
Submitted:
04 August 2023
Posted:
04 August 2023
You are already at the latest version
Abstract
Wilted hemp (Cannabis sativa L.) plants were observed in August 2019 in commercial fields around Osijek, Croatia. Plants and roots with disease symptoms were collected. The single-spored isolates produced septate cottony white to light pink aerial mycelium and purple undersurface on potato dextrose agar (PDA). Smooth and hyaline hyphae were branched and septate. Macroconidia were fusiform to sickle-shaped with foot-shaped basal cell, elongated apical cell and three to five septa. Sequencing of the internal transcribed spacer and the partial elongation factor 1-α gene identified species as Fusarium oxysporum. Artificial infection fulfills Koch’s postulates, producing plants which showed stunted growth and wilt symptoms similar to those observed in the commercial fields. Control seedlings remained symptomless and healthy. To the best of our knowledge, this is the first report of hemp Fusarium wilt causing F. oxysporum in Croatia. Considering that F. oxysporum has been reported in main field crops in Croatia, the presence of this pathogen could cause economically significant hemp production decreases, especially in humid and cold springs and susceptible varieties.
Keywords:
hemp
; Fusarium oxysporum
; wilt
; Croatia
1. Introduction
Hemp (Cannabis sativa L., Cannabaceae) is considered one of the oldest crops known to man [1]. It is estimated that its use dates back to 10,000 years ago [2] and a hypothesis of co-evolution of the genus Cannabis with the human species has been postulated [3]. The hemp plant is native to India and Persia, although it has been cultivated in nearly all temperate and tropical countries [4,5]. Hemp is a versatile herbaceous crop that has been used for fiber, food, and medicinal purposes [6,7,8,9]. Hemp is an inexpensive and available bast natural fiber [10]. Fiber production from hemp has been conducted over many centuries, for end uses ranging from textiles and papers (hemp paper was used in the first copies of the Bible) to ropes and sails [11].
The cultivation of hemp dates back to China around 2700 BC and is believed to have then expanded across Asia, making its way to Europe 2000–2200 years ago [3,4]. As a multi-use crop, hemp is considered one of the oldest plants cultivated to provide nutritional and medicinal benefits [7,12]. Also, hemp provides raw material to a large number of traditional and innovative industrial applications [1]. In recent years, the interest in investigating the potential use of industrial hemp in food and nutraceuticals has been growing [13]. Pressure from weeds, insects and diseases are among the major challenges in production of C. sativa [14]. Fusarium species can parasitize a wide range of plants, including vegetables, flowers, field crops (cotton, hemp, and tobacco), herbaceous perennial ornamentals, and plantation crops (banana, plantain, coffee, sugarcane) [15]. Also, they can parasite vertebrates, insects, humans, or even other fungi. Conducted mycological analysis of seed and grain of many crops from 2002 to 2008 in Croatia revealed the presence of Fusarium species on wheat, barley, oat, triticale, maize, tobacco, bean, pea, soybean, lupin, vetch, alfalfa, clover, flax, beet, spinach and lettuce seed and grain [16]. F. oxysporum has been confirmed as a cause of wilting soybeans in Croatia [17].
All parts of the plant at all growth stages are infected by one or more pathogens while pathogens in the genus Fusarium are among the most destructive pathogens of C. sativa [18]. Several Fusarium species attack hemp crops and cause damping off, including Fusarium solani, Fusarium oxysporum [19] and less frequently Fusarium sulphureum, Fusarium avenaceum, Fusarium graminearum, Fusarium culmorum, Fusarium avenaceum and Fusarium fujikuroi [18]. The time of infection and vegetative growth phases results in decreased plant quality or even total plant loss. Species of Fusarium also cause significant postharvest losses and produce mycotoxins that further limit the value of the crop [20]. Because, for many countries, the legal status of the plant changed from crop to illegal drug (1950s) and back again in the 2000s [21], the bulk of literature on C. sativa and Fusarium is written in two distinct time periods (prior to 1960 and after 1990). There are some exceptions to this bimodal distribution, particularly publications reporting the use of F. oxysporum f. sp. cannabis as a biological control of the plants [22].
F. oxysporum is a common fungal pathogen that causes wilt disease in nurseries and in field cultivation and causes high losses [23]. Some of these fungi also cause cotyledon drop [24]. In mature plants these pathogens cause Fusarium root rot, stem canker, and wilt [25]. Fusarium wilt is the name of a disease caused by two microscopic fungi that infect hemp roots and then move systemically through the plant. Symptoms of Fusarium wilt begin as small, dark, irregular spots on lower leaves [26] and pathogen-affected leaves suddenly become chlorotic (yellow). Wilt symptoms begin with an upward curling of leaf tips. Wilted leaves dry to a yellow-tan color and hang on the plant without falling off (Figure 1). Petioles, branches, and sometimes even the stalk may droop while stems also turn yellow-tan. Cutting into wilted stems reveals a reddish-brown discoloration of xylem tissue. Pulled-up roots show no external symptoms. Fusarium wilt is a warm-weather disease, and optimal temperature for fungal growth is 26°C. Disease symptoms may not become evident until the advent of hot summer temperature [27]. Fusarium conidia spread via splashed rain and water runoff.
