Submitted:
07 August 2023
Posted:
09 August 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Frontal Temporal Dementia (FTD) is a neurological disorder known to have less therapeutic options. So far only a few biomarkers are available for FTD that can be used as potential comorbidity targets. To this end, in the present study we aim to identify potential biomarkers or susceptible genes for FTD that show comorbidities with diseases such as COVID-19 and Breast Cancer. Gene expression Omnibus datasets containing FTD expression profiles from African American and white ethnicity background were included in our study. In FTD samples of GSE193391 dataset, we identified 305 DEGs with 168 genes being up-regulated and 137 genes being down-regulated. We conducted comorbidity analysis for COVID-19 and Breast cancer followed by analysis of potential drug interactions, pathogenicity, analysis of genetic variants and functional enrichment analysis. Resulting genes AKT3, GFAP, ADCYAP1R1, VDAC1, and C4A showed significant transcriptomic alterations in FTD and significant comorbidity status with COVID-19 and Breast cancer. Functional pathway analysis revealed that these comorbid genes were significantly enriched in the pathways such as glioma, JAK/STAT signaling, systematic lupus erythematosus, neurodegeneration-multiple diseases and neuroactive ligand-receptor interaction. Overall, from these results, we concluded that these genes could be recommended as potential therapeutic targets for the treatment of comorbidities in FTD.
Keywords:
Frontal Temporal Dementia
; Comorbidity
; Breast cancer
; COVID-19
; Differential Gene expression analysis
1. Introduction
Frontotemporal disorder or frontotemporal dementia (FTD) is a disease with symptoms such as impaired thinking, reasoning, memory, and other functions leading to interferences in performing daily activities causing diverse brain degeneration [1,2]. In the USA, FTD is prevalent neurodegenerative dementia with a lowest survival rate of only 3-14 years after the onset and is known to be most prevalent among individuals with age ≤ 65 years. To date, there are no effective therapies available for treating FTD or decreasing its progression [3].
Currently, there are several factors that confer susceptibility risk to FTD. Among them, genetic risk factors such as non-coding hexanucleotide repeat expansion (HRE) located in the C9orf72 gene is known to be prevalent [4,5]. Other risk factors include mutations in the genes such as GRN and MAPT. Several new therapeutic strategies are under development targeting these genes and some of them are moving into clinical trials [6]. Though clinical subtypes and genetic factors that confer risk for FTD have been recently identified, molecular mechanisms driving its pathogenesis are not much understood. In recent years, transcriptomic studies, and network biology methods targeting the disrupted pathways have been instrumental in neurodegenerative disorders.
FTD patients show a significantly lower prevalence of cancer [7] and exhibit reciprocal phenomena with worsening of their behavior and cognition abilities upon COVID-19 infection [8]. The molecular mechanisms driving such behavior or the genes conferring this comorbidity are unknown specifically among the racial/ethnic groups such as African Americans (AA) and white population. A recent study on gene co-expression network analysis of FTD-related genes in the frontal and temporal cortices of FTD patients showed enrichment of pathways related to DNA metabolism, transcriptional regulation, and DNA protection. These results indicate that network-based approaches could offer valuable insights into the pathogenesis of FTD thereby aiding the identification of potential treatment targets [9]. Therefore, in the current study, we aim to utilize gene expression data to elucidate some of the molecular features in FTD comparing AA and White ethnicities.
2. Results
2.1. Differential expression analysis of FTD genes
The flowchart of overall data analysis performed in our study was shown in Figure 1. Differential expression analysis between the control (n=8) and FTD (n=8) samples from African American (AA) and White population (Supplementary Table 1) from the GSE193391 dataset demonstrated 305 DEGs were found to be significant at a p-value<0.05 with 168 genes being up-regulated and 137 genes being down-regulated (Supplementary Table 2). Among these DEGs, genes RAB3A (1.91), NEFL (1.89), KCNV1 (1.88), BDNF (1.86), YWHAH (1.66), GABRG2 (1.62), SH3GL2 (1.58), SYN2 (1.57), RIMS1 (1.56), PAK1 (1.53) were found to be up-regulated and the genes GFAP (-3.08), C4A/B (-2.71), CD44 (-2.36), ENTPD2 (-2.27), CD163 (-1.94), PLXNB3 (-1.76), IL18 (-1.65), SPARC (-1.60), NWD1 (-1.60), GSN (-1.59) were found to be down-regulated based on log2fold change value. DEGs were represented using the box plot (Supplementary Figure 1A). DEGs were visualized using the Volcano Plots (Supplementary Figure 1B).
2.2. Analysis of DEGs involved in autophagy
Results from human autophagy database analysis showed that among the DEGs that are significantly expressed between AA and white population, MAPK9 was found to be involved in autophagy (Figure 1). Further, results from Variation effect on Protein Structure and function (VAPROS) [23] database analysis in Autophagy database, showed that the genes AKT3, GFAP, UBE2N, VDAC1 were also found to be involved in autophagy (Figure 1). Gene AKT3 showed a cluster of 540 located in the cytoplasm, nucleus, and cell membrane. Gene GFAP showed a cluster of 675 located in the cytoplasm. Gene UBE2N showed a cluster of 483 in an unknown location. Gene VDAC1 showed a cluster of 205 located in the mitochondrion outer membrane.
2.3. Analysis of the genes involved in Covid-19
For the genes that were found to be involved in autophagy, we performed DisGeNET analysis (Supplementary Table 3). Results from the DPI analysis showed that the genes GFAP and RTN4 were found to be involved in COVID-19 with a score of 0.885 and 0.692. DSI analysis showed that the genes GFAP had a DSI value of 0.421 and gene RTN4 had a DSI value of 0.621. Furthermore, gene GFAP had a pLI value of 0.00000146 and the gene RTN4 had a pLI value of 0.45318. Both the genes GFAP and RTN4 were found to be tolerable to protein truncating variation.
