
Submitted:
07 August 2023
Posted:
08 August 2023
You are already at the latest version
Abstract
Aim: Ectopic lipid accumulation, including pancreatic steatosis, exacerbates type 2 diabetes risk in susceptible individuals. Dysregulated circulating microRNAs (miRNAs) have been identified to correlate with clinical measures of pancreatitis, pancreatic cancer, and type 1 diabetes. The aim of the current study was, therefore, to examine the association between circulating abundances of candidate miRNAs and pancreatic and liver fat deposition as quantified using magnetic resonance imaging (MRI) and spectroscopy (MRS).
Methods: Asian Chinese (n=34; BMI=26.7±4.2 kg/m2) and European Caucasian (n=34; BMI=28.0±4.5 kg/m2) females from the TOFI_Asia cohort underwent MRI and MRS analysis of pancreas (MR-%pancreas) and liver (MR-%liver fat) respectively to quantify ectopic lipid deposition. Plasma miRNA abundances of a subset of circulatory miRNAs associated with pancreatic and liver steatosis were quantified by qRT-PCR.
Results: miR-21-3p and miR-320a-5p correlated with MR-%pancreas fat, plasma insulin and HOMA2-IR but not MR-%liver fat. MR-%pancreas fat remained associated with decreasing miR-21-3p abundance following multivariate regression analysis.
Conclusions: miR-21-3p and miR-320a were demonstrated to be negatively correlated with MR-%pancreas fat, independent of ethnicity. For miR-21-3p, this relationship persists with the inclusion of MR-%liver fat in the model, suggesting the potential for wider application as a specific circulatory correlate of pancreatic steatosis.
Keywords:
miRNA
; miR-21-3p
; miR-320a-5p
; ectopic
; steatosis
; biomarker
; pancreas
; liver
1. Introduction
Ectopic lipid infiltration and accumulation within the visceral organs are key underlying factors for a heightened risk of metabolic disease [1]. Collectively, the actions of pancreatic steatosis and non-alcoholic fatty liver disease (NAFLD) are major determinants of cardiometabolic disease (CVD) [2]. Ectopic lipid accumulation in the pancreas and liver is particularly notable as it has been demonstrated to be a key differentiator for the variation in type 2 diabetes mellitus (T2DM) and CVD risk across differing ethnicities [3].
Whilst considerable attention is directed towards the impact of NAFLD and its relationship to metabolic disease risk [2,4], the importance of pancreatic steatosis as a causal mechanism implicated in pancreatic β-cell dysfunction and apoptosis has also been established [5]. These impairments in pancreatic function are hallmark features for the loss of insulin secretory function present in T2DM [6]. Detailed analysis of pancreatic steatosis and its relationship to metabolic disease risk has been an ongoing challenge due to the need to accurately quantify lipid accumulation within this small organ located deep in the retroperitoneal abdominal region [7]. Recently described magnetic resonance imaging (MRI) and spectroscopy (MRS) methods are yielding advances in the precision of quantification [8,9]. However, these techniques remain technically challenging, precluding regular application clinically and experimentally [10]. Hence, there is a need for suitable informative proxy measures or biomarkers that can be used for preliminary screening or potentially to provide additional insight into the pathogenesis of pancreatic steatosis.
MicroRNAs (miRNAs) are integral molecular negative regulators of transcriptional processes, with evidence that dysregulated circulatory miRNA abundances contribute to pathophysiological processes associated with the onset of metabolic diseases such as pancreatitis, insulin resistance (IR), liver and pancreatic steatosis [11,12]. miRNA expression is reported to be tissue specific although there is evidence for bidirectional crosstalk between metabolically active organs such as pancreas and liver [13,14]. For example, circulatory miR-375 [15], an islet-cell enriched miRNA and a critical regulator of β-cell function [16], has been reported to be involved in the development of non-alcoholic liver steatosis [17] . Similarly, miR-122 [18,19], a liver specific miRNA, has been shown to exhibit significant differences in the blood of severe acute pancreatitis patients compared to healthy controls [20]. Whilst, such crosstalk exists between several miRNAs, it is expected that the identification of potential-specific biomarkers of miRNAs may help predict or detect the development and progression of steatosis in these organs at an early stage, and therefore allow timely intervention. However, it remains yet to be known if any correlation exists between the abundance of these circulatory miRNAs and the degree of pancreas and liver steatosis as quantified using MRI/MRS techniques.
The aim of the current study was, therefore, to examine in a cohort of women the correlation between the abundances of a subset of circulatory miRNAs that have previously been identified to be associated with pancreatic and lipid steatosis with MRI and MRS-determined pancreatic (MR-%pancreatic) and liver fat (MR-%liver) . We analysed the circulatory miRNA levels in plasma of a cohort of women from the previously described TOFI (Thin on the Outside, Fat on the Inside) cohort consisting of Asian Chinese and European Caucasian ethnicities, lean and overweight, normoglycaemic and prediabetic [21].
2. Methods
2.1. Study Design
The plasma samples utilised in this study were archived from the previously conducted TOFI cohort study [21], with ethical approval inclusive of these secondary analyses from the Health and Disabilities Ethics Committee (HDEC) Auckland, New Zealand (16/STH/23). The study was conducted according to the Declaration of Helsinki guidelines. The study is registered with the Australian New Zealand Clinical Trials Registry at anzctr.org.au (ACTRN12616000362493).
2.2. Study Population
2.3. Sample Collection
Fasting blood samples were collected from each participant in EDTA-coated tubes and immediately centrifuged at 1,300 × g for 10min at 4 °C for plasma separation. The resultant plasma was aliquoted and stored at −80 °C until further analysis.
