
Submitted:
08 August 2023
Posted:
09 August 2023
You are already at the latest version
Abstract
A relevant aspect in the epidemiology of Tegumentary Leishmaniasis (TL) is the Leishmania parasites carrying a viral endosymbiont Leishmania RNA Virus 1 (LRV1), a dsRNA virus. Leishmania parasites carrying LRV1 are prone to causing more severe TL symptoms, increasing the likelihood of unfavorable clinical outcomes. LRV1 has been observed in cultured strains of five L. (Viannia) species, and host specificity was suggested when studying LRV1 from L. braziliensis and L. guyanensis strains. The coevolution hypothesis of LRV1 and Leishmania was based on phylogenetic analyses, implying an association between LRV1 genotypes, Leishmania species, and their geographic origins. This study aimed to investigate LRV1 specificity to Leishmania (Viannia) species hosts by analyzing LRV1 from L. (Viannia) species. To this end, LRV1 was screened in L. (Viannia) species other than L. braziliensis or L. guyanensis, and it was detected in 11 out of 15 L. naiffi and in 2 out of 4 L. shawi. Phylogenetic analyses based on partial LRV1 genomic sequencing supported the hypothesis of host specificity, as LRV1 clustered according to their respective Leishmania species hosts. These findings underscore the importance of investigating Leishmania and LRV1 coevolution and its impact on Leishmania (Viannia) species dispersion and pathogenesis in the American Continent.
Keywords:
Leishmania (Viannia)
; Leishmania RNA Virus 1
; phylogeny
; host-specificity
; coevolution.
1. Introduction
Cutaneous leishmaniasis (CL) is a neglected disease that mainly affects the poorest populations in developing countries [1]. Brazil is the country with higher incidence of CL in the American Continent, where the disease is associated to at least seven species, one of the subgenus Leishmania: L. (Leishmania) amazonensis and the others of the subgenus Viannia: L. (Viannia) guyanensis, L. (Viannia) braziliensis, L. (Viannia) shawi, L. (Viannia) lainsoni, L. (Viannia) naiffi and L. (Viannia) lindenbergi [2].
A relevant aspect in the epidemiology of CL in Brazil, but not only observed in this region, is the circulation of parasites carrying a viral endosymbiont denominated Leishmania RNA Virus 1 (LRV1), found in L. (Viannia) sp. [3,4]. The outcome and severity of CL might be correlated with several factors, including the Leishmania species involved and the presence of LRV1 [4,5,6,7,8,9,10].
It was shown in experimental infections that Leishmania parasites carrying LRV1 are predisposed to aggravate the course of infection, leading to the mucosal form of leishmaniasis. The role of LRV1 in the exacerbation of the infection relies on Type I interferon (Type I IFNs) production by macrophages and in vivo signaling [11,12]. It is known that the virus can subvert innate immunity by acting as a negative regulator of the NLRP3 inflammasome, favoring parasites’ survival and disease chronification [13]. Furthermore, it was described that the presence of LRV1 increases the risk of mucosal development in human patients by almost three times compared to individuals who present CL without virus detection [4]. However, the association of LRV1 and mucosal manifestation was not observed in studies evaluating other cohorts, but association with therapeutic failure was demonstrated [14,15,16]. Differences in the parasites circulating in the different regions evaluated and/or in the LRV1 infecting these parasites are possible explanations for the observed differences among some studies.
Previous studies with the complete genome sequences of LRV1 and sequences obtained from a phylogenetically informative region of the viral genome from L. (V.) guyanensis and L. (V.) braziliensis showed evidence of host-specificity in L. (Viannia) species-LRV1 interaction, clustering LRV1 sequences according to their Leishmania species host [17,18]. LRV1 from one L. (V.) shawi strain was also analyzed, clustering close to LRV1 from L. guyanensis, mirroring what is observed in the phylogenetic relationship of these two Leishmania species [18]. Although LRV1 was detected already in cultivated strains of L. naiffi and L. panamensis [15,19,20,21], there is no information on phylogenetic relationship of viruses from these species comparing to LRV1 from other species. LRV1 was already associated with human infections caused by L. lainsoni [4] and L. peruviana [22], but not in cultivated strains from these species.
Due to the significance of the Leishmania-LRV1 symbiosis in the epidemiology of cutaneous and mucosal leishmaniasis, it is crucial to gain a comprehensive understanding of the viruses’s diversity and spread within parasite populations. Thus, our study aimed to examine the presence of LRV1 in various strains of Brazilian L. (Viannia) species, excluding L. braziliensis and L. guyanensis. LRV1 was not detected in different strains of three species with apparently restricted circulation in the Amazon region, and common in areas with reported circulation of LRV1: L. lainsoni, L. lindenbergi and L. utingensis. However, our findings revealed the presence of LRV1 in cultures identified as L. naiffi and L. shawi, confirming previous observations. Of note, phylogenetic analyses indicates L. naiffi, along with L. lainsoni, as the most divergent species of the Viannia subgenus [23]. Patients infected with L. naiffi experienced a poor response to antimonial or pentamidine therapy, indicating that the species could resist to first-line treatment. The presence of LRV1 in L. naiffi raised the possibility that this virus could increase Leishmania spp virulence and thereby influence therapeutic failure [15,24]. L. (V.) shawi is a species closely related to L. guyanensis [23] dispersed mainly in the Amazon region [25]. This species is responsible for CL, usually found as a single lesion, but cases of multiple lesions, clearly due to metastases, are occasionally seen [26]. A recent report showed the first L. shawi infection in mucosal secretion in Brazil, that represents a warning for the possible association between L. (V.) shawi and mucosal lesions [27]. In order to contribute with characterization of LRV1 from different L. (Viannia) species and strains and further investigate host-specificity in this combination, we conducted a comparative analyses of publicly available sequences of L. braziliensis and L. guyanensis to newly obtained LRV1 from both L. naiffi and L. shawi, reported herein.
2. Materials and Methods
2.1. Leishmania Culture
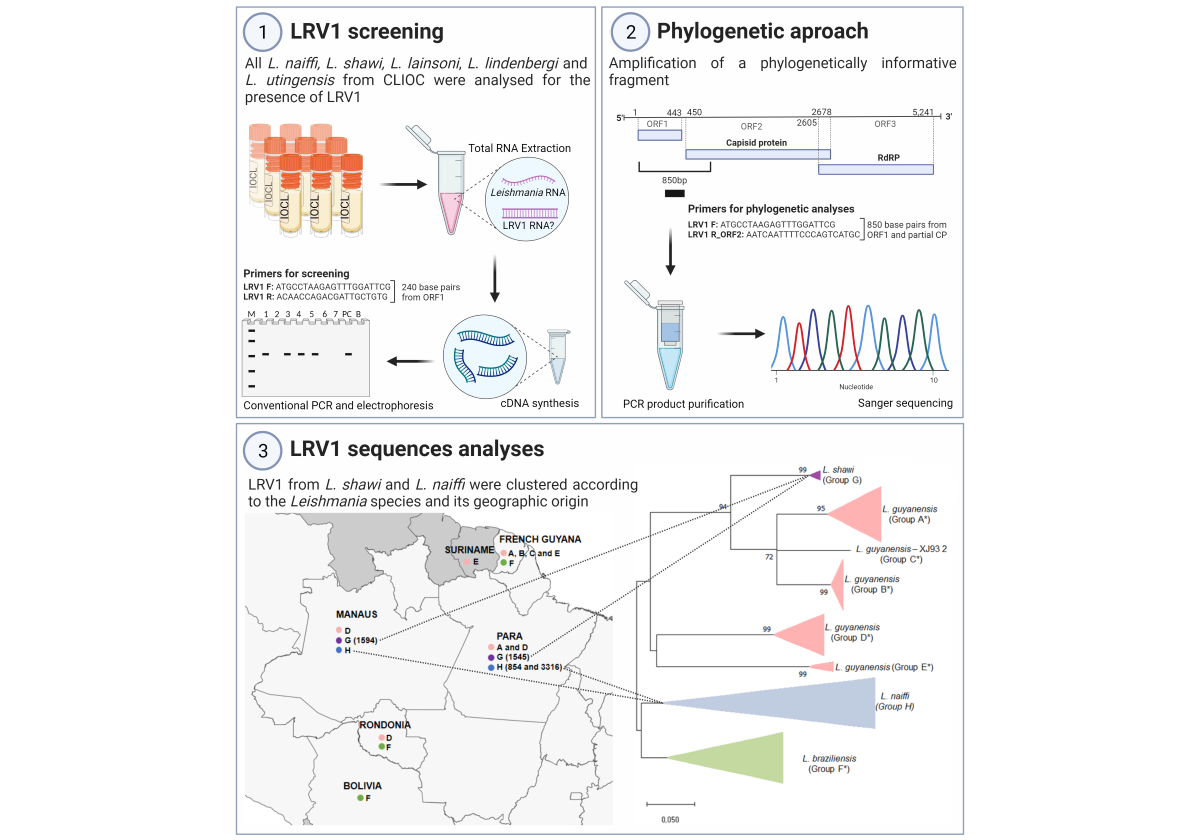
Strains from L. lainsoni (n=4), L. lindenbergi (n=3), L. naiffi (n= 18), L. shawi (n=4) and L. utingensis (n= 1), from different geographic regions, available at Leishmania Collection of Fiocruz (CLIOC), were screened (Table S1). Parasites were grown in NNN (Novy-MacNeal-Nicolle), and Schneider medium supplemented with 20% fetal bovine serum and incubated in a BOD (biochemical oxygen demand) incubator at 25º C until reaching the average amount of 5x105 parasites.
