
Submitted:
09 August 2023
Posted:
10 August 2023
You are already at the latest version
Abstract
Phytoalexins are naturally-occurring molecules with antimicrobial activity deriving from the secondary metabolism of plants, which are synthesized in response to physical agents or stresses, phytopathogenic agents (fungi, bacteria and viruses) as well as numerous chemical compounds and biological control agents. Among these, grapevine phytoalexins, which belong to the chem-ical group of stilbenes, exhibit biocidal activity against a large number and a variety of plant pathogens. An important issue in research on these compounds is to be able to stimulate their synthesis in planta and to determine whether the induction of their production can serve as a basis for protecting that plant against its pathogens. Quite a few chemical compounds, derivatives of phytohormones bio-elicitors as well as biocontrol agents have been used to induce the synthesis of stilbene phytoalexins with the aim of conferring protection to grapevine against its main dis-eases (gray mold, downy mildew, powdery mildew and esca). This article reviews the experi-ments that have been carried out in this direction during the last thirty years and shows that the observed protective effects towards pathogens are generally linked to induction and priming of the grapevine phytoalexin response, confirming the interest of using, in a more general way, stimulation of the production of phytoalexins in plants as a basis for crop protection.
Keywords:
stilbenes
; phytoalexins
; resveratrol
; elicitors
; plant defenses
; chemicals
; biological control agents
1. Introduction
According to the fundamental observation made by Müller and Börger [1], 80 years ago, infection with an incompatible strain of Phytophthora infestans capable of initiating hypersensitivity reactions in potato tubers (Solanum tuberosum) significantly prevented the effect of a subsequent infection with another strain (compatible) of P. infestans. This inhibition phenomenon was linked to the synthesis by the plant of a “chemical principle” making the tissues resistant to infection, which these two researchers named “phytoalexin”. Phytoalexins are produced in a certain number of plant families, mainly and not exhaustively, Leguminosae, Fabaceae, Solanaceae, Malvaceae, Poaceae, Brassicaceae (Cruciferae) and Vitaceae without knowing, due to a gap in knowledge, whether all plants possess phytoalexins [2,3]. Phytoalexins are compounds with an antimicrobial activity generally ranging between 10 and 100 μM, with a molecular mass over 1500 Da, high molecular-ordered stilbene oligomers such as pauciflorol D having a mass of 1587.624 Da) [4,5,6]. Their production essentially corresponds to a de novo synthesis in response to various biotic and abiotic stresses. Their study has given rise over the last thirty years with the progress of structural chemistry, biochemistry and molecular biology, to a ferment of activity on the elucidation of their biosynthetic pathways and the regulation of their syntheses, finding interesting applications in the field of the engineering of these pathways in microbes and plants [2,7,8,9]. Phytoalexins exhibit a very broad spectrum of biological activity against a number of living organisms, from viruses, bacteria, fungi, plants to animals also taking part in allelochemical processes between plants. Namely, the antimicrobial function of phytoalexins and the role they play in the defense mechanisms of plants have been a central topic of study among the plant pathology community. As a result, a need has arisen to study how to induce the synthesis of these compounds in plants with the aim of elucidating the implementation of their biosynthetic mechanisms and for technological/biotechnological purposes. Of prime concern for plant pathologists is to find out if the stimulation of the synthesis of phytoalexins by different means (chemical substances, biological elicitors, beneficial microorganisms) can be of interest in terms of crop protection [10].
In the phytoalexin world, stilbene phytoalexins have received a special attention, no doubt linked to their proven role in human health as anti-cancer, antioxidant, cardio-and neuroprotective agents. Although not strictly limited to the Vitaceae, it is nevertheless in this plant family that stilbene-type phytoalexins were first identified in 1976 following the pioneering works of Langcake and Pryce [11,12,13] and that many studies have been carried out there, since. Here, we will unveil the different ways of inducing the synthesis of these compounds in grapevine during experiments carried out in vitro and in vivo, on isolated leaves or on potted vines, also describing some experiments conducted in the vineyard. The challenge is to verify whether the stimulation of stilbene phytoalexin production can contribute to protecting this plant from pathogens’ attacks with possible applications in the vineyard. More generally, extending the results described in this review may also open the use of stimulating phytoalexin synthesis as a basis for crop protection.
