [CrossRef]
- J. W. V. de Azevedo et al., «Biology and pathogenesis of human osteosarcoma», Oncol Lett, vol. 19, fasc. 2, pp. 1099–1116, feb. 2020, doi: 10.3892/ol.2019.11229. [CrossRef]
- A. M. Czarnecka et al., «Molecular Biology of Osteosarcoma», Cancers (Basel), vol. 12, fasc. 8, p. 2130, lug. 2020, doi: 10.3390/cancers12082130. [CrossRef]
- M. Hameed e D. Mandelker, «Tumor Syndromes Predisposing to Osteosarcoma», Adv Anat Pathol, vol. 25, fasc. 4, pp. 217–222, lug. 2018, doi: 10.1097/PAP.0000000000000190. [CrossRef]
- X. Chen et al., «Recurrent Somatic Structural Variations Contribute to Tumorigenesis in Pediatric Osteosarcoma», Cell Reports, vol. 7, fasc. 1, pp. 104–112, apr. 2014, doi: 10.1016/j.celrep.2014.03.003. [CrossRef]
- L. L. Wang et al., «Association between osteosarcoma and deleterious mutations in the RECQL4 gene in Rothmund-Thomson syndrome», J Natl Cancer Inst, vol. 95, fasc. 9, pp. 669–674, mag. 2003, doi: 10.1093/jnci/95.9.669. [CrossRef]
- R. A. Kleinerman, S. J. Schonfeld, e M. A. Tucker, «Sarcomas in hereditary retinoblastoma», Clin Sarcoma Res, vol. 2, fasc. 1, p. 15, dic. 2012, doi: 10.1186/2045-3329-2-15. [CrossRef]
- K. Rickel, F. Fang, e J. Tao, «Molecular genetics of osteosarcoma», Bone, vol. 102, pp. 69–79, set. 2017, doi: 10.1016/j.bone.2016.10.017. [CrossRef]
- M. Fiedorowicz, E. Bartnik, P. Sobczuk, P. Teterycz, e A. M. Czarnecka, «Molecular biology of sarcoma», Oncol Clin Pract, vol. 14, fasc. 6, pp. 307–330, mar. 2019, doi: 10.5603/OCP.2018.0045. [CrossRef]
- D. Chen et al., «Super enhancer inhibitors suppress MYC driven transcriptional amplification and tumor progression in osteosarcoma», Bone Res, vol. 6, fasc. 1, p. 11, apr. 2018, doi: 10.1038/s41413-018-0009-8. [CrossRef]
- L. C. Kim, L. Song, e E. B. Haura, «Src kinases as therapeutic targets for cancer», Nat Rev Clin Oncol, vol. 6, fasc. 10, pp. 587–595, ott. 2009, doi: 10.1038/nrclinonc.2009.129. [CrossRef]
- P. Hingorani, W. Zhang, R. Gorlick, e E. A. Kolb, «Inhibition of Src Phosphorylation Alters Metastatic Potential of Osteosarcoma In vitro but not In vivo», Clinical Cancer Research, vol. 15, fasc. 10, pp. 3416–3422, mag. 2009, doi: 10.1158/1078-0432.CCR-08-1657. [CrossRef]
- M. L. Broadhead, J. C. M. Clark, D. E. Myers, C. R. Dass, e P. F. M. Choong, «The molecular pathogenesis of osteosarcoma: a review», Sarcoma, vol. 2011, p. 959248, 2011, doi: 10.1155/2011/959248. [CrossRef]
- S. Gagiannis et al., «Parathyroid hormone-related protein confers chemoresistance by blocking apoptosis signaling via death receptors and mitochondria», Int. J. Cancer, vol. 125, fasc. 7, pp. 1551–1557, ott. 2009, doi: 10.1002/ijc.24471. [CrossRef]
- S. Sharma, T. K. Kelly, e P. A. Jones, «Epigenetics in cancer», Carcinogenesis, vol. 31, fasc. 1, pp. 27–36, gen. 2010, doi: 10.1093/carcin/bgp220. [CrossRef]
- K. B. Jones et al., «miRNA Signatures Associate with Pathogenesis and Progression of Osteosarcoma», Cancer Research, vol. 72, fasc. 7, pp. 1865–1877, apr. 2012, doi: 10.1158/0008-5472.CAN-11-2663. [CrossRef]
- G. B. Andersen, A. Knudsen, H. Hager, L. L. Hansen, e J. Tost, «miRNA profiling identifies deregulated miRNAs associated with osteosarcoma development and time to metastasis in two large cohorts», Mol Oncol, vol. 12, fasc. 1, pp. 114–131, gen. 2018, doi: 10.1002/1878-0261.12154. [CrossRef]
- S. Zhou et al., «miR-421 is a diagnostic and prognostic marker in patients with osteosarcoma», Tumour Biol, vol. 37, fasc. 7, pp. 9001–9007, lug. 2016, doi: 10.1007/s13277-015-4578-5. [CrossRef]
- Z. Ren et al., «MiR-421 promotes the development of osteosarcoma by regulating MCPIP1 expression», Cancer Biol Ther, vol. 21, fasc. 3, pp. 231–240, 2020, doi: 10.1080/15384047.2019.1683331. [CrossRef]
- J. PosthumaDeBoer, M. A. Witlox, G. J. L. Kaspers, e B. J. van Royen, «Molecular alterations as target for therapy in metastatic osteosarcoma: a review of literature», Clin Exp Metastasis, vol. 28, fasc. 5, pp. 493–503, giu. 2011, doi: 10.1007/s10585-011-9384-x. [CrossRef]
- M. Felx et al., «Endothelin-1 (ET-1) promotes MMP-2 and MMP-9 induction involving the transcription factor NF-kappaB in human osteosarcoma», Clin Sci (Lond), vol. 110, fasc. 6, pp. 645–654, giu. 2006, doi: 10.1042/CS20050286. [CrossRef]
- J. Cui, D. Dean, F. J. Hornicek, Z. Chen, e Z. Duan, «The role of extracelluar matrix in osteosarcoma progression and metastasis», J Exp Clin Cancer Res, vol. 39, fasc. 1, p. 178, dic. 2020, doi: 10.1186/s13046-020-01685-w. [CrossRef]
- C. Chen, M. Zhao, A. Tian, X. Zhang, Z. Yao, e X. Ma, «Aberrant activation of Wnt/β-catenin signaling drives proliferation of bone sarcoma cells», Oncotarget, vol. 6, fasc. 19, pp. 17570–17583, lug. 2015, doi: 10.18632/oncotarget.4100. [CrossRef]
- F. Fang et al., «Targeting the Wnt/β-catenin pathway in human osteosarcoma cells», Oncotarget, vol. 9, fasc. 95, pp. 36780–36792, dic. 2018, doi: 10.18632/oncotarget.26377. [CrossRef]
- L. L. Worth, E. A. Lafleur, S.-F. Jia, e E. S. Kleinerman, «Fas expression inversely correlates with metastatic potential in osteosarcoma cells», Oncol Rep, vol. 9, fasc. 4, pp. 823–827, 2002. [CrossRef]
- E. A. Lafleur et al., «Increased Fas expression reduces the metastatic potential of human osteosarcoma cells», Clin Cancer Res, vol. 10, fasc. 23, pp. 8114–8119, dic. 2004, doi: 10.1158/1078-0432.CCR-04-0353. [CrossRef]
- G. Alloisio et al., «Effects of Extracellular Osteoanabolic Agents on the Endogenous Response of Osteoblastic Cells», Cells, vol. 10, fasc. 9, p. 2383, set. 2021, doi: 10.3390/cells10092383. [CrossRef]
- B. Navet et al., «The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases», Cancers (Basel), vol. 10, fasc. 11, p. 398, ott. 2018, doi: 10.3390/cancers10110398. [CrossRef]
- K. S. Nørregaard, H. J. Jürgensen, H. Gårdsvoll, L. H. Engelholm, N. Behrendt, e K. Søe, «Osteosarcoma and Metastasis Associated Bone Degradation—A Tale of Osteoclast and Malignant Cell Cooperativity», IJMS, vol. 22, fasc. 13, p. 6865, giu. 2021, doi: 10.3390/ijms22136865. [CrossRef]
- E. Grimaud et al., «Receptor Activator of Nuclear Factor κB Ligand (RANKL)/Osteoprotegerin (OPG) Ratio Is Increased in Severe Osteolysis», The American Journal of Pathology, vol. 163, fasc. 5, pp. 2021–2031, nov. 2003, doi: 10.1016/S0002-9440(10)63560-2. [CrossRef]
- Y. Chen et al., «RANKL blockade prevents and treats aggressive osteosarcomas», Sci Transl Med, vol. 7, fasc. 317, p. 317ra197, dic. 2015, doi: 10.1126/scitranslmed.aad0295. [CrossRef]
- E. R. Wagner et al., «Defective osteogenic differentiation in the development of osteosarcoma», Sarcoma, vol. 2011, p. 325238, 2011, doi: 10.1155/2011/325238. [CrossRef]
- E. R. Wagner et al., «Therapeutic Implications of PPARgamma in Human Osteosarcoma», PPAR Res, vol. 2010, p. 956427, 2010, doi: 10.1155/2010/956427. [CrossRef]
- L. Carpio, J. Gladu, D. Goltzman, e S. A. Rabbani, «Induction of osteoblast differentiation indexes by PTHrP in MG-63 cells involves multiple signaling pathways», Am J Physiol Endocrinol Metab, vol. 281, fasc. 3, pp. E489-499, set. 2001, doi: 10.1152/ajpendo.2001.281.3.E489. [CrossRef]
- A. Kallio et al., «Estrogen and the selective estrogen receptor modulator (SERM) protection against cell death in estrogen receptor alpha and beta expressing U2OS cells», Mol Cell Endocrinol, vol. 289, fasc. 1–2, pp. 38–48, lug. 2008, doi: 10.1016/j.mce.2008.03.005. [CrossRef]
- S. J. Cotterill, C. M. Wright, M. S. Pearce, A. W. Craft, e UKCCSG/MRC Bone Tumour Working Group, «Stature of young people with malignant bone tumors», Pediatr Blood Cancer, vol. 42, fasc. 1, pp. 59–63, gen. 2004, doi: 10.1002/pbc.10437. [CrossRef]
- S.-M. Wu, L.-H. Shih, J.-Y. Lee, Y.-J. Shen, e H.-H. Lee, «Estrogen enhances activity of Wnt signaling during osteogenesis by inducing Fhl1 expression», J Cell Biochem, vol. 116, fasc. 7, pp. 1419–1430, lug. 2015, doi: 10.1002/jcb.25102. [CrossRef]
- A. B. Khalid e S. A. Krum, «Estrogen receptors alpha and beta in bone», Bone, vol. 87, pp. 130–135, giu. 2016, doi: 10.1016/j.bone.2016.03.016. [CrossRef]
- P. V. Bodine et al., «Estrogen receptor-alpha is developmentally regulated during osteoblast differentiation and contributes to selective responsiveness of gene expression», Endocrinology, vol. 139, fasc. 4, pp. 2048–2057, apr. 1998, doi: 10.1210/endo.139.4.5897. [CrossRef]