2.4. Anthropometric and Biochemical Analysis
Height, weight, waist circumference and blood pressure were measured at fasting, as previously reported [23]. Body mass index (BMI) cut points were determined as Asian Chinese; overweight ≥24 kg/m2, obese ≥28kg/m2; European Caucasian: overweight ≥25kg/m2, obese ≥30kg/m2. Biochemical measures were assessed as described previously [21]. Among these measurements, glucose was analysed by the Hexokinase method, total plasma cholesterol and HDL cholesterol (HDL-C) measured using cholesterol-esterase/cholesterol-oxidase/peroxidase method, and low-density lipoprotein cholesterol (LDL-C) was calculated using the Friedwald formula [21]. Triglycerides (TAGs) were analysed by the lipase/ glycerol kinase method [21]. Plasma insulin (limit of detection (LOD): 87pg/ml) and C-peptide (LOD: 9.5pg/ml) were measured by multiplex immunoassay (MILLIPLEX®MAP Human Metabolic Hormone Magnetic Bead Panel, Germany). Glycosylated haemoglobin (HbA1c) was determined by capillary electrophoresis (CAP2FP, Sebia, France). Homeostasis model assessment of IR (HOMA2-IR) was determined online using the HOMA2 Calculator ©( The University of Oxford 2013, Version 2.2.3) [24].
Pancreatic and liver fat content quantification by magnetic resonance imaging (MRI) and spectroscopy (MRS)
Intra-pancreatic fat percentage was quantified using a 3T Magnetom Skyra scanner, VE 11A (Siemens, Germany), using a T1-weighted, 3D dual gradient-echo sequence (VIBE) 2-point Dixon method as previously described [25]. Two candidate pancreas (5 mm each) fat fraction (FF) maps with a visible head, body and tail were created using MRI images. To estimate MR-%pancreatic fat, three regions of interest (ROI) were placed in each image's head, body, and tail, respectively. Furthermore, a 1-20% threshold was applied to eliminate the potential inclusion of non-parenchymal tissue within the selected ROI. The average fat of both candidate pancreas FF images was utilised to calculate MR-%pancreas fat.
Liver fat content was measured using the MRS method as described previously [25]. Briefly, a 2×2×2 cm3 voxel was placed in the right lobe avoiding blood vessels and the biliary tree; spectra were obtained in transverse, coronal, and sagittal planes ± water suppression. Liver fat was expressed as % calculated vol/vol of fat and water.
2.5. Circulating Total RNA Extraction
Plasma (250μl) was used for total RNA (including miRNAs) isolation as described previously by D’Souza et al. [26].
2.6. cDNA Synthesis
A fixed volume of 2 μl of total RNA was used as input for each cDNA synthesis reaction using TaqMan™ Advanced miRNA cDNA Synthesis Kit (Catalogue number: A28007, Applied Biosystems) following the manufacturer's recommendations [27].
2.7. Circulating miRNA Real-Time Quantitative PCR (qPCR)
A total of ten [miR-375, miR-7, miR-361-5p and miR-15a-5p, miR-21-3p, miR-126-3p, miR-24-5p, miR-320a, miR-221, and miR-17-5p] custom human miRNA assays (TaqMan miRNA Assays, Applied Biosystems) were utilised. According to the manufacturer's instructions, a PCR reaction mixture of Master Mix and PCR-primer (TaqMan® Fast Advanced Master Mix and TaqMan miRNA Assays) was prepared, and amplification was performed on a Quant Studio™ 6 Flex Real-Time PCR System (Applied Biosystems) using cycling parameters recommended by Applied Biosystems. Samples with CT >35 were excluded from the analysis.
Two technical replicates were used for each sample. Haemolysis of samples was monitored by comparing miR-23a-3p expression, reported to be unaffected by haemolysis, with miR-451a, a highly expressed miRNA in red blood cells, ΔCt (miR-23a-3p-miR-451a), with a ΔCt of > 7 indicates a high risk of haemolysis [28]. For quality control of cDNA synthesis, an exogenous spike-in (cel-miR-238) was spiked in all samples. miRNA expression data were normalized using a geometric mean of two endogenous miRNAs miR-423-5p (Asian Chinese; Ct=22.14±2.15, European Caucasian; Ct=21.89±2.58 (p=0.66)) and miR-191-5p (Asian Chinese; Ct=24.90±1.91, European Caucasian; Ct=23.87±2.30 (p=0.11)) along with an exogenous spike-in cel-miR-238 (Asian Chinese; Ct= 20.95±0.20, European Caucasian; Ct= 20.48±0.25 (p=0.13)) was performed. The abundance of miRNAs was measured using the 2 (-ΔCt) method [29].
2.8. Statistical Analysis
Data are expressed as mean ± standard deviation (SD). Pearson's correlation coefficients assessed the association between circulatory miRNA expression, MR-%pancreas and MR-%liver fat. miRNAs that showed a significant correlation (p≤ 0.05) with MR-%pancreas fat were further used to construct a multivariate linear regression model. Regression models were adjusted for ethnicity and biochemical and physiological parameters, including BMI, fasting plasma glucose (FPG) and plasma insulin. The standard error (SE) was used to estimate the intercept and slope of linear regression equations, and analysis of variance (ANOVA) was used to demonstrate the significance of the whole model with statistical significance set at p≤ 0.05. All data were analysed using SPSS 25.0 (SPSS Inc., Chicago, IL), and all graphs were prepared using GraphPad Prism-7 (GraphPad Software, CA).
3. Results
3.1. Characteristics of the Study Population
The clinical and demographic characteristics of the study participants are detailed in Table 1.
Using ethnic specific cut-points, 18 women were classified as lean (Asian Chinese (n=10); European Caucasian (n=8)), 26 as overweight, (Asian Chinese (n=11); European Caucasian (n=15)) and 24 obese (Asian Chinese (n=13); European Caucasian (n=11)). Thirteen participants were classified with pre-diabetes (Asian Chinese (n=6); European Caucasian (n=7)) based on ADA-defined FPG concentrations [22]..
3.2. Expression of Circulating miRNAs Correlates with MR-%pancreas Fat But Not with MR-%liver Fat
Two of the ten targeted circulatory miRNAs exhibited significant inverse correlations with MR-%pancreatic fat (miR-21-3p; r= -0.26, p=0.04 and miR-320a-5p; r=-0.28, p=0.03) (Figure 1).
