
Submitted:
05 August 2023
Posted:
08 August 2023
You are already at the latest version
Abstract
In this review we extensively describe the main post-translational modifications that give rise to the multiple proteoforms characterized to date in the human salivary proteome and their potential role. Most of the data reported were obtained by our group in over twenty-five years of research carried out on human saliva mainly by applying a top-down strategy. At the beginning we describe the products generated by proteolytic cleavages, which can occur before and after secretion. In this section the most relevant families of salivary proteins are also described. Next, we report the current information concerning the human salivary phospho-proteome and the limited news available on sulfo-proteome. Three sections are dedicated to the description of the glycation and enzymatic glycosylation. Citrullination and N- and C- terminal PTMs and a miscellaneous of other modifications are described in the last two sections. Results highlighting the variation in the level of some proteoforms in local or systemic pathologies are also reviewed along the sections of manuscript to underline the impact and relevance of this information for the development of new diagnostic biomarkers useful in clinical practice.
Keywords:
salivary proteins
; top-down proteomics
; post-translational modifications
1. Introduction
Human whole saliva is a hypotonic fluid lining the oral cavity and composed of water (99%), a complex mixture of organic and inorganic compounds resulting from salivary gland secretion, oral flora, oropharynx, upper airway, gastrointestinal reflux, gingival crevicular fluid, food deposits, and mucosal surface secretion containing blood-derived components [1]. Saliva is essential for the accomplishment of multiple physiological func- tions encompassing lubrication, buffering action, maintenance of tooth integrity, chewing, initial digestion of some foods, swallowing, tissue hydration and lubrication, speech, wound healing, antibacterial, and antifungal activity [2]. The dramatic sequelae observed in patients suffering by the Sjögren syndrome clearly demonstrate the relevance of saliva and its components, particularly salivary proteins, in the protection of the mouth. Recent proteomic inventories report more than 3000 proteoforms in human saliva [3]. Before, during and after secretion, most salivary proteins undergo numerous post-translational modifications (PTMs), whose role has not yet been clearly elucidated. Over the last twenty-five years, our group has been able to characterize many PTMs of the salivary proteome, mainly by application of top-down proteomic platforms to saliva from subjects of various age and from patients affected by various pathologies. In this review, we report the PTMs most frequently observed, suggesting, when possible, some hypotheses on the possible role played in the protection of the mouth. We have done exhaustive research of these topics in the literature. Nonetheless, we apologize in advance for any possible omission.
2. Enzymatic cleavages (cryptides)
The most common PTM detectable in human saliva is the proteolytic cleavage of proteins. Indeed, most salivary proteins are submitted to the action of endogenous and exog enous proteinases (the last mainly from oral flora), which leads to the formation of a myriad of fragments. Many of these fragments could be considered important members of the cryptide family, defined as bioactive peptides encrypted inside a bigger functional polypeptide and released by a proteolytic event, with distinct or related function, but not superimposable, to that one of the parental polypeptides [4]. The action of proteases can occur before and after secretion, and in the following paragraphs this topic will be reviewed by describing separately pre-secretory and post-secretory cleavages.
2.1. Pre-secretory cleavages
The secretory pathway for many proteins includes transit in the Golgi apparatus and storage in secretory granules preceding their release from the cell into the duct system and secretion into the mouth [5]. Most of the pre-secretory cleavages of proteins occur during the transport towards the granules of trans-Golgi-network [6], which represents the major secretory pathway sorting station. Glandular secretions, protein extracts from secretory granule isolated from the major salivary glands, and whole saliva were investi gated by top-down proteomics to characterize and to distinguish between events occur ring prior the storage in the secretory granules, those taking place between granule release and secretion into the mouth during the passage through the secretory duct, and those occurring into the mouth [6]. The workflow applied in that study is represented in Figure 1. Proteases involved in the most relevant cleavages belong to the convertase and exopep tidase families [6,7,8,9].
2.1.1. Proline-rich proteins (PRPs)
Human PRPs constitute a very polymorphic family of proteins, missing in their se quence aromatic amino acids and characterized by high content of proline (as well as glu tamine and glycine). All PRPs are encoded by chromosome 12p13.2 and are divided in three classes based on their ionic properties, i.e. acidic proline-rich-proteins (aPRPs), en- coded by PRH1 and PRH2 loci, basic proline-rich proteins (bPRPs) encoded by the PRB1, PRB2 and PRB4 loci located (considering the direction of translation) immediately before the aPRPs loci, and the glycosylated (basic) proline-rich proteins (gPRPs) encoded by the PRB3 locus, that is the fourth locus of the bPRPs cluster. It is relevant to underline that except for the bPRPs named P-D, the other three proteoforms of PRB-4 locus, i.e. the IBa(Con 1+) of PRB-2 and all the products of PRB-3 locus are submitted to glycosylation, which will be described in more detail, if available, in the section devoted to this PTM.
2.1.1.1. Acidic proline-rich-proteins (aPRPs)
aPRPs exist as five principal isoforms called PRP-1, PRP-2 (coded by PRH-2 locus) and Pif-s (parotid isoelectric-focusing isoform-slow), Pa (parotid acidic isoform) and Db- s (double band isoform-slow), (coded by PRH-1 locus). The isoforms PRP-1, PRP-2, Pif-s and Pa are all residues long. Taking as reference PRP-2, the Pif-s and Pa isoforms have an asparagine (Asn4) instead of Asp4, PRP-1 has Asn50 instead of Asp50 present in the other isoforms (Figure 2). The Db-s isoform is a.a. residues long due to the insertion of 21 residues repeat after position 81 and, as Pa, it has a Leu27 instead of Ileu27. The PRP-1, PRP- and PIF-s isoforms are partially cleaved at the Arg106 by a convertase recognizing the ...R103XXR106… consensus sequence generating the PRP-3, PRP-4 and Pif-f (f stands for fast electrophoretic isoform) truncated isoforms and a terminal fragment of 44 amino acid res- idues called P-C (or IB-8b) peptide. The convertase consensus sequence of Db-s is shifted by the insertion, and it is cleaved at Arg127 generating the Db-f isoform and always the P C peptide. The consensus sequence of the convertase responsible for the cleavage is pre served because the Pa isoform, having a Cys103 residue instead of Arg103, is not cleaved. Indeed, a truncated Pa isoform was never detected. Rather it generates in the mouth a Pa 2-mer isoform due to the formation of a disulfide bridge between two Cys103 residues (Fig ure 2) that is the main Pa component detectable in whole saliva [9]. The cleavage of PRP 1, PRP-2, Pif-s, and Db-s is not complete and in whole saliva the entire and truncated isoforms are both detectable, approximatively in a proportion 70/30 %, respectively. All the entire isoforms are called PRP-1 type, while all the truncated are called PRP-3 type (obviously non-including the Pa proteoform). All these characteristics of aPRPs are sum- marized in Figure 2.
2.1.1.2. Basic-proline-rich-proteins (bPRPs)
bPRPs are encoded by the polymorphic PRB1, PRB2 and PRB4 loci, existing as three or four possible alleles, (S(mall), M(edium) and L(arge), or for the eventual fourth allele V(ery)L(arge)). Unlike aPRPs, they are completely cleaved by convertases during granule storage [6]. The proteoforms generated from the pre-pro-proteins are listed below:
- Products of Locus PRB-1:
II-2, P-E (or IB-9), P-Ko, IB-6, Ps-1, Ps-2,
- Products of Locus PRB-2:
IB-1, P-J, P-H (or IB-4), P-F (orIB-8c), IB-8a(Con 1+), IB-8a(Con 1-)
- Products of Locus PRB-4:
PGA, II-1, Cd-IIg, P-D (or IB-5).
The nomenclature reported for bPRPs is difficult and confusing, since it derives from two different criteria. Kauffmann and colleagues [10,11,12,13] have purified eleven basic-PRPs and determined the sequence of 10 of them. They named these proteoforms using the names of the different fractions obtained from an articulate multidimensional preparative chromatographic separation of whole saliva. In the same years, Saitoh and coll. [14,15,16] and Isemura and coll. [17,18] have identified nine basic-PRPs, naming them from P-A to P-I and sequenced seven of them. As reported in the list, the two nomenclatures have several overlaps. P-A and P-I are artifacts deriving from the proteolytic cleavage occurring 166 during purification of salivary proteins. Although included in the class of basic PRPs, P B, and P-C peptides are not codified by PRB1-PRB4 genes. Indeed, P-B peptide is encoded by PROL3 gene (PBI) clustered on chromosome 4q13.317. It is related to statherin and, therefore, it cannot be considered pertaining to bPRP family. As described in the previous 170 section on aPRPs, P-C peptide derives from the cleavage of four isoforms of aPRPs, namely PRP-1, PRP-2, Pif-s, and Db-s. In a top-down MS proteomic study we were able to characterize a new bPRP deriving from PRB-2 locus, which was named P-J [19,20].
The pre-secretory cleavage of bPRP pro-proteins usually occurs at the level of the consensus sequence KSRSXR↓, where X may be Pro, Ser, or Ala [6,21,22]. An extensive characterization of the different proteoforms and fragments of bPRPs has been recently published [23] where the reader interested can find the list of the bPRPs components de- tected until 2018. In this inventory a new classification of bPRPs was proposed, based on the similarities of their sequence, dividing them in three groups, reported in Figure 3(a) (b)-(c). The first group, which we named Group 1, includes P-E, P-Ko, IB-6, Ps-1, Ps-2, P H, P-F, P-J, and P-D (Figure 3(a)). The sequence of all these bPRPs starts with the same SPPGKPQGPP motif, followed by sequences somewhat similar but showing small varia- tions among the different components. The central part of the sequences shows similar repeats. Because P-E, IB-6, Ps-1, and Ps-2 sequences originate from DNA-length polymor- phisms in exon 3 of the PRB1 locus, they exhibit high similarity. The bPRP with a Mav of 10433.5 Da, detected in whole saliva and in parotid secretory granules and named P-Ko is encoded by cP4, a differentially spliced transcript of the PRB1-L allele.
Group 2 includes IB-1, II-2, and the glycosylated bPRPs codified by PRB3 and PRB4 genes, namely, Gl-1, Gl-2, Gl-3, GPA, II-1, and Cd-IIg (Figure 3(b)). Their sequences start with the similar motif (E/Q)XXXEDVSQEES, where XXX is LNE in IB-1, II-2, Gl-1, Gl-2, and Gl-3 and SSS in GPA, II-1, and Cd-IIg. The central part of the sequences comprises similar repeats with differences from the repeats of the members belonging to Group 1. Based on structural differences and similarities, members of Group 2 can be divided in three subgroups: Group 2A, including IB-1 and II-2, without glycosylation sequons; Group 2B, including the Gl proteins codified by the alleles of PRB3 locus; and Group 2C including the glycosylated proteins codified by the alleles of PRB4 locus. The small Group is a hybrid group, which includes the two proteoforms of IB-8a, Con1− and Con1+ (Figure 3(c)). The initial sequence of these two proteins resembles that of Group 1, while the terminal sequence is like the repeat responsible for the glycosylation of the bPRPs of Groups 2B and 2C [23].
2.1.2. Further pre-secretory cleavages of PRPs
All the PRP families above described are substrates of a carboxypeptidase, which re- moves the C-terminal residue, which commonly, as obvious, is an arginine. However, the enzyme is not specific. It removes other residues and sometimes a second C-terminal loss is detectable [22]. The C-terminal pre-secretory cleavages occurring on PRPs are reported in Table 1.
2.1.3. Role of PRPs
Salivary proline-rich proteins are highly conserved in mammalian saliva [24]. None- theless significant structural differences are present in the mammal families, suggesting that they play a relevant role in the oral protection, in the modulation of the activity of oral ions, as well as in the colonization of oral microbiota and the gastrointestinal tract and in the feeding habit [24]. However, while aPRPs and the gPRP called Gl 3M are de- tectable in the saliva of preterm newborns, the other bPRPs are not detectable in human saliva till the puberty [20,25,26] suggesting either a role in the perception of the taste of foods or a function in the secondary sexual maturation. As we are aware, this information is not available for other mammals. In human, bPRPs are secreted only by parotid glands, and this regioselectivity is puzzling too. Some bPRPs exhibit the ability to bind harmful tannins [27], others can modulate the oral flora [28,29], and some others are involved in bitter taste perception [30]. Some bPRP fragments are involved in enamel pellicle for mation [31] and others act as antagonists of the progesterone-induced cytosolic Ca2+ mo bilization [32].
The intrinsic propensity of some fragments to adopt a polyproline-II helix arrange ment joined to PxxP motifs was suggestive of the interaction with the SH3 domain family [33]. Interestingly, interactions were highlighted with Fyn, Hck, and c-Src SH3 domains [34], which are included in the Src kinases family, suggesting that some basic bPRPs can be involved in the signal transduction pathways modulated by these kinases. Only a small amount of data on correlations between genes of bPRPs and diseases linked to their allelic variants have been reported so far. In fact, for some of the alleles (PRB1VL, PRB2S, M, VL, PRB3VL) the genetic sequence is not reported. Moreover, for the small and large alleles of PRB1, the genetic sequence is incomplete [35] because the reference genome (NCBI Gene ID: 5542) encodes the medium allele. Regarding the primary structure of bPRP alleles, in the UniProtKB database (accession number P04280) is deposited the full amino acid se- quence of the large variant, deduced through experimental evidence at the protein level.
2.1.4. Statherin and P-B peptides
Statherin is encoded by STATH gene localized on chromosome 4q13.3 [36]. It is a 43- amino-acid-residues multifunctional phospho-peptide characterized by an anomalous high content of tyrosine, proline and glutamine. The roles of statherin in the oral cavity are various. Due to its great affinity for calcium phosphate minerals, such as hydroxyap- atite, it maintains the supersaturated state of calcium in human saliva, thus contributing to the mineral dynamics of tooth surface and stabilizing the acquired enamel [37]. Fur thermore, statherin is involved in bacterial colonization [38] and acts as a boundary lub ricant on the enamel surface [39]. P-B is the product of the specific PROL3 gene located on chromosome 4q13.3, very close to STATH gene [40]. Differently from bPRPs, which do not have aromatic amino acids in their sequences, and similarly to statherin, it has three tyro sine residues in its sequence. Furthermore, like statherin, P-B peptide is secreted mainly by submandibular/sublingual (SM/SL) glands and by parotid too [41]. It is important to recall that P-B is also called at the Swiss-Prot site (code P02814) “submaxillary gland an drogen-regulated protein 3B precursor”, but we were not able to find any reference justi fying the origin of this name. A specific role of P-B peptide in saliva is still far to be clari fied [42,43]. Nonetheless, its sequence resembles the ionic complement of statherin. In deed, statherin has a small N-terminal negative tail followed by a long neutral domain with many proline residues, while P-B peptide has a small N-terminal positive tail fol lowed by a long neutral poly-proline domain. The two peptides could interact for the for mation of the acquired enamel pellicle of the tooth. Different isoforms of statherin have been detected in human saliva; the variant called SV1 (Statherin variant 1) corresponds to statherin missing the C-terminal Phe residue (Statherin desPhe43) [41,44] (Figure 4(a)). The variant SV2, showing the deletion of the 6–15 internal residues with respect to statherin (Statherin des6-15), is originated by an alternative splicing that excludes an exon of 30 nucleotides [45]. SV3 corresponds to the SV2 variant lacking the C-terminal Phe residue. SV-1 lacking the C-terminal Phe43 and statherin desThr42Phe43 (Figure 4(a)) were detected in parotid and submandibular secretory granules, suggesting that C-terminal re moval occurs during maturation of the secretory granules [6]. On the contrary, the N terminal removal that generates statherin desAsp1 (Figure 4(a)) is probably an event oc curring after granule secretion because this derivative was not detected in granules [6]. Statherin desAsp1 was detected in human saliva also by Vitorino and coll. [46].
Similarly small amount of P-B des1–5 was found in whole saliva as well as in parotid and submandibular-sublingual saliva but not in secretory granules suggesting that also this N-terminal cleavage occurs after secretion [6]. Some P-B and statherin naturally oc- curring fragments are generated from a cleavage operated by a furin-like convertase and have been characterized by top-down approach in our previous studies [6,41]. Statherin undergoes to convertase cleavages at the consensus sequence …R9R10IGR13…. generating statherin des1-9, des1-10 and des1-13 (Figure 4(a)), while P-B des1-5 is a convertase frag- ment of P-B peptide generated at the consensus sequence …R2GPR5… (Figure 4(b)). P-B peptide undergo further cleavages at the level of various chymotryptic-like sequences generating several fragments detectable by a top-down proteomic approach [20] (Figure 4(b)).
2.1.5. Histatins (Hst)
Histatins are a family of small peptides deriving their name from the high number of histidine residues in their sequence. It is widely accepted that all the members of this fam- ily arise from two parent peptides, named histatin 1 and histatin 3, having very similar sequences and encoded by two genes (HIS1 and HIS2) located on chromosome 4q13 [47]. Despite the very high sequence similarity, these two peptides follow completely different PTM pathways. Histatin 3 (Hst3), differently from histatin 1 (Hst1), is submitted to a se quential cleavage generating at first Hst6 (Hst3 1/25), subsequently Hst5 (Hst 3 1/24) hav ing powerful anti-fungal activity [48] and then other fragments [49]. Their different sus ceptibility to cleavage derives from the presence in Hst3 of the …RGYR↓… convertase consensus sequence, which is absent in Hst1 and thus it is not cleaved. Hst1 today is considered a proangiogenic factor that may contribute to oral wound healing [50,51].
Some years ago, the group of Oppenheim [48] was able to establish the sequence of 12 histatins and named them from Hst1 to Hst12. The advent of the high throughput MS apparatus applied to proteomic studies allowed the determination of many other Hst frag ments, the majority deriving from Hst3 [49,52]. For this reason, the nomenclature of histat ins fragments has been recently modified and it is reported at the Swiss Prot site with the code numbers P15515 (Hst1) and P15516 (Hst3).
2.1.6. Cystatins
Cystatins include type 1 cystatins (cystatins A and B), type 2 cystatins (C, D, S, SN, SA), and kininogens, or type 3 cystatins. Various biological activities have been demon- strated for these proteins, but a major role is linked to the inhibitory action exerted against cysteine proteinases. Thus, for this ability to modulate the proteolytic system they are considered central in various diseases, including cancer [53,54,55,56]. Cystatins S, SN, SA, C and D are encoded by loci CST1–5 closely clustered on chromosome 20p11.21. It has been observed that S-type cystatins (S, SN and SA), present at higher concentration in SM/SL secretion than in parotid saliva [6], were absent or present at very low concentration from both parotid and SM/SL secretory granules. This finding suggested that S-type cystatins’ secretion is not granule mediated [6]. It is relevant to remark that cystatin A and B are leaderless [57] Cystatin C was found sporadically in parotid, SM/SL, and whole saliva, but it was absent from secretory granules, whereas Cystatin D was not found in any sample. Thus, it was suggested that cystatins C and D may have a different origin than the other salivary cystatins. The presence in human saliva of truncated proteoforms of cystatins have been suggested by Lupi and coll., which observed by HPLC-ESI-MS several masses possibly related to N-terminally truncated cystatins in the acidic soluble fraction of human saliva [58]. Indeed, a subsequent in-depth study performed both in the acidic supernatant of whole saliva and in RP-HPLC enriched fractions, by an integrated top-down/bottom-up pipeline, characterized some truncated proteoforms of cystatins [57]. The study evi denced that not all cystatins undergo proteolytic modifications. The following truncated proteoforms for the widespread cystatin SN and its natural variant SN Pro11 → Leu were detected and characterized: cystatin SN des1−4, and SN des1−7, cystatin SN Pro11 → Leu 354 des1−4, and SN Pro11 → Leu des1−7. A truncated form of Cystatin SA lacking the first seven amino acids from the N-terminus (cystatin SA des1−7) was also detected. Three truncated forms of the variant cystatin D Cys26 →Arg have been detected in human saliva, but none of the variant Arg26 →Cys. For easier reading we will utilize the name cystatin D to refer to the variant Cys26 →Arg. Cystatin D des1−4 and des1−8 have been characterized 359 by a top-down Fourier-transform ion cyclotron resonance mass spectrometry pipeline [59], and detected also in the study of Manconi and coll. [57] A third truncated proteoform of cystatin D lacking the first five amino acid residues with the N-terminal glutamine con verted to pyroglutamic acid (pGlu-cystatin D des1−5) was also characterized in the latter study. The authors speculated that the high abundance of the latter truncated proteoform was due to the greater resistance from degradation by amino peptidases due to N-termi nal pyroglutamination. Cystatin B in adult human saliva is commonly detectable as intact proteoform, and the two fragments 1-53 and 54-98 were characterized in saliva of human pre-term newborns [60].