- T. C. Spelsberg, M. Subramaniam, B. L. Riggs, e S. Khosla, «The Actions and Interactions of Sex Steroids and Growth Factors/Cytokines on the Skeleton», Molecular Endocrinology, vol. 13, fasc. 6, pp. 819–828, giu. 1999, doi: 10.1210/mend.13.6.0299. [CrossRef]
- D. G. Monroe, B. J. Getz, S. A. Johnsen, B. L. Riggs, S. Khosla, e T. C. Spelsberg, «Estrogen receptor isoform-specific regulation of endogenous gene expression in human osteoblastic cell lines expressing either ERalpha or ERbeta», J Cell Biochem, vol. 90, fasc. 2, pp. 315–326, ott. 2003, doi: 10.1002/jcb.10633. [CrossRef]
- M. K. Tee et al., «Estradiol and selective estrogen receptor modulators differentially regulate target genes with estrogen receptors alpha and beta», Mol Biol Cell, vol. 15, fasc. 3, pp. 1262–1272, mar. 2004, doi: 10.1091/mbc.e03-06-0360. [CrossRef]
- R. D. Roberts, «Is Estrogen the Answer for Osteosarcoma?», Cancer Research, vol. 79, fasc. 6, pp. 1034–1035, mar. 2019, doi: 10.1158/0008-5472.CAN-19-0209. [CrossRef]
- M. A. Lillo Osuna et al., «Activation of Estrogen Receptor Alpha by Decitabine Inhibits Osteosarcoma Growth and Metastasis», Cancer Res, vol. 79, fasc. 6, pp. 1054–1068, mar. 2019, doi: 10.1158/0008-5472.CAN-18-1255. [CrossRef]
- F. Stossi, D. H. Barnett, J. Frasor, B. Komm, C. R. Lyttle, e B. S. Katzenellenbogen, «Transcriptional Profiling of Estrogen-Regulated Gene Expression via Estrogen Receptor (ER) α or ERβ in Human Osteosarcoma Cells: Distinct and Common Target Genes for These Receptors», Endocrinology, vol. 145, fasc. 7, pp. 3473–3486, lug. 2004, doi: 10.1210/en.2003-1682. [CrossRef]
- O. Dohi et al., «Sex steroid receptors expression and hormone-induced cell proliferation in human osteosarcoma», Cancer Sci, vol. 99, fasc. 3, pp. 518–523, mar. 2008, doi: 10.1111/j.1349-7006.2007.00673.x. [CrossRef]
- B. J. Deroo e K. S. Korach, «Estrogen receptors and human disease», J Clin Invest, vol. 116, fasc. 3, pp. 561–570, mar. 2006, doi: 10.1172/JCI27987. [CrossRef]
- K. L. Auld et al., «Estrogen-related receptor α regulates osteoblast differentiation via Wnt/β-catenin signaling», J Mol Endocrinol, vol. 48, fasc. 2, pp. 177–191, apr. 2012, doi: 10.1530/JME-11-0140. [CrossRef]
- P.-I. Lin, Y.-T. Tai, W. P. Chan, Y.-L. Lin, M.-H. Liao, e R.-M. Chen, «Estrogen/ERα signaling axis participates in osteoblast maturation via upregulating chromosomal and mitochondrial complex gene expressions», Oncotarget, vol. 9, fasc. 1, pp. 1169–1186, gen. 2018, doi: 10.18632/oncotarget.23453. [CrossRef]
- J.-Y. Wang, C.-M. Chen, C.-F. Chen, P.-K. Wu, e W.-M. Chen, «Suppression of Estrogen Receptor Alpha Inhibits Cell Proliferation, Differentiation and Enhances the Chemosensitivity of P53-Positive U2OS Osteosarcoma Cell», IJMS, vol. 22, fasc. 20, p. 11238, ott. 2021, doi: 10.3390/ijms222011238. [CrossRef]
- Z.-X. Ouyang e X.-A. Li, «Inhibitory effects of tamoxifen and doxorubicin, alone and in combination, on the proliferation of the MG63 human osteosarcoma cell line», Oncol Lett, vol. 6, fasc. 4, pp. 970–976, ott. 2013, doi: 10.3892/ol.2013.1487. [CrossRef]
- T. Quist, H. Jin, J.-F. Zhu, K. Smith-Fry, M. R. Capecchi, e K. B. Jones, «The impact of osteoblastic differentiation on osteosarcomagenesis in the mouse», Oncogene, vol. 34, fasc. 32, pp. 4278–4284, ago. 2015, doi: 10.1038/onc.2014.354. [CrossRef]
- G. Lazennec, «Estrogen receptor beta, a possible tumor suppressor involved in ovarian carcinogenesis», Cancer Lett, vol. 231, fasc. 2, pp. 151–157, gen. 2006, doi: 10.1016/j.canlet.2005.01.021. [CrossRef]
- M. Gorska, R. M. Wyszkowska, A. Kuban-Jankowska, e M. Wozniak, «Impact of Apparent Antagonism of Estrogen Receptor β by Fulvestrant on Anticancer Activity of 2-Methoxyestradiol», Anticancer Res, vol. 36, fasc. 5, pp. 2217–2226, mag. 2016.
- M. Yang, B. Liu, L. Jin, H. Tao, e Z. Yang, «Estrogen receptor β exhibited anti-tumor effects on osteosarcoma cells by regulating integrin, IAP, NF-kB/BCL-2 and PI3K/Akt signal pathway», J Bone Oncol, vol. 9, pp. 15–20, nov. 2017, doi: 10.1016/j.jbo.2017.09.005. [CrossRef]
- Y. Zhang, C. Yin, X. Zhou, Y. Wu, e L. Wang, «Silencing of estrogen receptor β promotes the invasion and migration of osteosarcoma cells through activating Wnt signaling pathway», OTT, vol. Volume 12, pp. 6779–6788, ago. 2019, doi: 10.2147/OTT.S219222. [CrossRef]
- Z. Yang, W. Yu, B. Liu, M. Yang, e H. Tao, «Estrogen receptor β induces autophagy of osteosarcoma through the mTOR signaling pathway», J Orthop Surg Res, vol. 15, fasc. 1, p. 50, feb. 2020, doi: 10.1186/s13018-020-1575-1. [CrossRef]
- K. Polkowski e A. P. Mazurek, «Biological properties of genistein. A review of in vitro and in vivo data», Acta Pol Pharm, vol. 57, fasc. 2, pp. 135–155, 2000.
- H. S. Tuli et al., «Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances», Front. Pharmacol., vol. 10, p. 1336, dic. 2019, doi: 10.3389/fphar.2019.01336. [CrossRef]
- K. L. Chan, M. K. Y. Siu, Y.-X. Jiang, J.-J. Wang, T. H. Y. Leung, e H. Y. S. Ngan, «Estrogen receptor modulators genistein, daidzein and ERB-041 inhibit cell migration, invasion, proliferation and sphere formation via modulation of FAK and PI3K/AKT signaling in ovarian cancer», Cancer Cell Int, vol. 18, p. 65, 2018, doi: 10.1186/s12935-018-0559-2. [CrossRef]
- Cassidy Aedin, «Bioavailability of isoflavones in humans. In Flavonoids and Related Compounds: Bioavail-ability and Function», CRC Press: Boca Raton, 2012.
- K. Polkowski et al., «Cytostatic and cytotoxic activity of synthetic genistein glycosides against human cancer cell lines», Cancer Lett, vol. 203, fasc. 1, pp. 59–69, gen. 2004, doi: 10.1016/j.canlet.2003.08.023. [CrossRef]
- C. Spagnuolo et al., «Genistein and cancer: current status, challenges, and future directions», Adv Nutr, vol. 6, fasc. 4, pp. 408–419, lug. 2015, doi: 10.3945/an.114.008052. [CrossRef]
- L. Křížová, K. Dadáková, J. Kašparovská, e T. Kašparovský, «Isoflavones», Molecules, vol. 24, fasc. 6, p. 1076, mar. 2019, doi: 10.3390/molecules24061076. [CrossRef]
- T. Hertrampf, M. J. Gruca, J. Seibel, U. Laudenbach, K. H. Fritzemeier, e P. Diel, «The bone-protective effect of the phytoestrogen genistein is mediated via ER alpha-dependent mechanisms and strongly enhanced by physical activity», Bone, vol. 40, fasc. 6, pp. 1529–1535, giu. 2007, doi: 10.1016/j.bone.2007.02.006. [CrossRef]
- M. S. Kurzer, «Hormonal effects of soy in premenopausal women and men», J Nutr, vol. 132, fasc. 3, pp. 570S-573S, mar. 2002, doi: 10.1093/jn/132.3.570S. [CrossRef]
- F. H. Sarkar e Y. Li, «Mechanisms of cancer chemoprevention by soy isoflavone genistein», Cancer Metastasis Rev, vol. 21, fasc. 3–4, pp. 265–280, 2002, doi: 10.1023/a:1021210910821. [CrossRef]
- H. Jiang, J. Fan, L. Cheng, P. Hu, e R. Liu, «The anticancer activity of genistein is increased in estrogen receptor beta 1-positive breast cancer cells», OTT, vol. Volume 11, pp. 8153–8163, nov. 2018, doi: 10.2147/OTT.S182239. [CrossRef]
- A. Ranjan et al., «Role of Phytochemicals in Cancer Prevention», IJMS, vol. 20, fasc. 20, p. 4981, ott. 2019, doi: 10.3390/ijms20204981. [CrossRef]
- D. J. Rickard, D. G. Monroe, T. J. Ruesink, S. Khosla, B. L. Riggs, e T. C. Spelsberg, «Phytoestrogen genistein acts as an estrogen agonist on human osteoblastic cells through estrogen receptors alpha and beta», J Cell Biochem, vol. 89, fasc. 3, pp. 633–646, giu. 2003, doi: 10.1002/jcb.10539. [CrossRef]
- G. Lambrinidis, M. Halabalaki, E. S. Katsanou, A.-L. Skaltsounis, M. N. Alexis, e E. Mikros, «The estrogen receptor and polyphenols: molecular simulation studies of their interactions, a review», Environ Chem Lett, vol. 4, fasc. 3, pp. 159–174, ago. 2006, doi: 10.1007/s10311-006-0065-y. [CrossRef]
- S. Banerjee, Y. Li, Z. Wang, e F. H. Sarkar, «Multi-targeted therapy of cancer by genistein», Cancer Lett, vol. 269, fasc. 2, pp. 226–242, ott. 2008, doi: 10.1016/j.canlet.2008.03.052. [CrossRef]
- S. Nakashima, T. Koike, e Y. Nozawa, «Genistein, a protein tyrosine kinase inhibitor, inhibits thromboxane A2-mediated human platelet responses», Mol Pharmacol, vol. 39, fasc. 4, pp. 475–480, apr. 1991.