2.4. Evaluation of FTD DEGs expression in Breast Invasive Carcinoma and survival analysis
We then evaluated the expression of FTD DEGs in Breast invasive carcinoma and non-cancer tissues. Results demonstrated that the genes AKT3, UBE2N, VDAC1 were significantly upregulated (Figure 2A) and the genes ADCYAP1R1, C4A, GFAP were significantly downregulated (Figure 2B) in Breast Invasive Carcinoma patients compared to the normal controls at a p-value< 0.05. Results from the survival analysis showed that the up-regulated genes AKT3, VDAC1 and the downregulated genes ADCYAP1R1, GFAP, C4A were found to be significantly associated with the overall survival of the patients with breast invasive carcinoma (Figure 3). Overall heat map of FTD-Breast Cancer comorbid genes expression in metastatic, solid tissue of normal and primary tumors of the TCGA datasets was shown in Figure 4. Further, we evaluated the immunotherapeutic drugs interactions for these FTD-Breast Cancer comorbid genes. Results from immunotherapeutic drugs interactions analysis showed that AKT3 has interactions with the drugs XL-418, Ipatasertib, Afuresertib, Triciribine, MK-2201, Miransertib, AZD-5363, SR-13668, Archexin, MSC-2363318A, Triciribine Phosphate, Uprosertib, MK-2206, Capivasertib, LY-2780301, Perifosine, GSK-690693, Vemurafenib, Omipalisib, Fasudil, Everolimus, GSK-269962A (Supplementary Table 4). GFAP has interactions with the drugs Sorbitol, Permethrin, Fadrozole, Demecolcine, Oprelvekin, Dronabinol, Amphetamine, Pioglitazone, Capsaicin, Imodipine, Propranolol, Prasterone, Methimazole, Nifedipine, Melatonin, Caffeine, Staurosporine, Isoproterenol, Colchicine, Pentoxifylline, Disulfiram, Lithium, Genistein, Progesterone, Aspirin (Supplementary Table 4). VDAC1 and ADCYAP1R1 on the other hand, had interactions with Olesoxime, Naloxone and Insulin (Supplementary Table 4). Additionally, we evaluated the list of RNA binding proteins (RBPs) interacting with these FTD-Breast Cancer comorbid genes. Results showed that there are 11 RBPs (EIF4A3, FBL, FMR1, IGF2BP1, IGF2BP2, IGF2BP3, MOV10, NOP56, NOP58, RBM47, SRSF1) interacting with AKT3, 2 RBPs (EIF4A3, IGF2BP2) interacting with VDAC1, 1 RBP (DGCR8) interacting with ADCYAP1R1, 2 RBPs (DGCR8, FUS) interacting with GFAP and 27 RBPs (AUH, BCCIP, BUD13, CSTF2T, EIF4A3, FAM120A, GTF2F1, HNRNPA1, HNRNPC, HNRNPK, HNRNPUL1, IGF2BP1, IGF2BP3, LARP7, LIN28B, PRPF8, RBFOX2, SF3A3, SF3B4, SLTM, SMNDC1, SND1, SRSF1, SRSF7, SRSF9, TRA2A, U2AF1) interacting with C4A (Supplementary Table 5). We then conducted analysis of gene interactions using RNA. Our results showed that AKT3 has the highest whereas GFAP has the lowest interaction for RBP (Supplementary Table 6). We have also conducted mRNA-RNA Interaction analysis for the FTD-Breast comorbid genes. Results showed that VDAC1 had the highest interaction and the genes ADCYAP1R1 and GFAP had the lowest interaction among the five genes (Supplementary Table 7).
2.5. Pathogenicity analysis of FTD-Breast cancer comorbid genes
We analyzed the pathogenicity for the FTD-Breast cancer comorbid genes (Supplementary Table 8). Results showed that the tools CADD, DANN, ReMM, and fitCons expressed to have the most “likely pathogenic” pathogenicity scores for the FTD-Breast comorbid genes. Tool fitCons expressed all five genes as “likely pathogenic”. DANN expressed genes AKT3, VDAC1, GFAP, and C4A as “likely pathogenic”. CADD and ReMM tools expressed genes VDAC1, GFAP, C4A, ADCYAP1R1 as “likely pathogenic”. LINSIGHT, GenoCanyon, Eigen, Eigen_PC, regBase PAT, and FATHMM-MKL tools expressed one or two genes as “likely pathogenic”. Tool CDTS did not express any of the five genes as “likely pathogenic” (Supplementary Table 8).
2.6. Genetic Alteration analysis in FTD-Breast cancer comorbid genes
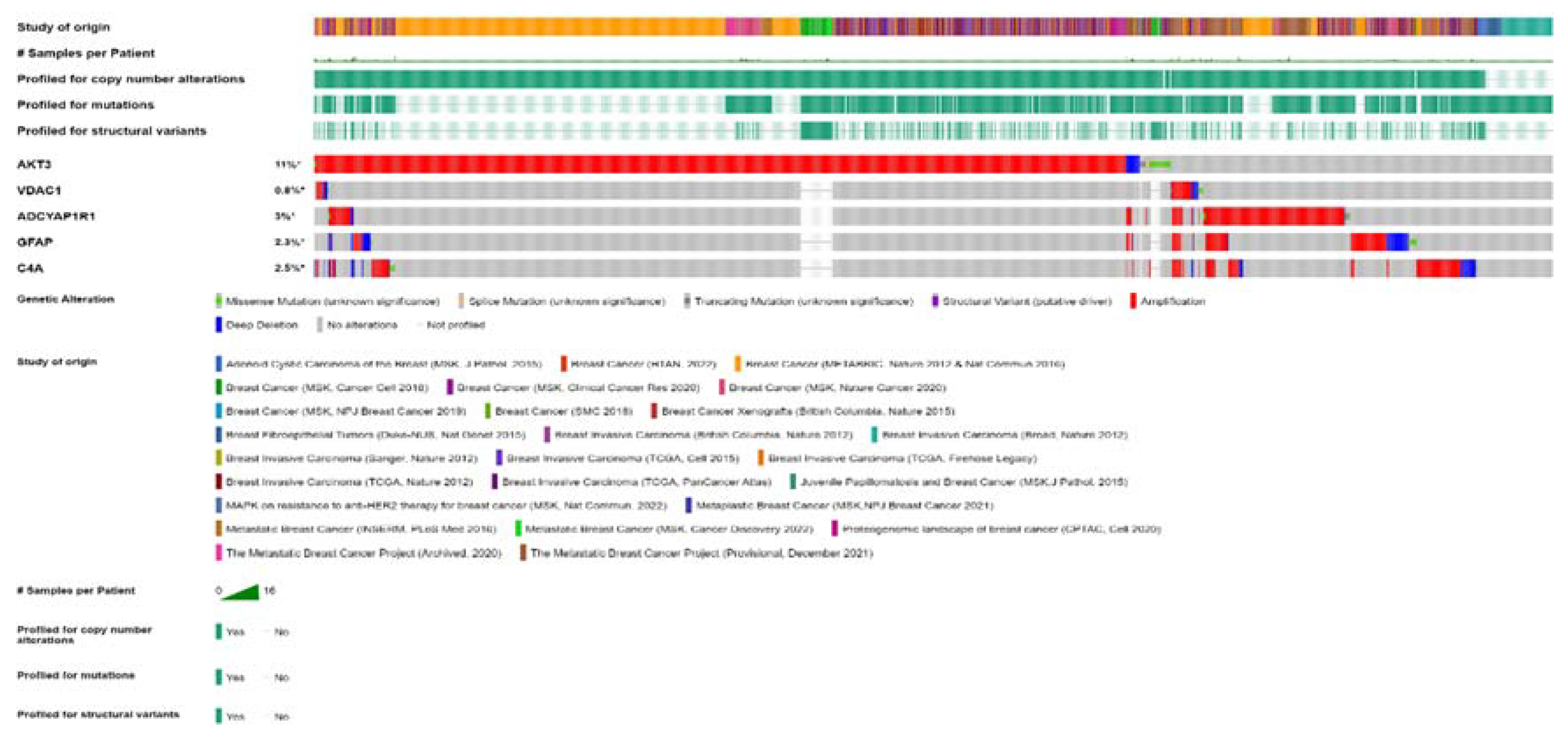
For the five FTD-Breast Cancer comorbid genes (AKT3, GFAP, C4A, VDAC1, ADCYAP1R1), we analyzed the alterations in the genetic profiles among the different subtypes of breast cancer. Results showed that AKT3 gene has the highest alterations in the genetic biomarker at 11% while VDAC1 has the lowest alterations at 0.8%. ADCYAP1R1 (3%), GFAP (2.5%) and C4A (2.3%) have similar percentages of genetic alterations. In all the genes, many of the alterations are mostly composed of amplification and deep deletion (Figure 5). To analyze these genes at single cell level, we performed the t-sne analysis in the breast cancer subtypes, Triple Negative A (TNA), Triple Negative B (TNB), Luminal A (LA), Luminal B (LB), Basal-like (BL) and H-subtype. Results showed that VDAC1 is highly expressed in all the subtypes whereas AKT3 is expressed in TNA, TNB and H-subtypes (Supplementary Figure 2).