Cultures were centrifuged at 1400 g for 10 minutes at 4°C, resuspended in DNA/RNA Shield™ (Zymo Research Corporation – Irvine, CA/USA), and stored at -20°C until RNA extraction.
2.2. RNA Extraction and cDNA Synthesis
RNA of the strains was extracted using the TRIzol® reagent (Invitrogen® – USA). RNA concentration and purity were determined using a NanoDrop® 2000 spectrophotometer (Thermo Scientific™ – Wilmington, MA/USA). Reverse transcription was performed using 2μg of RNA through the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems™ – Foster City, CA/USA), following the manufacturer's recommendations.
2.3. LRV1 Detection
To determine which strains deposited in the Fiocruz Leishmania Collection (CLIOC) were positive for LRV1, a screening protocol was performed using the primers LRV F - 5´- ATGCCTAAGAGTTTGGATTCG- 3’ and LRV R - 5´- ACAACCAGACGATTGCTGTG – 3’ [4] (Figure 1). For the HSP70 fragment amplification, which was used as an endogenous control of all RT-PCR reactions, primers Hsp70cF 5- GGACGAGATCGAGCGCATGGT-3’ and Hsp70cR 5´- TCCTTCGACGCCTCCTGGTTG-3’ were used [28]. For both reactions a final volume of 50 μl was used: 10X Buffer (1X), MgCl2 (1.5mM), dNTPs (0.2Mm), Primer F (0.2μM), Primer R (0.2mM), Taq Platinum (1.0U/μl) and 1 μl of cDNA. RT-PCR was performed at 94ºC for 2 minutes, 35 cycles at 94ºC for 30 seconds, 58ºC for 30 seconds and 72ºC for 45 seconds, with a final extension phase at 72ºC for 5 minutes.
Positive strains for LRV1 (n = 12; 11 L. naiffi and 1 L. shawi) were submitted to another PCR reaction that aimed to amplify a fragment of approximately 850 base pairs for LRV1, which is a phylogenetically informative fragment. For this, the primers LRV1 F orf1 5'-ATGCCTAAGAGTTTGGATTCG-3' and LRV1 R orf2 5'-AATCAATTTTCCCAGTCATGC-3' [18] were used, amplifying a fragment corresponding to part of the orf1 region and the beginning of the orf2 region, including the portion responsible for encoding the viral capsid protein (Figure 1). A final volume of 50 μl was used: 10X Buffer (1X), MgCl2 (1.5Mm), dNTPs (0.2Mm), Primer F (0.2μM), Primer Rg (0.2mM), Taq Platinum (1.0U /µl) and 3 µl of cDNA. RT-PCR was performed at 95ºC for 2 minutes, followed by 35 cycles at 95ºC for 30 seconds, 57ºC for 45 seconds and 72ºC for 45 seconds, with a final extension phase at 72ºC for 5 minutes. All RT-PCR products were stained with GelRed® (Biotium – Fremont, CA/USA) and visualized in a 2% agarose gel.
2.4. Sequencing
For sequencing, 45 μl of RT-PCR products were purified using the Wizard® SV Gel and RT-PCR Clean-Up System kit (Promega – Madison, WI/USA), following the manufacturer's recommendations, with a final elution volume of 45 μl. Sequencing was performed with the same primers for RT-PCR amplification on the Fiocruz DNA Sequencing Platform – Rio de Janeiro (RPT01A), where the technique employed was Sanger sequencing.
2.5. Analyses of LRV1 Sequences
The consensus sequences were created in the BioEdit program [29]. Then, the inference of the best model to build the phylogenetic tree was performed in MEGAX [30] software, where the Tamura 3-parameter model with gamma distribution parameter (G) and invariable sites (I) was the one presenting the lowest BIC (Bayesian Information Criterion) score. The tree was constructed using the maximum likelihood method employing 10.000 replicates (bootstrap).
In addition, to increase the chances of finding the most parsimonious connections, networks using NeighborNet were built by the SplitsTree program [31]. For that, we use the MEGAX function to exclude sites with missing/ambiguous data and gaps of the sequences, which resulted in sequences that were 509 nucleotides long and were common across groups of each species.
3. Results
3.1. LRV1 Was Not Detected in all L. (Viannia) Species Analyzed, but Was Frequent in L. naiffi Strains
All RT-PCR reactions were performed with promastigotes of available strains identified as L. lainsoni, L. lindenbergi, L. naiffi, L. shawi, and L. utingensis, in addition to L. guyanensis (MHOM/BR/1975/M4147) and L. braziliensis (MHOM/BR/1975/M2903) strains, which were used as positive and negative controls for the experiments, respectively. In total, 4 L. lainsoni, 3 L. lindenbergi, 18 L. naiffi, 4 L. shawi and 1 L. utingensis strains were submitted to screening protocol for LRV1 detection. Of these, 7 L. naiffi, 3 L. shawi, and all L. lainsoni, L. lindenbergi and L. utingensis strains were negative for the virus. LRV1 has been detected in 11 L. naiffi strains (61%) and in one L. shawi analyzed, in addition to the strain of this species that had already been described presenting LRV1 [18]. After quality checking, sequences ranging from 632nt to 799nt (Table 1) were employed in the analyses described below.
Figure 2.
Partial map of South and Central America indicating the countries and Brazilian states where LRV1 has been detected in Leishmania (Viannia) spp. analyzed in this study. Colored circles refer to species presenting LRV1: pink= L. guyanensis; green = L. braziliensis; purple = L. shawi and blue = L. naiffi. Letters (A to H) indicate groups defined accordingly the phylogenetic analyses (Figure 3); numbers indicate specific strains (Table 1) in such group.
Figure 2.
Partial map of South and Central America indicating the countries and Brazilian states where LRV1 has been detected in Leishmania (Viannia) spp. analyzed in this study. Colored circles refer to species presenting LRV1: pink= L. guyanensis; green = L. braziliensis; purple = L. shawi and blue = L. naiffi. Letters (A to H) indicate groups defined accordingly the phylogenetic analyses (Figure 3); numbers indicate specific strains (Table 1) in such group.

Figure 3.
