
Submitted:
07 August 2023
Posted:
09 August 2023
You are already at the latest version
Abstract
Ageing is associated with a decline in both Nrf2 activity and DNA repair efficiency, leading to the accumulation of DNA damage and an increased risk of cancer. Understanding the mechanisms behind the increased level of damaged DNA during ageing is crucial for developing interventions to mitigate age-related cancer risk and improve overall health in older individuals. (Poly)phenols are plant-derived bioactive compounds that undergo metabolism into simpler phenolic catabolites following ingestion. While these compounds have been associated with various health benefits, further analysis of their bioavailable catabolites is necessary. To this end, four phenolic catabolites derived from colonic microbiota were investigated for their ability to reduce DNA damage in response to oxidative challenge and modulate the Nrf2-Antixoidant Response Element pathway. Normal and adenocarcinoma colonocyte cell lines (CCD 841 CoN, & HT29 respectively) were exposed to sub cytotoxic concentrations (>10 µM,) of hydroxylated (4-hydroxybenzoic acid, 3-(3’-hydroxyphenyl) propionic acid) and non-hydroxylated (benzoic acid or 3-phenylpropionic acid) phenolic acids. Using the COMET assay, each catabolite demonstrated significant (p<0.001) geno-protective activity, albeit with varying potency. They also significantly modulated expression of key genes in the Nrf2-ARE pathway, including Nrf2, Heme oxygenase-1 (HO-1), and NAD(P)H dehydrogenase quinone 1 (NQO1). Hydroxylation of the phenolic acids appeared to have a limited effect with respect to bioactivity. Overall, the colon derived phenolic metabolites, when assessed at physiologically relevant concentrations, reduced DNA damage in both normal and adenocarcinoma colonic cells in response to oxidative challenge, mediated – at least in part via upregulation of the Nrf2-ARE pathway.
Keywords:
3-(3′-Hydroxyphenyl)propanoic acid
; 3-(Phenyl)propanoic acid
; 4-Hydroxybenzoic acid
; Benzoic acid
; Colonic cells
; COMET assay
; Nrf2 pathway
; Phenolic catabolites
1. Introduction
The association between ageing and tumorigenesis is well-established, with lifetime accumulation of DNA damage recognised as a significant contributing factor [1,2]. As individuals age, the progressive accumulation of DNA damage within tissues becomes increasingly evident, potentially triggering either the DNA repair mechanism apoptosis or cell senescence which is a contributing factor to the decline in tissue function observed with age [3,4]. In addition, the efficiency of DNA repair mechanisms declines over time, leading to the persistence of DNA damage within the genome as age advances [4-6]. This accumulation of DNA damage is believed to directly contribute to the observed increase in cancer prevalence among older individuals [1]. Expanding our knowledge of the mechanisms involved in DNA damage accumulation during ageing could provide valuable insights into this process and allow the development of rational interventions to mitigate age-related cancer risk and improve overall health in the ageing population.
Accumulating research suggests that (poly)phenols, a complex and diverse group of bioactive aromatic phytochemicals with up to 10,000 different known structures, may play a role in reducing DNA damage [7,8]. Flavonoids are the most abundant in our diet [9,10,11] and are present at high concentrations in foods such as berries, coffee, and cocoa [12], where they contribute to the estimated average intake of 580 – 1780 mg of (poly)phenols per day [13].
Over the last decades, advances in the sensitivity and selectivity of analytical techniques have facilitated significant improvements in our understanding of the metabolism and bioactivity potential of (poly)phenols [8,14,15,16]. Moreover, a paradigm shift in thinking as occurred upon the recognition that up to 95% of ingested (poly)phenols escape digestion in the stomach and small intestine, becoming biologically available only after considerable host and microbiota mediate catalytic transformation, which often also impacts on their bioactivity [12] (add reference from Del Rio). The focus now is on the kinetics and biological activities of (poly)phenol catabolites rather than the native structures found in plant foods themselves. Upon consumption, (poly)phenols undergo numerous structural changes during their transit through the gastrointestinal tract (GIT), including the production of bioactive, microbiota-derived phenolic catabolites [17], some of which have been reported to exert pleiotropic effects that likely contribute to reducing the risk for chronic diseases [12,18,19,20,21,22]. However, the specific sites and mechanisms of action remain elusive, with multiple pathways postulated and currently under investigation. For example, the ability of (poly)phenolic-rich foods to affect aspects of the colorectal carcinogenesis pathway in gut epithelial cells has been studied in vitro [18,23,24]. The microbiota-derived catabolities of dietary (poly)phenols have been reported to modulate the response of pathways involved in the cell cycle and apoptosis, by inducing variation in expression of genes such as Nuclear factor erythroid 2-related factor 2 (Nrf2), phorbol-12-myristate-13-acetate-induced protein 1 (PMAIP1), or BH3-interacting domain death agonist (BID) [15,25,26,27]. Of particular interest is Nrf2, a redox-sensitive transcription factor master regulator of oxidoreductive and immune homeostasis. Under normal conditions cytosolic Nrf2 is associated with the inhibitory protein Kelch-like ECH-associated Protein 1(Keap1), which acts as a sensor of endogenous and exogenous prooxidants [28]. Under a mild increase in oxidative stress, specific cysteine residues in Keap1 allow newly synthesized Nrf2 to escape Keap1-mediated ubiquitination and to activate the transcription of Nrf2-target genes. The protein then dimerizes with small Maf proteins and binds to the anti-oxidant response element (ARE) in the promoter region of these genes, activating their transcription and eliciting anti-oxidant defence and detoxification mechanisms [14]. Consequently, Nrf2 represents an important potential therapeutic target in human carcinogenesis [29] as it is activated by a range of diet derived (poly)phenolics [30]. Moreover, ageing is associated with a decline in Nrf2 activity and signalling, resulting in a compromised cellular response to DNA damage [31]. A comprehensive review by Zhou et al. highlights the importance of (poly)phenols as activators of KEAP1-NRF2 signalling pathway [31], with many (poly)phenolic-rich berry fractions and to a lesser degree, their colonic catabolites reported to reduce DNA damage in various cell models [33,34,35,36].
Dobani and colleagues recently reported an ileostomate-based ex vivo model in which 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA), 3-(phenyl)propanoic acid (3PPA), 4-hydroxybenzoic acid (4HBA), and benzoic acid (BA) were commonly produced as colonic catabolites following ingestion of raspberries, although they are not uniquely derived from that dietary source [16]. Consequently, we investigated the differential effects of these hydroxylated and non-hydroxylated catabolites in two contrasting colonic cell lines with greater and lesser degrees of transformation (HT29 & CCD 841 CoN cells), with respect to DNA damage and ability to modulate key genes associated with the cytoprotective Nrf2-ARE pathway.
2. Material and methods
2.1. Chemicals and reagents
Foetal Bovine Serum (FBS), Minimum Essential Media (MEM), Dulbecco’s Modified Eagle’s Medium (DMEM), and phosphate-buffered saline (PBS) were purchased from Gibco Life Technologies Ltd (Paisley, Scotland, UK). All remaining chemicals including the phenolic catabolites 4HBA, BA, 3'HPPA, and 3PPA were acquired from Sigma-Aldrich (Poole, Dorset, UK) unless otherwise stated.
2.2. Tissue culture
Both normal epithelial (CCD 841 CoN) and adenocarcinoma (HT29) colonic cells used for this investigation were acquired from the American Type Culture Collection (ATCC) and European Collection of Cell Cultures (ECACC), respectively. CCD 841 CoN cells were cultured in MEM supplemented with 10% FBS, 100 U/L penicillin/streptomycin, 1% sodium pyruvate, 1% non-essential amino acids. The HT29 cells were cultured in DMEM supplemented with 10% FBS and 100U/L penicillin/streptomycin. Both cell lines were grown as monolayers in tissue culture flasks as previously reported [23,36].
2.3. Cell treatments
Both cell lines were incubated individually with 3'HPPA, 3PPA, 4HBA, or BA at 10, 50, and 100 μM, prepared from 5 mM stock solutions as previously detailed [16].
2.4. Cytotoxicity assay
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay was used to determine cell viability. Following a previously established protocol [23], cells (1.5 × 104 cells/well) were seeded in a 96 multi-well plate and incubated for 48 h (37°C, 5% CO2) before replacement of media with 100 μL of each treatment and incubation (37°C, 5% CO2). After 24 h, cells were washed with PBS, treatments were replaced with an equal volume of media and 15 μL of MTT (5 mg/mL) was added to each well. After a 4 h incubation (37°C, 5% CO2) and addition of the solubilising solution, prepared as previously reported [17,28], absorbance at 570 nm was measured with a microplate-reader (VERSAmax). Experiments were carried out in triplicate and mean results expressed as a percentage (%) of cell viability normalized to untreated control. Deoxycholic acid (250 mM) was used as negative control and threshold cell viability level was set at 50%.
