
Submitted:
07 August 2023
Posted:
08 August 2023
You are already at the latest version
Abstract
Background: allergic diseases and, in particular food allergy (FA), have been identified as one of the major diseases in the world. The first 1000 days of life can be a “window of opportunity” or a “window of susceptibility” during which several factors can predispose children to FA development. Changes in gut microbiota composition, from pregnancy to infancy, could have a pivotal role in this regard: some bacterial genus such as Lactobacillus or Bifidobacterium seem to be protective from FA development, on the contrary Clostridium and Staphylococcus appear to be unprotective.
Methods: we conducted a research through the most recent literature (2013-2023), using Pubmed and Scopus as databases. We included English original papers, clinical trials, metanalysis and reviews. Case reports, series and letters have been excluded.
Results: during pregnancy, maternal diet can play a fundamental role in influencing gut microbiota composition of newborns. After birth, human milk can promote the development of protective microbial species by human milk oligosaccharides (HMOs), that play a prebiotic role. Also complementary feeding can modify gut microbiota composition.
Conclusions: the first two years of life are a critical period where several factors, in genetic predisposed children, can increase the risk of FA development.
Keywords:
gut microbiota
; human milk oligosaccharides (HMOs)
; complementary feeding
; food allergy (FA)
; breastfeeding
; first 1000 days of life
; dysbiosis
; newborns
1. Introduction
Allergic diseases have been identified by the World Health Organization (WHO) as one of the major diseases in the world. They include asthma, food allergy (FA), allergic rhinitis, eczema and others, that are traditionally referred to as type 1 hypersensitivity reactions [1]. The term “sensitization” refers to a process where T and B cells respond to an allergen leading to production of specific IgE antibodies. After a new allergen exposure, sensitization may or may not be associated with clinical allergic reaction [2]. In the last two decades, allergic diseases’ prevalence in childhood has been increasing [3]. In the USA, 1 in 13 children suffers from FA [4]. FA results from an abnormal response of the immune system to food antigens and can be life-threatening in children, by impacting their quality of life [4]. It can be either IgE-mediated or not, or both; nowadays, IgE-mediated FA is the best characterized and it is the classically recognized type in society [5]. IgE molecules occupy their receptors on the surface of basophils and mast cells; allergens, by cross-linking these complexes, stimulate the release of local bioactive mediators such as histamine [6]. Allergic diseases are related to enhanced responses from Th2 cells. During pregnancy Th1 immune response of the fetus is suppressed to prevent excessive response to maternal antigens; at birth, there is an increase toward Th2 response to novel antigens. Exposure to gut microbiota shifts this response to development of Th1 cells, which promotes immune tolerance and T-cells maturation [2,7]. Gut microbiota plays a key role in modulating and influencing immune tolerance [8], and early microbial exposure is very important for children’s immune development [4]. According to the “hygiene hypothesis”, proposed by Dr. David Strachan in the 1980s [9], an exposure to infection in the first months of life would be protective against allergic diseases. An early life microbial experience is associated with an improvement of immune development and with a reduced risk of allergic diseases’ onset [2,10]. In fact, a farm environment with a diverse range of environmental microbes to which children can be subjected, is associated with a reduced risk of allergic diseases, especially asthma [11]; on the contrary, an excessive sanitized Western lifestyle can disrupt children’s gut microbiota and disturb normal immune system development [12]. The “hygiene hypothesis” has been later expanded by the “microflora hypothesis” in 2005, which indicates that an early disturbance of the infants’ gut microbiota could alter immunological tolerance and make the immune system toward a state of allergic disease [1]. A dysbiosis of the gut microbiota is, in fact, associated with an increased intestinal permeability, an aberrant inflammatory response and with a different composition of the microbiota. Generally, it has been shown a reduction in Lactobacilli and Bifidobacteria that seem to correlate with protection against atopy by favoring Th1 immune response. On the contrary, early colonization by Clostridium difficile and Staphylococcus aureus is more likely to occur in children who develop allergy later in life [8]. In this context, breastfeeding plays a pivotal role in modulating immune system development of newborns and in the establishment of a healthy gut microbiota [13]. Several factors can influence and alter the composition of pregnants and newborns’ gut microbiota: mode of delivery, feeding practices, hygienic and social status, gestational age, use of antibiotics, maternal and infant’s diseases [14]. Maternal gut, the breast and the infant gut are linked by the so called “Gut-Breast Axis”: a correct functionality of gastrointestinal tract is necessary to assemble the components of human milk which modulates the development of newborn’s gastrointestinal tract, with lifelong effects [15]. Aim of this narrative review is to summarize the main changes of microbiota in the first years of life, during fetal, neonatal and early childhood periods, and to describe how these modifications may negatively predispose to FA onset in children.
2. Methods
A narrative review has been performed according to the most recent available literature (2013-2023). English original papers, clinical trials (both in humans and animals), metanalysis and reviews have been included, rather, case reports or series and letters have been excluded. The research has been conducted using the following keywords (alone or in combination): children, childhood, infants, newborns, gut microbiota, human milk, breastfeeding, complementary feeding, food allergy, diet, pregnancy, epigenetics, allergy, oligosaccharides, dysbiosis. The following electronic databases were used: PubMed and Scopus.
3. Epigenetics and Gut Microbiota
The term “microbiota” refers to a population of microorganisms that exist within a niche in the human body, with a mutualistic relationship with the host and includes bacteria, viruses, fungi, parasites and archaea. The term “microbiome” refers to the collection of genomes from all these microorganisms [16,17]. Bacterial component is the most prevalent (from 1013 to 1014 organisms) [2]. Bacterial communities are found in the gastrointestinal and urogenital tract, skin and oral cavity but gut microbiota is the most studied [18]. Two major characteristics of the gut microbiota are the richness and the diversity: the first term refers to the total number of bacterial species present; the second one refers to the number of individual bacteria from each of the bacterial species present [19]. It is possible to differentiate between groups at two levels: the alpha and beta diversity [20]. In the study of human microbiota, alpha diversity describes the compositional complexity of a single sample, whereas beta diversity describes taxonomical differences between samples [21]. Atopy seems to contribute to the gut diversity too [8]. A continuous exposure to allergy-protective microorganisms, especially during prenatal and postnatal period, results in a form of innate and adaptive immune training that makes individuals resistant to danger signals from the environment. This is the so-called “hypothesis of inflammatory resilience”. A lack of biodiversity is, in fact, associated with an inappropriate and exaggerated inflammatory response to danger signals [18]. The development of allergy is the result of gene-environment interactions with the epigenetic changes in one or more susceptibility genes [18]. Microbiota-derived metabolites can be used as epigenetic substrates [22]. Epigenetic mechanisms are able to modify gene expression without altering DNA sequence. The major epigenetic processes involved are: DNA methylation patterns, histone modification, chromatin remodeling and microRNAs (miRNAs), even if it is still debated if miRNAs can be considered a real epigenetic phenomenon [19,23,24,25]. Bifidobacteri and Lactobacilli are able to produce folate, an essential molecule involved in methylation processes: changes in bacterial composition can shift the DNA methylation status in the host. At the same time, short-chain fatty acids (SCFAs), produced by commensal microbes during fermentation patterns, are able to influence histone modifications [22]. A systematic review conducted by Hartwig et al. [26], has shown that breastfeeding might influence DNA methylation, although more studies are needed. For example, it has been demonstrated that duration of breastfeeding can be associated with DNA methylation in children. In particular, the hypermethylation of the gene encoding sorting nexin 25 (SNX25) is related to an increase of TGF-beta activity, a cytokine that promotes IgAs secretion [6]. Moreover, in a recent review conducted by Esch et al., [27] it has been demonstrated that allergic diseases are associated with changes in DNA methylation patterns in Th1, Th2, Th9, Th17 and Treg cells. In particular, there would be an alteration in FoxP3+ demethylation, molecule involved in tolerance induction [27,28]. At the same time, milk-derived miRNAs are able to regulate gene expression in a dose-dependent manner [29]. MiRNAs are short noncoding RNA molecules that are able to induce mRNA degradation and/or post-transcriptional inhibition of translation. Some of them are involved in the maturation of Treg cells [27]. Nevertheless, studies on breastfeeding and the role of epigenetic mechanisms in the imprinting effects of breastfeeding or its components in relation to allergic diseases are very limited [6], and further data are needed. It is therefore quite clear that the digestion and metabolism of molecules introduced with the diet can influence the state of health depending on the individual genotype (“nutrigenetics”) and the diet can change the gene expression (“nutritional epigenetics”) [25]. The regulation of gene expression by environmental and dietary exposures through epigenetic mechanisms has a demonstrated role in FA [4,24].