- Z. M. Shao, M. L. Alpaugh, J. A. Fontana, e S. H. Barsky, «Genistein inhibits proliferation similarly in estrogen receptor-positive and negative human breast carcinoma cell lines characterized by P21WAF1/CIP1 induction, G2/M arrest, and apoptosis», J Cell Biochem, vol. 69, fasc. 1, pp. 44–54, apr. 1998.
- S. Djiogue et al., «Estrogenic properties of naturally occurring prenylated isoflavones in U2OS human osteosarcoma cells: Structure-activity relationships», J Steroid Biochem Mol Biol, vol. 120, fasc. 4–5, pp. 184–191, giu. 2010, doi: 10.1016/j.jsbmb.2010.04.014. [CrossRef]
- L. Salvatori et al., «Down-regulation of epidermal growth factor receptor induced by estrogens and phytoestrogens promotes the differentiation of U2OS human osteosarcoma cells», J. Cell. Physiol., vol. 220, fasc. 1, pp. 35–44, lug. 2009, doi: 10.1002/jcp.21724. [CrossRef]
- X. W. Chen, S. C. Garner, e J. J. B. Anderson, «Isoflavones regulate interleukin-6 and osteoprotegerin synthesis during osteoblast cell differentiation via an estrogen-receptor-dependent pathway», Biochem Biophys Res Commun, vol. 295, fasc. 2, pp. 417–422, lug. 2002, doi: 10.1016/s0006-291x(02)00667-8. [CrossRef]
- A. De Wilde, M. Lieberherr, C. Colin, e A. Pointillart, «A low dose of daidzein acts as an ERbeta-selective agonist in trabecular osteoblasts of young female piglets», J Cell Physiol, vol. 200, fasc. 2, pp. 253–262, ago. 2004, doi: 10.1002/jcp.20008. [CrossRef]
- T.-L. Jia, H.-Z. Wang, L.-P. Xie, X.-Y. Wang, e R.-Q. Zhang, «Daidzein enhances osteoblast growth that may be mediated by increased bone morphogenetic protein (BMP) production», Biochem Pharmacol, vol. 65, fasc. 5, pp. 709–715, mar. 2003, doi: 10.1016/s0006-2952(02)01585-x. [CrossRef]
- C. Morris, J. Thorpe, L. Ambrosio, e M. Santin, «The soybean isoflavone genistein induces differentiation of MG63 human osteosarcoma osteoblasts», J Nutr, vol. 136, fasc. 5, pp. 1166–1170, mag. 2006, doi: 10.1093/jn/136.5.1166. [CrossRef]
- A. Nakamura et al., «Genistein inhibits cell invasion and motility by inducing cell differentiation in murine osteosarcoma cell line LM8», BMC Cell Biol, vol. 13, fasc. 1, p. 24, dic. 2012, doi: 10.1186/1471-2121-13-24. [CrossRef]
- J. Wei, M. Hu, K. Huang, S. Lin, e H. Du, «Roles of Proteoglycans and Glycosaminoglycans in Cancer Development and Progression», Int J Mol Sci, vol. 21, fasc. 17, p. 5983, ago. 2020, doi: 10.3390/ijms21175983. [CrossRef]
- M. A. Birch e T. M. Skerry, «Differential regulation of syndecan expression by osteosarcoma cell lines in response to cytokines but not osteotropic hormones», Bone, vol. 24, fasc. 6, pp. 571–578, giu. 1999, doi: 10.1016/s8756-3282(99)00088-5. [CrossRef]
- D. Nikitovic, A. M. Tsatsakis, N. K. Karamanos, e G. N. Tzanakakis, «The effects of genistein on the synthesis and distribution of glycosaminoglycans/proteoglycans by two osteosarcoma cell lines depends on tyrosine kinase and the estrogen receptor density», Anticancer Res, vol. 23, fasc. 1A, pp. 459–464, 2003.
- Z.-C. Dang, V. Audinot, S. E. Papapoulos, J. A. Boutin, e C. W. G. M. Löwik, «Peroxisome Proliferator-activated Receptor γ (PPARγ) as a Molecular Target for the Soy Phytoestrogen Genistein», Journal of Biological Chemistry, vol. 278, fasc. 2, pp. 962–967, gen. 2003, doi: 10.1074/jbc.M209483200. [CrossRef]
- M. Song et al., «Genistein exerts growth inhibition on human osteosarcoma MG-63 cells via PPARγ pathway», Int J Oncol, vol. 46, fasc. 3, pp. 1131–1140, mar. 2015, doi: 10.3892/ijo.2015.2829. [CrossRef]
- N. Engel et al., «Synergistic Action of Genistein and Calcitriol in Immature Osteosarcoma MG-63 Cells by SGPL1 Up-Regulation», PLoS One, vol. 12, fasc. 1, p. e0169742, 2017, doi: 10.1371/journal.pone.0169742. [CrossRef]
- W. B. Grant e C. F. Garland, «Vitamin D has a greater impact on cancer mortality rates than on cancer incidence rates», BMJ, vol. 348, p. g2862, apr. 2014, doi: 10.1136/bmj.g2862. [CrossRef]
- J. Lappe et al., «Effect of a combination of genistein, polyunsaturated fatty acids and vitamins D3 and K1 on bone mineral density in postmenopausal women: a randomized, placebo-controlled, double-blind pilot study», Eur J Nutr, vol. 52, fasc. 1, pp. 203–215, feb. 2013, doi: 10.1007/s00394-012-0304-x. [CrossRef]
- E. Degagné et al., «Sphingosine-1-phosphate lyase downregulation promotes colon carcinogenesis through STAT3-activated microRNAs», J Clin Invest, vol. 124, fasc. 12, pp. 5368–5384, dic. 2014, doi: 10.1172/JCI74188. [CrossRef]
- T. Ando, J. Ichikawa, A. Okamoto, K. Tasaka, A. Nakao, e Y. Hamada, «Gemcitabine inhibits viability, growth, and metastasis of osteosarcoma cell lines», J Orthop Res, vol. 23, fasc. 4, pp. 964–969, lug. 2005, doi: 10.1016/j.orthres.2005.01.010. [CrossRef]
- B. Zhang, Z.-L. Shi, B. Liu, X.-B. Yan, J. Feng, e H.-M. Tao, «Enhanced anticancer effect of gemcitabine by genistein in osteosarcoma: the role of Akt and nuclear factor-kappaB», Anticancer Drugs, vol. 21, fasc. 3, pp. 288–296, mar. 2010, doi: 10.1097/CAD.0b013e328334da17. [CrossRef]
- C. Nakanishi e M. Toi, «Nuclear factor-kappaB inhibitors as sensitizers to anticancer drugs», Nat Rev Cancer, vol. 5, fasc. 4, pp. 297–309, apr. 2005, doi: 10.1038/nrc1588. [CrossRef]
- C. Liang et al., «Genistein potentiates the anti-cancer effects of gemcitabine in human osteosarcoma via the downregulation of Akt and nuclear factor-κB pathway», Anticancer Agents Med Chem, vol. 12, fasc. 5, pp. 554–563, giu. 2012, doi: 10.2174/187152012800617867. [CrossRef]
- M. M. Alshehri et al., «Therapeutic Potential of Isoflavones with an Emphasis on Daidzein», Oxid Med Cell Longev, vol. 2021, p. 6331630, 2021, doi: 10.1155/2021/6331630. [CrossRef]
- P. J. Magee, P. Allsopp, A. Samaletdin, e I. R. Rowland, «Daidzein, R-(+)equol and S-(-)equol inhibit the invasion of MDA-MB-231 breast cancer cells potentially via the down-regulation of matrix metalloproteinase-2», Eur J Nutr, vol. 53, fasc. 1, pp. 345–350, feb. 2014, doi: 10.1007/s00394-013-0520-z. [CrossRef]
- V. Singh-Gupta et al., «Daidzein Effect on Hormone Refractory Prostate Cancer In Vitro and In Vivo Compared to Genistein and Soy Extract: Potentiation of Radiotherapy», Pharm Res, vol. 27, fasc. 6, pp. 1115–1127, giu. 2010, doi: 10.1007/s11095-010-0107-9. [CrossRef]
- Y.-S. Liang, W.-T. Qi, W. Guo, C.-L. Wang, Z.-B. Hu, e A.-K. Li, «Genistein and daidzein induce apoptosis of colon cancer cells by inhibiting the accumulation of lipid droplets», Food & Nutrition Research, vol. 62, fasc. 0, mag. 2018, doi: 10.29219/fnr.v62.1384. [CrossRef]
- Y. Zhu et al., «Investigation of inhibition effect of daidzein on osteosarcoma cells based on experimental validation and systematic pharmacology analysis», PeerJ, vol. 9, p. e12072, 2021, doi: 10.7717/peerj.12072. [CrossRef]
- G. Renda et al., «Comparative assessment of dermal wound healing potentials of various Trifolium L. extracts and determination of their isoflavone contents as potential active ingredients», J Ethnopharmacol, vol. 148, fasc. 2, pp. 423–432, lug. 2013, doi: 10.1016/j.jep.2013.04.031. [CrossRef]
- C. Yu, P. Zhang, L. Lou, e Y. Wang, «Perspectives Regarding the Role of Biochanin A in Humans», Front. Pharmacol., vol. 10, p. 793, lug. 2019, doi: 10.3389/fphar.2019.00793. [CrossRef]
- Z.-J. Feng e W.-F. Lai, «Chemical and Biological Properties of Biochanin A and Its Pharmaceutical Applications», Pharmaceutics, vol. 15, fasc. 4, p. 1105, mar. 2023, doi: 10.3390/pharmaceutics15041105. [CrossRef]
- W.-Y. Wu et al., «Biochanin A attenuates LPS-induced pro-inflammatory responses and inhibits the activation of the MAPK pathway in BV2 microglial cells», Int J Mol Med, vol. 35, fasc. 2, pp. 391–398, feb. 2015, doi: 10.3892/ijmm.2014.2020. [CrossRef]
- A. M. Jalaludeen et al., «Biochanin A Ameliorates Arsenic-Induced Hepato- and Hematotoxicity in Rats», Molecules, vol. 21, fasc. 1, p. 69, gen. 2016, doi: 10.3390/molecules21010069. [CrossRef]
- Y.-N. Hsu et al., «Anti-proliferative activity of biochanin A in human osteosarcoma cells via mitochondrial-involved apoptosis», Food Chem Toxicol, vol. 112, pp. 194–204, feb. 2018, doi: 10.1016/j.fct.2017.12.062. [CrossRef]
- Y. Zhao, L. Wang, X. Zhai, T. Cui, G. Wang, e Q. Pang, «The effect of biochanin A on cell growth, apoptosis, and migration in osteosarcoma cells», Pharmazie, vol. 73, fasc. 6, pp. 335–341, giu. 2018, doi: 10.1691/ph.2018.7351. [CrossRef]
- A. Caley e R. Jones, «The principles of cancer treatment by chemotherapy», Surgery (Oxford), vol. 30, fasc. 4, pp. 186–190, apr. 2012, doi: 10.1016/j.mpsur.2012.01.004. [CrossRef]
- M.-H. Lu, M.-F. Fan, e X.-D. Yu, «NSD2 promotes osteosarcoma cell proliferation and metastasis by inhibiting E-cadherin expression», Eur Rev Med Pharmacol Sci, vol. 21, fasc. 5, pp. 928–936, mar. 2017.