2.5. Genotoxicity assay
The COMET assay was used to determine each phenolic's efficacy in reducing DNA damage in CCD 841 CoN and HT29 cell lines. As outlined by De Santiago and co-workers [36], both cell lines were incubated with each phenolic compound individually for 24 h at 37°C in 5% CO2 before undergoing oxidative challenge with either 75 μM hydrogen peroxide (H2O2) for HT29 cells or 25 μM hydrogen peroxide (H2O2) for CCD841 Con cells to elicit an equivalent amount of damage (~50% Tail DNA) in the untreated controls. Images were analysed using a Nikon Eclipse 600 epifluorescence microscope at 400X magnification, with Komet 5.0 software used to record the tail DNA (%). The geno-protective potential was assessed by comparing the effects of the phenolics against the positive (H2O2) control and geno-toxic activity by comparing against the negative (PBS) experimental control. Experiments were carried out in triplicate and mean results expressed as a percentage% Tail DNA.
2.6. Real-time PCR (qPCR)
Following the previously established method [16], primers were designed for three genes of interest involved in the Nrf2-ARE pathway; Nrf2, Heme oxygenase-1 (HO-1), and NAD(P)H:quinone oxidoreductase 1 (NQO1), utilising the online software tools OligoPerfect (Thermo Fisher, UK) and NCBI Primer-BLAST [37]. Expression for each target gene was normalised against two reference genes (i.e., hypoxanthine phosphoribosyltransferase (HPRT) and β-actin) and analyses were performed in triplicate.
2.7. Statistical analysis
The results of three independent experiments were used for statistical analysis and presented as means ± standard error of the mean (SEM) for each type of dataset. Normality of datasets was assessed with the D’Agostino and Pearson omnibus test. One-way ANOVA, with Dunnett's Multiple comparison post-hoc test, was used to assessed significant differences between means (n = 3) compared to control and a Tukey’s Multiple comparison post-hoc test to compare concentration effects within compound for results of cytotoxicity, geno-protective activity, and gene expression. Additionally, Tukey’s multiple comparisons were also used to assess significance between the different compounds at the same concentrations for both geno-protective activity, and gene expression. An unpaired t-test was used to determine significance between cell lines for genotoxicity and gene expression. Significance was set at p<0.05. Analyses were performed using Prism 9 (Version 9.5.0 for Windows).
3. Results
3.1. Cell viability
Neither BA, 3'HPPA, 4HBA, or 3PPA at up to 100 µM influenced the viability of HT29 cells (Figure 1). In contrast, the viability of the CCD 841 CoN cell line was decreased to ~75% but only at the highest concentration tested (100 μM) when compared to the untreated control [for BA and 3'HPPA (p<0.01) and 4HBA and 3PPA (p<0.05)]. This indicates that the more highly transformed colonic cell line HT29 is less sensitive to the phenolic acids. As no dosage of the individual phenolics caused cell viability to decrease to the 50% viability threshold, they were considered sub-cytotoxic.
3.2. DNA damage reduction
All four phenolics, 3'HPPA, 3PPA, 4HBA, and BA, reduced oxidative-induced DNA damage in HT29 and CCD 841 CoN cells. In the HT29 cell line, the benzoic acids showed different patterns of effect (Figure 2A and B) with BA exerting a dose-dependent response with maximal effect at 100 µM, where BA reduced DNA damage by 23.3% tail DNA (p<0.001), followed by 50 µM BA (15.4%, p<0.001) and 10 µM BA (6.8%, p<0.05). In contrast, 4HBA was exhibited more geno-protection at the lowest concentration (10 μM) where it produced a threshold effect, resulting in a 20.4% (p<0.001) decrease in DNA damage and reduced activity seen at the higher concentrations (17.9% tail DNA at 50 μM and 18.5% tail DNA at 100 μM). For the phenylpropanoic acids (3PPA and 3'HPPA; Figure 2C and D), the most potent effect was observed at 50 µM with reductions of 17.9% and 15.7% respectively (p<0.001) while higher concentrations were less geno-protective. However, it was notable that 3'HPPA caused a significant reduction in DNA damage at 10 µM (10.7%, p<0.01) and was significantly (p<0.05) different in comparison to PPA which had no observable effect (1.7%).
In the CCD 841 CoN cell line, BA exhibited a dose-dependent response with maximal effect at 100 µM, reducing DNA damage by 25.4% (p<0.001), followed by 50 µM BA (15.0%, p<0.001) and 10 µM BA (9.6%, p<0.05). 4HBA was most potent at the highest 100 μM concentration (25.0% p<0.001) but interestingly was similarly effective at the lowest dose 10 μM (31.8%, p<0.001). The hydroxylated form is therefore more potent than the non-hydroxylated BA. For the phenylpropanoic acids, 3PPA and 3'HPPA, the most potent effect was observed at 100 µM (reductions of 27.1% and 25.0% respectively; p<0.001), however at the lowest concentration (10 µM) only 3'HPPA, exerted a significant reduction in DNA damage compared to control (42.0% vs 49.6%; p<0.05). This could indicate that 3'HPPA is the more potent compound, however, at 50 μM, 3PPA was significantly more effective in preventing DNA damage (p<0.05) than 3'HPPA.
The phenolic compounds under investigation did not exert significant geno-toxic damage compared to the negative experimental control in either cell line (data not shown) but did significantly reduce DNA damage (% Tail DNA) in response to oxidative challenge. 4HBA was more potent than BA in both cell lines, causing significant reduction in DNA damage even at lower concentrations. The effect of hydroxylation was, however, less clear for the phenylpropanoic acids as 3'HPPA, but not PPA, facilitated a significant reduction in DNA damage at 10 µM in both cell types. However, it is interesting to note that phenylpropanoic acids were markedly (p<0.05) less potent (Supplementary Table 1) at reducing DNA damage in the HT29 cell line at the highest doses compared to their effects on CCD 841 CoN.
3.3. Modulation of gene expression within the Nrf2-pathway
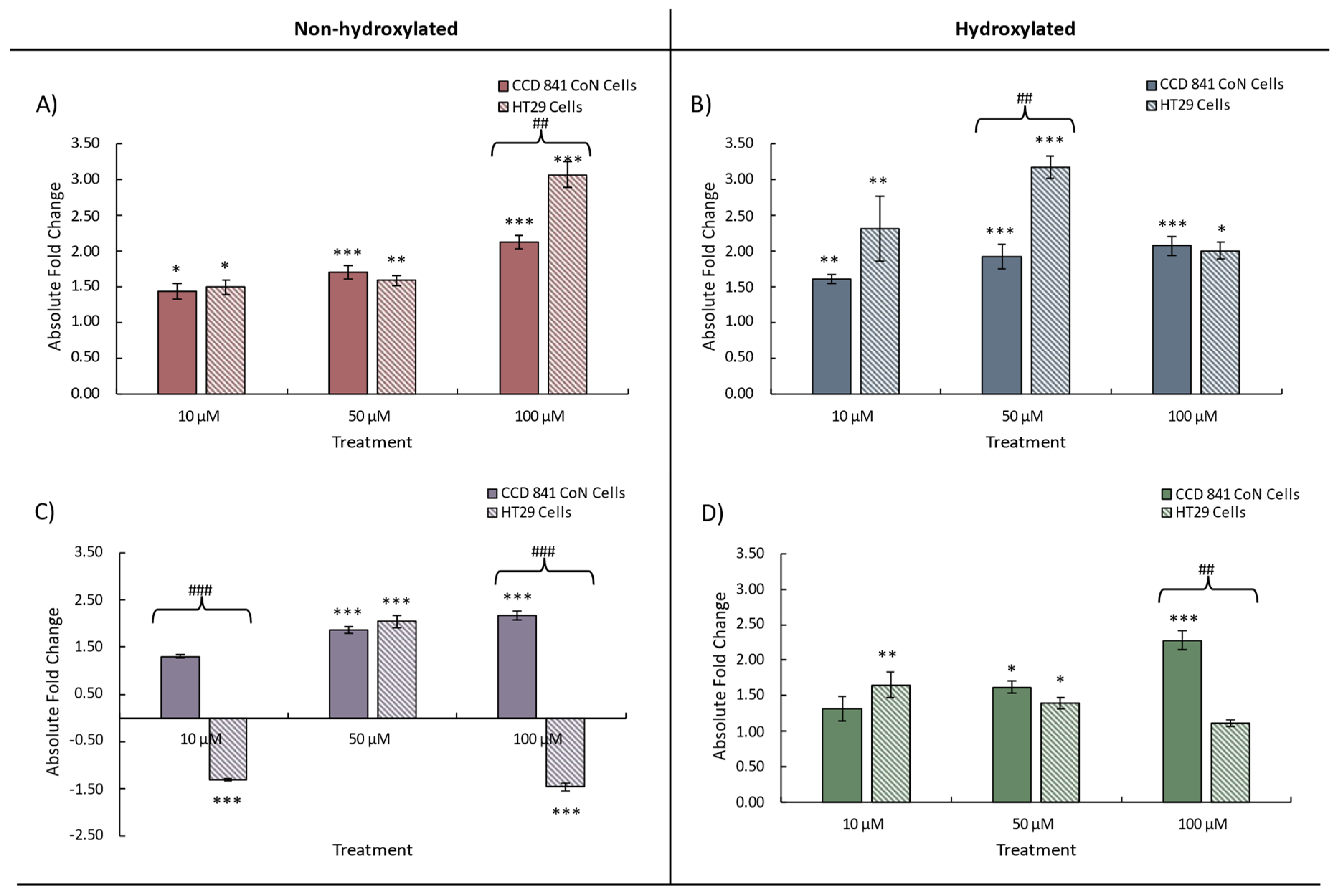
Addition of the phenolic acids resulted in an increase in the measured expression of Nrf2 in both cell lines compared to the untreated control (Figure 3). Changes in Nrf2 expression were generally similar between cell lines, however higher phenolic concentrations (50 and 100 μM) elicited significant increases (p<0.05) in Nrf2 expression above the controls. A dose effect was observed in HT29 cells treated with BA (maximum 2.37-fold increase, 100 mM, p<0.001), however an inverse correlation between dose and expression was seen for 4BA with no significant change from the control at 100 μM. An exception to these general observations was in HT29 adenocarcinoma cells, where the addition of 3PPA resulted in highly significant (p<0.001) decreases in Nrf2 expression at both 10 μM and 100 μM, but with a highly significant (p<0.001) increase at 50 μM (Figure 3C). By comparison, the CCD 841 CoN normal cell line exhibited increases in Nrf2 expression under the same treatments and the observed difference suggests less well-regulated and/or responsive gene expression in the carcinoma cell line – in the presence of 3PP – than normal CCD 841 CoN cells. It is known that Nrf2 binds to the ARE element upstream of target genes and increases expression of antioxidant defence/detoxification functions within the cell [28] and we therefore measured expression of NQO1 and HO-1 in the cell lines under investigation.