4. First 1000 days of life: “a window of opportunity” in gut microbiota
The first months of life are fundamental in the establishment of a healthy gut microbiome, so several factors can influence its normal composition and function. In this period, unhealthy cues can cause alterations of gene expression later in life increasing the risk for multifactorial environmentally driven diseases such as allergy [19]. In fact, according to Barker's theory [30], the first 1000 days after conception are a critical period in which most part of human development takes place [25]. This time frame corresponds to pregnancy, the neonatal period, and the first two years of life, and is the optimal time for protective factors to exert their beneficial effects (“window of opportunity”) and the risk factors to increase the chance of disease development (“window of susceptibility”), see Figure 1.
In this timeframe, epigenetic intervention (“window of intervention”) might prevent or even revert the negative effects of environmental risk factors [6]. Generally, vaginal delivery, breastfeeding, rural environment, exposure to pets and a high-fiber diet seem to be protective factors against FA onset. To date, no specific bacterial taxa would be associated with FA onset [31]. Anyway, a normal microbiota development involves early colonization by facultative anaerobes (predominantly belonging to the Enterobacteriaceae family), that deplete initial oxygen supplies to create a favorable environment for anaerobes colonization (especially belonging to the Bacteroidaceae family). Members of Bacteroidaceae families are able to stimulate the production of mucin that maintains an intact gut microbiota-mucin barrier. E/B ratio tends to decline over the time: its persistent elevation is very frequent in food-sensitized infants, as a marker of a delayed maturation of the gut microbiota [32] and as a marker of a less mature microbiome [4]. SCFAs, such as acetate, butyrate and propionate derive from bacteria fermentation and they seem to have many beneficial effects in autoimmune and inflammatory diseases [31]. Among these, butyrate is the most important: it might contribute to the development of immune oral tolerance and to the prevention and treatment of FA, by modulating Treg’s activity [24]. Butyrate inhibits the release of proinflammatory cytokines and promotes anti-inflammatory responses [19]. Demonstrating this, butyrate’s levels tend to be lower in children affected by cow’s milk allergy (CMA) at 1 year [4,31]. Moreover, Roduit et al. [33], have shown that children with the highest stool concentrations of butyrate at 1 year of age had significantly less atopic sensitization to food and/or inhalant allergens. At the same time, children were less likely to have a reported diagnosis of FA or allergic rhinitis between 3 and 6 years of age. Therefore, changes in gut microbiota composition during the first months of life could impair the future health of children.
4.1. Gut Microbiota of pregnant women
Environment plays a crucial role in modulating the gut microbiota composition of newborns already in the fetal period [3]. It has been hypothesized that nutritional exposure during the time of immune programming may play a role in the development of FA [34]. The role of maternal diet has been well evaluated: in a prospective cohort study conducted by Brzozowksa et al. [3], it has been shown that a reduced intake of vitamin C and magnesium during pregnancy can be associated to a greater risk of developing wheezing in the first 2 years of life. Moreover, an increased intake of vitamin D, retinoic acid, zinc and copper could positively influence epigenetic patterns associated to the allergic diseases in children [3,6,10,24,34]. At the same time, the perinatal supplementation of polyphenols seems to prevent allergies in offspring [6]. The immunomodulatory role of vitamin D is well known: lower plasmatic levels are associated with an increased risk of allergic diseases’ development [5]. Nevertheless, previous studies have shown a greater risk of overall allergy in children born by vitamin D supplemented mothers [35,36]. Probably, there is a U-shaped relationship between vitamin D levels and risk of allergy: both too little and too much vitamin D correlates with the greatest risk [5,37]. In animal studies, vitamin A supplementation during pregnancy can be a possible intervention for allergy prevention in the neonatal stage of life [38]. Also n-3 long chain-polyunsaturated fatty acids (LC-PUFAs) may modulate the development of IgE-mediated allergic disease and regulate immune responses, by influencing Th1/Th2 balance in infants [24,34]. In particular, in an observational study conducted by Best et al. [39], it has been hypothesized that an increased n-3 LC-PUFA intake during pregnancy, could be associated with a reduction in childhood allergic diseases’ prevalence [40]. In fact, ω-3 PUFAs are able to inhibit Th cell differentiation, moving away from development of allergy [10]. An increase in PUFAs’ levels is correlated to an abundance of Holdemania spp. in maternal feces during pregnancy: it is a gram-positive anaerobic bacterial genus associated to a reduced risk of FA in offspring and it can be used as a predictor marker [41]. Nevertheless, if it is true that n-3 LC-PUFA supplementation in pregnancy is associated with a reduced risk of allergy in children, it has not shown a positive influence when children are directly supplemented. Maybe, because pregnancy is an important time that influences the immune system development more than does the early postnatal period [27]. Since it has been shown that breastfeeding has a protective role against FA onset due to the significant levels of butyrate contained therein, it might be useful to increase the concentration of butyrate in human milk (HM) through the modulation of maternal diet [24,31]. A maternal diet rich in vegetables seems to be associated with a reduced risk of allergic disease in children, due to the increased diversity and richness of gut microbiota (Holdemania, Roseburia, Lachnospira and Coprococcus spp.) [11]. In a prospective population-based cohort study conducted by Tuokkola et al. [42], it has been demonstrated that diet during pregnancy could have a bigger role in influencing the development of FA rather than lactation period, suggesting antigen-specific induction of tolerance in subjects genetically not predisposed to the development of allergic disease. Nevertheless, no clear results have been established so, no recommendations for clinical practice are still available [3,10,38,42]. In fact, according to the most recent literature, the American Academy of Pediatrics (AAP) has concluded that there is not sufficient evidence to support maternal dietary restriction during pregnancy or lactation to prevent atopic diseases in offspring [4]. Also antibiotic exposure during pregnancy might have an important role in influencing gut microbiota composition of the offspring and, consequently, FA, eczema and asthma prevalence [11]; in murine models, it has been already shown that the use of antibiotics in pregnancy might enhance food sensitization in offspring [32]. For example, Prevotella copri is the predominant Prevotella species within the human gut microbiota: maternal antibiotic exposure is associated with its reduction and, consequently, with a minor protection against allergic diseases [11]. In fact, the abundance of Prevotella in the maternal gut microbiota during pregnancy has a protective effect on FA, independently from Prevotella abundance in the gut microbiota of the offspring [9,41]. Use of antibiotics in pregnancy has been associated with a lower count of Bifidobacteri in the neonatal gut within the first month of life [2]. Also the environment where the pregnant lives is important too: children whose mothers live in a farm environment would have an increasing number of Treg cells due to a higher level of stimulatory bacterial lipopolysaccharides (LPS) variants [43]. In a recent systematic review conducted by Venter et al. [44], it has been proposed an index of maternal diet during pregnancy by which we would be able to predict the risk of allergic diseases’ development in offspring. The frequency of consuming vegetables, yogurt, fried potatoes, rice, red meats, fruit juice and cold cereals was used to obtain data from which to derive the predictive index of development of allergic pathology in offspring. Vegetables and yogurt seem to be protective factors against allergy development, on the contrary, fried potatoes, rice, red meats, fruit juice and cold cereals would be risk factors [44]. Asnicar et al. [45] have shown that a higher intake of vegetables and yogurt is associated to a more diverse microbiome and also to higher levels of fecal butyrate, with protection from allergy development up to school age. Nevertheless, further studies are needed.