Correlation plot comparing the relative abundance of miR-21-3p (b) and miR-320a-3p to pancreatic fat percentage, r = 'Pearson's coefficient with exact p value listed. Each point represents an individual sample
A multivariate linear regression model for miR-21-3p and miR-320a was constructed to establish the association between these miRNAs and MR-%pancreatic fat adjusted for BMI, FPG, plasma insulin and ethnicity (Table 2). The regression model significantly explained the variance in MR-%-pancreatic fat in relation to miRNA expression (p < 0.01, R2= 0.43, adj. R2 =0.37)
To ascertain that the abundance of these miRNAs was specific to MR-%pancreatic fat, a multivariate linear regression model for miR-21-3p and miR-320a in relation to MR-%liver fat was constructed. No significant relationship was observed between these miRNA abundances and MR-%liver fat (Table 3).
3.3. Expression of Circulating miRNAs Correlates with HOMA2-IR, HbA1c and Fasting Plasma Insulin
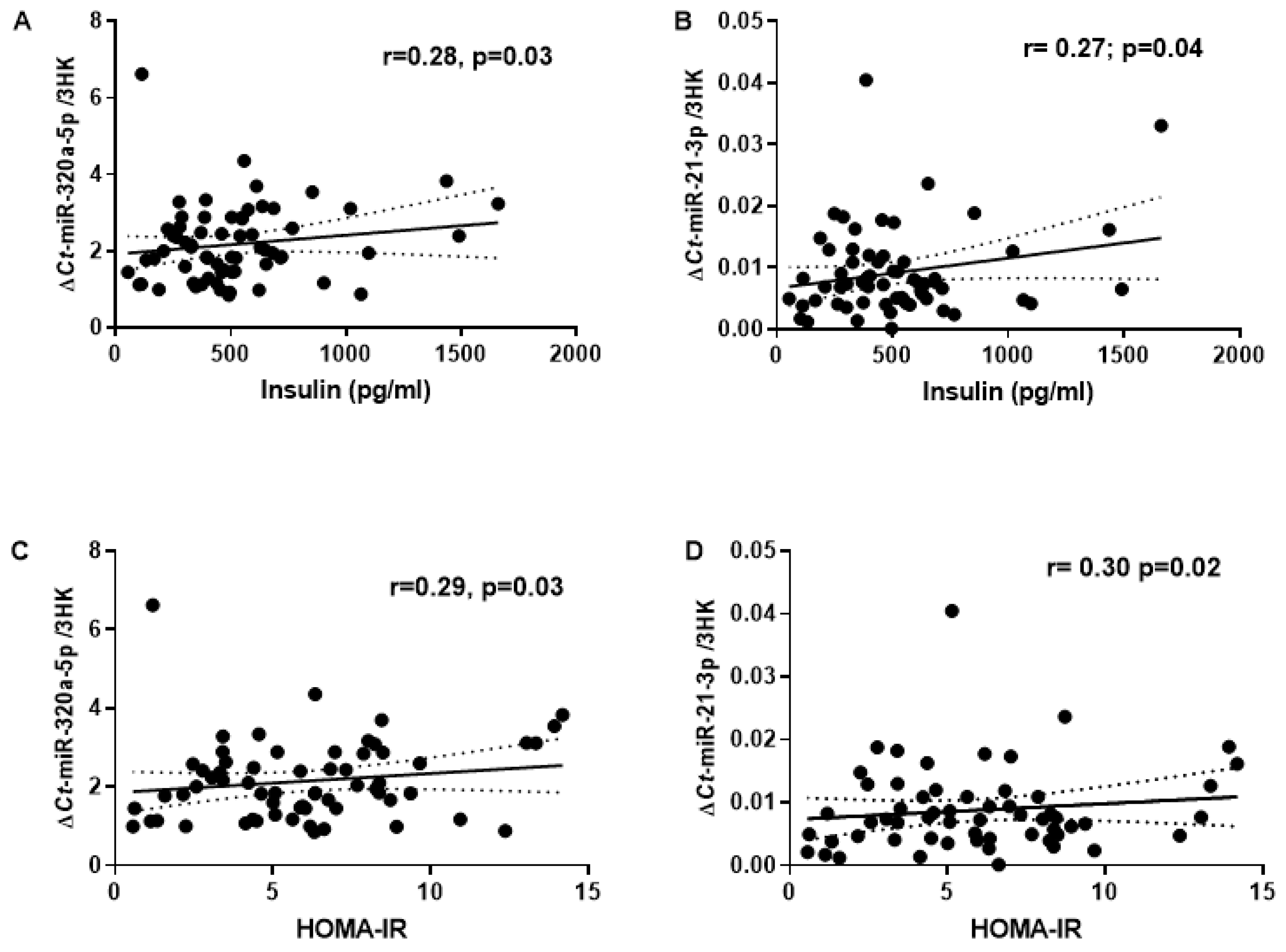
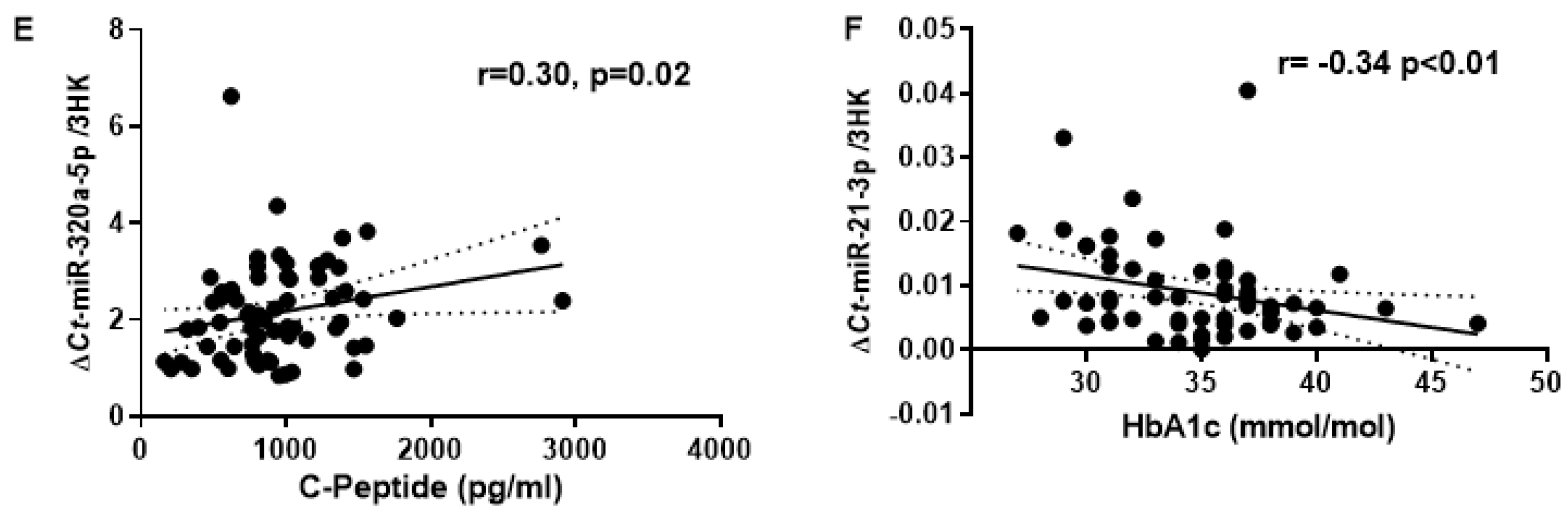
Additionally, biomarkers of T2DM risk showed a significant correlation with miR-21-3p and miR-320a (Figure 2). Expression of miR-21-3p and miR-320a significantly correlated with fasting plasma insulin (r=0.27, p=0.04 and r=0.28, p=0.03; respectively) and HOMA-IR (r= 0.30, p=0.02 and r=0.29, p=0.03; respectively). Furthermore, miR-320a positively correlated with C-peptide (r=0.30, p=0.02) while miR-21-3p negatively correlated with HbA1c (r=-0.34, p<0.01). No significant correlation was observed for FPG and circulatory triglycerides with MR-%pancreas fat.
4. Discussion
The circulating abundances of ten candidate miRNAs implicated in the regulation of insulin production and pancreatitis risk were analysed in relation to MRI-determined pancreas % fat and MRS-determined liver % fat. Of these miRNAs, miR-21-3p and miR-320a exhibited a negative correlation, specifically for MR-%pancreas fat. Further, the abundances of these miRNAs correlated with biomarkers of metabolic disease, which includes circulatory C-peptide, HOMA2-IR, plasma insulin and HbA1c. Moreover, in a multivariate linear regression model involving BMI, FPG and plasma insulin as covariates, miR-21-3p showed a significant negative association with pancreatic steatosis irrespective of ethnicity.
In recent years, studies have shown that compared to the liver, the pancreas might be more sensitive to fat infiltration [30]. However, there is no globally comparable cut-off for increased %pancreas fat, with evidence suggesting arbitrary values between 4.5%-6.5% in people with mild to morbid obesity [30]. Based on the previous literature, this study used an arbitrary value of 4.5% as a cut-off for %pancreas fat [30]. This threshold identified 39% of our cohort (n=25; mean age: 49±12y; BMI: 28.7±4.4 kg/m2) as above the cut point. Interestingly, an ethnicity-specific relationship was observed between pancreas fat and BMI; Chinese women at a fixed BMI of 20-25 kg/m2 had ~1% higher predicted pancreas fat