Fusarium spp. can live saprophytically on roots, stems, leaves, flowers, and seed of diseased and dead plants [28]. The fungus can survive on seed (internal and external) or as spores or mycelium in dead or infected tissues. The pathogen is disseminated by air, equipment, and water [15].
Fusarium wilts are of worldwide importance and disease severity is favored by warm climates and warm soil temperatures [18]. Wilt development is favored by air temperatures of 28°C, low levels of nitrogen and phosphorus, elevated levels of potassium, low soil pH, short day length, and low light intensity [15]. In other hosts, virulence of Fusarium can be enhanced with ammonium nitrogen, and decreased by nitrate nitrogen [29]. Many species of Fusarium co-exist with their hosts as endophytes and protect the host against pathogens and F. oxysporum has been reported as endophytic in C. sativa [20].
F. oxysporum is typically soil borne and is the most economically important and the most commonly isolated species of Fusarium [30]. Due to the large numbers of spores produced, oil infested with Fusarium may remain so indefinitely [31]. Seed-borne infections lay dormant until seedlings sprout the following spring. Mycoherbicide researchers reported that spore-coated hemp seeds effectively spread F. oxysporum through the soil [32].
Fusarium wilt reduces fiber quality and fiber yield and reduces seed production and seed quality. Seeds infested by the wilt fungus should not be used for human consumption, or for cultivation or breeding purposes. Mycotoxins, produced by many species of Fusarium, are of greatest agronomic importance limits the food and feed supply [18]. Mycotoxins can create serious health problems in humans and animals [33].
2. Results
2.1. Fungal Isolate and Identification
Numerous wilted hemp plants (~15%) were observed in August 2019 in commercial fields in Vladislavci, around Osijek (45.4646950° N, 18.5674770° E), Croatia. Plants and roots with symptoms of external and internal browning (Figure 1) at the base of stems, interveinal chlorosis of leaves and death of shoots were collected. Although according to literature pulled-up roots infected with F. oxysporum show no external symptoms [27], we found a reddish-brown discoloration on roots after removing external bark layer (Figure 1b).
Colony characteristics were observed after 7 days of incubation. Single-spore isolates produced septate cottony white to light pink aerial mycelium and colorless to light pink undersurface on potato dextrose agar (PDA) (Figure 2a). Smooth and hyaline hyphae were branched and septate. Macroconidia were fusiform to sickle-shaped with foot-shaped basal cell, elongated apical cell and three to five septa. In all our isolates three-septate macroconidia were commonly observed with average size 35.21±7.8 x 3.52±1.1 µm. Oval single-celled microconidia (Figure 2b) were produced only in false heads on short monophialides. The average size of microconidia was 8.91 x 2.17 µm.
2.2. Molecularal Identification
The internal transcribed spacers (ITS) sequences of the isolate HFox1 (524 bp) (GenBank accession No. OM475708 was amplified using primers ITS4/ITS5 [27]. The isolate was characterized by ITS sequencing and identified with 100% identity as F. oxysporum. Phylogenetic analyses (Figure 3) grouped HFox1 isolate with a high degree of sequence identity (99–100%) within the F. oxysporum complex. Isolate OM475708 showed 99.81% similarity with e-value of 0 to the reference sequences of F. oxysporum (GU724513). Because of its polyphyletic nature, F. oxysporum forms a cluster with different groups of the F. oxysporum species complex [34].
The sequences of the same isolate HFox1 (634 bp)(GenBank accession number OR149071) was amplified using primers EF1/EF2 [27]. The isolate was identified with 100% identity as F. oxysporum. Phylogenetic analyses (Figure 4) again grouped HFox1 isolate with a high degree of sequence identity (99–100%) within the F. oxysporum complex. Isolate OR149071 showed 100% similarity with e-value of 0 to the reference sequences of F. oxysporum (MK461973).
2.3. Artificial Inoculation
Two weeks after artificial inoculation, the inoculated seedlings showed stunted growth and wilt symptoms similar to those observed in the commercial fields. Stems of inoculated young plants showed the symptoms of external and internal browning at the base of stems, interveinal chlorosis of leaves, similarly to the diseased plants collected in the field (Figure 5a and 5).
The fungus was successfully re-isolated from infected plants on PDA with the same morphological characters as before, which fulfilled Koch’s postulates. The control plants remained symptomless and healthy (Figure 5c).
The isolates were stored in sterile distilled water at -18°C and archived at the Department of Phytomedicine, Faculty of Agrobiotechnical Sciences Osijek, Josip Juraj Strossmayer University of Osijek for further use.
3. Discussion
In this study, according to cultural and conidial morphology, ITS4/ITS5 and EF1/EF2 sequence based phylogenetic analysis and pathogenicity test, the pathogenic fungus was identified as F. oxysporum, and confirmed to be the causal agent of Fusarium wilt in hemp (C. sativa). To the best of our knowledge, this is the first report of hemp Fusarium wilt causing F. oxysporum in Croatia. Considering that F. oxysporum has been reported not only in field crops [16,17] but also on many weed species [35], the presence of this pathogen could cause economically significant hemp production decreases. F. oxysporum attacks hemp seeds causing a decrease in quality. Fibranova [36] hemp cultivar was used in the pathogenicity test because our collected isolates were found in the field sown with this cultivar. The authors did not conduct a statistical analysis during the trial because the aim of this study was nothing but to simply determine the existence of a new pathogen which causes disease on hemp plants in commercial field in the Republic of Croatia. Although is to be assumed that there may be differences in the susceptibility of cannabis varieties to this pathogen, the susceptibility of this variety was not the subject of this study.