4.2. Gut Microbiota in the first months of life
The colonization of the neonatal gut by the microbiota is a very vulnerable period [14,46]. The establishment of gut microbiota in infants initiates during pregnancy by translocation of maternal microorganisms through vagina, maternal gut, placental tissue and meconium [47]. At the beginning there is a prevalence of strict anaerobic bacteria such as Bifidobacteri, Clostridi and Bacteroides [14]. Although studies are contradictory, probably sensitization to milk and egg allergens is twice as likely to occur in children born by cesarean section (CS) [11,32]. Type of delivery, in fact, correlates to a different composition in gut microbiota: newborns via vaginal birth display a higher abundance of Bifidobacteri, Bacteroides, Lactobacilli and bacteria belonging to the Lachnospiraceae family, rather, bacteria belonging to Enterococcaceae and Enterobacteriaceae families are most abundant in CS-delivered newborns [47]. Gut microbiota composition appears healthier and more diverse in babies born vaginally when compared with infants born by CS. Moreover, maternal vaginal bacteria (i.e. Lactobacillus and Prevotella spp.) are commonly found in infants born vaginally, while maternal skin microbes (i.e Staphylococcus and Propionibacterium spp.) are prevalent in infants born by CS [19]. It is interesting that not only the mode but also the place of delivery can play a pivotal role: it has been demonstrated that in children with a positive family history for atopy, vaginal home delivery is more protective than vaginal hospital one [8]. Atopic dermatitis is associated with lower gut microbiota diversity and relative abundance of Bacteroides spp. by 1 month; an early colonization by Clostridium difficile at 1 month, could be predictive of atopic sensitization at 2 years. At the same time, anaerobes’ abundance is most frequent in infants with confirmed allergy to cow’s milk. At 3 months, lower microbiota richness is a risk factor for food sensitization at 1 year of age; on the contrary, it has been shown that richness at 1 year is not associated with food sensitization. In fact, the critical period for microbiota development is “early infancy”. Food-sensitized infants have an elevation in bacteria belonging to Enterobacteriaceae family (i.e. Escherichia, Shighella spp), even if there is a concomitant general reduction in Proteobacteria that is the phylum containing Enterobacteriaceae [32]. Maybe, this apparent contradiction is due to the concomitant reduction in other bacterial families belonging to Proteobacteria phylum [48,49]. At the same time, in this slice of population, a lower relative abundance of Bacteroidaceae bacterial family has been shown [24,32]. Nevertheless, in a recent population-based cohort study, gut microbiota composition of the first-pass meconium seems not to be associated with later atopic manifestations in children [50]. The increasing use of antibiotics in childhood is a topic of concern: in a case-control study conducted by Hirsch et al. [12], it has been shown that there is an association between antibiotics orders and several independent allergy diagnosis, especially regarding macrolides, in a dose-response relation. Probably, it is due to the capacity of antibiotics to modify the composition of gut microbiome, leading to functional changes that promote the development of allergic diseases. This effect seems to persist for a long period: penicillin and cephalosporins are more associated with FA in the first 2 years of life, rather macrolides are associated with FA later in childhood [51]. The increasingly massive use of antibiotics from the first months of life can affect the composition of gut microbiota and cause “dysbiosis”, predisposing children to develop FA [see Section 5].
4.2.1. Role of Human Milk Microbiota
For a long time, HM was thought to be sterile, but recently, many studies have demonstrated that it provides a rich source of microbes, whose changes can influence children's health. Two main sources of milk microbiota origin are known: the retrograde flow and the entero-mammary pathway. The first one is the consequence of the transmission of microbes from the infant’s oral cavity into the duct during suckling; the second pattern would be the consequence of maternal gut bacteria translocation through the intestinal epithelial barrier [52]. Staphylococcus, Streptococcus, Serratia, Pseudomonas, Corynebacterium, Ralstonia, Propionibacterium, Sphingomonas and Bradyrhizobium are the nine genera that constitute the “core” bacteriome of the HM microbiota. They represent approximately half of the microbial milk community, although their abundance can vary across milk samples [17,53,54]. The “Mother-HM-Infant” triad underlines the close connection between pregnant and newborn, by human milk, in modulating the trajectory of infant development [55]. Breast milk provides a quarter of the intestinal microbiota of infants and its composition could influence the development of FA in children. It has been shown that a relative abundance of Prevotella spp. in the breast milk microbiota is more common in mothers whose children will not develop FA [56]. Moreover, it has been shown that in the breastmilk of infants with allergic symptoms there is a relative abundance of Proteobacteria, especially Acinetobacter and Pseudomonas spp. [56]. Nevertheless, data about the protective role of HM against FA development are contradictory still now [57].
4.2.2. Role of Human Milk Oligosaccharides (HMOs)
HM is the golden standard for nutrition of newborns that can impact on FA development. “Enteromammary link” is an active connection between immune tissues in the maternal gut and the mammary glands that results in a human milk-specific IgA profile that can influence infants’ microbiome composition [38]. In early postnatal life, IgAs’ deficiency might be associated with a higher risk of atopic dermatitis [2]. Human milk is a dynamic bio fluid that contains a wide range of macro and micronutrients. In that respect, human milk oligosaccharides (HMOs) have gained considerable attention, especially for their prebiotic role in stimulating Bifidobacteria growth, the dominant bacterial genus in breastfed infants along with Lactobacilli and bacteria belonging to Enterobacteriaceae family [38]. Generally, in breastfed infants there is an abundance in Bacteroidota, on the contrary Firmicutes are prevalent in formula-fed infants [47]. A reduced number of Bifidobacterium spp. is associated with the development of atopic diseases later in life [14,58]. In fact, Bifidobacteria are able to induce mast cell apoptosis, reducing allergic symptoms [47]. In particular, Bifidobacterium infantis is able to digest HMOs (“inside eater”) [59], producing metabolites that support infant development at the cellular level [58]. In particular, B. infantis is able to produce indole-3-lactic acid, a metabolite which decreases enteric inflammation and exerts regulatory effects on Th2 and Th17 cells [4]. In a recent study conducted by Dai et al. [58], it has been pointed out that Bifidobacterium infantis is able to maintain diversity of gut microbiota after antibiotic exposure, making children less vulnerable to asthma onset. Nevertheless, it has been shown that microbial diversity is more beneficial in adults than in children [4]. HMOs also exert direct immune modulating effects, by promoting immune system maturation [14]. About 200 different compositions of HMOs have been characterized: they consist of a lactose core that can be elongated by glucose, galactose, N-acetylglucosamine, fucose and sialic acid monosaccharides. HMOs are not digested in the upper part of the gastrointestinal tract of infants, so they can reach the colon and are used as a substrate for specific microbes. Nowadays, cow-milk derived infant formulas are often supplemented with non-digestible carbohydrates (galacto-oligosaccharides, GOS; fructo-oligosaccharides, FOS) to substitute HMOs functions [14]. Moreover, it has been shown how a specific HMO profile in human milk can be associated with a reduced risk of food sensitization in the infant (for example, lacto-N-fucopentaose III, LNFP-III, would play a protective role) [38]. Another molecule contained in human milk which could have a potential role in preventing FA development is TGF-beta, a cytokine with potent tolerogenic properties, although studies are not conclusive [38,60]. Further studies are necessary to evaluate the real link between FA and breastfeeding.