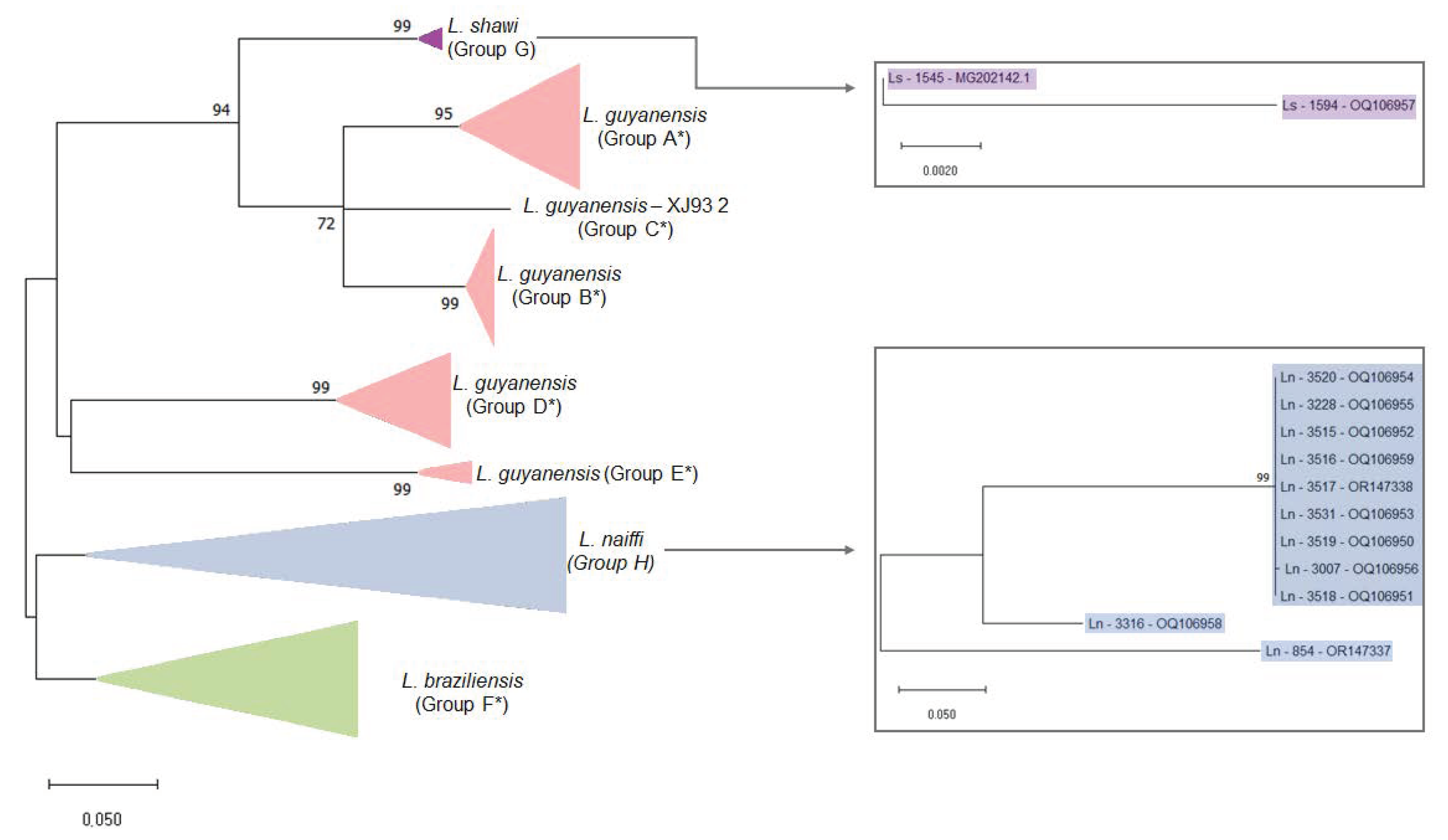
Maximum Likelihood phylogenetic tree of Leishmania RNA Virus 1 found in Leishmania (Viannia) species. The evolutionary history was inferred by using the Maximum Likelihood method and Tamura 3-parameter model [32]. The tree with the highest log likelihood (-4303.97) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Tamura 3 parameter model, and then selecting the topology with superior log likelihood value. A discrete Gamma distribution was used to model evolutionary rate differences among sites (5 categories (+G, parameter = 0.8242)). The rate variation model allowed for some sites to be evolutionarily invariable ([+I], 28.96% sites). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. This analyses involved 59 nucleotide sequences. There were a total of 442 positions in the final dataset. Ls = L. shawi; and Ln = L. naiffi. For details of each strains see Table 1 and Table S1. *Groups defined following the proposal of Tirera et al. [17]; Groups G and H were defined in the present study.
Figure 3.
Maximum Likelihood phylogenetic tree of Leishmania RNA Virus 1 found in Leishmania (Viannia) species. The evolutionary history was inferred by using the Maximum Likelihood method and Tamura 3-parameter model [32]. The tree with the highest log likelihood (-4303.97) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Tamura 3 parameter model, and then selecting the topology with superior log likelihood value. A discrete Gamma distribution was used to model evolutionary rate differences among sites (5 categories (+G, parameter = 0.8242)). The rate variation model allowed for some sites to be evolutionarily invariable ([+I], 28.96% sites). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. This analyses involved 59 nucleotide sequences. There were a total of 442 positions in the final dataset. Ls = L. shawi; and Ln = L. naiffi. For details of each strains see Table 1 and Table S1. *Groups defined following the proposal of Tirera et al. [17]; Groups G and H were defined in the present study.
3.2. Variability of LRV1 Diversity across Leishmania Host Species
In addition to the sequences obtained in the present study, another 47 LRV1 sequences from L. braziliensis (n=11), L. guyanensis (n=35) and L. shawi (n=1) strains available on GenBank were included in the analyses (Table S1), corresponding to sequences reported from studies in French Guiana [17], Bolivia [32,33] and Brazil [18,32,33]. Considering the LRV1 sequences analyzed, a higher diversity was observed within LRV1 from L. guyanensis, followed by LRV1 from L. braziliensis, L. naiffi, and L. shawi (Table 2). Of note, few differences were observed between the two LRV1 sequences from L. shawi analyzed despite being isolated from different hosts, in different regions. The high similarity among LRV1 sequences from L. naiffi is also interesting, since the two most divergent sequences correspond to strains from Pará. All the others (n=9), very close related, from Manaus (Amazonas, Brazil), despite of one strain from this group, IOCL 3007, was isolated ten years before the others from a patient who contracted the infection in Manaus, but it was attended in a referral center in Rio de Janeiro. The diversity of LRV1 from L. guyanensis is also depicted in the phylogenetic analyses, clustering these sequences in distinct groups (Figure 1) as previously demonstrated.
3.3. Higher Similarity Is Observed for LRV1 Sequences among Closely Related Leishmania Species
Putting together the results obtained in this study and sequences of LRV1 publicly available, it was possible to compare LRV1 from four different species: L. guyanensis, L. braziliensis, L. naiffi and L. shawi. The analyses between group mean distance reveals that L. shawi and L. guyanensis exhibit lower distances, particularly within Group A, B, and C (Table S2). However, there is considerable diversity among LRV1 from L. guyanensis, with L. braziliensis and L. naiffi showing a significant distance between them (Table 3).
3.4. Host-Specificity Is Clear Observed in the LRV1-L. (Viannia) Species Relationship
To investigate phylogenetic relationships between LRV1 from L. naiffi and L. shawi, along with other 47 LRV1 sequences available (from L. guyanensis, L. braziliensis and only one L. shawi), a Maximum Likelihood (ML) based tree (Figure 3) and phylogenetic network (Figure 4) were constructed. The Maximum Likelihood tree shows several groups with strong support (higher than 70%) and the results are in agreement with previously study [17,18], indicating LRV1 from L. guyanensis grouping in a cluster composed by different subclusters (here named accordingly to Tirera et al. [17]), and only one cluster for LRV1 from L. braziliensis, despite of the variability observed within this group (Figure 3 and Table 2). The new LRV1 sequence obtained for L. shawi clustered together with the other sequence available, keeping a close relationship to the L. guyanensis cluster (Figure 3). Although a clear group is observed for LRV1 sequences from L. naiffi, two strains presented very divergent LRV1 sequences and grouped with other LRV1 from L. naiffi with low bootstrap support, but the most parsimonious connections for these two sequences were observed with LRV1 from L. naiffi (Figure 4). As mentioned before, these two strains are from Pará and all the others are from another, but the same endemic region.
The phylogenetic network shows sequences of clustering similar to those observed with the ML tree. The network however suggests a structure of a common ancestor for all LRV1 (Figure 4). The structure could be the result of low amounts of recombination between sequences of the network, however, representing a bottleneck, or maybe simply due to divergent phylogenetic signals.