2.2. Post-secretory cleavages
Almost all the families of salivary proteins and peptides above described (and others) are substrates of several proteolytic enzymes present in the mouth deriving from oral flora [61,62]. Since they are a myriad, reporting a complete list is impossible and in the follow- ing we will refer to excellent published lists. Concerning PRPs and particularly bPRPs, many recent mass spectrometric studies allowed to report many bPRPs fragments [23,43,46,52], with high affinity for the tooth enamel [52]. However, the observation of a recurrent XPQ C-terminal sequence in many fragments detected induced the group of Oppenheim and Helmerhorst to characterize a glutamine endopeptidase from Rhotia sp [63]. Using the synthetic substrates KPQ-pNA and GGQ-pNA, the overall K(m) values were determined to be 97 +/.7 and 611 +/ micromolar, respectively, confirming glu tamine endoprotease activity in whole saliva and the influence of the amino acids in po sitions P(2) and P(3) on protease recognition. The pH optimum of KPQ-pNA hydrolysis was 7.0, and the activity was most effectively inhibited by antipain and 4-(2-aminoethyl) benzene sulfonyl fluoride. The enzyme is metal ion-dependent, and not inhibited by cys- teine protease inhibitors. A systematic evaluation of enzyme activities in various exocrine and non-exocrine contributors to whole saliva revealed that the glutamine endoprotease derives from Rhotia and it is localized in dental plaque [63,64].
A further protein submitted to fragmentation in the mouth is the polymeric immu- noglobulin receptor (PIgR), a type I transmembrane glycoprotein playing the main role in the adaptive immune response on mucosal surfaces [65,66]. It transports polymeric IgA across mucosal epithelial cells. A proteolytic cleavage occurring in the glycosylated extra- cellular portion of pIgR generates the secretory component (19–603 residues), which has been detected also in human saliva [67]. The cleavage occurs by action of unknown pro teases, probably released by activated neutrophils [66], and the highly conserved se quence 602–613 (PRLFAEEKAVAD) is believed to be the cleavage signal [65]. Two pep tides deriving from PIgR are detectable in saliva by a top-down proteomic approach, they are named AVAD and ASVD [68]. The peptide named AVAD originates by the cleavage occurring in this region at the level of Lys609, and the ASVD peptide derives from AVAD by the trypsin-like cleavage at Arg622. AVAD and ASVD peptides do not derive from the secretory component, and have a sequence partially overlapped to the transmembrane portion (639–661) of PIgR. Thus, they should originate by degradation of PIgR after its release from disrupted cell membranes.
2.3. Proteolytic cleavages: variations related to age and pathologies
Interestingly, it was observed that several proteolytic cleavages change according to the age and in several diseases. For instance, as previously reported in section 2.1.6, Iava rone and coll. [60] detected in saliva of human preterm newborns sensible amounts of cystatin B fragments 1-53 and 54-98, suggesting the presence in foetuses of high-active specific proteolytic events disappearing in the adults. The cleavage involves a tyrosine and phenylalanine couple (Tyr↓Phe) and it can be ascribed to a chymotrypsin-like enzy matic activity with strict specificity as suggested by the very precise consensus sequence necessary for the cleavage. The two fragments were detected at very high relative amounts with respect to intact cystatin B in very pre-term newborns, but its concentration de creased as a function of the post-conceptional age (PCA) and they were practically unde tectable when the age corresponded to that of full-term newborns. A search on the Merops data bank (http://merops.sanger.ac.uk/) returned various possibilities other than chymo trypsin A, such as chymosin, cathepsin E, metalloproteinase 2, ADAMTS4, endothelin converting enzyme 1, and some peptidases of the chymase class (mast cell chymotrypsin like proteinase). It is impossible to establish if cystatin B is a natural substrate of the en zyme, thereby implying a functional role for the fragments as potential cryptides, or rather if the fragments observed are by products, without any functional meaning, of a protein ase, whose activity is devoted to other specific foetal oral cleavage processes. Similarly, an increased activity of convertases and carboxypeptidases responsible for the cleavages of aPRPs, histatins and statherin was observed in preterm newborns with a low PCA of ap proximately 190 days. The activity decreased according to the PCA, and it reached the levels observed in the adult around the normal term of delivery [69]. Interestingly, this behaviour was observed for the release of the PRP3 type aPRPs and P-C peptide from PRP-1 type aPRPs, as well as for the release of histatin 6 and histatin 5 from histatin 3 and for the release of the fragments of statherin missing C-terminal residues, but not for sta therin des-D1, which showed low levels or was not detectable in preterm newborns at low PCA, but increased after birth reaching values similar to those determined in at-term new borns [70]. Because statherin desAsp1 is a fragment of statherin not detectable in granules preparations [6] this result is a further clue that the proteinases (convertases and carbox ypeptidases) involved in the cleavages of these families of salivary proteins are confined in the Golgi apparatus.
It has been reported that proteolytic fragmentations of salivary proteins may vary in some pathological states compared to physiological conditions. For instance, the study performed on a group of children and adolescent affected by type-1 diabetes revealed that several small peptides, most likely originated by post-secretory proteolytic cleavages oc- curring in the mouth, showed a higher concentration with respect to sex- and age-matched controls suggesting an increased activity of exogenous proteinases in the oral cavity of diabetics [70]. The peptides, characterized as fragments 1–14, 1–25, 5–25, 26–35, 26–44, and 36–44 of P-C peptide, have been already detected in saliva from healthy subjects [63], and fragments 1–14 e 26–44 were also detected in parotid saliva of healthy subjects by Hardt and coll. [71]. Furthermore, an association of fragments 1–14 and 26–44 to high numbers of dental caries and the presence of fragment 1–14 in saliva of subjects affected by Sjogrens syndrome have been demonstrated by Huq and coll. [72]. A top-down proteomic study investigated the salivary proteome of 49 multiple sclerosis (MuSc) patients and 54 healthy controls quantifying 119 salivary peptides/proteins [73]. Among the observed differences, the fragments 1–14, 26–44, and 36–44 of P-C peptide, SV1 fragment of statherin, and cys- tatin SN des1–4 showed higher levels in patients with respect to controls.
In a study performed to characterize possible differences of the salivary proteome of subjects affected by the Wilson’s disease (WD) with respect to healthy controls, increased levels of the AVAD and ASVD fragments of PIgR (see above) were observed in patients probably because of an increased disruption of cell membranes due to the high production of ROS typical of WD [68]. Higher level of ASVD peptide was also determined in MuSc patients with respect to healthy controls [73]. Interestingly, a study performed on patients affected by systemic mastocytosis (SM) and grouped into SM with (SM+C) or without (SM-C) additional cutaneous lesions evidenced that the two SM forms were distinguished by the lower levels of PRP-3, PRP-3 desArg106, statherin desPhe43, P-B des1-5, and cystatin D des1-5 and des1-8 in SM-C patients with respect to SM+C [74]. It should be outlined that 460 lower levels of cystatin D des1-5, and des1-8 have been also observed by comparing saliva of healthy elderly subjects with respect to adults [75].
The study performed on patients affected by Autoimmune hepatitis (AIH) and pri mary biliary cholangitis (PBC), two autoimmune liver diseases characterized by chronic hepatic inflammation and progressive liver fibrosis, to establish a panel of salivary pro- teins/peptides able to classify with good accuracy PBC patients vs HCs, AIH patients vs HCs, and PBC vs AIH patients, among the other data revealed significant different levels of PRP-3 desArg106 by comparing the two patient groups [76].
Higher levels of statherin desPhe43, and two others naturally occurring fragments (des1–9, and des1–13) were measured in saliva of a group of Alzheimer disease patients compared to controls [77].
3. Phosphorylation
Phosphorylation is probably (after proteolytic cleavage) the most common PTM in human saliva. Generally, phosphorylation is a reversible PTM, catalyzed by more than 500 different human protein kinases, while de-phosphorylation is due to enzymes called phosphatases [78]. The detection of phosphorylated proteoforms of human saliva is strongly dependent on the proteomic pipeline utilized: bottom-up or top-down. Com- monly, the bottom-up strategies are always accomplished by enrichment capture in order to increase the number of phosphorylated fragments.
A study of the group of Oppenheim and Helmerhorst [79] utilized a chemical deri vatization using dithiothreitol (DTT) of the phospho-serine/threonine-containing pep tides obtained after trypsin digestion of whole saliva samples. The DTT-phospho-peptides were enriched by covalent disulfide-thiol interchange chromatography and analysis by nanoflow liquid chromatography and electrospray ionization tandem mass spectrometry (LC-ESI-MS/MS). The specificity of DTT chemical derivatization was evaluated separately under different base-catalyzed conditions with NaOH and Ba(OH)2, blocking cysteine res- idues by iodoacetamide and enzymatic O-deglycosylation prior to DTT reaction. Further analysis of whole saliva samples that were subjected to either of these conditions provided supporting evidence for phosphoprotein identifications. The combined chemical strate- gies and mass spectrometric analyses identified 65 phosphoproteins in whole saliva; of these, 28 were based on two or more peptide identification criteria with high confidence and 37 were based on a single phospho-peptide identification. Most of the identified pro- teins (∼80%) were previously unknown phosphoprotein components.
A phospho-proteomic study based on a bottom-up pipeline on human saliva gener- ated a large-scale catalog of phosphorylated protein fragments too [80]. To circumvent the wide dynamic range of phosphoprotein abundance in whole saliva, the proteomic plat form combined dynamic range compression using hexapeptide beads, strong cation ex change HPLC peptide fractionation, and immobilized metal affinity chromatography prior to mass spectrometry. In total, 217 unique phospho-peptides sites were identified representing 85 distinct phosphoproteins at 2.3% global FDR. From these peptides, 129 distinct phosphorylation sites were identified of which only 57 were previously known. Cellular localization analysis revealed salivary phosphoproteins had a distribution like all known salivary proteins, but with less relative representation in “extracellular” and “plasma membrane” categories compared to salivary glycoproteins. Sequence alignment showed that phosphorylation is mainly linked to the action of the Golgi casein kinase called Fam20c (see below), but occurred also at acidic-directed kinase, proline-directed, and basophilic motifs. [80].
The top-down pipelines are more conservative, giving precise information on the sal- ivary protein substrate of kinase and on their sites of modification. All the phosphorylated proteins pertaining to the secretory pathway are phosphorylated by a pleiotropic Golgi casein kinase which, until few years ago, has been elusive. An interesting study performed by Tagliabracci and coll. [81,82] was able to establish that the enzyme is the kinase named Fam20C. The main consensus sequence of Fam20C is a serine with a +2 specific negative residue, either glutamic acid or phospho-serine (SXE/S(phos)). The salivary proteins and peptides submitted to the action of Fam20C in agreement with this consensus sequence are reported in Table 3.
Phosphorylation of cystatin S, whom N-terminal sequence is S1SS3KE5… is interest- ing. Mono-phosphorylated cystatin S is called cystatin S1, and due to the presence of the glutamic acid residue in position +2, Ser3 is necessarily the first site of phosphorylation. Di-phosphorylated cystatin S is called cystatin S2 and the second phosphorylation at Ser1 is strictly hierarchical, occurring only after that of Ser3. Cystatin S1 is the most abundant in adult human saliva and the approximated ratio between the relative percentages of the three components (S, S1 and S2) is 5:80:15, respectively [58,83]. A more complex situation concerns the phosphorylation of aPRPs, which are commonly di-phosphorylated having Ser8 and Ser22 as the two main sites of phosphorylation [9,84]. However, by mass spec trometry it is possible to detect also in small amount non-, mono- and three-phosphory lated aPRPs (on Ser8, Ser22 and Ser17) [9] (Figure 2). The consensus sequence recognized for Ser8 phosphorylation is the canonical S8QE, with a negatively charged residue at position +2, while the phosphorylation of Ser22 by Fam20c is ensured by the recognition of the sec- ondary consensus sequence S(X)3-4 (D/E/S(phos))3 [84]. To emphasize that phosphorylation of Ser22 allows the hierarchical phosphorylation of Ser17 which is located in the sequence …S17DGGDS22EQFIDEE…[9]. Hence, the mono-phosphorylated aPRPs can be phosphor ylated either on Ser8 or on Ser22 (with a different proportion in favour of Ser8) [9]. The di phosphorylated components (the most abundant) are commonly phosphorylated on Ser8 and Ser22, but a very small percentage of aPRPs phosphorylated on Ser17 and Ser22 could be present. The hierarchical phosphorylation on Ser17 is further substantiated by a study characterizing the aPRP-1 Roma-Boston Ser22(phos)→Phe variant, which was never de- tected as di-phosphorylated proteoform, also using high resolution HPLC-MS apparatus [85]. Statherin is mainly di-phosphorylated on Ser2 and Ser3 but being the N-terminal se- quence DS2S3EE… the two phosphorylation sites are independent and the phosphorylated Ser of mono-phosphorylated proteoform, always detectable in adult human saliva, can be either Ser2 or Ser3.
The N-terminal sequence of Hst1 is DS2HE… and about 90% of the peptide is phos- phorylated on Ser2, 10% of the non-phosphorylated peptide is detectable in adult human saliva [49]. Even though the Fam20C consensus sequence was not respected, a study of Halgand [86] and coll., carried out with a top-down pipeline evidenced a second minor phosphorylation site on Ser20.
The canonical consensus sequence of Fam20C is responsible for the phosphorylation of several bPRPs too. Indeed, the IB-1 and II-2 proteoforms are both phosphorylated on Ser8. Among the gPRPs Gl-1, Gl-2, Gl-3, GPA, II-1, and Cd-IIg, which have an N-terminal sequence motif similar to IB-1 and II-2, ((E/Q)XXXEDVS8QEES…, where XXX is LNE in IB-1, II-2, Gl-1, Gl-2, and Gl-3 and SSS in GPA, II-1, and Cd-IIg) Gl-2 is phosphorylated on Ser8 [23]. Phosphorylation is an almost complete event because <1% of the non-phosphor- ylated forms can be detected in parotid granules, parotid, and whole saliva and probably occurs after the cleavage of the proprotein [6]. It can be supposed, by sequence similarity, that also Gl-1 and Gl-3 undergo the same PTMs, although experimental evidence is miss- ing. The presence of the …S8QE consensus sequence of GPA, II-1, and Cd-IIg (Group 2C of bPRPs) suggests phosphorylation of Ser8 for these bPRPs, too, even though these mod- ifications have not been experimentally evidenced until now. A second potential, but not demonstrated, phosphorylation site at Ser3 is present in the sequence of Group 2C bPRPs, (<ESS3SED….). The activity of other kinases can be revealed in human saliva by using a top-down proteomic approach, among them the MAPK14, a kinase pertaining to the p38 mitogen-activated protein kinase pathway, which can partly phosphorylate the protein S100A9 on the penultimate Thr residue of its sequence [87].
3.3. Variation of phosphorylation as a function of age and for the diagnosis of different diseases
Some bottom-up studies suggested that the analysis of the phospho-proteome of sal- ivary extracellular vesicles could offer a possibility for the diagnosis either of the lung cancer [88] or to distinguish oral squamous cell carcinoma patients from healthy individ- uals [89]. Our studies were able to evidence that, during the last months of foetal growth the phosphorylation of secretory salivary proteins and therefore the activity if Fam20C is very low, if not completely absent [90]. It increases slowly reaching the level observed in the adult few weeks after the normal time of delivery [91]. Interestingly, a top-down study of the salivary proteome performed in a group of children with autism spectrum disorders evidenced significant lower phosphorylation levels of four salivary peptides, in compari son with age and gender matched healthy controls, giving a clue of the molecular patho genesis responsible for these disorders [92].
4. Sulfation
Sulfonation (or sulfation) is usually an irreversible PTM. It consists in the transfer of a sulfate group (-SO3-1) from the only known sulfate donor, i.e. 3-phosphoadenosine-5 phospho-sulfate (PAPS), to endogenous substances such as proteins, carbohydrates, cate cholamines as well as estrogenic steroids and xenobiotics [93]. 3’-phosphoadenosine-5’ phosphosulfate synthase (PAPSS) is the enzyme responsible to biosynthesize PAPS dur ing two reactions: inorganic sulphate is first converted to adenosine-5-phosphosulphate (APS) by ATP sulfurylase (EC 2.7.7.4) and this intermediate molecule is then phosphory- lated by the APS kinase (EC 2.7.1.25) to form PAPS [94]. Both APS and PAPS are activated sulphuryl donors that possess a phospho-sulphate anhydride bond [95]. In prokaryotes, fungi, and plants, synthesis of PAPS is performed by two separate enzymes [94]. In the animal kingdom, however, the ATP sulfurylase and the APS kinase are encoded by the same gene and translated into a single polypeptide which forms the dual-functional en- zyme PAPSS [95]. PAPSS1 and PAPSS2 are two characterized isoforms of this enzyme according to their different localization. Moreover, relation of various pathological condi- tions to deficiencies of PAPSS (both isoforms) has been demonstrated [96]. While PAPSS1 and PAPSS2 are responsible for the bioactivation of sulphate, sulfo-conjugation reactions are catalysed by enzymes known as sulfotransferases [97]. Sulfotransferases are mainly divided into two groups, as they are either cytosolic or membrane-bound [98]. Cytosolic sulfotransferases constitute the superfamily of enzymes known as SULTs which are in volved in the sulfonation of xenobiotics and small endogenous compounds such as neu rotransmitters and hormones [99]. The membrane-bound sulfotransferases are found in the Golgi apparatus and are responsible for post-translational sulfation of endogenous 605 macromolecules such as proteins, lipids, and glycosaminoglycans [100]. Currently, 12 SULT isoforms have been identified and detected in human tissues [101]. Availability of PAPS in different tissues can highly affect the sulfation pathway by modifying the affinity of sulfotransferases [100,101,102].