In the study of Ivic et al [16] the authors list the crops in Croatia on which they found the presence of Fusarium species, but do not determine them to Form species (formae specialis) depending on what hosts they are to infect.
Using the described methodology, morphological characteristics and based on molecular analyzes, we determined new isolate HFox1. We confirmed pathogenicity of HFox 1 isolate to hemp by applying Koch's postulates. The sequences [21,37] of isolate (GenBank accession no. OM475708 and OR149071) was compared with those available in the NCBI (National Center for Biotechnology Information, http://www.ncbi.nlm.gov/; accessed on 22 March 2022 and 12 July 2023) genetic database by using the Basic Local Alignment Tool (BLAST) algorithm showed 100% similarity with e-value of 0 to the reference sequences of Fusarium oxysporum (GU724513 and KX196809), as well as F. oxysporum (MK461973).
Hemp was regularly grown in eastern Croatia after, and especially between the two Great Wars. By the middle of the twentieth century, the largest producers will remain concentrated in parts of Russia, Ukraine, Hungary and former Yugoslavia [38]. During the former Yugoslavia, hemp was one of the leading cultures, and the country itself was 3rd in the World in hemp production [39]. The area of Slavonia was significantly oriented towards cultivation, processing and export of industrial hemp [40]. In the 1960s last century, hemp production was almost completely abandoned by passing laws that prescribed strict conditions for industrial cultivation hemp [40]. With cessation of production as was the accompanying processing industry.
The industry hemp in Croatia has had a sharp increase in the past 5 years [39]. Production has been in constant increase, from 1560 ha in 2015 to 2476 ha in 2019 [41]. Upon the resumption of cannabis production in recent years in Croatia, the diseases have not been identified, or they have not been a problem nor have they been reported by producers. With the liberalization of the legislation, hemp production is expected to grow in the coming years. Narrowing the crop rotation could potentially increase the problem with diseases in general as well as the disease caused by Fusarium, especially in humid and warm springs and more susceptible varieties [15,27].
4. Materials and Methods
4.1. Fungal Strain Collection, Isolation and Growth Conditions
The stems and roots of the infected hemp plants were collected in August 2019 in commercial fields in Vladislavci, around Osijek (45.4646950° N, 18.5674770° E), Croatia. Several pieces of the stem and root cortex (2- to 3 cm pieces) were sampled with sterile blades, placed in sterile plastic bags and transferred to the laboratory. Small pieces of tissue (0.5-1 cm) were treated with 1% sodium hypochlorite solution for 5 min and rinsed three times with sterile water. Pieces were left to dry under hood flow and placed on half-strength potato dextrose agar (PDA, Biolife Italiana, Italy) amended with streptomycin stock solution (Sigma-Aldrich, St. Louis, MO, USA) to inhibit bacterial growth.
The culture plates were incubated at 25 ± 2°C for 7 days and purified to obtain single-spore isolates by sterile needles using a Nikon E400 Eclipse stereomicroscope under M18 laminar flow hood. The single-spore isolates produced septate cottony white to light pink aerial mycelium and purple undersurface on PDA (Figure 2a). Smooth and hyaline hyphae were branched and septate. Morphological identification was done according to The Fusarium Laboratory Manual [42].
4.2. Fungal Strain Molecular Isolation
4.2.1. Internal Transcribed Spacer Region Analysis
The Fusarium oxysporum isolate was identified at the species level by ITS rDNA sequencing. Mycelium was harvested form the colony surface by a sterile medicine spoon [43]. The primers ITS4/ITS5 (ITS4 5’-TCCTCCGCTTATTGATATGC-3’ and ITS5 5’-GAAAGTAAAAGTCGTAACAAGG-3’) [43,44] were used in the following reaction mixture: ~150 mg of fresh mycelium, 2 μL of 20 mg μL-1 bovine sieroalbumin solution; 1.5 μL of 50U μL-1 Taq polymerase solution; 5 μL Buffer 10x; 1 μL od 10 mM dNTP, 4 μL of 50 Mn MgCl2 solution; 2 μL ITS4 Primer forward; 2 μL ITS5 primer reverse; and sterile destilled water up to 50 μL. Polymerase chain reaction (PCR) amplification was carried out in a thermal cycler (SimpliAmpTM Thermal Cycler – Applied Biosystems) with the following program: 95 °C for 3 min, followed by 34 cycles of denaturation at 95 °C for 30 s, annealing at 54-57 °C for 30 s, extension at 72 °C for 1 min and final extension 72 °C for 8 min. Sequencing was carried out by Microsynth AB Company (Balgach, Switzerland), starting from the solution of amplicons obtained by PCR checked on 1.5% agarose gel.
The ITS sequences were compared with those available in the National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.gov/; accessed on 25 April 2023) genetic database by using the Basic Local Alignment Tool (BLAST) algorithm and using only sequence identity values above 99%.