2. Chemistry and antifungal activity of some stilbenes of grapevine: A brief overview
A recent and comprehensive study on these aspects has recently been published [5]. Stilbenes belong to a restricted chemical group compared to the large family of flavonoids [14]. But it is in grapevine that this class of compounds presents a spectacular diversity [5,15]. In fact, 48 different coding genes have been counted in the stilbene synthase family in grapevine, of which 33 are potentially active [16,17]. We will only mention here the elements necessary to the understanding of the data presented in this review.
Building of the resveratrol core begins with phenylalanine, itself deriving from the shikimic acid and arogenic acid pathways [5,18]. Phenylalanine undergoes oxidative deamination catalyzed by phenylalanine ammonia lyase (PAL) and leading to trans-cinnamic acid (Figure 1). The latter is hydroxylated to para-coumaric acid (trans isomer) by a specific cytochrome P450 hydroxylase, C4H, para-coumaric acid being then converted into its coenzyme A thioester, para-coumaroyl-CoA, by binding with a Coenzyme A molecule via a cinnamoyl-CoA ligase (4CL). A dichotomous system unique in the plant kingdom and comprising two enzymes, chalcone synthase (CHS) and stilbene synthase (STS) using the same substrates, p-coumaroyl-CoA and three malonyl-CoA units (formed from glycolysis), affords flavonoids, on the one hand and stilbenes, on the other (Figure 1). Resveratrol is a hub, seat of an intense metabolism involving in grapevine, methylation, hydroxylation, glycosylation reactions and especially peroxidation reactions leading to the formation of the stilbene oligomers, which makes the originality of this class of compounds [6].
Certain methylated, glycosylated and hydroxylated stilbene derivatives as well as numerous oligomers have been identified in grapevine [5,15]. For all that, few enzymes catalyzing the transformation of resveratrol into its derivatives have been characterized both at the genomic and functional level. Piceatannol, a hydroxylated derivative of resveratrol, was characterized for the first time in grapevine [19] (not pictured). Though its structural relationship with resveratrol suggests that it could be synthesized directly by hydroxylation of the latter by a flavonoid-3-hydroxylase-like enzyme (F3H) as in flavonoid biosynthesis, no experimental evidence of the genesis of piceatannol from resveratrol has been reported to date. Pterostilbene, which is a dimethylated stilbene identified in 1979 by Langcake’s group [20], is undoubtedly the best-known resveratrol derivative (Figure 2). The direct biosynthesis of pterostilbene from resveratrol is catalyzed by a resveratrol-O-methyl transferase (ROMT), which was cloned from mildew-infected grape leaves and functionally characterized. This enzyme ensured the methylation of resveratrol into pterostilbene in vitro and in vivo [21]. Isorhapontigenin (not pictured) is a monomethylated stilbene whose probable origin could result from the direct methylation of piceatannol. Glycosylation (addition of one or more sugars to a given compound) is a very common reaction in polyphenol chemistry. Piceid, the 3-O-resveratrol glucoside, plays an important role in the metabolism of resveratrol where it is considered a storage form (Figure 2). Glycosylation reactions are catalyzed by glycosyl-transferases generally using uridine diphospho-glucose (UDPG) as a glucose donor and although there are 240 putative genes coding for glycosyltransferases (GTs) in grapevine [22], only two genes (VLRSgt and VvUGT72B27) encoding GTs active on resveratrol and leading to piceid synthesis have been identified in grapevine [23,24].