2.7. Clinical association of FTD-Breast comorbid genes
We investigated whether common genetic variants of AKT3 and GFAP genes have a direct relationship or clinical association with the pathology of FTD by identifying 3’aQTR on 3′aQTL-atlas (https://wlcb.oit.uci.edu/3aQTLatlas/index.php). Data of 3’aQTL variants from Brain Frontal Cortex tissues for genes AKT3 and GFAP was collected (Supplementary Table 9). Using the Gene/SNP search query in Brain Frontal Cortex tissue, the AKT3 gene yielded 11 significant variants with single nucleotide variants (rs2502342, rs2998662, rs2998661, rs12077950, rs9725721, rs6429391, rs6429389, rs12075066, rs12078540, rs12073551, rs12087532) (Supplementary Figure 3). None of the transcript variants mentioned of the AKT3 gene posed a significant association with FTD [20]. The GFAP gene yielded 10 significant variants (rs12941832, rs34902223, rs9895349, rs4793148, rs3764840, rs3760382, rs4426386, rs2337848, rs9893320, rs9911454) in human Brain Frontal Cortex tissue (Supplementary Figure 4). The genetic consequence of several of these 3’UTR variants affects the promoter region of the gene and encodes the candidate tumor suppressor gene, ADAM11. ADAM11 is a highly conserved gene involved in biological pathways including fertilization, muscle development, and neurogenesis (https://www.ncbi.nlm.nih.gov/gene/4185). The ADAM11 gene is associated with the molecular pathology of breast cancer, epilepsy, and familial frontotemporal lobe. [24]
2.8. Pathway enrichment analysis
We further explored the interaction between the FTD-Breast comorbid genes (AKT3, GFAP, C4A, VDAC1, ADCYAP1R1) and their different aspects of biological pathways in relation to neurological changes by finding the pathways on https://www.kegg.jp/kegg/pathway.html [22]. Results showed that AKT3 gene is enriched with multiple pathways of glial cells impacting the brain (Supplementary Figure 5) such as Cytokine-Cytokine receptor interaction, ErbB signaling, mTOR signaling, Calcium signaling, MAPK signaling, p53 signaling, cell cycle and cell growth and proliferation. GFAP is enriched with the JAK/STAT signaling pathway (Supplementary Figure 6), C4A was enriched with the pathways such as JAK/STAT signaling, cytokine-cytokine receptor interaction, B cell/T cell receptor signaling (Supplementary Figure 7), VDAC1 was enriched with neurodegeneration pathways of multiple diseases such as Alzheimer's, Parkinson’s, Amyotrophic lateral sclerosis, Huntington’s, Spinocerebellar ataxia, and Prion’s (Supplementary Figure 8) and ADCYAP1R1 gene was enriched with neuroactive ligand-receptor interaction pathway (Supplementary Figure 9).
3. Discussion
FTD is a neurodegenerative disease affecting behavior, language and connects to pathology in the brain’s frontal and temporal lobes. [25] FTD patients show a decline in survival rates and show comorbidities with several other diseases. The correlation of these diseases such as COVID-19 and Breast cancer to FTD and survival at the molecular level is unknown [26]. So far there is no known cause for FTD and no actual therapies for the treatment of FTD patients.
Additionally, breast cancer affects 230, 480 women in the USA alone. 39, 520 of these women will die from breast cancer. [27] These statistical factors are important for our study as we measured comorbidity of FTD and Breast cancer to find the possible cause and prognosis for these diseases.
Therefore, the development of new diagnostic and prognostic genetic biomarkers for patients with FTD is urgently needed.
Differentially expressed genes among the racial/ethnic groups have been extensively explored in recent years and may harbor as prognostic biomarkers. Thus, we performed DEG analysis for the FTD among the AA and white patients. We found 305 DEGs to be significant at a p-value<0.05(Supplementary Table 2). We then analyzed their potential to be involved in autophagy and found the genes MAPK9, AKT3, GFAP, UBE2N and VDAC1 were involved in autophagy (Figure 1). We then validated whether these 5 genes have molecular underpinnings of COVID-19 specific comorbidities and identified two genes GFAP and RTN4 involved in COVID-19 and FTD (Supplementary Table 3). Then we analyzed whether any of these FTD DEGs play a key role in breast cancer prognosis and survival. We found that the FTD DEGs, AKT3, UBE2N, VDAC1, ADCYAP1R1, C4A and GFAP showed significant comorbidity in Breast Invasive Carcinoma patients at a p-value< 0.05 (Figure 2) We analyzed immunotherapeutic drugs interactions (Supplementary Table 4), RBP interactions (Supplementary Table 5), pathogenicity (Supplementary Table 8), genetic alterations (Figure 5), Clinical association (Supplementary Table 9), pathway enrichment (Supplementary Figures 5–9) of these five novel FTD-Breast Cancer comorbid genes. Our findings from this analysis suggested that these five FTD-Breast Cancer comorbid genes were novel and play a crucial role in FTD-Breast cancer comorbidity.
