
Submitted:
09 August 2023
Posted:
09 August 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
SARS-CoV-2, the virus that causes theCOVID-19 disease, has been demonstrated to cause immune suppression in certain individuals. This can manifest as a reduced ability for the host's immune system to effectively control the infection. Studies have reported that patients with COVID-19 can exhibit a decline in white blood cell counts, including natural killer cells and T cells, which are integral components of the immune system's response to viral pathogens. These cells play critical roles in the immune response to viral infections, and their depletion can make it harder for the body to mount an effective defense against the virus. Additionally, the virus can also directly infect immune cells, further compromising their ability to function. Some individuals with severe COVID-19 pneumonia may develop a "cytokine storm," an overactive immune response that may result in tissue damage and organ malfunction. The underlying mechanisms of immune suppression in SARS-CoV-2 are not entirely comprehended at this time, and ongoing research is being conducted to gain a more comprehensive understanding. Research has shown that severe SARS-CoV-2 infection promotes the synthesis of IgG4 antibodies. In this work, we propose the hypothesis that the IgG4 antibody produced by B cells in response to infection by SARS-CoV2 generates immunological tolerance that prevents its elimination, and leads to persistence and chronic infection. In sum, we believe that this constitutes another immune evasion mechanism that bears striking similarities to that developed by cancer cells to evade immune surveillance.
Keywords:
SARS-CoV-2
; IgG4
; Tregs
; long COVID
; immune tolerance
1. Introduction
Viruses are extremely complex molecular entities that can go through intra-host evolution to evolve tactics that successfully subvert the host immune system and sustain chronic infection by constantly reproducing, creating latent reservoirs, or merging into the genome of the host cell. The virus's persistent immune stimulation and/or pathologic effects will last until the infection resolves or the host is killed if the immune system is unable to eliminate the virus [1]. Some viruses can persist within a host for long periods by establishing a balance with the host's immune response. This is known as a “metastable virus-host immune response interaction equilibrium”. Examples of such viruses include the hepatitis B virus (HBV), the human immunodeficiency virus (HIV), and the hepatitis C virus (HCV). These viruses have the ability to evade the host's antiviral immunity by remaining in a latent state within host cells, which enables them to avoid identification and destruction by the immune system of the host. This allows the virus to persist within the host for prolonged periods, making it difficult to eliminate the infection [2].
In support of the possibility that SARS-CoV-2 could also establish this persistence within the host, it has been recently reported that a small number of COVID-19 patients appeared to heal from acute disease, but still progressed in their condition and eventually died [3]. Autopsies were conducted on 27 patients who appeared to have recovered from COVID-19 but whose clinical conditions had gotten progressively worse, regardless of nasopharyngeal swab viral negativity or PCR tests. Notwithstanding the notorious virological recovery, pathological changes in the lungs was comparable to that detected in acute COVID-19 patients. Forensic analysis of the lungs indicated that pneumonia was frequently the main cause of death. This research demonstrated that these individuals still had cells infected with SARS-CoV-2 in their lungs, specially in the para-bronchial glands and bronchial cartilage, notwithstanding the negative results in tests. This substantiates the hypothesis that such patients did not completely recovered from the infection. The lack of SARS-CoV-2 in the respiratory epithelium may explain why these patients appeared negative in PCR tests conducted on bronchoalveolar lavage [3]. It is also possible that the observed symptoms were caused not by the virion itself but by the viral S and/or N proteins that stimulated some subtype of marcorphages, which realsed pathological mediators and cutokines.
2. IgG4 antibodies induced by SARS-CoV-2 may help it to evade the immune system
Human Immunoglobulin G (IgG) consists of four subcategories (IgGl, IgG2, IgG3, and IgG4), which being highly conserved, differ in their constant regions, particularly in their hinges and upper CH2 domains, and are distinguished by the immunogenicity of their heavy chains [4-6]. It has been demonstrated that intrinsic characteristics of the antibody immune response, such as the antigen specificity, antigen persistence, antibody glycosylation, antibody affinity, antibody avidity, neutralizing antibody, and immunoglobulin (Ig) G subclass; i.e., specific characteristics that primarily determine the antibody durability and neutralization capacity, affect the course of SARS-CoV-2 infection and COVID-19 clinical prognosis [7-10]. A poor COVID-19 clinical evolution partly relies on an unbalanced antibody response, resulting in the increased systemic inflammation due to the defective viral neutralization [8].
There are not many publications on IgG4 production following SARS-CoV-2 infection. The IgG1 and IgG3 subclasses were the most prevalent in the COVID-19 patients [10-12]. Since IgG4 was not expected to have a role in this viral infection, many published work did not included it during estimation, especially during 2019- 2022. It has been reported that Omicron breakthrough infections further increased IgG4 production in three times mRNA-vaccinated individuals, showing that SARS-CoV-2 infection can also stimulate IgG4 synthesis [13].