One of the most fascinating aspects in stilbene chemistry is the formation of oligomers; it has been brilliantly demonstrated by Stephenson and his group that resveratrol constitutes the building block of all oligomers whose levels of condensation can reach seven resveratrol units in the case of pauciflorol D. Oligomer formation initially proceeds by the oxidation of resveratrol into several radicals which then condense with each other according to defined coupling modes [4]. Oxidation of resveratrol is linked to the action of various peroxidases, some of which have previously been described in grapevine, the peroxidases A1, B2 located in the apoplastic compartment and the vacuolar peroxidase B5 [25,26,27]. The horseradish peroxidase (HRP) was employed in Langcake’s group’s pioneering works on stilbene phytoalexins reporting in this case the formation of δ-viniferin [28]. It is unknown, however, whether peroxidases are able to orientate the polymerization of resveratrol radicals towards specific oligomers, the use of these enzymes such as HRPs in vitro generally affording complex mixtures of stilbene oligomers [4,5,29]. Current hypotheses favor the involvement of chemical reactions between the radicals formed (Friedel-Crafts reactions, oxa-Michael additions) to explain these oligomerization processes [4].
Many grapevine pathogenic microorganisms can elicit both the synthesis and the accumulation of stilbene phytoalexins. Following the pioneering work of Langcake’s group, other researchers have described the synthesis of resveratrol and its derivatives in response to B. cinerea [30,31], Plasmopara viticola [32,33] Aspergillus carbonarius, an ochratoxin A-producing fungus [34] or in response to the Esca complex of diseases responsible for the leaf stripe disease symptoms in grapevine [35,36], this list obviously not being exhaustive. Conversely, the produced stilbenes exerted a biocidal activity against these pathogens.
The antifungal activity of stilbenes has recently been described elsewhere [5]. We will limit ourselves here to presenting the activity of the main stilbene monomers and of some oligomers described in grapevine. Due to their low water-solubility, the antifungal activity of stilbenes has greatly been underestimated. Langcake and Pryce [11] solubilizing resveratrol in water alone reported concentrations > 870 μM for obtaining 50% inhibition of the spore germination of B. cinerea (ED50), for example, though lower values of 438 μM were reported for the inhibition of the development of the mycelium of this fungus when minute quantities of an organic solvent (acetone) was added to the medium and even less (88 μM) for Fusarium oxysporum ([11]. Data regarding inhibition of the spore germination of B. cinerea have since been revisited by incorporating a minimum amount of ethanol into the spore incubation medium to ensure resveratrol solubility. Under these conditions, ED50 values of 390 μM were noted [37]. Fairly low inhibition values have been reported concerning the inhibition of zoospore motility of the oomycete Plasmopara viticola, responsible for grapevine downy mildew, respectively ED50 of 192 μM and ED100 500 μM [38,39]. Methylated stilbenes such as pterostilbene and isorhapontigenin, respectively derived from resveratrol and piceatannol, exhibited higher biocidal activities, in the order of 2.5 to 10 times compared to the hydroxylated compounds, depending on the type of pathogen. Several studies have reported ED50 for pterostilbene around 70 μM for the inhibition of the germination of B. cinerea spores [28,37,40] and 39–390 μM for inhibition of the fungal growth of Esca disease-associated fungi [41]. The biocidal activity of pterostilbene was more accentuated regarding the inhibition of the motility of the zoospores of P. viticola with ED50 of 9 μM [13] and 14.6 μM [38]. As confirmation of the greater biocidal activity of methylated stilbenes compared to their non-methylated counterparts, isorhapontigenin has been reported to be 2.5 times more inhibitory to the development of downy mildew (ED50 of 116 μM) than the highly hydroxylated piceatannol (ED50 of 254 μM) [42].