- A. Labernadie et al., «A mechanically active heterotypic E-cadherin/N-cadherin adhesion enables fibroblasts to drive cancer cell invasion», Nat Cell Biol, vol. 19, fasc. 3, pp. 224–237, mar. 2017, doi: 10.1038/ncb3478. [CrossRef]
- S. Ong et al., «Focus on Formononetin: Anticancer Potential and Molecular Targets», Cancers, vol. 11, fasc. 5, p. 611, mag. 2019, doi: 10.3390/cancers11050611. [CrossRef]
- J. Machado Dutra, P. J. P. Espitia, e R. Andrade Batista, «Formononetin: Biological effects and uses - A review», Food Chem, vol. 359, p. 129975, ott. 2021, doi: 10.1016/j.foodchem.2021.129975. [CrossRef]
- Y. Liu et al., «The Proapoptotic Effect of Formononetin in Human Osteosarcoma Cells: Involvement of Inactivation of ERK and Akt Pathways», Cell Physiol Biochem, vol. 34, fasc. 3, pp. 637–645, 2014, doi: 10.1159/000363029. [CrossRef]
- W. Hu e Z. Xiao, «Formononetin induces apoptosis of human osteosarcoma cell line U2OS by regulating the expression of Bcl-2, Bax and MiR-375 in vitro and in vivo», Cell Physiol Biochem, vol. 37, fasc. 3, pp. 933–939, 2015, doi: 10.1159/000430220. [CrossRef]
- K. Li, H. Shen, M. Lu, J. Chen, Q. Yin, e P. Li, «Formononetin inhibits osteosarcoma cell proliferation and promotes apoptosis by regulating the miR-214-3p/phosphatase and tensin homolog pathway», Transl Cancer Res, vol. 9, fasc. 8, pp. 4914–4921, ago. 2020, doi: 10.21037/tcr-20-2296. [CrossRef]
- X. Zhao et al., «RNA Sequencing of Osteosarcoma Gene Expression Profile Revealed that miR-214-3p Facilitates Osteosarcoma Cell Proliferation via Targeting Ubiquinol-Cytochrome c Reductase Core Protein 1 (UQCRC1)», Med Sci Monit, vol. 25, pp. 4982–4991, lug. 2019, doi: 10.12659/MSM.917375. [CrossRef]
- W. Hu et al., «Anti-cancer targets of formononetin and molecular mechanisms in osteosarcoma: Findings of bioinformatic and experimental assays», J Cell Mol Med, vol. 23, fasc. 5, pp. 3505–3511, mag. 2019, doi: 10.1111/jcmm.14248. [CrossRef]
- I. Domínguez-López, M. Yago-Aragón, A. Salas-Huetos, A. Tresserra-Rimbau, e S. Hurtado-Barroso, «Effects of Dietary Phytoestrogens on Hormones throughout a Human Lifespan: A Review», Nutrients, vol. 12, fasc. 8, p. 2456, ago. 2020, doi: 10.3390/nu12082456. [CrossRef]
- M. G. Hertog, P. C. Hollman, M. B. Katan, e D. Kromhout, «Intake of potentially anticarcinogenic flavonoids and their determinants in adults in The Netherlands», Nutr Cancer, vol. 20, fasc. 1, pp. 21–29, 1993, doi: 10.1080/01635589309514267. [CrossRef]
- G. D’Andrea, «Quercetin: A flavonol with multifaceted therapeutic applications?», Fitoterapia, vol. 106, pp. 256–271, ott. 2015, doi: 10.1016/j.fitote.2015.09.018. [CrossRef]
- M. Harwood, B. Danielewska-Nikiel, J. F. Borzelleca, G. W. Flamm, G. M. Williams, e T. C. Lines, «A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties», Food Chem Toxicol, vol. 45, fasc. 11, pp. 2179–2205, nov. 2007, doi: 10.1016/j.fct.2007.05.015. [CrossRef]
- X. Wang, D. Ha, R. Yoshitake, Y. S. Chan, D. Sadava, e S. Chen, «Exploring the Biological Activity and Mechanism of Xenoestrogens and Phytoestrogens in Cancers: Emerging Methods and Concepts», Int J Mol Sci, vol. 22, fasc. 16, p. 8798, ago. 2021, doi: 10.3390/ijms22168798. [CrossRef]
- D. Yang, T. Wang, M. Long, e P. Li, «Quercetin: Its Main Pharmacological Activity and Potential Application in Clinical Medicine», Oxid Med Cell Longev, vol. 2020, p. 8825387, 2020, doi: 10.1155/2020/8825387. [CrossRef]
- F. Virgili, F. Acconcia, R. Ambra, A. Rinna, P. Totta, e M. Marino, «Nutritional flavonoids modulate estrogen receptor alpha signaling», IUBMB Life, vol. 56, fasc. 3, pp. 145–151, mar. 2004, doi: 10.1080/15216540410001685083. [CrossRef]
- M. Marino, M. Pellegrini, P. La Rosa, e F. Acconcia, «Susceptibility of estrogen receptor rapid responses to xenoestrogens: Physiological outcomes», Steroids, vol. 77, fasc. 10, pp. 910–917, ago. 2012, doi: 10.1016/j.steroids.2012.02.019. [CrossRef]
- P. Galluzzo et al., «Quercetin-induced apoptotic cascade in cancer cells: antioxidant versus estrogen receptor alpha-dependent mechanisms», Mol Nutr Food Res, vol. 53, fasc. 6, pp. 699–708, giu. 2009, doi: 10.1002/mnfr.200800239. [CrossRef]
- P. Bulzomi e M. Marino, «Environmental endocrine disruptors: does a sex-related susceptibility exist?», Front Biosci (Landmark Ed), vol. 16, fasc. 7, pp. 2478–2498, giu. 2011, doi: 10.2741/3867. [CrossRef]
- K. Berndt, C. Campanile, R. Muff, E. Strehler, W. Born, e B. Fuchs, «Evaluation of quercetin as a potential drug in osteosarcoma treatment», Anticancer Res, vol. 33, fasc. 4, pp. 1297–1306, apr. 2013.
- X. Zhang, Q. Guo, J. Chen, e Z. Chen, «Quercetin Enhances Cisplatin Sensitivity of Human Osteosarcoma Cells by Modulating microRNA-217-KRAS Axis», Mol Cells, vol. 38, fasc. 7, pp. 638–642, lug. 2015, doi: 10.14348/molcells.2015.0037. [CrossRef]
- W. Liang et al., «Quercetin-mediated apoptosis via activation of the mitochondrial-dependent pathway in MG-63 osteosarcoma cells», Mol Med Rep, vol. 4, fasc. 5, pp. 1017–1023, 2011, doi: 10.3892/mmr.2011.533. [CrossRef]
- H. Lan, W. Hong, P. Fan, D. Qian, J. Zhu, e B. Bai, «Quercetin Inhibits Cell Migration and Invasion in Human Osteosarcoma Cells», Cell Physiol Biochem, vol. 43, fasc. 2, pp. 553–567, 2017, doi: 10.1159/000480528. [CrossRef]
- S. Li, Y. Pei, W. Wang, F. Liu, K. Zheng, e X. Zhang, «Quercetin suppresses the proliferation and metastasis of metastatic osteosarcoma cells by inhibiting parathyroid hormone receptor 1», Biomed Pharmacother, vol. 114, p. 108839, giu. 2019, doi: 10.1016/j.biopha.2019.108839. [CrossRef]
- D. K. Suh, E. J. Lee, H. C. Kim, e J. H. Kim, «Induction of G(1)/S phase arrest and apoptosis by quercetin in human osteosarcoma cells», Arch Pharm Res, vol. 33, fasc. 5, pp. 781–785, mag. 2010, doi: 10.1007/s12272-010-0519-4. [CrossRef]
- N. Delepine, G. Delepine, G. Bacci, G. Rosen, e J. C. Desbois, «Influence of methotrexate dose intensity on outcome of patients with high grade osteogenic osteosarcoma. Analysis of the literature», Cancer, vol. 78, fasc. 10, pp. 2127–2135, nov. 1996.
- X. Xie et al., «Quercetin induces apoptosis in the methotrexate-resistant osteosarcoma cell line U2-OS/MTX300 via mitochondrial dysfunction and dephosphorylation of Akt», Oncol Rep, vol. 26, fasc. 3, pp. 687–693, set. 2011, doi: 10.3892/or.2011.1328. [CrossRef]
- J. Yin et al., «[Effect and mechanism of quercetin on proliferation and apoptosis of human osteosarcoma cell U-2OS/MTX300]», Zhongguo Zhong Yao Za Zhi, vol. 37, fasc. 5, pp. 611–614, mar. 2012.