than their Caucasian counterparts [21]. For liver fat, where a cut-off of ≥5.6% can be considered elevated [31], the mean %liver fat measured across the full cohort of 68 women was below this threshold (mean: 4.2±0.8%, Table 1), with 22% (n=15; mean age: 50.1±13.0 y; BMI: 28.9±3.6 kg/m2) of women were identified with elevated liver fat [21]. Although the average of both %pancreas fat and liver fat were lower than the adopted thresholds, this study observed a significant correlation between %-pancreatic fat and circulatory miRNAs, possibly demonstrating that even small changes in the circulatory miRNAs would likely reflect the early stages of steatosis.
Upregulated expression of miR-21-3p is reported to be a general feature of tissue inflammation and fibrosis [32]. miR-21-3p is shown to have pro-adipogenic characteristics [33], with an upregulated expression in subcutaneous adipose tissues of obese human and animal models [34]. Interestingly, under normal physiological conditions, miR-21-3p is highly expressed in the hepatocytes of mice, although minimally active [35]. However, upon challenge with a high-fat diet in mice, miR-21-3p promotes metabolic derangements, including glucose intolerance, IR and steatosis [36]. Furthermore, in rat models of pancreatitis, an upregulation of miR-21-3p expression is reported to aggravate the disease and promote pancreatic injury by activating the transient receptor potential (TRP) signalling pathway [32]. In contrast, the current study observed a negative association between the measured circulatory abundance of miR-21-3p with that of MR-%pancreas fat. Although understanding the mechanism behind this discrepancy is out of the scope for this study, we assume the possible reason involved could be due to different species and/or different techniques used to quantify the abundance of miRNAs in this current study.
Although little is known about the role of miR-320a and pancreatic steatosis, altered expression of miR-320a has been observed in pancreatic cancer, IR, and pancreatic fibrosis [37,38].The observed correlation between miR-320a and pancreatic steatosis in this study might indicate a wide range of pathophysiological events within the pancreas and not specifically the measured steatosis. Clearly, mechanistic analysis is required to confirm the function of these miRNAs in the complex regulation of pancreatic function.
Variation in the deposition of pancreatic fat has been reported in different ethnicities [39], with additional evidence for ethnicity impacting the expression patterns of miRNAs [40]. Therefore, to elucidate the effect of different ethnicities of our study participants on the expression of miR-21-3p and miR-320a, a partial correlation of these miRNAs with MR-%pancreas fat while controlling for ethnicity was performed. Interestingly, the expression of these miRNAs still showed an inverse correlation with the MR-%pancreas fat, thus suggesting that miR-21-3p and miR-320a are associated with pancreatic fat independent of ethnicity.