Surprisingly, dimeric stilbenes such as the well-known ε-viniferin showed quite high biocidal activity with ED50 of 220 and 230 μM for the respective inhibition of spore germination and mycelial development of B. cinerea [13] and even more strong activity with regard to sporulation inhibition in P. viticola (ED50 of 12.7 and 70 μM) [38,43]. Finally, several high molecular-ordered stilbenes, the trimer, miyabenol C and three tetramers, hopeaphenol, vitisin A and vitisin B exhibited remarkable inhibitory activities on the development of downy mildew with very low ED50 values: 18 μM for hopeaphenol; 40 μM for miyabenol C and, respectively, 20 and 12 μM for the vitisins A and B [43]. These data are not so surprising because stilbene oligomers possess biological activities, for example, anti-cancer, anti-arthritic properties as well as protective activities against neuroinflammatory processes, superior to resveratrol [6].
The fact that the synthesis of phytoalexins is induced by a large number of pathogenic fungi, their synthesis being possibly associated with an increase in the resistance of plants to infection, has led researchers to wonder about ways to stimulate the production of these compounds with different types of elicitors or by the use of microorganisms (essentially of bacterial origin) through numerous assays carried out in potted plants or in vivo [44]. The crucial question of transferring the data acquired in the field has been more rarely addressed [45,46]. These questions will be developed in the following sections.
3. Induction of stilbene phytoalexin synthesis by chemicals and possible applications in the vineyard
3.1. Organic and Inorganic Metallic Salts
Among the chemicals which can induce the production of phytoalexins, numerous metallic salts have been described as having the ability to trigger the synthesis of these compounds with more or less applications in the vineyard (Table 1), in particular. Fairly old studies have reported the use of heavy metallic salts such as, among the most active, silver (AgCl), mercury (HgCl2) and copper (CuCl2) salts for the induction of pisatin synthesis in pea (Pisum sativum) [47]. Also, in pea, copper chloride at a concentration of 5 mM was found to induce the activity of a methyltransferase catalyzing the methylation of (+)-6α-hydroxymaackiain into (+)-pisatin [48].
Copper chloride and other metallic salts such as copper sulfate (CuSO4), which is widely used in viticulture, have been recognized as inducers of the synthesis of several stilbene phytoalexins and flavonoid glucosides in Veratrum grandiflorum [49] and the synthesis of resveratrol, ε-viniferin and piceid in grapevine plantlets in vitro [50]. Organic or inorganic aluminum-based metallic salts have also been employed in grapevine with potential applications for the protection of this plant in the field.
The specific mechanisms of activation of the grapevine defenses by aluminium, particularly implementation of the phytoalexin system, have been deciphered very recently as we will see below. Adrian et al. [51] reported a phenomenon of hyper-elicitation of the resveratrol synthesis in isolated grapevine shoots with aluminum chloride (AlCl3) at a concentration of 7 mM in V. rupestris. In a similar way to what has been described for the biosynthesis of pisatin in pea, methylation of resveratrol to its dimethylated derivative, pterostilbene, was observed on grapevine leaf discs by induction of a resveratrol-O-methyltransferase in the presence of 1% AlCl3 [21]. Among the organic aluminum salts, fosetyl-Al has been the subject of numerous studies in plants with applications in the vineyard.