NQO1 scavenges ROS and is implicated in reducing DNA damage [38] and, in our investigation, NQO1 expression, similar to Nrf2 expression, was increased by the addition, individually, of all four phenolic acids to both cell lines (Figure 4). The less transformed CCD 841 CoN cells exhibited increased NQO1 expression with increasing phenolic concentration with the most pronounced dose effect observed with 3PPA (Figure 4C). Similar to the Nrf2 data, decreases in NQO1 expression were measured in HT29 cells in the presence of 3PPA, possibly as a consequence of the apparently dysregulated Nrf2 expression also observed in these cells with this treatment. We also noted decreased expression (1.5-fold) of NQO1 in HT29 cells exposed to 100 μM 4HBA, contrasting with a 1.5-fold increase in expression in CCD 841 CoN cells exposed to the same treatment (Figure 4B). Statistically significant differences in NQO1 expression between CCD841 and HT29 cells again mapped – broadly – to the same phenolic treatments where such differences were observed in Nrf2 expression and the data suggests that the greatest differences in gene expression between cells occurs at both the lowest and highest phenolic concentrations.
Heme oygenase (HO-1) is an inducible enzyme that exerts cytoprotective effects, as the products of it’s catabolism of haeme possess antioxidative, anti-inflammatory and antiapoptotic activities (see, e.g. PMC8547446) [39]. For this gene, we again observed a similar response to treatment with increases in expression arising from treatment of both cell lines (Figure 5). Divergence in HO-1 expression between CCD 841 CoN (increase) and HT29 cells (decrease) with 10 μM and 100 μM 3PPA was observed (Figure 5C), consistent with our observation for Nrf2 and NQO1. We also noted a relatively greater maximum increase in HO-1 expression with BA and 4HBA, and for 3’HPPA, with the maximal increases – with the exception of 4PPA (50 μM) – observed following treatment with 100 μM of a given phenolic. With the exception of the divergence in expression between cell lines, expressional changes following treatment with 3PPA were more modest (~1.5-fold) (Figure 5C) an observation made also for both Nrf2 (Figure 3) and NQO1 (Figure 4) expression. Taken together, the gene expression data indicate that at the lowest phenolic concentration (10 μM) no significant changes to gene expression may be expected in the normal cell line CCD 841 CoN, possibly indicating a more tightly regulated Nrf2-ARE cascade. This contrasts with the HT29 adenocarcinoma cell line in which significant changes in expression were observed – with the exception of BA – at 10 μM phenolic acid concentration. It is notable also that only the HT29 cell line exhibited decreases in gene expression in response to phenolic acid treatment whereas the CCD 841 CoN cells responded by increasing gene expression or exhibiting no significant change above that of the control. Limited differences between the hydroxylated and non-hydroxylated phenolics' ability to modulate the expression of each of the genes were evident and no clear pattern was discernible (Supplementary Table 1).
Taken together our data indicates that each of the phenolic compounds tested reduce DNA damage in colonic cells in response to an oxidative stress challenge. This effect appears mediated, in part, by altered gene expression of Nrf2, NQO1, and HO-1 genes. Some concentration and or structure dependent effects were apparent in the response of the more and less transformed cell lines to the phenolic acids tested. For example, phenylpropanoic acid treatments (PPA and 3'HPPA) caused noticeably different patterns of response in both cell lines (except 50 μM), while benzoic acids elicited similar levels of gene expression except at the highest concentration of 4HBA (Figure 3B).
4. Discussion
Reducing damage to DNA is a critical cellular defence mechanism that safeguards against mutations and maintains normal cellular functions [40]. Extensive research has established that (poly)phenols could positively contribute in maintaining a healthy status by influencing this mechanism along with exerting anti-proliferative, anti-inflammatory, and anti-oxidative effects [41-44]. As a result, there has been increased interest in determining the mechanism of actions of these plant-derived compounds and more recently their catabolites, at both the cellular and systemic level.
Due to their extensive biotransformation, especially in the distal gut, which alters (poly)phenol bioavailability and biological activity, it is important to test dietarily relevant phytochemicals at physiologically relevant doses and in the chemical forms encountered by different human tissues [12,19]. Phenolic acids are present both in the food sources and as catabolites derived from their in planta parent (poly)phenols [18-24] formed during catabolism by the intra-luminal microbiota [17], as well as from the putative metabolism of other compounds such as phenylalanine and catecholamines [44]. Compositional analyses of in vivo digestion or in vitro faecal fermentations can determine the physiological relevance of these compounds to the colon [16,45]. For example, Dobani et al. [16] highlighted a clear differentiation between pre- and post-fermentation ileal fluid samples, with eight raspberry-derived phenolic acid catabolites identified as key discriminators of the fermentation process, four of which were assessed for their geno-protective potential and the ability to modulate genes in the cytoprotective Nrf2 mediated ARE pathway in normal colonocytes (CCD 841 CoN).
In the current study, the work by Dobani and colleagues on the geno-protective effects of the simple phenolics (3PPA, 3'HPPA, BA, 4’HBA) on the CCD 841 CoN cells [16] has been expanded using the classic and more transformed colonocyte cell line HT29. This cell line exhibits rapid growth, irregular morphology, and carries genetic alterations associated with colorectal adenocarcinoma [46]. In contrast, CCD 841 CoN cells are derived from normal cells, display slower growth, epithelial-like morphology, and retain the genetic and phenotypic characteristics of healthy colonic epithelium [47]. Heterogeneities in both physiological and biochemical response have been reported between these cell lines when testing the same compound under similar conditions [48] and, in the current work, the individual phenolic acid treatments exerted greater geno-protective capacity in the CCD 841 CoN cell line, particularly at the highest concentrations of 4HBA, 3PPA and 3'HPPA. By contrast, a recent ex vivo study investigating strawberry enriched ileal fluid fermentates in the same cell lines, determined that a complex mixture of (poly)phenolic compounds was less effective in reducing DNA damage in CCD 841 CoN cells than HT29 cells in response to oxidative challenge [24]. An investigation that assessed a colonically-relevant raspberry-derived triterpenoid extract elucidated that the extract exerted greater anti-genotoxic capacity in the CCD 841 CoN cells than HT29 cells, reducing DNA damage by ~55% and ~45% respectively. Overall, the data from the current investigation are generally consistent with previous work in our laboratory [45], in which HT29 cells treated with 5 μg/mL of 3'HPPA (30 μM) or 4HBA (~36 μM) exhibited a reduction in DNA damage of ca. 30% in response to oxidative challenge (75 μM H2O2), similar to the magnitude of effect reported here. It has been reported that açai and blueberry extracts (2.5 mg/mL), which are also sources of simple phenolic compounds like 4HBA, in addition to other polyphenolics such as pelargonidin-3-O-glucoside, reduced DNA damage in HT29 cells by ~60% (p<0.001) compared to the untreated control [49]. What are arguably the physiologically more relevant effects were revealed when the in vitro digested and fermented acai extract was tested, where it significantly reduced DNA damage by ~31.5% [50].