- J. Yuan et al., «Osteoblastic and osteolytic human osteosarcomas can be studied with a new xenograft mouse model producing spontaneous metastases», Cancer Invest, vol. 27, fasc. 4, pp. 435–442, mag. 2009, doi: 10.1080/07357900802491477. [CrossRef]
- K. Husmann et al., «Matrix Metalloproteinase 1 promotes tumor formation and lung metastasis in an intratibial injection osteosarcoma mouse model», Biochim Biophys Acta, vol. 1832, fasc. 2, pp. 347–354, feb. 2013, doi: 10.1016/j.bbadis.2012.11.006. [CrossRef]
- B. Wu et al., «Quercetin induced NUPR1-dependent autophagic cell death by disturbing reactive oxygen species homeostasis in osteosarcoma cells», J. Clin. Biochem. Nutr., vol. 67, fasc. 2, pp. 137–145, 2020, doi: 10.3164/jcbn.19-121. [CrossRef]
- C. W. Yun e S. H. Lee, «The Roles of Autophagy in Cancer», Int J Mol Sci, vol. 19, fasc. 11, p. 3466, nov. 2018, doi: 10.3390/ijms19113466. [CrossRef]
- P. Bhat, J. Kriel, B. Shubha Priya, null Basappa, N. S. Shivananju, e B. Loos, «Modulating autophagy in cancer therapy: Advancements and challenges for cancer cell death sensitization», Biochem Pharmacol, vol. 147, pp. 170–182, gen. 2018, doi: 10.1016/j.bcp.2017.11.021. [CrossRef]
- T. Martin et al., «NUPR1 and its potential role in cancer and pathological conditions (Review)», Int J Oncol, vol. 58, fasc. 5, p. 21, mar. 2021, doi: 10.3892/ijo.2021.5201. [CrossRef]
- D. Hanahan e R. A. Weinberg, «Hallmarks of cancer: the next generation», Cell, vol. 144, fasc. 5, pp. 646–674, mar. 2011, doi: 10.1016/j.cell.2011.02.013. [CrossRef]
- H.-Y. Ren et al., «Prognostic role of hypoxia-inducible factor-1 alpha expression in osteosarcoma: a meta-analysis», OTT, p. 1477, mar. 2016, doi: 10.2147/OTT.S95490. [CrossRef]
- D. Nikitovic et al., «Parathyroid hormone/parathyroid hormone-related peptide regulate osteosarcoma cell functions: Focus on the extracellular matrix (Review)», Oncology Reports, vol. 36, fasc. 4, pp. 1787–1792, ott. 2016, doi: 10.3892/or.2016.4986. [CrossRef]
- N. Oršolić e N. Car, «Quercetin and hyperthermia modulate cisplatin-induced DNA damage in tumor and normal tissues in vivo», Tumour Biol, vol. 35, fasc. 7, pp. 6445–6454, lug. 2014, doi: 10.1007/s13277-014-1843-y. [CrossRef]
- L. Yi, Y. Zongyuan, G. Cheng, Z. Lingyun, Y. GuiLian, e G. Wei, «Quercetin enhances apoptotic effect of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) in ovarian cancer cells through reactive oxygen species (ROS) mediated CCAAT enhancer-binding protein homologous protein (CHOP)-death receptor 5 pathway», Cancer Sci, vol. 105, fasc. 5, pp. 520–527, mag. 2014, doi: 10.1111/cas.12395. [CrossRef]
- G. Scambia et al., «Synergistic antiproliferative activity of quercetin and cisplatin on ovarian cancer cell growth», Anticancer Drugs, vol. 1, fasc. 1, pp. 45–48, ott. 1990, doi: 10.1097/00001813-199010000-00008. [CrossRef]
- J. Guo, Z. Feng, Z. Huang, H. Wang, e W. Lu, «MicroRNA-217 functions as a tumour suppressor gene and correlates with cell resistance to cisplatin in lung cancer», Mol Cells, vol. 37, fasc. 9, pp. 664–671, set. 2014, doi: 10.14348/molcells.2014.0121. [CrossRef]
- E. Mohammadi, F. Alemi, M. Maleki, F. Malakoti, N. Farsad-Akhtar, e B. Yousefi, «Quercetin and Methotrexate in Combination have Anticancer Activity in Osteosarcoma Cells and Repress Oncogenic MicroRNA-223», Drug Res (Stuttg), vol. 72, fasc. 4, pp. 226–233, apr. 2022, doi: 10.1055/a-1709-0658. [CrossRef]
- Y. C. Jung et al., «Anti-inflammatory effects of galangin on lipopolysaccharide-activated macrophages via ERK and NF-κB pathway regulation», Immunopharmacol Immunotoxicol, vol. 36, fasc. 6, pp. 426–432, dic. 2014, doi: 10.3109/08923973.2014.968257. [CrossRef]
- T. P. T. Cushnie e A. J. Lamb, «Assessment of the antibacterial activity of galangin against 4-quinolone resistant strains of Staphylococcus aureus», Phytomedicine, vol. 13, fasc. 3, pp. 187–191, feb. 2006, doi: 10.1016/j.phymed.2004.07.003. [CrossRef]
- J. J. Meyer, A. J. Afolayan, M. B. Taylor, e D. Erasmus, «Antiviral activity of galangin isolated from the aerial parts of Helichrysum aureonitens», J Ethnopharmacol, vol. 56, fasc. 2, pp. 165–169, apr. 1997, doi: 10.1016/s0378-8741(97)01514-6. [CrossRef]
- L. Zhu, Q. Luo, J. Bi, J. Ding, S. Ge, e F. Chen, «Galangin inhibits growth of human head and neck squamous carcinoma cells in vitro and in vivo», Chem Biol Interact, vol. 224, pp. 149–156, dic. 2014, doi: 10.1016/j.cbi.2014.10.027. [CrossRef]
- J. Cao et al., «Galangin inhibits cell invasion by suppressing the epithelial-mesenchymal transition and inducing apoptosis in renal cell carcinoma», Mol Med Rep, vol. 13, fasc. 5, pp. 4238–4244, mag. 2016, doi: 10.3892/mmr.2016.5042. [CrossRef]
- W.-W. Zou e S.-P. Xu, «Galangin inhibits the cell progression and induces cell apoptosis through activating PTEN and Caspase-3 pathways in retinoblastoma», Biomedicine & Pharmacotherapy, vol. 97, pp. 851–863, gen. 2018, doi: 10.1016/j.biopha.2017.09.144. [CrossRef]
- Z. Yang et al., «Galangin suppresses human osteosarcoma cells: An exploration of its underlying mechanism», Oncology Reports, vol. 37, fasc. 1, pp. 435–441, gen. 2017, doi: 10.3892/or.2016.5224. [CrossRef]
- C. Liu, M. Ma, J. Zhang, S. Gui, X. Zhang, e S. Xue, «Galangin inhibits human osteosarcoma cells growth by inducing transforming growth factor-β1-dependent osteogenic differentiation», Biomed Pharmacother, vol. 89, pp. 1415–1421, mag. 2017, doi: 10.1016/j.biopha.2017.03.030. [CrossRef]
- K. Janssens, P. ten Dijke, S. Janssens, e W. Van Hul, «Transforming growth factor-beta1 to the bone», Endocr Rev, vol. 26, fasc. 6, pp. 743–774, ott. 2005, doi: 10.1210/er.2004-0001. [CrossRef]
- D. Patel, S. Shukla, e S. Gupta, «Apigenin and cancer chemoprevention: progress, potential and promise (review)», Int J Oncol, vol. 30, fasc. 1, pp. 233–245, gen. 2007. [CrossRef]
- Cos et al., «Structure−Activity Relationship and Classification of Flavonoids as Inhibitors of Xanthine Oxidase and Superoxide Scavengers», J. Nat. Prod., vol. 61, fasc. 1, pp. 71–76, gen. 1998, doi: 10.1021/np970237h. [CrossRef]
- T.-D. Way, M.-C. Kao, e J.-K. Lin, «Apigenin induces apoptosis through proteasomal degradation of HER2/neu in HER2/neu-overexpressing breast cancer cells via the phosphatidylinositol 3-kinase/Akt-dependent pathway», J Biol Chem, vol. 279, fasc. 6, pp. 4479–4489, feb. 2004, doi: 10.1074/jbc.M305529200. [CrossRef]
- M. E. Gonzalez-Mejia, O. H. Voss, E. J. Murnan, e A. I. Doseff, «Apigenin-induced apoptosis of leukemia cells is mediated by a bimodal and differentially regulated residue-specific phosphorylation of heat-shock protein–27», Cell Death Dis, vol. 1, fasc. 8, pp. e64–e64, ago. 2010, doi: 10.1038/cddis.2010.41. [CrossRef]
- C.-C. Lin et al., «Apigenin induces apoptosis through mitochondrial dysfunction in U-2 OS human osteosarcoma cells and inhibits osteosarcoma xenograft tumor growth in vivo», J Agric Food Chem, vol. 60, fasc. 45, pp. 11395–11402, nov. 2012, doi: 10.1021/jf303446x. [CrossRef]
- X. Liu, L. Li, L. Lv, D. Chen, L. Shen, e Z. Xie, «Apigenin inhibits the proliferation and invasion of osteosarcoma cells by suppressing the Wnt/β-catenin signaling pathway», Oncol Rep, vol. 34, fasc. 2, pp. 1035–1041, ago. 2015, doi: 10.3892/or.2015.4022. [CrossRef]
- X. Du, J. Yang, D. Yang, W. Tian, e Z. Zhu, «The genetic basis for inactivation of Wnt pathway in human osteosarcoma», BMC Cancer, vol. 14, fasc. 1, p. 450, dic. 2014, doi: 10.1186/1471-2407-14-450. [CrossRef]
- F. Zhang, A. Chen, J. Chen, T. Yu, e F. Guo, «SiRNA-mediated silencing of beta-catenin suppresses invasion and chemosensitivity to doxorubicin in MG-63 osteosarcoma cells», Asian Pac J Cancer Prev, vol. 12, fasc. 1, pp. 239–245, 2011.