4.3. Gut Microbiota and Complementary Feeding
Gut bacterial colonization is a crucial event in infancy to establish oral tolerance and to set a functional digestive tract [61]. Before weaning, gut microbiota is generally enriched by lactate-producing bacteria, on the contrary when solid foods are introduced in the diet, there is a growth of bacteria able to use a larger variety of carbohydrates and vitamins [61]. During complementary feeding, gut microbiota is characterized by a greater diversity in bacteria species, especially Enterococci, Enterobacteria, Clostridi and Bacteroides [24,38], and by a relative abundance in anaerobic bacteria [2]. The fluctuations in abundance of Actinobacteria and Firmicutes in the first 6 months of life seem to be beneficial for FA prevention [41]. Moreover, progression in complementary feeding is associated with abundance in Lachnospiraceae and Ruminococcaceae spp., and with a reduction in Bifidobacterium spp. This is the result of a transition from a breast-milk promoted Bifidobacteria gut community toward a fiber and protein promoted gut microbial community (more diverse) [62]. Timing of introduction of complementary feeding is important in influencing infant gut microbiota diversity [63]. In a prospective cohort study conducted by Differding et al., it has been shown that an early introduction of complementary feeding (before 3 months vs later) was associated with higher gut microbiome diversity and more abundance of fecal SCFAs (especially butyrate) at 12 months [64]. To date, higher fecal concentrations of butyrate and propionate in adults would be related to worse metabolic outcomes, on the contrary higher serum levels of SCFAs would be related to better health [65,66]. Anyway, studies are not conclusive in infants. A prospective longitudinal study conducted by Pannaraj et al. [67], has demonstrated that early weaning is associated with a faster maturation in gut microbiome composition: for example, there would be a reduction in Bifidobacteria spp. and an increase in more adult-associated bacteria (i.e genus Bacteroides) [68]. The establishment and development of the infant microbiome would continue even after complementary feeding, up to 3 years of life, when the child's gut microbiota appears more similar to the adult's one [2]. Recent works suggest that a typical adult pattern of the gut microbiota may not be established before adolescence [57]. The delay in introducing allergenic foods during complementary feeding (beyond 4 to 6 months of life), is not recommended to date [4,60]. American Academy of Pediatrics asserts that there is no sufficient evidence to support delayed introduction of potential food allergens into infants’ diet; on the contrary, an early introduction of these foods could be even protective against FA development [69]. For example, Learning Early About Peanut Allergy (LEAP) study shows that early introduction (4-11 months) of peanuts into children’s diet at high risk of developing FA is effective in reducing peanut allergy development compared to avoidance [70]. Nevertheless, there are not other convincing studies regarding other foods. It has been proposed that 17 weeks of life is a crucial time point, with solid food introduction before this time appearing to facilitate allergic diseases whereas solid food introduction after this time seems to promote tolerance. Probably, it is due to the fact that foods introduced after 17 weeks of age are mainly fruit and vegetables, which are not considered allergenic and which are able to promote nonallergen-specific tolerogenic immunologic mechanisms. Fruit and vegetables, in fact, are able to induce Treg cells maturation by epigenetics mechanisms [71]. Finally, also complementary feeding practice (“a baby-led” approach vs “traditional spoon-fed” one) seems to influence gut microbiota composition in children; in particular, baby-led weaning is associated with lower diversity and lower levels of Lachnospiraceae [63,72], traditionally related to egg sensitization [73].
5. Microbiota’s dysbiosis: mechanisms associated with FA
Dysbiosis is an imbalance in the gut microbial community; recent data confirm that gut dysbiosis precedes FA onset [18,31]. In fact, disruption in early gut microbiota development may alter gut epithelial integrity, affect the immune system and potentiate the allergic response [2]. In particular, the massive use of antibiotics is an important risk factor in inducing intestinal dysbiosis. Gut microbiota dysbiosis is correlated to systemic and local inflammation, with a consequent intestinal barrier damage. A modified microbiota, with a reduction of beneficial bacteria, increases the susceptibility to and severity of FA [74]. So, it is important to create and maintain a symbiosis between humans and their commensal gut organisms: if this relationship becomes unbalanced, several pathological processes can happen [2]. Gut microbiota of children affected by FA is characterized by a reduction in Bacteroides, Bifidobacteri and Clostridi spp., with a consensual abundance in Anaerobacter spp. [48,57]. At the same time, Azad et al. [32], demonstrated that there was a low diversity in bacterial species in gut microbiota of food-sensitized children, along with a relative abundance in Bacteroides spp. It has been shown that in the gut microbiota of infants with FA there is a reduction in butyrate-producing bacteria [31,47], with colonization by Clostridium paraputrificum and tertium too [61]. Moreover, there would be a difference in gut microbiota composition of children who resolve FA or not in the first 8 years of life: in the first group, Firmicutes are most prevalent, in the second one there would be a higher abundance of Bacteroidota [61]. In future, the use of prebiotics and probiotics (i.e. Lactobacillus rhamnosus) could help to modulate gut microbiota composition [47,60], especially in children with a familiar history of atopy and born by CS [43]. At the same time, innovative approaches as use of synbiotics and fecal microbiota transplantation could create a breakthrough in this area [75].
In Table 1 a taxonomic classification of the main bacterial phyla, families and genus involved in allergic diseases.
In Figure 2, a summary of the main protective and unprotective bacterial genus in children’s gut microbiota.
6. Conclusions
The first 1000 days of life are a critical period where several factors, in genetic predisposed children, can increase the risk of FA development. It is important, where possible, to try to act on the modifiable risk factors which in the very early stages of life (pregnancy-infancy-weaning) can outline a “healthy” rather that “unhealthy” intestinal microbiota, in order to reduce the incidence of dysbiosis, that is related to the severity of FA. Breastfeeding is fundamental to reduce intestinal inflammation, especially due to HMOs’ prebiotic role. In the future, the use of probiotics and synbiotics could also positively modulate gut microbiota composition especially in children with a family history of atopy.