In human saliva we were able to determine that Hst1 is partly sulfated on the last four tyrosines (out of five) of its sequence [103] and until now it is the unique phospho- sulfo-peptide detected in human saliva. As previously reported, phosphorylation of Hst1 is not a complete event, because in whole human saliva it is possible to detect about 10% of the non-phosphorylated peptide. Further studies performed by high-resolution HPLC MS apparatus suggested that: a) as supposed in our work, the sulfation process is hierar chical, being Tyr27 the first residue sulfated, followed by Tyr30, Tyr34 and Tyr36. Indeed MS CID fragmentation data on the mono-sulfated (non-phosphorylated) Hst1 clearly indi- cated Tyr27 as the unique residue involved c) the sulfation process is strictly confined to the submandibular gland, because sublingual gland does not express Hst1 (manuscript in preparation) and Hst1 secreted by parotid gland is not sulfated; d) the phosphorylation and the sulfation processes are independent, because, by the study of the neutral losses generated during the MS/MS CID induced fragmentation it is possible to discriminate non-phosphorylated poly-sulfated derivatives from the phosphorylated ones; f) the per centages of the different polysulfated derivatives varies sensibly in human submandibu lar saliva of human g) sulfation of Hst1 is not detectable in children till the puberty; h) potential changes in physio-pathological conditions as well as during pharmacological treatments require extensive statistical analysis on a large population.
5. S-modifications
The thiol side chain of cysteine residues present in the proteins is characterized by high redox sensitivity. This redox sensitivity derives from the pKa of the thiol group, which due to the environment of the protein, even at physiological pH may be partly pre- sent in the thiolate form (RS-) most reactive than the thiol form (RSH). Indeed, RS- is prone to donate an electron pair to target reagents generating the oxidized derivative sulfenic acid (RSOH), mainly present as sulfenate (RSO-) by a reversible reaction. Then, sulfenate may undergo other reversible, and irreversible oxidative modifications (Figure 5) [104]. For instance, it can rapidly react with another thiol (RSH) by generating the corre sponding disulfide (RSSR), or with GSH or free Cys to form S-glutathionyl (RSSG) or S cysteinyl derivatives. This is not the only formation pathway for disulfide derivatives, since the latter can also be generated in a process termed thiol-disulfide exchange, in which a thiol form reacts with a disulfide to generate a different thiol and disulfide.
In the presence of strong oxidants, sulfenate can undergo further oxidation and gen- erate sequentially the sulfinic (RSO2H) and sulfonic acid (RSO3H) derivatives. The latter two modifications are regarded as irreversible and associated with oxidative damage. Sul- fenate can also react with hydrogen sulfide (HS-) to form a cysteinyl persulfide (R–SS-), which can be oxidized to a cysteinyl thiosulfate(R–SSO3-). The thiol side chain of cysteine may also react with nitric oxide resulting in the formation of nitrosothiol derivatives. [105]. These modifications have been observed and characterized on specific salivary pro teins as cystatins and S100A proteins.
Cabras and coll. evidenced that in human whole saliva cystatin B is present mostly as S-modified derivatives on Cys3, being the S-unmodified proteoform rarely detectable in saliva of healthy adults. Generally, more than half of cystatin B (55%) is found S-gluta- thionylated, 30% is present in the dimeric form, and the remaining 15% is S-cysteinylated [106]. In an in-depth study, performed both in the acidic supernatant of whole saliva and in enriched fractions obtained by preparative RP-HPLC, the structures of all these S-mod ified proteoforms of cystatins were confirmed by an integrated top-down/bottom-up pro teomic approach [57]. In the study, a carboxymethylated Cys3 derivative of cystatin B was also detected and characterized. Carboxymethylation, like the formation of sulfinic and sulfonic acid derivatives, is a nonenzymatic irreversible and stable modification generated by the endogenously formed glyoxal with Cys sulfhydryl groups (Figure 7). This modifi cation was novel not only for cystatin B but also for any other human salivary protein. It should be outlined that Cys residue present in the N-terminal region at C3 position of cystatin B is involved in the formation of these and other derivatives. The N-terminal re gion of cystatin B is crucial for the biological function of the protein, being involved in the binding of cysteine proteinases [107]. Indeed, it has been shown that Cys3 is the most im portant residue for the interaction with papain and cathepsin H and is also a major con tributor to cathepsin L binding [108]. Differently from adults, the S-unmodified cystatin B represented the main proteoform in saliva of preterm newborns. Interestingly, it was ob served that the high relative amount the unmodified cystatin B observed in preterm new borns decreased as a function of the PCA, reaching at the normal term of delivery values 675 like those determined in at-term newborns, children, and adults [60]. As reported in sec tion 1.3 describing the proteolytic cleavages, in the same study high relative amounts of fragments 1−53 and 54−98 of cystatin B were detected in very preterm newborns. The N terminal 1-53 fragment was detected both as unmodified and modified at the level of Cys3 residue, being the percentage of the different forms like those determined for the intact protein [60].
S100 proteins constitute the largest family of calcium binding proteins with more than 3000 related entries in the NCBI Reference Sequences Data Bank. They are EF-hand calcium binding proteins and bind calcium via helix-loop-helix motifs often present in multiple copies. The “S100” name originates from the solubility of the first identified S100 proteins in 100% ammonium sulfate solution. The members of the EF-hand superfamily can be divided according to their calcium affinity and their ability to change conformation following binding of calcium. Intracellular functions of S100 proteins include: i) regulation of protein phosphorylation by interaction with the substrates of the kinases, thus they play a role in signal transduction; ii) regulation of enzyme activity [109].
In the extracellular milieu, S100 proteins do not function as Ca2+ sensors as they are saturated by the mM range Ca2+ concentration, however they are recognized to play an important role in mediating inflammatory responses through the activation of several cell surface receptors, after release from activated or necrotic cells [110]. S100A8, S100A9, and S100A12 are constitutively expressed in high amounts in neutrophils and are inducible in macrophages, cells that generate high amounts of ROS, while expression of the complex S100A8/S100A9 (calprotectin), S100A12 and S100A7 can be induced in keratinocytes, en- dothelial cells, and epithelial cells during inflammation [111].
Human S100A9 is encoded by a single copy gene with two isoforms: full-length and truncated S100A9, which is translated from an alternate start site at codon 4 of the full- length form and lacks the single Cys3 residue, making it less susceptible to oxidation. In the studies of Lim and coll. [112,113] the many pro-inflammatory functions described for S100A8 and S100A9, as well as its anti-inflammatory roles in wound-healing and protec tion against excessive oxidative tissue damage, are discussed and an explanation that ox idative modifications may act as a regulatory switch for the disparate, functional roles of S100A8 and S100A9 is suggested.
As far as it concerns biofluids, in a series of studies carried out to investigate possible salivary biomarkers of pathologies both confined to the oral cavity and systemic, different oxidized derivatives of S100A8 and S100A9 proteins were revealed and characterized both in healthy and diseased subjects. Structure of these new proteoforms was established by high-resolution HPLC-ESI-MS/MS analysis of the mixture of peptides obtained by trypsin digestion of salivary fractions enriched with S100A8 and S100A9 oxidized derivatives [68]. In this study the long form of S100A9 was detected as glutathionylated at Cys3 in 15/32 subjects and cysteinylated only in 3/32, while S100A8 was sporadically detected as unmodified S100A8 (4/32), and as sulfonic derivative at Cys42 (5/32).
Glutathionylated and cysteinylated S100A9 derivatives were also detected in saliva of human preterm newborns [114]. Finally, the nitrosylated derivative of S100A8 was also observed in saliva from adult healthy subjects, but sporadically [73,74].
5.1. S-modifications of salivary proteins in pathologies
A wider variety and abundance of S-modified proteoforms has been revealed in sa- liva of subjects affected by various pathologies. For instance, a study performed on saliva collected from patients with schizophrenia and bipolar disorder, compared to healthy non-smokers and smokers control groups, revealed more than 10-fold increase in salivary levels of S-cysteinylated and S-glutathionylated cystatin B, in addition to α-defensins 1-4, S100A12, and cystatin A, suggesting dysregulation of the peripheral white blood cell im mune pathway associated with the pathologies [115]. S-glutathionylated cystatin B was 726 found at high levels also in Predominantly Antibody Deficiencies [116].
In WD patients, top-down proteomic analysis of saliva revealed significant higher levels of S100A9 and S100A8, and some of its oxidized proteoforms, with respect to con- trols [68]. S100A8 oxidized at Cys42 to sulfinic acid (S100A8-SO2H) was detected only in patients, while the sulfonic derivative (S100A8-SO3H) oxidized also at Trp54 was detected both in patients and healthy controls, even if with different levels. The latter proteoform was subjected to further oxidation to give the so-called hyperoxidized S100A8, with the second oxidation located at either Met1 or Met78 (position to be determined). S100A8 was also shown to undergo glutathionylation, nitrosylation, and formation of a disulfide bridge with Cys3 of long S100A9 (S100A8/A9-SS dimer), but only in the patient group. In the same group for the first time the homodimer proteoform of long S100A9 was charac- terized. Overall, the salivary proteome of WD patients reflected the oxidative stress and inflammatory conditions characteristic of the pathology, highlighting differences that could be useful clues of disease exacerbation [68].
Oxidative cross-linking via disulfide bonds of S100A9 and S100A8 has been observed also in saliva from healthy adults, and in lavage fluid from the lungs of patients with res- piratory diseases by Hoskin et al. [117]. The authors showed that reactive halogen species promote cross-linking of the non-covalent heterodimer of S100A8/S100A9 and hypothe- sised that the cross-linking detected in the saliva samples was most likely mediated by hypothiocyanous acid produced by lactoperoxidase. They also demonstrated that for- mation of the disulfide cross linked derivative enhanced susceptibility to proteolysis by neutrophil proteases. Furthermore, Gomes et al. observed that sulfinic and sulfonic acid derivatives of monomeric S100A8, together with novel oxathiazolidine oxide/dioxide forms, were present in asthmatic sputum [118].
The salivary proteome of Alzheimer disease (AD) patients highlighted also elevated levels of some S100A8 and S100A9 oxidized proteoforms with respect to age and gender matched healthy controls. They were the hyperoxidized proteoform of S100A8, S100A8 SNO, and glutathionylated long S100A9 [77]. This finding was not surprising since oxida tive modifications of proteins are common in neurological disorders, due to the strongly oxidizing characteristics of the extracellular milieu because of generation of ROS and re active nitric oxide species [119]. Higher levels of S100A8-SNO were also determined in MuSc patients with respect to healthy controls while levels of mono- and di-oxidized cys tatin SN, mono- and di-oxidized cystatin S1, mono-oxidized cystatin SA were lower [73]. The reduced level in the patients of oxidized derivatives of S-type cystatins was an intri guing result. Indeed, high levels of oxidative stress markers and lower antioxidant status have been reported in saliva and plasma of MuSc patients under corticosteroid therapy by Karlik and coll. [120]. However, different results were obtained by Schipper and coll. [121] that highlighted a positive effect of the immunotherapy in oxidative stress evidenced by a reduction of oxidative stress markers in MuSc. Manconi et al. speculated that reduced S-type cystatin oxidation in MuSc patients could be related to the treatment of 32/49 pa- tients enrolled for the study. Indeed, S100A8-SNO was found at significantly higher level only in the group of untreated patients with respect to controls [73].
Recently, analysis of the salivary proteome of patients affected by AD compared to healthy adult and elderly controls highlighted significant higher levels of S100A8-SNO and hyperoxidized S100A8, as well as glutathionylated S100A9 (long) in healthy adults with respect to elderly, and in AD patients with respect to healthy elderly. The same trend was observed for both glutathionylated and dimeric cystatin B, but not for the cysteinyl- ated proteoform which showed a significant difference only by comparing patients and elderly subjects [75].
6. Transglutamination
Several salivary proteins are involved in the formation of proteins layers, i.e. the so called “oral pellicles” The general term for these layers is pellicle, but due to the different characteristics of the coated surfaces the enamel pellicle and mucosal pellicle are their own entities. These protein films have a dual role because one “the acquired enamel” is im portant for the integrity of tooth and the second “mucosal pellicle” for the protection of the oral mucosa. There is considerable information on the enamel pellicle: many proteins 782 and their fragments are involved with a particular concern for PRPs, statherin, histatins and P-B peptide [122,123]. On the contrary, only limited data are available on the mucosal pellicle [124]. This can be attributed to the difficult to develop a standardized preparation of this latter biological structure. However, it has a completely different ultrastructure as 786 compared with the enamel pellicle. Since it is comprised of larger glycoproteins retaining water, it might be considered as a hydrogel, and it appears to have a lower tenacity than the enamel pellicle. Maturation and turnover are influenced by the delivery of salivary proteins, by the flow of saliva and the underlying desquamating oral epithelium. Its prob able functions include lubrication and moisture retention. Furthermore, interactions be tween mucosal pellicle proteins and bacterial surfaces are responsible for specificity of the 792 bacterial colonization during the earliest stage of plaque formation [125]. The in vivo pel licle is thought to be an insoluble network of proteins generated by post-secretory pro cessing of proteins mainly due to cross-linking. Cross-links for the formation of oral mu cosal pellicle were demonstrated firstly by Bradway and coll., which highlighted the ex istence of a network of proteins formed by components of saliva adsorbed onto buccal epithelial cell surfaces that cover the oral mucosal surface [126]. Recently, a new proteomic protocol was optimized to investigate the proteins participating to the composition of the oral mucosal pellicle, among them proteins of the PLUNC family were identified [127].
The oral mucosal pellicle is a thin lubricating layer generated by the binding of saliva proteins on epithelial oral cells [128]. This protein molecular network interacts with the oral epithelial-cell plasma membrane and its associate cytoskeleton and contributes to the mucosal epithelial flexibility and turnover. It was demonstrated that acidic-proline-rich proteins, statherin, the major histatins as well mucins are substrates of oral transglutami nase 2 (TG2) and they participate in cross-linking reactions [126] as putative pellicle pre cursor proteins. Whatever the structure of these protein networks may be, oral transglu taminases (mainly type 2 transglutaminase) are the pivotal enzymes for pellicle formation. TG2 can cross-link acidic PRP-1 and statherin in vitro [122]. TG2 is the ubiquitous tissue enzyme expressed also in oral epithelial cells [129], which catalyzes different biological processes and generates a cross-link between two peptide chains, typically between ε amine of the lysine residue acting as lone-pair donor and a glutamine residue, the lone pair acceptor, and the reaction is accomplished by the loss of an ammonia molecule. TG2 is a Ca2+-dependent enzyme, released by the epithelial oral cells, negatively modulated by GTP [129] and affected by the reversible formation of an intramolecular disulfide bridge. [130]. Recently, a study of our group [131] showed that also bPRPs and P-C peptide are potential substrates of TG2. Nonetheless, they showed a very different reactivity for mon- odansyl-cadaverine (used as lone pair donor). Mass spectrometry analyses of the reaction products highlighted that P-C, P-H and P-D (both Pro32 and Ala32 variants) peptides were active substrates of TG2, II-2 was less reactive, while P-F and P-J peptides showed negli- gible activity [131]. MS characterization suggested that the consensus sequence for the linking is connected more to the environment of glutamine residue, than to the donor ammine. The pivotal residues characterized for P-H, II-2, and both variants of P-D evi denced …GNPQ… as the consensus sequence recognized by TG2 on bPRP peptides [131]. This consensus sequence is not present in P-F and P-J peptides and, probably for this rea son, they were poor substrates for TG2.
P-C, P-H, P-D peptides formed cyclo-derivatives after TG2 reaction, and only specific glutamine residues were involved in the cycle formation and reacted with specific mono- dansyl-cadaverine [131]. The stereospecificity of TG2 was at first recognized on statherin, which under the action of TG2 forms a cyclic derivative involving almost only the Gln37 (out of 7 glutamine residues) and Lys6 as unique lysine residue in the sequence (Figure 4) [132]. The detection of small amount of cyclo-statherin in adult human whole saliva [132] and the high reactivity of secondary glutamine residues after the formation of cyclo-sta therin and cyclo-P-C were suggestive for the in vivo formation of ring structures with a pivotal role in the architecture of the oral mucosal and enamel pellicles [133,134]. What ever the molecular mechanism towards the structure of the oral pellicles, they have a rel evant role in the protection of the mucosal from the mechanical and thermal high stresses that the mouth undergoes during the human life.
7. Glycosylation
7.1. Non enzymatic glycosylation (glycation)
Non-enzymatic glycosylation or glycation is a ubiquitous process involving proteins, peptides, lipids, and nucleic acids. It consists in a nucleophilic addiction of a sugar (com- monly glucose) to the free amino group of a biological molecule (in protein, typically the ε-amino group of lysine). First studies regarding glycation processes were carried out by food industry in quality control of the foodstuffs [135] and only forty years later, this pro cess was observed in human body monitoring the trend of glycated hemoglobin in dia betic patients [136,137,138]. Glycation process starts with formation of reversible but unstable Shiff’s base, followed by intermolecular rearrangement into stable Amadori’s products. When large amounts of Amadori products are formed, they rearrange to form a heteroge neous group of protein-bound moieties, termed as Advanced Glycated End products (AGEs) [139]. Fluorescent spectroscopy is usually used to monitor the rate of formation of AGEs considering that most of the advanced products of glycation are fluorophores [140]. 852 Rate of these reactions are quite slow and only proteins with large amounts of lysine res idues undergo glycation with significant amounts of AGEs leading to reduced protein functionality and stability [141,142].
Proteins with a long metabolic turnover, as extracellular matrix proteins, are the most susceptible to non-enzymatic glycosylation, compared to most proteins that counteract the impact of glycation thanks to the high turnover rate and short half-life [143]. For in- stance, the fast hematic turnover of serum albumin offers chance to monitor short term diabetes [144]. It’s known that proteins from saliva show a very quick turnover, neverthe- less few studies point out how the loss of functional activity of some salivary proteins is to be related to glycation processes. The antibacterial activity of lactoferrin or lysozyme and the antioxidant capacity of superoxide dismutase, all detectable in human saliva, de- creased after the formation of AGEs [142,145,146].
AGEs accumulate into the human body by binding to AGE-specific receptors (RAGEs) which play an important role in the development of age-related organ hypofunction [139]. The presence of RAGEs was also evidenced in the oral cavity, particularly in the minor salivary glands [147]. Manig and coll. in 2019 [148] recently carried out the first study focused on monitoring the free salivary glycated compounds predominant during later reaction stages of AGEs production in saliva. Specifically, by LC MS-MS analysis, the concentration of Nɛ-carboxymethyllysine (CML), pirralyne (Pyr), methylglyoxal-derived hydroimidazolone 1 (MG-H 1) and Nɛ-carboxyethyllysine (CEL) was evaluated [148]. The presence of AGEs in saliva increases the risk of developing periodontitis and pulpitis [147]. Furthermore, the expression of RAGEs appeared to be correlated within the histo- logic differentiation of oral squamous cell carcinomas [149]. Maciejczyk M. and coll. [150] evidenced the role of saliva, as diagnostic biofluid alternative to blood for assessing the severity of protein glycation, correlated to the aging processes. Several proteomic studies, mainly performed by top-down mass spectrometry approaches, have shown that saliva can be a mirror of the aging process. Indeed, age-related changes in the salivary proteome profiles have been observed in individuals from 180 days to elderly [75]. Among the sali- vary proteins and peptides affected by the aging process, S100A8 and S100A9 have been found to participate in the toll-like receptor-4- or RAGE-mediated multiple inflammatory pathways [75]. In fact, S100A8 and S100A9 represent the main protein content of the neu- trophils and actively participate in modulating the inflammation [151], by acting also as ROS/RNS scavengers. The maintaining of normal salivary function is important for a healthy oral environment. In this context the study of glycated proteins and peptides in saliva could be, given the low-invasiveness and easy collection of this biofluid, a potential tool to study, aging process as well as diseases progression or patient follow-up during treatments [152].
It is relevant to remark that improper sample manipulation can generate artefactual detection of non-enzymatic glycation of proteins can be generated by improper sample manipulation. For instance, freeze-drying of salivary solutions containing glucose and/or fructose can generate artefactual amounts of glycated proteins [22].