A recent study sought to determine whether there would exist a connection among COVID-19-associated mortality and IgG subclasses given their various inflammatory qualities. A hospital in Italy received 138 patients with COVID-19 (41 females, 31.3%) with COVID-19 from June to December 2020. Daily data collection was used to continuously follow up on these patients. IgG1, IgG2, IgG3, and IgG4 subtypes were all increased at baseline in 8 (6%), 6, (5%), 6 (5%), and 13 (10%) individuals, respectively. 30 patients (23% of the total) passed away after 30 days [14]. Interleukin (IL)-6, C-reactive protein (CRP), and serum IgG4 levels were significantly higher in non-survivors. Notably, an IgG4/IgG1 ratio > 0.05 and a serum IgG4 concentration > 700 mg/dl were linked to a considerably higher 30-day death rate, and serum IgG4 concentration was observed to be significantly associated with IL-6 levels [14], a recognized forecaster of COVID-19-linked mortality [15-17], and which synthesis is induced by the open reading frame protein 8 (ORF8) from SARS-CoV-2 [18]. It is also known that IL-6 raises serum levels of IgG4 and other IgG subtypes [19,20]. However, we should also mention here that although Della-Torre et al. reported a compelling story showing that the serum IgG4 levels can potentially predict COVID-19 related mortality [14], it was also indicated [21] that this conclusion could be considered as quite far-reaching due to the low number of enrolled patients (128 participants) and the unique properties of IgG4 protein, levels of which may vary significantly in healthy individuals [22].
According to the research that is currently accessible, IgG4 antibodies could aggravate COVID-19 cases via at least two pathways [14]. One hypothesis is that individuals with high IgG4 levels may be more susceptible to the SARS-CoV-2 infection because anti-spike IgG4 has demonstrated weak in vitro neutralizing ability compared to IgG1, IgG2, and IgG3 antibodies [13]. On the other hand, the anti-IFN-γ auto-antibodies were identified as IgG4 subclass in people with adult-onset immunodeficiency similar to advanced HIV infection. Researchers detected a disproportionately high concentration of anti-IFN-γ auto-antibodies in 88% of Asian adults with numerous opportunistic infections [23]. It is therefore probable that the anti-IFN antibodies linked to decreased anti-SARS-CoV-2 immunity and potentially fatal COVID-19 pneumonia may be an IgG4 subclass [14]. From the start of the COVID-19 pandemic, several studies have found a relationship between preexisting autoantibodies against type I IFNs and the severity of COVID-19 [7,24-26], and these autoantibodies can inhibit the effectiveness of the corresponding type I IFNs to prevent SARS-CoV-2 infection in vitro [7] and in vivo. Although SARS-CoV-2 infection induces the synthesis of IgG4, in about 10 % of severe cases of COVID-19 pneumonia, the patients had high pre-existing levels of neutralizing auto-antibodies against type I IFNs, wich were responsible for the severity of the disease [7].
Importantly, it has been demonstrated that IFNγ significantly inhibits IL-6-induced Ig secretion [19]. This evidence suggests that in non-complicated COVID-19 cases, IFNγ is able to halt excessive IL-6 production. However, in severe cases, pre-existent autoanti-IFNγ antibodies contribute to a severe clinical course because these auto-antibodies block the normal INFγ function. In such conditions, SARS-CoV-2-induced IL-6 release rises uncontrolled, thus contributing to the development of the cytokine storm associated with severe cases of COVID-19, being one of many strategies for induction and propagation of cytokine storm.
A prospective study demonstrated that blood IgG4 levels reflect a poor COVID-19 prognosis. That research identified IgG4 antibodies as a potential additional underappreciated characteristic of antibody response against SARS-CoV-2 related to COVID-19 prognosis [14]. Another study found that patients with COVID-19 who passed away between 8-14 and 15-21 days also had higher anti-RBD (receptor binding domain) IgG4 levels than those who recovered (P < 0.05), proposing that some individuals who are in a life-threatening condition can trigger an IgG4 to RBD antibody response in the first weeks after the appearance of symptoms [11]. In that investigation, more than half of the blood samples from patients who died tested positive for IgG4 antibodies; in contrast, the majority of patients who recovered from COVID-19 tested negative for IgG4 throughout the same period [11].
The principal antibody responses to many viral illnesses are mainly represented by the IgG1 and IgG3 subclasses [27,28]. More recently, these subclasses have also been linked to the SARS-CoV-2 infection [29,30]. On the other hand, it was pointed out that IgG4 generally represents a minor component of the total immunoglobulin response and is mostly induced in response to continuous antigenic stimulation [31]. In fact, in viral respiratory illness, IgG1 and IgG3 responses are connected to immunological processes like viral neutralization, opsonization, and complement activation [27]. One research group examined the SARS-CoV-2 RBD IgG isotypes in sera from non-critical COVID-19 patients. As predicted, a strong IgG1 and IgG3 antibody response specific to RBD predominated in contrast to weaker IgG4 responses [11]. Similar to these results, it was shown that non-severe COVID-19 patients evaluated in the USA generated RBD-specific IgG1 and IgG3 early during acute infection with no discernible IgG2 or IgG4 production [29]. Comparable antibody reactions with significant IgG1 and IgG3 reactivity in sera from individuals who tested positive for SARS-CoV-2 infection were also documented in Italy [30]. On the other hand, it was also indicated that SARS-CoV-2 infection induces IgG4 production, and high serum IgG4 concentration > 700 mg/dl were linked to a considerably higher 30-day death rate, and it was significantly correlated with IL-6 levels [14], a known forecaster of COVID-19-related mortality [15-17]. We suggest that the IgG4 antibodies induced by infection with SARS-CoV-2 exacerbate a pre-existing condition with elevated levels of IgG4 autoantibodies in some individuals, thus complicating the clinical outcome and causing a higher mortality rate.
