
Submitted:
04 August 2023
Posted:
07 August 2023
You are already at the latest version
Abstract
Drought stress imposes substantial constraints on the growth and production of wheat (Triticum aestivum L.), a globally important cereal crop that is essential for food security. To mitigate these adverse effects, researchers are intensifying their efforts to comprehend how different genotypes respond to drought stress, aiding in the development of sustainable breeding and management strategies. This review summarized past and recent research on genotype-dependent responses of wheat to drought stress, encompassing morphological, physiological, biochemical, molecular, genetic, and epigenetic reactions in plants. Screening of drought-affected features at early developmental stages can provide valuable insights into the adult growth stages that are closely linked to plant productivity. The review underscores the importance of identifying key traits associated with drought resistance, and the potential of leveraging wheat diversity to select cultivars with desirable agronomic characteristics. It also highlights recent advancements in investigating Bulgarian wheat genotypes with varying levels of drought tolerance, specifically in detecting essential features contributing to drought tolerance. Cultivating drought-resistant wheat genotypes and understanding stress stability determinants could markedly contribute to enhancing wheat production and ensuring stable yields under changing climate conditions.
Keywords:
drought stress
; wheat genotypes
; drought-resistant cultivars
; genotype-dependent responses
; wheat diversity
1. Introduction
The increasing global population and intensifying weather extremes present a significant threat to global food security and sustainability. Among the many environmental challenges, water shortage is a major constraint that affects all aspects of plant growth and development, and agricultural productivity [1,2]. Drought can occur across all of Europe in any season, irrespective of whether the regions experience high or low rainfall. The impacts of drought become particularly pronounced in areas with limited water resources or inadequate management practices, leading to an imbalance between water demands and water supply. In recent decades, the frequency and severity of drought events have increased, particularly in Central and Southern Europe including Bulgaria [3,4,5]. The increase in summer temperatures by up to 2.4 °C in combination with dry spells lead to prolonged drought periods, which worsen the agrometeorological conditions in the initial stages of the development of winter crops, such as wheat [6]. Improving our understanding of the complex mechanisms behind plant drought responses and survival requires extensive research efforts because drought outcomes are affected by various external and internal factors, such as duration and severity of water loss [7], the age and stage of development during drought exposure [8], and the affected organ and cell types [9]. Collaborative endeavors involving scientists, breeders, and farmers over the next decade will have a pivotal role in identifying and comprehending traits linked to drought resistance in plants, and breeding drought-resilient crop cultivars [10].
Wheat (Triticum aestivum L.) is a highly cultivated and widely consumed food crop that provides more proteins than any other cereal crop. In the span from 2018 to 2021, wheat grain yields have ranged between 732.1 and 760.9 million tons per year [11]. However, this high production is not sufficient to prevent a depletion of global inventories due to dry weather spells and other unpredictable climate events that can lead to crop losses. Compared to other crops like rice and soybean, wheat displays higher sensitivity to water supply anomalies, resulting in reduced productivity [12]. Water scarcity can affect wheat at all developmental stages: germination, early seedling growth, flowering, and grain filling [13]. Wheat plants rely mainly on stored soil moisture [14], which becomes depleted during prolonged dry periods and replenished from rainwater [15]. Enhancing water uptake and water use efficiency (WUE) through breeding can provide higher yields and better crop quality [16,17,18]. Thus, in light of the growing global population and the ongoing climate fluctuations, it is crucial to continue research efforts aimed at improving the drought resistance of wheat.
2. Plant Defense Strategies against Drought
Plants have evolved various defense mechanisms to withstand environmental variations, such as stress escape, stress avoidance, and stress tolerance (Figure 1). These mechanisms can operate independently of adverse conditions or be specifically adapted to a particular stressor and facilitate a heritable plastic response [19]. By leveraging the stress adaptive strategies, plants can better cope with extreme weather and improve their chances of survival and successful reproduction.
2.1. Drought Escape Strategy
Drought escape is a survival mechanism that allows plants to successfully undergo their entire life cycle before experiencing drought-induced stress, entering a state of dormancy during periods of favourable weather conditions [20]. Drought-escaping plants typically do not undergo special morphological, physiological, or biochemical modifications. Instead, developmental plasticity is the key mechanism that enables these plants to escape from drought. Plant plasticity is associated with varying the duration of the transition from vegetative to reproductive stages, and fast phenological development, including early flowering and early maturation [21].
To achieve rapid plant development necessary for drought escape, several variables come into play, such as decreased stomatal conductance, low transpiration, and high photosynthetic carbon gain [22]. These factors promote metabolic activity, and lead to increased cell division and expansion, and the formation of new seeds before the drought-induced end of the plant life cycle [23]. Drought escape strategy is widely employed by natural plants [24] and also by crops like wheat [20], rice [25], and Brassica rapa [22]. This strategy can be highly effective in coping with short or recurrent drought periods, but it may not be as efficient under prolonged water shortages [20]. This is because the strategy relies on plants completing their life cycle before the onset of drought stress, which can be challenging during prolonged drought periods.
Early flowering is a critical aspect of drought escape as it shortens the vegetative development phase but it can potentially decrease crop yield. Therefore, the timing of the crop life cycle, primarily determined by the flowering time, is essential for yield production in water-limited environments. Under normal conditions, a longer crop cycle generally promotes productivity by allowing extended photosynthetic seasons and greater opportunities to harness solar energy [26]. Severe terminal drought can reduce yields due to depleted soil water before the end of the crop cycle [27]. For instance, wheat genotypes selected in Mediterranean environments, where terminal drought stress frequently occurs, show substantially lower production due to the higher risk of water stress during reproductive or grain filling stages [28].
2.2. Drought Avoidance Strategy
Drought avoidance, also known as the ‘succulent strategy’, is a plant adaptation that enhances WUE in dry environments. This strategy can be achieved through two main mechanisms: efficient water uptake by the plant root system, and reduced water loss from the shoot parts [21]. Values of WUE may vary depending on plant species, and duration and severity of drought. A higher WUE is generally linked to increased yields under water shortage [29]. Drought avoidance strategy is typically connected with limited vegetative growth and involves specific plant adaptations, such as maintaining low metabolic rates, small or closed stomata to reduce water loss, decreased transpiration and photosynthesis rates, and morphological adjustments, like leaf curling and increased wax deposition on the leaf surface [30].
Plants that use drought avoidance strategy can be broadly classified into two categories: water savers and water spenders [31]. Water savers limit water loss from the plant canopy by developing thicker cuticles, closing stomata, decreasing transpiration area and radiation absorption, and conserving water in specialized tissues for later use during grain filling and yield formation. In contrast, water spenders achieve high tissue water status by maintaining water uptake through increased rooting and enhanced hydraulic conductance [32,33].
The drought avoidance strategy is observed in natural herbaceous populations [24] as well as in cultivated crops [34,35]. Certain wheat cultivars possess specific traits, such as thicker cuticles, reduced stomatal density, and deep root systems that allow them to minimize water loss through transpiration and survive in drought conditions [36,37]. However, the success of this strategy depends on the interplay between plant adaptations, soil moisture conditions, and environmental factors. Although drought avoidance could be effective in conserving water, it should be considered that limited vegetative growth may potentially reduce crop productivity.
2.3. Drought Tolerance Strategy
Drought tolerance is a complex phenomenon that enables plants to survive drought exposure by employing various physiological, biochemical, or morphological adaptations that prevent or mitigate the harmful effects of dehydration [38]. This defense strategy involves drought-induced responses at multiple levels, ranging from the molecular to whole-plant levels, such as the regulation of gene expression, protein turnover, DNA or protein repair, the maintenance of tissue turgor through osmotic adjustment, changes in cell wall extensibility, enhancement of protoplasmic drought resistance, the preservation of redox homeostasis [27,39].
One central mechanism for drought tolerance is osmotic adjustment that can be achieved through the accumulation of soluble molecules, known as osmoprotectants (proline, polyamines, ammonium compounds, soluble sugars, and ions) that reduce cell dehydration [40]. The accumulation of osmoprotectants leads to a decrease in the inner osmotic potential of the cells, which enhances water retention and maintains homeostasis, thus ensuring proper plant functioning under drought stress [41].
Water scarcity promotes oxidative stress in plants due to the excessive production of Reactive Oxygen Species (ROS) that often cause cellular damage. Plant drought tolerance depends on the mobilization of endogenous antioxidant systems, which can alleviate the damaging effects [42]. The defense systems include enzymatic components, such as catalase (CAT), peroxidase (POX), superoxide dismutase (SOD), ascorbate peroxidase (APX), and glutathione reductase (GR), as well as nonenzymatic antioxidants like flavonoids, ascorbic acid, α-tocopherol, reduced glutathione, and β-carotene [43]. Modulation of antioxidant defense systems represents a major strategy for enhancing drought tolerance. Plant species resistant to dehydration generally contain higher concentrations of osmoprotectants and possess a more robust antioxidant system [42,44].
Drought-tolerant wheat cultivars have the capacity to sustain productivity even in water-limited conditions by employing various mechanisms, such as increased root growth, osmotic adjustment, and improved WUE to achieve drought tolerance [27,45,46], ensuring stable crop yield despite water scarcity. Selecting drought-tolerant wheat genotypes is not only economically viable but also a sustainable approach to supporting wheat production in drought-affected regions. To facilitate the selection of such genotypes, it is essential to identify relevant plant features that confer advantages under drought stress. Screening for such traits, particularly during the early stages of wheat development, can provide valuable hints for the subsequent growth stages linked to plant productivity.
Hence, wheat plants can employ a combination of drought escape, drought avoidance, and drought tolerance strategies. The choice of the most suitable strategy relies on the specific genetic makeup of the wheat cultivar and the prevailing environmental conditions. Besides, the categorization of plants into drought escape or drought avoidance types is not entirely precise in practice because they occur at different times. Drought escape happens prior to a drought event, while drought avoidance occurs during or after a drought period. Researchers and breeders should actively pursue the development of wheat cultivars with a combination of strategies to increase their resilience to drought stress in changing environments.
3. Genotype-Dependent Morphological Responses
Figure 2 presents the genotype-specific effects of drought stress on the root and shoot morphology of wheat plants.
3.1. Root System Traits
The ability of plants to thrive in arid environments depends on root system traits (Figure 2), vital to absorption of soil water and nutrients under favourable and extreme conditions [47]. Monocots, such as wheat, possess a complex root system that comprises seminal roots originating from the embryo of germinating seeds, nodal roots developing from the base of the tiller, and fine roots, like lateral roots and root hairs that further enhance the root absorption capability [48,49]. Seminal roots possess the ability to grow and absorb water from deeper soil layers, providing an advantage in terms of water absorption [50]. A greater root density and deeper root distribution can enhance water acquisition from lower soil layers [51,52,53], and lead to increased grain yield even under conditions of terminal drought stress [54]. Comparative analysis of two wheat genotypes with differing root depth has shown that the genotype with an extensive root system generally provides higher yields in drought-prone environments, possibly due to improved water supply during grain filling stage [55].
A deeper root system is often related with a higher rate of root elongation [45,52] and/or narrower root angles [56,57]. Narrow root growth angles facilitate vertical growth, allowing roots to penetrate deeper into the soil and access water and nutrients during drought periods [58]. In support of this, Fang et al. [59] observed that the recently developed cultivar CH1 exhibits accelerated root growth, enabling improved water uptake from subsoil layers after anthesis, which substantially contributes to achieving high grain yield under drought conditions. A cluster analysis of 29 wheat cultivars based on their seminal root traits revealed that cultivars better adapted to drought conditions tend to have a narrower angle of seminal axes resulting in higher yields, which is attributed to the enhanced plant ability to access soil water [55]. On the other hand, after evaluating six genotypes with varying drought susceptibility index, Grzesiak et al. [60] have shown that a more downward orientation of root growth can be beneficial, but not mandatory for mitigating drought impacts on tolerant cultivars. McDonald [61] did not find a direct correlation between root angles and higher yield. Nevertheless, wheat genotypes with narrower root angles demonstrate a tendency to produce slightly higher yield.
Xylem vessel diameter [62] and root-to-shoot ratio [59] are other genotype-dependent traits related to wheat aridity resistance and productivity. Drought-adapted wheat cultivars utilize the plasticity of wheat root stele and xylem number/diameter features to improve WUE [63]. Narrower xylem vessels have been successfully employed in breeding programs for enhancing wheat drought tolerance and developing advanced wheat lines [64]. These lines have a decreased size and higher number of metaxylem vessels at the root-shoot junction, and the opposite pattern near the root tips. Such a configuration facilitates efficient utilization of soil water resources.