4. Discussion
Virus-like particles were demonstrated in Leishmania parasites in the late 1970s [34] but the first molecular description of Leishmania RNA Virus was in 1988, for viruses found in the cytoplasm of a L. (V.) guyanensis strain [35]. Ever since LRV1 has been detected in clinical isolates from Peru [36], Brazil [4,15], Colombia [37], Bolivia [16], Costa Rica [38], French Guiana [39], and more recently in Panamá [19]. These viruses were observed not only in L. guyanensis, but also in strains identified as L. braziliensis, L. shawi, L. naiffi and L. panamensis [4,18,19,20,21,40,41], indicating an old relationship between LRV1 and L. (Viannia) subgenus. Furthermore, LRV1 was detected in clinical samples collected from patients infected by L. lainsoni [4] and L. peruviana [22], but no cultivated strains of these two strains positive for LRV1 are available yet, limiting our analyses. Considering the epidemiological and medical importance of the symbiosis between Leishmania and LRV, there is a distinct requirement to comprehend the variety and spread of the virus within parasite populations.
The coevolution hypothesis for LRV-Leishmania species emerged in 1995 when Widmer and Dooley performed a phylogenetic analyses and found that genetic distances between LRV types mirror the heterogeneity observed for Leishmania species based on random amplified polymorphic DNA (RAPD) fingerprints [42]. More than ten years later a study presenting a genetic characterization and phylogenetic analyses of LRV1 sequences from 27 L. guyanensis strains and two L. braziliensis was published and host-specificity for LRV1 began to be revealed [17]. A year later a robust phylogenetic analyses was presented including 35 LRV1 sequences from L. guyanensis, 11 from L. braziliensis and, for the first time, a sequence of LRV1 found in a L. shawi strain [18]. Both studies presented evidence corroborating the hypothesis of the coevolution of LRV1 and L. (Viannia) parasites, grouping LRV1 sequences according to the parasite species and geographical clustering.
The mentioned above studies suggest a specific relationship of LRV1 to such L. (Viannia) species, combined with the observation of LRV1 in other species, motivate the present study to screen for LRV1 in strains representing L. (Viannia) species not analyzed so far and available at the Leishmania Collection from Fiocruz. To this end, we analyzed all available strains for L. lainsoni, L. lindenbergi, L. naiffi, L. shawi and L. utingensis. As previously demonstrated, LRV1 was detected in L. naiffi and L. shawi strains [18,24], but not in L. lainsoni, despite the fact that LRV1 was previously detected in clinical samples collected from a patient presenting cutaneous leishmaniasis caused by this species [4]. Of note, Leishmania parasites were isolated from this patient and the identified strain was included in our analyses (IOCL 3398), but it was negative for LRV1. There are some possibilities to explain these results, including the possibility of mixed infection by two or more Leishmania species, but with the isolation and growth in culture medium of L. lainsoni to the detriment of another L. (Viannia) species that does not grow very well in culture medium as L. lainsoni [43] Loss of LRV1 during the process of cultivation is also another possibility [44]. Thus, herein we were able to screen LRV1 in different strains from different L. (Viannia) species and to analyze nucleotide sequences of LRV1 from L. naiffi and L. shawi. Interestingly was the fact that more than 50% of L. naiffi strains analyzed were positive for LRV1, but we don't know yet if this symbiotic relationship confers any advance to L. naiffi parasites, such as the capacity of interacting with different sandfly species and/or dispersion in different geographic regions [45]. It has been demonstrated that L. naiffi infection could not have a self-healing nature as it was described years ago [46,47]. Patients could experience a poor response to antimonial or pentamidine therapy [24]. Only in 2019 was first reported a case of a patient infected by L. naiffi carrying LRV1, raising the possibility that the presence of this virus could increase Leishmania spp. virulence and thereby influence therapeutic failure [15], aspects already observed for L. braziliensis and L. guyanensis, but that must be better investigated.
Although it is still important to investigate the specificity of LRV1 to other Leishmania species such as L. panamensis, since LRV1 was already detected in cultivated strains of this species [19,20,21], our results strongly support this kind of relationship, keeping L. shawi in a separate cluster close related to L. guyanensis. Here we assumed the groups suggested by Tirera et al 2017 [17], where L. guyanensis was divided into five subclusters (A-E). The divergence within L. guyanensis is higher than the observed between L. guyanensis and L. shawi, corroborating the assumption of these as a complex of species [23]. Following this, LRV1 sequences from L. shawi formed another subcluster, named here as Group G, closely related to the L. guyanensis subclusters A and B and C. Although LRV1 sequences were obtained for only two L. shawi strains so far, the fact that they clustered together, despite the strains being from different geographic regions, is also an important aspect to support host-specificity for LRV1. The diversity of LRV1 from L. guyanensis, forming several subclusters, must be better explored, but the number of LRV1 sequences from L. guyanensis analyzed and the geographic dispersion of these parasites in the Amazon region might be contributing to this result.
Host specificity was also observed for LRV1 genotypes from L. naiffi. The phylogenetic tree and the NeighborNet (Figs 3 and 4) show that LRV1 sequences from L. naiffi clustered in a well-supported monophyletic clade. Of note, most of LRV1 sequences from L. naiffi analyzed were very similar. The diversity observed within LRV1 sequences from the same species must be further investigated, but it is important to consider that all but one L. naiffi strains presenting highly similar LRV1 were obtained from patients infected in the same endemic region and included in the same study, suggesting a possibility of problems during laboratory manipulation. However, this very similar group contained one strain isolated previously to the mentioned study and was not manipulated together with the other strains, suggesting a homogeneity for the L. naiffi population circulating in this area and causing human disease, which can represent an epidemic clone. LRV1 sequences obtained for two L. naiffi strains, IOCL 3316 and IOCL 854 strains, both isolated in Pará, showed a different phylogenetic pattern from the above-mentioned L. naiffi strains isolated from Manaus, Amazonas. The strain IOCL 854 was obtained from a sandfly species, Lutzomyia squamiventris, and LRV1 from this strain showed a basal position into the L. naiffi clade, despite the close relationship of this strain to other L. naiffi, including IOCL 3007, as previously demonstrated [23,48]. Considering Leishmania (Viannia) species depicted by microsatellite analyses, it is more expected that parasites from populations circulating in the Amazon basin (POP2 and POP3 after Kuhls et al. [48]) carries LRV1 and each subpopulation has an association with specific LRV1 genotypes.
By analyzing LRV1 sequences from several strains representing different L. (Viannia) species we demonstrate that LRV1 genotypes form distinct clusters corresponding to their Leishmania species host, suggesting that the transfer of viral particles between strains from different species does not occur frequently. Altogether, our results reinforce the concordance between the phylogenetic patterns of LRV1 and Leishmania (Viannia) species, providing support for the prevailing hypothesis that LRV1 is an ancient virus that has undergone co-evolution with their hosts [42,49]. Recently it was shown that parasite hybridization might explain the high occurrence of the symbiotic interaction of LRV1 with L. braziliensis in Peru and Bolivia [50]. It is possible that this also explains the high frequency of LRV1 in parasites from the Brazilian Amazon Region, since many possible hybrids were described in the region [51], and analyses of microsatellite markers have shown extensive diversity in the subgenus L. (Viannia), with an indication of both clonality and recombination as a strategy of reproduction [48].
5. Conclusions
Our study adds to the growing body of evidence supporting a specific relationship between Leishmania RNA Virus 1 (LRV1) and species within the L. (Viannia) subgenus. LRV1 has been detected in various clinical isolates from different species, including L. guyanensis, L. braziliensis, L. shawi, L. naiffi, and L. panamensis, but not in other species as well as not in L. braziliensis circulating outside the Amazon Basin, raising intriguing questions about host specificity and the potential impact on virulence and therapeutic response.