The gut microbiota plays a pivotal role in the modulation of (poly)phenols and their colonic catabolites [22,23,24]. The phenolic acids used in this study are present during colonic transit at differing concentrations and in differing forms, differentiated through the presence/absence of a hydroxyl group. Hydroxylated phenolics have been reported to have higher bioavailability and bioactivity [51] and this may be corroborated by comparison of DNA damage between the hydroxylated and non-hydroxylated phenolics in each cell line. Treatment with both 4HBA and 3'HPPA resulted in greater reductions in DNA damage, particularly at lower concentrations, than either BA or 3PPA, in addition to their higher level of gene modulation, particularly in the adenocarcinoma cells. This observation may be due to greater antioxidant activity and indeed a recent study [52] reported that the microbially-derived 3′-hydroxylated soy isoflavone derivatives, 3′-hydroxydaidzin and 3′-hydroxygenistin, exhibited higher free radical scavenging activity than their non-hydroxylated counterparts. In addition, Cao et al. [53] reported stronger antioxidant activity in flavonoids with hydroxyl groups at both the 3′ and 4′ positions of the B-ring in comparison to their dehydroxylated counterparts. Our observations, therefore, may be attributable to the increased antioxidant activity seen with the hydroxylated phenolic acids, as hydroxyl groups are reported as having strong radical-scavenging and antioxidative effects [54,55]. As both the hydroxylated and non-hydroxylated forms provide geno-protective effects in vitro at physiological levels, this suggests that they could provide synergistic protection following their colonic generation via the successive microbial degradation of intact flavonoids to the simple phenolic acids that accumulate and are excreted in the faeces.
The underlying mechanism behind the antigenotoxic effects of these compounds is not fully understood, however, accumulating evidence [56,57,58,59,60,61] suggests that the Nrf2-ARE pathway may play a role in protecting against DNA damage by activating downstream targets such as NQO1 and HO-1, potent anti-inflammatory enzymes that alleviate oxidative stress in cells [28]. The current study clearly highlights the role each phenolic acid may play in modulating the expression of Nrf2 and its target genes, NQO1 and HO-1 in both normal colonic epithelial cells (CCD 841 CoN) and carcinogenic colonic epithelial cells (HT29). The hydroxylation status of the phenolic compound tested appears to exert some differential effects on gene expression for HO1 & NQO1 but not for Nrf2. For Nrf2, no clear pattern of effect was evident, for example PPA and 3'HPPA caused noticeably different patterns (increase vs decrease) while benzoic acids elicited similar patterns and levels of gene expression in both cell lines. This is in broad agreement with the previous finding of McDougall et al. [24] who demonstrated that a raspberry triterpenoid-rich fraction caused increased expression of Nrf2 in HT29 cells but, conversely, decreased it in CCD 841 CoN cells, while measured NQO1 (increase) and HO-1 (decrease) gene expression was similar in both cell lines. More recently, research in hepatocyte cells (HepG2) [62] demonstrated that a solvent-based raspberry extract (200 μg/mL) – albeit at physiologically questionable levels – increased Nrf2, NQO1 and HO-1 gene expression by 38.4% (p<0.01), 18.7% (p<0.01) and 9.4% relative to the control. (Poly)phenols, such as resveratrol, cyanidin, chlorogenic acid and luteolin also activate the Nrf2-ARE pathway and exert antigenotoxic effects [63,64]. Indeed, luteolin (60 μM) reduced H2O2 induced ROS generation (at 24 h) by 50% in a dose-dependent manner, mediated in part by significant (p<0.05) time dependent (12, 24 & 48 h) increases in Nrf2 expression (~15-, 50- & 145-fold increased mRNA levels, respectively) in HT29 cells [64]. Therefore, the current study builds upon our previous work, and that of others ,to emphasize the role of (poly)phenols in inducing phase II detoxifying and antioxidant enzymes, regulating oxidative stress in both normal and adenocarcinoma cells. However, the effects vary depending on the compound, concentration, and cell type.
5. Conclusions
This study provides evidence that microbially-mediated phenolic catabolites possesses the ability to reduce DNA damage induced by oxidative challenge in two colonic epithelial models. The protective effect is partially attributed to the upregulation of the Nrf2-ARE pathway and its downstream targets, NQO1 and HO-1. Despite this, the hydroxylation state of the phenolic acids appears to have limited impact on the bioactivity overall. Consequently, the presence of abundant amounts of low molecular weight phenolic catabolites from diets rich in (poly)phenols may exert protective effects against colonic epithelial damage in vivo by reducing DNA damage, a key early-stage event in colorectal carcinogenesis.
Author Contributions
C.I.R.G., G.McD., R.L., I.R. and A.C. were involved in study design. B.O.M. S.D., M.F. and C.L., were involved in experimental and data analysis. The manuscript was prepared by B.O.M., T.M.A, C.L., S.D., G.P.-C., N.G.T., L.K.P., D.D.R., C.L., I.R., G.McD., A.C. and C.I.R.G with contributions from all the authors.
Acknowledgments
C.I.R.G., R.L. and A.C. acknowledge funding from the National Processed Raspberry Council who had no involvement in study design; in the collection, analysis and interpretation of data; in the writing of the report; and in the decision to submit the article for publication. G.McD., and J.W.A. acknowledge funding from the Scottish Government's Rural and Environment Science and Analytical Services (RESAS) Division. G.McD. acknowledge funding from BachBerry (Project No. FP7-613793). G.P.-C. was supported by a postdoctoral research contract “Juan de la Cierva-Incorporación” funded by the Spanish Ministry of Economy and Competitiveness (FJCI-2015-26433). C.I.R.G., A.C., G.P.-C. and T.M.A. were funded by the Distinguished Scientist Fellowship Program (DSFP) of King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors report no conflicts of interest.
Abbreviations
3'HPPA, 3-(3′-hydroxyphenyl)propanoic acid; 3PPA, 3-(phenyl)propanoic acid; 4HBA, 4-hydroxybenzoic acid; ATCC, American Type Culture Collection; BA, benzoic acid; BID, BH3-interacting domain death agonist; CRC, colorectal cancer; DMEM, Dulbecco’s Modified Eagle’s Medium; ECACC, European Collection of Cell Cultures; FBS, Foetal Bovine Serum; GIT, Gastrointestinal tract; H2O2, Hydrogen peroxide; HO-1, Heme oxygenase 1; HPRT, Hypoxanthine phosphoribosyltransferase; MEM, Minimum Essential Media; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; NQO1, NAD(P)H dehydrogenase, quinone 1; Nrf2, Nuclear factor erythroid 2-related factor 2; PBS, phosphate-buffered saline; PMAIP1, phorbol-12-myristate-13-acetate-induced protein 1; SEM, Standard error of the mean.