Adaptive phenotypic plasticity of the root system, changing the root-to-shoot ratio can mitigate the negative impacts of drought stress [65]. In water-limited environments, the growth of shoot plant parts is constrained, resulting in a shift in the root-to-shoot ratio. This shift reduces the evaporation area of shoots compared to the absorptive area of roots, thus improving WUE [66]. However, some researchers argue that having a larger root biomass may not necessarily be advantageous for drought adaptation, as it can increase the risk of depleting available soil water before completing grain filling [54].
Although the relationship between belowground characteristics and drought response is complex, the root system undeniably plays a critical role in the plant capacity to absorb soil water and nutrients, thereby influencing crop productivity and adaptation to harsh environmental conditions. Further research exploring the specific mechanisms by which root traits of various wheat genotypes impact drought resistance could provide valuable insights for optimizing agricultural practices and enhancing plant resilience.
3.2. Aboveground Traits
Under drought stress, wheat plants notably change their aboveground traits (Figure 2), encompassing plant height, coleoptile length, leaf size, shape and perimeter, leaf surface morphology and waxiness, and various anatomical adjustments [67,68,69,70]. These variations are contingent upon the drought susceptibility of the genotypes, the duration and severity of drought events, as well as the growth stage of the plants [13,60,71,72].
Typically, drought stress causes a reduction of leaf area, which can subsequently decrease the photosynthetic capacity of the leaves [73]. Modern semi-dwarf wheat varieties maintain better water balance even under severe drought, as compared to older tall bread wheat varieties. These variations can be partly ascribed to a smaller leaf size and more rounded leaf shape, which contribute to enhanced drought tolerance by reducing evaporation and preserving the integrity of cell membranes [69]. In contrast, when comparing two wheat genotypes with differing levels of drought resistance under moderate dehydration, the drought-tolerant cultivar accumulates higher shoot dry biomass, has larger flag leaf size and water content, and a higher harvest index and WUE, compared to the susceptible genotype [74]. Micromorphological features of leaves, such as stomatal and trichome density and size, as well as leaf waxiness, are also considered important traits related to drought tolerance and adaptation [69,75]. Screening five wheat cultivars, David et al. [76] have revealed substantial variations in the number of trichomes and stomata among the cultivars. Interestingly, the two genotypes with higher drought tolerance exhibited a greater abundance of trichomes on both the upper (adaxial) and lower (abaxial) leaf surfaces, and a lower number of stomata. During drought exposure at anthesis and early kernel development, Jäger et al. [77] identified two drought-tolerant wheat cultivars using yield parameters, RWC in flag leaves, and leaf micromorphological features. The tolerant cultivars displayed a reduced occurrence of stomata on the flag leaf surfaces along with larger stomatal guard cells and high membrane integrity. In contrast, the drought-susceptible cultivars exhibited lower membrane integrity and a higher number of stomata on both leaf surfaces.
Additionally, under dehydration, the length of coleoptiles has notable variations among wheat cultivars [68] with longer coleoptiles that enhance the frequency of seedling emergence during desiccation, and potentially resulting in higher yields [78,79]. Conversely, shorter coleoptiles can negatively impact plant growth and productivity under dry conditions [68,80].
Utilizing these simple belowground and aboveground traits as selection criteria for drought tolerance holds promise for supporting breeding efforts aimed at developing drought-resistant wheat genotypes. By focusing on these traits, breeders can effectively identify and select genotypes that are more likely to withstand water scarcity; however, to achieve optimal results, it is important to conduct comprehensive research that considers not only the individual traits but also their interactions with one another.
4. Genotype-Dependent Physiological and Biochemical Responses
Drought stress commonly triggers various physiological and biochemical responses in wheat plants (Figure 3), such as changes in relative water content (RWC), leaf gas exchange rates, production of ROS, oxidative stress, peroxidation of membrane lipids, accumulation of osmolytes, and induction of leaf senescence [81,82]. Certain cultivars can sustain their biological functions under conditions of limited water supply and rapidly recover upon restoration of normal watering [46,83,84,85]. Relative water content serves as a vital indicator of plant water status and susceptibility to drought, since it closely correlates with moisture levels in the soil and atmosphere. The extent of drought-induced reduction in RWC typically depends on the stress tolerance level of a specific wheat genotype [84,85]. Under severe water stress, drought-sensitive Bulgarian wheat cultivars experience a more significant decline in leaf water potential, while the drought-tolerant Katya cultivar sustains a higher water potential [46]. After the rewatering phase, Katya water potential returns to its original control values as early as the first day, whereas the drought-sensitive cultivars take until the third day of recovery to approach similar values. Furthermore, Katya exhibits nearly complete recovery in leaf gas exchange and photosynthetic activity, which could be attributed to its more dynamic stomatal responses.
Several parameters have been proposed for the selection of drought-tolerant wheat germplasm, such as stomatal conductance [86], water retention capability [87], leaf chlorophyll content, fluorescence induction parameters in leaves at grain filling stage [88,89], and leaf organelle ultrastructure [90]. Drought stress greatly impacts water dynamics, hampering inherent ability of plants to retain water effectively, primarily through stomatal and cuticular transpiration, which regulate water loss via leaves [91,92]. Drought-tolerant plants have evolved mechanisms to balance stomatal and cuticular transpiration, allowing them to conserve water and mitigate the negative impacts of water scarcity. Cultivation of three elite wheat cultivars in a dry environment has revealed that the tolerant cultivar has lower stomatal conductance, reduced intercellular CO2 concentration, transpiration and photosynthetic rates, but higher WUE, compared to the other two genotypes [93]. The tolerant cultivar has also exhibited significantly greater grain yield, emphasizing the important role of leaf gas exchange characteristics in wheat productivity.
Another potential drought-induced limitation to photosynthesis could be attributed to alterations in Rubisco (Ribulose-1,5-bisphosphate carboxylase/oxygenase) activity, the enzyme responsible for CO2 assimilation during photosynthesis, as well as in Rubisco activase, a molecular chaperone essential for the light activation of Rubisco. In field studies, relatively stable or elevated levels of Rubisco large subunit, Rubisco small subunit, and Rubisco activase are associated with drought tolerance and higher productivity. Drought-tolerant cultivars have the ability to achieve greater yields even under severe drought conditions [84,94,95].
Water shortage often leads to an increase in the production of ROS through the transfer of photosynthetic electrons towards molecular O2 [83], which subsequently increase lipid peroxidation [96]. The relationship between leaf gas exchange parameters, RWC, malondialdehyde (MDA) content, and drought tolerance displays genotype-dependent changes among six winter wheat cultivars. However, internal CO2 concentration and MDA content did not consistently correlate with drought tolerance across the genotypes [97]. In general, MDA, a byproduct of lipid peroxidation, is used as an indicator of membrane damage and wheat drought tolerance [98]. While some studies did not find a clear correlation between drought tolerance and MDA accumulation in wheat cultivars [97,99], others reported that genotypes with lower MDA content under dehydration are more drought-resistant [83,100].
As aforementioned, plants have developed enzymatic and non-enzymatic mechanisms to protect themselves from stress-induced ROS accumulation [101]. The activities of antioxidant enzymes, such as SOD and CAT, have been suggested as selection criteria for drought resistance and productivity in wheat varieties [102]. A drought-tolerant genotype with high activity of POX and CAT, as well as with high levels of ascorbic acid, exhibit lower accumulation of hydrogen peroxide and lipid peroxidation compared to a sensitive genotype [103]. Fan et al. [104] categorized the drought adaptation strategies of old and modern wheat varieties into different defense phases, based on drought severity. The second and third phases are connected with increased levels of ROS and greater activity of antioxidant enzymes, resulting in membrane lipid peroxidation. Modern wheat varieties possess a greater capacity to resist drought than the old variety, which is partially related to different antioxidant capacity. Genotype-dependent differences in the activities of APX and GR, as well as non-enzymatic protection responses such as the accumulation of reducing sugars and sucrose, have been identified in two winter wheat cultivars with contrasting productivity [99].
Wheat plants employ a range of osmoprotectants as a means of defense to bolster their resistance against drought [41,105,106]. Proline and soluble sugars are the primary molecules that contribute to enhancing wheat drought tolerance [107]. In a study assessing 25 bread wheat genotypes, four showed improved growth under dehydration due to the accumulation of free proline and soluble phenolics, which improves tissue water status and photosynthetic capacity [108]. Drought-tolerant genotypes accumulate higher levels of proline in the leaves, which partially mitigates the detrimental drought effects [109]. While glycine betaine does not directly scavenge ROS [110], its excessive accumulation can activate the antioxidant defense system in transgenic wheat plants [111]. Moreover, under drought stress tolerant cultivars typically deposit more soluble sugars like sucrose than susceptible cultivars [105,112,113]. Khoshro et al. [114] have also reported that drought sensitive wheat cultivars accumulate significantly lower levels of soluble sugars, proline, glycine betaine, and potassium compared to tolerant cultivars. Additionally, dehydration tolerance in wheat is linked to a higher proportion of raffinose, which serves as a protective molecule, aiding in the stabilization of cellular structures and preventing damages caused by dehydration [115].
Hence, the extensive array of physiological, biochemical, and metabolite responses during dehydration demonstrates the existence of diverse mechanisms employed by various wheat genotypes to endure drought conditions. As dehydration tolerance is a multifaceted adaptation that is controlled at various levels of plant organization and varies depending on the developmental stage of the plant, the identification of a comprehensive set of independent and interrelated physiological and biochemical traits can serve as direct and indirect selection criteria for wheat drought tolerance.
5. Genotype-Dependent Molecular Responses
Understanding the intricate molecular mechanisms that underlie wheat responses to dry environments is essential for developing wheat genotypes with enhanced drought resistance [116,117]. Comparative transcriptomic analysis of drought-induced gene expression in wheat cultivars with varying stress tolerance revealed that differentially expressed genes (DEGs) are mainly linked to carbon metabolism, flavonoid biosynthesis, and phytohormone signaling (Figure 3) [116,118,119]. Drought-tolerant winter wheat cultivar exposed to water stress during various growth stages has shown that DEGs are associated with floral development, photosynthetic activity, and stomatal movement [120]. Study of root transcriptomes in two wheat genotypes with contrasting root growth under drought stress reveals the downregulation of hormone pathway genes in both genotypes. However, the tolerant genotype demonstrates the upregulation of genes related to ROS metabolism, chaperones, transcription factors, and cell wall synthesis, enabling more effective drought adaptation [121].
Another transcriptome profiling during the early seedling stage of wheat under water deficit has shown that a drought-susceptible genotype has numerous genotype-dependent DEGs with predominantly upregulated metabolism-related genes, such as those involved in carbohydrate, lipid, amino acid, and terpenoid pathways. This implies the presence of an energy-intensive adaptation strategy to cope with water deficit [122]. Conversely, in the tolerant genotype, a notable downregulation of metabolism-related genes, including those, implicated in secondary metabolite synthesis, indicates an energy-saving response to the stress. The same authors identified genes exclusively upregulated in the tolerant genotype, with seven genes involved in flavonoid biosynthesis, six in phenylpropanoid biosynthesis pathways, and seven associated with plant signal transduction pathways. Two of these genes encode SAUR (Small Auxin Upregulated RNA) proteins responsible for cell enlargement, and plant growth and adaptation.
In two pairs of near-isogenic lines (NILs), DEGs linked to drought tolerance are involved in protein phosphorylation, oxidation-reduction, and transcription regulation [123]. The DEGs primarily function in protein and ATP binding, protein kinase activity, and DNA binding. The tolerant wheat genotype rapidly upregulates homologous genes connected with phospholipase C, components of the mitogen-activated protein kinase (MAPK) cascade, as well as the ethylene and abscisic acid (ABA) signaling pathways under drought stress [119]. Moreover, through gene enrichment analysis, elite wheat cultivars demonstrate distinct drought response strategies, with the tolerant cultivar featuring genes related to intracellular signal transduction, the MAP kinase pathway, and cellular response to abiotic stress, granting it better drought adaptability compared to the sensitive genotype [116]. Other molecular components related to genotype-dependent variation in wheat drought responses and adaptation are chloroplast genes encoding photosystem II core proteins [124], heat shock protein [125] and dehydrin [126] genes, as well as genes encoding late embryogenesis abundant (LEA) proteins, proteases, protease inhibitors [127], and the transcription factors AP2/EREBP, bZIP, MYB/MYC, NAC, and WRKY [128]. The overexpression of a gene called MORE ROOT (TaMOR-D), which encodes a wheat auxin responsive transcription factor from the LBD family, increases root growth and enhance grain yield in rice [129]. Additionally, ASYMMETRIC LEAVES2/LATERAL ORGAN BOUNDARIES DOMAIN (AS2/LOB) wheat genes has led to the identification of a specific transcription factor that plays a role in optimizing root architecture, thereby aiding in drought adaptation [130].
Overall, these findings emphasize the complexity of wheat response to drought and the involvement of distinct molecular mechanisms in drought adaptation across different wheat cultivars, which suggest potential targets for improving drought resistance through genetic engineering.