3. Mechanisms of the IgG4-induced immune evasion in SARS-CoV-2 infection
In this work, we propose the hypothesis that SARS-CoV-2 induces IgG4 synthesis to promote immune tolerance, thus evading immune surveillance and permitting unopposed viral replication. To gain insights into how IgG4 could be associated with a virus-induced immune suppression mechanism, it is necessary to comprehend how this antibody is produced, which cells produce it, and what the other immune mechanisms involved are. IgG4 is known to be produced by a variety of immune cells, including T helper 2 (Th2) cells, regulatory T cells (Treg), and regulatory B cells (Breg) when the cytokine IL-10 levels are elevated [32]. Regulatory T cells (Tregs) are necessary to preserve immunologic homeostasis, self-tolerance, and to stop uncontrolled immunological reactions [33]. Through a variety of effector pathways, Tregs control the activity of various innate and adaptive immune system branches [34]. Additionally, specific populations of "tissue Tregs" regulate homeostasis in a variety of nonimmunological tissues, reducing inflammation and fostering proper tissue regeneration [35]. Tregs, on the other hand, can be harmful, as indicated by their suppression of efficient cytotoxic responses in cancer, where they assume a unique phenotype [36-38]. They may also produce contradictory outcomes on antiviral responses [39,40].
In severe COVID-19 patients admitted to the intensive care unit (ICU), increased numbers of Tregs were found, and these cells expressed high levels of the transcription factor FoxP3, which were associated with negative results. These aberrant Tregs contribute to COVID-19 physiopathology, as patients with fewer Tregs and FoxP3 had better clinical outcomes [41]. Interestingly, these abnormal T-regs had a startling resemblance to T-regs that infiltrate tumors and suppress anticancer responses. These findings imply that Tregs may have negative effects on COVID-19 by inhibiting antiviral T-cell responses through the disease's acute phase and by directly promoting inflammation. The increase in Tregs was induced by IL-6 [41], a cytokine-induced by the ORF8 protein from SARS-CoV-2 [18]. IL-6 is necessary to increase the immune suppression ability of the RORγ+ Treg subpopulation [42].
All these data suggest that SARS-CoV-2 has developed another immune evasion mechanism that is very similar to that of cancer cells by promoting RORγ+ Treg-mediated immune tolerance. In the previously cited study [41], researchers focused their investigation on elements that Tregs may coincide in both malignancies and SARS-CoV2 infection. One such component was hypoxia, which is a characteristic of cancers and a significant contributor to severe COVID-19 [43] and can enhance Treg suppressive activity [44]. Accordingly, significant levels of lactic acid are found in COVID-19 patients [45], as well as in tumors, where they have been shown to impact Tregs function [46]. Finally, since all samples from severely affected patients profiled were obtained during the cytokine storm phase, it is possible that COVID-19 Tregs excessively inhibit the antiviral response, leading to a secondary re-expansion of the disease. This possibility is supported by the overexpression of FoxP3 and Treg effector molecules as well as by their resemblance to suppressive tumor Tregs [41].
4. Conclusions
To combat infections, the immune system has developed a variety of innate and adaptive strategies. Similarly, to guarantee their permanence within the host, viruses have developed sophisticated mechanisms to elude the immune system, generating chronic infections that are rarely eliminated [47]. We propose that when SARS-CoV infects the respiratory system, the viral ORF8 protein induces IL-6 production, thus altering the Breg lymphocytes normal phenotype. These cells then synthesize high amounts of IgG4 antibodies. This antibody has a weak in vitro neutralizing potential compared to IgG1, IgG2, and IgG3 antibodies [13,28,29]. Recent studies confirmed that a class change from IgG3 to IgG4 was linked to a decreased ability of the spike-specific antibodies to trigger complement deposition and antibody-dependent cellular phagocytosis [13]. The proposed mechanism by which the IgG4 antibody can block IgG3 attachment to its Fc receptor, thus inhibiting viral phagocytosis, is shown in Figure 1. This model is supported by the known capability of human IgG4 to interact with the Fc parts of all IgG subclasses [48].
Figure 1.
A. In normal conditions, the IgG3 antibody binds to the spike protein using its variable region. This antibody has a constant region (Fc) that is recognized by the corresponding receptor found on macrophages and other immune cells. This mechanism is called “opsonization”, and it marks foreign pathogens for phagocyte destruction. B. SARS-CoV-2 induces IL-6 production, altering the normal Treg or Breg cells phenotype and making these cells produce IgG4 antibodies (depicted in red). The constant region of the IgG4 antibody binds to the constant region of IgG3, thus preventing the binding of said antibody to its receptor located on the macrophage. In this way, the IgG3 effector functions are blocked.
Figure 1.
A. In normal conditions, the IgG3 antibody binds to the spike protein using its variable region. This antibody has a constant region (Fc) that is recognized by the corresponding receptor found on macrophages and other immune cells. This mechanism is called “opsonization”, and it marks foreign pathogens for phagocyte destruction. B. SARS-CoV-2 induces IL-6 production, altering the normal Treg or Breg cells phenotype and making these cells produce IgG4 antibodies (depicted in red). The constant region of the IgG4 antibody binds to the constant region of IgG3, thus preventing the binding of said antibody to its receptor located on the macrophage. In this way, the IgG3 effector functions are blocked.