Author Contributions
V.N., M.C., S.A., M.G.: conceptualization and methodology; V.N.: writing original draft; M.G.: review and editing, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
none.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hu, T.; Dong, Y.; Yang, C.; Zhao, M.; He, Q. Pathogenesis of Children’s Allergic Diseases: Refocusing the Role of the Gut Microbiota. Front Physiol 2021, 12, 749544. [Google Scholar] [CrossRef]
- Molloy, J.; Allen, K.; Collier, F.; Tang, M.L.K.; Ward, A.C.; Vuillermin, P. The potential link between gut microbiota and IgE-mediated food allergy in early life. Int J Environ Res Public Health 2013, 10, 7235–7256. [Google Scholar] [CrossRef]
- Brzozowska, A.; Podlecka, D.; Jankowska, A.; Król, A.; Kaleta, D.; Trafalska, E.; Nowakowska-Swirta, E.; Kaluzny, P.; Hanke, W.; Bal-Gierańczyk, K.; Kowalska, M.; Polańska, K.; Jerzyńska, J. Maternal diet during pregnancy and risk of allergic diseases in children up to 7–9 years old from Polish Mother and Child Cohort study. Environ Res 2022, 208, 112682. [Google Scholar] [CrossRef]
- Davis, E.C.; Jackson, C.M.; Ting, T.; Harizaj, A.; Järvinen, K.M. Predictors and biomarkers of food allergy and sensitization in early childhood. Ann Allergy Asthma Immunol 2022, 129, 292–300. [Google Scholar] [CrossRef]
- Poole, A.; Song, Y.; Brown, H.; Hart, P.H.; Zhang, G.B. Cellular and molecular mechanisms of vitamin D in food allergy. J Cell Mol Med 2018, 22, 3270–3277. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, N.; Alhamwe, B.A.; Caraballo, L.; Ding, M.; Ferrante, A.; Garn, H.; Garssen, J.; Hii, C.S.; Irvine, J.; Llinás-Caballero, K.; et al. Perinatal and Early-Life Nutrition, Epigenetics, and Allergy. Nutrients 2021, 13, 724. [Google Scholar] [CrossRef] [PubMed]
- Rackaityte, E.; Halkias, J. Mechanisms of Fetal T Cell Tolerance and Immune Regulation. Front Immunol 2020, 11, 588. [Google Scholar] [CrossRef]
- Tsabouri, S.; Priftis, K.N.; Chaliasos, N.; Siamopoulou, A. Modulation of gut microbiota downregulates the development of food allergy in infancy. Allergol Immunopathol (Madr) 2014, 42, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Vuillermin, P.J.; O’Hely, M.; Collier, F.; Allen, K.J.; Tang, M.L.K.; Harrison, L.C.; Carlin, J.B.; Saffery, R.; Ranganathan, S.; Sly, P.D.; et al. Maternal carriage of Prevotella during pregnancy associates with protection against food allergy in the offspring. Nat Commun 2020, 11, 1452. [Google Scholar] [CrossRef]
- Netting, M.J.; Middleton, P.F.; Makrides, M. Does maternal diet during pregnancy and lactation affect outcomes in offspring? A systematic review of food-based approaches. Nutrition 2014, 30, 1225–1241. [Google Scholar] [CrossRef]
- Gao, Y.; Nanan, R.; Macia, L.; Tan, J.; Sominsky, L.; Quinn, T.P.; O’Hely, M.; Ponsonby, Al.; Tang, M.L.K.; Collier, F.; et al. The maternal gut microbiome during pregnancy and offspring allergy and asthma. J Allergy Clin Immunol 2021, 148, 669–678. [Google Scholar] [CrossRef]
- Hirsch, A.G.; Pollak, J.; Glass, T.A.; Poulsen, M.N.; Bailey-Davis, L.; Mowery, J.; Schwartz, B.S. Early Life Antibiotic Use and Subsequent Diagnosis of Food Allergy and Allergic Diseases. Clin Exp Allergy 2017, 47, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, G.; Carta, M.; Montante, C.; Notarbartolo, V.; Corsello, G; Giuffrè, M. Current Insights on Early Life Nutrition and Prevention of Allergy. Front Pediatr 2020, 8, 448. [Google Scholar] [CrossRef] [PubMed]
- Akkerman, R.; Faas, M.M.; de Vos, P. Non-digestible carbohydrates in infant formula as substitution for human milk oligosaccharide functions: Effects on microbiota and gut maturation. Crit Rev Food Sci Nutr 2019, 59, 1486–1497. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Fernández, L.; Verhasselt, V. The Gut-Breast Axis: Programming Health for Life. Nutrients 2021, 13, 606. [Google Scholar] [CrossRef]
- Groer, M.W.; Morgan, K.H.; Louis-Jacques, A.; Miller, E.M. A scoping review of research on the human milk microbiome. J Hum Lact 2020, 36, 628–643. [Google Scholar] [CrossRef] [PubMed]
- Notarbartolo, V.; Giuffrè, M.; Montante, C.; Corsello, G.; Carta, M. Composition of Human Breast Milk Microbiota and Its Role in Children’s Health. Pediatr Gastroenterol Hepatol Nutr 2022, 25, 194–210. [Google Scholar] [CrossRef]
- Renz, H.; Skevaki, C. Early life microbial exposures and allergy risks: Opportunities for prevention. Nat Rev Immunol 2021, 21, 177–191. [Google Scholar] [CrossRef]
- Gabbianelli, R.; Bordoni, L.; Morano, S.; Calleja-Agius, J.; Lalor, J.G. Nutri-Epigenetics and Gut Microbiota: How Birth Care, Bonding and Breastfeeding Can Influence and Be Influenced? Int J Mol Sci 2020, 21, 5032. [Google Scholar] [CrossRef]
- Kers, J.G.; Saccenti, E. The Power of Microbiome Studies: Some Considerations on Which Alpha and Beta Metrics to Use and How to Report Results. Front Microbiol 2022, 12, 796025. [Google Scholar] [CrossRef]
- Finotello, F.; Mastrorilli, E.; Di Camillo, B. Measuring the diversity of the human microbiota with targeted next-generation sequencing. Brief Bioinform 2018, 19, 679–692. [Google Scholar] [CrossRef]
- Woo, V.; Alenghat, T. Epigenetic regulation by gut microbiota. Gut Microbes 2022, 14, 2022407. [Google Scholar] [CrossRef]
- Hong, X.; Wang, X. Epigenetics and Development of Food Allergy (FA) in Early Childhood. Curr Allergy Asthma Rep. 2014, 14, 460. [Google Scholar] [CrossRef]
- Di Costanzo, M.; De Paulis, N.; Capra, M.E.; Biasucci, G. Nutrition during Pregnancy and Lactation: Epigenetic Effects on Infants’ Immune System in Food Allergy. Nutrients 2022, 14, 1766. [Google Scholar] [CrossRef] [PubMed]
- Verduci, E.; Banderali, G.; Barberi, S.; Radaelli, G.; Lops, A.; Betti, F.; Riva, E.; Giovannini, M. Epigenetic Effects of Human Breast Milk. Nutrients 2014, 6, 1711–1724. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, F.P.; Loret de Mola, C.; Davies, N.M.; Victora, C.G.; Relton, C.L. Breastfeeding effects on DNA methylation in the offspring: A systematic literature review. PLoS ONE 2017, 12, e0173070. [Google Scholar] [CrossRef]