7.2. Enzymatic glycosylation
Enzymatic glycosylation is one of the most common and prominent PTMs, modulating the activity of cellular and extracellular proteins, implicated in complex and articulate physiological processes [153,154] such as cell adhesion, signaling and cell-to-cell interac tion, to cite a few, and many proteins detectable in human saliva are enzymatically glyco sylated. The two principal types of protein glycosylation in humans are N- and O-glyco sylation, while C-mannosylation of tryptophan occurs rarely with function still not well understood [155].
N-glycosylation occurs as a co-translational modification on the Asn residue at the Asn-X-Ser/Thr sequon, even though some recent glycoproteomic studies evidenced that several N-glycosylation sites do not adhere to this canonical consensus sequence [156]. Glycan structure is not under direct genetic control, rather it results from the cooperative and competitive interactions between several glycosyltransferases. These enzymes trans- fer a monosaccharide unit from an activated sugar donor (frequently a nucleotide-sugar) to an acceptor (such as an amino acid, a lipid, or another sugar). Consequently, glycosyl- ation must be considered a stochastic rather than a deterministic process. The biological role of glycans is frequently mediated by sugar-binding molecules (lectins) which, upon recognition of specific carbohydrate structures, trigger a broad range of cellular effects, including proliferation, apoptosis, and cell migration [157]. Galectins (beta-galactoside binding proteins) and siglecs (Sialic acid-binding immunoglobulin-type lectins) are among these sugar-binding molecules. The activities of specific galectins and siglecs can be inhibited or promoted by non-coding RNAs (ncRNAs), as those implicated in tumor invasion and cell proliferation [158], as well as of many glycosyltransferases [158]. Indeed, ncRNAs form an extremely complex non-deterministic network of gene expression regulation, in which have a key role in modulating the expression of several glycosyltransferase genes, especially promoting those ones implicated in the progression and invasion of various kind of tumors [158]. Rlevant role of several classes of ncRNAs is nowadays recognized in regulating both physiological and pathological mechanisms, moreover, the ncRNA aberrant expression appears to be associated to different human disorders, such as reproductive disorders and cancer [159]. The term ncRNAs comprises a wide and heterogeneous group of RNA molecules differing for length, biological function, and cellular localization. ncRNAs are classified, based on their size, in long-ncRNAs (lncRNA, >200 nucleotides) and short-ncRNAs (<200 nucleotides), which include, among the others, microRNA (miRNA). Circulating miRNA are largely studied as the Next-Gen class diagnostic and prognostic biomarkers. lncRNAs are tissue-specific and can directly interact with other macromolecules, such as DNA, RNA and protein, moreover, they can regulate gene expression, both at the transcriptional and the post-transcriptional level. In addition, glycosylation has been demonstrated, on the other hand, to modulate ncRNA expression in some instances. Finally, short-ncRNAs have recently been shown to undergo canonical N-glycosylation [160]. These glyco-RNAs bear terminal sialic acid and fucose residues and can interact with sugar-binding molecules, such as siglecs. Thus, ncRNA network and glycosylation can be considered as two stochastic mechanisms affecting the role of proteins: the first and the second acting before and after the protein axis biosynthesis, respectively. The N-linked glycans present a common penta-saccharide core to which different types of monosaccharide residues can be linked in different positions, generating diverse classes of branched glycans [161]. O-glycosylation occurs on either Ser or Thr residue exposed on the protein surface. The O-linked glycans can be very simple and are constituted by one or few monosaccharide residues, such as N-acetyl-galactosamine, mannose, glucose, and fucose. The glycosylation of salivary proteins has been studied several years ago by bottom-up pipeline identifying about 1500 glycosites [162].
Due to the stochastic mechanism of synthesis above described, glycoproteins are characterized by high microheterogeneity, because each glycoprotein may exist as a com plex family of glycoforms sharing the amino acid sequence and differing for various fea tures: (a) occupancy of the multiple glycosites; (b) type of glycan (only N-linked, only O linked, N- and O-linked on the same protein); (c) composition and stereoisomery of the glycans that can change on the same glycosite.
We were able to characterize by an integrated top-down/bottom-up strategy the structure of six glycoforms of IB-8a (Con 1+), so called because interact with concanavalin A [163]. MS analyses on the intact glycoproteins before and after N-deglycosylation with PNGase F and MS/MS sequencing of peptides and glycopeptides from tryptic digests al lowed the structural characterization of the glycan moieties and the polypeptide back bone, as well as to establish the glycosylation site at the asparagine residue at 98th position. Five of the glycoforms carry a biantennary N-linked glycan fucosylated in the innermost N-acetylglucosamine of the core and showed from zero to four additional fucoses in the antennal region. The sixth glycoform carries a monoantennary monofucosylated oli gosaccharide. The glycoform cluster was detected in 28 out of the 71 adult saliva speci mens analyzed [163].
The level of fucosylation showed interindividual variability with the major relative abundance for the tri-fucosylated glycoform. Non-glycosylated IB-8a (Con 1+) and the var iant IB-8a (Con 1−), lacking the glycosylation site for the substitution, have been also de tected in human saliva.
The surprising detection of the gPRP protein 3M (encoded by the PRB3 human gene) in the saliva of preterm newborns at an age when the other bPRPs are absent in human saliva [20,25,26] allowed to purify the 3M protein and to establish the structures of the in- tact proteoforms before and after N-deglycosylation with Peptide-N-Glycosidase F and MS/MS sequencing of peptides obtained after endoproteinase GluC digestion [162]. The heterogeneous mixture of the proteoforms derives from the combination of 8 different neutral and sialylated glycans O-linked to Thr50, and 33 different glycans N-linked to var- ious Asn residues at positions 66, 87, 108, 129, 150, 171, 192, and 213 of the 3M protein.
In a study carried out to evidence potential salivary diagnostic markers of MuSc [72] we were able to evidence that prolactin inducible protein exists in two glycosylated pro teoforms, one with a N-acetyl-hexosamine residue and the other with a further deoxy hexose moiety. Although the site of modification was not established, these two glycosyl ated proteoforms, were significantly increased in the patients with MuSc with respect to, healthy controls. Indeed, altered glycoprotein expression and glycosylation aberrations, consisting in increased glycan size, extra branching of glycan chains, over-sialylation, or over-fucosylation, were associated to many diseases, especially cancer and neurological disorders [165,166]. A recent magistral review [158] evidences the complexity of the rela tionships between enzymatic glycosylation and other factors and their molecular altera tions as potential diagnostic markers in many diseases. Therefore, glycoproteomic studies are of great interest for the development of disease biomarkers. Nonetheless, they are a complex and challenging field of research, requiring the evaluation of the glycoprotein profiling of a biological sample in physiological conditions and the characterization of the qualitative and quantitative changes related to a specific pathological condition [167,168].
8. Citrullination
Citrullination is a PTM consisting in the conversion of peptidylarginine to peptidyl- citrulline in a calcium-dependent reaction catalysed by peptidylarginine deiminases (PADs), a family of five isoenzymes (PAD 1-4 and 6) with tissue specific expression [169,170]. The reaction of PAD enzyme with the arginine residue of a peptide/polypeptide chain forms an adduct with release of ammonia and subsequent formation of a ketone functional group following the cleavage of the adduct by a water molecule. The result of the deimination reaction is the loss of a positive charge, i.e. the immino mojety, resulting in a generation of a neutral amino acid, the citrulline, lacking the strong basic character proper of arginine, and producing a delta mass increase of the protein/peptide molecular mass of +0.9840276 and +0.98476 Da, monoisotopic and average, respectively. Interest- ingly, the rate of peptidylarginine citrullination by PADs enzymes is depending on the amino acid position along the protein/peptide sequence, with about the 80-90% of citrul- linated arginines positioned after aspartic acid residues. Arginines close to glutamic acid or to the N-terminal sequence trait are largely citrullinated too [169,170,171].
The citrullination affects the physico-chemical properties of a protein because at neu- tral pH the positive charge of arginine is lost by the modification and causes changes of the overall charge and charge distribution of the protein, altering the isoelectric point, ionic bonds, protein structure, activity, and protein-protein interactions [169,170,171]. The in crease of hydrophobicity produced by citrullination allows to easily distinguish the un modified from the modified form of a small protein or a peptide by reverse phase liquid chromatographic separations. In addition, because citrulline is not included in the list of natural amino acids incorporated in proteins, it was hypothesized to possibly induce im mune response and suggested to deeply investigate its role in autoimmune inflammatory diseases such as rheumatoid arthritis (RA) [172]. In fact, citrullination PTM was studied in relation to various physio-pathological processes in addition to RA, such as apoptosis, multiple sclerosis, Alzheimer disease, and psoriasis, systemic lupus erythematosus, peri odontitis, COVID-19, cancer, and thromboembolism [169,170,171]. Citrullination has been the topic of a very recent review that outlines the diagnostic as well as the therapeutic ap proaches based on this PTM and the future perspectives of “citrullinome” characterization and disclosure [172]. Studies and clinical trials are nowadays based on the detection of anticitrullinated protein antibodies as high specificity tool for RA diagnosis also before clinical evidence [173,174].
Several are the proteins that are substrates of PAD enzymes and that can exhibit cit rullination PTM in physiological- as well as in inflammatory pathological states, and in clude filaggrin, keratin, fibronectin, actin, tubulin, vimentin, glial fibrillary acidic protein and histones [173]. PAD 2 is the isoenzyme acting in salivary and secretory glands, as well as in other cell types and tissues, targeting myelin basic protein, C-X-C motif chemokine 10 and 11, vimentin, actin, glial fibrillary acidic protein, S100-A3, histones H3 and H4 [173].
The importance of anti-citrullinated protein antibodies (ACPAs) detection in serum for the diagnosis of RA [173,175], has increased the interest of their application to saliva analysis for the characterization of citrullinated proteins.
Yasuda et al. identified citrullinated cytokeratin 13 in saliva samples of RA patients and healthy subjects by two-dimensional electrophoresis, silver staining and immunopre- cipitation-western blotting [175]. In the same paper the origin of pre- or post-secretion of citrullinated cytokeratin 13 in saliva is also discussed based on calcium concentration data inside salivary gland cells and secreted saliva since PAD enzyme catalysis is calcium de pendent. According to Genotype-Tissue Expression database PAD 1-4 and 6 calcium bind ing enzymes are reported to be expressed in human minor salivary glands [176]. Citrulli nation could take place either in saliva, i.e. post-secretion, because the biofluid contains calcium levels proper to PAD enzyme activity, or intracellularly depending on the calcium concentration increasing. Indeed, intracellular citrullination of histones and other proteins was recognized [177]. Citrullination of salivary proteins could therefore occur either in tracellularly, followed by secretion of the modified protein, or extracellularly in the bio fluid, after secretion.
A recent paper compared the levels of citrullinated proteins levels in saliva of RA patients with respect to healthy controls also studying their correlation with the periodon- tal status and temporomandibular joint associated disorder [178]. Confirming previous data [175], no differences in citrullinated protein levels were found between RA patients and healthy controls apart differences in periodontal status observed between groups. However, in RA the salivary anti-cyclic citrullinated peptide levels were in correlation with periodontal disease staging, considering that bacteria involved in its pathogenesis produce non-physiological citrullination by a family of PAD enzymes very similar to hu- man PADs [179].
A study investigated the influence of inflammation from periodontal disease in RA model mice analysis in serum and saliva samples of both anti-citrullinated protein anti- body analysis and citrullinated proteins by gel electrophoresis [180]. Saliva and serum showed similar results evidencing protein citrullination around 55 kDa molecular weight electrophoretic band. Periodontitis exacerbated RA symptoms, indicating a relationship between the Porphyromonas gingivalis periodontitis bacterial infection and RA.
Patients affected by Sjögren’s syndrome have been found positive for ACPAs and showed increased levels of PAD 2 enzyme, suggesting a possible role of citrullination in the disease that could be responsive for autoantigens triggering [180]. However, in the same study it was outlined that only approximately 7.5% of Sjögren’s syndrome patients are positive for ACPAs.
Although the poor knowledge on the protein pattern that underwent to citrullination in saliva, the indications of the high potential of this biofluid in the development of new high specificity and sensitivity diagnostic tools of low invasiveness targeting selected cit- rullinated proteins in different pathologies are clear. However, deeper research still needs to be done to decipher the saliva citrullinome associated to physiological as well as to pathological states and to clearly understand where the modification occurs, i.e. inside salivary gland cells or extracellularly or both. To the best of our knowledge, combined top-down/bottom-up proteomic platforms have never been applied to the study of saliva citrullinome. Their application coupled to high resolution mass spectrometry detection could finally characterize the proteins subjected to citrullination in saliva in a wide mo- lecular range and abundance, and, especially, could localize the modification inside the sequence and better define its molecular features.
9. N-terminal modifications
The N-terminal of a polypeptide chain is a structural site which must be often modi- fied and protected during the ribosome synthesis. The positive charge of the terminal amino group can be an obstacle for the proper structural folding of the protein which starts in the nascent protein [181]. The most common modifications in human are the ex- cision of the initiatory methionine (iMet), which is commonly carried out by enzymes called methionine aminopeptidases (MAPs). In any case the polypeptide chain, with or without iMet can be submitted to N-terminal acetylation induced by enzymes called N- terminal acetyltransferases (NATs). To date, 12 NATs have been identified, possessing different compositions, substrate specificities, and modes of regulation. In humans, NATs are present as group of six enzymes (called from NAT-A to NAT-F), with slightly different specificity [182].
Another possibility, less common, of N-terminal modification occurs when the ter- minal residue is either a glutamic acid or a glutamine, which can generate a cyclic amide, called pyroglutamic N-terminus, with the loss of a molecule of water (Glu) or ammonia (Gln), respectively. This PTM can proceed spontaneously, at reasonable rates, or by the action of enzymes called glutaminyl cyclases, in a reaction commonly much faster with N-terminal glutamines than with glutamate residues [183].
Our top-down mass spectrometry pipelines allowed to verify that all these three PTMs are largely detectable in human saliva, while C-terminal modifications are rarely detectable, probably because at the end of its synthesis the proteins have assumed a fold- ing close to the functional conformation. Few minor examples of C-terminal modifications of human salivary proteins will be reported at the end of this section.
9.1. Excision of initiatory methionine and Nt-acetylation
It is relevant to outline that when the methionine residue is at the first position of a leader peptide at turn responsible for the docking of the protein to the ribosome at the endoplasmic reticulum, acetylation is not carried out. This is observed for cystatins A, B, and S-type (S, SA and SN), and all the PRPs, suggesting the absence of NATs into the cisternae of cytoplasmic reticulum. In all other proteins, the action of MAPs is not man- datory and seems to depend on the type of the residue present at second position in the sequence. Therefore, Nt-acetylation may occur either on the iMet or on the first residue after iMet excision by MAPs. N-termini of proteins with small amino acid residues (Ser, Ala, Thr, Val, Gly, and Cys) at second position are mostly processed by MAPs, and the newly generated N-termini may be acetylated by NAT-A, as observed for the serine resi- due of thymosins β4 and β10, of S100A7, and of the two proteoforms of small proline-rich protein 3 [87,184,185,186]. N-termini of proteins with larger amino acid residues in the second position are not cleaved by MAPs, but potentially acetylated directly on the iMet by a variety of NATs depending on the N-terminal sequence. NAT-B potentially acetylates Met-N-termini when the second residue is Asp, Glu, Asn. NAT-C potentially acetylates Met-N-termini when the second residue is a hydrophobic amino acid (mainly Leu, Ile, Phe). Hydrophobic terminals are also recognized by NAT-E and NAT-F, suggesting that redundancy in activity exists between NATs [187]. Moreover, NAT-F, showed a broader specificity than NAT-C in acetylating Met-N-terminals when the second residue is Met, Lys and Gln, and the potential to acetylate the same N-terminals recognized by NAT-A in proteins in which iMet has not been cleaved [188]. Examples of human salivary proteins are represented by cystatin A, which has an isoleucine residue in position 2 and it is prob ably acetylated on Met1 by NAT-C (or NAT-F) and cystatin B, which has a second methi onine residue in position 2 and it is acetylated on Met1 probably by NAT-F. [57,87].
A peculiar example concerns the acetylation of S100A9, which among the five N terminal residues has two Met, one at position 1 and the other at position 5 (MTCKM…). Because of this N-terminal sequence, MAP catalyzes the cleavage not only at the level of the first Met residue, generating the long proteoform (1-113 residues), but also at the level of the fifth Met, giving rise to the short proteoform (1-109 residues), being the percentages of the two proteins similar in human saliva [25,87]. The Cys3 residue, present only in the long form, is responsible for the formation of the S-cysteinyl and S-glutathionyl deriva- tives of S100A9, above described in the paragraph 5. Since these four proteoforms of S100A9 can be also phosphorylated in the penultimate threonine residue of the sequence, the number of possible proteoforms of S100A9 detectable in human saliva are eight [188]. As we are aware, no information exists about specific function of these different pro teoforms of S100A9.
9.2. Pyroglutamic acid modification
As above reported, several human salivary proteins undergo N-terminal cyclization of glutamine and glutamic acid, as a protection for the N-terminal amino group. This PTM has been characterized for all the proteoforms of aPRPs (PRP-1 and PRP-3 types), [90,189] and bPRPs (II-2, IB-1), all the gPRPs [23] as well as of P-B peptide [41]. It is relevant to remark that although this PTM allows the integrity of the protein structure by protecting it from the action of aminopeptidases [190], on the other hand it significantly alters its hydrophobicity and solubility [183], facilitating protein aggregation. For this reason, many authors hypothesized that it plays a relevant role in several amyloidopathies [191]. Indeed, experiments carried out in a mouse model using glutaminyl cyclase inhibitors demonstrated a reduced level of pGlu-modified Aβ peptides that appeared to attenuate Alzheimer’s disease [192]. Nonetheless, the enzyme does not seem essential for the cyclization, which according to Bersin and coll. [193], is spontaneous and observed in ab sence of the enzyme too.
10. C-terminal modifications
Except for the removal of C-terminal residues by carboxy-peptidases, described in section 1.1.2, only three C-terminal modifications were detected during our top-down ex periments. One is the formation of a cyclic anhydride bond for the loss of one water mol ecule from two carboxylic groups in the presence of a C-terminal Glu residue of the frag ment 1-26 of β-globin, the second is the formation of one cyclic ester for the loss of a water molecule from the C-terminal serine of the fragment 1 of P-C peptide, the third is the formation of a cyclic N-substituted amide for the loss of one water molecule from the C- terminal glutamine of the fragment 1-41 of P-C peptide.
11. Conclusions
This review, which provides many answers regarding the complexity of the PTMs that human salivary proteins undergo, nevertheless prompts even more questions regarding the roles of many of them.
The first question concerns the possible functional role played by the products gen erated by the multiple proteolytic cleavages to which the components of almost all protein 1152 families are subjected.
It is relevant to underline that they could be evidenced only by a top-down strategy. Since evolution is addressed towards the selection of molecular events relevant to the functions of cells, organs and tissues, particularly puzzling is the functional meaning of the cleavages to which undergo acidic and basic PRPs, as well as challenging is the role of P-B peptide. Particularly challenging is defining the role of the myriad of recurrent fragments generated from bigger salivary proteins by exogenous proteinases. Therefore, the development of new strategies to study the biological role of the fragments is demanding.