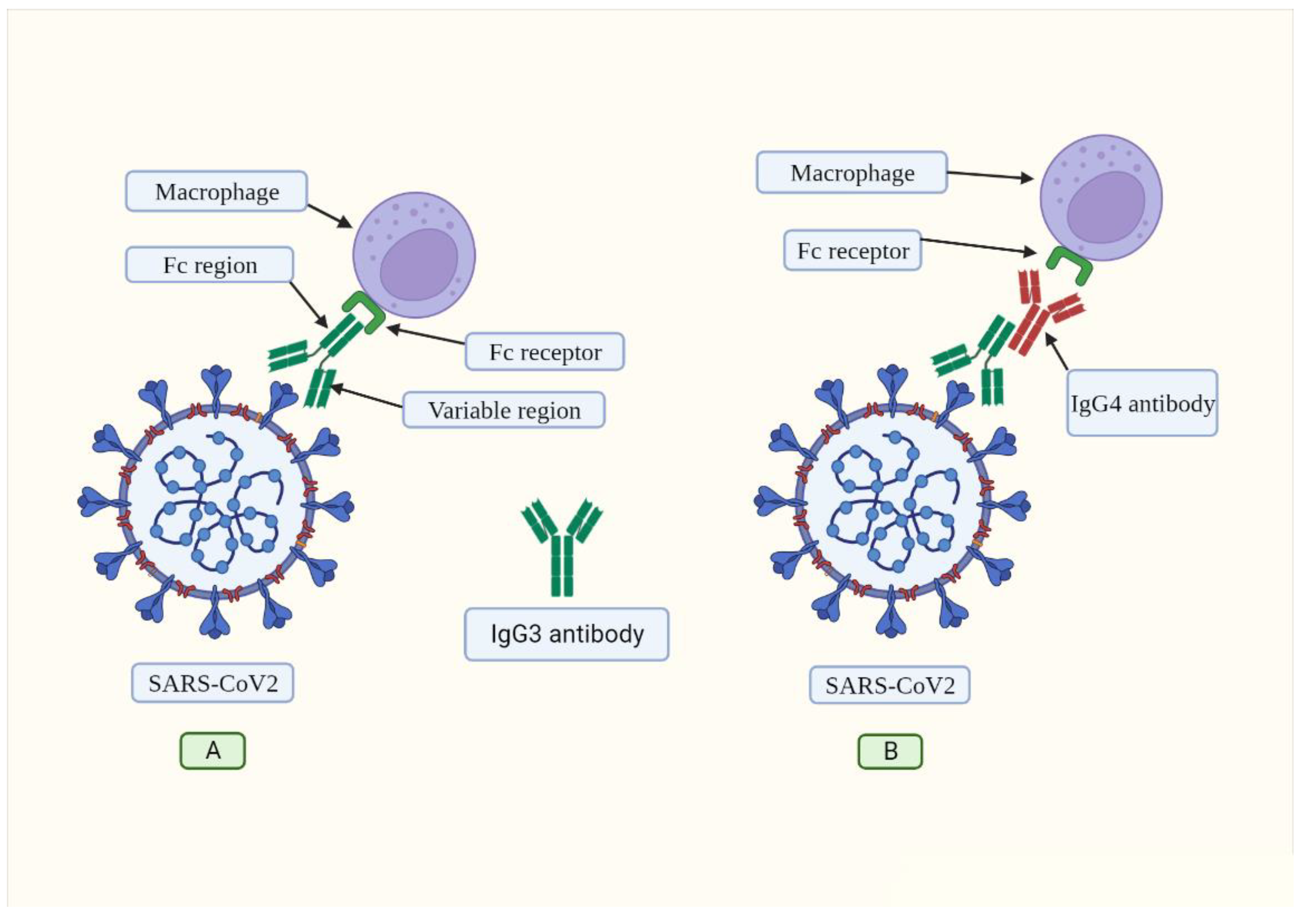
By using this IgG4-mediated tolerance mechanism, SARS-CoV-2 avoids being detected and attacked by the immune system. In other words, our immune system is hijacked and forced to “tolerate” or “ignore” the virus, thus possibly allowing chronic infection. The IgG4-induced tolerance could produce a poor immune response against SARS-CoV-2 when these patients suffer a re-infection. The virus could infect cells for a long time, causing chronic infection.
Finally, the probable IgG4-mediated evasion mechanism induced by SARS-CoV2 is remarkably similar to that evolved by cancer cells to avoid immune surveillance and attack. Researchers have investigated malignant melanoma and discovered that tumor-specific IgG4 was produced locally in tumor tissues. Additionally, they discovered that, in opposition to cancer-specific IgG1, cancer-specific IgG4 did not activate two immunological mechanisms that use antibodies to recognize and eliminate cancer cells [49]. Furthermore, the IgG1 antibody prevented the spread of cancer in an in vivo model but IgG4 did not. While IgG1 antibodies are responsible for mediating tumor cell death, IgG4 antibodies can obstruct this process. These results highlight an aspect of tumor-driven immunological escape that has not previously been studied: IgG4 production generated by tumors restricts effector immune cell activity against them [49]. The IgG4 antibody is significant and required for cancer immune evasion, according to another study [50]. B cells with high IgG4 concentrations were found in both patient serum samples and malignant cells in a cohort of people with esophageal cancer. Both increased cancer malignancy and a bad prognosis were highly correlated with higher IgG4 levels, which appear to be linked to more aggressive cancer growth. It was found that, when attaching to Fc receptors found in some immune cells in vitro, IgG4 can compete with IgG1 (as depicted in Figure 2). The normal immune responses against cancer cells, such as cell and complement cytotoxicity and cell phagocytosis, which are mediated by IgG1 antibodies, are inhibited as a result of this competition [50].