- Van Esch, B.C.A.M.; Porbahaie, M.; Abbring, S.; Garssen, J.; Potaczek, D.P.; Savelkoul, H.F.J.; Joost van Neerven, R.J. The Impact of Milk and Its Components on Epigenetic Programming of Immune Function in Early Life and Beyond: Implications for Allergy and Asthma. Front Immunol 2020, 11, 2141. [Google Scholar] [CrossRef]
- Wang, M.; Yang, I.V.; Davidson, E.J.; Joetham, A.; Takeda, K.; O’Connor, B.P.; Gelfand, E.W. Forkhead Box Protein 3 (FoxP3) Demethylation is Associated with Tolerance Induction in Peanut-Induced Intestinal allergy. J Allergy Clin Immunol 2018, 141, 659–670. [Google Scholar] [CrossRef]
- Briollais, L.; Rustand, D.; Allard, C.; Wu, Y.; Xu, J.; Rajan, S.G.; Hivert, MF.; Doyon, M.; Bouchard, L.; O McGowan, P.; et al. DNA methylation mediates the association between breastfeeding and early-life growth trajectories. Clin Epigenetics 2021, 13, 231. [Google Scholar] [CrossRef]
- Barker, D.J.P. Sir Richard Doll Lecture. Developmental origins of chronic disease. Public Health 2012, 126, 185–189. [Google Scholar] [CrossRef]
- Di Costanzo, M.; De Paulis, N.; Biasucci, G. Butyrate: A Link between Early Life Nutrition and Gut Microbiome in the Development of Food Allergy. Life (Basel) 2021, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Guttman, D.S.; Fiedl, C.J.; Sears, M.R.; HayGlass, K.T.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Becker, A.B.; et al. Infant gut microbiota and food sensitization: Associations in the first year of life. Clin Exp Allergy 2015, 45, 632–643. [Google Scholar] [CrossRef]
- Roduit, C.; Frei, R.; Ferstl, R.; Loeliger, S.; Westermann, P.; Ehyner, C.; Schiavi, E.; Barcik, W.; Rodriguez-Perez, N.; Wawrzyniak, M.; et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 2019, 74, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Kim, H. Update on Early Nutrition and Food Allergy in Children. Yonsei Med J 2016, 57, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Junge, K.M.; Bauer, T.; Geissler, S.; Hirche, F.; Thürmann, L.; Bauer, M.; Trump, S.; Bieg, M.; Weichenhan, D.; Gu, L.; et al. Increased vitamin D levels at birth and in early infancy increase offspring allergy risk-evidence for involvement of epigenetic mechanisms. J Allergy Clin Immunol. 2016, 137, 610–613. [Google Scholar] [CrossRef] [PubMed]
- Weisse, K.; Winkler, S.; Hirche, F.; Herberth, G.; Hinz, D.; Bauer, M.; Rӧder, S.; Rolle-Kampczyk, U.; von Bergern, M.; Olek, S.; et al. Maternal and newborn vitamin D status and its impact on food allergy development in the German LINA cohort study. Allergy 2013, 68, 220–228. [Google Scholar] [CrossRef]
- Venter, C.; Agostoni, C.; Arshad, S.H.; Ben-Abdallah, M.; Du Toit, G.; Fleischer, D.M.; Greenhawt, M.; Glueck, D.H.; Groetch, M.; Lunjani, N.; et al. Dietary factors during pregnancy and atopic outcomes in childhood: A systematic review from the European Academy of Allergy and Clinical Immunology. Pediatr Allergy Immunol 2020, 31, 889–912. [Google Scholar] [CrossRef]
- Järvinen, K.M.; Martin, H.; Oyoshi, M.K. Immunomodulatory Effects of Breast Milk on Food Allergy. Ann Allergy Asthma Immunol. 2019, 123, 133–143. [Google Scholar] [CrossRef]
- Best, K.P.; Gold, M.; Kennedy, D.; Martin, J.; Makrides, M. Omega-3 long-chain PUFA intake during pregnancy and allergic disease outcomes in the offspring: A systematic review and meta-analysis of observational studies and randomized controlled trials. Am J Clin Nutr 2016, 103, 128–143. [Google Scholar] [CrossRef]
- Venter, C.; Brown, K.R.; Maslin, K.; Palmer, D.J. Maternal dietary intake in pregnancy and lactation and allergic disease outcomes in offspring. Pediatr Allergy Immunol 2017, 28, 135–143. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, R.; Li, X.; Gao, Y.; Dai, N.; We, Y.; Liu, L.; Xing, Y.; Li, Z. Relationship between maternal-infant gut microbiota and infant food allergy. Front Microbiol 2022, 13, 933152. [Google Scholar] [CrossRef]
- Tuokkola, J.; Luukkainen, P.; Tapanainen, H.; Kaila, M.; Vaarala, O.; Kenward, M.G.; Virta, L.J.; Veijola, R.; Simell, O.; Ilonen, J.; et al. Maternal diet during pregnancy and lactation and cow’s milk allergy in offspring. Eur J Clin Nutr 2016, 70, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Vuillermin, P.J.; Macia, L.; Nanan, R.; Lk Tang, M.; Collier, F.; Brix, S. The maternal microbiome during pregnancy and allergic disease in the offspring. Semin Immunopathol 2017, 39, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Venter, C.; Palumbo, M.P.; Glueck, D.H.; Sauder, K.A.; O’Mahony, L.; Fleischer, D.M.; Ben-Abdallah, M.; Ringham, B.M.; Dabelea, D. The maternal diet index in pregnancy is associated with offspring allergic diseases: The Healthy Start study. Allergy 2022, 77, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Al Khatib, H.; et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat Med. 2021, 27, 321–332. [Google Scholar] [CrossRef]
- Shen, X.; Wang, M.; Zhang, X.; He, M.; Li, M.; Cheng, G.; Wan, C.; He, F. Dynamic construction of gut microbiota may influence allergic diseases of infants in Southwest China. BMC Microbiol. 2019, 19, 123. [Google Scholar] [CrossRef]
- Méndez, C.S.; Bueno, S.M.; Kalergis, A.M. Contribution of Gut Microbiota to Immune Tolerance in Infants. J immunol Res. 2021, 2021, 7823316. [Google Scholar] [CrossRef]
- Ling, Z.; Li, Z.; Liu, X.; Cheng, Y.; Luo, Y.; Tong, X.; Yuan, L.; Wang, Y.; Sun, J.; Li, L.; et al. Altered fecal microbiota composition for food allergy in infants. Appl Environ Microbiol 2014, 80, 2546–2554. [Google Scholar] [CrossRef]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Bjorksten, B.; Engstrand, L.; Jenmalm, M.C. Low diversity of the gut microbiota in infants with atopic eczema. J Allergy Clin.Immunol. 2012, 129, 434–440. [Google Scholar] [CrossRef]
- Kielleniva, K.; Ainonen, S.; Vänni, P.; Paalanne, N.; Renko, M.; Salo, J.; Tejesvi, M.V.; Pokka, T.; Pirttilä, A.M.; Tapiainen, T. Microbiota of the first-pass meconium and subsequent atopic and allergic disorders in children. Clin Exp Allergy 2022, 52, 684–696. [Google Scholar] [CrossRef]