The role of almost all the other PTMs described is obscure too. Proteomic searches, devoted to the discovery of new disease biomarkers, are observational studies, supported by powerful statistical tools, able to suggest potential biomarkers, but not able to suggest their specific physio-pathological role. Therefore, new experimental designs are needed to help researchers to connect the information obtained on structural modifications and the role of these changes in health and disease. This knowledge is fundamental for the biotechnological utilization of biologically active salivary proteins [194]. The recent pandemia evidenced the utility of saliva as human biofluid for painless non-invasive free at-home diagnostic purposes [1,195,196]. Indeed, saliva is to date also used for the fast check of illicit drugs abuse on the road [196]. We hope that this review could be a stimulus for further investigations and clinical appli cations.
Author Contributions
Conceptualization, MC, IM, GF, TC, AF, BM; methodology, All authors; software, AO, TC, CC, GG, CD, DR; validation, All authors; formal analysis, All; investigation; data curation, All authors; writing—original draft preparation, All the authors; writing—review and editing, All the authors. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Ethical review and approval were not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boroumand, M.; Olianas, A.; Cabras, T.; Manconi, B.; Fanni, D.; Faa, G.; Desiderio, C.; Messana, I.; Castagnola, M. Saliva, abodily fluid with recognized and potential diagnostic applications. J. Sep. Sci. 2021, 44, 3677–3690. [Google Scholar] [CrossRef] [PubMed]
- Helmerhorst, E.J.; Oppenheim, F.G. Saliva: a dynamic proteome. J. Dent. Res. 2007, 86, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.W.; Hardt, M.; Zhang, Y.H.; Freire, M.; Ruhl, S. The Human Salivary Proteome Wiki: A Community-Driven Research Platform. J. Dent. Res. 2021, 100, 1510–1519. [Google Scholar] [CrossRef] [PubMed]
- Iavarone, F.; Desiderio, C.; Vitali, A.; Messana, I.; Martelli, C.; Castagnola, M.; Cabras, T. Cryptides: latent peptides eve- rywhere. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 246–263. [Google Scholar] [CrossRef]
- Castle, D.; Castle, A. Intracellular transport and secretion of salivary proteins. Crit. Rev. Oral Biol. Med. 1998, 9, 4–22. [Google Scholar] [CrossRef]
- Messana, I.; Cabras, T.; Pisano, E.; Sanna, M. T.; Olianas, A.; Manconi, B.; Pellegrini, M.; Paludetti, G.; Scarano, E.; Fiorita, A.; et al. Trafficking and postsecretory events responsible for the formation of secreted human salivary peptides: a proteomics ap- proach. Mol. Cell. Proteomics 2008, 7, 911–926. [Google Scholar] [CrossRef]
- Chan, M.; Bennick, A. Proteolytic processing of a human salivary proline-rich protein precursor by proprotein convertases. Eur. J. Biochem. 2001, 268, 3423–3431. [Google Scholar] [CrossRef]
- Cai, K.; Bennick, A. Processing of acidic proline-rich proprotein by human salivary gland convertase. Arch. Oral Biol. 2004, 49, 871–879. [Google Scholar] [CrossRef]
- Inzitari, R.; Cabras, T.; Onnis, G.; Olmi, C.; Mastinu, A.; Sanna, M.T.; Pellegrini, M.G.; Castagnola, M.; Messana, I. Different isoforms and post-translational modifications of human salivary acidic proline-rich proteins. Proteomics 2005, 5, 805–815. [Google Scholar] [CrossRef]
- Kauffman, D. L.; Keller, P.J. The basic proline-rich proteins in human parotid saliva from a single subject. Arch. Oral Biol. 1979, 24, 249–256. [Google Scholar] [CrossRef]
- Kauffman, D.; Wong, R.; Bennick, A.; Keller, P. Basic proline-rich proteins from human parotid saliva: complete covalent struc- ture of protein IB-9 and partial structure of protein IB-6, members of a polymorphic pair. Biochemistry 1982, 21, 6558–6562. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, D.; Hofmann, T.; Bennick, A.; Keller, P. Basic proline-rich proteins from human parotid saliva: complete covalent structures of proteins IB-1 and IB-6. Biochemistry 1986, 25, 2387–2392. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, D. L.; Bennick, A.; Blum, M.; Keller, P. J. Basic proline-rich proteins from human parotid saliva: relationships of the covalent structures of ten proteins from a single individual. Biochemistry 1991, 30, 3351–3356. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, E.; Isemura, S.; Sanada, K. Complete amino acid sequence of a basic proline-rich peptide, P-D, from human parotid saliva. J. Biochem. 1983, 93, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, E.; Isemura, S.; Sanada, K. Complete amino acid sequence of a basic proline-rich peptide, P-F, from human parotid saliva. J. Biochem. 1983, 93, 883–888. [Google Scholar] [CrossRef]
- Saitoh, E.; Isemura, S.; Sanada, K. Further fractionation of basic proline-rich peptides from human parotid saliva and complete amino acid sequence of basic proline-rich peptide P-H. J. Biochem. 1999. [Google Scholar] [CrossRef]
- Isemura, S.; Saitoh, E.; Sanada, K. Isolation and amino acid sequences of proline-rich peptides of human whole saliva. J. Biochem. 1979, 86, 79–86. [Google Scholar]
- Isemura, S.; Saitoh, E.; Sanada, K. Fractionation and characterization of basic proline-rich peptides of human parotid saliva and the amino acid sequence of proline-rich peptide P-E. J. Biochem. 1982, 91, 2067–2075. [Google Scholar] [CrossRef]
- Messana, I.; Cabras, T.; Inzitari, R.; Lupi, A.; Zuppi, C.; Olmi, C.; Fadda, M. B.; Cordaro, M.; Giardina, B.; Castagnola, M. Char- acterization of the human salivary basic proline-rich protein complex by a proteomic approach. J. Proteome Res. 2004, 3, 792–800. [Google Scholar] [CrossRef]
- Cabras, T.; Pisano, E.; Boi, R.; Olianas, A.; Manconi, B.; Inzitari, R.; Fanali, C.; Giardina, B.; Castagnola, M.; Messana, I. Age- dependent modifications of the human salivary secretory protein complex. J. Proteome Res. 4134. [Google Scholar] [CrossRef]
- Maeda, N.; Kim, H.S.; Azen, E.A.; Smithies, O. Differential RNA splicing and post-translational cleavages in the human salivary proline-rich protein gene system. J. Biol. Chem. 1985, 260, 11123–11130. [Google Scholar] [CrossRef]
- Messana, I.; Inzitari, R.; Fanali, C.; Cabras, T.; Castagnola, M. Facts and artifacts in proteomics of body fluids. What proteomics of saliva is telling us? J. Sep. Sci. 2008, 31, 1948–1963. [Google Scholar] [CrossRef]
- Padiglia, A.; Orrù, R.; Boroumand, M.; Olianas, A.; Manconi, B.; Sanna, M.T.; Desiderio, C.; Iavarone, F.; Liori, B.; Messana, I.; et al. Extensive Characterization of the Human Salivary Basic Proline-Rich Protein Family by Top-Down Mass Spectrometry. J. Proteome Res. 2018, 17, 3292–3307. [Google Scholar] [CrossRef] [PubMed]
- Manconi, B.; Castagnola, M.; Cabras, T.; Olianas, A.; Vitali, A.; Desiderio, C.; Sanna, M.T.; Messana, I. The intriguing heteroge- neity of human salivary proline-rich proteins: Short title: Salivary proline-rich protein species. J. Proteomics 2016, 134, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Messana, I.; Cabras, T.; Iavarone, F.; Manconi, B.; Huang, L.; Martelli, C.; Olianas, A.; Sanna, M.T.; Pisano, E.; Sanna, M.; et al. Chrono-proteomics of human saliva: variations of the salivary proteome during human development. J. Proteome Res. 2015, 14, 1666–1677. [Google Scholar] [CrossRef] [PubMed]
- Manconi, B.; Cabras, T.; Pisano, E.; Sanna, M.T.; Olianas, A.; Fanos, V.; Faa, G.; Nemolato, S.; Iavarone, F.; Castagnola, M.; Messana, I. Modifications of the acidic soluble salivary proteome in human children from birth to the age of 48 months investi- gated by a top-down HPLC-ESI-MS platform. J. Proteomics 2013, 91, 536–543. [Google Scholar] [CrossRef]
- Lu, Y.; Bennick, A. Interaction of tannin with human salivary proline-rich proteins. Arch. Oral Biol. 1998, 43, 717–728. [Google Scholar] [CrossRef]
- Cannon, R.D.; Chaffin, W.L. Oral colonization by Candida albicans. Crit. Rev. Oral Biol Med. 1999, 359–383. [Google Scholar] [CrossRef]
- Ruhl, S.; Sandberg, A.L.; Cisar, J.O. Salivary receptors for the proline-rich protein-binding and lectin-like adhesins of oral acti- nomyces and streptococci. J. Dent. Res. 2004, 83, 505–510. [Google Scholar] [CrossRef]
- Cabras, T.; Melis, M.; Castagnola, M.; Padiglia, A.; Tepper, B.J.; Messana, I.; Tomassini Barbarossa, I. Responsiveness to 6-n- propylthiouracil (PROP) is associated with salivary levels of two specific basic proline-rich proteins in humans. PLoS One 2012, 7, e30962. [Google Scholar] [CrossRef]
- Vitorino, R.; Calheiros-Lobo, M.J.; Williams, J.; Ferrer-Correia, A.J.; Tomer, K.B.; Duarte, J.A.; Domingues, P.M.; Amado, F.M. (2007). Peptidomic analysis of human acquired enamel pellicle. Biomed. Chromatogr. 2007, 21, 1107–1117. [Google Scholar] [CrossRef]
- Palmerini, C.A.; Mazzoni, M.; Radicioni, G.; Marzano, V.; Granieri, L.; Iavarone, F.; Longhi, R.; Messana, I.; Cabras, T. , Sanna, M.T.; et al. Antagonistic Effect of a Salivary Proline-Rich Peptide on the Cytosolic Ca2+ Mobilization Induced by Progesterone in Oral Squamous Cancer Cells. PLoS One, 0147. [Google Scholar] [CrossRef]
- Macias, M. J.; Wiesner, S.; Sudol, M. WW and SH3 domains, two different scaffolds to recognize proline-rich ligands. FEBS Lett. 2002, 513, 30–37. [Google Scholar] [CrossRef]
- Righino, B.; Pirolli, D.; Radicioni, G.; Marzano, V.; Longhi, R.; Arcovito, A.; Sanna, M.T.; De Rosa, M.C.; Paoluzi, S.; Cesareni, G.; et al. Structural studies and SH3 domain binding properties of a human antiviral salivary proline-rich peptide. Biopolymers 2016, 106, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Azen, E.A. Genetics of salivary protein polymorphisms. Crit. Rev. Oral Biol. Med. [CrossRef]
- Sabatini, L.M.; Carlock, L.R.; Johnson, G.W.; Azen, E.A. cDNA cloning and chromosomal localization (4q11-13) of a gene for statherin, a regulator of calcium in saliva. Am. J. Hum. Genet. 1987, 41, 1048–1060. [Google Scholar] [PubMed]
- Raj, P.A.; Johnsson, M.; Levine, M.J.; Nancollas, G.H. Salivary statherin. Dependence on sequence, charge, hydrogen bonding potency, and helical conformation for adsorption to hydroxyapatite and inhibition of mineralization. J. Biol. Chem. 1992, 267, 5968–5976. [Google Scholar] [CrossRef] [PubMed]
- Niemi, L.D.; Johansson, I. Salivary statherin peptide-binding epitopes of commensal and potentially infectious Actinomyces spp. delineated by a hybrid peptide construct. Infect. Immun. 2004, 72, 782–787. [Google Scholar] [CrossRef]
- Gibbons, R.J.; Hay, D.I. Human salivary acidic proline-rich proteins and statherin promote the attachment of Actinomyces vis- cosus LY7 to apatitic surfaces. Infect. Immun. 1988, 56, 439–445. [Google Scholar] [CrossRef]
- Isemura, S. Nucleotide sequence of gene PBII encoding salivary proline-rich protein P-B. J. Biochem. 2000, 127, 393–398. [Google Scholar] [CrossRef]
- Inzitari, R.; Cabras, T.; Rossetti, D.V.; Fanali, C.; Vitali, A.; Pellegrini, M.; Paludetti, G.; Manni, A.; Giardina, B.; Messana, I.; et al. Detection in human saliva of different statherin and P-B fragments and derivatives. Proteomics 2006, 6, 6370–6379. [Google Scholar] [CrossRef]
- Strawich, E.; Glimcher, M.J. Tooth ‘enamelins’ identified mainly as serum proteins. Major ‘enamelin’ is albumin. Eur. J. Biochem. 1990, 191, 47–56. [Google Scholar] [CrossRef]
- Ayad, M.; Van Wuyckhuyse, B.C.; Minaguchi, K.; Raubertas, R.F.; Bedi, G.S.; Billings, R.J.; Bowen, W.H.; Tabak, L.A. The asso ciation of basic proline-rich peptides from human parotid gland secretions with caries experience. J. Dent. Res. 2000, 79, 976–982. [Google Scholar] [CrossRef]
- Jensen, J.L.; Lamkin, M.S.; Troxler, R.F.; Oppenheim, F.G. Multiple forms of statherin in human salivary secretions. Arch. Oral Biol. 1991, 36, 529–534. [Google Scholar] [CrossRef]
- Sabatini, L.M.; He, Y.Z.; Azen, E.A. Structure and sequence determination of the gene encoding human salivary statherin. Gene 1990, 89, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, R.; Lobo, M.J.; Duarte, J.R.; Ferrer-Correia, A.J.; Domingues, P.M.; Amado, F.M. The role of salivary peptides in dental caries. Biomed. Chromatogr. 2005, 19, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, L.M.; Azen, E.A. Histatins, a family of salivary histidine-rich proteins, are encoded by at least two loci (HIS1 and HIS2). Biochem. Biophys. Res. Commun. 1989, 160, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Oppenheim, F.G.; Xu, T.; McMillian, F.M.; Levitz, S.M.; Diamond, R.D.; Offner, G.D.; Troxler, R.F. Histatins, a novel family of histidine-rich proteins in human parotid secretion. Isolation, characterization, primary structure, and fungistatic effects on Can dida albicans. J. Biol. Chem. 1988, 263, 7472–7477. [Google Scholar] [CrossRef] [PubMed]
- Castagnola, M.; Inzitari, R.; Rossetti, D.V.; Olmi, C.; Cabras, T.; Piras, V.; Nicolussi, P.; Sanna, M.T.; Pellegrini, M.; Giardina, B.; et al. A cascade of 24 histatins (histatin 3 fragments) in human saliva. Suggestions for a pre-secretory sequential cleavage path way. J. Biol. Chem. 2004, 279, 41436–41443. [Google Scholar] [CrossRef]
- Torres, P.; Díaz, J.; Arce, M.; Silva, P.; Mendoza, P.; Lois, P.; Molina-Berríos, A.; Owen, G.I.; Palma, V.; Torres, V.A. The salivary peptide histatin-1 promotes endothelial cell adhesion, migration, and angiogenesis. FASEB J. 2017, 31, 4946–4958. [Google Scholar] [CrossRef]
- Oudhoff, M.J.; Bolscher, J.G.; Nazmi, K.; Kalay, H.; van ‘t Hof, W.; Amerongen, A.V.; Veerman, E.C. Histatins are the major 1317 wound-closure stimulating factors in human saliva as identified in a cell culture assay. FASEB J. 2008, 22, 3805–3812. [Google Scholar] [CrossRef]
- Amado, F.; Lobo, M.J.; Domingues, P.; Duarte, J.A.; Vitorino, R. Salivary peptidomics. Expert. Rev. Proteomics 2010, 7, 709–721. [Google Scholar] [CrossRef]
- Castagnola, M.; Scarano, E.; Passali, G.C.; Messana, I.; Cabras, T.; Iavarone, F.; Di Cintio, G.; Fiorita, A.; De Corso, E.; Paludetti, G. Salivary biomarkers and proteomics: future diagnostic and clinical utilities. Acta Otorhinolaryngol. Ital. 2017, 37, 94–101. [Google Scholar] [CrossRef]
- Kos, J.; Lah, T.T. Cysteine proteases and their endogenous inhibitors: target proteins for prognosis, diagnosis and therapy in cancer (review). Oncol. Rep. 1998, 5, 1349–13461. [Google Scholar] [CrossRef]
- Henskens, Y.M.; Veerman, E.C.; Nieuw Amerongen, A.V. Cystatins in health and disease. Biol. Chem. Hoppe Seyler 1996, 377, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Calkins, C.C.; Sloane, B.F. Mammalian cysteine protease inhibitors: biochemical properties and possible roles in tumor progres sion. Biol. Chem. Hoppe Seyler 1995, 376, 71–80. [Google Scholar] [PubMed]
- Manconi, B.; Liori, B.; Cabras, T.; Vincenzoni, F.; Iavarone, F.; Castagnola, M.; Messana, I.; Olianas, A. Salivary cystatins: explor ing new post-translational modifications and polymorphisms by top-down mass spectrometry. J. Proteome Res. 2017, 16, 4196–4207. [Google Scholar] [CrossRef]
- Lupi, A.; Messana, I.; Denotti, G.; Schininà, M.E.; Gambarini, G.; Fadda, M.B.; Vitali, A.; Cabras, T.; Piras, V.; Patamia, M.; et al. 1334 Identification of the human salivary cystatin complex by the coupling of high-performance liquid chromatography and ion trap mass spectrometry. Proteomics 2003, 3, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.M.; Souda, P.; Halgand, F.; Wong, D.T.; Loo, J.A.; Faull, K.F.; Whitelegge, J.P. Confident assignment of intact mass tags to human salivary cystatins using top-down Fourier-transform ion cyclotron resonance mass spectrometry. J. Am. Soc. Mass Spectrom. 2010, 21, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Iavarone, F.; Cabras, T.; Pisano, E.; Sanna, M.T.; Nemolato, S.; Vento, G.; Tirone, C.; Romagnoli, C.; Cordaro, M.; Fanos, V.; et al. Top-down HPLC-ESI-MS detection of S-glutathionylated and S-cysteinylated derivatives of cystatin B and its 1-53 and 54 98 fragments in whole saliva of human preterm newborns. J. Proteome Res. 2013, 12, 917–926. [Google Scholar] [CrossRef]
- Requena, T.; Velasco, M. The human microbiome in sickness and in health. Rev. Clin. Esp. (Barc). 2021, 221, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Chimenos-Küstner, E.; Giovannoni, M.L.; Schemel-Suárez, M. Dysbiosis as a determinant factor of systemic and oral pathology: importance of microbiome. Med. Clin. (Barc). 2017, 149, 305–309. [Google Scholar] [CrossRef]