6. Genotype-Dependent Genetic Basis of Drought Tolerance
Wheat drought tolerance is a complex trait governed by multiple genetic loci known as quantitative trait loci (QTLs), as well as by environmental factors and their interactions. Study of wheat drought tolerance genetics entails examining both phenotypic (observable characteristics) and genotypic (genetic makeup) data to pinpoint chromosomal regions that harbor the pertinent loci (Figure 3). However, the identification of QTLs for drought tolerance is challenging due to the large size of the wheat genome, the numerous genes affecting drought resistance, epistatic QTL interactions, and the instability of certain QTLs [131,132,133,134]. To address these challenges, researchers have adopted diverse approaches, encompassing studies conducted under various environmental conditions to identify loci connected with drought stress tolerance in wheat [135,136], utilization of different types of bi-parental populations [137,138,139], and the implementation of a wide array of DNA molecular markers [137,140]. To reveal genomic regions governing wheat drought tolerance, various methodologies have been applied encompassing genome-wide association studies (GWAS) and QTL mapping. In addition, association mapping has been used to assess genetic diversity among wheat genotypes from different geographic regions [141,142]. Recent research using simple sequence repeat (SSR) markers has revealed the genetic diversity among a population of 117 modern wheat varieties (T. aestivum) from Bulgaria and several Western, Central and Eastern European countries. In this study, several drought-tolerant Bulgarian cultivars are grouped together, forming a branch within one of the sub-clusters linked with drought stress tolerance, thus highlighting their unique genetic makeup in relation to drought tolerance [143]. Furthermore, genomic regions related to drought tolerance-related traits have been successfully detected through QTL mapping [81,144]. Most mapping studies in wheat have primarily focused on identifying QTLs responsible for the final yield components under drought stress conditions. These studies have successfully pinpointed major genomic regions on chromosomes 1B, 2B, 3B, 7B, 4A, 5A, 1D, and 7D that control various productivity traits, such as grain yield, kernel number per spike, thousand-kernel weight, heading time, among others. The genomic regions have also shown associations with QTLs related to drought-adaptive traits, which suggest their importance in conferring wheat drought tolerance [145,146,147]. It is noteworthy that that some QTLs may have pleiotropic effects, meaning they can simultaneously influence multiple agronomic traits in either a synergistic or antagonistic manner. Additionally, the same QTLs can exhibit neutral effects on these traits in different environmental conditions. QTLs with pleiotropic effects on agronomic traits have been identified on chromosomes 1B, 4B, 5A, 4A, 7A, 2D, 3D, 5D, and 6D [148,149,150,151,152]. Later, the identification of QTLs associated with morphological and physiological traits linked to wheat drought tolerance has become a priority area of research. This shift in focus is driven by the belief that indirect selection based on these traits can be more efficient than direct selection for higher yield, and provide a better understanding of the drought impacts at different growth stages [3]. Using microsatellite markers, Malik et al. [153] effectively mapped QTLs related to photosynthesis, cell membrane stability, and RWC on chromosome 2A, reporting their connection with the plant developmental stage. During the early stages (earing), marker-trait associations have been detected on chromosomes 1B, 4B, 5B, and 7A, while in the later stages (maturing), they have been observed on chromosomes 1B, 2A, 3D, 4B, 5B, and 6D. Another study confirmed the critical role of chromosome 2A in conferring drought-tolerant status to wheat plants. The researchers identified candidate genes associated with hormonal signaling pathways mediated by gibberellic, jasmonic, abscisic, salicylic acids and ethylene [154]. Moreover, ABA responsiveness at the seedling stage has been reported on chromosomes 1B, 2A, 3A, 5A, 6D, and 7B [155,156].
The seedling stage plays a key role for drought tolerance, as it greatly impacts all subsequent growth stages and, ultimately, the grain yield. Several studies have been dedicated to exploring genetic variations during the seedling stage, leading to the identification of numerous QTLs with varying effects on seedling traits. These QTLs have been mapped to specific chromosomes, namely 1B, 1D, 2B, 3A, 3B, 3D, 4A, 6A, 6B, 6D, 7A, and 7B [157]. Further analysis of these QTLs has has shed light on a specific region on chromosome 7B, which is linked to drought susceptibility. Gene annotation analysis by Ahmed et al. [157] has revealed the presence of gene clusters within this region that encode for ubiquitin-associated (UBA)-like superfamily proteins, which act as negative regulators of drought stress responses. In addition to the above findings, several other studies have identified QTLs related to shoot dry weight, number of culms, plant height, root dry weight, root volume, root length, root surface area, and the number of root forks and tips. Kocheva et al. [158] observed improved adaptation to drought stress in wheat seedlings carrying Rht-B1 alleles. Further analysis has revealed that one of the QTLs on chromosome 4B corresponds to the Rht-B1 locus, which substantially influences shoot and root traits [159]. Recently, Schierenbeck et al. [160] conducted an association analysis to identify quantitative trait nucleotides (QTNs) influencing drought tolerance traits during the seedling stage. They discovered 70 stable QTNs across 17 chromosomes, with eight of them specifically located on chromosomes 1B, 2A, 2B, 2D, 4B, 7A, and 7B, all related to multiple seedling growth-related traits.
To recapitulate, wheat drought resistance is a multifaceted phenomenon governed by numerous genetic factors and strongly influenced by environmental conditions. Recent research has made significant progress in mapping QTLs linked to various drought-related traits, however, only a limited number of these QTLs have consistent effects across different environments and populations [161]. Leveraging genomic information for predicting the performance of individuals, holds great potential in enhancing breeding efforts aimed at developing drought-resistant varieties. Additionally, the discovery and utilization of epistatic QTLs (QTL interactions) can further improve breeding strategies for drought resistance [133].
7. Genotype-Dependent Epigenetic Responses
Epigenetic modifications, which alter gene expression patterns without changing the underlying DNA sequence, have also emerged as principal regulators of wheat responses to drought stress (Figure 3).
Water deficit exerts a profound impact on the DNA methylation status of plants, shaping gene expression and plant drought stress tolerance. Dynamic DNA methylation and demethylation events target specific genetic loci, and result in intricate patterns of plant methylation [162]. Mounting evidence indicate that different wheat genotypes exhibit distinct DNA methylation patterns under drought stress, suggesting a potential correlation between methylation status and drought tolerance [163]. DNA methylation emerges as the primary mechanism activated in wheat that actively shapes the plant ability to cope with the adverse conditions, while demethylation events play a complementary and supportive role [164]. It has been observed that hypermethylation is mainly associated with increased vulnerability to drought, whereas hypomethylation is indicative of enhanced resilience and tolerance to arid conditions [163,165]. Furthermore, the impact of drought stress extends to organ-specific alterations in the methylation levels of the wheat genome, as elucidated through studies comparing the methylation patterns of two contrasting genotypes: the drought-tolerant genotype C306 and drought-sensitive genotype HUW468. Interestingly, while some similarities in DNA methylation patterns are observed between the leaves and roots of these genotypes, notable differences in methylation and demethylation events are still apparent [163]. The drought-tolerant genotype C306 displays a higher frequency of demethylation events, whereas the drought-sensitive genotype HUW468 exhibits a higher occurrence of methylation events. Moreover, under drought stress, wheat leaves display a higher content of 5-methyl cytosine and greater overall methylation levels compared to root tissues [164,166].
Alterations in histone acetylation levels represent another crucial mechanism that affects the expression of stress-responsive genes. In general, wheat tissues that grow faster show higher expression levels of histone acetyltransferase (HAT) genes. This suggests that the way TaHATs are expressed varies among tissues and is connected to the plant growth [167]. In the drought-resistant wheat variety BN207, the levels of HAT genes, including TaHAG2, TaHAG3, TaHAC2 and TaHDT1, have been upregulated compared to the levels observed in the drought-sensitive varieties BN64 and ZM16 [168]. The dynamic regulation of HATs and histone deacetylases (HDACs) controls multiple signaling pathways invloved in drought responses, such as those involving ABA, acetic and jasmonic acids [169,170]. A comprehensive analysis of the wheat genome has identified a total of 166 SET domain genes SET [Su(var)3–9, Enhancer-of-zeste and Trithorax] domain genes (SDGs)]. Among these genes, 30 have been downregulated, and six exhibited upregulation in different plant organs during the seedling stage. Under drought conditions, only TaSDG23b-1BI has been upregulated, emphasizing its role as an epigenetic mark involved in the methylation of H3K9 [171]. The expression of TaSDG1a-7A and TaSDG20-3D genes, responsible for the methylation of H3K9 and H3K27, respectively, have been also induced by drought stress. Importantly, the methylation of histones mediated by these genes ultimately leads to the downregulation of specific target genes, which is associated with enhanced stress tolerance of wheat plants.
Chromatin remodeling processes involve the dynamic change of histone-DNA interactions, leading to the unfolding of compact DNA structures and facilitating the access of transcription factors [172]. These processes can be achieved through two distinct mechanisms: the action of chromatin-remodeling factors (e.g., SWI/SNF, ISWI, INO80, and CHD) or specialized enzymes [173]. The involvement of these factors in chromatin remodeling has already been demonstrated in Arabidopsis [174,175]. The first indication of transcription factor-mediated recruitment for chromatin remodeling in wheat has been provided by Wang et al. [176]. They found that the wheat CHD-type chromatin remodeling factor, known as TaCHR729, interacts with the promoter regions of wheat 3-KETOACYL-CoA SYNTHASE (TaKCS6). This enzyme plays a pivotal role in wheat cuticular wax biosynthesis. Additionally, TaKPAB1, a bHLH type transcription factor, has been also associated with the TaKCS6 promoter in conjunction with TaCHR729. Wang et al. [176] further revealed that the silencing of TaCHR729 regulates the interplay between wheat and powdery mildew by modulating histone methylation and finely adjusting the biosynthesis of cuticular wax.
Small interfering RNAs (siRNAs) are a predominant class of small RNAs (sRNAs) in plants [177,178]. Alongside microRNAs (miRNAs), they play vital roles in epigenetic regulatory pathways and modulate the expression of genes associated with development and stress tolerance [179,180]. Using deep sequencing technology, Ma et al. [181] analyzed two wheat genotypes with contrasting stress tolerance and identified 367 differentially expressed miRNAs in leaves under drought stress. Among these, 13 miRNAs are downregulated in the drought-tolerant cultivar Hanxuan10, but upregulated in the drought-susceptible cultivar Zhengyin1. Moreover, a comparative transcriptome analysis of the roots in modern durum wheat and its wild relatives (Triticum turgidum ssp. durum variety Kızıltan and two Triticum turgidum ssp. dicoccoides genotypes TR39477 and TTD-22) displays genotype- and/or stress-specific associations of miRNAs under drought conditions [182]. The levels of miRNA expression vary between stress-tolerant and stress-sensitive genotypes based on the type of stress, genotype and time-point. Through transcriptome sequencing, DEGs associated with hormone homeostasis, photosynthesis and signaling are discovered [183]. Furthermore, miRNA regulation in durum wheat controls physiological parameters, yield performance, and grain quality traits in the subsequent generations following drought stress [184]. Li et al. [185] have focused on noncoding RNAs (lncRNAs and miRNAs) and their role in regulating gene expression during drought stress using two wheat genotypes with different drought tolerance. Analyzing the relationships and expression patterns of lncRNAs, miRNAs, and DEGs, they have identified 10 regulatory modules responsible for wheat response to drought stress.
In summary, epigenetic modifications are important regulatory mechanisms that fine-tune gene expression and ultimately contribute to the plant capacity to withstand and adapt to drought challenges. The exploration of these epigenetic processes holds great promise for unraveling novel strategies to enhance drought resilience in wheat and other essential crops. Moreover, studies on the transgenerational effects of drought stress have provided valuable insights into the long-term impact of epigenetic modifications on phenotypic traits, highlighting the significance of the epigenetic marks on crop performance and quality. Understanding the specific epigenetic modifications unique to each genotype offers valuable insights into the molecular mechanisms governing plant responses to water scarcity, thereby paving the way for targeted approaches to enhance drought resilience in agricultural systems.
8. Exploring the Drought Resistance of Bulgarian Wheat Genotypes
Maximizing the assessment and characterization of wheat genotypes in various repositories is instrumental in optimizing the utilization of these invaluable resources. Bulgaria, known for its rich history in wheat cultivation, has been curating collections of wheat genotypes, primarily in the breeding centers of Dobrudzha Agricultural Institute in General Toshevo, and the Institute of Plant Genetic Resources in Sadovo [186,187,188]. These collections represent vital repositories for exploring and identifying cultivars with improved traits, particularly in relation to their ability to withstand drought conditions.