- Shu, S.A.; Yuen, A.W.T.; Woo, E.; Chu, K.H.; Kwan, H.S.; Yang, GX.; Yang, Y.; Leung, P.S.C. Microbiota and Food Allergy. Clin Rev Allergy Immunol 2019, 57, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Leyva, L.L.; Brereton, N.J.B.; Koski, K.G. Emerging frontiers in human milk microbiome research and suggested primers for 16S rRNA gene analysis. Comput Struct Biotechnol J 2020, 19, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Moubareck, C.A. Human Milk Microbiota and Oligosaccharides: A Glimpse into Benefits, Diversity, and Correlations. Nutrients 2021, 13, 1123. [Google Scholar] [CrossRef] [PubMed]
- Demmelmair, H.; Jiménez, E.; Collado, M.C.; Salminen, S.; McGuire, M.K. Maternal and Perinatal Factors Associated with the Human Milk Microbiome. Curr Dev Nutr 2020, 4, nzaa027. [Google Scholar] [CrossRef] [PubMed]
- Verduci, E.; Giannì, M.L.; Vizzari, G.; Vizzuso, S.; Cesarani, J.; Mosca, F.; Zuccotti, G.V. The Triad Mother-Breast Milk-Infant as Predictor of Future Health: A Narrative Review. Nutrients 2021, 13, 486. [Google Scholar] [CrossRef]
- Wang, S.; Wei, Y.; Liu, L.; Li, Z. Association Between Breastmilk Microbiota and Food Allergy in Infants. Front Cell Infect Microbiol 2022, 11, 770913. [Google Scholar] [CrossRef]
- Rey-Mariňo, A.; Pilar Francino, M. Nutrition, Gut Microbiota, and Allergy Development in Infants. Nutrients 2022, 14, 4316. [Google Scholar] [CrossRef]
- Dai, D.L.Y.; Petersen, C.; Hoskinson, C.; Del Bel, K.L.; Becker, A.B.; Moraes, T.J.; Mandhane, P.J.; Finlay, B.B.; Simons, E.; Kozyrskyj, A.L.; et al. Breastfeeding enrichment of B. longum subsp. infantis mitigates the effect of antibiotics on the microbiota and childhood asthma risk. Med 2023, 4, 92–112. [Google Scholar] [CrossRef]
- Charbonneau, M.R.; O’Donnell, D.; Blanton, L.V.; Totten, S.M.; Davis, J.C.C.; Barratt, M.J.; Cheng, J.; Guruge, J.; Talcott, M.; Bain, J.R.; et al. Sialylated Milk Oligosaccharides Promote Microbiota-Dependent Growth in Models of Infant Undernutrion. Cell 2016, 164, 859–871. [Google Scholar] [CrossRef]
- Yokanovich, L.T.; Newberry, R.D.; Knoop, K.A. Regulation of oral antigen delivery early in life: Implications for oral tolerance and food allergy. Clin Exp Allergy 2021, 51, 518–526. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol Int 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Laursen, M.F. Gut Microbiota Development: Influence of Diet from Infancy to Toddlerhood. Ann Nutr Metab 2021, 30, 1–14. [Google Scholar] [CrossRef]
- Di Profio, E.; Magenes, V.C.; Fiore, G.; Agostinelli, M.; La Mendola, A.; Acunzo, M.; Francavilla, R.; Indrio, F.; Bosetti, A.; D’Auria, E.; et al. Special Diets in Infants and Children and Impact on Gut Microbioma. Nutrients 2022, 14, 3198. [Google Scholar] [CrossRef]
- Differding, M.K.; Benjamin-Neelon, S.E.; Hoyo, C.; Østbye, T.; Mueller, N.T. Timing of complementary feeding is associated with gut microbiota diversity and composition and short chain fatty acid concentrations over the first year of life. BMC Microbiol 2020, 20, 56. [Google Scholar] [CrossRef] [PubMed]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Alvarez-Quintero, R.; Velasquez-Mejia, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher Fecal Short-Chain Fatty Acid Levels Are Associated with Gut Microbiome Dysbiosis, Obesity, Hypertension and Cardiometabolic Disease Risk Factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef]
- Kim, K.N.; Yao, Y.; Ju, S.Y. Short Chain Fatty Acids and Fecal Microbiota Abundance in Humans with Obesity: A Systematic Review and MetaAnalysis. Nutrients 2019, 11, 2512. [Google Scholar] [CrossRef]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association Between Breast Milk Bacterial Communities and Establishment and Development of the Infant Gut Microbiome. JAMA Pediatr 2017, 171, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.L.; Monteagudo-Mera, A.; Cadenas, M.B.; Lampl, M.L.; Azcarate-Peril, M.A. Milk- and solid-feeding practices and daycare attendance are associated with differences in bacterial diversity, predominant communities, and metabolic and immune function of the infant gut microbiome. Front Cell Infect Microbiol. 2015, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Comberiati, P.; Costagliola, G.; D’Elios, S.; Peroni, D. Prevention of Food Allergy: The Significance of Early Introduction. Medicina (Kaunas) 2019, 55, 323. [Google Scholar] [CrossRef]
- Du Toit, G.; Roberts, G.; Sayre, P.H.; Bahnson, H.T.; Radulovic, S.; Santos, A.F.; Brough, H.A.; Phippard, D.; Basting, M.; Feeney, M.; et al. Randomized trial of peanut consumption in infants at risk for peanut allergy. N Engl J Med 2015, 372, 803–813. [Google Scholar] [CrossRef]
- Grimshaw, K.E.C.; Maskell, J.; Oliver, E.M.; Morris, R.C.G.; Foote, K.D.; Clare Mills, E.N.; Roberts, G.; Margetts, B.M. Introduction of complementary foods and the relationship to food allergy. Pediatrics 2013, 132, e1529–e1538. [Google Scholar] [CrossRef] [PubMed]
- Leong, C.; Haszard, J.J.; Lawley, B.; Otal, A.; Taylor, R.W.; Szymlek-Gay, E.A.; Fleming, E.A.; Daniels, L.; Fangupo, L.J.; Tannock, G.W.; et al. Mediation Analysis as a Means of Identifying Dietary Components That Differentially Affect The Fecal Microbiota of Infants Weaned by Modified Baby-Led and Traditional Approaches. Appl Environ Microbiol. 2018, 84, e00914–e00918. [Google Scholar] [CrossRef] [PubMed]
- Fazlollahi, M.; Chun, Y.; Grishin, A.; Wood, R.A.; Burks, A.W.; Dawson, P.; Jones, S.M.; Leung, D.Y.M.; Sampson, H.A.; Sicherer, S.H.; et al. Early-life gut microbiome and egg allergy. Allergy 2018, 73, 1515–1524. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, L.; Wang, J.; Hao, M.; Che, H. Antibiotic-Induced Gut Microbiota Dysbiosis Damages the Intestinal Barrier, Increasing Food Allergy in Adult Mice. Nutrients 2021, 13, 3315. [Google Scholar] [CrossRef]
- Nance, C.L.; Deniskin, R.; Diaz, V.C.; Paul, M.; Anvari, S.; Anagnostou, A. The Role of the Microbiome in Food Allergy: A Review. Children (Basel) 2020, 7, 50. [Google Scholar] [CrossRef]
Figure 1.
Main factors influencing children’s gut microbiota composition in the first 1000 days of life.
Figure 1.
Main factors influencing children’s gut microbiota composition in the first 1000 days of life.
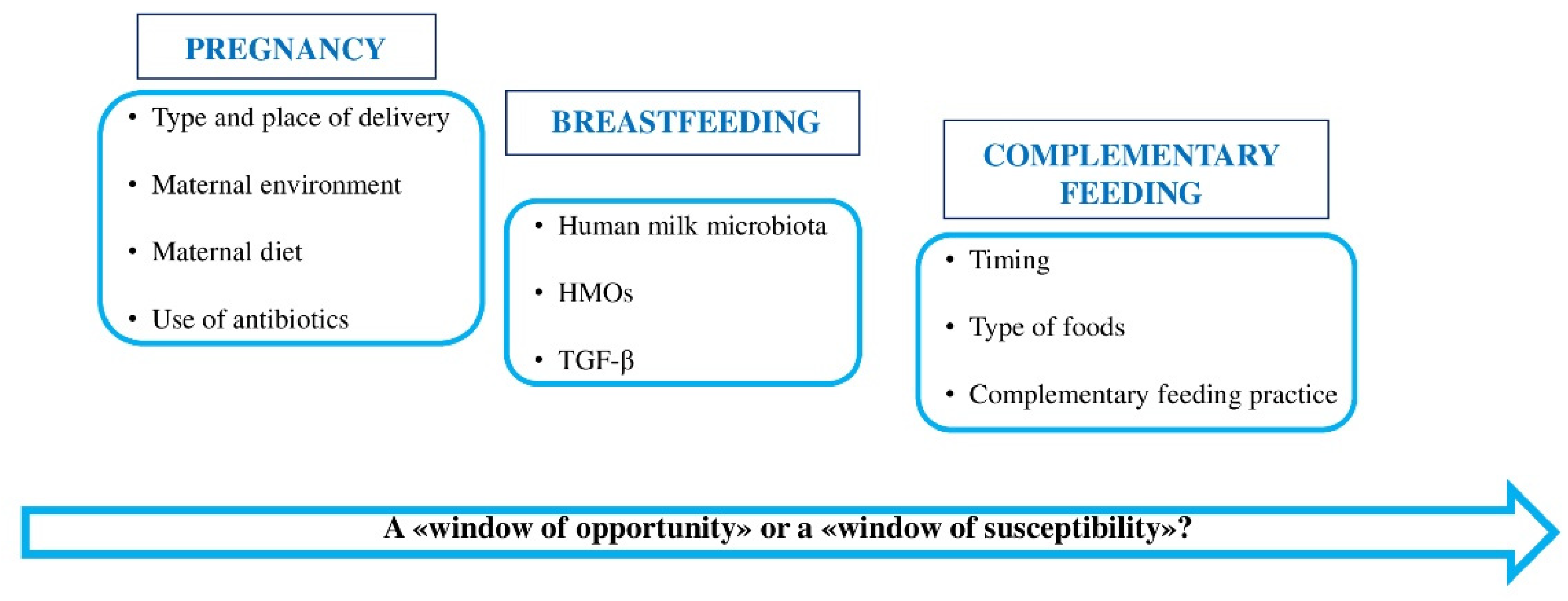
Figure 2.
Protective and unprotective bacterial genus in children’s gut microbiota.