- Helmerhorst, E.J.; Sun, X.; Salih, E.; Oppenheim, F.G. Identification of Lys-Pro-Gln as a novel cleavage site specificity of saliva associated proteases. J. Biol. Chem. 2008, 283, 19957–19966. [Google Scholar] [CrossRef]
- Zamakhchari, M.; Wei, G.; Dewhirst, F.; Lee, J.; Schuppan, D.; Oppenheim, F.G.; Helmerhorst, E.J. Identification of Rothia bac teria as gluten-degrading natural colonizers of the upper gastro-intestinal tract. PloS One 2011, 6, e24455. [Google Scholar] [CrossRef]
- Asano, M.; Komiyama, K. Polymeric immunoglobulin receptor. J. Oral Sci. 2011, 53, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Kaetzel, C.S. The polymeric immunoglobulin receptor: bridging innate and adaptive immune responses at mucosal sur faces. Immunol. Rev. 2005, 206, 83–99. [Google Scholar] [CrossRef]
- Ramachandran, P.; Boontheung, P.; Xie, Y.; Sondej, M.; Wong, D.T.; Loo, J.A. Identification of N-linked glycoproteins in human saliva by glycoprotein capture and mass spectrometry. J. Proteome Res. 2006, 5, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Cabras, T.; Sanna, M.; Manconi, B.; Fanni, D.; Demelia, L.; Sorbello, O.; Iavarone, F.; Castagnola, M.; Faa, G.; Messana, I. Proteo mic investigation of whole saliva in Wilson’s disease. J. Proteomics 2015, 128, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Boroumand, M.; Iavarone, F.; Manconi, B.; Pieroni, L.; Greco, V.; Vento, G.; Tirone, C.; Desiderio, C.; Fiorita, A.; Faa, G.; et al. HPLC-ESI-MS top-down analysis of salivary peptides of preterm newborns evidenced high activity of some exopeptidases and convertases during late fetal development. Talanta 2021, 222, 121429. [Google Scholar] [CrossRef] [PubMed]
- Cabras, T.; Pisano, E.; Mastinu, A.; Denotti, G.; Pusceddu, P.P.; Inzitari, R.; Fanali, C.; Nemolato, S.; Castagnola, M.; Messana, I. 1362 Alterations of the salivary secretory peptidome profile in children affected by type 1 diabetes. Mol. Cell. Proteomics 2010, 9, 2099–2108. [Google Scholar] [CrossRef]
- Hardt, M.; Thomas, L.R.; Dixon, S.E.; Newport, G.; Agabian, N.; Prakobphol, A.; Hall, S.C.; Witkowska, H.E.; Fisher, S.J. Toward defining the human parotid gland salivary proteome and peptidome: identification and characterization using 2D SDS-PAGE, ultrafiltration, HPLC, and mass spectrometry. Biochemistry 2005, 44, 2885–2899. [Google Scholar] [CrossRef]
- Huq, N.L.; Cross, K.J.; Ung, M.; Myroforidis, H.; Veith, P.D.; Chen, D.; Stanton, D.; Huiling, H.; Ward, B.R.; Reynolds, E.C. A Review of the Salivary Proteome and Peptidome and Saliva-derived Peptide Therapeutics. Int. J. Pept. Res. Ther. [CrossRef]
- Manconi, B.; Liori, B.; Cabras, T.; Vincenzoni, F.; Iavarone, F.; Lorefice, L.; Cocco, E.; Castagnola, M.; Messana, I.; Olianas, A. Top-down proteomic profiling of human saliva in multiple sclerosis patients. J. Proteomics. [CrossRef]
- Serrao, S.; Firinu, D.; Olianas, A.; Deidda, M.; Contini, C.; Iavarone, F.; Sanna, M.T.; Boroumand, M.; Amado, F.; Castagnola, M. 1374 Top-Down Proteomics of Human Saliva Discloses Significant Variations of the Protein Profile in Patients with Mastocytosis. J. 1375 Proteome. Res. 2020, 19, 3238–3253. [Google Scholar] [CrossRef]
- Contini, C. , Serrao, S., Manconi, B., Olianas, A., Iavarone, F., Bizzarro, A., Masullo, C., Castagnola, M., Messana, I., Diaz, G., et al. Salivary Proteomics Reveals Significant Changes in Relation to Alzheimer’s Disease and Aging. J. Alzheimers Dis. 2022, 89, 605–622. [Google Scholar] [CrossRef]
- Olianas, A.; Guadalupi, G.; Cabras, T.; Contini, C.; Serrao, S.; Iavarone, F.; Castagnola, M.; Messana, I.; Onali, S.; Chessa, L.; et al. Top-Down Proteomics Detection of Potential Salivary Biomarkers for Autoimmune Liver Diseases Classification. Int. J. Mol. Sci. 2023, 24, 959. [Google Scholar] [CrossRef]
- Contini, C.; Olianas, A.; Serrao, S.; Deriu, C.; Iavarone, F.; Boroumand, M.; Bizzarro, A.; Lauria, A.; Faa, G.; Castagnola, M.; et al. Top-Down Proteomics of Human Saliva Highlights Anti-inflammatory, Antioxidant, and Antimicrobial Defense Responses in Alzheimer Disease. Front. Neurosci. 2021, 15, 668852. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.J.; Linley, A.; Hammond, D.E.; Hood, F.E.; Coulson, J.M.; MacEwan, D.J.; Ross, S.J.; Slupsky, J.R.; Smith, P.D.; Eyers, P.A. New Perspectives, Opportunities, and Challenges in Exploring the Human Protein Kinome. Cancer Res. 2018, 78, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Salih, E.; Siqueira, W.L.; Helmerhorst, E.J.; Oppenheim, F.G. Large-scale phosphoproteome of human whole saliva using disul fidethiol interchange covalent chromatography and mass spectrometry. Anal. Biochem. [CrossRef]
- Stone, M.D. .; Chen, X.; McGowan, T.; Bandhakavi, S.; Cheng, B.; Rhodus, N.L.; Griffin, T.J. Large-scale phosphoproteomics analysis of whole saliva reveals a distinct phosphorylation pattern. J. Proteome Res. Tagliabracci, V.S.; Pinna L.A.; Dixon, J.E. Secreted protein kinases. Trends Biochem. Sci. , , -130, doi:10.1016/j.tibs.2012.11.008. 2011, 10, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Tagliabracci, V.S.; Wiley, S.E.; Guo, X.; Kinch, L.N.; Durrant, E.; Wen, J.; Xiao, J.; Cui, J.; Nguyen, K.B.; Engel, J.L.; et al. A Single Kinase Generates the Majority of the Secreted Phosphoproteome. Cell 2015, 161, 1619–1632. [Google Scholar] [CrossRef]
- Isemura, S.; Saitoh, E.; Sanada, K.; Minakata, K. Identification of full-sized forms of salivary (S-type) cystatins (cystatin SN, cystatin SA, cystatin S, and two phosphorylated forms of cystatin S) in human whole saliva and determination of phosphory lation sites of cystatin S. J Biochem. 1991, 110, 648–654. [Google Scholar] [CrossRef]
- Madapallimattam, G.; Bennick, A. Phosphorylation of salivary proteins by salivary gland protein kinase. J. Dent. Res. 1986, 65, 405–411. [Google Scholar] [CrossRef]
- Iavarone, F.; D’Alessandro, A.; Tian, N.; Cabras, T.; Messana, I.; Helmerhorst, E.J.; Oppenheim, F.G.; Castagnola, M. Castagnola, M. High resolution high-performance liquid chromatography with electrospray ionization mass spectrometry and tandem mass spec trometry characterization of a new isoform of human salivary acidic proline-rich proteins named Roma-Boston Ser22 (Phos). 1405. [Google Scholar]
- Phe variant. J Sep Sci. 2014, 37, 1896–1902. [CrossRef]
- Halgand, F.; Zabrouskov, V.; Bassilian, S.; Souda, P.; Loo, J.A.; Faull, K.F.; Wong, D.T.; &, *!!! REPLACE !!!*; Whitelegge, J.P. ; & Whitelegge, J.P. Defining intact protein primary structures from saliva: a step toward the human proteome project. Anal. Chem. [CrossRef]
- Castagnola, M.; Inzitari, R.; Fanali, C.; Iavarone, F.; Vitali, A.; Desiderio, C.; Vento, G.; Tirone. C.; Romagnoli, C.; Cabras T.; et al. The surprising composition of the salivary proteome of preterm human newborn. Mol. Cell Proteomics 2011, 10, M110–003467. [Google Scholar] [CrossRef]
- Wahid, A.; Sohail, A.; Wang, H.; Guo, M.; Zhang, L.; Ji, Y.; Wang, P.; Xiao, H. Titanium(IV) immobilized affinity chromatog raphy facilitated phosphoproteomics analysis of salivary extracellular vesicles for lung cancer. Anal. Bioanal. Chem. 2022, 414, 3697–3708. [Google Scholar] [CrossRef]
- Sun, J.; Wang, X.; Ding, Y.; Xiao, B.; Wang, X.; Ali, M.M.; Ma, L.; Xie, Z.; Gu, Z.; Chen, G.; et al. Proteomic and phosphoproteomic landscape of salivary extracellular vesicles to assess OSCC therapeutical outcomes. Proteomics 2023, 23, e2200319. [Google Scholar] [CrossRef]
- Inzitari, R.; Vento, G.; Capoluongo, E.; Boccacci, S.; Fanali, C.; Cabras, T.; Romagnoli, C.; et al. Proteomic analysis of salivary acidic proline-rich proteins in human pre-term and at-term new-borns. J. Proteome Res. 2007, 6, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Messana, I.; Cabras, T.; Iavarone, F.; Manconi, B.; Huang, L.; Martelli, C.; Olianas, A.; Sanna, M.T.; Pisano, E.; Sanna, M.; et al. Chrono-proteomics of human saliva: variations of the salivary proteome during human development. J. Proteome Res. 2015, 14, 1666–1677. [Google Scholar] [CrossRef] [PubMed]
- Castagnola, M.; Messana, I.; Inzitari, R.; Fanali, C.; Cabras, T.; Morelli, A.; Pecoraro, A.M.; Neri, G.; Torrioli, M.G. , Gurrieri, F. Hypo-phosphorylation of salivary peptidome as a clue to the molecular pathogenesis of autism spectrum disorders. J. Proteome Res. 2008, 7, 5327–5332. [Google Scholar] [CrossRef]
- Gamage, N.; Barnett, A.; Hempel, N.; Duggleby, R.G.; Windmill, K.F.; Martin, J.L.; McManus, M.E. Human sulfotransferases and their role in chemical metabolism. Toxicol. Sci. 2006, 90, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Leyh, T.S. The physical biochemistry and molecular genetics of sulfate activation. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 515–542. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.V. Human 3’-phosphoadenosine 5’-phosphosulfate (PAPS) synthase: biochemistry, molecular biology and genetic deficiency. IUBMB Life 2003, 55, 1–11. [Google Scholar] [CrossRef]
- Schröder, E.; Gebel, L.; Eremeev, A.A.; Morgner, J.; Grum, D.; Knauer, S.K.; Bayer, P.; Mueller, J.W. Human PAPS synthase 1433 isoforms are dynamically regulated enzymes with access to nucleus and cytoplasm PLoS One. 7, 2955; e9. [Google Scholar] [CrossRef]
- Gamage, N.; Barnett, A.; Hempel, N.; Duggleby, R.G.; Windmill, K.F.; Martin, J.L.; McManus, M.E. Human sulfotransferases and their role in chemical metabolism. Toxicol Sci. 2006, 90, 5–22. [Google Scholar] [CrossRef]
- Negishi, M.; Pedersen, L.G.; Petrotchenko, E.; Shevtsov, S.; Gorokhov, A.; Kakuta, Y.; Pedersen, L.C. Structure and function of sulfotransferases. Arch. Biochem. Biophys. 2001, 390, 149–157. [Google Scholar] [CrossRef]
- Falany, C.N. Enzymology of human cytosolic sulfotransferases. FASEB J. 1997, 11, 206–216. [Google Scholar] [CrossRef]
- Pedersen, L.C.; Yi, M.; Pedersen, L.G.; Kaminski, A.M. From steroid and drug metabolism to glycobiology, using sulfotransfer ase structures to understand and tailor function. Drug Metab Dispos. 2022, 50, 1027–1041. [Google Scholar] [CrossRef]
- Kurogi, K.; Rasool, M.I.; Alherz, F.A.; El Daibani, A.A.; Bairam, A.F.; Abunnaja, M.S.; Yasuda, S.; Wilson, L.J.; Hui, Y.; Liu, M.C. SULT genetic polymorphisms: physiological, pharmacological and clinical implications. Expert Opin. Drug Metab. Toxicol. 2021, 17, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Riches, Z.; Stanley, E.L.; Bloomer, J.C.; Coughtrie, M.W. Quantitative evaluation of the expression and activity of five major sulfotransferases (SULTs) in human tissues: the SULT “pie”. Drug Metab. Dispos. [CrossRef]
- Cabras, T.; Fanali, C.; Monteiro, J.A.; Amado, F.; Inzitari, R.; Desiderio, C.; Giardina, B.; Castagnola, M.; Messana, I. Tyrosine polysulfation of human salivary histatin 1. A post-translational modification specific of the submandibular gland. J. Proteome Res. 2007, 6, 2472–2480. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B. The basics of thiols and cysteines in redox biology and chemistry. Free Radic. Biol. Med. 2015, 80, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B. Thiol chemistry and specificity in redox signaling. Free Radic. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef]
- Cabras, T.; Manconi, B.; Iavarone, F.; Fanali, C.; Nemolato, S.; Fiorita, A.; Scarano, E.; Passali, G.C.; Manni, A.; Cordaro, M.; et al. RP-HPLC-ESI-MS evidenced that salivary cystatin B is detectable in adult human saliva mostly as S-modified derivatives: S glutathionyl, S-cysteinyl and S-S 2-mer. J. Proteomics 2012, 75, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Pol, E.; Björk, I. Contributions of individual residues in the N-terminal region of cystatin B (stefin B) to inhibition of cysteine proteinases. Biochim. Biophys. Acta 2003, 1645, 105–112. [Google Scholar] [CrossRef]
- Grek, C.L.; Zhang, J.; Manevich, Y.; Townsend, D.M.; Tew, K.D. Causes and consequences of cysteine S-glutathionylation. J. Biol. Chem. 2013, 288, 26497–26504. [Google Scholar] [CrossRef]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef]
- Leclerc, E.; Fritz, G.; Vetter, S.W.; Heizmann, C.W. Binding of S100 proteins to RAGE: an update. Biochim. Biophys. Acta 2009, 1793, 993–1007. [Google Scholar] [CrossRef]
- Zackular, J.P.; Chazin, W.J.; Skaar, E.P. Nutritional Immunity: S100 Proteins at the Host-Pathogen Interface. J. Biol. Chem. 2015, 290, 18991–18998. [Google Scholar] [CrossRef]
- Lim, S.Y.; Raftery, M.J.; Goyette, J.; Hsu, K.; Geczy, C.L. Oxidative modifications of S100 proteins: functional regulation by redox. J. Leukoc. Biol. 2009, 86, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Raftery, M.J.; Goyette, J.; Geczy, C.L. S-glutathionylation regulates inflammatory activities of S100A9. J. Biol. Chem. 2010, 285, 14377–14388. [Google Scholar] [CrossRef] [PubMed]
- Boroumand, M.; Manconi, B.; Serrao, S.; Iavarone, F.; Olianas, A.; Cabras, T.; Contini, C.; Pieroni, L.; Sanna, M.T.; Vento, G.; et al. Investigation by top-down high-performance liquid chromatography–mass spectrometry of glutathionylation and cysteinyl ation of salivary S100A9 and cystatin B in preterm newborns. Sep. Sci. plus 2022, 5, 17–27. [Google Scholar] [CrossRef]
- Iavarone, F.; Melis, M.; Platania, G.; Cabras, T.; Manconi, B.; Petruzzelli, R.; Cordaro, M.; Siracusano, A.; Faa, G.; Messana, I.; et al. Characterization of salivary proteins of schizophrenic and bipolar disorder patients by top-down proteomics. J. Proteomics 2014, 103, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Contini, C.; Firinu, D.; Serrao, S.; Manconi, B.; Olianas, A.; Cinetto, F.; Cossu, F.; Castagnola, M.; Messana, I.; Del Giacco, S.; et al. RP-HPLC-ESI-IT Mass Spectrometry Reveals Significant Variations of the Human Salivary Protein Profile Associated with Predominantly Antibody Deficiencies. J. Clin. Immunol. 2020, 40, 329–339. [Google Scholar] [CrossRef]
- Hoskin, T.S.; Crowther, J.M.; Cheung, J.; Epton, M.J.; Sly, P.D.; Elder, P.A.; Dobson, R.C.J.; Kettle, A.J.; Dickerhof, N. Oxidative cross-linking of calprotectin occurs in vivo, altering its structure and susceptibility to proteolysis. Redox Biol. 2019, 24, 101202. [Google Scholar] [CrossRef]
- Gomes, L.H.; Raftery, M.J.; Yan, W.X.; Goyette, J.D.; Thomas, P.S.; Geczy, C.L. S100A8 and S100A9-oxidant scavengers in in flammation. Free Radic. Biol. Med. 2013, 58, 170–186. [Google Scholar] [CrossRef]
- Smith, T.S.; Bennett, J.P.Jr. Mitochondrial toxins in models of neurodegenerative diseases. I: In vivo brain hydroxyl radical production during systemic MPTP treatment or following microdialysis infusion of methylpyridinium or azide ions. Brain Res. 1997, 765, 183–188. [Google Scholar] [CrossRef]
- Karlík, M.; Valkovič, P.; Hančinová, V.; Krížová, L.; Tóthová, L.; Celec, P. Markers of oxidative stress in plasma and saliva in patients with multiple sclerosis. Clin. Biochem. 2015, 48, 24–28. [Google Scholar] [CrossRef]
- Schipper, H.M.; Arnold, D.; GrandʼMaison, F.; Melmed, C.; Moore, F.; Levental, M. Constantin Su, M., Stril, J.L. Godin J., Tol erability and safety of combined glatiramer acetate and N-acetylcysteine in relapsing-remitting multiple sclerosis. Clin. Neuro pharmacol, 38. [CrossRef]
- Yao, Y.; Berg, E.A.; Costello, C.E.; Troxler, R.F.; Oppenheim, F.G. Identification of protein components in human acquired enamel pellicle and whole saliva using novel proteomics approaches. J. Biol. Chem. [CrossRef]
- Schüpbach, P.; Oppenheim, F.G.; Lendenmann, U.; Lamkin, M.S.; Yao, Y.; Guggenheim, B. Electron-microscopic demonstration of proline-rich proteins, statherin, and histatins in acquired enamel pellicles in vitro. Eur J Oral Sci. 2001, 109, 60–68. [Google Scholar] [CrossRef]
- Hannig, C.; Hannig, M.; Kensche, A.; Carpenter, G. The mucosal pellicle - An underestimated factor in oral physiology. Arch. Oral Biol. 2017, 80, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Presland, R.B.; Dale, B.A. Epithelial structural proteins of the skin and oral cavity: function in health and disease. Crit. Rev. Oral Biol. Med. 2000, 11, 383–408. [Google Scholar] [CrossRef] [PubMed]
- Bradway, S. D.; Bergey, E.J.; Jones, P.C.; Levine, M.J. Oral mucosal pellicle. Adsorption and transpeptidation of salivary com ponents to buccal epithelial cells. Biochem. J. 1989, 261, 887–896. [Google Scholar] [CrossRef]