Figure 2.
Schematic representation of the hypothesized mechanism for immune evasion by cancer cells using B lymphocyte-produced IgG4 . Long-term contact with tumour antigens induces B cells to switch class and produce IgG4. Because of its unique structural and biological characteristics, increased IgG4 in the cancer microenvironment creates an effective immune evasion mechanism for the disease. The terms antibody-dependent cell mediated cytotoxicity (ADCC), antibody-dependent cell phagocytosis (ADCP), complement-dependent cytotoxicity (CDC), and natural killer cells (NK) are abbreviations for these processes, respectively. Adapted from [50]. This article is open access and is distributed under the Creative Commons Attribution Non-Commercial (CC BY-NC 4.0) license, which enables others to distribute, remix, adapt, and build upon it for non-commercial purposes and to license their derivative works under different conditions as long as the original work is properly cited, due credit, any changes are noted, and the use is for non-commercial purposes.
Figure 2.
Schematic representation of the hypothesized mechanism for immune evasion by cancer cells using B lymphocyte-produced IgG4 . Long-term contact with tumour antigens induces B cells to switch class and produce IgG4. Because of its unique structural and biological characteristics, increased IgG4 in the cancer microenvironment creates an effective immune evasion mechanism for the disease. The terms antibody-dependent cell mediated cytotoxicity (ADCC), antibody-dependent cell phagocytosis (ADCP), complement-dependent cytotoxicity (CDC), and natural killer cells (NK) are abbreviations for these processes, respectively. Adapted from [50]. This article is open access and is distributed under the Creative Commons Attribution Non-Commercial (CC BY-NC 4.0) license, which enables others to distribute, remix, adapt, and build upon it for non-commercial purposes and to license their derivative works under different conditions as long as the original work is properly cited, due credit, any changes are noted, and the use is for non-commercial purposes.
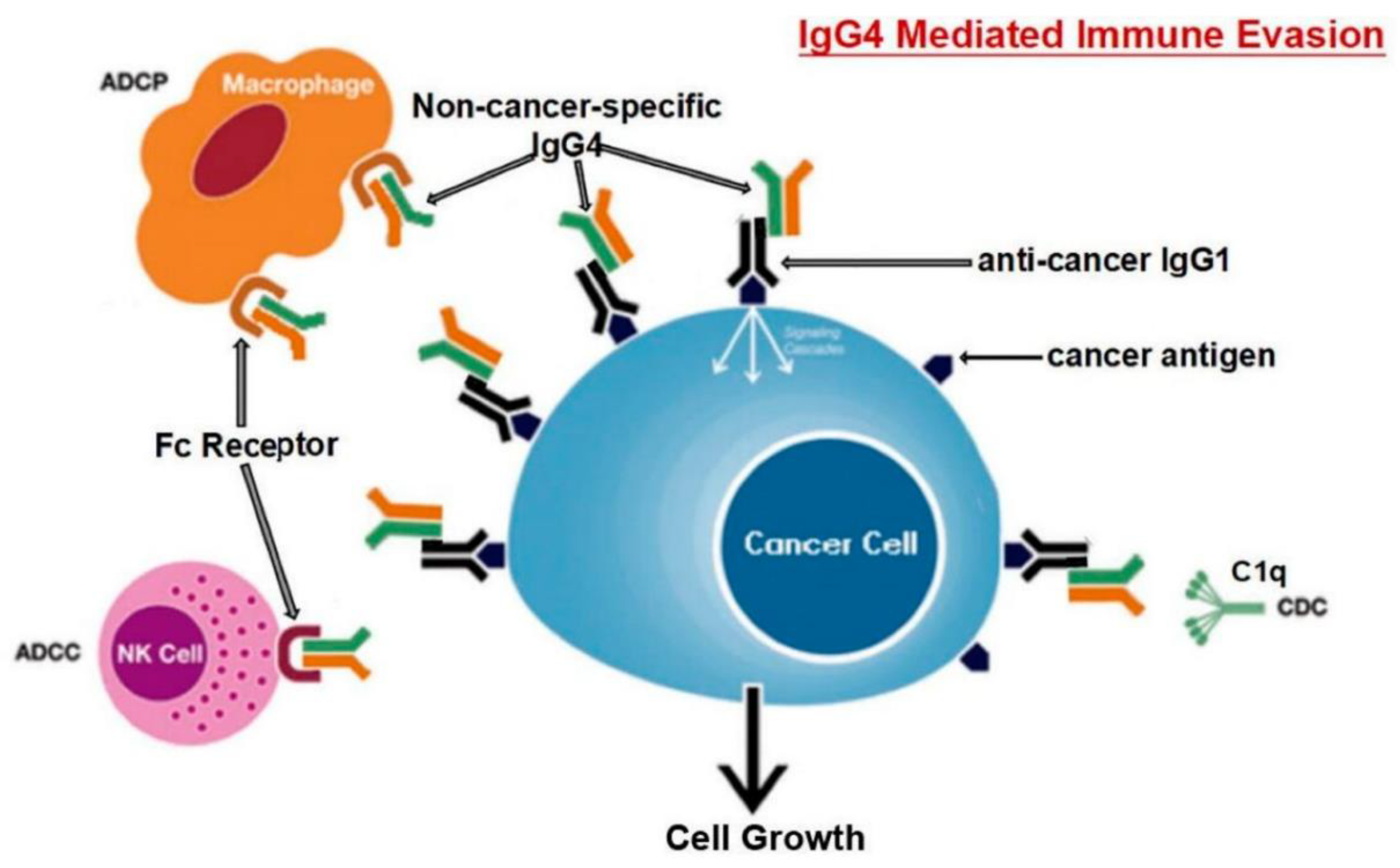
The similarity between our proposed mechanism for IgG4-mediated immune evasion by SARS-CoV-2 and that evolved by cancer cells suggest that viruses are more complex than they are usually believed. It is not known if it were viruses or cancer cells that first developed this ingenious evasion mechanism, but we are convinced that the study of the molecular mechanisms that govern these processes could have therapeutic implications both for the treatment of viral infections and for cancer.