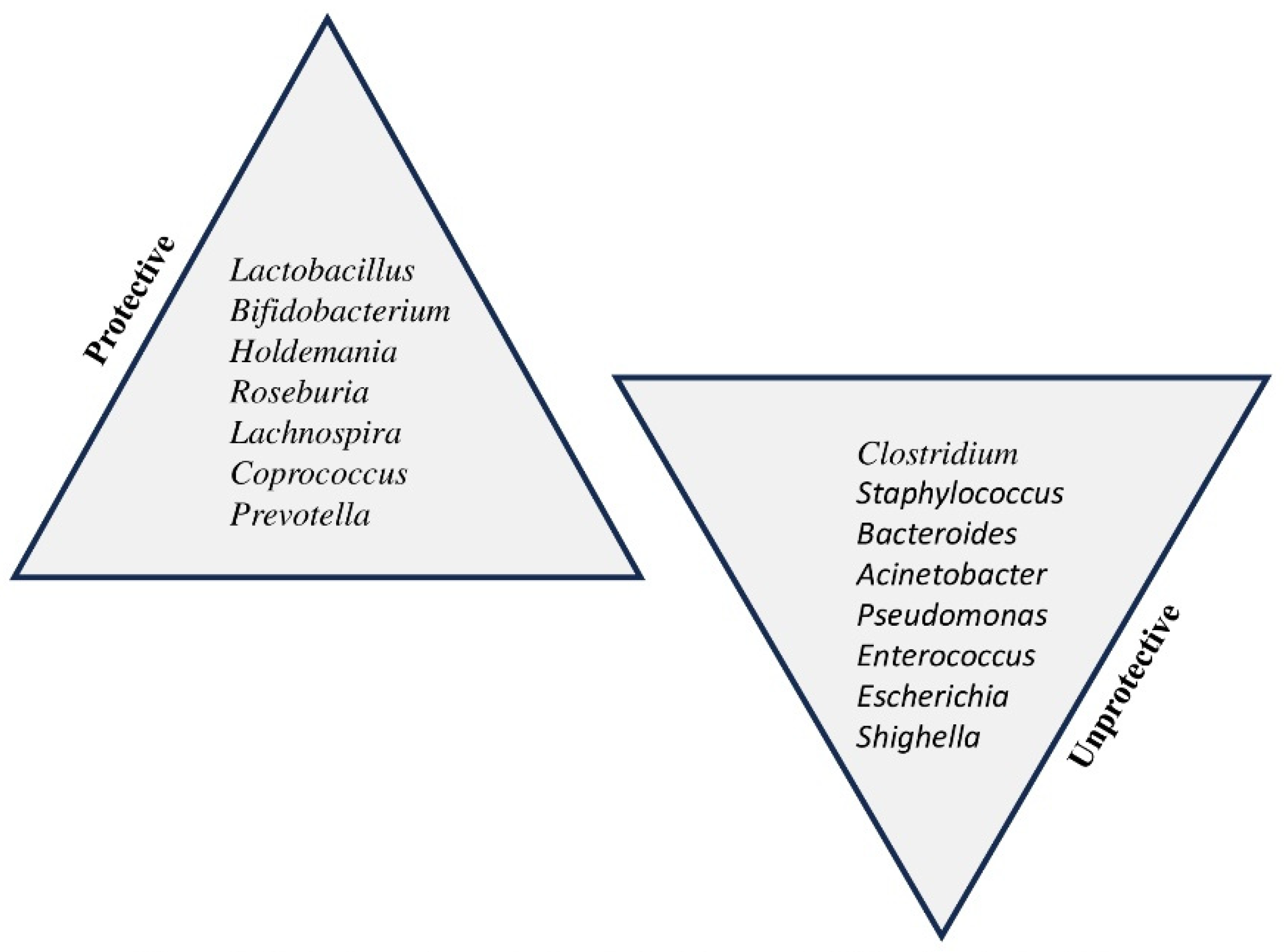

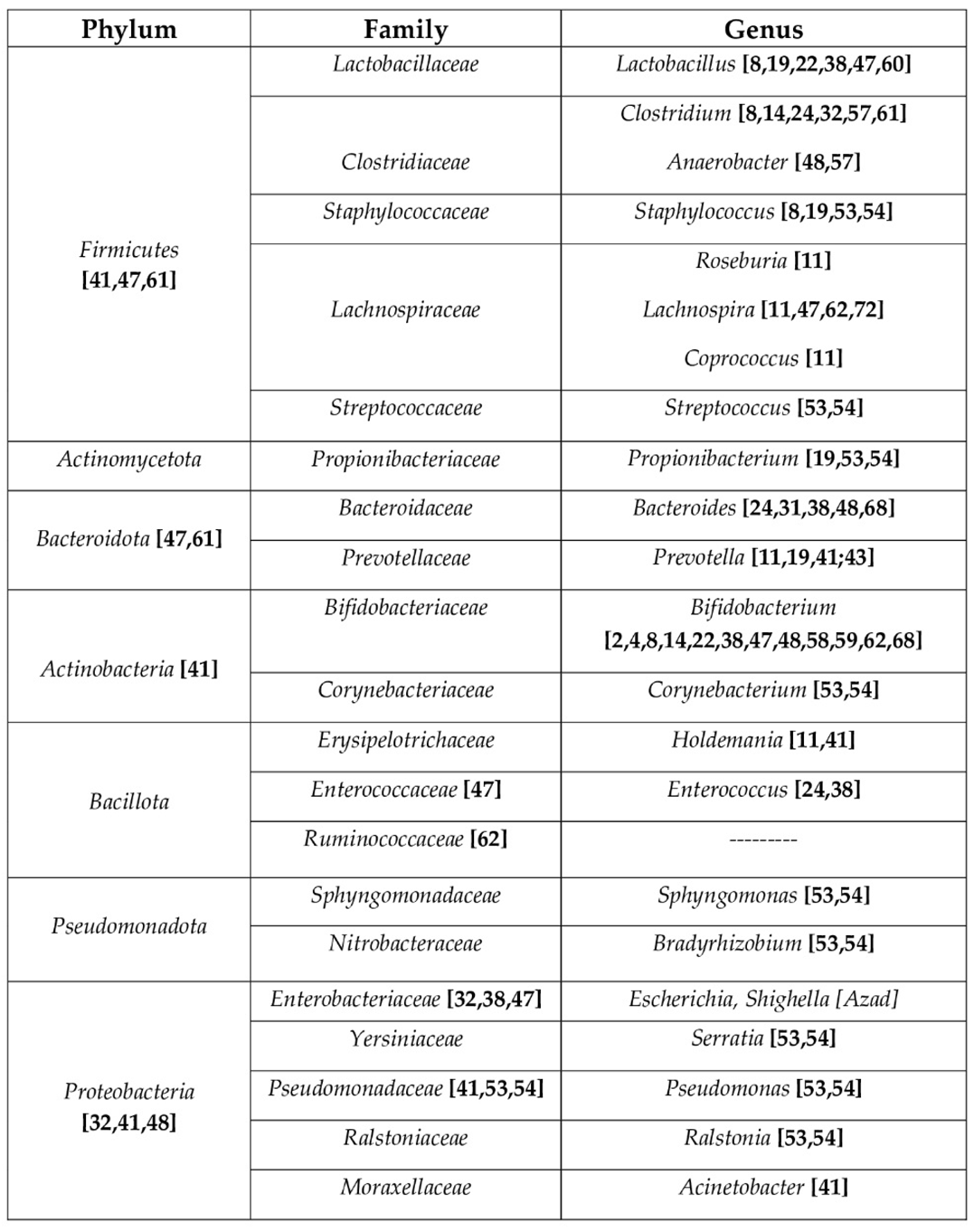
Table 1.
taxonomic classification of the main bacterial phyla, families and genus involved in allergic diseases.
Table 1.
taxonomic classification of the main bacterial phyla, families and genus involved in allergic diseases.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