Overall, we found two novel FTD-COVID-19 comorbid genes GFAP, RTN4 and five novel FTD-Breast Cancer comorbid genes AKT3, GFAP, ADCYAP1R1, VDAC1, C4A in this study. AKT3 gene plays a role in AKT-Kinase, which correlates with serine and threonine stressors. In relation to FTD, this causes deterioration in keeping balance and destroys DNA [28]. VDAC1 gene interacts with IP3 receptors that work with sigma-1 receptors and mutation SOD1 acts upon these receptors. The sigma-1 receptor disrupts IP3 receptors pathway of taking Ca2+ from the endoplasmic reticulum to the mitochondria and this disturbance from the sigma-1 receptor causes the mutation SOD1 to take place and impair the endoplasmic-reticulum mitochondria signaling [29]. GFAP is known to be correlated to the process of FTD, called astrogliosis, which is the process of trying to amend neurological damage to the brain [30]. C4A gene is involved in inflammatory processes associated with Alzheimer’s and plays a role in the pathology of FTD. Several studies, such as mice and human postmortem, have shown to express upregulation in FTD. C4A gene works in changing the structure and the number of copies in genetic biomarkers. In response to the exposure of C4A gene expression, the nuclear ribonucleoprotein gives a transactive response to DNA protein binding [31]. ADCYAP1R1 gene is involved in the regulation of adrenocorticotropin and catecholamine hormones. Depletion of catecholamine compounds is linked to various underlying neurological health complications, including loss of or impaired motor function and distinct behavioral changes [32] which are consistent in FTD phenotypes. Upregulation of the ADCYAP1R1 gene is likely to influence the pathology of FTD by causing a disruption of adrenocorticotropin and catecholamine levels in the body.
Collectively, the above findings demonstrated that alterations of the FTD-COVID-19 and FTD-Breast Cancer comorbid genes AKT3, GFAP, ADCYAP1R1, VDAC1, and C4A were associated with the pathogenesis of FTD, COVID-19 and Breast cancer. These genes could be utilized as a potential therapeutic target for the treatment of comorbidities among these three diseases.
4. Materials and Methods
4.1. Data collection and identification of differentially expressed genes in FTD
Gene Expression Omnibus (GEO) dataset (GSE193391) from GEO database [10] with AA and white ethnic background (Supplementary Table 1) was selected for our study. Differential gene expression (DEG) analysis was performed using the GEO2R tool available on the GEO database. For the identification of differentially expressed genes, log2(fold change [FC]) value>2, p-value<0.05, and Bonferroni false discovery rate (FDR) < 0.05 parameters were considered as the cut-off threshold values in our study. Box plots between the samples from Control and FTD groups were created using the GEO2R (Supplementary Figure 1A).
4.2. Identification of Autophagy genes
DEGs that are significant between the healthy controls and the FTD samples were submitted to the Human Autophagy Database (http://www.autophagy.lu/index.html) (http://www.tanpaku.org/autophagy/) [11] for the identification of genes associated with Autophagy, a lysosomal degradation pathway required for survival, differentiation, development, homeostasis and known to play a crucial role in diverse pathologies, such as cancer, infections, aging and neurodegeneration.
4.3. Identification of COVID-19-associated genes
DEGs that are significant between the healthy controls and the FTD samples were submitted to DisGeNET (http://www.disgenet.org/) for identification of comorbid genes between FTD and COVID-19. DisGeNET is a discovery platform that integrates data from curated repositories of experts, GWAS catalogs, animal models, and their respective scientific literature. It contains 1, 134, 942 gene-disease associations (GDAs), between various genes, diseases, disorders, traits, clinical or abnormal human phenotypes, and their respective variant associations [12]. DisGNET measures Disease Pleiotropy Index (DPI), Disease Specificity Index (DSI), and probability of being loss-of-function intolerant (pLI). DPI is measured by two variables, the value of different disease classes of the diseases in accordance with FTD and the total number of disease classes in DisGeNET, which is 29. DPI ranges from 0 to 1. DSI is the variable that measures the gene or variant association with a disease. DSI ranges from 0 to 1 and has an inverse proportional relationship to the disease number in accordance with the specified gene. pLI on the other hand, measures how tolerant or intolerant a gene is for the protein truncating variation (variation of nonsense, splice acceptor/donor). pLI value close to 1 means the gene is intolerant.
4.4. Verification of FTD DEGs in breast cancer
To verify the expression of identified FTD DEGs with statistically significant differences in breast cancer based on a large sample size, we performed analysis using GEPIA [13] (http://gepia.cancer-pku.cn/). For the genes that were significantly expressed in breast cancer, we used the KM-plotter [14] (https://kmplot.com/analysis/) to analyze the influence of genes on breast cancer survival under low and high expression.
4.5. Identification of the alteration in the DEGs
To analyze the alterations in the identified DEGs, we submitted them to cBioPortal (http://cbioportal.org), a web server for integrative analysis of complex cancer genomics and clinical profiles (http://cbioportal.org) [15]. The Genomic Alteration Types and putative copy-number alterations were downloaded for each of the identified DEGs.
4.6. Expression analysis of FTD-breast cancer comorbid genes
To integrate, analyze and visualize the expression of public genomic data with the FTD-breast cancer comorbid genes we used UCSC Xena server [16] (https://xena.ucsc.edu/welcome-to-ucsc-xena/). Xena browser allows us to explore the functional genomic datasets to perform the correlations between variables related to genotype or phenotype. To determine the differences in FTD-Breast Cancer comorbid gene expression between tumor and normal tissues, we performed the clustering analysis of the FTD-breast cancer comorbid genes using heatmaps to compare their gene expression, exon expression, and DNA methylation.
4.7. Identification of RNA interactions for the FTD-breast cancer comorbid genes
To identify the RNA Interactomes for the FTD-breast cancer comorbid genes, we used ENCORI (The Encyclopedia of RNA Interactomes) (https://rnasysu.com/encori/index.php), a database with millions of RBP-RNA, RNA-RNA interactions, functions, and mechanisms in human diseases assessed through CLIP-seq and various high-throughput sequencing data [17].
4.8. Single-cell transcriptome profiling of FTD-breast cancer comorbid genes
To identify the transcriptome profile of FTD-breast cancer comorbid genes at single cell level, we have submitted the genes to single-cell atlas (http://bcatlas.tigem.it). Single cell atlas is a web-based server consisting of 35, 276 individual cells from 32 breast cancer cell lines clustered according to either genomic variants or copy number variations. It allows us to study tumor heterogeneity and drug response of breast cancer cell lines at single-cell level [15].
4.9. Prediction of potential therapeutic drugs
To identify interaction information about the approved and immunotherapeutic drugs for the FTD-breast cancer comorbid genes we used drug-gene interaction database (DGIdb, https://www.dgidb.org/search_interactions), a web resource that provides information on drug-gene interactions, databases, druggable genes and other web-based sources [18]. We selected the drugs with an interaction score greater than 1 as the cut-off criterion [19].