Previous investigations have extensively investigated the performance of Bulgarian wheat genotypes under varying degrees of water scarcity and subsequent recovery phases. Through detailed analysis, these studies have successfully identified several traits associated with enhanced wheat drought tolerance [46,84,94,189,190,191]. Generous funding provided by the National Science Fund of Bulgaria for the project titled ‘Study on adaptive mechanisms to drought in Bulgarian winter wheat varieties’ has enabled these studies to progress seamlessly, and encompass an assessment of a wide range of agronomic, physiological, molecular, and genetic traits of genotypes with different stress tolerance levels. Examination of the genetic and phenotypic diversity among multiple wheat cultivars have provided valuable insights into plant adaptability to diverse agro-climatic regions. The prevalence of specific SSR alleles in sub-populations indicates that different cultivars have developed specific genetic traits to thrive in their respective environments [143]. Genotype-dependent impacts of drought and subsequent rewatering have been noticed in various aspects, including leaf water deficit and transcriptional changes in DNA methyltransferase coding genes [192], leaf morphology (trichome density, stomatal frequency, and guard cell length), alterations in phytohormone levels and their derivatives, mobilization of antioxidant defense components, and modifications in the functioning of the photosynthetic apparatus, evident through changes in the mRNA content of chloroplast-encoded photosynthetic genes psbA and rbcL. Through GC-MS analysis, cultivar-dependent drought adaptation strategies were found, with noticeable changes in primary and secondary metabolites. Furthermore, the activation and suppression of protease isoenzymes showing cultivar-dependent variations, particularly in root proteases. Dehydration also triggered the mobilization of different components of the antioxidant defense system, displaying organ- and genotype-specific responses. Cultivars displaying higher tolerance to water deficit exhibited greater total antioxidant activity and ROS scavenging potential. Additionally, dehydration induced primary DNA damage, including single- and double-stranded breaks in all wheat cultivars, with more pronounced effects observed in sensitive genotypes (unpublished data).
Overall, the comprehensive assessment and characterization of Bulgarian wheat genotypes highlight their diversity and offer potential markers for distinguishing between resistant and sensitive cultivars. This knowledge can facilitate the development and selection of local wheat cultivars with enhanced drought tolerance.
9. Conclusions
Understanding the responses of different wheat genotypes to drought stress provides valuable knowledge for developing drought-resistant wheat cultivars and optimizing agricultural practices in regions prone to water scarcity. By studying these responses researchers can identify specific genes and mechanisms that confer drought tolerance. This knowledge could serve as a base for generation of drought-tolerant wheat cultivars through conventional breeding techniques or advanced biotechnological approaches like gene editing. In addition, the data gathered can assists in devising effective agronomic strategies by adopting appropriate management practices to minimize the drought effect on wheat production, which in turn, will contribute to ensuring food security in an ever-changing climate.
Author Contributions
Conceptualization, V.V.; writing—original draft preparation, V.V., A.D., M.G. and G.Z.; writing—review and editing, V.V.; visualization, M.G. and G.Z.; supervision, V.V.; project administration, A.D.; funding acquisition, V.V. and A.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Fund of Bulgaria (BNSF), grant number DN06/12 ‘Study on adaptive mechanisms to drought in Bulgarian winter wheat varieties’.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 2009 103, 551–560. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Khadka, K.; Raizada, M.N.; Navabi, A. Recent progress in germplasm evaluation and gene mapping to enable breeding of drought-tolerant wheat. Front. Plant Sci. 2020, 11, 1149. [Google Scholar] [CrossRef] [PubMed]
- Moravec, V.; Markonis, Y.; Rakovec, O.; Svoboda, M.; Trnka, M.; Kumar, R.; Hanel, M. Europe under multi-year droughts: How severe was the 2014-2018 drought period? Environ. Res. Lett. 2021, 16, 034062. [Google Scholar] [CrossRef]
- Kazandjiev, V.; Georgieva, V.; Malasheva, P.; Atanassov, D. Evapotranspiration and drought in different agricultural zones of Bulgaria. In Challenges in Agro-Climate and Ecosystem, Saifullah, M., Tardio, G.B., Mickovski, S., Eds.; IntechOpen, 2022. [CrossRef]
- Georgieva, V.; Kazandjiev, V.; Bozhanova, V.; Mihova, G.; Ivanova, D.; Todorovska, E.; Uhr, Z.; Ilchovska, M.; Sotirov, D.; Malasheva, P. Climatic changes—A challenge for the Bulgarian farmers. Agriculture 2022, 12, 2090. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [PubMed]
- De Leonardis, A.M.; Marone, D.; Mazzucotelli, E.; Neffar, F.; Rizza, F.; Di Fonzo, N.; Cattivelli, L.; Mastrangelo, A.M. Durum wheat genes up-regulated in the early phases of cold stress are modulated by drought in a developmental and genotype dependent manner. Plant Sci. 2007, 172, 1005–1016. [Google Scholar] [CrossRef]
- Pastori, G.M.; Foyer, C.H. Common components, networks, and pathways of cross-tolerance to stress. The central role of “redox” and abscisic acid-mediated controls. Plant Physiol. 2002, 129, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Tuberosa, R. Dissection and modelling of abiotic stress tolerance in plants. Curr. Opin. Plant Biol. 2010, 13, 206–212. [Google Scholar] [CrossRef]
- Mitura, K.; Cacak-Pietrzak, G.; Feledyn-Szewczyk, B.; Szablewski, T.; Studnicki, M. Yield and grain quality of common wheat (Triticum aestivum L.) depending on the different farming systems (organic vs. integrated vs. conventional). Plants 2023, 12, 1022. [Google Scholar] [CrossRef] [PubMed]
- Rockström, J.; Falkenmark, M. Agriculture: Increase water harvesting in Africa. Nature 2015, 519, 283–285. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhang, Q.; Sun, P.; Song, C. Impact of droughts on winter wheat yield in different growth stages during 2001–2016 in Eastern China. Int. J. Disaster Risk Reduct. 2018, 3, 376–391. [Google Scholar] [CrossRef]
- Rich, S.M.; Wasson, A.P.; Richards, R.A.; Katore, T.; Prashar, R.; Chowdhary, R.; Saxena, D.C.; Mamrutha, H.M.; Zwart, A.; Misra, S.C.; et al. Wheats developed for high yield on stored soil moisture have deep vigorous root systems. Funct. Plant Biol. 2016, 43, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Portmann, F.T.; Siebert, S.; Döll, P. MIRCA2000-Global monthly irrigated and rainfed crop areas around the year 2000: A new high-resolution data set for agricultural and hydrological modeling. Glob. Biogeochem. Cycles 2010, 24, 1–24. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Christopher, J.T.; Hammer, G.L.; Devoil, P. Experimental and modelling studies of drought-adaptive root architectural traits in wheat (Triticum aestivum L.). Plant Biosyst. 2010, 144, 458–462. [Google Scholar] [CrossRef]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.S.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.S.; Rebetzke, G.J.; Reynolds, M. Integration of phenotyping and genetic platforms for a better understanding of wheat performance under drought. J. Exp. Bot. 2014, 65, 6167–6177. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. Response and defence mechanisms of vegetable crops against drought, heat and salinity stress. Agriculture, 2021, 11, 463. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; De Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Res. 2016, 5, F1000. [Google Scholar] [CrossRef]
- Franks, S.J. Plasticity and evolution in drought avoidance and escape in the annual plant Brassica rapa. New Phytol. 2011, 190, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Sherrard, M.E.; Maherali, H. The adaptive significance of drought escape in Avena barbata, an annual grass. Evolution 2006, 60, 2478–2489. [Google Scholar] [CrossRef] [PubMed]
- Kooyers, N.J. The evolution of drought escape and avoidance in natural herbaceous populations. Plant Sci. 2015, 234, 155–162. [Google Scholar] [CrossRef]
- Du, H.; Huang, F.; Wu, N.; Li, X.; Hu, H.; Xiong, L. Integrative regulation of drought escape through ABA dependent and independent pathways in rice. Mol. Plant. 2018, 11, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Dohleman, F.G.; Long, S.P. More productive than maize in the midwest: How does miscanthus do it? Plant Physiol. 150, 2104–2115. [CrossRef] [PubMed]
- Tardieu, F.; Simonneau, T.; Muller, B. The physiological basis of drought tolerance in crop plants: A scenario-dependent probabilistic approach. Annu. Rev. Plant Biol. 2018, 69, 733–759. [Google Scholar] [CrossRef] [PubMed]
- Álvaro, F.; Isidro, J.; Villegas, D.; García del Moral, L.F.; Royo, C. Breeding effects on grain filling, biomass partitioning, and remobilization in Mediterranean durum wheat. Agron. J. 2008, 100, 361–370. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Park, S.H.; Jenks, M.A. Changes in leaf cuticular waxes of sesame (Sesamum indicum L.) plants exposed to water deficit. J. Plant Physiol. 2007, 164, 1134–1143. [Google Scholar] [CrossRef] [PubMed]
- Delfin, E.F.; Drobnitch, S.T.; Comas, L.H. Plant strategies for maximizing growth during water stress and subsequent recovery in Solanum melongena L. (eggplant). PLoS ONE 2021, 16, e0256342. [Google Scholar] [CrossRef]
- Mori, M.; Inagaki, M.N.; Inoue, T.; Nachit, M.M. Association of root water-uptake ability with drought adaptation in wheat. Cereal Res. Commun. 2011, 39, 551–559. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Turner, N.C.; Liu, Y.X.; Kadambot, H.M.S.; Xiong, Y.C. Effects of drought stress on morphological, physiological and biochemical characteristics of wheat species differing in ploidy level. Funct. Plant Biol. 2016, 44, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ma, B.; Palta, J.A.; Ding, T.; Cheng, Z.; Lv, G.; Xiong, Y. Wheat breeding highlights drought tolerance while ignores the advantages of drought avoidance: A meta-analysis. Eur. J. Agron. 2021, 122, 126196. [Google Scholar] [CrossRef]
- Loss, S.P.; Siddique, K.H.M. Morphological and physiological traits associated with wheat yield increases in Mediterranean environments. Adv. Agron. 1994, 52, 229–276. [Google Scholar] [CrossRef]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5, 106. [Google Scholar] [CrossRef] [PubMed]
- Puijalon, S.; Bouma, T.J.; Douady, C.J.; van Groenendael, J.; Anten, N.P.; Martel, E.; Bornette, G. Plant resistance to mechanical stress: Evidence of an avoidance-tolerance trade-off. New Phytol. 2011, 191, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.M. Osmoregulation as a selection criterion for drought tolerance in wheat. Aust. J. Agric. Res. 1983, 34, 607–614. [Google Scholar] [CrossRef]
- Izanloo, A.; Condon, A.G.; Langridge, P.; Tester, M.; Schnurbusch, T. Different mechanisms of adaptation to cyclic water stress in two South Australian bread wheat cultivars. J. Exp. Bot. 2008, 59, 3327–3346. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought stress in wheat during flowering and grain-filling periods. CRC Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Velinov, V.; Vaseva, I.; Zehirov, G.; Zhiponova, M.; Georgieva, M.; Vangheluwe, N.; Beeckman, T.; Vassileva, V. Overexpression of the NMig1 gene encoding a NudC domain protein enhances root growth and abiotic stress tolerance in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 815. [Google Scholar] [CrossRef] [PubMed]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Vassileva, V.; Signarbieux, C.; Anders, I.; Feller, U. Genotypic variation in drought stress response and subsequent recovery of wheat (Triticum aestivum L.). J. Plant Res. 2011, 124, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Meister, R.; Rajani, M.S.; Ruzicka, D.; Schachtman, D.P. Challenges of modifying root traits in crops for agriculture. Trends Plant Sci. 2014, 19, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Manske, G.G.B.; Vlek, P.L.G. Root architecture - wheat as a model plant. In Plant Roots: The Hidden Half, 3rd ed.; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 249–259. [Google Scholar]
- Placido, D.F.; Sandhu, J.; Sato, S.J.; Nersesian, N.; Quach, T.; Clemente, T.E.; Staswick, P.E.; Walia, H. The LATERAL ROOT DENSITY gene regulates root growth during water stress in wheat. Plant Biotechnol. J. 2020, 18, 1955–1968. [Google Scholar] [CrossRef] [PubMed]
- Golan, G.; Hendel, E.; Mendez Espitia, G.E.; Schwartz, N.; Peleg, Z. Activation of seminal root primordia during wheat domestication reveals underlying mechanisms of plant resilience. Plant Cell Environ. 2018, 41, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Whalley, P.A.; Ashton, R.W.; Evans, J.; Hawkesford, M.J.; Griffiths, S.; Huang, Z.D.; Zhou, H.; Mooney, S.J.; Whalley, W.R. A comparison between water uptake and root length density in winter wheat: Effects of root density and rhizosphere properties. Plant Soil 2020, 451, 345–356. [Google Scholar] [CrossRef]
- Palta, J.; Watt, M. Vigorous crop root systems: Form and function for improving the capture of water and nutrients. In Applied Crop Physiology: Boundaries between Genetic Improvement and Agronomy. Academic, San Diego, 2009; pp. 309–325.