4.2.2. Translation Elongation Factor 1-α Gene Analysis
A second PCR analysis was performed using fungi-specific tef pair of primers EF1 (5′ ATGGGTAAGGAAGGACAAG 3′) and EF 2 (5′ GGAGAGTACCAGTGCATCAT 3′) [45]. Total DNA from isolate HFox1 was extracted with OmniPrep™ for Fungi PCR Kit (G-Biosciences Co., USA). Amplification of the translation elongation factor 1α was performed with primer pair EF1 TEF -1α/EF2 [1] using EmeraldAmp MAX PCR Master Mix (Takara Bio Inc.) in a reaction mixture containing 0.2 µM µL of each primer and 2 µL of undiluted genomic DNA. Polymerase chain reaction (PCR) was conducted using MiniAmp Plus Thermal Cycler (Applied Biosystems) with the following conditions: an initial denaturation step at 95°C for 5 min, 35 amplification cycles with denaturation at 94°C for 30 s, primer annealing at 55°C for 45 s, and elongation at 72°C for 1 min 30 s, followed by a final elongation step at 72°C for 10 min. Obtained PCR products were visualized on 1% agarose gel pre-stained with SYBR Safe DNA Gel Stain, and documented using the Kodak EDAS 290 system with a UV transilluminator (UVITEC). PCR products were purified Wizard® SV Gel and PCR Clean-Up System (Promega Corporation, Madison, USA), and sequenced by Macrogen Europe (Amsterdam, The Netherlands) on both strands with the same primers used for amplification. The sequence assembling was performed with DNADynamo (BlueTractorSoftware), and nucleotide sequence data was deposited in GenBank under accession number OR149071. The result was confirmed with an additional primer pair Fa+7/Ra+6 [46] which gave an identical nucleotide sequence for the translation elongation factor 1α of isolate HFox1 Amplification with primers Fa+7/Ra+6 was the same as described for EF1 TEF -1α/EF2 except that primer annealing was at 67°C for 1 min.
The EF sequences were compared with those available in the National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.gov/; accessed on 12 July 2023) genetic database by using the Basic Local Alignment Tool (BLAST) algorithm and using only sequence identity values above 99%.
4.3. Phylogenetic Analysis
This analysis involved 42 nucleotide sequences for ITS and 36 nucleotide sequences for TEF. Both, ITS and TEF sequence of HFox1 isolate was submitted to National Center for Biotechnology Information (NCBI) – GenBank accession numbers OM475708 and OR149071. The bioinformatics algorithm basic local alignment search tool (BLASTn) program [49] was used for sequence analysis. Phylogenetic trees were constructed using ClustalW sequence alignment and evolutionary analyses were conducted in MEGA11 [47]. Bayesian analyses were performed in MrBayes v.3.2.7a [48]. The Markov Chain Monte Carlo sampling (MCMC) analyses were conducted with six simultaneous Markov chain. They were run for 120000 generations; sampling the trees at every 100th generation. Resulting phylogenetic trees (Figure 3 and 4) were drawn using FigTree v.1.4.4 [50].
4.4. Pathogenicity Test
To confirm their pathogenicity, isolate HFox1 was used for preparing inoculum. A mixture of wheat and barley seeds (3:1, v/v) was soaked in water overnight. The following day, excess water was decanted and seeds were autoclaved and inoculated with F. oxysporum isolate. Inoculated grains were incubated for two weeks at 25± 2 °C, protected from sunlight. Macroconidia were washed off the colonized grains, and conidial suspension was diluted to a final concentration of 1 × 106 conidia mL−1 [36,51]. Control inoculum was prepared by applying sterile distilled water to seed. After 2 weeks, 6 g of the inoculum was placed 3-4 cm below the surface of sterile soil in each 12x10 cm pot. Twenty-seven hemp seeds of cultivar Fibranova [36] were sown in pots with universal natural substrate Florafin (three seeds / pot in 3 replications). Several pieces of the stems cortex (1- to 2 cm pieces) were samples with sterile blades were treated with 1% sodium hypochlorite solution for 5 min and rinsed three times with sterile water. Pieces were left to dry under hood flow and placed on half-strength potato dextrose agar (PDA) amended with streptomycin stock solution (Sigma-Aldrich, St. Louis, MO, USA) to inhibit bacterial growth.