There are some limitations to the findings of this study. There has been evidence of differences in the physiological processes of the pancreas based on the sex of an individual. For example, while analysing the secretin-induced exocrine pancreatic response in healthy men and women above and below the age of 45, it was observed that women over the age limit, compared to the males, secreted significantly less water and bicarbonate [41]. Moreover, the authors also observed age and sex-dependent variations for lipase secretion, with a decreased secretion in women per mg of pancreatic tissue [41]. There is no consensus around sex-specific miRNA expression and pancreatic steatosis. The present study was undertaken in females only and further studies examining potential sex-specificity in circulatory miRNA profiles and associations with pancreatic steatosis are required.. In addition, plasma miRNA profiling provides limited insights into tissue-specific exosomal miRNA expression as it cannot conclusively identify the tissue source of these miRNAs[42]. Therefore, further elucidation of the clinical implications of the altered miRNA abundances on pancreatic steatosis and other metabolically active organs is required to ascertain systemic effects.
This study showed a correlation between MR-%pancreas fat and the abundance of circulating miRNAs. Irrespective of ethnicity, miR-21-3p was negatively correlated with the MR-%pancreas fat but not with MR-%liver fat. A significant correlation of both miRNAs was also observed with biomarkers related to impaired pancreatic endocrine function, including HOMA2-IR, HbA1c, and fasting plasma insulin. These analyses were exploratory and preliminary. Therefore, the clinical utility of these miRNAs as biomarkers of organ steatosis remains uncertain. However, this study highlights important observations that would help bridge the gap in understanding the underlying mechanisms linking miRNAs and the pathophysiology of pancreatic and liver steatosis.
Author Contributions
FR, DCS, CJM, RFD, and SDP designed the research. IRS, LWL and SDP enrolled the participants and conducted the clinical assessments. FR conducted the lab experiments. FR conducted the statistical analysis. FR wrote the paper. FR, IVS, CJM, RFD, MHV, JOM, SDP and DCS provided content and feedback on the manuscript. FR had primary responsibility for the final content of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by AgResearch Limited through the Strategic Science Investment (Nutritional strategies for an aging population, Contracts A19079 and A21246). Funding was also provided by the New Zealand National Science Challenge, High-Value Nutrition Program, Ministry for Business, Innovation and Employment (MBIE, grant no. 3710040).
Data Availability Statement
Approval of Research Protocol: The study was approved by the Health and Disabilities Ethics Committee (HDEC) Auckland, New Zealand (16/STH/23). Informed Consent: Written informed consent was obtained from all participating women in accordance with the Declaration of Helsinki. Registry and the registration no. of the study/trial: The study is registered with the Australian New Zealand Clinical Trials Registry at anzctr.org.au (ACTRN12616000362493).
Acknowledgements
The authors would like to acknowledge all the participants involved in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Neeland, I.J.; Ross, R.; Després, J.P.; Matsuzawa, Y.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; et al. Visceral and Ectopic Fat, Atherosclerosis, and Cardiometabolic Disease: A Position Statement. Lancet Diabetes Endocrinol. 2019, 7, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Godoy-Matos, A.F.; Silva Júnior, W.S.; Valerio, C.M. NAFLD as a Continuum: From Obesity to Metabolic Syndrome and Diabetes. Diabetol. Metab. Syndr. 2020, 12, 1–20. [Google Scholar] [CrossRef]
- Catanzaro, R.; Cuffari, B.; Italia, A.; Marotta, F. Exploring the Metabolic Syndrome: Nonalcoholic Fatty Pancreas Disease. World J. Gastroenterol. 2016, 22, 7660. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Zhang, X.J.; Ji, Y.X.; Zhang, P.; She, Z.G.; Li, H. Nonalcoholic Fatty Liver Disease Pandemic Fuels the Upsurge in Cardiovascular Diseases. Circ. Res. 2020, 126, 679–704. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Marine, M.; Lu, D.; Swartz-Basile, D.A.; Saxena, R.; Zyromski, N.J.; Pitt, H.A. Nonalcoholic Fatty Pancreas Disease. HPB 2007, 9, 312–318. [Google Scholar] [CrossRef]
- Unger, R.H. Lipid Overload and Overflow: Metabolic Trauma and the Metabolic Syndrome. Trends Endocrinol. Metab. 2003, 14, 398–403. [Google Scholar] [CrossRef]