4.10. Pathogenicity Gene Score Analysis
For each of the DEGs, we performed tolerance or pathogenicity gene score analysis using Vanno Portal (http://www.mulinlab.org/vportal). Respective pathogenicity scores were taken from various tools such as Combined Annotation Dependent Deletion (CADD), Deep and Neural Network (DANN), base wise annotation (regBase PAT), spectral approach for coding and noncoding genetic variants (Eigen), finds point mutation (FATHMM-MKL), simpler meta score based on direct, known genes (Eigen_PC), Regulatory Mendelian Mutation score (ReMM), fitness consequence (fitCons), whole genome functional annotation (GenoCanyon), linear model for functional genomic data with probabilistic model (LINSIGHT), and Context-Dependent Tolerance Score (CDTS) in the Vanno portal.
4.11. Interpretation of genetic variants
To identify phenotypic variation influenced by DEGs, we conducted genetic variant analysis using 3’aQTL-atlas (https://wlcb.oit.uci.edu/3aQTLatlas) [20] search by gene/SNP across human Brain Frontal Cortex tissue. The resulting variants of the queried genes were refined for association with changes in untranslated regions or 3’UTR alternative polyadenylation (APA) site usage based on statistical significance p-value<0.05. Boxplots depicting the allele frequencies of the common genetic variants were extracted from the 3’aQTL-atlas browser to determine genotypic influence and functional interpretation of genetic variations of the genes.
4.12. Enrichment of functional pathways
To perform functional and pathway enrichment analysis, we have submitted the FTD-breast cancer comorbid genes to the Database for Annotation, Visualization, and Integrated Discovery (DAVID, https://david.ncifcrf.gov/) [21]. DAVID offers systematic and integrative functional annotation for researchers to unravel biological meaning of the submitted genes. Using DAVID, gene ontology (GO) annotation including the biological process (BP), molecular function (MF) and cellular component and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis [22] were performed on the selected genes. The value of p <0.05 was chosen as the threshold.
5. Conclusions
In conclusion, the genes AKT3, VDAC1, ADCYAP1R1, C4A and GFAP play a crucial role in the pathogenesis and clinical association of FTD. These genes could be used as novel diagnostic and prognostic biomarkers and therapeutic targets for FTD patients.
Author Contributions
Conceptualization, K.N.C. ; Data curation, I.D. M., J.R., K.N.C; Formal analysis, I.D. M., J.R., K.N.C ; Funding acquisition, K.N.C.; Investigation, I.D. M., J.R.; Methodology, I.D. M., J.R., K.N.C.; Project administration, K.N.C.; Resources, K.N.C.; Software, I.D. M., J.R., K.N.C; Supervision, K.N.C.; Validation, I.D. M., J.R.; Visualization, I.D. M., J.R., K.N.C.; Writing—original draft, I.D. M., J.R., K.N.C.; Writing—review & editing, K.N.C.
Funding
Work on the manuscript and its revisions were supported by the National Institute on Minority Health and Health Disparities (NIMHD) of the National Institutes of Health (NIH) to the University of Houston under Award Number U54MD015946. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khan I, De Jesus O: Frontotemporal Lobe Dementia. In: StatPearls. Treasure Island (FL): StatPearls Publishing Copyright © 2023, StatPearls Publishing LLC.; 2023.
- Onyike, C.U.; Diehl-Schmid, J. The epidemiology of frontotemporal dementia. Int Rev Psychiatry 2013, 25, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Magrath Guimet N, Zapata-Restrepo LM, Miller BL: Advances in Treatment of Frontotemporal Dementia. J Neuropsychiatry Clin Neurosci 2022, 34, 316–327. [CrossRef] [PubMed]
- Zanella I, Zacchi E, Piva S, Filosto M, Beligni G, Alaverdian D, Amitrano S, Fava F, Baldassarri M, Frullanti E et al.: C9orf72 Intermediate Repeats Confer Genetic Risk for Severe COVID-19 Pneumonia Independently of Age. Int J Mol Sci 2021, 22.
- Smeyers J, Banchi EG, Latouche M: C9ORF72: What It Is, What It Does, and Why It Matters. Front Cell Neurosci 2021, 15, 661447. [CrossRef]
- Rohrer JD, Warren JD, Fox NC, Rossor MN: Presymptomatic studies in genetic frontotemporal dementia. Rev Neurol (Paris) 2013, 169, 820–824. [CrossRef]
- Katisko K, Haapasalo A, Koivisto A, Krüger J, Hartikainen P, Korhonen V, Helisalmi S, Herukka SK, Remes AM, Solje E: Low Prevalence of Cancer in Patients with Frontotemporal Lobar Degeneration. J Alzheimers Dis 2018, 62, 789–794.
- Ng KP, Chiew HJ, Hameed S, Ting SKS, Ng A, Soo SA, Wong BYX, Lim L, Yong ACW, Mok VCT et al. : Frontotemporal dementia and COVID-19: Hypothesis generation and roadmap for future research. Alzheimers Dement (N Y) 2020, 6, e12085. [Google Scholar] [CrossRef]
- Bottero V, Alrafati F, Santiago JA, Potashkin JA: Transcriptomic and Network Meta-Analysis of Frontotemporal Dementias. Front Mol Neurosci 2021, 14, 747798. [CrossRef]
- Edgar R, Domrachev M, Lash AE: Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res 2002, 30, 207–210. [CrossRef]
- Homma K, Suzuki K, Sugawara H: The Autophagy Database: an all-inclusive information resource on autophagy that provides nourishment for research. Nucleic Acids Res 2011, 39, D986–D990. [CrossRef]
- Piñero J, Bravo À, Queralt-Rosinach N, Gutiérrez-Sacristán A, Deu-Pons J, Centeno E, García-García J, Sanz F, Furlong LI: DisGeNET: a comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res 2017, 45, D833–d839. [CrossRef] [PubMed]
- Tang Z, Li C, Kang B, Gao G, Zhang Z: GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res 2017, 45, W98–w102. [CrossRef] [PubMed]
- Lánczky A, Győrffy B: Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J Med Internet Res 2021, 23, e27633. [CrossRef] [PubMed]
- Gambardella G, Viscido G, Tumaini B, Isacchi A, Bosotti R, di Bernardo D: A single-cell analysis of breast cancer cell lines to study tumour heterogeneity and drug response. Nat Commun 2022, 13, 1714. [CrossRef]
- Goldman MJ, Craft B, Hastie M, Repečka K, McDade F, Kamath A, Banerjee A, Luo Y, Rogers D, Brooks AN et al. : Visualizing and interpreting cancer genomics data via the Xena platform. In: Nat Biotechnol. 2020, 38, 675–678. [Google Scholar]
- Kr Z: An encyclopedia of RNA interactomes in ENCORI. In. Edited by Huang JH LS, Li B, Liu SR, Zheng WJ, cai L, et al.