Author Contributions
Conceptualization, T.D., K.V. and J.C.; methodology, T.D. and K.V.; software, T.D. and K.V.; validation, M.M.K., J.C., T.S. and A.S.; investigation, T.D., T.S. and K.V.; resources, T.D. and K.V.; data curation, K.V.; writing— T.D. and K.V.; writing—review and editing, T.D., M.M.K. and A.S.; visualization, T.D.; supervision, A.S. and J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key cultivation techniques for hemp in Europe and China. Ind. Crops and Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Schultes, R.E.; Klein, W.M.; Plowman, T.; Lockwood, T.E. Cannabis: an example of taxonomic neglect. Bot. Mus. Leaflets. 1974, 23, 337–367. [Google Scholar] [CrossRef]
- McPartland, J.M.; Guy, G. The evolution of Cannabis and coevolution with the cannabinoid receptor–a hypothesis. In The medicinal use of cannabis; Guy, G., Robson, R., Strong, K., Whittle, B., Eds.; Royal Society of Pharmacists: London, 2004; pp. 71–102. [Google Scholar]
- Thamae, T.; Aghedo, S.; Baillie, C.; Matovic, D. Tensile Properties of Hemp and Agave Americana Fibres. Handb. Tensile Prop. Text. Tech. Fibres. 2009, 73–99. [Google Scholar]
- Salentijn, E.M.J.; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L.M. New Developments in Fiber Hemp (Cannabis Sativa, L.) Breeding. Ind. Crops Prod. 2015, 68, 32–41. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.F.; Guerriero, G. Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef]
- Vonapartis, E.; Aubin, M.P.; Seguin, P.; Mustafa, A.F.; Charron, J.B. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Composit. Anal. 2015, 39, 8–12. [Google Scholar] [CrossRef]
- Callaway, J.C. Hempseed as a nutritional resource: An overview. Euphytica. 2004, 140, 65–72. [Google Scholar] [CrossRef]
- Struik, P.C.; Amaducci, S.; Bullard, M.J.; Stutterheim, N.C.; Venturi, G.; Cromack, H.T.H. Agronomy of fibre hemp (Cannabis sativa L. ) in Europe. Ind. Crops Prod. 2000, 11, 107–118. [Google Scholar] [CrossRef]
- Manaia, J.P.; Manaia, A.T.; Rodriges, L. Industrial Hemp Fibers: An Overview. Fibers. 2019, 7, 106. [Google Scholar] [CrossRef]
- Ranalli, P.; Venturi, G. Hemp as a raw material for industrial applications. Euphytica. 2004, 149, 1–6. [Google Scholar] [CrossRef]
- Matthäus, B.; Brühl, L. Virgin hemp seed oil: An interesting niche product. Eur. J. Lip. Sci. Technol. 2008, 110, 655–661. [Google Scholar] [CrossRef]
- Rupasinghe, V.H.P.; Davis, A.; Kumar, S.K.; Murray, B.; Zheljazkov, V.Z. Industrial hemp (Cannabis sativa subsp. sativa) as an emerging source for value-added functional food ingredients and nutraceuticals. Molecules. 2020, 25, 4078. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K. Epidemiology of Fusarium oxysporum causing root and crown rot of cannabis (Cannabis sativa L., marijuana) plants in commercial greenhouse production. Can. J. Plant Pathol. 2021, 43, 216–235. [Google Scholar] [CrossRef]
- Agrios, G. N. Plant Pathology, 5th ed.; Agrios, G. N., Ed.; Elsevier Academic Press: Burlington, MA, 2005. [Google Scholar]
- Ivić, D.; Domijan, A.M.; Peraica, M.; Miličević, T.; Cvjetković, B. Fusarium spp. on wheat, maize, soybean and pea. Arh Hig Rada Toksikol. 2009, 60, 435–442. [Google Scholar] [CrossRef]
- Duvnjak, T.; Sudaric, A.; Matosa Kocar, M.; Cosic, J.; Vrandecic, K. First report of soybean fusarium wilt caused by Fusarium oxysporum in Croatia. Plan. Dis. 2017, 101, 249. [Google Scholar] [CrossRef]
- Gwinn, K.D.; Hansen, Z.; Kelly, H.; Ownley, B.H. Diseases of Cannabis sativa caused by diverse Fusarium species. Front. Agron. 2022, 3, 796062. [Google Scholar] [CrossRef]
- Patschke, K.; Gottwald, R.; Muller, R. Erste Ergebnisse phytopathologischer Beobachtungen im Hanfanbau im Land Brandenburg. Nach. des Deut. Pflan. 1997, 49, 296–290. [Google Scholar]
- Punja, Z. K.; Collyer; Scott, C.; Lung, S.; Holmes, J.; Sutton, D. Pathogens and molds affecting production and quality of Cannabis sativa L. Front. Plant Sci. 2019, 10, 1120. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N. “American weed: a history of Cannabis cultivation in the United States”. EchoGéo. 2019, 48, 17650. [Google Scholar] [CrossRef]
- McCain, A. H.; Noviello, C. “Biological control of Cannabis sativa,” in Proceedings of the VI International Symposium on Biological Control of Weeds (Vancouver, BC). ed E. S. Delfosse. 1985, 635–642. [Google Scholar]
- Noviello, C.; McCain, A. H.; Aloj, B.; Scalcione, M.; Marziano, F. Lotta biologica contro Cannabis sativa medlante l'impiego di Fusarium oxysporum f. sp. cannabis. Annali delta Facolta di Scienze Agrarie della Universita degli Studi di Napoli. Portici. 1990, 24, 33–44. [Google Scholar]
- Rataj, K. Skodivi cinitele pradnych rostlin. Pram. lit. 1957, 2, 1–123. [Google Scholar]