- Singh, R.G.; Yoon, H.D.; Poppitt, S.D.; Plank, L.D.; Petrov, M.S. Ectopic Fat Accumulation in the Pancreas and Its Biomarkers: A Systematic Review and Meta-Analysis. Diabetes. Metab. Res. Rev. 2017, 33, e2918. [Google Scholar] [CrossRef]
- Heber, S.D.; Hetterich, H.; Lorbeer, R.; Bayerl, C.; Machann, J.; Auweter, S.; Storz, C.; Schlett, C.L.; Nikolaou, K.; Reiser, M.; et al. Pancreatic Fat Content by Magnetic Resonance Imaging in Subjects with Prediabetes, Diabetes, and Controls from a General Population without Cardiovascular Disease. PLoS One 2017, 12, e0177154. [Google Scholar] [CrossRef]
- Al-Mrabeh, A.; Hollingsworth, K.G.; Steven, S.; Tiniakos, D.; Taylor, R. Quantification of Intrapancreatic Fat in Type 2 Diabetes by MRI. PLoS One 2017, 12, e0174660. [Google Scholar] [CrossRef]
- Dong, Z.; Luo, Y.; Cai, H.; Zhang, Z.; Peng, Z.; Jiang, M.; Li, Y.; Li, C.; Li, Z.P.; Feng, S.T. Noninvasive Fat Quantification of the Liver and Pancreas May Provide Potential Biomarkers of Impaired Glucose Tolerance and Type 2 Diabetes. Medicine (Baltimore). 2016, 95, e3858. [Google Scholar] [CrossRef]
- LaPierre, M.P.; Stoffel, M. MicroRNAs as Stress Regulators in Pancreatic Beta Cells and Diabetes. Mol. Metab. 2017, 6, 1010–1023. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.; Otgonsuren, M.; Younoszai, Z.; Allawi, H.; Raybuck, B.; Younossi, Z. Circulating MiRNA in Patients with Non-Alcoholic Fatty Liver Disease and Coronary Artery Disease. BMJ Open Gastroenterol. 2016, 3, e000096. [Google Scholar] [CrossRef]
- Goncalves, B.d.S.; Meadows, A.; Pereira, D.G.; Puri, R.; Pillai, S.S. Insight into the Inter-Organ Crosstalk and Prognostic Role of Liver-Derived MicroRNAs in Metabolic Disease Progression. Biomedicines 2023, 11, 1597. [Google Scholar] [CrossRef] [PubMed]
- López-Bermudo, L.; Luque-Sierra, A.; Maya-Miles, D.; Gallego-Durán, R.; Ampuero, J.; Romero-Gómez, M.; Berná, G.; Martín, F. Contribution of Liver and Pancreatic Islet Crosstalk to β-Cell Function/Dysfunction in the Presence of Fatty Liver. Front. Endocrinol. (Lausanne). 2022, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Calderari, S.; Diawara, *!!! REPLACE !!!*; Garaud, A.; Gauguier, D. Biological Roles of MicroRNAs in the Control of Insulin Secretion and Action. Physiol. Genomics 2017, 49, 1–10. [Google Scholar] [CrossRef]
- Song, I.; Roels, S.; Martens, G.A.; Bouwens, L. Circulating MicroRNA-375 as Biomarker of Pancreatic Beta Cell Death and Protection of Beta Cell Mass by Cytoprotective Compounds. PLoS One 2017, 12, e0186480. [Google Scholar] [CrossRef]
- Lei, L.; Zhou, C.; Yang, X.; Li, L. Down-Regulation of MicroRNA-375 Regulates Adipokines and Inhibits Inflammatory Cytokines by Targeting AdipoR2 in Non-Alcoholic Fatty Liver Disease. Clin. Exp. Pharmacol. Physiol. 2018, 45, 819–831. [Google Scholar] [CrossRef]
- Gatfield, D.; Le Martelot, G.; Vejnar, C.E.; Gerlach, D.; Schaad, O.; Fleury-Olela, F.; Ruskeepää, A.-L.; Oresic, M.; Esau, C.C.; Zdobnov, E.M.; et al. Integration of MicroRNA MiR-122 in Hepatic Circadian Gene Expression. Genes Dev. 2009, 23, 1313–1326. [Google Scholar] [CrossRef]
- Lynn, F.C. Meta-Regulation: MicroRNA Regulation of Glucose and Lipid Metabolism. Trends Endocrinol. Metab. 2009, 20, 452–459. [Google Scholar] [CrossRef]
- Qu, Y.; Ding, Y.; Lu, J.; Jia, Y.; Bian, C.; Guo, Y.; Zheng, Z.; Mei, W.; Cao, F.; Li, F. Identification of Key MicroRNAs in Exosomes Derived from Patients with the Severe Acute Pancreatitis. Asian J. Surg. 2023, 46, 337–347. [Google Scholar] [CrossRef]
- Sequeira, I.R.; Yip, W.C.; Lu, L.W.W.; Jiang, Y.; Murphy, R.; Plank, L.D.; Cooper, G.J.S.; Peters, C.N.; Lu, J.; Hollingsworth, K.G.; et al. Pancreas Fat, an Early Marker of Metabolic Risk? A Magnetic Resonance Study of Chinese and Caucasian Women: TOFI_Asia Study. Front. Physiol. 2022, 13, 421. [Google Scholar] [CrossRef] [PubMed]
- Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2018. Diabetes Care 2018, 41, S13–S27. [CrossRef] [PubMed]
- Sequeira, I.R.; Yip, W.; Lu, L.; Jiang, Y.; Murphy, R.; Plank, L.; Zhang, S.; Liu, H.; Chuang, C.L.; Vazhoor-Amarsingh, G.; et al. Visceral Adiposity and Glucoregulatory Peptides Are Associated with Susceptibility to Type 2 Diabetes: The TOFI_Asia Study. Obesity 2020, 28, 2368–2378. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis Model Assessment: Insulin Resistance and β-Cell Function from Fasting Plasma Glucose and Insulin Concentrations in Man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.E.; Fraser, K.; Kruger, M.C.; Sequeira, I.R.; Yip, W.; Lu, L.W.; Plank, L.D.; Murphy, R.; Cooper, G.J.S.; Martin, J.C.; et al. Untargeted Metabolomics Reveals Plasma Metabolites Predictive of Ectopic Fat in Pancreas and Liver as Assessed by Magnetic Resonance Imaging: The TOFI_Asia Study. Int. J. Obes. 2021, 45, 1844–1854. [Google Scholar] [CrossRef]