Author Contributions
Conceptualization, A.R.C.; Investigation, A.R.C. and V.N.U.; Data curation, A.R.C. and V.N.U.; Formal analysis, A.R.C., E.M.R., and V.N.U.; Validation, A.R.C., E.M.R., and V.N.U.; Visualization, A.R.C.; Project administration, A.R.C.; Writing – original draft, A.R.C., E.M.R., and V.N.U.; Writing – review and editing, A.R.C., E.M.R., and V.N.U. The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Desimmie, B.A.; Raru, Y.Y.; Awadh, H.M.; He, P.; Teka, S.; Willenburg, K.S. Insights into SARS-CoV-2 Persistence and Its Relevance. Viruses 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Y.; Moorman, J.P.; Yao, Z.Q.; Jia, Z.S. Viral (hepatitis C virus, hepatitis B virus, HIV) persistence and immune homeostasis. Immunology 2014, 143, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Bussani, R.; Zentilin, L.; Correa, R.; Colliva, A.; Silvestri, F.; Zacchigna, S.; Collesi, C.; Giacca, M. Persistent SARS-CoV-2 infection in patients seemingly recovered from COVID-19. J Pathol 2023, 259, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Grey, H.M.; Kunkel, H.G. H Chain Subgroups of Myeloma Proteins and Normal 7s Gamma-Globulin. J Exp Med 1964, 120, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Terry, W.D.; Fahey, J.L. Subclasses of Human Gamma-2-Globulin Based on Differences in the Heavy Polypeptide Chains. Science 1964, 146, 400–401. [Google Scholar] [CrossRef] [PubMed]
- Mayumi, M.; Kuritani, T.; Kubagawa, H.; Cooper, M.D. IgG subclass expression by human B lymphocytes and plasma cells: B lymphocytes precommitted to IgG subclass can be preferentially induced by polyclonal mitogens with T cell help. J Immunol 1983, 130, 671–677. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Beziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.E.; Katsaounou, P.; et al. Complex Immune Dysregulation in COVID-19 Patients with Severe Respiratory Failure. Cell Host Microbe 2020, 27, 992–1000. [Google Scholar] [CrossRef]
- Chakraborty, S.; Gonzalez, J.; Edwards, K.; Mallajosyula, V.; Buzzanco, A.S.; Sherwood, R.; Buffone, C.; Kathale, N.; Providenza, S.; Xie, M.M.; et al. Proinflammatory IgG Fc structures in patients with severe COVID-19. Nat Immunol 2021, 22, 67–73. [Google Scholar] [CrossRef]
- Yates, J.L.; Ehrbar, D.J.; Hunt, D.T.; Girardin, R.C.; Dupuis, A.P., 2nd; Payne, A.F.; Sowizral, M.; Varney, S.; Kulas, K.E.; Demarest, V.L.; et al. Serological analysis reveals an imbalanced IgG subclass composition associated with COVID-19 disease severity. Cell Rep Med 2021, 2, 100329. [Google Scholar] [CrossRef]
- Moura, A.D.; da Costa, H.H.M.; Correa, V.A.; de, S.L.A.K.; Lindoso, J.A.L.; De Gaspari, E.; Hong, M.A.; Cunha-Junior, J.P.; Prudencio, C.R. Assessment of avidity related to IgG subclasses in SARS-CoV-2 Brazilian infected patients. Sci Rep 2021, 11, 17642. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Jia, T.; Chen, J.; Zeng, S.; Qiu, Z.; Wu, S.; Li, X.; Lei, Y.; Wang, X.; Wu, W.; et al. The Characterization of Disease Severity Associated IgG Subclasses Response in COVID-19 Patients. Front Immunol 2021, 12, 632814. [Google Scholar] [CrossRef]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schafer, S.; Zhong, J.; et al. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci Immunol 2023, 8, eade2798. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Lanzillotta, M.; Strollo, M.; Ramirez, G.A.; Dagna, L.; Tresoldi, M.; group, C.O.-B.s. Serum IgG4 level predicts COVID-19 related mortality. Eur J Intern Med 2021, 93, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Campochiaro, C.; Cavalli, G.; De Luca, G.; Napolitano, A.; La Marca, S.; Boffini, N.; Da Prat, V.; Di Terlizzi, G.; Lanzillotta, M.; et al. Interleukin-6 blockade with sarilumab in severe COVID-19 pneumonia with systemic hyperinflammation: an open-label cohort study. Ann Rheum Dis 2020, 79, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Della-Torre, F.; Kusanovic, M.; Scotti, R.; Ramirez, G.A.; Dagna, L.; Tresoldi, M. Treating COVID-19 with colchicine in community healthcare setting. Clin Immunol 2020, 217, 108490. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Lanzillotta, M.; Campochiaro, C.; Cavalli, G.; De Luca, G.; Tomelleri, A.; Boffini, N.; De Lorenzo, R.; Ruggeri, A.; Rovere-Querini, P.; et al. Respiratory Impairment Predicts Response to IL-1 and IL-6 Blockade in COVID-19 Patients With Severe Pneumonia and Hyper-Inflammation. Front Immunol 2021, 12, 675678. [Google Scholar] [CrossRef]