While the results provide strong support for the association of LRV1 with specific Leishmania species, more investigations are needed to understand its specificity to other species, such as L. panamensis. The divergence observed within LRV1 sequences from the same species warrants further scrutiny, especially considering potential issues during laboratory manipulation and the homogeneity of L. naiffi populations in certain endemic regions. Overall, the identification and characterization of LRV1 in different Leishmania species shed light on the complex interactions between these viruses and the parasites they infect. Future research in this area may uncover novel insights into the biology and pathogenesis of Leishmania infections, offering new perspectives on therapeutic strategies and disease management.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Information on LRV1 sequences from previous studies used for phylogenetic analyses; Table S2: Estimates of average evolutionary divergence over sequence pairs within groups and between groups using Tamura 3-parameter model and number of differences.
Author Contributions
Conceptualization, Elisa Cupolillo, Khaled Chourabi, Lilian Cantanhêde and Mayara Santana; Formal analyses, Elisa Cupolillo and Mayara Santana; Funding acquisition, Elisa Cupolillo; Methodology, Khaled Chourabi and Mayara Santana; Writing – original draft, Mayara Santana and Lilian Cantanhêde; Writing – review & editing, Elisa Cupolillo, Khaled Chourabi and Lilian Cantanhêde. All authors have read and agreed to the published version of the manuscript.
Funding
Elisa Cupolillo: (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Finance Code 001; Conselho Nacional de Desenvolvimento Científico e Tecnológico, Research Fellow, 302622/2017-9; Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro, CNE, E26-202.569/2019; Temáticos, E26-210.038/2020; Fundação de Amparo ao Desenvolvimento das Ações Científicas e Tecnológicas e à Pesquisa-FAPERO, Edital 16/2014). Lilian Motta Cantanhêde: (Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro, Pós-Doutorado Nota 10 E-26/205.730/2022 and 205.731/2022).
Data Availability Statement
The graphical abstract was created using BioRender (www.biorender.com).
Acknowledgments
We sincerely appreciate the Leishmania collection (CLIOC) from Oswaldo Cruz Institute, for providing all Leishmania strains analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization Control of the Leishmaniases. 2010.
- OPAS, O. Situação Epidemiológica Leishmaniose Cutânea e Mucosa; 2021.
- Zamora, M.; Stuart, K.; Salinas, G.; Saravia, N. Leishmania RNA Viruses in Leishmania of the Viannia Subgenus. Am J Trop Med Hyg 1996, 54, 425–429. [Google Scholar] [CrossRef]
- Cantanhêde, L.M.; da Silva Júnior, C.F.; Ito, M.M.; Felipin, K.P.; Nicolete, R.; Salcedo, J.M.V.; Porrozzi, R.; Cupolillo, E.; Ferreira, R. de G.M. Further Evidence of an Association between the Presence of Leishmania RNA Virus 1 and the Mucosal Manifestations in Tegumentary Leishmaniasis Patients. PLoS Negl Trop Dis 2015, 9, e0004079. [Google Scholar] [CrossRef] [PubMed]
- Ramasawmy, R.; Menezes, E.; Magalhães, A.; Oliveira, J.; Castellucci, L.; Almeida, R.; Rosa, M.E.A.; Guimarães, L.H.; Lessa, M.; Noronha, E.; et al. The -2518bp Promoter Polymorphism at CCL2/MCP1 Influences Susceptibility to Mucosal but Not Localized Cutaneous Leishmaniasis in Brazil. Infection, Genetics and Evolution 2010, 10, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Castellucci, L.; Jamieson, S.E.; Miller, E.N.; De Almeida, L.F.; Oliveira, J.; Magalhães, A.; Guimarães, L.H.; Lessa, M.; Lago, E.; De Jesus, A.R.; et al. FLI1 Polymorphism Affects Susceptibility to Cutaneous Leishmaniasis in Brazil. Genes Immun 2011, 12, 589–594. [Google Scholar] [CrossRef]
- Silveira, F.T.; Lainson, R.; Corbett, C.E.P. Clinical and Immunopathological Spectrum of American Cutaneous Leishmaniasis with Special Reference to the Disease in Amazonian Brazil - A Review. Mem Inst Oswaldo Cruz 2004, 99, 239–251. [Google Scholar] [CrossRef]
- Alvar, J.; Croft, S.; Olliaro, P. Chemotherapy in the Treatment and Control of Leishmaniasis. Adv Parasitol 2006, 61, 223–274. [Google Scholar] [CrossRef]
- Ives, A.; Ronet, C.; Prevel, F.; Ruzzante, G.; Fuertes-Marraco, S.; Schutz, F.; Zangger, H.; Revaz-Breton, M.; Lye, L.-F.; Hickerson, S.M.; et al. Leishmania RNA Virus Controls the Severity of Mucocutaneous Leishmaniasis. Science (1979) 2011, 331, 775–778. [Google Scholar] [CrossRef]
- Hartley, M.A.; Ronet, C.; Zangger, H.; Beverley, S.M.; Fasel, N. Leishmania RNA Virus: When the Host Pays the Toll. Front Cell Infect Microbiol 2012, 2, 99. [Google Scholar] [CrossRef]
- Goyal, G.; Phukan, A.C.; Hussain, M.; Lal, V.; Modi, M.; Goyal, M.K.; Sehgal, R. Virion Structure of Leishmania RNA Virus 1. J Neurol Sci 2019, 116544. [Google Scholar] [CrossRef]
- Rossi, M.; Castiglioni, P.; Hartley, M.A.; Eren, R.O.; Prével, F.; Desponds, C.; Utzschneider, D.T.; Zehn, D.; Cusi, M.G.; Kuhlmann, F.M.; et al. Type I Interferons Induced by Endogenous or Exogenous Viral Infections Promote Metastasis and Relapse of Leishmaniasis. Proc Natl Acad Sci U S A 2017, 114, 4987–4992. [Google Scholar] [CrossRef]
- de Carvalho, R.V.H.; Lima-Junior, D.S.; da Silva, M.V.G.; Dilucca, M.; Rodrigues, T.S.; Horta, C. V.; Silva, A.L.N.; da Silva, P.F.; Frantz, F.G.; Lorenzon, L.B.; et al. Leishmania RNA Virus Exacerbates Leishmaniasis by Subverting Innate Immunity via TLR3-Mediated NLRP3 Inflammasome Inhibition. Nat Commun 2019, 10, 5273. [Google Scholar] [CrossRef]