- D’Souza, R.F.; Markworth, J.F.; Aasen, K.M.M.; Zeng, N.; Cameron-Smith, D.; Mitchell, C.J. Acute Resistance Exercise Modulates MicroRNA Expression Profiles: Combined Tissue and Circulatory Targeted Analyses. PLoS One 2017, 12, e0181594. [Google Scholar] [CrossRef]
- Ramzan, F.; D’Souza, R.F.; Durainayagam, B.R.; Milan, A.M.; Markworth, J.F.; Miranda-Soberanis, V.; Sequeira, I.R.; Roy, N.C.; Poppitt, S.D.; Mitchell, C.J.; et al. Circulatory MiRNA Biomarkers of Metabolic Syndrome. Acta Diabetol. 2020, 57, 203–214. [Google Scholar] [CrossRef]
- Shah, J.S.; Soon, P.S.; Marsh, D.J. Comparison of Methodologies to Detect Low Levels of Hemolysis in Serum for Accurate Assessment of Serum MicroRNAs. PLoS One 2016, 11, e0153200. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Singh, R.G.; Yoon, H.D.; Wu, L.M.; Lu, J.; Plank, L.D.; Petrov, M.S. Ectopic Fat Accumulation in the Pancreas and Its Clinical Relevance: A Systematic Review, Meta-Analysis, and Meta-Regression. Metabolism 2017, 69, 1–13. [Google Scholar] [CrossRef]
- Petäjä, E.M.; Yki-Järvinen, H. Definitions of Normal Liver Fat and the Association of Insulin Sensitivity with Acquired and Genetic NAFLD-A Systematic Review. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, L.; Wei, X.; Liu, B.; Zhao, J.; Xie, P.; Yang, B.; Wang, L. MiR-21-3p Aggravates Injury in Rats with Acute Hemorrhagic Necrotizing Pancreatitis by Activating TRP Signaling Pathway. Biomed. Pharmacother. 2018, 107, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Kuryłowicz, A.; Wicik, Z.; Owczarz, M.; Jonas, M.I.; Kotlarek, M.; Świerniak, M.; Lisik, W.; Jonas, M.; Noszczyk, B.; Puzianowska-Kuźnicka, M. NGS Reveals Molecular Pathways Affected by Obesity and Weight Loss-Related Changes in MiRNA Levels in Adipose Tissue. Int. J. Mol. Sci. 2018, 19, 66. [Google Scholar] [CrossRef] [PubMed]
- Keller, P.; Gburcik, V.; Petrovic, N.; Gallagher, I.J.; Nedergaard, J.; Cannon, B.; Timmons, J.A. Gene-Chip Studies of Adipogenesis-Regulated MicroRNAs in Mouse Primary Adipocytes and Human Obesity. BMC Endocr. Disord. 2011, 11, 7. [Google Scholar] [CrossRef]
- Androsavich, J.R.; Chau, B.N.; Bhat, B.; Linsley, P.S.; Walter, N.G. Disease-Linked MicroRNA-21 Exhibits Drastically Reduced MRNA Binding and Silencing Activity in Healthy Mouse Liver. RNA 2012, 18, 1510–1526. [Google Scholar] [CrossRef]
- Calo, N.; Ramadori, P.; Sobolewski, C.; Romero, Y.; Maeder, C.; Fournier, M.; Rantakari, P.; Zhang, F.-P.; Poutanen, M.; Dufour, J.-F.; et al. Stress-Activated MiR-21/MiR-21* in Hepatocytes Promotes Lipid and Glucose Metabolic Disorders Associated with High-Fat Diet Consumption. Gut 2016, 65, 1871–1881. [Google Scholar] [CrossRef]
- Xin, L.; Gao, J.; Wang, D.; Lin, J.H.; Liao, Z.; Ji, J.T.; Du, T.T.; Jiang, F.; Hu, L.H.; Li, Z.S. Novel Blood-Based MicroRNA Biomarker Panel for Early Diagnosis of Chronic Pancreatitis. Sci. Rep. 2017, 7, 40019. [Google Scholar] [CrossRef]
- Ling, H.-Y.; Ou, H.-S.; Feng, S.-D.; Zhang, X.-Y.; Tuo, Q.-H.; Chen, L.-X.; Zhu, B.-Y.; Gao, Z.-P.; Tang, C.-K.; Yin, W.-D.; et al. Changes in MicroRNA (MiR) Profile and Effects of Mir-320 in Insulin-Resistant 3t3-L1 Adipocytes. Clin. Exp. Pharmacol. Physiol. 2009, 36, e32–e39. [Google Scholar] [CrossRef]
- Lê, K.A.; Ventura, E.E.; Fisher, J.Q.; Davis, J.N.; Weigensberg, M.J.; Punyanitya, M.; Hu, H.H.; Nayak, K.S.; Goran, M.I. Ethnic Differences in Pancreatic Fat Accumulation and Its Relationship with Other Fat Depots and Inflammatory Markers. Diabetes Care 2011, 34, 485–490. [Google Scholar] [CrossRef]
- Huang, R.S.; Gamazon, E.R.; Ziliak, D.; Wen, Y.; Im, H.K.; Zhang, W.; Wing, C.; Duan, S.; Bleibel, W.K.; Cox, N.J.; et al. Population Differences in MicroRNA Expression and Biological Implications. RNA Biol. 2011, 8, 692–701. [Google Scholar] [CrossRef]
- Tiscornia, O.M.; Cresta, M.A.; de Lehmann, E.S.; Celener, D.; Dreiling, D.A. Effects of Sex and Age on Pancreatic Secretion. Int. J. Pancreatol. 1986, 1, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and Exosomal MicroRNA: Trafficking, Sorting, and Function. Genomics. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Correlation of circulatory miRNAs with pancreatic fat percentage.
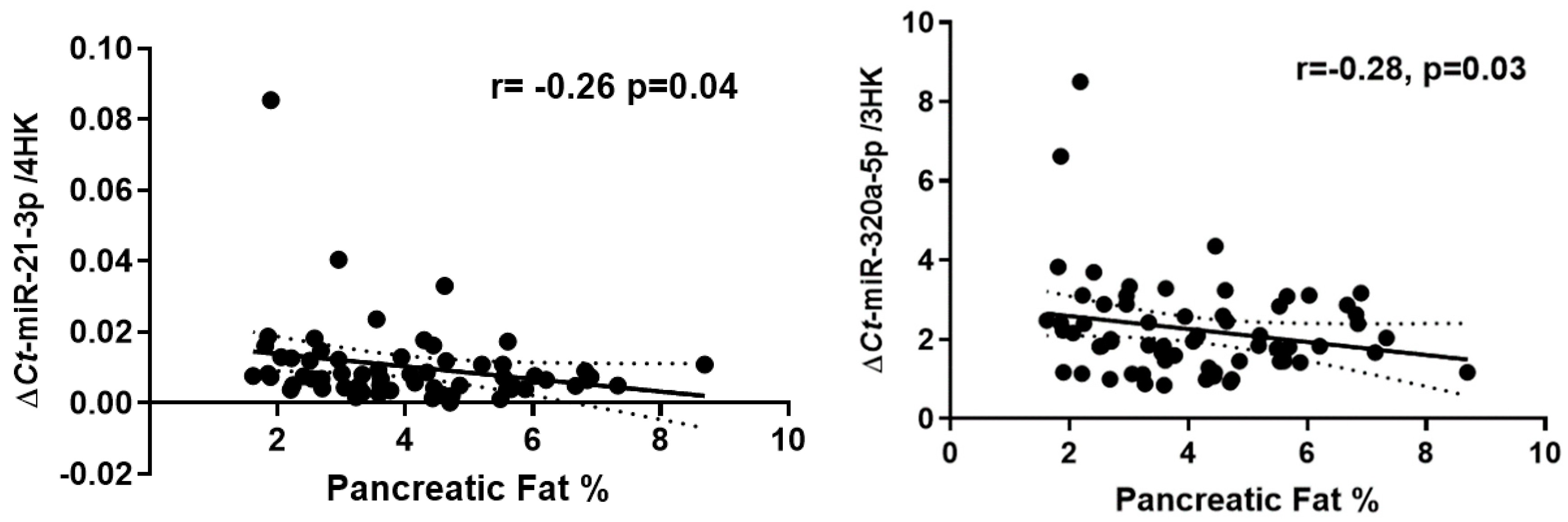
Figure 2.