- Wu, X.; Xia, T.; Shin, W.J.; Yu, K.M.; Jung, W.; Herrmann, A.; Foo, S.S.; Chen, W.; Zhang, P.; Lee, J.S.; et al. Viral Mimicry of Interleukin-17A by SARS-CoV-2 ORF8. mBio 2022, 13, e0040222. [Google Scholar] [CrossRef]
- Bertolini, J.N.; Benson, E.M. The role of human interleukin-6 in B-cell isotype regulation and differentiation. Cell Immunol 1990, 125, 197–209. [Google Scholar] [CrossRef]
- Kawano, Y.; Noma, T.; Kou, K.; Yoshizawa, I.; Yata, J. Regulation of human IgG subclass production by cytokines: human IgG subclass production enhanced differentially by interleukin-6. Immunology 1995, 84, 278–284. [Google Scholar]
- Dulak, N.A.; Trzcinski, R. Disguised aspects of IgG4. Eur J Intern Med 2022, 95, 97. [Google Scholar] [CrossRef] [PubMed]
- Nirula, A.; Glaser, S.M.; Kalled, S.L.; Taylor, F.R. What is IgG4? A review of the biology of a unique immunoglobulin subtype. Curr Opin Rheumatol 2011, 23, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Browne, S.K.; Burbelo, P.D.; Chetchotisakd, P.; Suputtamongkol, Y.; Kiertiburanakul, S.; Shaw, P.A.; Kirk, J.L.; Jutivorakool, K.; Zaman, R.; Ding, L.; et al. Adult-onset immunodeficiency in Thailand and Taiwan. N Engl J Med 2012, 367, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bastard, P.; Bolze, A.; Jouanguy, E.; Zhang, S.Y.; Effort, C.H.G.; Cobat, A.; Notarangelo, L.D.; Su, H.C.; Abel, L.; et al. Life-Threatening COVID-19: Defective Interferons Unleash Excessive Inflammation. Med 2020, 1, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370. [Google Scholar] [CrossRef] [PubMed]
- Arrestier, R.; Bastard, P.; Belmondo, T.; Voiriot, G.; Urbina, T.; Luyt, C.E.; Gervais, A.; Bizien, L.; Segaux, L.; Ben Ahmed, M.; et al. Auto-antibodies against type I IFNs in > 10% of critically ill COVID-19 patients: a prospective multicentre study. Ann Intensive Care 2022, 12, 121. [Google Scholar] [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Mendez, N.V.; Landin, A.M.; Blomberg, B.B. Effects of age on H1N1-specific serum IgG1 and IgG3 levels evaluated during the 2011-2012 influenza vaccine season. Immun Ageing 2013, 10, 14. [Google Scholar] [CrossRef]
- Cavacini, L.A.; Kuhrt, D.; Duval, M.; Mayer, K.; Posner, M.R. Binding and neutralization activity of human IgG1 and IgG3 from serum of HIV-infected individuals. AIDS Res Hum Retroviruses 2003, 19, 785–792. [Google Scholar] [CrossRef]
- Suthar, M.S.; Zimmerman, M.G.; Kauffman, R.C.; Mantus, G.; Linderman, S.L.; Hudson, W.H.; Vanderheiden, A.; Nyhoff, L.; Davis, C.W.; Adekunle, O.; et al. Rapid Generation of Neutralizing Antibody Responses in COVID-19 Patients. Cell Rep Med 2020, 1, 100040. [Google Scholar] [CrossRef]
- Mazzini, L.; Martinuzzi, D.; Hyseni, I.; Benincasa, L.; Molesti, E.; Casa, E.; Lapini, G.; Piu, P.; Trombetta, C.M.; Marchi, S.; et al. Comparative analyses of SARS-CoV-2 binding (IgG, IgM, IgA) and neutralizing antibodies from human serum samples. J Immunol Methods 2021, 489, 112937. [Google Scholar] [CrossRef]
- Aalberse, R.C.; Dieges, P.H.; Knul-Bretlova, V.; Vooren, P.; Aalbers, M.; van Leeuwen, J. IgG4 as a blocking antibody. Clin Rev Allergy 1983, 1, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Satoguina, J.S.; Weyand, E.; Larbi, J.; Hoerauf, A. T regulatory-1 cells induce IgG4 production by B cells: role of IL-10. J Immunol 2005, 174, 4718–4726. [Google Scholar] [CrossRef] [PubMed]
- Josefowicz, S.Z.; Lu, L.F.; Rudensky, A.Y. Regulatory T cells: mechanisms of differentiation and function. Annu Rev Immunol 2012, 30, 531–564. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat Rev Immunol 2008, 8, 523–532. [Google Scholar] [CrossRef]
- Panduro, M.; Benoist, C.; Mathis, D. Tissue Tregs. Annu Rev Immunol 2016, 34, 609–633. [Google Scholar] [CrossRef]