Fosetyl®-Al (Aluminum Tris-O-ethylphosphonate) is a well-known fungicide (Aliette, Bayer Crop Science) which has shown in practice a fair efficacy against downy mildews and, particularly, grapevine downy mildew (P. viticola) [52,53,54,55,56] but which was also applied for the treatment of Esca [56]. Very rapidly, works sought to highlight a relationship between the protective action of fosetyl-Al and a stimulation of the defense mechanisms in tobacco [57,58] and more specifically of the phytoalexin system in tobacco [59], citrus [60] and in grapevine [61,62]. These latest works, carried out on detached grapevine leaves or on leaf discs, have reported that it was difficult to distinguish which part could be attributed to the stimulation of phytoalexin synthesis following infection by sporangial suspensions of P. viticola and which part was due to the sole application of the fungicide. For Raynal et al. [61], fosetyl-Al alone did not induce phytoalexin synthesis in grapevine (resveratrol and cis-ε-viniferin), whereas these compounds saw their concentration increasing in the tissues when this fungicide was applied simultaneously at—or 24 h after—infection (priming effect) (Table 1). For Dercks and Creasy [62], interpretation of the results was even more complex. The sole application of fosetyl-Al induced resveratrol production but at modest levels (4.7 μg/g fresh weight). Post-infectional applications of fosetyl-Al, that is, 22 h after inoculation with P. viticola, led to a significant rise in resveratrol amounts (19.1 μg/g fresh weight in the susceptible variety, Riesling; 69 μg/g fresh weight in the mid-resistant species, V. rupestris and 107.38 μg/g fresh weight in the resistant variety, Castor). On the other hand, the 2 to 4 times increase in the content of resveratrol compared to the control during the pre-infectional application of fosetyl-Al (24 h before inoculation by P. viticola), was particularly interesting since it suggested that prior application of the fungicide displayed a priming effect on the synthesis of resveratrol and implementation of the phytoalexin response [62]. Fosetyl-Al efficacy towards the sporulation of P. viticola depended on the date of application of the treatment, the dose used and above all, the grape vine variety. The doses of fosetyl-Al required to totally suppress sporulation at the pre-infectional stage varied from 200 μg/mL for V. rupestris (mid-tolerant and the cv Castor (resistant) and 400 μg/mL for the cv Riesling (susceptible). At the post-infectional stage, the sensitive variety was no longer protected by fosetyl-al regardless of the dose, V. rupestris was protected only with the highest dose while the resistant variety (Castor) remained protected by both treatment doses.
To appreciate, the capacity of aluminium to trigger stilbene phytoalexin synthesis in grapevine, tests were carried out in the 90 s jointly by the University of Dijon and Goemar Laboratories on a fungicide called, Synermix (Table 1). This contained aluminum chloride in combination with a seaweed extract. Previously, trials conducted in compliance with regional treatment programs for the fight against gray mold in the vineyards of Bordeaux, Champagne and the Val de Loire over an eight-year period revealed an average efficacy of this fungicide of 33.6% against B. cinerea [63,64].

Table 1.
Induction of stilbene phytoalexin synthesis by chemicals associated with resistance of grapevine to diseases.
Table 1.
Induction of stilbene phytoalexin synthesis by chemicals associated with resistance of grapevine to diseases.
| Chemicals | Plant Material | Biological Inputs | References |
|---|---|---|---|
| Fosetyl-Al | Potted grapevine (cv Carignan) | Protection towards P. viticola infection. No effect of fosetyl-Al alone on stilbene accumulation but active when applied at the time of infection or shortly after | [61] |
| Fosetyl-Al | cv Riesling (susceptible), V. rupestris (mid-tolerant) and cv Castor (resistant) leaf discs | Total suppression of P. viticola sporulation for pre-infectional applications. No protection towards the susceptible cultivar with post-infectional applications. Induction of stilbene production and priming effect | [62] |
| Synermix (aluminum chloride in combination with a seaweed extract) | Eight-year period experiments in the vineyard (V. vinifera) | Efficacy of 33.6% towards B. cinerea infection vs. Iprodione (42.2%). 70.7% efficacy for the combination Synermix/Iprodione. High AlCl3-induced resveratrol production | [51,64] |
| Copper sulfate | In vitro plantlets and potted grapevine (cv Chardonnay) | Protection against B. cinerea infection (> 60%) and only 38% towards P. viticola associated with an induction of chitinase and β-1,3-glucanases along with stilbene production | [50] |