Correlation of circulatory miRNAs with the established T2D risk markers. (A-B); Correlation plot comparing the relative abundance of miR-320a-5p and miR-21-3p with Plasma Insulin. (C-D) Relative abundance of miR-320a-5p and miR-21-3p with HOMA2-IR. (E) Relative abundance of miR-320a-5p with C-Peptide. (F) Relative abundance of miR-21-3p and miR-21-3p with HbA1c; r = 'Pearson's coefficient with exact p value listed. Each point represents an individual sample.
Figure 2.
Correlation of circulatory miRNAs with the established T2D risk markers. (A-B); Correlation plot comparing the relative abundance of miR-320a-5p and miR-21-3p with Plasma Insulin. (C-D) Relative abundance of miR-320a-5p and miR-21-3p with HOMA2-IR. (E) Relative abundance of miR-320a-5p with C-Peptide. (F) Relative abundance of miR-21-3p and miR-21-3p with HbA1c; r = 'Pearson's coefficient with exact p value listed. Each point represents an individual sample.

| Participant Characteristics | Asian Chinese (n=34) | European Caucasian (n=34) |
| Age (years) | 41.0 ± 13.0 | 47.8 ± 15.4 |
| BMI (kg/m2) | 26.7 ± 4.2 | 28.0 ± 4.5 |
| Waist Circumference (cm) | 85.6 ± 11.1 | 91.7± 13.9 |
| BP-Systolic (mmHg) | 120 ± 22 | 120 ± 19 |
| BP-Diastolic (mmHg) | 65 ± 12 | 64 ± 8 |
| HbA1c (%NGSP) | 5.4 | 5.3 |
| Fasting Plasma Glucose (FPG) (mmol/L) | 5.2 ± 0.5 | 5.1 ± 0.7 |
| Total Cholesterol (mmol/L) | 4.5 ± 0.9 | 5.2 ± 0.9 |
| LDL-C (mmol/L) | 2.5 ± 0.7 | 2.9 ± 0.9 |
| HDL-C (mmol/L) | 1.4 ± 0.4 | 1.8 ± 0.4 |
| Triglycerides (mmol/L) | 1.3 ± 0.7 | 1.0 ± 0.5 |
| HOMA2-IR | 1.8 ± 1.0 | 1.6 ± 1.2 |
| MR-%pancreas fat | 4.3 ± 2.0 | 4.1 ± 1.9 |
| MR-%liver fat | 4.6 ± 0.8 | 3.8 ± 0.8 |
Data are expressed as Mean ± SD, BMI, body mass index; BP, blood pressure; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; HOMA2-IR, -homeostasis model assessment of insulin resistance; HbA1c, glycosylated haemoglobin; MR, magnetic resonance; NGSP, National Glycohemoglobin Standardization Program.
Table 2.
Multivariate linear regression model showing association of MR-%pancreatic fat with alterations in the circulatory miR-21-3p and miR-320a-5p expression.
Table 2.
Multivariate linear regression model showing association of MR-%pancreatic fat with alterations in the circulatory miR-21-3p and miR-320a-5p expression.
| Model | B | SEM | Expected (B) | P-value | |
|---|---|---|---|---|---|
| (Constant) | -0.29 | 0.19 | -1.48 | 0.14 | |
| miR-21-3p | -3.87 | 1.63 | -2.37 | 0.02 | |
| miR-320a-5p | -0.02 | 0.01 | -1.53 | 0.13 | |
| BMI (kg/m2) | 0.01 | 0.00 | 3.07 | 0.00 | |
| FPG (mmol/L) | 0.11 | 0.03 | 3.60 | 0.00 | |
| Insulin (pg/ml) | -0.00 | 0.00 | -0.48 | 0.62 | |
| Ethnicity | 0.00 | 0.03 | 0.23 | 0.81 | |
Model was fitted for BMI (body mass index); FPG (fasting plasma glucose); plasma insulin and Ethnicity, B-regression β coefficient.
Table 3.
Multivariate linear regression model showing association of MR-%liver fat with alterations in the circulatory miR-21-3p and miR-320a-5p expression.
Table 3.
Multivariate linear regression model showing association of MR-%liver fat with alterations in the circulatory miR-21-3p and miR-320a-5p expression.
| Model | B | SEM | Expected (B) | P-value |
|---|---|---|---|---|
| (Constant) | -1.04 | 0.47 | -2.21 | 0.03 |
| miR-21-3p | -4.92 | 3.82 | -1.28 | 0.20 |
| miR-320a-5p | -0.02 | 0.04 | -0.50 | 0.61 |
| BMI (kg/m2) | 0.02 | 0.01 | 2.16 | 0.03 |
| FPG (mmol/L) | 0.12 | 0.08 | 1.52 | 0.13 |
| Insulin (pg/ml) | 0.00 | 0.00 | 1.86 | 0.06 |
| Ethnicity | 0.05 | 0.09 | 0.56 | 0.57 |
Model was fitted for BMI (body mass index); FPG (fasting plasma glucose); plasma insulin and Ethnicity, B-regression β coefficient.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