- B. Salehi et al., «The Therapeutic Potential of Naringenin: A Review of Clinical Trials», Pharmaceuticals, vol. 12, fasc. 1, p. 11, gen. 2019, doi: 10.3390/ph12010011. [CrossRef]
- A. Arafah et al., «Multi-Therapeutic Potential of Naringenin (4’,5,7-Trihydroxyflavonone): Experimental Evidence and Mechanisms», Plants (Basel), vol. 9, fasc. 12, p. 1784, dic. 2020, doi: 10.3390/plants9121784. [CrossRef]
- G.-Y. Liou e P. Storz, «Reactive oxygen species in cancer», Free Radic Res, vol. 44, fasc. 5, pp. 479–496, mag. 2010, doi: 10.3109/10715761003667554. [CrossRef]
- L. Zhang et al., «Citrus aurantium Naringenin Prevents Osteosarcoma Progression and Recurrence in the Patients Who Underwent Osteosarcoma Surgery by Improving Antioxidant Capability», Oxidative Medicine and Cellular Longevity, vol. 2018, pp. 1–16, 2018, doi: 10.1155/2018/8713263. [CrossRef]
- S.-I. Kanno et al., «Inhibitory effects of naringenin on tumor growth in human cancer cell lines and sarcoma S-180-implanted mice», Biol Pharm Bull, vol. 28, fasc. 3, pp. 527–530, mar. 2005, doi: 10.1248/bpb.28.527. [CrossRef]
- P. Totta, F. Acconcia, S. Leone, I. Cardillo, e M. Marino, «Mechanisms of naringenin-induced apoptotic cascade in cancer cells: involvement of estrogen receptor alpha and beta signalling», IUBMB Life, vol. 56, fasc. 8, pp. 491–499, ago. 2004, doi: 10.1080/15216540400010792. [CrossRef]
- C.-W. Lee et al., «Naringenin Induces ROS-Mediated ER Stress, Autophagy, and Apoptosis in Human Osteosarcoma Cell Lines», Molecules, vol. 27, fasc. 2, p. 373, gen. 2022, doi: 10.3390/molecules27020373. [CrossRef]
- L.-P. Xiang et al., «Suppressive Effects of Tea Catechins on Breast Cancer», Nutrients, vol. 8, fasc. 8, p. 458, lug. 2016, doi: 10.3390/nu8080458. [CrossRef]
- X. Chen et al., «Effects of Tea-Polysaccharide Conjugates and Metal Ions on Precipitate Formation by Epigallocatechin Gallate and Caffeine, the Key Components of Green Tea Infusion», J Agric Food Chem, vol. 67, fasc. 13, pp. 3744–3751, apr. 2019, doi: 10.1021/acs.jafc.8b06681. [CrossRef]
- Y. Shirakami e M. Shimizu, «Possible Mechanisms of Green Tea and Its Constituents against Cancer», Molecules, vol. 23, fasc. 9, p. 2284, set. 2018, doi: 10.3390/molecules23092284. [CrossRef]
- C.-Y. Tsai et al., «Epigallocatechin-3-Gallate Suppresses Human Herpesvirus 8 Replication and Induces ROS Leading to Apoptosis and Autophagy in Primary Effusion Lymphoma Cells», IJMS, vol. 19, fasc. 1, p. 16, dic. 2017, doi: 10.3390/ijms19010016. [CrossRef]
- S. Stadlbauer et al., «Impact of Green Tea Catechin ECG and Its Synthesized Fluorinated Analogue on Prostate Cancer Cells and Stimulated Immunocompetent Cells», Planta Med, vol. 84, fasc. 11, pp. 813–819, lug. 2018, doi: 10.1055/s-0044-102099. [CrossRef]
- J. Wang, G. C. W. Man, T. H. Chan, J. Kwong, e C. C. Wang, «A prodrug of green tea polyphenol (-)-epigallocatechin-3-gallate (Pro-EGCG) serves as a novel angiogenesis inhibitor in endometrial cancer», Cancer Lett, vol. 412, pp. 10–20, gen. 2018, doi: 10.1016/j.canlet.2017.09.054. [CrossRef]
- J.-W. Oh, M. Muthu, S. S. C. Pushparaj, e J. Gopal, «Anticancer Therapeutic Effects of Green Tea Catechins (GTCs) When Integrated with Antioxidant Natural Components», Molecules, vol. 28, fasc. 5, p. 2151, feb. 2023, doi: 10.3390/molecules28052151. [CrossRef]
- A. Kale et al., «Studies on the effects of oral administration of nutrient mixture, quercetin and red onions on the bioavailability of epigallocatechin gallate from green tea extract: QUERCETIN/RED ONIONS INCREASE BIOAVAILABILITY OF EGCG», Phytother. Res., vol. 24, fasc. S1, pp. S48–S55, gen. 2010, doi: 10.1002/ptr.2899. [CrossRef]
- Y. Wang et al., «In Vitro and in Vivo Mechanism of Bone Tumor Inhibition by Selenium-Doped Bone Mineral Nanoparticles», ACS Nano, vol. 10, fasc. 11, pp. 9927–9937, nov. 2016, doi: 10.1021/acsnano.6b03835. [CrossRef]
- W. Aljohani, M. W. Ullah, X. Zhang, e G. Yang, «Bioprinting and its applications in tissue engineering and regenerative medicine», International Journal of Biological Macromolecules, vol. 107, pp. 261–275, feb. 2018, doi: 10.1016/j.ijbiomac.2017.08.171. [CrossRef]
- L. P. Gabriel et al., «Bio-based polyurethane for tissue engineering applications: How hydroxyapatite nanoparticles influence the structure, thermal and biological behavior of polyurethane composites», Nanomedicine: Nanotechnology, Biology and Medicine, vol. 13, fasc. 1, pp. 201–208, gen. 2017, doi: 10.1016/j.nano.2016.09.008. [CrossRef]
- S. Khan et al., «Catechins-Modified Selenium-Doped Hydroxyapatite Nanomaterials for Improved Osteosarcoma Therapy Through Generation of Reactive Oxygen Species», Front Oncol, vol. 9, p. 499, 2019, doi: 10.3389/fonc.2019.00499. [CrossRef]
- A.-S. Hönicke, S. A. Ender, e J. Radons, «Combined administration of EGCG and IL-1 receptor antagonist efficiently downregulates IL-1-induced tumorigenic factors in U-2 OS human osteosarcoma cells», Int J Oncol, vol. 41, fasc. 2, pp. 753–758, ago. 2012, doi: 10.3892/ijo.2012.1498. [CrossRef]
- K. Zhu e W. Wang, «Green tea polyphenol EGCG suppresses osteosarcoma cell growth through upregulating miR-1», Tumour Biol, vol. 37, fasc. 4, pp. 4373–4382, apr. 2016, doi: 10.1007/s13277-015-4187-3. [CrossRef]
- C. Dong, Z. Wang, P. Shen, Y. Chen, J. Wang, e H. Wang, «Epigallocatechin-3-gallate suppresses the growth of human osteosarcoma by inhibiting the Wnt/β-catenin signalling pathway», Bioengineered, vol. 13, fasc. 4, pp. 8490–8502, apr. 2022, doi: 10.1080/21655979.2022.2051805. [CrossRef]
- W. Wang, D. Chen, e K. Zhu, «SOX2OT variant 7 contributes to the synergistic interaction between EGCG and Doxorubicin to kill osteosarcoma via autophagy and stemness inhibition», J Exp Clin Cancer Res, vol. 37, fasc. 1, p. 37, feb. 2018, doi: 10.1186/s13046-018-0689-3. [CrossRef]
- B. Zhang, Z.-L. Shi, B. Liu, X.-B. Yan, J. Feng, e H.-M. Tao, «Enhanced anticancer effect of gemcitabine by genistein in osteosarcoma: the role of Akt and nuclear factor-kappaB», Anticancer Drugs, vol. 21, fasc. 3, pp. 288–296, mar. 2010, doi: 10.1097/CAD.0b013e328334da17. [CrossRef]
- A. Y. Berman, R. A. Motechin, M. Y. Wiesenfeld, e M. K. Holz, «The therapeutic potential of resveratrol: a review of clinical trials», NPJ Precis Oncol, vol. 1, p. 35, 2017, doi: 10.1038/s41698-017-0038-6. [CrossRef]
- P. Jeandet, A.-C. Douillet-Breuil, R. Bessis, S. Debord, M. Sbaghi, e M. Adrian, «Phytoalexins from the Vitaceae: Biosynthesis, Phytoalexin Gene Expression in Transgenic Plants, Antifungal Activity, and Metabolism», J. Agric. Food Chem., vol. 50, fasc. 10, pp. 2731–2741, mag. 2002, doi: 10.1021/jf011429s. [CrossRef]
- X. Chen et al., «Stereospecific determination ofcis- andtrans-resveratrol in rat plasma by HPLC: application to pharmacokinetic studies», Biomed. Chromatogr., vol. 21, fasc. 3, pp. 257–265, mar. 2007, doi: 10.1002/bmc.747. [CrossRef]
- B. B. Aggarwal, Y. Takada, e O. V. Oommen, «From chemoprevention to chemotherapy: common targets and common goals», Expert Opinion on Investigational Drugs, vol. 13, fasc. 10, pp. 1327–1338, ott. 2004, doi: 10.1517/13543784.13.10.1327. [CrossRef]
- C.-H. Cottart, V. Nivet-Antoine, e J.-L. Beaudeux, «Review of recent data on the metabolism, biological effects, and toxicity of resveratrol in humans», Mol Nutr Food Res, vol. 58, fasc. 1, pp. 7–21, gen. 2014, doi: 10.1002/mnfr.201200589. [CrossRef]
- F. Hajizadeh-Sharafabad, A. Sahebkar, F. Zabetian-Targhi, e V. Maleki, «The impact of resveratrol on toxicity and related complications of advanced glycation end products: A systematic review», BioFactors, vol. 45, fasc. 5, pp. 651–665, set. 2019, doi: 10.1002/biof.1531. [CrossRef]
- Q. Xiao et al., «A Review of Resveratrol as a Potent Chemoprotective and Synergistic Agent in Cancer Chemotherapy», Front Pharmacol, vol. 9, p. 1534, 2018, doi: 10.3389/fphar.2018.01534. [CrossRef]
- C. Cal, H. Garban, A. Jazirehi, C. Yeh, Y. Mizutani, e B. Bonavida, «Resveratrol and cancer: chemoprevention, apoptosis, and chemo-immunosensitizing activities», Curr Med Chem Anticancer Agents, vol. 3, fasc. 2, pp. 77–93, mar. 2003, doi: 10.2174/1568011033353443. [CrossRef]
- J.-H. Ko et al., «The Role of Resveratrol in Cancer Therapy», IJMS, vol. 18, fasc. 12, p. 2589, dic. 2017, doi: 10.3390/ijms18122589. [CrossRef]
- K. T. Noh, S. H. Chae, S. H. Chun, I. D. Jung, H. K. Kang, e Y.-M. Park, «Resveratrol suppresses tumor progression via the regulation of indoleamine 2,3-dioxygenase», Biochem Biophys Res Commun, vol. 431, fasc. 2, pp. 348–353, feb. 2013, doi: 10.1016/j.bbrc.2012.12.093. [CrossRef]
- Z.-H. Xin et al., «Finding an efficient tetramethylated hydroxydiethylene of resveratrol analogue for potential anticancer agent», BMC Chem, vol. 14, fasc. 1, p. 13, dic. 2020, doi: 10.1186/s13065-020-00667-5. [CrossRef]
- T. Sakamoto, H. Horiguchi, E. Oguma, e F. Kayama, «Effects of diverse dietary phytoestrogens on cell growth, cell cycle and apoptosis in estrogen-receptor-positive breast cancer cells», J Nutr Biochem, vol. 21, fasc. 9, pp. 856–864, set. 2010, doi: 10.1016/j.jnutbio.2009.06.010. [CrossRef]