- Ober, E.S.; Alahmad, S.; Cockram, J.; Forestan, C.; Hickey, L.T.; Kant, J.; Maccaferri, M.; Marr, E.; Milner, M.; Pinto, F.; et al. Wheat root systems as a breeding target for climate resilience. Theor. Appl. Genet. 2021, 134, 1645–1662. [Google Scholar] [CrossRef] [PubMed]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Manschadi, A.M.; Hammer, G.L.; Christopher, J.T.; Devoil, P. Genotypic variation in seedling root architectural traits and implications for drought adaptation in wheat (Triticum aestivum L.). Plant Soil 2008, 303, 115–129. [Google Scholar] [CrossRef]
- Nakamoto, T.; Oyanagi, A. The direction of growth of seminal roots of Triticum aestivum L. and experimental modification thereof. Ann. Bot. 1994, 73, 363–367. [Google Scholar] [CrossRef]
- Slack, S.; York, L.M.; Roghazai, Y.; Lynch, J.; Bennett, M.; Foulkes, J. Wheat shovelomics II: Revealing relationships between root crown traits and crop growth. BioRxiv 2018, 0917. [Google Scholar] [CrossRef]
- Alahmad, S.; El Hassouni, K.; Bassi, F.M.; Dinglasan, E.; Youssef, C.; Quarry, G.; Aksoy, A.; Mazzucotelli, E.; Juhász, A.; Able, J.A.; et al. A major root architecture QTL responding to water limitation in durum wheat. Front. Plant Sci. 2019, 10, 436. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Du, Y.; Wang, J.; Wu, A.; Qiao, S.; Xu, B.; Zhang, S.; Siddique, K.H.; Chen, Y. Moderate drought stress affected root growth and grain yield in old, modern and newly released cultivars of winter wheat. Front. Plant Sci. 2017, 8, 672. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, M.T.; Hordyńska, N.; Maksymowicz, A.; Grzesiak, S.; Szechyńska-Hebda, M. Variation among spring wheat (Triticum aestivum L.) genotypes in response to the drought stress. II-Root system structure. Plants 2019, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- McDonald, G. The effects of root angle on root growth and yield of wheat in the Australian cereal belt. In Food Security from Sustainable Agriculture: Proceedings of 15th Agronomy Conference.; p. 2010.
- Ouyang, W.; Yin, X.; Yang, J.; Struik, P.C. Comparisons with wheat reveal root anatomical and histochemical constraints of rice under water-deficit stress. Plant Soil 2020, 2020 452, 547–568. [Google Scholar] [CrossRef]
- Kadam, N.N.; Yin, X.; Bindraban, P.S.; Struik, P.C.; Jagadish, K.S. Does morphological and anatomical plasticity during the vegetative stage make wheat more tolerant of water deficit stress than rice? Plant Physiol. 2015, 167, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.A.; Passioura, J.B. A breeding program to reduce the diameter of the major xylem vessel in the seminal roots of wheat and its effect on grain yield in rain-fed environments. Aust. J. Agric. Res. 1989, 40, 943–950. [Google Scholar] [CrossRef]
- Ehdaie, B.; Layne, A.P.; Waines, J.G. Root system plasticity to drought influences grain yield in bread wheat. Euphytica 2012, 186, 219–232. [Google Scholar] [CrossRef]
- Correa, J.; Postma, J.A.; Watt, M.; Wojciechowski, T. Soil compaction and the architectural plasticity of root systems. J. Exp. Bot. 2019, 70, 6019–6034. [Google Scholar] [CrossRef] [PubMed]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 2013, 610721. [Google Scholar] [CrossRef] [PubMed]
- Farhad, M.; Hakim, M.A.; Alam, M.A.; Barma, N.C.D. Screening wheat genotypes for coleoptile length: A trait for drought tolerance. Am. J. Agric. Forest 2014, 2, 237–245. [Google Scholar] [CrossRef]
- Petrov, P.; Petrova, A.; Dimitrov, I.; Tashev, T.; Olsovska, K.; Brestic, M.; Misheva, S. Relationships between leaf morpho-anatomy, water status and cell membrane stability in leaves of wheat seedlings subjected to severe soil drought. J. Agron. Crop Sci. 2018, 204, 219–227. [Google Scholar] [CrossRef]
- Sewore, B.M.; Abe, A.; Nigussie, M. Evaluation of bread wheat (Triticum aestivum L.) genotypes for drought tolerance using morpho-physiological traits under drought-stressed and well-watered conditions. PLoS ONE 2023, 18, p–e0283347. [Google Scholar] [CrossRef] [PubMed]
- Kilic, H.; Yagbasanlar, T. The effect of drought stress on grain yield, yield components and some quality traits of durum wheat (Triticum turgidum) cultivars. Not. Bot. Horti. Agrobot. 2010, 38, 164–170. [Google Scholar] [CrossRef]
- Liwani, U.; Magwaza, L.S.; Odindo, A.O.; Sithole, N.J. Growth, morphological and yield responses of irrigated wheat (Triticum aestivum L.) genotypes to water stress. Acta Agric. Scand. B Soil Plant Sci. 2019, 69, 369–376. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Han, X.; Tian, F.; Wu, J. Effects of water stress on photosynthesis, yield, and water use efficiency in winter wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Willick, I.R.; Lahlali, R.; Vijayan, P.; Muir, D.; Karunakaran, C.; Tanino, K.K. Wheat flag leaf epicuticular wax morphology and composition in response to moderate drought stress are revealed by SEM, FTIR-ATR and synchrotron X-ray spectroscopy. Physiol. Plant. 2018, 162, 316–332. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.A.; Rawson, H.M.; Johnson, D.A. Glaucousness in wheat: Its development and effect on water-use efficiency, gas exchange and photosynthetic tissue temperatures. Aust. J. Plant Physiol. 1986, 13, 465–473. [Google Scholar] [CrossRef]
- David, O.A.; Osonubi, O.; Olaiya, C.O.; Agbolade, J.O.; Ajiboye, A.A.; Komolafe, R.J.; Chukwuma, D.M.; Akomolafe, G.F. Anatomical response of wheat cultivars to drought stress. Ife J. Sci. 2017, 19, 323–331. [Google Scholar] [CrossRef]
- Jäger, K.; Fábián, A.; Eitel, G.; Szabó, L.; Deák, C.; Barnabás, B.; Papp, I. A morpho-physiological approach differentiates bread wheat cultivars of contrasting tolerance under cyclic water stress. J. Plant Physiol. 2014, 171, 1256–1266. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Bruce, S.E.; Kirkegaard, J.A. Longer coleoptiles improve emergence through crop residues to increase seedling number and biomass in wheat (Triticum aestivum L.). Plant Soil 2005, 272, 87–100. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Richards, R.A.; Fettell, N.A.; Long, M.; Condon, A.G.; Forrester, R.I.; Botwright, T.L. Genotypic increases in coleoptile length improves stand establishment, vigour and grain yield of deep-sown wheat. Field Crops Res. 2007, 100, 10–23. [Google Scholar] [CrossRef]
- Schillinger, W.F.; Donaldson, E.; Allan, R.E.; Jones, S.S. Winter wheat seedling emergence from deep sowing depths. Agron. J. 1998, 90, 582–586. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.; Baenziger, P.S.; Börner, A. Drought stress tolerance in wheat and barley: Advances in physiology, breeding and genetics research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [PubMed]
- Ghaffar, A.; Hussain, N.; Ajaj, R.; Shahin, S.M.; Bano, H.; Javed, M.; Khalid, A.; Yasmin, M.; Shah, K.H.; Zaheer, M.; et al. Photosynthetic activity and metabolic profiling of bread wheat cultivars contrasting in drought tolerance. Front. Plant Sci. 2023, 14, 1123080. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed]
- Vassileva, V.; Demirevska, K.; Simova-Stoilova, L.; Petrova, T.; Tsenov, N.; Feller, U. Long-term field drought affects leaf protein pattern and chloroplast ultrastructure of winter wheat in a cultivar-specific manner. J. Agron. Crop Sci. 2012, 198, 104–117. [Google Scholar] [CrossRef]
- Qayyum, A.; Al Ayoubi, S.; Sher, A.; Bibi, Y.; Ahmad, S.; Shen, Z.; Jenks, M.A. Improvement in drought tolerance in bread wheat is related to an improvement in osmolyte production, antioxidant enzyme activities, and gaseous exchange. Saudi J. Biol. Sci. 2021, 28, 5238–5249. [Google Scholar] [CrossRef] [PubMed]
- Khamssi, N.N.; Najaphy, A. Agro-morphological and phenological attributes under irrigated and rain-fed conditions in bread wheat genotypes. Afr. J. Agric. Res. 2012, 7, 51–57. [Google Scholar] [CrossRef]
- Clarke, J.M.; McCaig, T.N. Evaluation of techniques for screening for drought resistance in wheat. Crop Sci. 1982, 22, 503–506. [Google Scholar] [CrossRef]
- Roostaei, M.; Mohammadi, S.A.; Amri, A.; Majidi, E.; Nachit, M.; Haghparast, R. Chlorophyll fluorescence parameters and drought tolerance in a mapping population of winter bread wheat in the highlands of Iran. Russ. J. Plant Physiol. 2011, 58, 351–358. [Google Scholar] [CrossRef]
- Larouk, C.; Gabon, F.; Kehel, Z.; Djekoun, A.; Nachit, M.; Amri, A. Chlorophyll fluorescence and drought tolerance in a mapping population of durum wheat. Contemporary Agric. 2021, 70, 123–134. [Google Scholar] [CrossRef]
- Grigorova, B.; Vassileva, V.; Klimchuk, D.; Vaseva, I.; Demirevska, K.; Feller, U. Drought, high temperature, and their combination affect ultrastructure of chloroplasts and mitochondria in wheat (Triticum aestivum L.) leaves. J. Plant Interact. 2012, 7, 204–213. [Google Scholar] [CrossRef]
- Basal, H.; Smith, C.W.; Thaxton, P.S.; Hemphill, J.K. Seedling drought tolerance in upland cotton. Crop Sci. 2005, 45, 766–771. [Google Scholar] [CrossRef]
- Kane, C.N.; Jordan, G.J.; Jansen, S.; McAdam, S.A. A permeable cuticle, not open stomata, is the primary source of water loss from expanding leaves. Front. Plant Sci. 2020, 11, 774. [Google Scholar] [CrossRef]
- Thapa, S.; Reddy, S.K.; Fuentealba, M.P.; Xue, Q.; Rudd, J.C.; Jessup, K.E.; Devkota, R.N.; Liu, S. Physiological responses to water stress and yield of winter wheat cultivars differing in drought tolerance. J. Agron. Crop Sci. 2018, 204, 347–358. [Google Scholar] [CrossRef]
- Demirevska, K.; Simova-Stoilova, L.; Vassileva, V.; Feller, U. Rubisco and some chaperone protein responses to water stress and rewatering at early seedling growth of drought sensitive and tolerant wheat varieties. Plant Growth Regul. 2008, 56, 97–106. [Google Scholar] [CrossRef]
- Thakur, P.; Prasad, L.C.; Prasad, R.; Chandra, K. Estimation of genetic variability, heat susceptibility index and tolerance efficiency of wheat (Triticum aestivum L.) for timely and late sown environments. Electron. J. Plant Breed. 2020, 11, 769–775. [Google Scholar] [CrossRef]
- Khalilzadeh, R.; Seyed Sharifi, R.; Jalilian, J. Antioxidant status and physiological responses of wheat (Triticum aestivum L.) to cycocel application and bio fertilizers under water limitation condition. J. Plant Interact. 2016, 11, 130–137. [Google Scholar] [CrossRef]
- Marček, T.; Hamow, K.A.; Végh, B.; Janda, T.; Darko, E. Metabolic response to drought in six winter wheat genotypes. PLoS ONE 2019, 14, e0212411. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Kumar, A.; Kaur, N. ROS and oxidative burst: Roots in plant development. Plant Divers. 2020, 42, 33–43. [Google Scholar] [CrossRef]
- Maevskaya, S.N.; Nikolaeva, M.K. Response of antioxidant and osmoprotective systems of wheat seedlings to drought and rehydration. Russ. J. Plant Physiol. 2013, 60, 343–350. [Google Scholar] [CrossRef]
- Simova-Stoilova, L.; Demirevska, K.; Petrova, T.; Tsenov, N.; Feller, U. Antioxidative protection and proteolytic activity in tolerant and sensitive wheat (Triticum aestivum L.) varieties subjected to long-term field drought. Plant Growth Regul. 2009, 58, 107–117. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef]