| Fosetyl-Al | Nine-year period experiments in the vineyard (cvs Albana and Sangiovese) | Reduction of the foliar symptoms of esca decorrelated with stilbene production | [56] |
At the same time, application of a fungicide known for the treatment of gray mold, iprodione, demonstrated an effectiveness close to 42.2%. Interestingly, use of the two fungicide products in combination showed an efficacy in the vineyard of 70.7%, that is, a synergistic effect of +9.1% compared to the calculated theoretical efficacy of the combination of the two fungicides according to Colby’s formula [65], where TE is the theoretical efficacy of the combination of the two fungicides; α and β are respectively the efficacy of fungicides A and B measured in the fields. The synergy is deduced by subtracting the efficacy of the fungicide combination in the fields to the theoretical efficacy of the fungicide combination:
In attempting to explain the effectiveness of Synermix in the control of gray mold in the vineyard, the hypothesis of a stimulation by this fungicide of the natural defenses of grapevine (phytoalexin system) has been put forward. On the basis of previous works carried out with metallic salts (salts of copper and mercury), the inducing effect of aluminum chloride, one of the components of Synermix, was evaluated on the synthesis of phytoalexins of this plant. A study published in 1996 clearly demonstrated a hyper-elicitation of resveratrol production, resulting in quantities of the order of a few hundred micrograms per gram of fresh weight, on grapevine shoots (V. rupestris) dipped in tubes containing solutions of AlCl3.6H2O ranging from 7 to 90 mM within 15 h of incubation [51]. In contrast, no eliciting activity was observed on resveratrol synthesis using the seaweed extract. Synermix was marketed as an adjuvant for fungicidal products and not as a fungicide but was withdrawn from the market probably due to the possible toxicity of the aluminum salt present in the formulation.
Wang et al. [66] recently elucidated the mechanisms by which the aluminum cation Al3+ triggered phytoalexin production in grapevine cell cultures. Aluminum initiated intracellular actin remodeling (Al-actin bundling) through the activation of a NADPH oxidase located at the plasma membrane level, the membrane-associated NADPH oxidase Reactive burst Oxidase Homologue, RbOH. An induction of two subfamilies of genes encoding resveratrol synthase (STS47 and STS27) as well as the gene coding for the transcription factor MyB14 involved in the transcription of the stilbene synthase promoter, was observed through aluminum action, a phenomenon which was RbOH-actin-dependent. This resulted in resveratrol synthesis. Interestingly, rigorously similar observations have been reported in planta in two cultivars of V. vinifera and V. sylvestris demonstrating that these mechanisms are validated from the cell to the whole plant. The fact that RbOH mediates the remodeling of actin in response to aluminum involving the activation of several genes coding for the synthesis of stilbene phytoalexins in grapevine, would make this enzyme a privileged target to prime host defenses upstream of the infection process. The authors therefore suggested looking for ways other than aluminum, whose toxicity is proven, to stimulate the RbOH protein as alternative approaches for plant protection [66].
With copper sulfate (CuSO4) applied at a concentration of 50 μg/mL to in vitro grapevine plantlets or potted plants (cv Chardonnay), a protective effect of more than 60% in the extension of B. cinerea necrosis and only 38% in the case of downy mildew development, was respectively observed [50]. This protection was associated with an induction of chitinase and β-1,3-glucanase activities of the plant as well as a significant rise in stilbene synthesis in the leaves (resveratrol: 20 μg/g FW; ε-viniferin: 45 μg/g FW and piceid: 25 μg/g FW). These experiments showed that pre-treatment with non-toxic concentrations of CuSO4 led to increased protection towards grey mold and, to a lesser extent, against downy mildew, correlated with a stimulation of the plant defense mechanisms. This effect can be potentiated by a co-treatment with chitosan (see Section 3.3).
Application of fosetyl-Al for the control of the main pathogens responsible for esca in grapevine showed a protective activity of this treatment against the incidence of the disease foliar symptoms of esca disease in an experiment carried out in the vineyard from 1999 to 2007 on cvs Albana and Sangiovese [56]. However, the protective effect of fosetyl-Al appeared to be decorrelated from the production of stilbenes (resveratrol and ε-viniferin), the application of fosetyl-Al alone not inducing their synthesis.