- E. Pozo-Guisado, A. Alvarez-Barrientos, S. Mulero-Navarro, B. Santiago-Josefat, e P. M. Fernandez-Salguero, «The antiproliferative activity of resveratrol results in apoptosis in MCF-7 but not in MDA-MB-231 human breast cancer cells: cell-specific alteration of the cell cycle», Biochem Pharmacol, vol. 64, fasc. 9, pp. 1375–1386, nov. 2002, doi: 10.1016/s0006-2952(02)01296-0. [CrossRef]
- J. C. Tou, «Resveratrol supplementation affects bone acquisition and osteoporosis: Pre-clinical evidence toward translational diet therapy», Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease, vol. 1852, fasc. 6, pp. 1186–1194, giu. 2015, doi: 10.1016/j.bbadis.2014.10.003. [CrossRef]
- D. Bellavia et al., «Non-flavonoid polyphenols in osteoporosis: preclinical evidence», Trends Endocrinol Metab, vol. 32, fasc. 7, pp. 515–529, lug. 2021, doi: 10.1016/j.tem.2021.03.008. [CrossRef]
- Y. Li, C.-M. Bäckesjö, L.-A. Haldosén, e U. Lindgren, «Resveratrol inhibits proliferation and promotes apoptosis of osteosarcoma cells», Eur J Pharmacol, vol. 609, fasc. 1–3, pp. 13–18, mag. 2009, doi: 10.1016/j.ejphar.2009.03.004. [CrossRef]
- Y. Liu et al., «Pterostilbene exerts antitumor activity against human osteosarcoma cells by inhibiting the JAK2/STAT3 signaling pathway», Toxicology, vol. 304, pp. 120–131, feb. 2013, doi: 10.1016/j.tox.2012.12.018. [CrossRef]
- Y. Zou, J. Yang, e D. Jiang, «Resveratrol inhibits canonical Wnt signaling in human MG-63 osteosarcoma cells», Molecular Medicine Reports, vol. 12, fasc. 5, pp. 7221–7226, nov. 2015, doi: 10.3892/mmr.2015.4338. [CrossRef]
- L. Peng e D. Jiang, «Resveratrol eliminates cancer stem cells of osteosarcoma by STAT3 pathway inhibition», PLoS ONE, vol. 13, fasc. 10, p. e0205918, ott. 2018, doi: 10.1371/journal.pone.0205918. [CrossRef]
- N. Xu, L. Wang, S. Fu, e B. Jiang, «Resveratrol is cytotoxic and acts synergistically with NF-κB inhibition in osteosarcoma MG-63 cells», Arch Med Sci, vol. 17, fasc. 1, pp. 166–176, gen. 2021, doi: 10.5114/aoms.2020.100777. [CrossRef]
- A. De Luca et al., «Multiple Effects of Resveratrol on Osteosarcoma Cell Lines», Pharmaceuticals (Basel), vol. 15, fasc. 3, p. 342, mar. 2022, doi: 10.3390/ph15030342. [CrossRef]
- T. Mengie Ayele, Z. Tilahun Muche, A. Behaile Teklemariam, A. Bogale Kassie, e E. Chekol Abebe, «Role of JAK2/STAT3 Signaling Pathway in the Tumorigenesis, Chemotherapy Resistance, and Treatment of Solid Tumors: A Systemic Review», J Inflamm Res, vol. 15, pp. 1349–1364, 2022, doi: 10.2147/JIR.S353489. [CrossRef]
- C.-F. Chiu et al., «NF-κB-driven suppression of FOXO3a contributes to EGFR mutation-independent gefitinib resistance», Proc Natl Acad Sci U S A, vol. 113, fasc. 18, pp. E2526-2535, mag. 2016, doi: 10.1073/pnas.1522612113. [CrossRef]
- A. Mitra, L. Mishra, e S. Li, «EMT, CTCs and CSCs in tumor relapse and drug-resistance», Oncotarget, vol. 6, fasc. 13, pp. 10697–10711, mag. 2015, doi: 10.18632/oncotarget.4037. [CrossRef]
- D. Xie et al., «Antitumor activity of resveratrol against human osteosarcoma cells: a key role of Cx43 and Wnt/β-catenin signaling pathway», Oncotarget, vol. 8, fasc. 67, pp. 111419–111432, dic. 2017, doi: 10.18632/oncotarget.22810. [CrossRef]
- Y. Shen, P. R. Khusial, X. Li, H. Ichikawa, A. P. Moreno, e G. S. Goldberg, «Src Utilizes Cas to Block Gap Junctional Communication Mediated by Connexin43», Journal of Biological Chemistry, vol. 282, fasc. 26, pp. 18914–18921, giu. 2007, doi: 10.1074/jbc.M608980200. [CrossRef]
- H. Clevers, «Wnt/beta-catenin signaling in development and disease», Cell, vol. 127, fasc. 3, pp. 469–480, nov. 2006, doi: 10.1016/j.cell.2006.10.018. [CrossRef]
- P. J. Morin et al., «Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC», Science, vol. 275, fasc. 5307, pp. 1787–1790, mar. 1997, doi: 10.1126/science.275.5307.1787. [CrossRef]
- V. Korinek et al., «Constitutive transcriptional activation by a beta-catenin-Tcf complex in APC-/- colon carcinoma», Science, vol. 275, fasc. 5307, pp. 1784–1787, mar. 1997, doi: 10.1126/science.275.5307.1784. [CrossRef]
- S.-O. Yoon, S.-J. Park, C.-H. Yun, e A.-S. Chung, «Roles of matrix metalloproteinases in tumor metastasis and angiogenesis», J Biochem Mol Biol, vol. 36, fasc. 1, pp. 128–137, gen. 2003, doi: 10.5483/bmbrep.2003.36.1.128. [CrossRef]
- S.-F. Yang et al., «Upregulation of miR-328 and inhibition of CREB-DNA-binding activity are critical for resveratrol-mediated suppression of matrix metalloproteinase-2 and subsequent metastatic ability in human osteosarcomas», Oncotarget, vol. 6, fasc. 5, pp. 2736–2753, feb. 2015, doi: 10.18632/oncotarget.3088. [CrossRef]
- G. Ranieri, R. Patruno, E. Ruggieri, S. Montemurro, P. Valerio, e D. Ribatti, «Vascular endothelial growth factor (VEGF) as a target of bevacizumab in cancer: from the biology to the clinic», Curr Med Chem, vol. 13, fasc. 16, pp. 1845–1857, 2006, doi: 10.2174/092986706777585059. [CrossRef]
- L. C. J. Baker et al., «The HIF-pathway inhibitor NSC-134754 induces metabolic changes and anti-tumour activity while maintaining vascular function», Br J Cancer, vol. 106, fasc. 10, pp. 1638–1647, mag. 2012, doi: 10.1038/bjc.2012.131. [CrossRef]
- Z. Liu, Y. Li, e R. Yang, «Effects of resveratrol on vascular endothelial growth factor expression in osteosarcoma cells and cell proliferation», Oncol Lett, vol. 4, fasc. 4, pp. 837–839, ott. 2012, doi: 10.3892/ol.2012.824. [CrossRef]
- Q. Liu, M. Li, S. Wang, Z. Xiao, Y. Xiong, e G. Wang, «Recent Advances of Osterix Transcription Factor in Osteoblast Differentiation and Bone Formation», Front. Cell Dev. Biol., vol. 8, p. 601224, dic. 2020, doi: 10.3389/fcell.2020.601224. [CrossRef]
- V. T. Baron, R. Pio, Z. Jia, e D. Mercola, «Early Growth Response 3 regulates genes of inflammation and directly activates IL6 and IL8 expression in prostate cancer», Br J Cancer, vol. 112, fasc. 4, pp. 755–764, feb. 2015, doi: 10.1038/bjc.2014.622. [CrossRef]
- H. Jayatilaka et al., «Synergistic IL-6 and IL-8 paracrine signalling pathway infers a strategy to inhibit tumour cell migration», Nat Commun, vol. 8, fasc. 1, p. 15584, mag. 2017, doi: 10.1038/ncomms15584. [CrossRef]
- T. H. Kim et al., «Resveratrol enhances chemosensitivity of doxorubicin in multidrug-resistant human breast cancer cells via increased cellular influx of doxorubicin», Biochimica et Biophysica Acta (BBA) - General Subjects, vol. 1840, fasc. 1, pp. 615–625, gen. 2014, doi: 10.1016/j.bbagen.2013.10.023. [CrossRef]
- A. S. Barros, E. C. Costa, A. S. Nunes, D. De Melo-Diogo, e I. J. Correia, «Comparative study of the therapeutic effect of Doxorubicin and Resveratrol combination on 2D and 3D (spheroids) cell culture models», International Journal of Pharmaceutics, vol. 551, fasc. 1–2, pp. 76–83, nov. 2018, doi: 10.1016/j.ijpharm.2018.09.016. [CrossRef]
- M. Ren et al., «Resveratrol synergizes with cisplatin in antineoplastic effects against AGS gastric cancer cells by inducing endoplasmic reticulum stress-mediated apoptosis and G2/M phase arrest», Oncol Rep, lug. 2020, doi: 10.3892/or.2020.7708. [CrossRef]
- D. Şöhretoğlu, M. Y. Baran, R. Arroo, e A. Kuruüzüm-Uz, «Recent advances in chemistry, therapeutic properties and sources of polydatin», Phytochem Rev, vol. 17, fasc. 5, pp. 973–1005, ott. 2018, doi: 10.1007/s11101-018-9574-0. [CrossRef]
- P. Jeandet et al., «Phytostilbenes as agrochemicals: biosynthesis, bioactivity, metabolic engineering and biotechnology», Nat Prod Rep, vol. 38, fasc. 7, pp. 1282–1329, lug. 2021, doi: 10.1039/d0np00030b. [CrossRef]
- S. Chen et al., «Polydatin down-regulates the phosphorylation level of Creb and induces apoptosis in human breast cancer cell», PLoS ONE, vol. 12, fasc. 5, p. e0176501, mag. 2017, doi: 10.1371/journal.pone.0176501. [CrossRef]
- J. Ye et al., «Polydatin inhibits mast cell-mediated allergic inflammation by targeting PI3K/Akt, MAPK, NF-κB and Nrf2/HO-1 pathways», Sci Rep, vol. 7, fasc. 1, p. 11895, set. 2017, doi: 10.1038/s41598-017-12252-3. [CrossRef]
- Y.-S. Shen et al., «Polydatin improves osteogenic differentiation of human bone mesenchymal stem cells by stimulating TAZ expression via BMP2-Wnt/β-catenin signaling pathway», Stem Cell Res Ther, vol. 11, fasc. 1, p. 204, mag. 2020, doi: 10.1186/s13287-020-01705-8. [CrossRef]
- A. Karami, S. Fakhri, L. Kooshki, e H. Khan, «Polydatin: Pharmacological Mechanisms, Therapeutic Targets, Biological Activities, and Health Benefits», Molecules, vol. 27, fasc. 19, p. 6474, ott. 2022, doi: 10.3390/molecules27196474. [CrossRef]
- Y. Chen et al., «Anti-oxidant polydatin (piceid) protects against substantia nigral motor degeneration in multiple rodent models of Parkinson’s disease», Mol Neurodegeneration, vol. 10, fasc. 1, p. 4, dic. 2015, doi: 10.1186/1750-1326-10-4. [CrossRef]
- M. Chen, «Enzymatic transformation of polydatin to resveratrol by piceid-β-d-glucosidase from Aspergillus oryzae».