References
- Patel, J.; Baptiste, B.A.; Kim, E.; Hussain, M.; Croteau, D.L.; Bohr, V.A. DNA damage and mitochondria in cancer and aging. Carcinogenesis 2020, 41, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Soria-Valles, C.; López-Soto, A.; Osorio, F.G.; López-Otín, C. Immune and inflammatory responses to DNA damage in cancer and aging. Mech Ageing Dev 2017, 165, 10–16. [Google Scholar] [CrossRef]
- Niedernhofer, L.J.; Gurkar, A.U.; Wang, Y.; Vijg, J.; Hoeijmakers, J.H.J.; Robbins, P.D. Nuclear genomic instability and aging. Annu Rev Biochem 2018, 87, 295–322. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, B.; Pothof, J.; Vijg, J.; Hoeijmakers, J.H.J. The central role of DNA damage in the ageing process. Nature 2021, 592, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Heinen, C.D. The mismatch repair-dependent DNA damage response: mechanisms and implications. DNA Repair 2019, 78, 60–69. [Google Scholar] [CrossRef]
- Rashid, S.; Freitas, M.O.; Cucchi, D.; Bridge, G.; Yao, Z.; Gay, L.; Williams, M.; Wang, J.; Suraweera, N.; Silver, A.; McDonald, S.A.C.; Chelala, C.; Szabadkai, G.; Martin, S.A. MLH1 deficiency leads to deregulated mitochondrial metabolism. Cell Death Dis 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E. , Santini, A. Polyphenols: a concise overview on the chemistry, occurrence, and human health. Phytother Res 2019, 33, 2221. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and human health: the role of bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.H.; Parihar, A.; Doseff, A.I. Dietary flavonoids for immunoregulation and cancer: food design for targeting disease. Antioxidants 2019, 8, 202. [Google Scholar] [CrossRef]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary flavonoids: cardioprotective potential with antioxidant effects and their pharmacokinetic, toxicological and therapeutic concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef]
- Rufino, A.T.; Costa, V.M.; Carvalho, F.; Fernandes, E. Flavonoids as antiobesity agents: a review. Med Res Rev 2021, 41, 556–585. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid Redox Signal 2013, 18, 1818–1892. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.; Touillaud, M.; Katzke, V.; Kühn, T.; Boeing, H.; Förster, J.; Trichopoulou, A.; Valanou, E.; Peppa, E.; Palli, D.; Agnoli, C.; Ricceri, F.; Tumino, R.; de Magistris, M.S.; Peeters, P.H.M.; Bueno-de-Mesquita, H.B.; Engeset, D.; Skeie, G.; Hjartåker, A.; Menéndez, V.; Agudo, A.; Molina-Montes, E.; Huerta, J.M.; Barricarte, A.; Amiano, P.; Sonestedt, E.; Nilsson, L.M.; Landberg, R.; Key, T.J.; Khaw, K.; Wareham, N.J.; Lu, Y.; Slimani, N.; Romieu, I.; Riboli, E.; Scalbert, A. Dietary polyphenol intake in Europe: the European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur J Nutr 2016, 55, 1359–1375. [Google Scholar] [CrossRef]
- Gavrilas, L.I.; Cruceriu, D.; Ionescu, C.; Miere, D.; Balacescu, O. Pro-apoptotic genes as new targets for single and combinatorial treatments with resveratrol and curcumin in colorectal cancer. Food Funct 2019, 10, 3717–3726. [Google Scholar] [CrossRef]
- Song, M.; Lee, D.; Chun, K.; Kim, E. The Role of NRF2/KEAP1 Signaling Pathway in Cancer Metabolism. Int J Mol Sci 2021, 22, 4376. [Google Scholar] [CrossRef]
- Dobani, S.; Latimer, C.; McDougall, G.J.; Allwood, J.W.; Pereira-Caro, G.; Moreno-Rojas, J.M.; Ternan, N.G.; Pourshahidi, L.K.; Lawther, R.; Tuohy, K.M.; Del Rio, D.; O'Connor, G.; Rowland, I.; Almutairi, T.M.; Crozier, A.; Gill, C.I.R. Ex vivo fecal fermentation of human ileal fluid collected after raspberry consumption modifies (poly)phenolics and modulates geno-protective effects in colonic epithelial cells. Redox Biol 2021, 40, 101862. [Google Scholar] [CrossRef]
- Carregosa, D.; Pinto, C.; Ávila-gálvez, M.Á.; Bastos, P.; Berry, D.; Santos, C.N. A look beyond dietary (poly)phenols: the low molecular weight phenolic metabolites and their concentrations in human circulation. Compr Rev Food Sci Food Saf 2022, 21, 3931. [Google Scholar] [CrossRef]
- Wang, M.; Yu, F.; Zhang, Y.; Chang, W.; Zhou, M. The effects and mechanisms of flavonoids on cancer prevention and therapy: focus on gut microbiota. J Agric Food Chem 2022, 70, 1451–1475. [Google Scholar] [CrossRef]
- Mena, P.; Bresciani, L.; Brindani, N.; Ludwig, I.A.; Pereira-Caro, G.; Angelino, D.; Calani, L.; Brighenti, F.; Clifford, M.N.; Del Rio, D. Phenyl-γ-valerolactones and phenylvaleric acids, the main colonic metabolites of flavan-3-ols: synthesis, analysis, bioavailability, and bioactivity. Nat Protoc 2019, 36, 714–752. [Google Scholar] [CrossRef]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-mediated gut microbiota modulation: toward prebiotics and further. Front Nutr 2021, 8, 689456. [Google Scholar] [CrossRef]
- Diotallevi, C.; Fontana, M.; Latimer, C.; Ternan, N.G.; Pourshahidi, L.K.; Lawther, R.; O’Connor, G.; Conterno, L.; Gasperotti, M.; Angeli, A.; Lotti, C.; Bianchi, M.; Vrhovsek, U.; Fava, F.; Gobbetti, M.; Gill, C.I.R.; Tuohy, K.M. Ex Vivo Faecal fermentation of human ileal fluid collected after wild strawberry consumption modulates human microbiome community structure and metabolic output and protects against DNA damage in colonic epithelial cells. Mol Nutr Food Res 2021, 66, e2100405. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. bioactivity of dietary polyphenols: the role of metabolites. Crit Rev Food Sci Nutr 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Brown, E.M.; Nitecki, S.; Pereira-Caro, G.; McDougall, G.J.; Stewart, D.; Rowland, I.R.; Crozier, A.; Gill, C.I. Comparison of in vivo and in vitro digestion on polyphenol composition in lingonberries: potential impact on colonic health. Food Funct 2014, 5, 611–623. [Google Scholar] [CrossRef]
- McDougall, G.J.; Allwood, J.W.; Pereira-Caro, G.; Brown, E.M.; Verrall, S.; Stewart, D.; Latimer, C.; McMullan, G.; Lawther, R.; O’Connor, G.; Rowland, I.; Crozier, A.; Gill, C.I. Novel colon-available triterpenoids identified in raspberry fruits exhibit antigenotoxic activities in vitro. Mol Nutr Food Res 2017, 61, 1600327. [Google Scholar] [CrossRef]
- Jin, H.; Tan, X.; Liu, X.; Ding, Y. Study of effect of tea polyphenols on microsatellite instability colorectal cancer and its molecular mechanism. Int J Colorectal Dis 2010, 25, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Shan, S.; Li, H.; Shi, J.; Zhang, X.; Li, Z. Reversal effects of bound polyphenol from foxtail millet bran on multidrug resistance in human hct-8/fu colorectal cancer cell. J Agric Food Chem 2018, 66, 5190–5199. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Pawlik, N.; Polka, D.; Nowak, A.; Koziołkiewicz, M.; Podsędek, A. Viburnum opulus fruit phenolic compounds as cytoprotective agents able to decrease free fatty acids and glucose uptake by caco-2 cells. Antioxidants 2019, 8, 262. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An overview of Nrf2 signaling pathway and its role in inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Telkoparan-Akillilar, P.; Panieri, E.; Cevik, D.; Suzen, S.; Saso, L. Therapeutic targeting of the Nrf2 signaling pathway in cancer. Molecules 2021, 26, 1417. [Google Scholar] [CrossRef]
- Islam, M.A.; Medha, M.M.; Nahar, A.U.; Al Fahad, M.A.; Siraj, M.A. Cancer protective role of selected dietary polyphenols via modulating Keap1/Nrf2/ARE and interconnected signaling pathways. Nutr Cancer 2023, 75, 1065–1102. [Google Scholar] [CrossRef]
- Schmidlin, C.J.; Dodson, M.B.; Madhavan, L.; Zhang, D.D. Redox regulation by NRF2 in aging and disease. Free Radic Biol Med 2019, 134, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jiang, Z.; Lu, H.; Xu, Z.; Tong, R.; Shi, J.; Jia, G. Recent advances of natural polyphenols activators for keap1-Nrf2 signaling pathway. Chem Biodivers 2019, 16, e1900400. [Google Scholar] [CrossRef]
- Prasad, R.; Katiyar, S.K. Polyphenols from green tea inhibit the growth of melanoma cells through inhibition of class I histone deacetylases and induction of DNA damage. Genes Cancer 2015, 6, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Majidinia, M.; Bishayee, A.; Yousefi, B. Polyphenols: Major regulators of key components of DNA damage response in cancer. DNA Repair 2019, 82, 102679. [Google Scholar] [CrossRef]