- Bourreau, E.; Ginouves, M.; Prévot, G.; Hartley, M.-A.; Gangneux, J.-P.; Robert-Gangneux, F.; Dufour, J.; Marie, D. Sainte; Bertolotti, A.; Pratlong, F.; et al. Leishmania-RNA Virus Presence in L. Guyanensis Parasites Increases the Risk of First-Line Treatment Failure and Symptomatic Relapse. J Infect Dis 2015, 1–28. [Google Scholar] [CrossRef]
- Vieira-Gonçalves, R.; Fagundes-Silva, G.A.; Heringer, J.F.; Fantinatti, M.; Da-Cruz, A.M.; Oliveira-Neto, M.P.; Guerra, J.A.O.; Gomes-Silva, A. First Report of Treatment Failure in a Patient with Cutaneous Leishmaniasis Infected by Leishmania (Viannia) Naiffi Carrying Leishmania RNA Virus: A Fortuitous Combination? Rev Soc Bras Med Trop 2019, 52, 10–12. [Google Scholar] [CrossRef]
- Adaui, V.; Lye, L.-F.; Akopyants, N.S.; Zimic, M.; Llanos-Cuentas, A.; Garcia, L.; Maes, I.; De Doncker, S.; Dobson, D.E.; Arevalo, J.; et al. Association of the Endobiont Double-Stranded RNA Virus LRV1 With Treatment Failure for Human Leishmaniasis Caused by Leishmania Braziliensis in Peru and Bolivia. J Infect Dis 2015, 1–32. [Google Scholar] [CrossRef]
- Tirera, S.; Ginouves, M.; Donato, D.; Caballero, I.S.; Bouchier, C.; Lavergne, A.; Bourreau, E.; Mosnier, E.; Vantilcke, V.; Couppié, P.; et al. Unraveling the Genetic Diversity and Phylogeny of Leishmania RNA Virus 1 Strains of Infected Leishmania Isolates Circulating in French Guiana. PLoS Negl Trop Dis 2017, 11, 1–20. [Google Scholar] [CrossRef]
- Cantanhêde, L.M.; Fernandes, F.G.; Ferreira, G.E.M.; Porrozzi, R.; Ferreira, R. de G.M.; Cupolillo, E. New Insights into the Genetic Diversity of Leishmania RNA Virus 1 and Its Species-Specific Relationship with Leishmania Parasites. PLoS One 2018, 13, e0198727. [Google Scholar] [CrossRef]
- Gonzalez, K.; De León, S.S.; Pineda, V.; Samudio, F.; Capitan-Barrios, Z.; Suarez, J.A.; Weeden, A.; Ortiz, B.; Rios, M.; Moreno, B.; et al. Detection of Leishmania RNA Virus 1 in Leishmania (Viannia) Panamensis Isolates, Panama. Emerg Infect Dis 2023, 29. [Google Scholar] [CrossRef]
- Kariyawasam, R.; Mukkala, A.N.; Lau, R.; Valencia, B.M.; Llanos-Cuentas, A.; Boggild, A.K. Virulence Factor RNA Transcript Expression in the Leishmania Viannia Subgenus: Influence of Species, Isolate Source, and Leishmania RNA Virus-1. Trop Med Health 2019, 47, 25. [Google Scholar] [CrossRef]
- Kariyawasam, R.; Grewal, J.; Lau, R.; Purssell, A.; Valencia, B.M.; Llanos-Cuentas, A.; Boggild, A.K. Influence of Leishmania RNA Virus 1 on Proinflammatory Biomarker Expression in a Human Macrophage Model of American Tegumentary Leishmaniasis. J Infect Dis 2017, 216, 877–886. [Google Scholar] [CrossRef]
- Valencia, B.M.; Lau, R.; Kariyawasam, R.; Jara, M.; Ramos, A.P.; Chantry, M.; Lana, J.T.; Boggild, A.K.; Llanos-Cuentas, A. Leishmania RNA Virus-1 Is Similarly Detected among Metastatic and Non-Metastatic Phenotypes in a Prospective Cohort of American Tegumentary Leishmaniasis. PLoS Negl Trop Dis 2022, 16, e0010162. [Google Scholar] [CrossRef]
- Boité, M.C.; Mauricio, I.L.; Miles, M.A.; Cupolillo, E. New Insights on Taxonomy, Phylogeny and Population Genetics of Leishmania (Viannia) Parasites Based on Multilocus Sequence Analyses. PLoS Negl Trop Dis 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Fagundes-Silva, G.A.; Sierra Romero, G.A.; Cupolillo, E.; Gadelha Yamashita, E.P.; Gomes-Silva, A.; De Oliveira Guerra, J.A.; Da-Cruz, A.M. Leishmania (Viannia) Naiffi: Rare Enough to Be Neglected? Mem Inst Oswaldo Cruz 2015, 110, 797–800. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R.; Braga, R.R.; De Souza, A.A.; Pôvoa, M.M.; Ishikawa, E.A.; Silveira, F.T. Leishmania (Viannia) Shawi Sp. n., a Parasite of Monkeys, Sloths and Procyonids in Amazonian Brazil. Ann Parasitol Hum Comp 1989, 64, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R. The Neotropical Leishmania Species: A Brief Historical Review of Their Discovery, Ecology and Taxonomy. Rev Panamazonica Saude 2010, 1. [Google Scholar] [CrossRef]
- Oliveira, L.P.; Nascimento, L.C.S.; Santos, F.S.; Takamatsu, J.L.C.; Sanchez, L.R.P.; Santos, W.S.; Garcez, L.M. First Report of an Asymptomatic Leishmania (Viannia) Shawi Infection Using a Nasal Swab in Amazon, Brazil. Int J Environ Res Public Health 2022, 19. [Google Scholar] [CrossRef]
- da Graça, G.C.; Volpini, A.C.; Romero, G.A.S.; Neto, M.P. de O.; Hueb, M.; Porrozzi, R.; Boité, M.C.; Cupolillo, E. Development and Validation of PCR-Based Assays for Diagnosis of American Cutaneous Leishmaniasis and Identification of the Parasite Species. Mem Inst Oswaldo Cruz 2012, 107, 664–674. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analyses Program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analyses across Computing Platforms. Mol Biol Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol Biol Evol 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Brettmann, E.A.; Shaik, J.S.; Zangger, H.; Lye, L.F.; Kuhlmann, F.M.; Akopyants, N.S.; Oschwald, D.M.; Owens, K.L.; Hickerson, S.M.; Ronet, C.; et al. Tilting the Balance between RNA Interference and Replication Eradicates Leishmania RNA Virus 1 and Mitigates the Inflammatory Response. Proc Natl Acad Sci U S A 2016, 113, 11998–12005. [Google Scholar] [CrossRef]
- Parmentier, L.; Cusini, A.; Müller, N.; Zangger, H.; Hartley, M.A.; Desponds, C.; Castiglioni, P.; Dubach, P.; Ronet, C.; Beverley, S.M.; et al. Case Report: Severe Cutaneous Leishmaniasis in a Human Immunodeficiency Virus Patient Coinfected with Leishmania Braziliensis and Its Endosymbiotic Virus. American Journal of Tropical Medicine and Hygiene 2016, 94, 840–843. [Google Scholar] [CrossRef] [PubMed]
- Croft, S.L.; Molyneux, D.H. Studies on the Ultrastructure, Virus-like Particles and Infectivity of Leishmania Hertigi. Ann Trop Med Parasitol 1979, 73, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Aline, R.F.; Smiley, B.L.; Scholler, J.; Keithly, J.; Stuart, K. LR1: A Candidate RNA Virus of Leishmania. Proc Natl Acad Sci U S A 1988, 85, 9572–9275. [Google Scholar] [CrossRef]