- Tabarzad, A.; Ayoubi, B.; Riasat, M.; Saed-Moucheshi, A.; Pessarakli, M. Perusing biochemical antioxidant enzymes as selection criteria under drought stress in wheat varieties. J. Plant Nutr. 2017, 40, 2413–2420. [Google Scholar] [CrossRef]
- Zhang, W.; Gu, J.; Wang, Z.; Wei, C.; Yang, J.; Zhang, J. Comparison of structural and functional properties of wheat starch under different soil drought conditions. Sci. Rep. 2017, 7, 12312. [Google Scholar] [CrossRef]
- Fan, X.W.; Li, F.M.; Song, L.; Xiong, Y.C.; An, L.Z.; Jia, Y.; Fang, X.W. Defense strategy of old and modern spring wheat varieties during soil drying. Physiol. Plant. 2009, 136, 310–323. [Google Scholar] [CrossRef]
- Erdei, L.; Tari, I.; Csiszar, J.; Pecsvaradi, A.; Horvath, F.; Szabo, M.; Ordog, M.; Cseuz, L.; Zhiponova, M.; Szilak, L.; et al. Osmotic stress responses of wheat species and cultivars differing in drought tolerance: Some interesting genes (advices for gene hunting). Acta Biol. Szeged 2002, 46, 63–65. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Camaille, M.; Fabre, N.; Clément, C.; Ait Barka, E. Advances in wheat physiology in response to drought and the role of plant growth promoting rhizobacteria to trigger drought tolerance. Microorganisms 2021, 9, 687. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Al-Busaidi, W.M.; Al-Sadi, A.M.; Farooq, M. Bread wheat genotypes accumulating free proline and phenolics can better tolerate drought stress through sustained rate of photosynthesis. J. Soil Sci. Plant Nutr. 2022, 22, 165–176. [Google Scholar] [CrossRef]
- Saeedipour, S. Relationship of grain yield, ABA and proline accumulation in tolerant and sensitive wheat cultivars as affected by water stress. Proc. Natl. Acad. Sci. U.S.A. 2013, 83, 311–315. [Google Scholar] [CrossRef]
- Pecetti, L.; Annicchiarico, P.; Gorham, J. Field heterogeneity of the stress affects genotypic response to salinity in durum wheat. Cereal Res. Commun. 1995, 23, 173–177. [Google Scholar]
- Wang, G.P.; Zhang, X.Y.; Li, F.; Luo, Y.; Wang, W. Overaccumulation of glycine betaine enhances tolerance to drought and heat stress in wheat leaves in the protection of photosynthesis. Photosynthetica 2010, 48, 117–126. [Google Scholar] [CrossRef]
- Bowne, J.B. , Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef]
- Wang, X.; Mao, Z.; Zhang, J.; Hemat, M.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Osmolyte accumulation plays important roles in the drought priming induced tolerance to post-anthesis drought stress in winter wheat (Triticum aestivum L.). Environ. Exp. Bot. 2019, 166, 103804. [Google Scholar] [CrossRef]
- Khoshro, H.H.; Taleei, A.; Bihamta, M.R.; Shahbazi, M.; Abbasi, A. Expression analysis of the genes involved in osmotic adjustment in bread wheat (Triticum aestivum L.) cultivars under terminal drought stress conditions. J. Crop Sci. Biotechnol. 2013, 16, 173–181. [Google Scholar] [CrossRef]
- Bogdan, J.; Zagdańska, B. Changes in the pool of soluble sugars induced by dehydration at the heterotrophic phase of growth of wheat seedlings. Plant Physiol. Biochem. 2006, 44, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Feng, K.; Peng, S.; Wang, J.; Zhang, Y.; Bian, J.; Nie, X. Comparative analysis of the transcriptional response of tolerant and sensitive wheat genotypes to drought stress in field conditions. Agronomy 2018, 8, 247. [Google Scholar] [CrossRef]
- Shamloo-Dashtpagerdi, R.; Shahriari, A.G.; Tahmasebi, A.; Vetukuri, R.R. Potential role of the regulatory miR1119-MYC2 module in wheat (Triticum aestivum L.) drought tolerance. Front. Plant Sci. 2023, 14, 1161245. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Xie, Y.; Fan, S.; Wang, Z.; Wang, F.; Zhang, B.; Li, H.; Song, J.; Kong, L. Comparative analysis of root transcriptome profiles between drought-tolerant and susceptible wheat genotypes in response to water stress. Plant Sci. 2018, 272, 276–293. [Google Scholar] [CrossRef]
- Ergen, N.Z.; Thimmapuram, J.; Bohnert, H.J.; Budak, H. Transcriptome pathways unique to dehydration tolerant relatives of modern wheat. Funct. Integr. Genom. 2009, 9, 377–396. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, R.; Wang, H.; Li, D.; Wang, X.; Zhang, Y.; Zhen, W.; Duan, H.; Yan, G.; Li, Y. Transcriptomics analyses reveal wheat responses to drought stress during reproductive stages under field conditions. Front. Plant Sci. 2017, 8, 592. [Google Scholar] [CrossRef]
- Dalal, M.; Sahu, S.; Tiwari, S.; Rao, A.R.; Gaikwad, K. Transcriptome analysis reveals interplay between hormones, ROS metabolism and cell wall biosynthesis for drought-induced root growth in wheat. Plant Physiol. Biochem. 2018, 130, 482–492. [Google Scholar] [CrossRef]
- Mia, M.S.; Liu, H.; Wang, X.; Zhang, C.; Yan, G. Root transcriptome profiling of contrasting wheat genotypes provides an insight to their adaptive strategies to water deficit. Sci. Rep. 2020, 10, 4854. [Google Scholar] [CrossRef]
- Nouraei, S.; Mia, M.S.; Liu, H.; Turner, N.C.; Yan, G. Transcriptome analyses of near isogenic lines reveal putative drought tolerance controlling genes in wheat. Front. Plant Sci. 2022, 13, 857829. [Google Scholar] [CrossRef]
- Liu, W.J.; Yuan, S.; Zhang, N.H.; Lei, T.; Duan, H.G.; Liang, H.G.; Lin, H.H. Effect of water stress on photosystem 2 in two wheat cultivars. Biol. Plant. 2006, 50, 597–602. [Google Scholar] [CrossRef]
- Yu, T.F.; Xu, Z.S.; Guo, J.K.; Wang, Y.X.; Abernathy, B.; Fu, J.D.; Chen, X.; Zhou, Y.B.; Chen, M.; Ye, X.G.; et al. Improved drought tolerance in wheat plants overexpressing a synthetic bacterial cold shock protein gene SeCspA. Sci. Rep. 2017, 7, 44050. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.G.; Banowetz, G.M.; Peterson, C.J.; Kronstad, W.E. Dehydrin expression and drought tolerance in seven wheat cultivars. Crop Sci. 2003, 43, 577–582. [Google Scholar] [CrossRef]
- Vaseva, I.I.; Grigorova, B.S.; Simova-Stoilova, L.P.; Demirevska, K.N.; Feller, U. Abscisic acid and late embryogenesis abundant protein profile changes in winter wheat under progressive drought stress. Plant Biol. 2010, 12, 698–707. [Google Scholar] [CrossRef]
- Gahlaut, V.; Jaiswal, V.; Kumar, A.; Gupta, P.K. Transcription factors involved in drought tolerance and their possible role in developing drought tolerant cultivars with emphasis on wheat (Triticum aestivum L.). Theor. Appl. Genet. 2016, 129, 2019–2042. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liu, D.; Li, Q.; Mao, X.; Li, A.; Wang, J.; Chang, X.; Jing, R. Overexpression of wheat gene TaMOR improves root system architecture and grain yield in Oryza sativa. J. Exp. Bot. 2016, 67, 4155–4167. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, R.; Cheng, Y.; Lei, P.; Song, W.; Zheng, W.; Nie, X. Genome-wide identification, evolution, and expression analysis of LBD transcription factor family in bread wheat (Triticum aestivum L.). Front. Plant Sci. 2021, 12, 721253. [Google Scholar] [CrossRef]
- Des Marais, D.L.; Hernandez, K.M.; Juenger, T.E. Genotype-by-environment interaction and plasticity: Exploring genomic responses of plants to the abiotic environment. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 5–29. [Google Scholar] [CrossRef]
- Ishikawa, A. A strategy for identifying quantitative trait genes using gene expression analysis and causal analysis. Genes 2017, 8, 347. [Google Scholar] [CrossRef]
- Gupta, P.K.; Balyan, H.S.; Gahlaut, V. QTL analysis for drought tolerance in wheat: Present status and future possibilities. Agronomy 2017, 7, 5. [Google Scholar] [CrossRef]
- Bapela, T.; Shimelis, H.; Tsilo, T.J.; Mathew, I. Genetic improvement of wheat for drought tolerance: Progress, challenges and opportunities. Plants 2022, 11, 1331. [Google Scholar] [CrossRef]
- Malosetti, M.; Voltas, J.; Romagosa, I.; Ullrich, S.E.; Van Eeuwijk, F.A. Mixed models including environmental covariables for studying QTL by environment interaction. Euphytica 2004, 137, 139–145. [Google Scholar] [CrossRef]
- Mathews, K.L.; Malosetti, M.; Chapman, S.; McIntyre, L.; Reynolds, M.; Shorter, R.; Van Eeuwijk, F. Multi-environment QTL mixed models for drought stress adaptation in wheat. Theor. Appl. Genet. 2008, 117, 1077–1091. [Google Scholar] [CrossRef] [PubMed]
- Quarrie, S.A.; Gulli, M.; Calestani, C.; Steed, A.; Marmiroli, N. Location of a gene regulating drought-induced abscisic acid production on the long arm of chromosome 5A of wheat. Theor. Appl. Genet. 1994, 89, 794–800. [Google Scholar] [CrossRef]
- McIntyre, C.L.; Mathews, K.L.; Rattey, A.; Chapman, S.C.; Drenth, J.; Ghaderi, M.; Reynolds, M.; Shorter, R. Molecular detection of genomic regions associated with grain yield and yield-related components in an elite bread wheat cross evaluated under irrigated and rainfed conditions. Theor. Appl. Genet. 2010, 120, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Czyczyło-Mysza, I.; Marcińska, I.; Skrzypek, E.; Cyganek, K.; Juzoń, K.; Karbarz, M. QTL mapping for germination of seeds obtained from previous wheat generation under drought. Open Life Sci. 2014, 9, 374–382. [Google Scholar] [CrossRef]
- Maccaferri, M.; Mantovani, P.; Tuberosa, R.; DeAmbrogio, E.; Giuliani, S.; Demontis, A.; Massi, A.; Sanguineti, M.C. A major QTL for durable leaf rust resistance widely exploited in durum wheat breeding programs maps on the distal region of chromosome arm 7BL. Theor. Appl. Genet. 2008, 117, 1225–1240. [Google Scholar] [CrossRef]
- Nielsen, H.B.; Almeida, M.; Juncker, A.S.; Rasmussen, S.; Li, J.; Sunagawa, S.; Plichta, D.R.; Gautier, L.; Pedersen, A.G.; Le Chatelier, E.; et al. Identification and assembly of genomes and genetic elements in complex metagenomic samples without using reference genomes. Nature Biotechnol. 2014, 32, 822–828. [Google Scholar] [CrossRef]
- Yang, Y.; Chai, Y.; Zhang, X.; Lu, S.; Zhao, Z.; Wei, D.; Chen, L.; Hu, Y.G. Multi-locus GWAS of quality traits in bread wheat: Mining more candidate genes and possible regulatory network. Front. Plant Sci. 2020, 11, 1091. [Google Scholar] [CrossRef]
- Tsonev, S.; Christov, N.K.; Mihova, G.; Dimitrova, A.; Todorovska, E.G. Genetic diversity and population structure of bread wheat varieties grown in Bulgaria based on microsatellite and phenotypic analyses. Biotechnol. Biotechnol. Equip. 2021, 35, 1520–1533. [Google Scholar] [CrossRef]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef]
- Peleg, Z.V.I.; Fahima, T.; Krugman, T.; Abbo, S.; Yakir, D.A.N.; Korol, A.B.; Saranga, Y. Genomic dissection of drought resistance in durum wheat×wild emmer wheat recombinant inbreed line population. Plant, Cell & Environ. 2009, 32, 758–779. [Google Scholar] [CrossRef]
- Tahmasebi, S.; Heidari, B.; Pakniyat, H.; McIntyre, C.L. Mapping QTLs associated with agronomic and physiological traits under terminal drought and heat stress conditions in wheat (Triticum aestivum L.). Genome 2017, 60, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Zandipour, M.; Majidi Hervan, E.; Azadi, A.; Khosroshahli, M.; Etminan, A. A QTL hot spot region on chromosome 1B for nine important traits under terminal drought stress conditions in wheat. Cereal Res. Commun. 2020, 48, 17–24. [Google Scholar] [CrossRef]