- Maleki Dana, P.; Sadoughi, F.; Asemi, Z.; Yousefi, B. The role of polyphenols in overcoming cancer drug resistance: a comprehensive review. Cell Mol Biol Lett 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- De Santiago, E.; Gill, C.I.R.; Carafa, I.; Tuohy, K.M.; De Peña, M.-P.; Cid, C. Digestion and colonic fermentation of raw and cooked Opuntia ficus-indica Cladodes impacts bioaccessibility and bioactivity. J Agric Food Chem 2019, 67, 2490. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: a tool to design target-specific primers for polymerase chain reaction. BMC Bioinform 2012, 13, 134. [Google Scholar] [CrossRef]
- Rashid, M.H.; Babu, D.; Siraki, A.G. Interactions of the antioxidant enzymes NAD(P)H: Quinone oxidoreductase 1 (NQO1) and NRH: Quinone oxidoreductase 2 (NQO2) with pharmacological agents, endogenous biochemicals and environmental contaminants. Chem Biol Interact 2021, 345, 109574. [Google Scholar] [CrossRef]
- Zhang, A.; Suzuki, T.; Adachi, S.; Naganuma, E.; Suzuki, N.; Hosoya, T.; Itoh, K.; Sporn, M.B.; Yamamoto, M. Distinct Regulations of HO-1 Gene Expression for Stress Response and Substrate Induction. Mol Cell Biol 2021, 41, e0023621. [Google Scholar] [CrossRef]
- Izquierdo-Vega, J.A.; Morales-González, J.A.; SánchezGutiérrez, M.; Betanzos-Cabrera, G.; Sosa-Delgado, S.M.; Sumaya-Martínez, M.T.; Morales-González, Á.; Paniagua-Pérez, R.; Madrigal-Bujaidar, E.; Madrigal-Santillán, E. Evidence of some natural products with antigenotoxic effects. Part 1: Fruits and polysaccharides. Nutrients 2017, 9, 102. [Google Scholar] [CrossRef]
- Patra, S. , Pradhan, B., Nayak, R., Behera, C., Das, S., Patra, S., Efferth, T., Jena, M., and Bhutia, S. Dietary polyphenols in chemoprevention and synergistic effect in cancer: clinical evidences and molecular mechanisms of action. Phytomedicine 2021, 90, 153554. [Google Scholar] [CrossRef]
- Coșarcă, S.; Moacă, E.; Tanase, C.; Muntean, D.; Pavel, I.; Dehelean, C. Spruce and beech bark aqueous extracts: source of polyphenols, tannins and antioxidants correlated to in vitro antitumor potential on two different cell lines. Wood Sci Technol 2019, 53, 313–333. [Google Scholar] [CrossRef]
- Moura, C.; Noguti, J.; Jesus, G.; Ribeiro, F.; Garcia, F.; Gollucke, A.; Aguiar, O.; Ribeiro, D. Polyphenols as a chemopreventive agent in oral carcinogenesis: putative mechanisms of action using in-vitro and in-vivo test systems. Eur J Cancer Prev 2013, 22, 467–472. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Cáceres-Jimenez, S.; Bresciani, L.; Mena, P.; Almutairi, T.M.; Dobani, S.; Pourshahidi, L.K.; Gill, C.I.R.; Moreno Rojas, J.M.; Clifford, M.N.; Crozier, A. Excretion by subjects on a low (poly)phenol diet of phenolic gut microbiota catabolites sequestered in tissues or associated with catecholamines and surplus amino acids. Int J Food Sci Nutr 2023, 74, 532–543. [Google Scholar] [CrossRef]
- Brown, E.M.; McDougall, G.J.; Stewart, D.; Pereira-Caro, G.; González-Barrio, R.; Allsopp, P.; Magee, P.; Crozier, A.; Rowland, I.; Gill, C.I.R. Persistence of anticancer activity in berry extracts after simulated gastrointestinal digestion and colonic fermentation. PLOS ONE 2012, 7, e49740. [Google Scholar] [CrossRef]
- Pearce, S.C.; Coia, H.G.; Karl, J.P.; Pantoja-Feliciano, I.G.; Zachos, N.C.; Racicot, K. Intestinal in vitro and ex vivo models to study host-microbiome interactions and acute stressors. Front Physiol 2018, 9, 1584. [Google Scholar] [CrossRef]
- Zoetemelk, M.; Rausch, M.; Colin, D.J.; Dormond, O.; Nowak-Sliwinska, P. Short-term 3d culture systems of various complexity for treatment optimization of colorectal carcinoma. Sci Rep 2019, 9, 7103. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, Z.; Wang, Y.; Zhang, L.; Chua, N.; Dai, L.; Chen, J.; Ho, C.L. Evaluation of the anti-inflammatory and anti-oxidative effects of therapeutic human lactoferrin fragments. Front Bioeng Biotechnol 2021, 9. [Google Scholar] [CrossRef]
- Randah, A.; Daniel, C.; Rowland, I.R. Acai fruit as a source of bioactive phytochemicals. Nutrients 2018, 10, 496. [Google Scholar] [CrossRef]
- Alqurashi, R.M.; Alarifi, S.N.; Walton, G.E.; Costabile, A.F.; Rowland, I.R.; Commane, D.M. In vitro approaches to assess the effects of açai (Euterpe oleracea) digestion on polyphenol availability and the subsequent impact on the faecal microbiota. Food Chem 2017, 234, 190–198. [Google Scholar] [CrossRef]
- Lee, U.-J.; Sohng, J.K.; Kim, B.-G.; Choi, K.-Y. Recent trends in the modification of polyphenolic compounds using hydroxylation and glycosylation. Curr Opin Biotechnol 2023, 80, 102914. [Google Scholar] [CrossRef]
- Chiang, C.-M.; Wang, D.-S.; Chang, T.-S. Improving free radical scavenging activity of soy isoflavone glycosides daidzin and genistin by 3′-hydroxylation using recombinant Escherichia coli. Molecules 2016, 21, 1723. [Google Scholar] [CrossRef]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: structure-activity relationships. Free Radic Biol Med 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Umeno, A.; Horie, M.; Murotomi, K.; Nakajima, Y.; Yoshida, Y. Antioxidative and antidiabetic effects of natural polyphenols and isoflavones. Molecules 2016, 21, 708. [Google Scholar] [CrossRef]
- Kostić, K.; Brborić, J.; Delogu, G.; Simić, M.R.; Samardžić, S.; Maksimović, Z.; Dettori, M.A.; Fabbri, D.; Kotur-Stevuljević, J.; Saso, L. Antioxidant activity of natural phenols and derived hydroxylated biphenyls. Molecules 2023, 28, 2646. [Google Scholar] [CrossRef]
- Mou, S.; Zhou, Z.; He, Y.; Liu, F.; Gong, L. Curcumin inhibits cell proliferation and promotes apoptosis of laryngeal cancer cells through Bcl-2 and PI3K/Akt, and by upregulating miR-15a. Mol Med Rep 2017, 14, 4937–4942. [Google Scholar] [CrossRef]
- Yoshimura, H.; Yoshida, H.; Matsuda, S.; Ryoke, T.; Ohta, K.; Ohmori, M. The therapeutic potential of epigallocatechin 3 gallate against human oral squamous cell carcinoma through inhibition of cell proliferation and induction of apoptosis: In vitro and in vivo murine xenograft study. Clin Oral Investig 2016, 20, 1139–1148. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Alam, M.B.; Ju, M.-K.; Kwon, K.-R.; Huh, Y.S.; Han, Y.-K.; Lee, S.H. Antioxidant mechanism of polyphenol-rich Nymphaea nouchali leaf extract protecting DNA damage and attenuating oxidative stress-induced cell death via Nrf2-mediated heme-oxygenase-1 induction coupled with ERK/p38 signaling pathway. Biomolecules 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Silva, M.M.; Rocha, C.R.R.; Kinker, G.S.; Pelegrini, A.L.; Menck, C.F.M. The balance between NRF2/GSH antioxidant mediated pathway and DNA repair modulates cisplatin resistance in lung cancer cells. Sci Rep 2019, 9, 17639. [Google Scholar] [CrossRef]
- Darband, S.G.; Sadighparvar, S.; Yousefi, B.; Kaviani, M.; Ghaderi-Pakdel, F.; Mihanfar, A.; Rahimi, Y.; Mobaraki, K.; Majidinia, M. Quercetin attenuated oxidative DNA damage through NRF2 signaling pathway in rats with DMH induced colon carcinogenesis. Life Sci 2020, 253, 117584. [Google Scholar] [CrossRef]
- Moratilla-Rivera, I.; Sánchez, M.; Valdés-González, J.A.; Gómez-Serranillos, M.P. Natural products as modulators of Nrf2 signaling pathway in neuroprotection. Int J Mol Sci 2023, 24, 3748. [Google Scholar] [CrossRef]
- Chen, L.; Li, K.; Liu, Q.; Quiles, J.L.; Filosa, R.; Kamal, M.A.; Wang, F.; Kai, G.; Zou, X.; Teng, H.; Xiao, J. Protective effects of raspberry on the oxidative damage in HepG2 cells through Keap1/Nrf2-dependent signalling pathway. Food Chem Toxicol 2019, 133, 110781. [Google Scholar] [CrossRef]
- Teng, H.; Fang, T.; Lin, Q.; Song, H.; Liu, B.; Chen, L. Red raspberry and its anthocyanins: bioactivity beyond antioxidant capacity. Trends Food Sci Technol 2017, 66, 153–165. [Google Scholar] [CrossRef]
- Kang, K.A.; Piao, M.J.; Hyun, Y.J.; Zhen, A.X.; Cho, S.J.; Ahn, M.J.; Yi, J.M.; Hyun, J.W. Luteolin promotes apoptotic cell death via upregulation of Nrf2 expression by DNA demethylase and the interaction of Nrf2 with p53 in human colon cancer cells. Exp Mol Med 2019, 51, 1–14. [Google Scholar] [CrossRef]
Figure 1.
Cytotoxic effect of different concentrations of phenolics on CCD 841 CoN and HT29 cells. Data presented as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *p<0.05 **p<0.01. Benzoic acid, BA; 4-hydroxybenzoic acid, 4HBA; 3-(3′-hydroxyphenyl)propanoic acid, 3'HPPA; 3-(phenyl)propanoic acid, 3PPA.
Figure 1.
Cytotoxic effect of different concentrations of phenolics on CCD 841 CoN and HT29 cells. Data presented as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *p<0.05 **p<0.01. Benzoic acid, BA; 4-hydroxybenzoic acid, 4HBA; 3-(3′-hydroxyphenyl)propanoic acid, 3'HPPA; 3-(phenyl)propanoic acid, 3PPA.