- Saiz, M.; Llanos-Cuentas, A.; Echevarria, J.; Roncal, N.; Cruz, M.; Tupayachi Muniz, M.; Lucas, C.; Wirth, D.F.; Scheffter, S.; Magill, A.J.; et al. Short Report: Detection of Leishmaniavirus in Human Biopsy Samples of Leishmaniasis from Peru. American Journal of Tropical Medicine and Hygiene 1998, 58, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Salinas, G.; Zamora, M.; Stuart, K.; Saravia, N. Leishmania RNA Viruses in Leishmania of the Viannia Subgenus. Am J Trop Med Hyg 1996, 54, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Mata-somarribas, C.; Quesada-lópez, J.; Matamoros, M.F.; Cervantes-gómez, C. Raising the Suspicion of a Non-Autochthonous Infection : Identification of Leishmania Guyanensis from Costa Rica Exhibits a Leishmaniavirus Related to Brazilian North-East and French Guiana Viral Genotypes. 2022, 117, 1–7. [Google Scholar] [CrossRef]
- Couppié, P.; Nacher, M.; Demar, M.; Ginouvès, M.; Simon, S.; Prévot, G.; Bourreau, E.; Ronet, C.; Lacoste, V. Prevalence and Distribution of Leishmania RNA Virus 1 in Leishmania Parasites from French Guiana. Am J Trop Med Hyg 2016, 94, 102–106. [Google Scholar] [CrossRef]
- Ito, M.M.; Catanhêde, L.M.; Katsuragawa, T.H.; da Silva Junior, C.F.; Camargo, L.M.A.; Mattos, R. de G.; Vilallobos-Salcedo, J.M. Correlation between Presence of Leishmania RNA Virus 1 and Clinical Characteristics of Nasal Mucosal Leishmaniosis. Braz J Otorhinolaryngol 2015, 81, 533–540. [Google Scholar] [CrossRef]
- de Oliveira Ramos Pereira, L.; Maretti-Mira, A.C.; Rodrigues, K.M.; Lima, R.B.; de Oliveira-Neto, M.P.; Cupolillo, E.; Pirmez, C.; de Oliveira, M.P. Severity of Tegumentary Leishmaniasis Is Not Exclusively Associated with Leishmania RNA Virus 1 Infection in Brazil. Mem Inst Oswaldo Cruz 2013, 108, 665–667. [Google Scholar] [CrossRef]
- Widmer, G.; Dooley, S. Phylogenetic Analyses of Leishmania RNA Virus and Leishmania Suggests Ancient Virus-Parasite Association. Nucleic Acids Res 1995, 23, 2300–2304. [Google Scholar] [CrossRef]
- das Chagas, B.D.; Pereira, T.M.; Cantanhêde, L.M.; da Silva, G.P.; Boité, M.C.; Pereira, L. de O.R.; Cupolillo, E. Interspecies and Intrastrain Interplay among Leishmania Spp. Parasites. Microorganisms 2022, 10, 1883. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, F.M.; Robinson, J.I.; Bluemling, G.R.; Ronet, C.; Fasel, N.; Beverley, S.M. Antiviral Screening Identifies Adenosine Analogs Targeting the Endogenous DsRNA Leishmania RNA Virus 1 (LRV1) Pathogenicity Factor. Proceedings of the National Academy of Sciences 2017, 114. [Google Scholar] [CrossRef] [PubMed]
- Cantanhêde, L.M.; Cupolillo, E. Leishmania (Viannia) Naiffi Lainson & Shaw 1989. Parasit Vectors 2023, 16. [Google Scholar]
- Naiff, R.D.; Freitas, R.A.; Naiff, M.F.; Arias, J.R.; Barrett, T. V.; Momen, H.; Grimaldi Júnior, G. Epidemiological and Nosological Aspects of Leishmania Naiffi Lainson & Shaw, 1989. Mem Inst Oswaldo Cruz 1991, 86, 317–321. [Google Scholar] [PubMed]
- Van Der Snoek, E.M.; Lammers, A.M.; Kortbeek, L.M.; Roelfsema, J.H.; Bart, A.; Jaspers, C.A.J.J. Spontaneous Cure of American Cutaneous Leishmaniasis Due to Leishmania Naiffi in Two Dutch Infantry Soldiers. Clin Exp Dermatol 2009, 34, 889–891. [Google Scholar] [CrossRef]
- Kuhls, K.; Cupolillo, E.; Silva, S.O.; Schweynoch, C.; Côrtes Boité, M.; Mello, M.N.; Mauricio, I.; Miles, M.; Wirth, T.; Schönian, G. Population Structure and Evidence for Both Clonality and Recombination among Brazilian Strains of the Subgenus Leishmania (Viannia). PLoS Negl Trop Dis 2013, 7, e2490. [Google Scholar] [CrossRef]
- Cantanhêde, L.M.; Mata-Somarribas, C.; Chourabi, K.; Pereira da Silva, G.; Dias Das Chagas, B.; de Oliveira, R. Pereira, L.; Côrtes Boité, M.; Cupolillo, E. The Maze Pathway of Coevolution: A Critical Review over the Leishmania and Its Endosymbiotic History. Genes (Basel) 2021, 12. [Google Scholar] [CrossRef]
- Heeren, S.; Maes, I.; Sanders, M.; Lye, L.-F.; Arevalo, J.; Garcia, L.; Lemey, P.; Beverley, S.M.; Cotton, J.A.; Dujardin, J.-C.; et al. Parasite Hybridization Promotes Spreading of Endosymbiotic Viruses. [CrossRef]
- Tojal da Silva, A.C.; Cupolillo, E.; Volpini, A.C.; Almeida, R.; Sierra Romero, G.A. Species Diversity Causing Human Cutaneous Leishmaniasis in Rio Branco, State of Acre, Brazil. Tropical Medicine and International Health 2006, 11, 1388–1398. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the genome of LRV1 showing the annealing region for each primer used for screening (240bp) and phylogenetic analyses (850bp).
Figure 1.
Schematic representation of the genome of LRV1 showing the annealing region for each primer used for screening (240bp) and phylogenetic analyses (850bp).
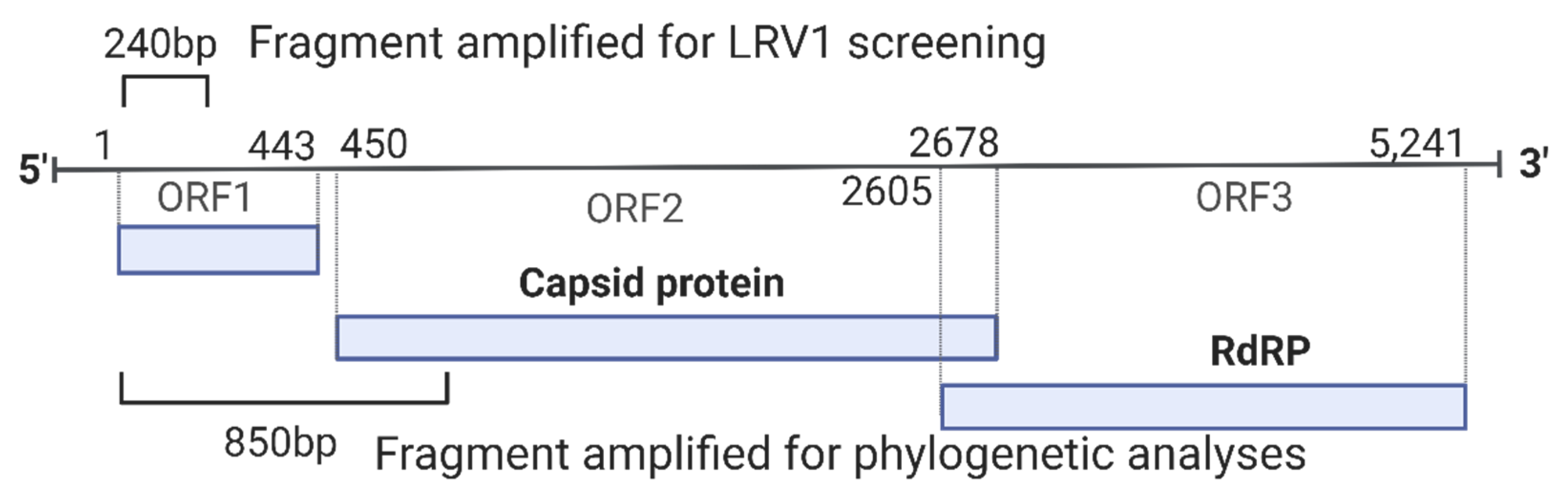
Figure 4.
NeighborNet showing the relationship among LRV1 sequences from different Leishmania (Viannia) species. The network was computed using SplitsTree software. Text colors refer to each group analyzed. Pink = L. guyanensis; purple = L. shawi; blue = L. naiffi and green = L. braziliensis. Strains 854 and 3316, belonging to the species of L. naiffi, presented a divergent profile from the other strains of the same group.
Figure 4.
NeighborNet showing the relationship among LRV1 sequences from different Leishmania (Viannia) species. The network was computed using SplitsTree software. Text colors refer to each group analyzed. Pink = L. guyanensis; purple = L. shawi; blue = L. naiffi and green = L. braziliensis. Strains 854 and 3316, belonging to the species of L. naiffi, presented a divergent profile from the other strains of the same group.