- Deng, S.; Wu, X.; Wu, Y.; Zhou, R.; Wang, H.; Jia, J.; Liu, S. Characterization and precise mapping of a QTL increasing spike number with pleiotropic effects in wheat. Theor. Appl. Genet. 2011, 122, 281–289. [Google Scholar] [CrossRef]
- Jin, J.; Liu, D.; Qi, Y.; Ma, J.; Zhen, W. Major QTL for seven yield-related traits in common wheat (Triticum aestivum L.). Front. Genet. 2020, 11, 1012. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Xu, Z.; Fan, X.; Zhou, Q.; Yu, Q.; Liu, X.; Liao, S.; Feng, B.; Wang, T. Identification of a major and stable QTL on chromosome 5A confers spike length in wheat (Triticum aestivum L.). Mol. Breed. 2021, 41, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Itam, M.O.; Mega, R.; Gorafi, Y.S.; Yamasaki, Y.; Tahir, I.S.; Akashi, K.; Tsujimoto, H. Genomic analysis for heat and combined heat-drought resilience in bread wheat under field conditions. Theor. Appl. Genet. 2022, 135, 337–350. [Google Scholar] [CrossRef]
- Said, A.A.; Moursi, Y.S.; Sallam, A. Association mapping and candidate genes for physiological non-destructive traits: Chlorophyll content, canopy temperature, and specific leaf area under normal and saline conditions in wheat. Front. Genet. 2022, 13, 980319. [Google Scholar] [CrossRef]
- Malik, S.; Malik, T.A. Genetic mapping of potential QTLs associated with drought tolerance in wheat. JAPS: J. Anim. Plant Sci. 2015, 25, 1032–1040. [Google Scholar]
- Pshenichnikova, T.A.; Osipova, S.V.; Smirnova, O.G.; Leonova, I.N.; Permyakova, M.D.; Permyakov, A.V.; Rudikovskaya, E.G.; Konstantinov, D.K.; Verkhoturov, V.V.; Lohwasser, U.; et al. Regions of chromosome 2A of bread wheat (Triticum aestivum L.) associated with variation in physiological and agronomical traits under contrasting water regimes. Plants 2021, 10, 1023. [Google Scholar] [CrossRef]
- Kobayashi, F.; Takumi, S.; Handa, H. Identification of quantitative trait loci for ABA responsiveness at the seedling stage associated with ABA-regulated gene expression in common wheat. Theor. Appl. Genet. 2010, 121, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Iehisa, J.C.; Matsuura, T.; Mori, I.C.; Takumi, S. Identification of quantitative trait locus for abscisic acid responsiveness on chromosome 5A and association with dehydration tolerance in common wheat seedlings. J. Plant Physiol. 2014, 171, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.; Mohamed, E.A.; Hussein, M.Y.; Sallam, A. Genomic regions associated with leaf wilting traits under drought stress in spring wheat at the seedling stage revealed by GWAS. Environ. Exp. Bot. 2021, 184, 104393. [Google Scholar] [CrossRef]
- Kocheva, K.; Nenova, V.; Karceva, T.; Petrov, P.; Georgiev, G.I.; Börner, A.; Landjeva, S. Changes in water status, membrane stability and antioxidant capacity of wheat seedlings carrying different Rht-B1 dwarfing alleles under drought stress. J. Agron. Crop Sci. 2014, 200, 83–91. [Google Scholar] [CrossRef]
- Iannucci, A.; Marone, D.; Russo, M.A.; De Vita, P.; Miullo, V.; Ferragonio, P.; Blanco, A.; Gadaleta, A.; Mastrangelo, A.M. Mapping QTL for root and shoot morphological traits in a durum wheat × T. dicoccum segregating population at seedling stage. Int. J. Genomics 2017, 2017, 6876393. [Google Scholar] [CrossRef]
- Schierenbeck, M.; Alqudah, A.M.; Thabet, S.G.; Lohwasser, U.; Simón, M.R.; Börner, A. Association mapping unravels the genetics controlling seedling drought stress tolerance in winter wheat. Front. Plant Sci. 2023, 14, 1061845. [Google Scholar] [CrossRef]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Fei, Y.; Xue, Y.; Du, P.; Yang, S.; Deng, X. Expression analysis and promoter methylation under osmotic and salinity stress of TaGAPC1 in wheat (Triticum aestivum L). Protoplasma 2017, 254, 987–996. [Google Scholar] [CrossRef]
- Kaur, A.; Grewal, A.; Sharma, P. Comparative analysis of DNA methylation changes in two contrasting wheat genotypes under water deficit. Biol. Plant. 2018, 62, 471–478. [Google Scholar] [CrossRef]
- Duan, H.; Li, J.; Zhu, Y.; Jia, W.; Wang, H.; Jiang, L.; Zhou, Y. Responsive changes of DNA methylation in wheat (Triticum aestivum) under water deficit. Sci. Rep. 2020, 10, 7938. [Google Scholar] [CrossRef]
- Lim, A. Drought-induced epigenetic modulation and transcriptional variation of winter wheat. BS Thesis, Oklahoma State University, USA, 2020. Available online: Hdl.handle.net/11244/329964.
- Li, J.; Jia, W.; Wang, H.; Zhu, Y.; Duan, Z.; Jiang, L.; Zhou, Y.; Duan, H. Morpho-physiological adaptation and DNA methylation of wheat seedlings under osmotic stress. Crop Pasture Sci. 2020, 71, 349–355. [Google Scholar] [CrossRef]
- Gao, S.; Li, L.; Han, X.; Liu, T.; Jin, P.; Cai, L.; Xu, M.; Zhang, T.; Zhang, F.; Chen, J.; et al. Genome-wide identification of the histone acetyltransferase gene family in Triticum aestivum. BMC Genom. 2021, 22, 1–17. [Google Scholar] [CrossRef]
- Li, H.; Liu, H.; Pei, X.; Chen, H.; Li, X.; Wang, J.; Wang, C. Comparative genome-wide analysis and expression profiling of histone acetyltransferases and histone deacetylases involved in the response to drought in wheat. J. Plant Growth Regul. 2022, 41, 1065–1078. [Google Scholar] [CrossRef]
- Li, S.; He, X.; Gao, Y.; Zhou, C.; Chiang, V.L.; Li, W. Histone acetylation changes in plant response to drought stress. Genes 2021, 12, 1409. [Google Scholar] [CrossRef] [PubMed]
- Demetriou, K.; Kapazoglou, A.; Tondelli, A.; Francia, E.; Stanca, M.A.; Bladenopoulos, K.; Tsaftaris, A.S. Epigenetic chromatin modifiers in barley: I. Cloning, mapping and expression analysis of the plant specific HD2 family of histone deacetylases from barley, during seed development and after hormonal treatment. Physiol Plant. 2009, 136, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Batra, R.; Gautam, T.; Pal, S.; Chaturvedi, D.; Rakhi, Jan, I. ; Balyan, H.S.; Gupta, P.K. Identification and characterization of SET domain family genes in bread wheat (Triticum aestivum L.). Sci. Rep. 2020, 10, 14624. [Google Scholar] [CrossRef]
- Kong, L.; Liu, Y.; Wang, X.; Chang, C. Insight into the role of epigenetic processes in abiotic and biotic stress response in wheat and barley. Int. J. Mol. Sci. 2020, 21, 1480. [Google Scholar] [CrossRef]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patterns and paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef]
- Saez, A.; Rodrigues, A.; Santiago, J.; Rubio, S.; Rodriguez, P.L. HAB1–SWI3B interaction reveals a link between abscisic acid signaling and putative SWI/SNF chromatin-remodeling complexes in Arabidopsis. Plant Cell 2008, 2008 20, 2972–2988. [Google Scholar] [CrossRef]
- Umezawa, T.; Sugiyama, N.; Takahashi, F.; Anderson, J.C.; Ishihama, Y.; Peck, S.C.; Shinozaki, K. Genetics and phosphoproteomics reveal a protein phosphorylation network in the abscisic acid signaling pathway in Arabidopsis thaliana. Sci. Signal. 2013, 6, rs8. [Google Scholar] [CrossRef]
- Wang, L.; Gao, Y.; Zheng, X.; Liu, C.; Dong, S.; Li, R.; Zhang, G.; Wei, Y.; Qu, H.; Li, Y.; et al. Histone modifications regulate chromatin compartmentalization by contributing to a phase separation mechanism. Mol. Cell 2019, 76, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Bhardwaj, A.R.; Agarwal, M.; Katiyar-Agarwal, S. Discovery of small RNAs in wheat: A survey. Indian J. Plant Physiol. 2017, 22, 411–421. [Google Scholar] [CrossRef]
- Shanker, A.K.; Maheswari, M. Small RNA and drought tolerance in crop plants. Indian J. Plant Physiol. 2017, 22, 422–433. [Google Scholar] [CrossRef]
- Bakhshi, B.; Fard, E.M.; Gharechahi, J.; Safarzadeh, M.; Nikpay, N.; Fotovat, R.; Azimi, M.R.; Salekdeh, G.H. The contrasting microRNA content of a drought tolerant and a drought susceptible wheat cultivar. J. Plant Physiol. 2017, 216, 35–43. [Google Scholar] [CrossRef]
- Gómez-Martín, C.; Zhou, H.; Medina, J.M.; Aparicio-Puerta, E.; Shi, B.; Hackenberg, M. Genome-wide analysis of microRNA expression profile in roots and leaves of three wheat cultivars under water and drought conditions. Biomolecules 2023, 13, 440. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xin, Z.; Wang, Z.; Yang, Q.; Guo, S.; Guo, X.; Cao, L.; Lin, T. Identification and comparative analysis of differentially expressed miRNAs in leaves of two wheat (Triticum aestivum L.) genotypes during dehydration stress. BMC Plant Biol. 2015, 15, 1–15. [Google Scholar] [CrossRef]
- Akpinar, B.A.; Kantar, M.; Budak, H. Root precursors of microRNAs in wild emmer and modern wheats show major differences in response to drought stress. Funct. Integr. Genom. 2015, 15, 587–598. [Google Scholar] [CrossRef]
- Liu, H. , Able, A.J. and Able, J.A. Integrated analysis of small RNA, transcriptome, and degradome sequencing reveals the water-deficit and heat stress response network in durum wheat. Int. J. Mol. Sci. 2020, 21, 6017. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Small RNAs and their targets are associated with the transgenerational effects of water-deficit stress in durum wheat. Sci. Rep. 2021, 11, 3613. [Google Scholar] [CrossRef]
- Li, N.; Liu, T.; Guo, F.; Yang, J.; Shi, Y.; Wang, S.; Sun, D. Identification of long non-coding RNA-microRNA-mRNA regulatory modules and their potential roles in drought stress response in wheat (Triticum aestivum L.). Front. Plant Sci. 2022, 13, 1011064. [Google Scholar] [CrossRef]
- Todorovska, E.; Abu Mhadi, N.; Christov, N.; Bozhanova, V.; Atanassov, A. Cereal genetics and genomics in Bulgaria - challenges and perspectives. C. R. Acad. Bulg. Sci. 2018, 71, 143–160. [Google Scholar]
- Mihova, G.; Baychev, V.; Alexandrov, T.; Petrova, T.; Stanoeva, Y.; Ivanova, V. Breeding of cereal crops at Dobrudzha Agricultural Institute-General Toshevo, Bulgaria. J. Agricult. Food Environmen. Sci., JAFES.
- Dimitrov, E.; Uhr, Z.; Chipilski, R. Study of yield and stability by common winter wheat varieties by changing climatic conditions in Sadovo region. Bulg. J. Agric. Sci. 2022, 28. [Google Scholar]
- Simova-Stoilova, L.; Vaseva, I.; Grigorova, B.; Demirevska, K.; Feller, U. Proteolytic activity and cysteine protease expression in wheat leaves under severe soil drought and recovery. Plant Physiol. Biochem. 2010, 48, 200–206. [Google Scholar] [CrossRef]
- Vassileva, V.; Simova-Stoilova, L.; Demirevska, K.; Feller, U. Variety-specific response of wheat (Triticum aestivum L.) leaf mitochondria to drought stress. J. Plant Res. 2009, 122, 445–454. [Google Scholar] [CrossRef]
- Vaseva, I.; Akiscan, Y.; Simova-Stoilova, L.; Kostadinova, A.; Nenkova, R.; Anders, I.; Feller, U.; Demirevska, K. Antioxidant response to drought in red and white clover. Acta Physiol. Plant. 2012, 34, 1689–1699. [Google Scholar] [CrossRef]
- Vassileva, V.; Vaseva, I.; Dimitrova, A. Expression profiling of DNA methyltransferase genes in wheat genotypes with contrasting drought tolerance. Bulg. J. Agric. Sci. 2019, 25, 845–851. [Google Scholar]
Figure 1.