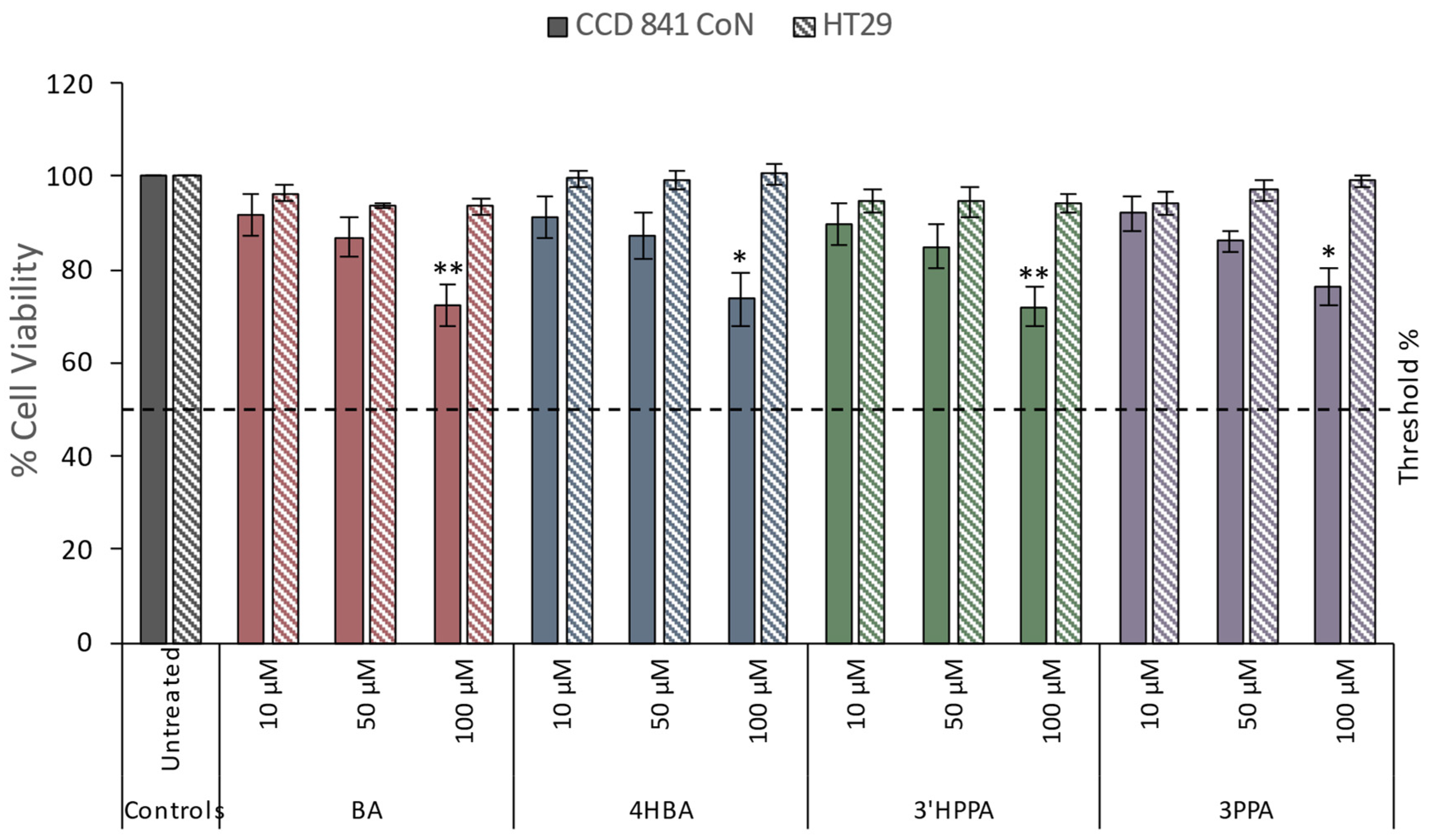
Figure 2.
DNA damage reducing capacity of 4 individual phenolics on CCD 841 CoN and HT29 cells in response to oxidative challenge. Data for A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA), at 10, 50, or 100 μM after 24 h pre-incubation of cells at 37°C and challenge with H2O2 (75 µM - HT29 or 25 µM - CDD 841 CoN) are presented. Data presented is as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *p<0.05, **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05.
Figure 2.
DNA damage reducing capacity of 4 individual phenolics on CCD 841 CoN and HT29 cells in response to oxidative challenge. Data for A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA), at 10, 50, or 100 μM after 24 h pre-incubation of cells at 37°C and challenge with H2O2 (75 µM - HT29 or 25 µM - CDD 841 CoN) are presented. Data presented is as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *p<0.05, **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05.
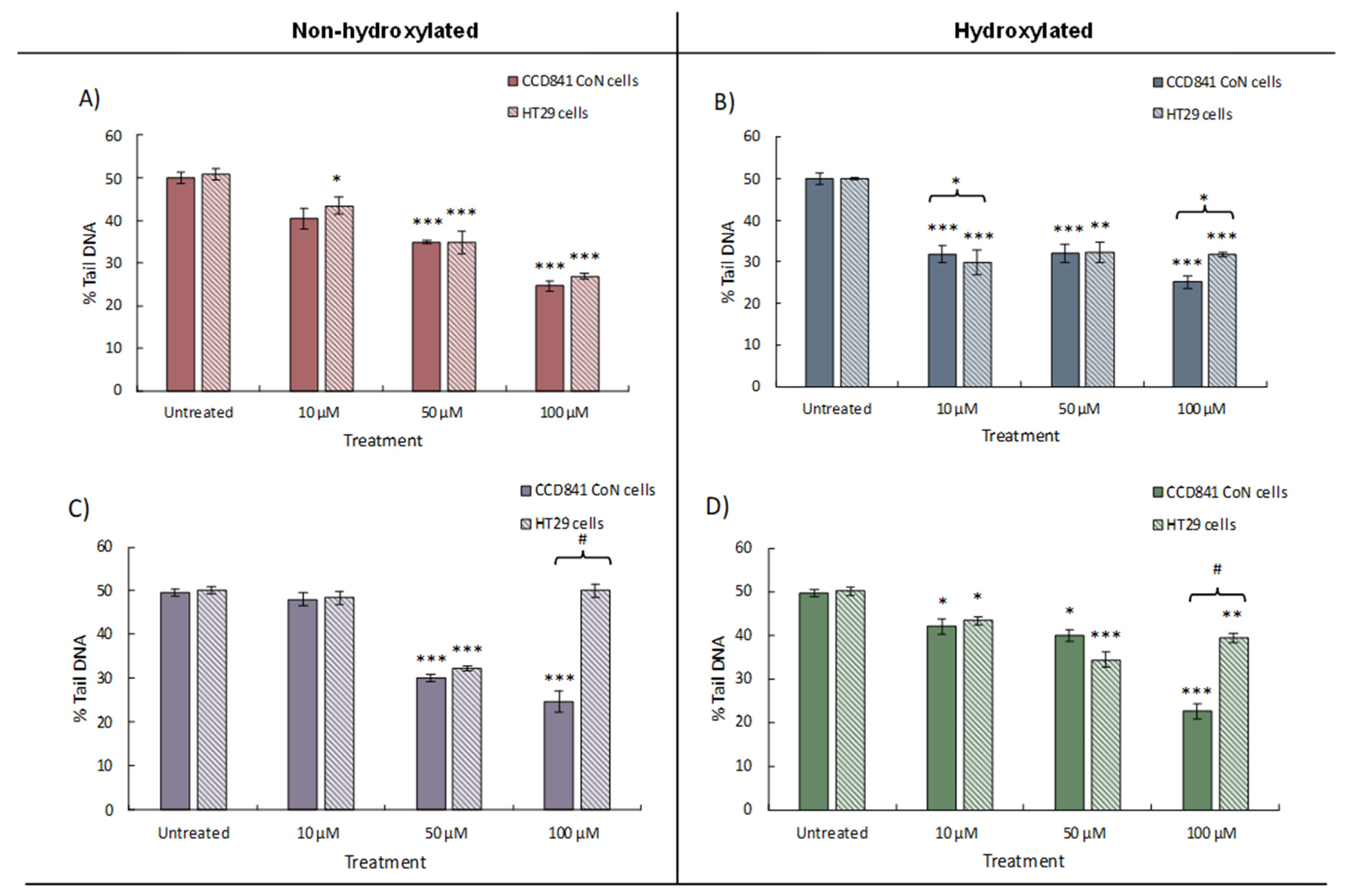
Figure 3.
Change in Nrf2 expression in CCD 841 CoN and HT29 cells treated with either 10, 50, or 100 μM of individual phenolic compounds. Data for A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA) is presented as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *<p0.05 **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05, ##p<0.01, ###p<0.001.
Figure 3.
Change in Nrf2 expression in CCD 841 CoN and HT29 cells treated with either 10, 50, or 100 μM of individual phenolic compounds. Data for A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA) is presented as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *<p0.05 **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05, ##p<0.01, ###p<0.001.