- Freshour SL, Kiwala S, Cotto KC, Coffman AC, McMichael JF, Song JJ, Griffith M, Griffith OL, Wagner AH: Integration of the Drug-Gene Interaction Database (DGIdb 4. 0) with open crowdsource efforts. Nucleic Acids Res 2021, 49, D1144–d1151. [Google Scholar] [CrossRef]
- Chen Z, Zhong Z, Zhang W, Su G, Yang P: Integrated Analysis of Key Pathways and Drug Targets Associated With Vogt-Koyanagi-Harada Disease. Front Immunol 2020, 11, 587443. [CrossRef]
- Cui Y, Peng F, Wang D, Li Y, Li JS, Li L, Li W: 3'aQTL-atlas: an atlas of 3'UTR alternative polyadenylation quantitative trait loci across human normal tissues. Nucleic Acids Res 2022, 50, D39–d45. [CrossRef]
- Sherman BT, Hao M, Qiu J, Jiao X, Baseler MW, Lane HC, Imamichi T, Chang W: DAVID: a web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res 2022, 50, W216–w221. [CrossRef]
- Kanehisa M, Goto S: KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res 2000, 28, 27–30. [CrossRef]
- Gojobori T, Ikeo K, Katayama Y, Kawabata T, Kinjo AR, Kinoshita K, Kwon Y, Migita O, Mizutani H, Muraoka M et al. : VaProS: a database-integration approach for protein/genome information retrieval. J Struct Funct Genomics 2016, 17, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Wolfsberg TG, Primakoff P, Myles DG, White JM: ADAM, a novel family of membrane proteins containing A Disintegrin And Metalloprotease domain: multipotential functions in cell-cell and cell-matrix interactions. J Cell Biol 1995, 131, 275–278. [CrossRef] [PubMed]
- Finger EC: Frontotemporal Dementias. Continuum (Minneap Minn) 2016, 22, 464–489.
- Devenney EM, Ahmed RM, Hodges JR: Frontotemporal dementia. Handb Clin Neurol 2019, 167, 279–299.
- DeSantis C, Siegel R, Bandi P, Jemal A: Breast cancer statistics, 2011. CA Cancer J Clin 2011, 61, 409–418.
- Palluzzi F, Ferrari R, Graziano F, Novelli V, Rossi G, Galimberti D, Rainero I, Benussi L, Nacmias B, Bruni AC et al. : A novel network analysis approach reveals DNA damage, oxidative stress and calcium/cAMP homeostasis-associated biomarkers in frontotemporal dementia. PLoS One 2017, 12, e0185797. [Google Scholar]
- Martín-Guerrero SM, Markovinovic A, Mórotz GM, Salam S, Noble W, Miller CCJ: Targeting ER-Mitochondria Signaling as a Therapeutic Target for Frontotemporal Dementia and Related Amyotrophic Lateral Sclerosis. Front Cell Dev Biol 2022, 10, 915931.
- Heller C, Foiani MS, Moore K, Convery R, Bocchetta M, Neason M, Cash DM, Thomas D, Greaves CV, Woollacott IO et al. : Plasma glial fibrillary acidic protein is raised in progranulin-associated frontotemporal dementia. J Neurol Neurosurg Psychiatry 2020, 91, 263–270. [Google Scholar] [CrossRef]
- Reus LM, Pasaniuc B, Posthuma D, Boltz T, Pijnenburg YAL, Ophoff RA: Gene Expression Imputation Across Multiple Tissue Types Provides Insight Into the Genetic Architecture of Frontotemporal Dementia and Its Clinical Subtypes. Biol Psychiatry 2021, 89, 825–835. [CrossRef]
- Nagatsu T: The catecholamine system in health and disease -Relation to tyrosine 3-monooxygenase and other catecholamine-synthesizing enzymes. Proc Jpn Acad Ser B Phys Biol Sci 2007, 82, 388–415.
Figure 1.
Flow chart of the analysis Bioinformatics analysis performed in this study.
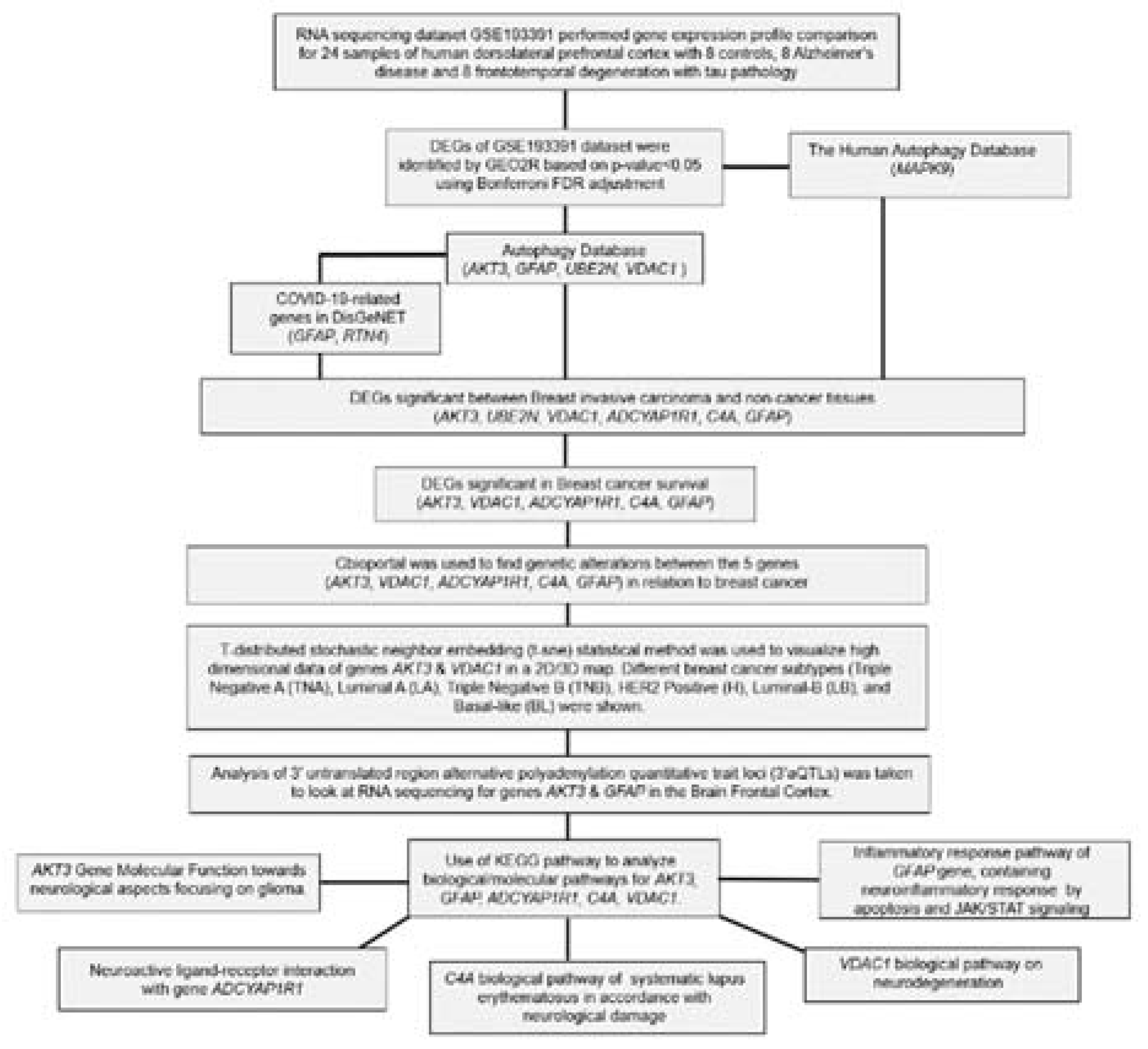
Figure 2.