- De Simone, M.; Arrigoni, A.; Rossetti, G.; Gruarin, P.; Ranzani, V.; Politano, C.; Bonnal, R.J.P.; Provasi, E.; Sarnicola, M.L.; Panzeri, I.; et al. Transcriptional Landscape of Human Tissue Lymphocytes Unveils Uniqueness of Tumor-Infiltrating T Regulatory Cells. Immunity 2016, 45, 1135–1147. [Google Scholar] [CrossRef]
- Plitas, G.; Konopacki, C.; Wu, K.; Bos, P.D.; Morrow, M.; Putintseva, E.V.; Chudakov, D.M.; Rudensky, A.Y. Regulatory T Cells Exhibit Distinct Features in Human Breast Cancer. Immunity 2016, 45, 1122–1134. [Google Scholar] [CrossRef]
- Magnuson, A.M.; Kiner, E.; Ergun, A.; Park, J.S.; Asinovski, N.; Ortiz-Lopez, A.; Kilcoyne, A.; Paoluzzi-Tomada, E.; Weissleder, R.; Mathis, D.; et al. Identification and validation of a tumor-infiltrating Treg transcriptional signature conserved across species and tumor types. Proc Natl Acad Sci U S A 2018, 115, E10672–E10681. [Google Scholar] [CrossRef]
- Lund, J.M.; Hsing, L.; Pham, T.T.; Rudensky, A.Y. Coordination of early protective immunity to viral infection by regulatory T cells. Science 2008, 320, 1220–1224. [Google Scholar] [CrossRef]
- Almanan, M.; Raynor, J.; Sholl, A.; Wang, M.; Chougnet, C.; Cardin, R.D.; Hildeman, D.A. Tissue-specific control of latent CMV reactivation by regulatory T cells. PLoS Pathog 2017, 13, e1006507. [Google Scholar] [CrossRef]
- Galvan-Pena, S.; Leon, J.; Chowdhary, K.; Michelson, D.A.; Vijaykumar, B.; Yang, L.; Magnuson, A.M.; Chen, F.; Manickas-Hill, Z.; Piechocka-Trocha, A.; et al. Profound Treg perturbations correlate with COVID-19 severity. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Hagenstein, J.; Melderis, S.; Nosko, A.; Warkotsch, M.T.; Richter, J.V.; Ramcke, T.; Herrnstadt, G.R.; Scheller, J.; Yan, I.; Mittrucker, H.W.; et al. A Novel Role for IL-6 Receptor Classic Signaling: Induction of RORgammat(+)Foxp3(+) Tregs with Enhanced Suppressive Capacity. J Am Soc Nephrol 2019, 30, 1439–1453. [Google Scholar] [CrossRef] [PubMed]
- Dhont, S.; Derom, E.; Van Braeckel, E.; Depuydt, P.; Lambrecht, B.N. The pathophysiology of 'happy' hypoxemia in COVID-19. Respir Res 2020, 21, 198. [Google Scholar] [CrossRef] [PubMed]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.P.; Gimotty, P.A.; Gilks, C.B.; Lal, P.; Zhang, L.; et al. Tumour hypoxia promotes tolerance and angiogenesis via CCL28 and T(reg) cells. Nature 2011, 475, 226–230. [Google Scholar] [CrossRef] [PubMed]
- McElvaney, O.J.; McEvoy, N.L.; McElvaney, O.F.; Carroll, T.P.; Murphy, M.P.; Dunlea, D.M.; Ni Choileain, O.; Clarke, J.; O'Connor, E.; Hogan, G.; et al. Characterization of the Inflammatory Response to Severe COVID-19 Illness. Am J Respir Crit Care Med 2020, 202, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.J.; Vignali, P.D.A.; Mullett, S.J.; Overacre-Delgoffe, A.E.; Peralta, R.M.; Grebinoski, S.; Menk, A.V.; Rittenhouse, N.L.; DePeaux, K.; Whetstone, R.D.; et al. Metabolic support of tumour-infiltrating regulatory T cells by lactic acid. Nature 2021, 591, 645–651. [Google Scholar] [CrossRef]
- Janeway Jr, C.A.; Travers, P.; Walport, M.; Shlomchik, M.J. Pathogens have evolved various means of evading or subverting normal host defenses. In Immunobiology: The Immune System in Health and Disease. 5th edition, Garland Science: 2001.
- Rispens, T.; Meesters, J.; den Bleker, T.H.; Ooijevaar-De Heer, P.; Schuurman, J.; Parren, P.W.; Labrijn, A.; Aalberse, R.C. Fc-Fc interactions of human IgG4 require dissociation of heavy chains and are formed predominantly by the intra-chain hinge isomer. Mol Immunol 2013, 53, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, P.; Gilbert, A.E.; Josephs, D.H.; Ali, N.; Dodev, T.; Saul, L.; Correa, I.; Roberts, L.; Beddowes, E.; Koers, A.; et al. IgG4 subclass antibodies impair antitumor immunity in melanoma. J Clin Invest 2013, 123, 1457–1474. [Google Scholar] [CrossRef]
- Wang, H.; Xu, Q.; Zhao, C.; Zhu, Z.; Zhu, X.; Zhou, J.; Zhang, S.; Yang, T.; Zhang, B.; Li, J.; et al. An immune evasion mechanism with IgG4 playing an essential role in cancer and implication for immunotherapy. J Immunother Cancer 2020, 8. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