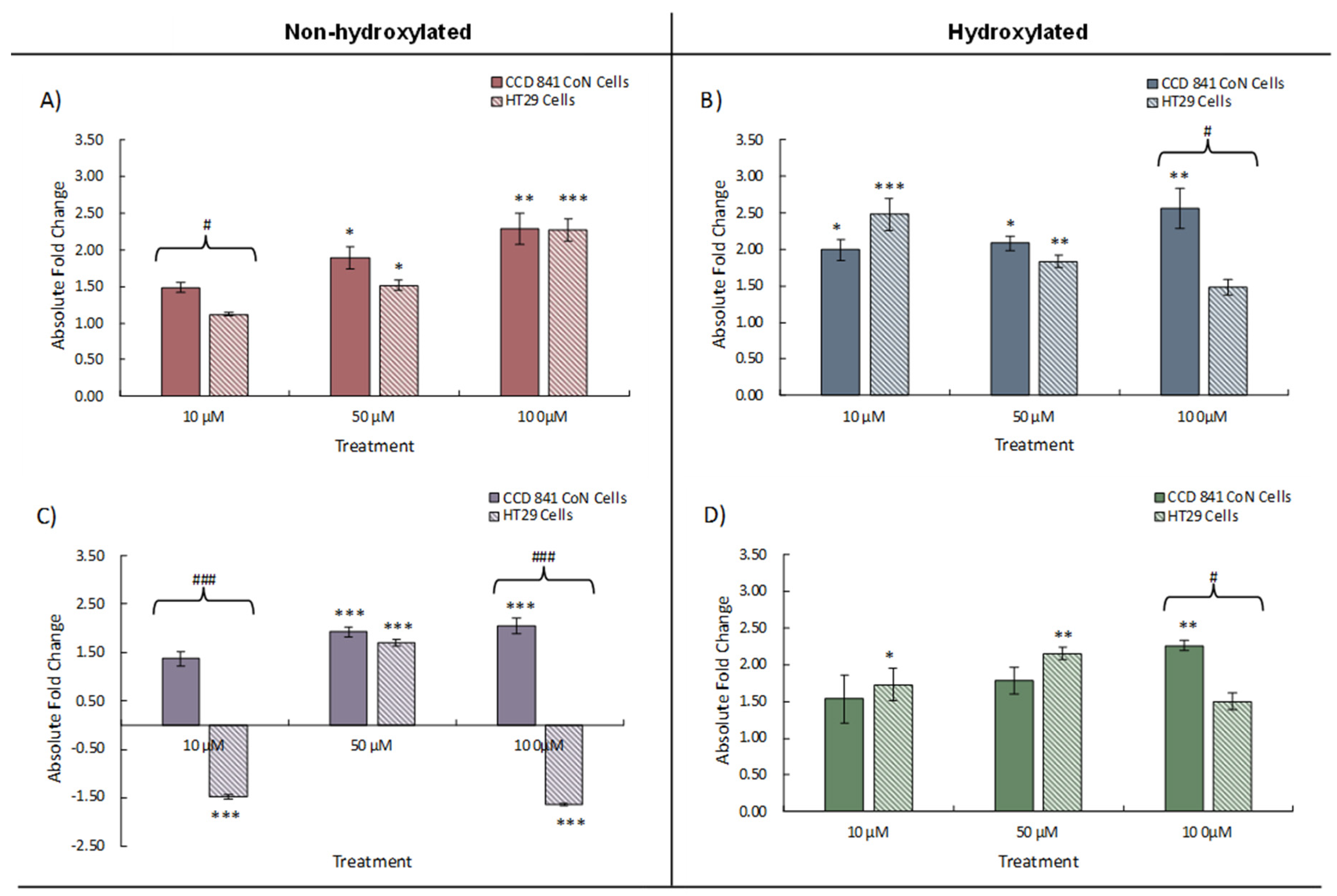
Figure 4.
Change in NQO1 expression in CCD 841 CoN and HT29 cells treated with either 10, 50, or 100 μM individual phenolics; A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA). Data presented is as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *<p0.05 **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05, ##p<0.01, ###p<0.001.
Figure 4.
Change in NQO1 expression in CCD 841 CoN and HT29 cells treated with either 10, 50, or 100 μM individual phenolics; A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA). Data presented is as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *<p0.05 **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05, ##p<0.01, ###p<0.001.
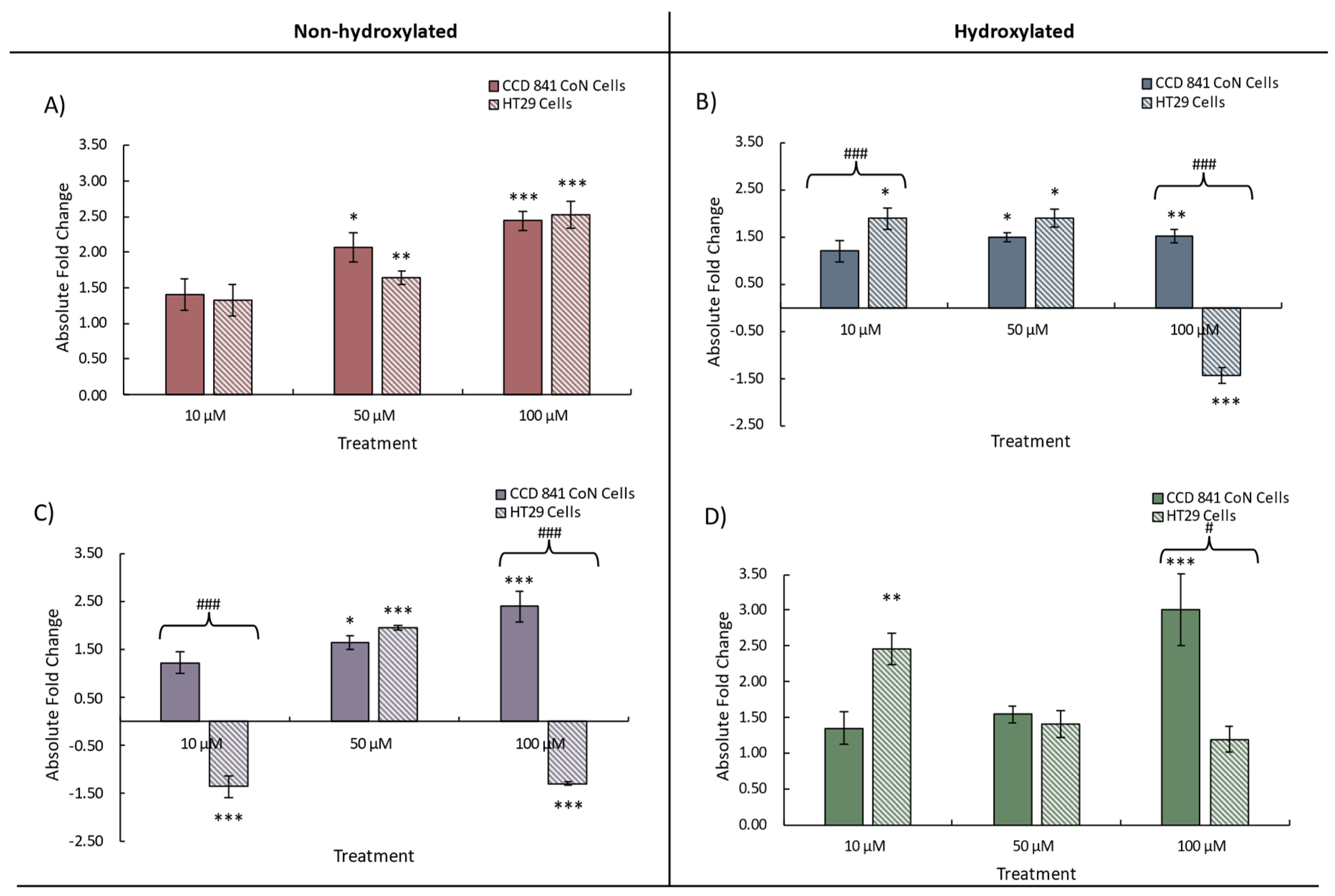
Figure 5.
Change in HO-1 expression in CCD 841 CoN and HT29 cells treated with either 10, 50, or 100 μM individual phenolics; A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA). Data presented is as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *p<0.05 **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05, ##p<0.01, ###p<0.001.
Figure 5.
Change in HO-1 expression in CCD 841 CoN and HT29 cells treated with either 10, 50, or 100 μM individual phenolics; A) benzoic acid (BA), B) 4-hydroxybenzoic acid (4HBA), C) 3-(phenyl)propanoic acid (3PPA), D) 3-(3′-hydroxyphenyl)propanoic acid (3'HPPA). Data presented is as mean of 3 independent experiments ± SEM. significance is compared to untreated control using One-way ANOVA and Dunnett’s Multiple Comparison test, *p<0.05 **p<0.01, ***p<0.001, significance compared between cell types using an independent t-test, #p<0.05, ##p<0.01, ###p<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