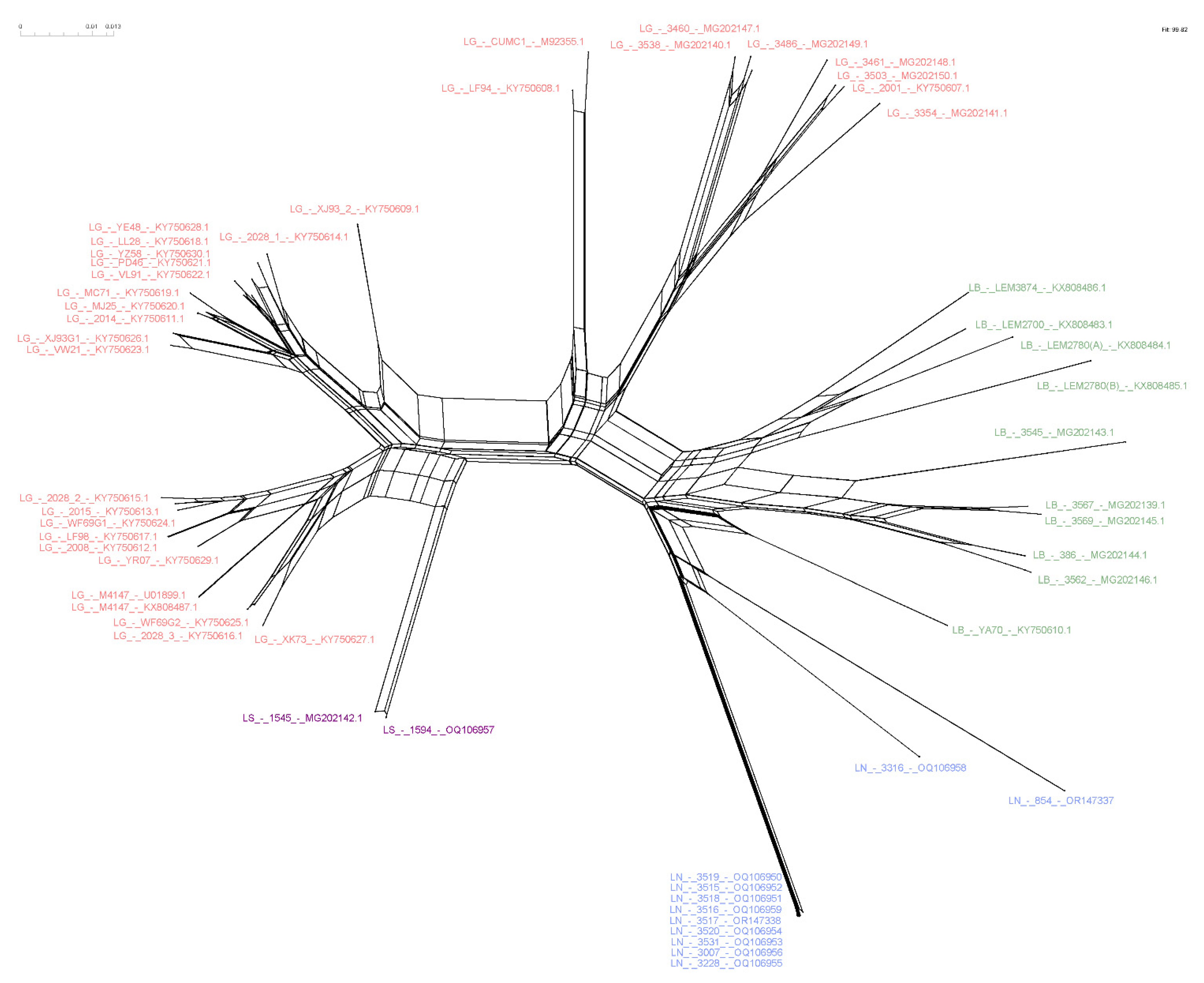

Table 1.
Leishmania (Viannia) strains from different species screened for the presence of the viral endosymbiont LRV1 and information on accession for LRV1 sequences obtained for the positive strains.
Table 1.
Leishmania (Viannia) strains from different species screened for the presence of the viral endosymbiont LRV1 and information on accession for LRV1 sequences obtained for the positive strains.
| Leishmania Strain ID (IOCL) | Parasite Species | Leishmania International Code | Geographic Origin (City, State) | Sequence Length | GenBank Accession Number |
|---|---|---|---|---|---|
| 854 | L. naiffi | ISQU/BR/1985/IM2264 | Cachoeira Porteira, Pará | 759 | OR147337 |
| 3007 | L. naiffi | MHOM/BR/2003/IRCF | Manaus, Amazonas | 701 | OQ106956 |
| 3228 | L. naiffi | MHOM/BR/2010/MS | Manaus, Amazonas | 699 | OQ106955 |
| 3316 | L. naiffi | MHOM/BR/2011/58-AMS | Mojuí dos Campos, Pará | 706 | OQ106958 |
| 3515 | L. naiffi | MHOM/BR/2013/49UAS | Manaus, Amazonas | 709 | OQ106952 |
| 3516 | L. naiffi | MHOM/BR/2013/63DDL | Manaus, Amazonas | 799 | OQ106959 |
| 3517 | L. naiffi | MHOM/BR/2013/65HCC | Manaus, Amazonas | 710 | OR147338 |
| 3518 | L. naiffi | MHOM/BR/2013/66CPS | Manaus, Amazonas | 679 | OQ106951 |
| 3519 | L. naiffi | MHOM/BR/2013/51FRS | Manaus, Amazonas | 660 | OQ106950 |
| 3520 | L. naiffi | MHOM/BR/2013/62FJFM | Manaus, Amazonas | 632 | OQ106954 |
| 3531 | L. naiffi | MHOM/BR/2013/56EGP | Manaus, Amazonas | 708 | OQ106953 |
| 991 | L. naiffi | MDAS/BR/1987/IM3307 | São Félix do Xingu, Pará | - | --- |
| 992 | L. naiffi | MDAS/BR/1987/IM3280 | São Félix do Xingu, Pará | - | --- |
| 993 | L. naiffi | MDAS/BR/1987/IM3281 | São Félix do Xingu, Pará | - | --- |
| 1123 | L. naiffi | MHOM/BR/1986/IM2736 | Manaus, Amazonas | - | --- |
| 1365 | L. naiffi | MDAS/BR/1979/M5533 | Almeirim, Pará | - | --- |
| 3310 | L. naiffi | MHOM/BR/2011/S50 | Santarém, Pará | - | --- |
| 3541 | L. naiffi | MHOM/BR/2014/61AAM | Manaus, Amazonas | - | --- |
| 1594 | L. shawi | MHOM/BR/1990/IM2842 | Manaus, Amazonas | 781 | OQ106957 |
| 1067 | L. shawi | IWHI/BR/1985/IM2324 | Tucuruí, Pará | - | --- |
| 1068 | L. shawi | IWHI/BR/1985/IM2326 | Tucuruí, Pará | - | --- |
| 3481 | L. shawi | MHOM/BR/2013/18 | Manaus, Amazonas | - | --- |
| 1023 | L. lainsoni | MHOM/BR/1981/M6426 | Benevides, Pará | - | --- |
| 1266 | L. lainsoni | MCUN/BR/1983/IM1721 | Tucuruí, Pará | - | --- |
| 2497 | L. lainsoni | MHOM/BR/2002/NMT-RBO 027P | Rio Branco, Acre | - | --- |
| 3398 | L. lainsoni | MHOM/BR/2012/AP60A | Porto Velho, Rondônia | - | --- |
| 2690 | L. lindenbergi | MHOM/BR/1966/M15733 | Belém, Pará | - | --- |
| 3645 | L. lindenbergi | MHOM/BR/2015/RO514 | Porto Velho, Rondônia | - | --- |
| 3746 | L. lindenbergi | MHOM/BR/2014/RO285 | Porto Velho, Rondônia | - | --- |
| 2689 | L. utingensis | ITUB/BR/1977/M4964 | Belém, Pará | - | --- |
In gray are highlighted the negative strains for LRV1. MHOM = Mammalia, Homo sapiens; ISQU = Insecta, Lutzomyia squamiventris; MDAS = Mammalia, Dasypus sp.; IWHI = Insecta, Lutzomyia whitmani.
Table 2.
Estimates of average evolutionary divergence, based on number of differences and Tamura-3-parameter, over sequence pairs within mean groups.
Table 2.
Estimates of average evolutionary divergence, based on number of differences and Tamura-3-parameter, over sequence pairs within mean groups.
| L. guyanensis | L. braziliensis | L. naiffi | L. shawi | |
|---|---|---|---|---|
| Number of differences | 448.96 | 295.15 | 31.75 | 7 |
| Tamura-3-parameter model | 0.18 | 0.16 | 0.06 | 0.01 |
Table 3.
Estimates of evolutionary divergence over sequence pairs between groups based on Tamura 3-parameter model (below diagonal) and on number of differences (above diagonal).
Table 3.
Estimates of evolutionary divergence over sequence pairs between groups based on Tamura 3-parameter model (below diagonal) and on number of differences (above diagonal).
| L. naiffi | L. shawi | L. guyanensis | L. braziliensis | |
|---|---|---|---|---|
| L. naiffi | - | 109 | 109 | 100 |
| L. shawi | 0.208 | - | 88 | 114 |
| L. guyanensis | 0.214 | 0.147 | - | 550 |
| L. braziliensis | 0.190 | 0.204 | 0.260 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