- Cabiddu, G.; Maes, P.; Hyvrier, F.; Olianas, A.; Manconi, B.; Brignot, H.; Canon, F.; Cabras, T.; Morzel, M. Proteomic character ization of the mucosal pellicle formed in vitro on a cellular model of oral epithelium. J. Proteomics 2020, 222, 103797. [Google Scholar] [CrossRef]
- Neyraud, E.; Morzel, M. Biological films adhering to the oral soft tissues: Structure, composition, and potential impact on taste perception. J. Texture Stud. 2019, 50, 19–26. [Google Scholar] [CrossRef]
- Perez Alea, M.; Thomas, V.; Martin, G.; El Alaoui, S. Identification of human salivary transglutaminases. Amino Acids 2013, 44, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Griffin, M. TG2, a novel extracellular protein with multiple functions. Amino Acids 2012, 42, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Boroumand, M.; Olianas, A.; Manconi, B.; Serrao, S.; Iavarone, F.; Desiderio, C.; Pieroni, L.; Faa, G.; Messana, I.; Castagnola, M.; et al. Mapping of transglutaminase 2 sites of human salivary small basic proline-rich proteins by HPLC high resolution ESI MS/MS. J. Proteome Res. 2020, 19, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Cabras, T.; Inzitari, R.; Fanali, C.; Scarano, E.; Patamia, M. Sanna, M.T.; Pisano, E.; Giardina, B.; Castagnola, M.; Messana, I. HPLC-MS characterization of cyclo-statherin Q-37, a specific cyclization product of human salivary statherin generated by transglutaminase 2. J. Sep. Sci. 2006, 29, 2600–2608. [Google Scholar] [CrossRef]
- Hannig, C.; Hannig, M.; Attin, T. Enzymes in the acquired enamel pellicle. Eur. J. Oral Sci. 2005, 113, 2–13. [Google Scholar] [CrossRef]
- Siqueira, W.L.; Custodio, W.; McDonald, E.E. New insights into the composition and functions of the acquired enamel pellicle. J. Dent. Res. 2012, 91, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ames, J.M.; Smith, R.D.; Baynes, J.W.; Metz, T.O. A Perspective on the Maillard Reaction and the Analysis of protein Glycation by Mass Spectrometry: Probing the Pathogenesis of Chronic Disease. J. Proteome Res. 2009, 8, 754–769. [Google Scholar] [CrossRef] [PubMed]
- Rahbar, S.; Blumenfe, O.; Ranney, H.M. Studies of an unusual hemoglobin in patients with diabetes mellitus. Biochem. Biophys. Re.s Commun. 1969, 36, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, I.; Morimoto, K.; Takeshita, T.; Toda, M. Correlation between Saliva Glycated and Blood Glycated Proteins. Environ. Health Prev. Med. 2003, 8, 95–99. [Google Scholar] [CrossRef]
- Goldstein, D.E.; Little, R.R.; Lorenz, R.A.; Malone, J.I.; Nathan, D.; Peterson, C.M. Tests of glycemia in diabetes. Diabetes Care 1995, 18, 896–909. [Google Scholar] [CrossRef]
- Ramasamy. R.; Vannucci, S.J.; Yan, S.S.; Herold, K.; Yan, S.F.; Schmidt, A.M. Advanced glycation end products and RAGE: a common thread in aging, diabetes, neurodegeneration, and inflammation. Glycobiology 2005, 15, 16R–28R. [Google Scholar] [CrossRef]
- Corica, D.; Pepe, G.; Currò, M.; Aversa, T.; Tropeano, A.; Ientile, R.; Wasniewska, M. Methods to investigate advanced glycation end-product and their application in clinical practice. Methods. 2022, 203, 90–102. [Google Scholar] [CrossRef]
- McCarthy, A.D.; Cortizo, A.M.; Giménez Segura, G.; Bruzzone, L.; Etcheverry, S.B. Non-enzymatic glycosylation of alkaline phosphatase alters its biological properties. Mol. Cell. Biochem. 1998, 181, 63–69. [Google Scholar] [CrossRef]
- Muraoka, M.Y.; Justino, A.B; Caixeta, D.C.; Queiroz, J.S.; Sabino-Silva, R.; Espindola, F.S. Fructose and methylglyoxal-induced glycation alters structural and functional properties of salivary proteins, albumin and lysozyme. PLoS One 2022, 17, e0262369. [Google Scholar] [CrossRef]
- Tessier, F.J. The Maillard reaction in the human body. The main discoveries and factors that affect glycation. Pathol. Biol. (Paris) 2010, 58, 214–219. [Google Scholar] [CrossRef]
- Shin, A.; Connolly, S.; Kabytaev, K. Protein glycation in diabetes mellitus. Adv. Clin. Chem. 2023, 113, 101–156. [Google Scholar] [CrossRef] [PubMed]
- Li. Y.M. Glycation ligand binding motif in lactoferrin. Implication in diabetic infection. Adv. Exp. Med. Biol. 1998, 443, 57–63. [Google Scholar] [CrossRef]
- Belce, A.; Uslu, E.; Kucur, M.; Umut, M.; Ipbuker, A.; Seymen, H.O. Evaluation of salivary sialic acid level and Cu-Zn superoxide dismutase activity in type 1 diabetes mellitus. Tohoku J. Exp. Med. 2000, 192, 219–225. [Google Scholar] [CrossRef]
- Ilea, A.; Băbţan, A.M.; Boşca, B.A.; Crişan, M.; Petrescu, N.B.; Collino, M.; Sainz, R.M.; Gerlach, J.Q.; Câmpian, R.S. Advanced Glycation end products (AGEs) in oral pathology. Arch. Oral Biol. 2018, 93, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Manig, F.; Hellwig, M.; Pietz, F.; Henle, T. Quantitation of free glycation compounds in saliva. PLoS One 2019, 14, e0220208. [Google Scholar] [CrossRef]
- Landesberg, R.; Woo, V.; Huang, L.; Cozin, M.; Lu, Y.; Bailey, C.; Qu, W.; Pulse, C.; Schmidt, A.M. The expression of the receptor for glycation endproducts (RAGE) in oral squamous cell carcinomas. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2008, 105, 617–624. [Google Scholar] [CrossRef]
- Maciejczyk, M.; Nesterowicz, M.; Szulimowska, J.; Zalewska, A. Oxidation, Glycation, and Carbamylation of Salivary Biomol ecules in Healthy Children, Adults, and Elderly: Can Saliva Be Used in the assessment of Aging? J. Inflam. Res. 2022, 15, 2051–2073. [Google Scholar] [CrossRef]
- Goyette, J.; Geczy, C.L. Inflammation-associated S100 proteins: new mechanisms that regulate function. Amino Acids 2011, 41, 821–842. [Google Scholar] [CrossRef]
- Agho, E.T.; Owotade, F.J.; Kolawole, B.A.; Oyetola, E.O.; Adedeji, T.A. ; Salivary inflammatory biomarkers and glycated hae moglobin among patients with type 2 diabetic mellitus. BMC Oral Health 2021, 21, 101. [Google Scholar] [CrossRef]
- Helenius, A.; Aebi, M. Intracellular functions of N-linked glycans. Science 2001, 291, 2364–2369. [Google Scholar] [CrossRef]
- Xu, C.; Ng, D.T.W. Glycosylation-directed quality control of protein folding. Nat. Rev. Mol. Cell. Biol. 2015, 16, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Minakata, S.; Manabe, S.; Inai, Y.; Ikezaki, M.; Nishitsuji, K.; Ito, Y.; Ihara, Y. Protein C-Mannosylation and C-Mannosyl Tryp- tophan in Chemical Biology and Medicine. Molecules 2021, 26, 5258. [Google Scholar] [CrossRef] [PubMed]
- Franc, V.; Yang, Y.; Heck, A.J. Proteoform profile mapping of the human serum complement component C9 revealing unex- pected new features of N-, O-, and C-glycosylation. Anal. Chem. 2017, 89, 3483–3491. [Google Scholar] [CrossRef] [PubMed]
- Siew, J.J.; Chern, Y. Microglial Lectins in Health and Neurological Diseases. Front. Mol. Neurosci. 2018, 11, 158. [Google Scholar] [CrossRef] [PubMed]
- Duca, M.; Malagolini, N.; Dall’Olio, F. The Mutual Relationship between Glycosylation and Non-Coding RNAs in Cancer and Other Physio-Pathological Conditions. Int. J. Mol. Sci. 2022, 23, 15804. [Google Scholar] [CrossRef] [PubMed]
- Borga, C.; Meeran, S.M.; Fassan, M. Non-coding RNAs, a real Next-Gen Class of Biomarkers? Noncoding RNA Res. 2019, 4, 80–81. [Google Scholar] [CrossRef]
- Flynn, R.A.; Pedram, K.; Malaker, S.A.; Batista, P.J.; Smith, B.A.H.; Johnson, A.G.; George, B.M. ; Majzoub. K.; Villalta, P.W.; Carette, J.E.; et al. Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell, 3109. [Google Scholar] [CrossRef]
- Morelle, W.; Canis, K.; Chirat, F.; Faid, V.; Michalski, J.C. The use of mass spectrometry for the proteomic analysis of glycosyl- ation. Proteomics 2006, 6, 3993–4015. [Google Scholar] [CrossRef]
- Sondej, M.; Denny, P.A.; Xie, Y.; Ramachandran, P.; Si, Y.; Takashima, J.; Shi, W.; Wong, D.T.; Loo, J.A.; Denny, P.C. Glycop ro- filing of the Human Salivary Proteome. Clin. Proteomics. 2009, 5, 52–68. [Google Scholar] [CrossRef]
- Cabras, T.; Boi, R.; Pisano, E.; Iavarone, F.; Fanali, C.; Nemolato, S.; Faa, G.; Castagnola, M.; Messana, I. HPLC-ESI-MS and MS/MS structural characterization of multifucosylated N-glycoforms of the basic proline-rich protein IB-8a CON1(+) in human saliva. J. Sep. Sci. 2012, 35, 1079–1086. [Google Scholar] [CrossRef]
- Manconi, B.; Cabras, T.; Sanna, M.; Piras, V.; Liori, B.; Pisano, E.; Iavarone, F.; Vincenzoni, F.; Cordaro, M.; Faa, G.; et al. N- and O-linked glycosylation site profiling of the human basic salivary proline-rich protein 3M. J. Sep. Sci. 2016, 39, 1987–1997. [Google Scholar] [CrossRef]
- Liu, S.; Wang, H.; Jiang, X.; Ji, Y.; Wang, Z.; Zhang, Y.; Wang, P.; Xiao, H. Integrated N-glycoproteomics Analysis of Human Saliva for Lung Cancer. J. Proteome Res. 2022, 21, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Kobeissy, F.; Mondello, S.; Barsa, C.; Mechref, Y. MS-based glycomics: An analytical tool to assess nervous system diseases. Front. Neurosci. 2022, 16, 1000179. [Google Scholar] [CrossRef] [PubMed]
- Robertson, L.A.; Moya, K.L.; Breen, K.C. The potential role of tau protein O-glycosylation in Alzheimer’s disease. J. Alzheimers Dis. 2004, 6, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: mechanisms and clinical implications. Nat. Rev. Cancer. 2015, 15, 540–555. [Google Scholar] [CrossRef]
- György, B.; Tóth, E.; Tarcsa, E.; Falus, A.; Buzás, E.I. Citrullination: a posttranslational modification in health and disease. Int. J. Biochem. Cell. Biol. 2006, 38, 1662–1677. [Google Scholar] [CrossRef]
- Anzilotti, C.; Pratesi, F.; Tommasi, C.; Migliorini, P. Peptidylarginine deiminase 4 and citrullination in health and disease. Au- toimmun. Rev. 9,. [CrossRef]
- Alghamdi, M.; Alasmari, D.; Assiri, A.; Mattar, E.; Aljaddawi, A.A.; Alattas, S.G.; Redwan, E.M. An Overview of the Intrinsic Role of Citrullination in Autoimmune Disorders. J. Immunol. Res. 2019, 2019, 7592851. [Google Scholar] [CrossRef]
- van Venrooij, W.J.; Pruijn, G.J. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2000, 2, 249–251. [Google Scholar] [CrossRef]
- Ciesielski, O.; Biesiekierska, M.; Panthu, B.; Soszyński, M.; Pirola, L.; Balcerczyk, A. Citrullination in the pathology of inflam matory and autoimmune disorders: recent advances and future perspectives. Cell. Mol. Life Sci. 2022, 79, 94. [Google Scholar] [CrossRef]
- Bay-Jensen, A.C.; Sand, J.M.B.; Genovese, F.; Siebuhr, A.S.; Nielsen, M.J.; Leeming, D.J.; Manon-Jensen, T.; Karsdal, M.A. Chap ter 31 - Structural Biomarkers. In Biochemistry of Collagens, Laminins and Elastin, 1st ed.; Karsdal, M., Ed.; Elsevier: Radarweg 29, 1043 NX Amsterdam, The Netherlands, 2016; pp. 203–233. [Google Scholar] [CrossRef]
- Yasuda, T.; Tahara, K.; Sawada, T. Detection of salivary citrullinated cytokeratin 13 in healthy individuals and patients with rheumatoid arthritis by proteomics analysis. PLoS One. 2022, 17, e0265687. [Google Scholar] [CrossRef]
- GTEx Consortium. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science. 2020, 369, 1318–1330. [Google Scholar] [CrossRef]
- Anzilotti, C.; Pratesi, F.; Tommasi, C.; Migliorini, P. Peptidylarginine deiminase 4 and citrullination in health and disease. Au toimmun. Rev. 2010, 9, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Tar, I.; Csősz, É.; Végh, E.; Lundberg, K.; Kharlamova, N.; Soós, B.; Szekanecz, Z.; Márton, I. Salivary citrullinated proteins in rheumatoid arthritis and associated periodontal disease. Sci. Rep. 2021, 11, 13525. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, W.; To, M.; Yamamoto, Y.; Inaba, K.; Yakeishi, M.; Saruta, J.; Fuchida, S.; Hamada, N.; Tsukinoki, K. Detection of anti-citrullinated protein antibody (ACPA) in saliva for rheumatoid arthritis using DBA mice infected with Porphyromonas gingivalis. Arch. Oral. Biol. 2019, 108, 104510. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Esparza, R; Rodríguez-Rodríguez, M. ; Pérez-Pérez, M.E.; Badillo-Soto, M.A.; Torres-Del-Muro, F.; Bollain-Y-Goytia, J.J.; Pacheco-Tovar, D.; Avalos-Díaz, E. Posttranslational Protein Modification in the Salivary Glands of Sjögren’s Syndrome Patients. Autoimmune Dis. 2013, 2013, 548064. [Google Scholar] [CrossRef]
- Giglione, C.; Fieulaine, S.; Meinnel, T. N-terminal protein modifications: bringing back into play the ribosome. Biochimie 2015, 114, 134–146. [Google Scholar] [CrossRef]
- Deng, S.; Marmorstein, R. Protein N-Terminal Acetylation: Structural Basis, Mechanism, Versatility, and Regulation. Trends Biochem. Sci. 2021, 46, 15–27. [Google Scholar] [CrossRef]
- Vijayan, D.K.; Zhang, K.Y.J. Human glutaminyl cyclase: Structure, function, inhibitors and involvement in Alzheimer’s disease. Pharmacol. Res. 2019, 147, 104342. [Google Scholar] [CrossRef]
- Castagnola, M.; Cabras, T.; Iavarone, F.; Vincenzoni, F.; Vitali, A.; Pisano, E.; Nemolato, S.; Scarano, E.; Fiorita, A.; Vento, G.; et al. Top-down platform for deciphering the human salivary proteome. J. Matern. Fetal Neonatal Med. 2012, 25, 27–43. [Google Scholar] [CrossRef]
- Manconi, B.; Cabras, T.; Pisano, E.; Nemolato, S.; Inzitari, R.; Iavarone, F.; Fanali, C.; Sanna, M.T.; Tirone, C.; Vento, G.; et al. 1637 Characterization of two isoforms of SPRR3 from preterm human newborn saliva, autoptic fetal oral mucosa, parotid and sub mandibular gland samples. Biochem. Biophys, Res. Commun. 2010, 398, 477–481. [Google Scholar] [CrossRef]
- Cabras, T.; Iavarone, F.; Manconi, B.; Olianas, A.; Sanna, M.T.; Castagnola, M.; Messana, I. Top-down analytical platforms for the characterization of the human salivary proteome. Bioanalysis 2014, 6, 563–581. [Google Scholar] [CrossRef]
- Van Damme, P.; Hole, K.; Pimenta-Marques, A.; Helsens, K.; Vandekerckhove, J.; Martinho, R.G.; Gevaert, K.; Arnesen, T. NatF Contributes to an Evolutionary Shift in Protein N-Terminal acetylation and is important for normal chromosome segrega tion. PLoS Genet. 2011, 7, e1002169. [Google Scholar] [CrossRef] [PubMed]
- Varland, S.; Osberg, C.; Arnesen, T. N-terminal modifications of cellular proteins: the enzymes involved, their substrate speci ficity and biological effects. Proteomics 2015, 15, 2385–2401. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, R.; Alves, R.; Barro, A.; Caseiro, A.; Ferreira, R.; Lobo, M.C.; Bastos, A.; Duarte, J.; Carvalho, D.; Santos, L.L. Finding new translational modifications in salivary proline-rich proteins. Proteomics, 3742. [Google Scholar] [CrossRef]
- Schilling, S.; Wasternack, C.; Demuth, H.U. Glutaminyl cyclase from animals and plants: a case of functionally convergent protein evolution. Biol. Chem. 2008, 389, 983–991. [Google Scholar] [CrossRef]
- Schlenzig, D.; Manhart, S.; Cinar, Y.; Kleinschmidt, M.; Hause, G.; Willbold, D.; Funke, S.A.; Schilling, S.; Demuth, H.U. Py roglutamate formation influences solubility and amyloidogenicity of amyloid peptides. Biochemistry 2009, 48, 7072–7078. [Google Scholar] [CrossRef] [PubMed]
- Wirths, O.; Breyhan, H.; Cynis, H.; Schilling, S.; Demuth, H.U.; Bayer, T.A. Intraneuronal pyroglutamate-Abeta 3-42 triggers neurodegeneration and lethal neurological deficits in a transgenic mouse model. Acta Neuropathol. 2009, 118, 487–496. [Google Scholar] [CrossRef]
- Bersin, L.M.; Patel, S.M.; Topp, E.M. Effect of ‘pH’ on the rate of pyroglutamate formation in solution and lyophilized sol ids. Mol. Pharm. 2021, 18, 3116–3124. [Google Scholar] [CrossRef]
- Castagnola, M.; Cabras, T.; Vitali, A.; Sanna, M.T.; Messana, I. Biotechnological implication of the salivary proteome. Trends Biotechnol. 2011, 29, 409–418. [Google Scholar] [CrossRef]
- Schulte, S.J.; Huang, J.; Pierce, N.A. ; Hybridization Chain Reaction Lateral Flow Assays for Amplified Instrument-Free At Home SARS-CoV-2 Testing. ACS Infect. Dis. 2023, 9, 450–458. [Google Scholar] [CrossRef]
- Yu, H.; Lee, H.; Cheong, J.; Woo, S.W.; Oh, J.; Oh, H.K.; Lee, J.H.; Zheng, H.; Castro, C.M.; Yoo, Y.E.; et al. A rapid assay provides on-site quantification of tetrahydrocannabinol in oral fluid. Sci. Transl. Med. 2021, 13, eabe2352. [Google Scholar] [CrossRef]
Figure 1.
Workflow applied for studying the origin of PTMs of salivary peptides and proteins secreted by the major salivary glands by a top-down proteomic approach.
Figure 1.
Workflow applied for studying the origin of PTMs of salivary peptides and proteins secreted by the major salivary glands by a top-down proteomic approach.