Expression of FTD DEGs in Breast invasive carcinoma and non-cancer tissues. (a) Upregulated DEGs expression in Breast invasive carcinoma (BRCA) patients and normal controls (b) Down regulated DEGs expression in Breast invasive carcinoma (BRCA) patients and normal controls. Red color boxes represent Cancer whereas black color boxes represent normal, * represents significance at a p-value<0.05.
Figure 2.
Expression of FTD DEGs in Breast invasive carcinoma and non-cancer tissues. (a) Upregulated DEGs expression in Breast invasive carcinoma (BRCA) patients and normal controls (b) Down regulated DEGs expression in Breast invasive carcinoma (BRCA) patients and normal controls. Red color boxes represent Cancer whereas black color boxes represent normal, * represents significance at a p-value<0.05.
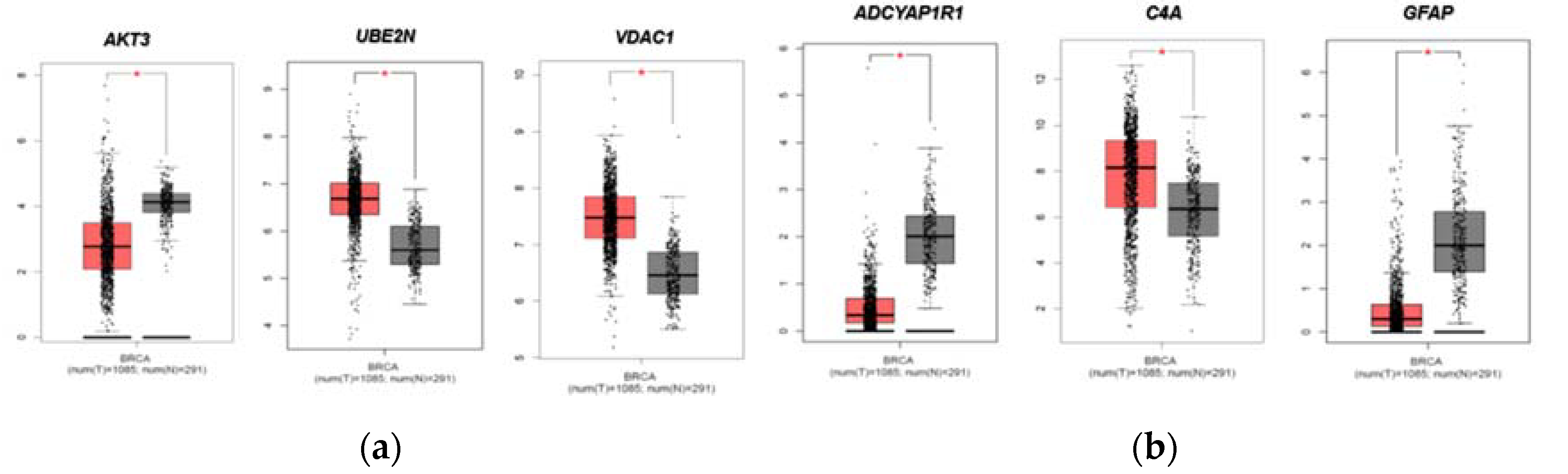
Figure 3.
FTD DEGs significantly affecting the overall survival of the patients with breast invasive carcinoma (a) Upregulated genes (b) Downregulated genes.
Figure 3.
FTD DEGs significantly affecting the overall survival of the patients with breast invasive carcinoma (a) Upregulated genes (b) Downregulated genes.