Plant defense mechanisms against drought stress.
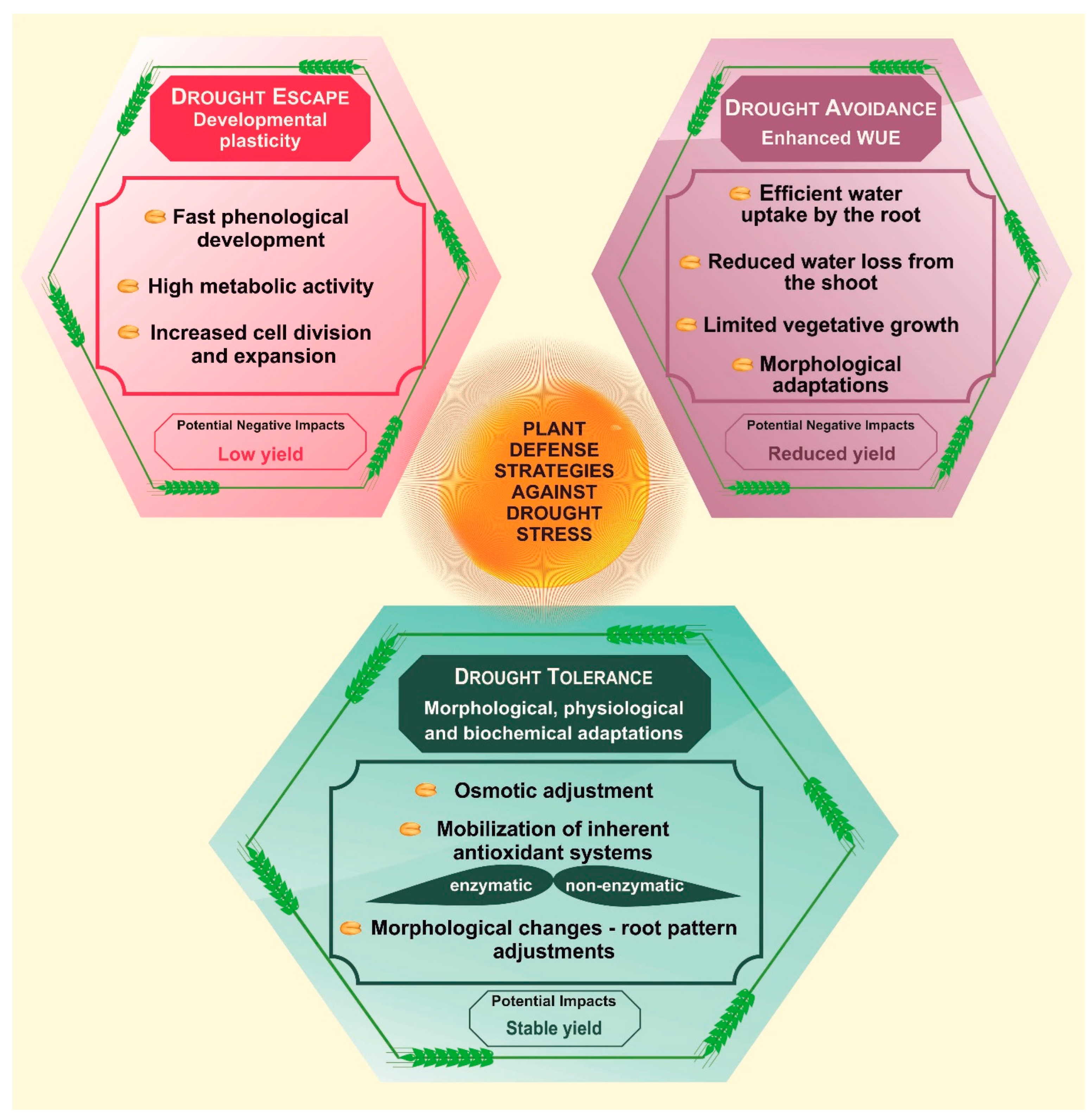
Figure 2.
Genotype-dependent morphological changes in the root and shoot of wheat plants under drought stress.
Figure 2.
Genotype-dependent morphological changes in the root and shoot of wheat plants under drought stress.
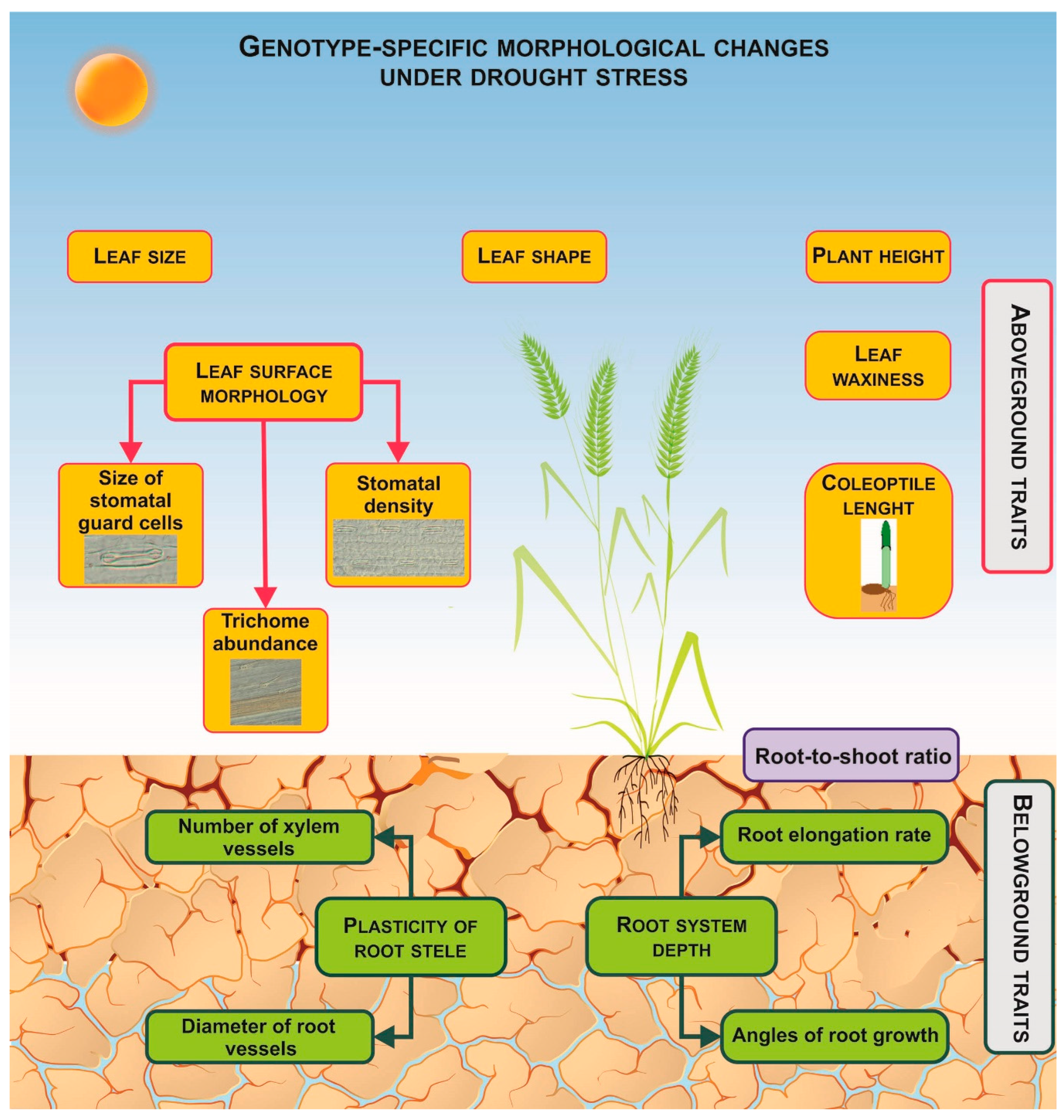
Figure 3.
Genotype-dependent physiological, biochemical, molecular, genetic and epigenetic changes in wheat plants under drought stress. Abbreviations: DEGs, differentially expressed genes; LEA proteins, late embryogenesis abundant proteins; MDA, malondialdehyde; miRNAs, microRNAs; PSII, Photosystem II; ROS, reactive oxygen species; RWC, relative water content; QTLs, quantitative trait loci.
Figure 3.
Genotype-dependent physiological, biochemical, molecular, genetic and epigenetic changes in wheat plants under drought stress. Abbreviations: DEGs, differentially expressed genes; LEA proteins, late embryogenesis abundant proteins; MDA, malondialdehyde; miRNAs, microRNAs; PSII, Photosystem II; ROS, reactive oxygen species; RWC, relative water content; QTLs, quantitative trait loci.
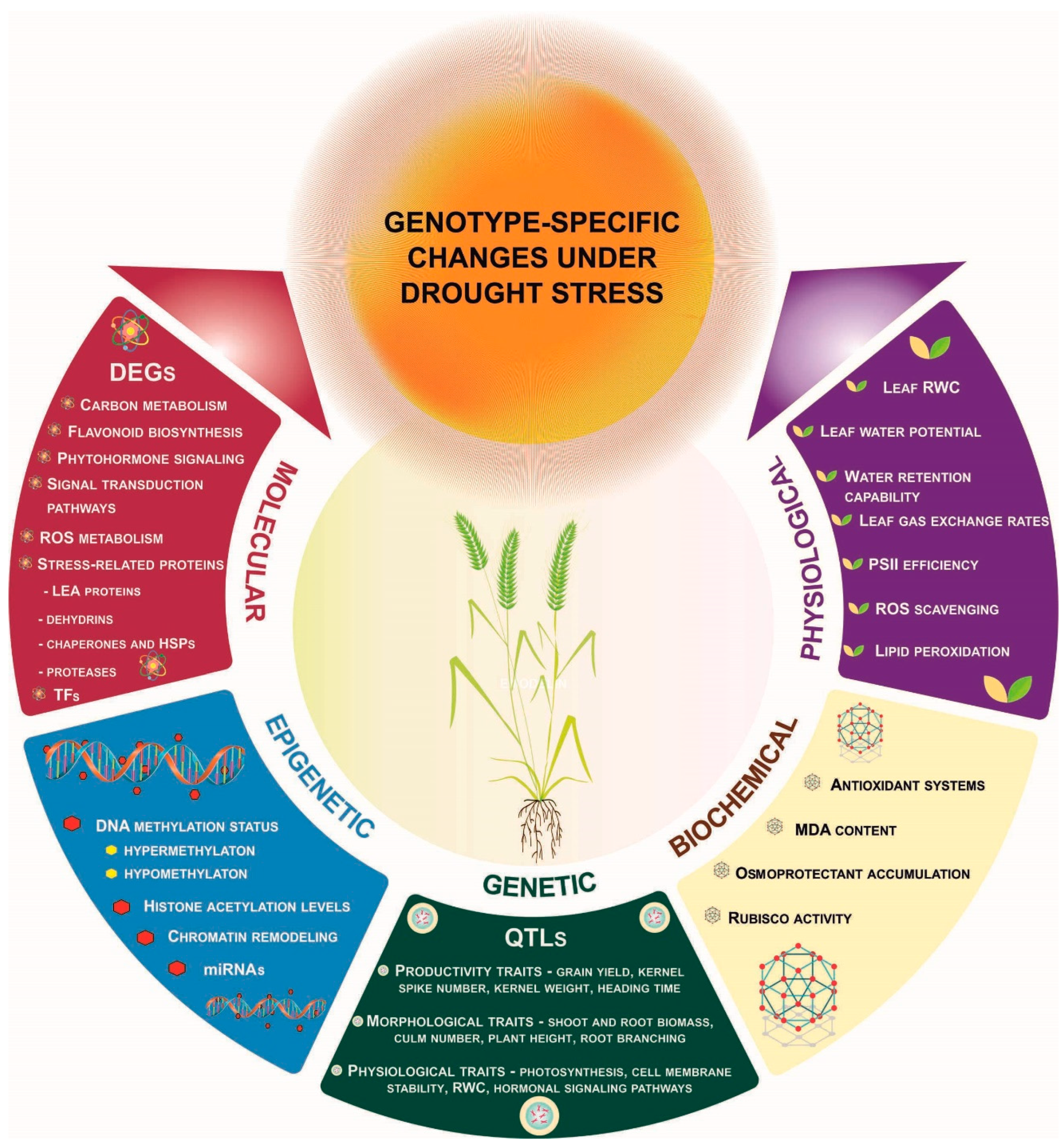
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