- McPartland, J.M.; Clarke, R.C.; Watson, D.P. Fungal Diseases. In Hemp Diseases and Pests, Management and Biological Control; McPartland, J.M., Clarke, R.C., Watson, D.P., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 93–135. [Google Scholar]
- McPartland, J.M.; Hillig, K.W. Cannabis clinic Fusarium Wilt, J. of Ind. Hemp. 2004, 9, 2–67. [Google Scholar] [CrossRef]
- Noviello, C.; Snyder, W.C. Fusarium wilt of hemp. Phytopathology 1962, 52, 1315–1317. [Google Scholar]
- Summerell, B. A.; Salleh, B. .; Leslie, J. F. A utilitarian approach to Fusarium identification. Plant Dis. 2003, 87, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Huber, D. M.; Thompson, I. A. Nitrogen and plant disease. In Mineral Nutrition and Plant Disease; Datnoff, L. E., Elmer, W. H., Huber, D. M., Eds.; The American Phytopathological Society: St. Paul, MN, 2009; pp. 31–44. [Google Scholar]
- Lombard, L.; Sandoval-Denis, S. C.; Crous, P. W. Epitypification of Fusarium oxysporum – clearing the taxonomic chaos. Persoonia 2019, 43, 1–47. [Google Scholar] [CrossRef]
- Bruehl, G. W. Soilborne Plant Pathogens; Macmillan Pub. Co.: New York, NY, 1987. [Google Scholar]
- Tiourebaev, KS.; Pilgeram, A.L.; Anderson, T.W.; Sands, D.C. Soil penetration of a mycoherbicide facilitated by carrier seedlings. Phytopathology. 1997, 87 (6 supplement), S:57. [Google Scholar]
- Agostinelli, A.M.; Clark, A.J.; Brown-Guedira, G.; Van Sanford, D.A. Optimizing phenotypic and genotipis selection for Fusatium head blight resistance in wheat. Euphytica 2012, 186, 115–126. [Google Scholar] [CrossRef]
- O’Donnell, K.; Gueidan, C.; Sink, S.; Johnston, P.R.; Crous, P.W.; Glenn, A.; Riley, R.; Zitomer, N.C.; Colyer, P.; Waalwijk, C.; van der Lee, T.; Moretti, A.; Kang, S.; Kim, H.S.; Geiser, D.M.; Juba, J.H.; Baayen, R.P.; Cromey, M.G.; Sean Bithell, S.; Sutton, D.A.; Skovgaard, K.; Ploetz, R.; Kistler, H.C.; Elliott, M.; Davis, M.; Sarver, B.A.J. A two-locus DNA sequence database for typing plant and human pathogens within the Fusarium oxysporum species complex. Fun. Gen. and Biol. 2009, 46, 936–948. [Google Scholar] [CrossRef]
- Ilić, J.; Ćosić, J.; Jurković, D.; Vrandečić, K. Vegetative compatibility of Fusarium oxysporum isolated from weeds in eastern croatia. Poljoprivreda 2013, 19, 20–24. [Google Scholar]
- European Commission EU Plant Variety Database (v.3.2). Available online: https://ec.europa.eu/food/plant/plant_propagation_material/plant_variety_catalogues_databases/search/public/index.cfm?event=SearchVariety&ctl_type=A&species_id=240&variety_name=&listed_in=0&show_current=on&show_deleted= (accessed on 15 March 2020).
- Pellegrini, M.; Ercole, C.; Gianchino, C.; Bernardi, M.; Pace, L.; Del Gallo, M. Fusarium oxysporum f. sp. cannabis isolated from Cannabis sativa L.: In vitro and in planta biocontrol by a plant growth promoting-bacteria consortium. Plants. 2021, 10, 2436. [Google Scholar] [CrossRef] [PubMed]
- Herrer, J. The Emperor Wears No Clothes: The Authoritative Historical Record of Cannabis Conspiracy Against Marijuana; Publishers Group West: New Jersey, 1998; pp. 1–181. [Google Scholar]
- Mihelčić, D. Primjena konoplje u poljoprivredi i tržišne perspektive njezinih prerađevina i proizvoda.Master's thesis / Diplomski rad, 2017; Josip Juraj Strossmayer University of Osijek, Faculty of agriculture / Permanent link https://urn.nsk.hr/urn:nbn:hr:151:098050.
- The Government of the Republic of Croatia https://vlada.gov.hr/UserDocsImages//2016/Sjednice/2019/O%C5%BEujak/148%20sjednica%20VRH//148 %20-%2016.pdf 27 April 2022. 27 April.
- Paying Agency for Agriculture, Fisheries and Rural Development (PAAFRD). Available online: www.apprrr.hr/agronet (accessed on 14 March 2022).
- Leslie, J. L.; Summerell, B. A. The Fusarium Laboratory Manual; Blackwell Publishing Professional: Hoboken, NJ, 2006; p. 212. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, 1990; p. 315. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R. C. Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Sciences, 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, I.; Edel-Hermann, V.; Gautheron, N.; Durling, M.B.; Kolseth, A.K.; Steinberg, C.; Persson, P.; Friberg, H. Genus-Specific Primers for Study of Fusarium Communities in Field Samples. Appl. Environ. Microbiol. 2015, 82, 491–501. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. and Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Fredrik Ronquist, R.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–10. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v.1.4.4., 2018.
- Matić, M.; Vuković, R.; Vrandečić, K.; Štolfa Čamagajevac, I.; Ćosić, J.; Vuković, A.; Sabljić, K.; Sabo, N.; Dvojković, K.; Novoselović, D. Oxidative status and antioxidative response to Fusarium attack and different nitrogen levels in winter wheat varieties. Plants 2021, 10, 611. [Google Scholar] [CrossRef]
Figure 1.