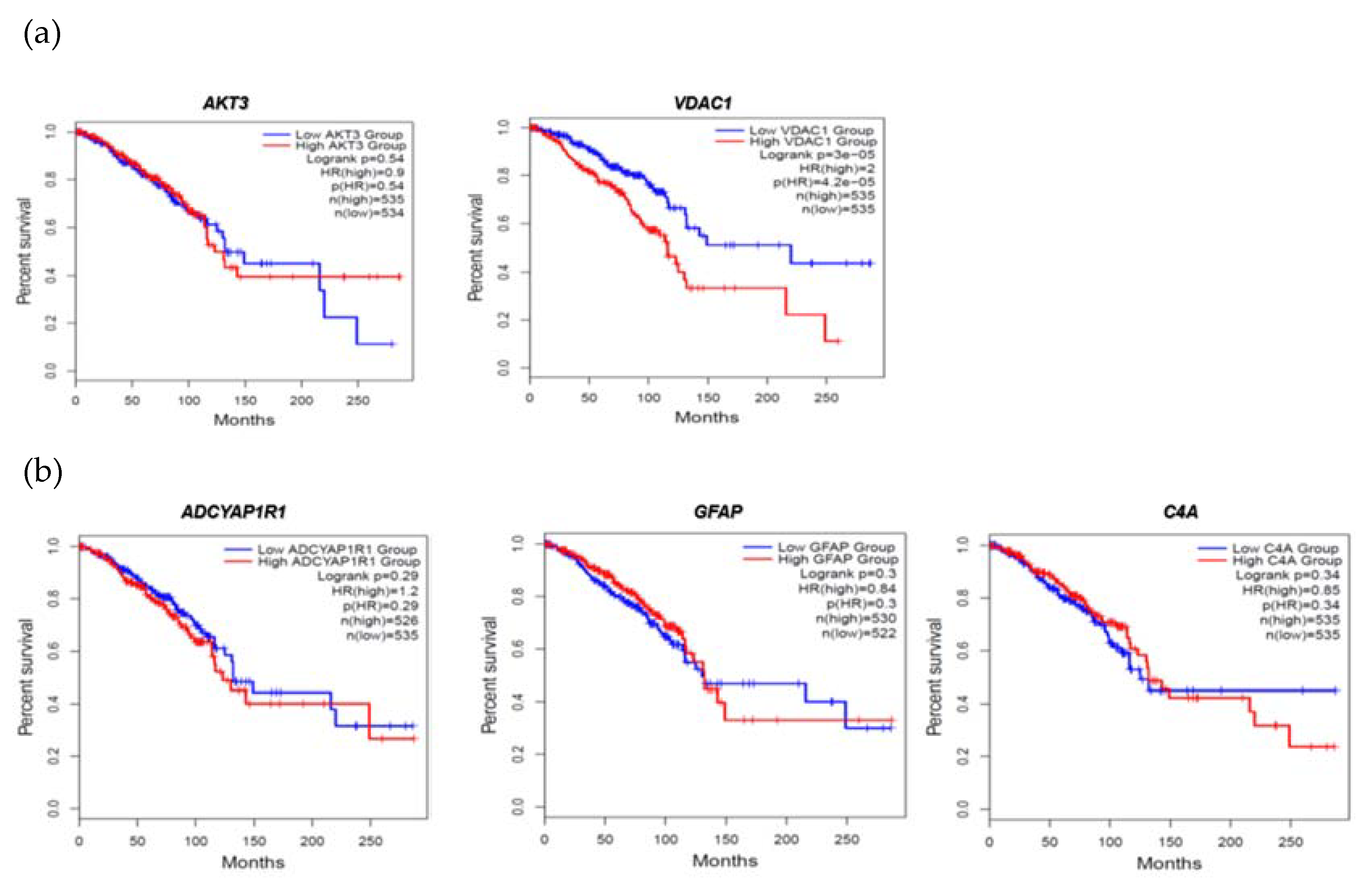
Figure 4.
Heat map of FTD-breast cancer comorbid genes expression in the metastatic, solid tissue normal and primary tumors of TCGA datasets. Metastatic is shown in purple, Solid Tissue Normal in shown in red and Primary Tumor is shown in blue. The significance of the DEGs were measured in log2(normal count +1) representing red as the stronger the correlation and blue as the weaker the correlation.
Figure 4.
Heat map of FTD-breast cancer comorbid genes expression in the metastatic, solid tissue normal and primary tumors of TCGA datasets. Metastatic is shown in purple, Solid Tissue Normal in shown in red and Primary Tumor is shown in blue. The significance of the DEGs were measured in log2(normal count +1) representing red as the stronger the correlation and blue as the weaker the correlation.
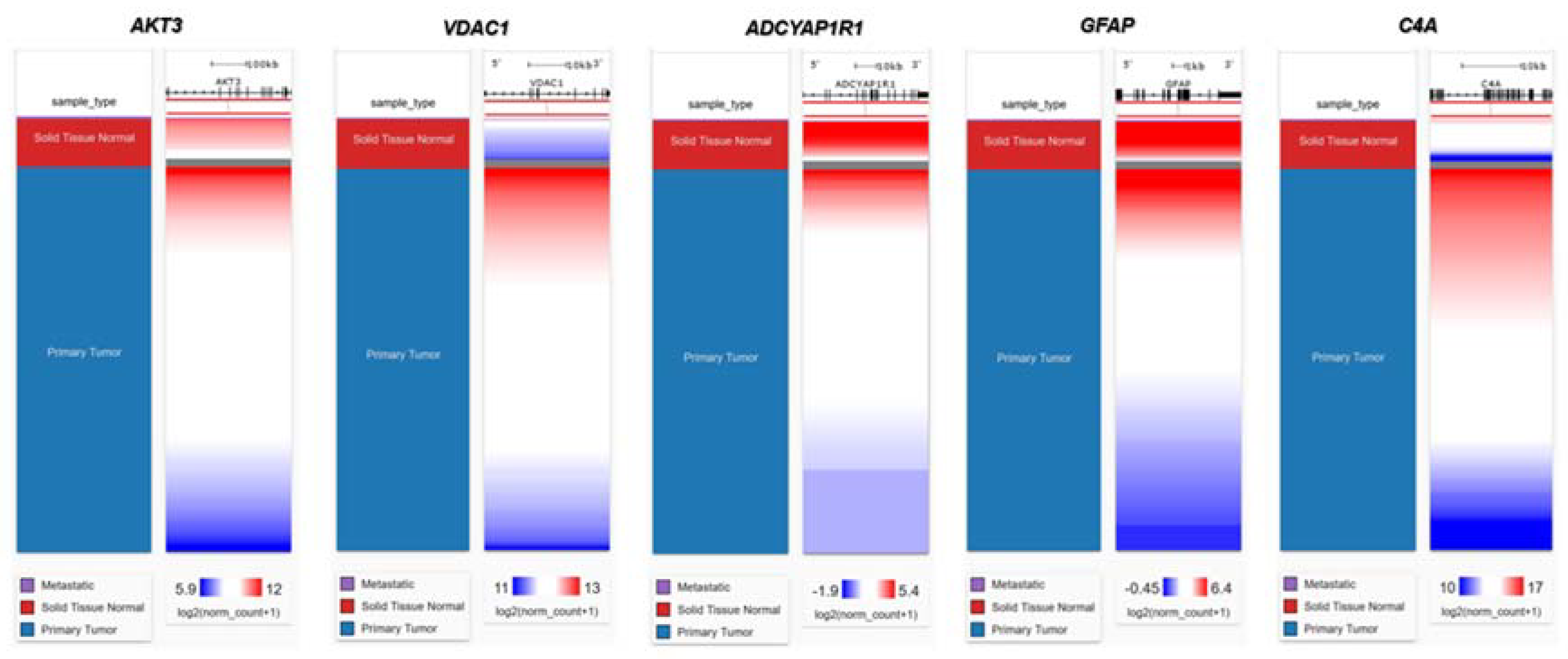
Figure 5.
cBioPortal "oncoprint" representation of alterations in FTD genes identified in breast cancer consisting of 31, 500 samples (amplification, red; deletion, blue; mutation, green). Numbers represent the combined frequency of all alterations. TCGA datasets selected were shown above.
Figure 5.
cBioPortal "oncoprint" representation of alterations in FTD genes identified in breast cancer consisting of 31, 500 samples (amplification, red; deletion, blue; mutation, green). Numbers represent the combined frequency of all alterations. TCGA datasets selected were shown above.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