- W.-J. Cao, K. Wu, C. Wang, e D.-M. Wan, «Polydatin-induced cell apoptosis and cell cycle arrest are potentiated by Janus kinase 2 inhibition in leukemia cells», Mol Med Rep, vol. 13, fasc. 4, pp. 3297–3302, apr. 2016, doi: 10.3892/mmr.2016.4909. [CrossRef]
- H.-L. Wang et al., «Comparative studies of polydatin and resveratrol on mutual transformation and antioxidative effect in vivo», Phytomedicine, vol. 22, fasc. 5, pp. 553–559, mag. 2015, doi: 10.1016/j.phymed.2015.03.014. [CrossRef]
- H. Zhang et al., «Anticancer effects and potential mechanisms of ginsenoside Rh2 in various cancer types (Review)», Oncol Rep, vol. 45, fasc. 4, p. 33, feb. 2021, doi: 10.3892/or.2021.7984. [CrossRef]
- M. A. Shah et al., «Uncovering the Anticancer Potential of Polydatin: A Mechanistic Insight», Molecules, vol. 27, fasc. 21, p. 7175, ott. 2022, doi: 10.3390/molecules27217175. [CrossRef]
- B. Wang, Y. Zhang, Z. Mao, D. Yu, e C. Gao, «Toxicity of ZnO nanoparticles to macrophages due to cell uptake and intracellular release of zinc ions», J Nanosci Nanotechnol, vol. 14, fasc. 8, pp. 5688–5696, ago. 2014, doi: 10.1166/jnn.2014.8876. [CrossRef]
- C. Hong et al., «Novel ginsenoside-based multifunctional liposomal delivery system for combination therapy of gastric cancer», Theranostics, vol. 9, fasc. 15, pp. 4437–4449, 2019, doi: 10.7150/thno.34953. [CrossRef]
- L. Mele et al., «A new inhibitor of glucose-6-phosphate dehydrogenase blocks pentose phosphate pathway and suppresses malignant proliferation and metastasis in vivo», Cell Death Dis, vol. 9, fasc. 5, p. 572, mag. 2018, doi: 10.1038/s41419-018-0635-5. [CrossRef]
- G. Xu, G. Kuang, W. Jiang, R. Jiang, e D. Jiang, «Polydatin promotes apoptosis through upregulation the ratio of Bax/Bcl-2 and inhibits proliferation by attenuating the β-catenin signaling in human osteosarcoma cells», Am J Transl Res, vol. 8, fasc. 2, pp. 922–931, 2016.
- C.-Q. Jiang, L.-L. Ma, Z.-D. Lv, F. Feng, Z. Chen, e Z.-D. Liu, «Polydatin induces apoptosis and autophagy via STAT3 signaling in human osteosarcoma MG-63 cells», J Nat Med, vol. 74, fasc. 3, pp. 533–544, giu. 2020, doi: 10.1007/s11418-020-01399-5. [CrossRef]
- W. Zhao, Z. Chen, e M. Guan, «Polydatin enhances the chemosensitivity of osteosarcoma cells to paclitaxel», J Cell Biochem, vol. 120, fasc. 10, pp. 17481–17490, ott. 2019, doi: 10.1002/jcb.29012. [CrossRef]
- T. Hu, Z. Fei, H. Su, R. Xie, e L. Chen, «Polydatin inhibits proliferation and promotes apoptosis of doxorubicin-resistant osteosarcoma through LncRNA TUG1 mediated suppression of Akt signaling», Toxicol Appl Pharmacol, vol. 371, pp. 55–62, mag. 2019, doi: 10.1016/j.taap.2019.04.005. [CrossRef]
- A. Luce et al., «Polydatin Induces Differentiation and Radiation Sensitivity in Human Osteosarcoma Cells and Parallel Secretion through Lipid Metabolite Secretion», Oxidative Medicine and Cellular Longevity, vol. 2021, pp. 1–11, lug. 2021, doi: 10.1155/2021/3337013. [CrossRef]
- V. Kota e H. Hama, «2′-Hydroxy ceramide in membrane homeostasis and cell signaling», Advances in Biological Regulation, vol. 54, pp. 223–230, gen. 2014, doi: 10.1016/j.jbior.2013.09.012. [CrossRef]
- E. Matsuzaki et al., «Sphingosine-1-phosphate promotes the nuclear translocation of β-catenin and thereby induces osteoprotegerin gene expression in osteoblast-like cell lines», Bone, vol. 55, fasc. 2, pp. 315–324, ago. 2013, doi: 10.1016/j.bone.2013.04.008. [CrossRef]
- F. Mostafa, A. Abdel-Moneim, M. Abdul-Hamid, S. R. Galaly, e H. M. Mohamed, «Polydatin and polydatin-loaded chitosan nanoparticles attenuate diabetic cardiomyopathy in rats», J Mol Histol, vol. 52, fasc. 2, pp. 135–152, apr. 2021, doi: 10.1007/s10735-020-09930-4. [CrossRef]
- Q. Guan, W. Chen, X. Hu, X. Wang, e L. Li, «Novel nanoliposomal delivery system for polydatin: preparation, characterization, and in vivo evaluation», DDDT, p. 1805, mar. 2015, doi: 10.2147/DDDT.S77615. [CrossRef]
- L. Lin et al., «Nanodrug with ROS and pH Dual-Sensitivity Ameliorates Liver Fibrosis via Multicellular Regulation», Adv Sci (Weinh), vol. 7, fasc. 7, p. 1903138, apr. 2020, doi: 10.1002/advs.201903138. [CrossRef]
- J. Xiao, F. Wang, J. Liu, L. Wang, G. Kai, e X. Yu, «Effect of ZnO#ZnS QDs heterojunctures on the stilbenes–plasma proteins interactions», Mol. BioSyst., vol. 7, fasc. 8, p. 2452, 2011, doi: 10.1039/c1mb05087g. [CrossRef]
- A. Basta-Kaim et al., «Protective effects of polydatin in free and nanocapsulated form on changes caused by lipopolysaccharide in hippocampal organotypic cultures», Pharmacol Rep, vol. 71, fasc. 4, pp. 603–613, ago. 2019, doi: 10.1016/j.pharep.2019.02.017. [CrossRef]
- A. M. Abd El-Hameed, A. I. Yousef, S. M. Abd El-Twab, A. A. G. El-Shahawy, e A. Abdel-Moneim, «Hepatoprotective Effects of Polydatin-Loaded Chitosan Nanoparticles in Diabetic Rats: Modulation of Glucose Metabolism, Oxidative Stress, and Inflammation Biomarkers», Biochemistry (Mosc), vol. 86, fasc. 2, pp. 179–189, feb. 2021, doi: 10.1134/S0006297921020061. [CrossRef]
- E. D. Moyers-Montoya, R. G. Escobedo-González, C. L. Vargas-Requena, P. E. Garcia-Casillas, e C. A. Martínez-Pérez, «Epithelial Growth Factor-Anchored on Polycaprolactone/6-deoxy-6-amino-β-cyclodextrin Nanofibers: In Vitro and In Vivo Evaluation», Polymers, vol. 13, fasc. 8, p. 1303, apr. 2021, doi: 10.3390/polym13081303. [CrossRef]
- S. Lama et al., «Polydatin Incorporated in Polycaprolactone Nanofibers Improves Osteogenic Differentiation», Pharmaceuticals (Basel), vol. 15, fasc. 6, p. 727, giu. 2022, doi: 10.3390/ph15060727. [CrossRef]
- Á. Peirotén, D. Bravo, e J. M. Landete, «Bacterial metabolism as responsible of beneficial effects of phytoestrogens on human health», Crit Rev Food Sci Nutr, vol. 60, fasc. 11, pp. 1922–1937, 2020, doi: 10.1080/10408398.2019.1622505. [CrossRef]
- J. Feng, Z. Shi, e Z. Ye, «Effects of metabolites of the lignans enterolactone and enterodiol on osteoblastic differentiation of MG-63 cells», Biol Pharm Bull, vol. 31, fasc. 6, pp. 1067–1070, giu. 2008, doi: 10.1248/bpb.31.1067. [CrossRef]
- S. C. Thomasset, D. P. Berry, G. Garcea, T. Marczylo, W. P. Steward, e A. J. Gescher, «Dietary polyphenolic phytochemicals—promising cancer chemopreventive agents in humans? A review of their clinical properties», Int. J. Cancer, vol. 120, fasc. 3, pp. 451–458, feb. 2007, doi: 10.1002/ijc.22419. [CrossRef]
- R. H. Dashwood, «Frontiers in Polyphenols and Cancer Prevention3», The Journal of Nutrition, vol. 137, fasc. 1, pp. 267S-269S, gen. 2007, doi: 10.1093/jn/137.1.267S. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