Fusarium wilt of hemp caused by Fusarium oxysporum. (a) Leaf symptoms of F. oxysporum on hemp in the field; (b) Root symptoms.
Figure 1.
Fusarium wilt of hemp caused by Fusarium oxysporum. (a) Leaf symptoms of F. oxysporum on hemp in the field; (b) Root symptoms.

Figure 2.
(a) 12-day-old colony of F. oxysporum on Potato Dextrose Agar (PDA), front and back; (b) microconidia of F. oxysporum;.
Figure 2.
(a) 12-day-old colony of F. oxysporum on Potato Dextrose Agar (PDA), front and back; (b) microconidia of F. oxysporum;.
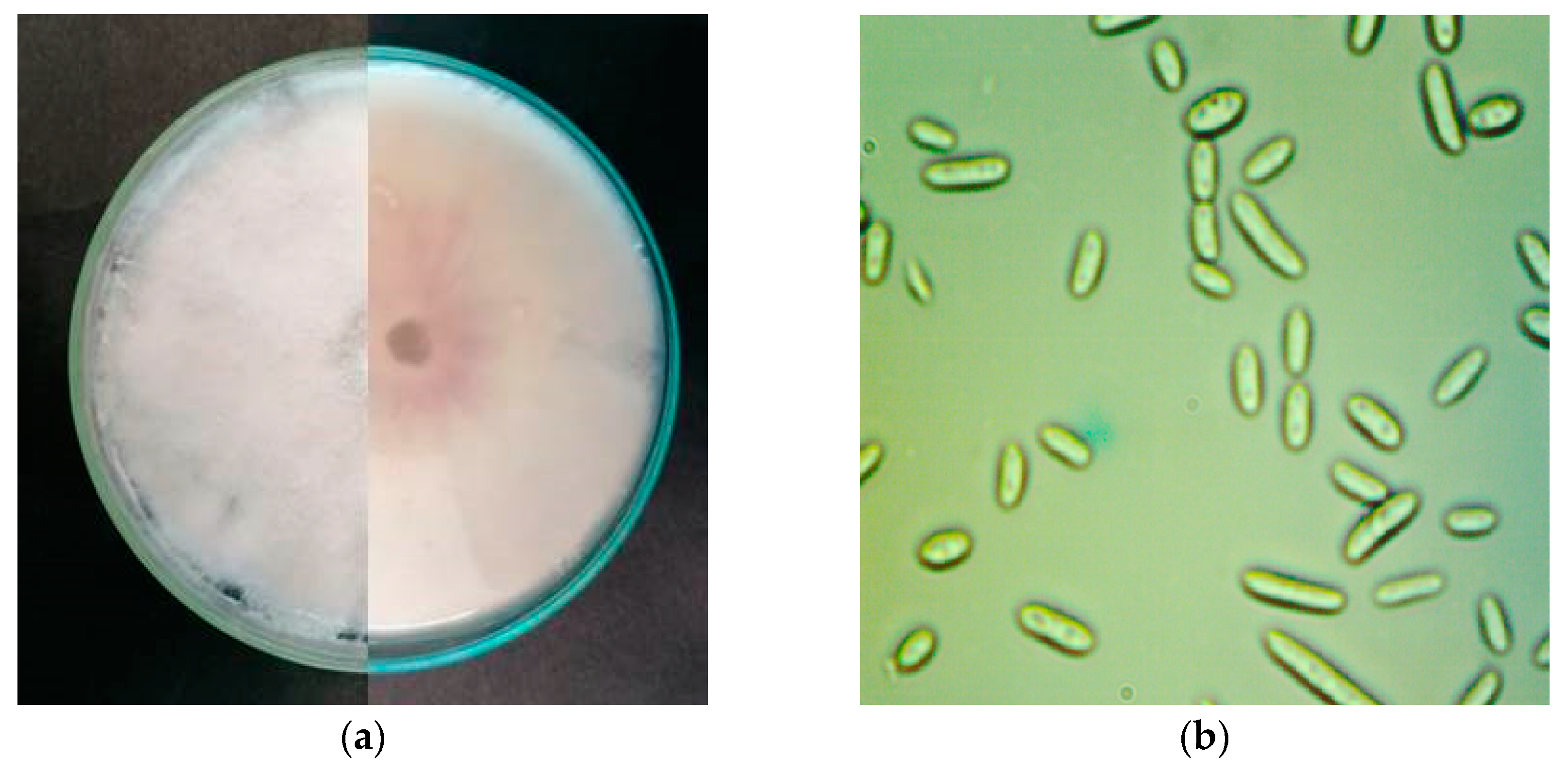
Figure 3.
Phylogenetic tree inferred from Bayesian analysis on sequences of the internal transcribed spacer (ITS) region of 42 representative species of Fusarium. Bayesian posterior probabilities are indicated beside nodes. Fusarium staphyleae (NRRL22316) obtained from GenBank was treated as the outgroup.
Figure 3.