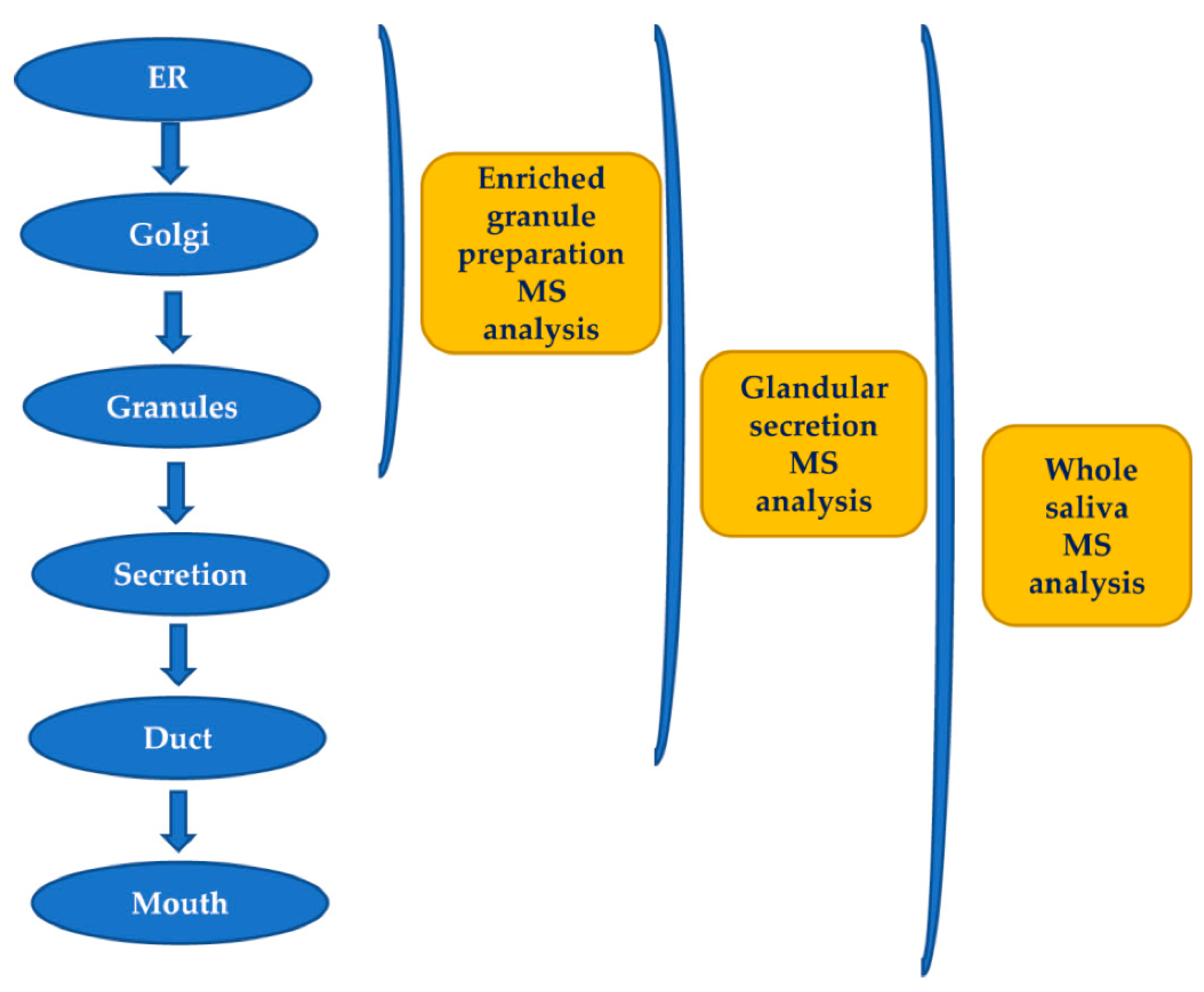
Figure 2.
Principal aPRP proteoforms, in blue the entire proteoforms, in red the truncated ones. Red arrows indicate the cleavage site recognized by convertase activity, from which PRP-3 type truncated forms and P-C peptide are generated from the entire PRP-1 type forms. Grey arrows indicate the possible loss of C-terminal residue by carboxypeptidase activity. Pyroglutamination N-terminal is evidenced in yellow, while the Ser residues that can undergo to phosphorylation are evidenced in light green. The different phosphorylated proteoforms obtainable from the entire and truncated aPRPs are shown, as well as the disulfide dimeric form of Pa.
Figure 2.
Principal aPRP proteoforms, in blue the entire proteoforms, in red the truncated ones. Red arrows indicate the cleavage site recognized by convertase activity, from which PRP-3 type truncated forms and P-C peptide are generated from the entire PRP-1 type forms. Grey arrows indicate the possible loss of C-terminal residue by carboxypeptidase activity. Pyroglutamination N-terminal is evidenced in yellow, while the Ser residues that can undergo to phosphorylation are evidenced in light green. The different phosphorylated proteoforms obtainable from the entire and truncated aPRPs are shown, as well as the disulfide dimeric form of Pa.
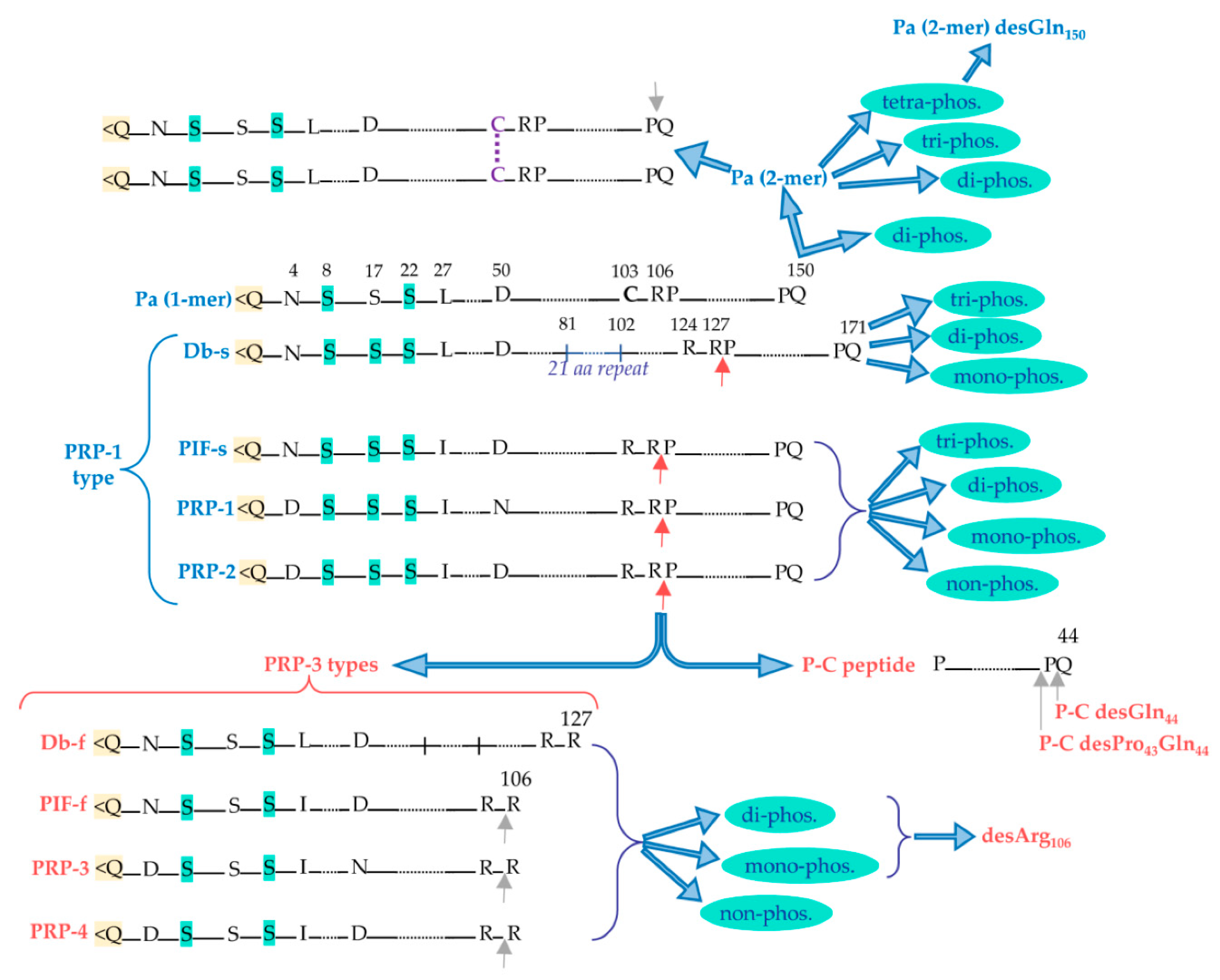
Figure 3.
Classification of bPRPs based on similarity of the N-terminal motif and on the presence of glycosylations. Group 1 in panel (a), group 2 in panel (b), group 3 in panel (c).
Figure 3.
Classification of bPRPs based on similarity of the N-terminal motif and on the presence of glycosylations. Group 1 in panel (a), group 2 in panel (b), group 3 in panel (c).
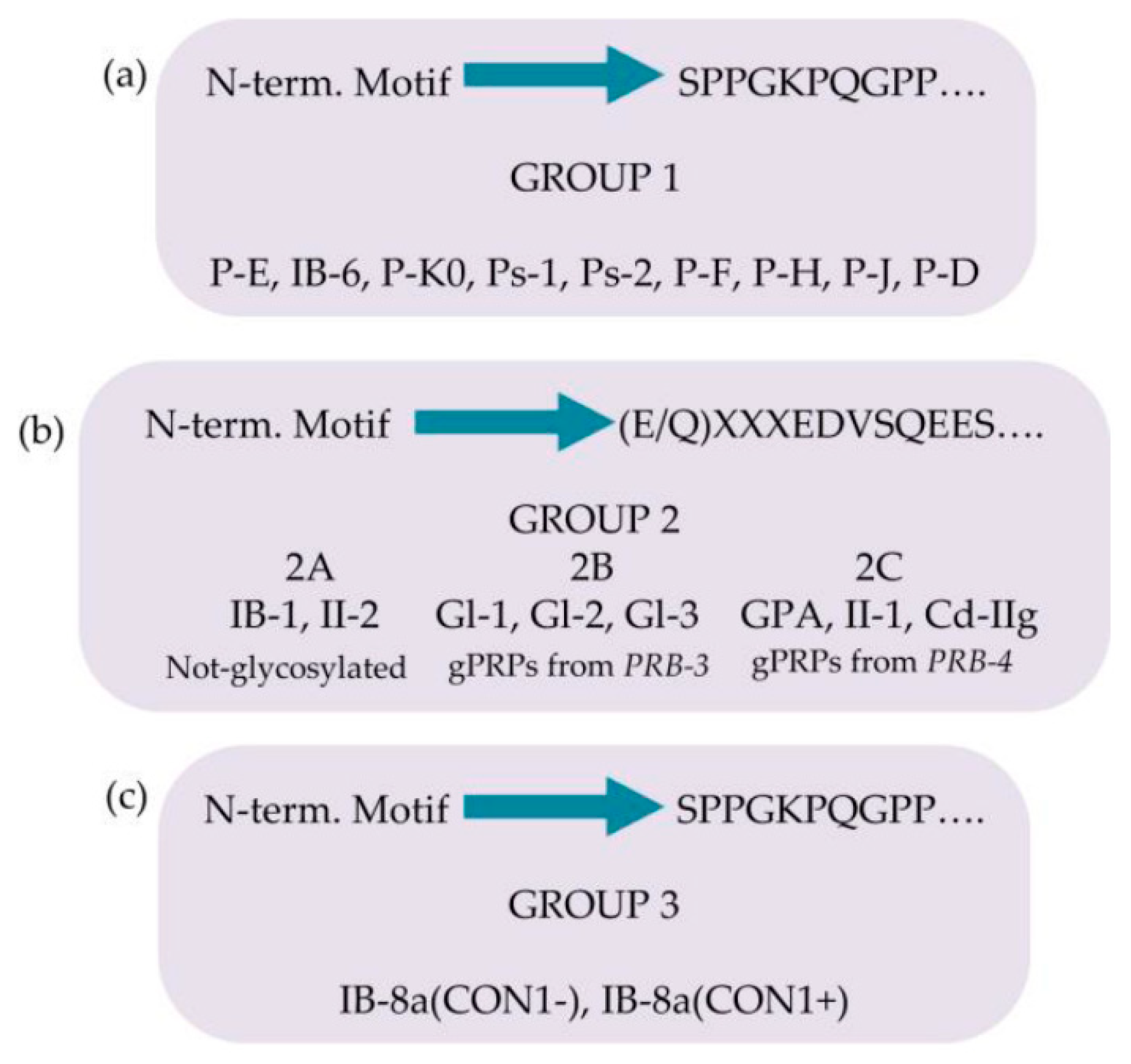
Figure 4.
Pattern of proteoforms, mainly proteolytic fragments, originating from statherin (a), and 321 P-B peptide (b), characterized in human saliva by a top-down approach based on RP-HPLC-ESI MS/MS analysis. The arrows indicate the main cleavage sites. The sequences recognized by furin like convertase are evidenced in red.
Figure 4.
Pattern of proteoforms, mainly proteolytic fragments, originating from statherin (a), and 321 P-B peptide (b), characterized in human saliva by a top-down approach based on RP-HPLC-ESI MS/MS analysis. The arrows indicate the main cleavage sites. The sequences recognized by furin like convertase are evidenced in red.

Figure 5.
Map of reversible, and irreversible oxidative modifications that can occur on the thiol group of the Cys residue.
Figure 5.
Map of reversible, and irreversible oxidative modifications that can occur on the thiol group of the Cys residue.
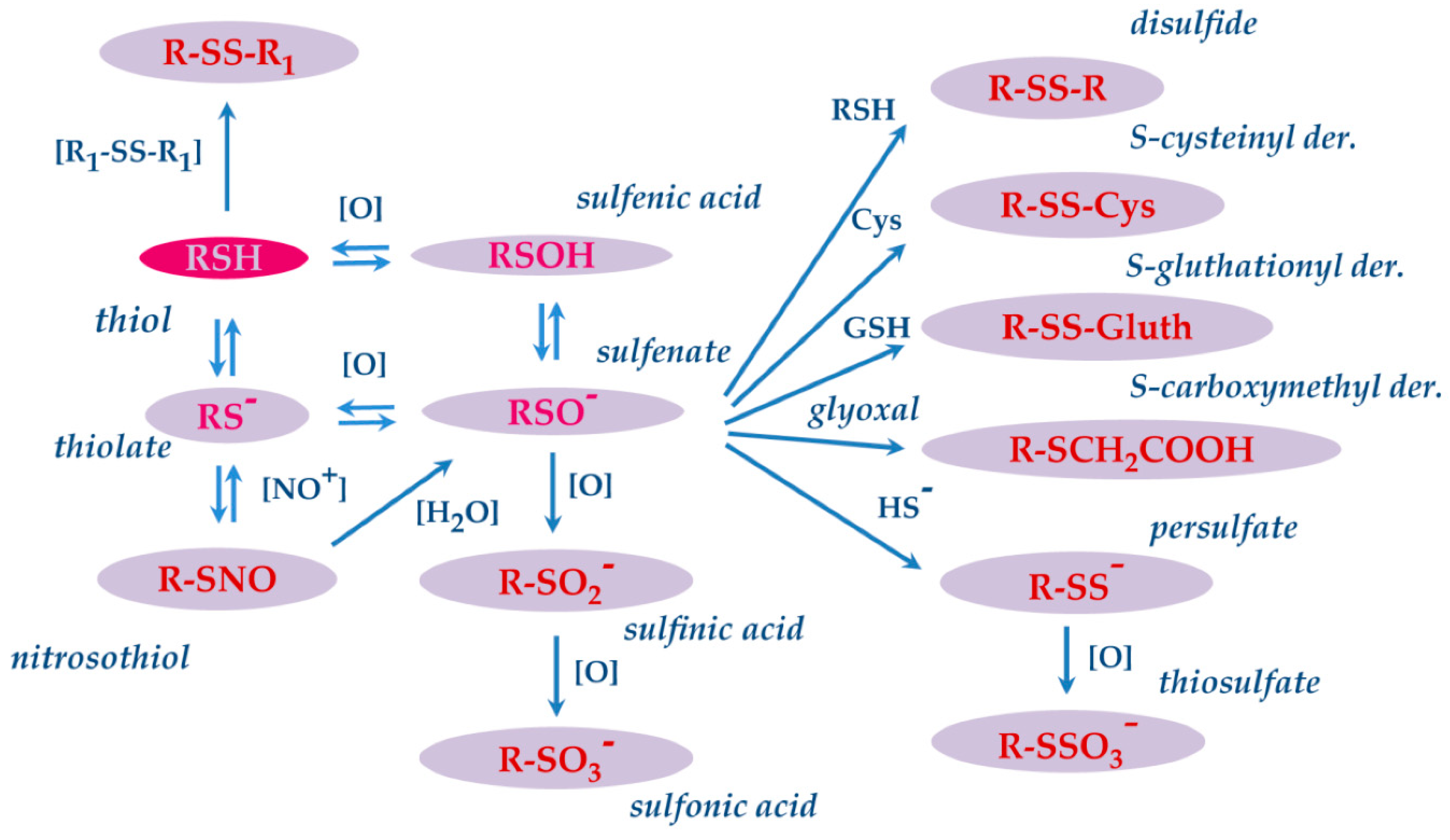

Table 1.
Some terminal losses detected in salivary peptides/proteins.
| Protein and Peptides | C-term. cleavage |
|---|---|
| aPRP Pa-dimer | …..QSP↓Q |
| PRP3/PRP4/PIF-f | …..RPP↓R |
| P-C (or IB-8b) | ….QS↓P↓Q |
| bPRP II-2 | …..RSP↓R |
| bPRP IB-1 | .….RSP↓R |
| bPRP PF (or IB-8c) precursor | .…RSA↓R |
| bPRP PE (or IB-9) | ….RSP↓R |
Table 3.
Phosphorylation sites recognized by Fam20C of human specific secretory proteins.
| Protein/peptide | Partial sequence | Consensus | Ref.s |
|---|---|---|---|
| Statherin (Ser2, Ser3) | DSSEEKF… | SXE | [41] |
| Histatin 1 (Ser2) | DSHEKR… | SXE | [103] |
| Cystatin S (Ser3) | SSSKEENR… | SXE | [57,58,59,83] |
| Cystatin S1 (Ser1) | SSS(Phos)KEENR… | SXS(Phos) | “ |
| aPRP (Types 1 and 3) (Ser8) | …DEDVSQEDV…. | SXE | [9] |
| aPRP (Types 1 and 3) (Ser22) | …GGDSEQFIDEER… | SX(3/4)(E/D/S(phos))3 | “ |
| aPRP (Types 1 and 3) (Ser17) | …LVISDGG DS(phos)EQFI… | SX(3/4)(E/D/S(phos))3 | “ |
| bPRP II-2 (Ser8) | …NEDVSQEESPS… | SXE | [23] |
| bPRP IB-1 (Ser8) | …NEDVSQEESPS… | SXE | “ |
| gPRP Gl-2 or PRP-3M (Ser8) | …NEDVSQEESPS… | SXE | “ |
| gPRP Gl-1 or PRP-3L (Ser8)1 | …NEDVSQEESPS… | SXE | “ |
| gPRP Gl-3 or PRP-3S (Ser8)1 | …NEDVSQEESPS… | SXE | “ |
| gPRP Glycosyl. Protein A (Ser8)1 | …SEDVSQEESLFL… | SXE | “ |
| gPRP II-1 (Ser8)1 | …SEDVSQEESLFL… | SXE | “ |
| gPRP Cd-IIg (Ser8)1 | …SEDVSQEESLFL… | SXE | “ |
1Hypothetical (by similarity).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