Phylogenetic tree inferred from Bayesian analysis on sequences of the internal transcribed spacer (ITS) region of 42 representative species of Fusarium. Bayesian posterior probabilities are indicated beside nodes. Fusarium staphyleae (NRRL22316) obtained from GenBank was treated as the outgroup.
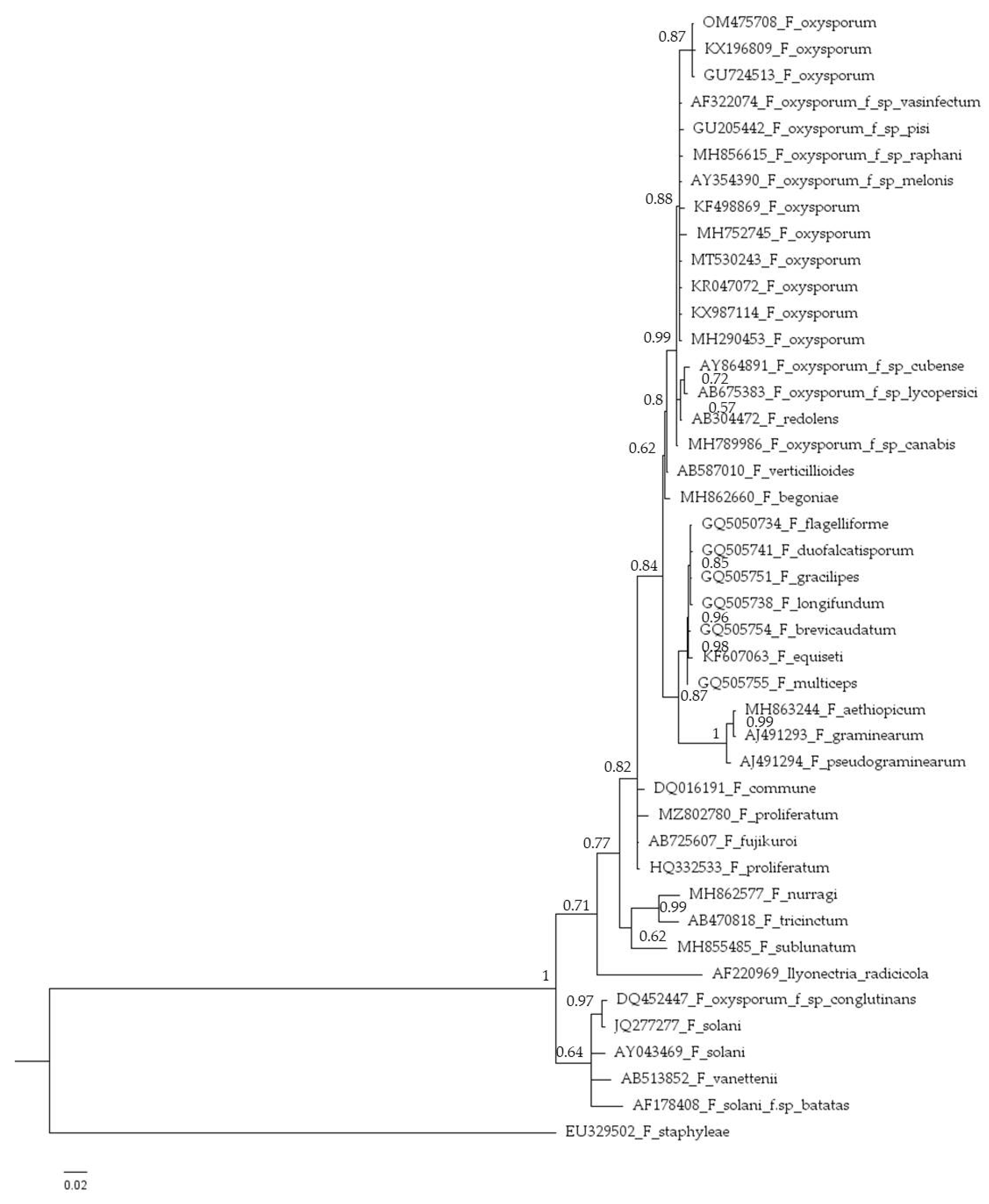
Figure 4.
Phylogenetic tree inferred from Bayesian analysis on sequences of the translation elongation factor 1-α (TEF1-α) region of 36 representative species of Fusarium. Bayesian posterior probabilities are indicated beside nodes. Fusarium staphyleae (NRRL22316) obtained from GenBank was treated as the outgroup.
Figure 4.
Phylogenetic tree inferred from Bayesian analysis on sequences of the translation elongation factor 1-α (TEF1-α) region of 36 representative species of Fusarium. Bayesian posterior probabilities are indicated beside nodes. Fusarium staphyleae (NRRL22316) obtained from GenBank was treated as the outgroup.

Figure 5.
Symptoms of F. oxysporum artificial infection in laboratorial conditions (a) Wilt symptoms begin with an upward curing of leaf tips; (b) Wilted leaves dry to a yellow-tan color and hang on the plant without falling off; (c) Control plants.
Figure 5.
Symptoms of F. oxysporum artificial infection in laboratorial conditions (a) Wilt symptoms begin with an upward curing of leaf tips; (b) Wilted leaves dry to a yellow-tan color and hang on the plant without falling off; (c) Control plants.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