3.2. Phytohormone derivatives with a stimulating effect on grapevine natural defenses
A certain number of phytohormones along with their derivatives (salicylic acid, SA, jasmonic acid, JA, and its methylated derivative, methyljasmonate, MeJA, ethylene, etc…) display an inducing effect on plant systemic acquired resistance (SAR) including active responses such as accumulation of pathogenesis-related proteins, phytoalexins or plant cell wall strengthening. It is beyond the scope of this review to deal with all the trials that have been carried out in grapevine. Some of these molecules (benzothiadiazole, metyljasmonate alone or in combination) have been used in particular with the aim of modifying the metabolism of polyphenols in grapes, increasing the biosynthesis of stilbenes through induction of the genes encoding enzymes of the phenypropanoid/stilbene pathway [67,68]. Only works linking protection against pathogens and stimulation of the phytoalexin system will be discussed here (also excluding those carried out on cell suspensions).
Benzothiadiazole (S-methyl benzo [1–3]thiadiazole-7-carbothioate, Bion, Syngenta) (BTH), is a salicylic acid (SA) mimic which provides plants with protection against pathogens without possessing a direct antifungal activity, but which acts on the induction of systemic acquired resistance (SAR) by SA-dependent pathways [69,70]. In the study conducted by Iriti et al. [71], BTH (0.3 mM) was applied in three different treatments within one week to detached grape bunches (V. vinifera cv Merlot) at veraison time (Table 2).
Subsequent infections of the bunches by B. cinerea carried out at the fourth week after treatment with BTH clearly showed a protective effect of the treatment regarding contamination of the grape clusters by gray mold since 87% of them only exhibited between 0 and 10% infection while 92% of the untreated ones showed a contamination rate of between 50 and 100%. The authors then attempted to explain the observed protective effect of BTH by an increase, particularly, in the synthesis of resveratrol in the berries. A 40% increase in the resveratrol content of the berries treated with BTH compared to the control (0.546 mg/kg FW versus 0.390 mg/kg FW) was effectively observed, but these very low levels in absolute values were in this case not high enough to support the hypothesis of a stimulation of the phytoalexin system by BTH [71]. This study therefore remained inconclusive as to a possible inducing effect of BTH on phytoalexin synthesis. Another work has reported the protective effect of BTH used at a dose of 0.05% on the grapevine V. vinifera cv Grüner Veltiner, a variety susceptible to downy mildew (P. viticola), against the infection by this pathogen, resulting in a reduction in the incidence of the disease to only 15.8% in greenhouses [72]. The priming effect of BTH was linked to the activation of various genes coding for PR proteins, peroxidase, but also PAL (phenylpropanoid pathway) and STS (resveratrol synthesis) (Table 2).
Application of BTH (1.9 mM) to the leaves of grapevine plants of V. vinifera cv Cabernet Sauvignon was carried out prior to inoculation with different isolates of downy mildew (P. viticola) and powdery mildew (Erysiphe necator) [73]. BTH exhibited a growth inhibition for all the tested isolates ranging from 60 to 98% (P. viticola) and from 65 to 75% (E. necator), thus conferring grapevine a very interesting level of protection. Besides an overexpression of a number of genes coding for PR proteins, lipoxygenase (LOX-9) and glutathione-S-transferase (GST) following pretreatment with BTH and at different times following inoculation by the two isolates of each of the two pathogens, an overexpression of the STS gene was observed as well as a significant accumulation of piceid (up to 60 μg/g dry weight (DW)) whereas the levels of resveratrol remained very low (2 to 8 μg/g DW). Quite surprisingly and taking account of the fact that pterostilbene is generally only a minor component in quantitative terms of the phytoalexin response of grapevine [74,75], the latter accumulated significantly in the leaves. under BTH treatment (whether contaminated or not) reaching, for example, 16 to 21 μg/g DW in plants treated with BTH and inoculated with E. necator. The high fungistatic and fungitoxic activities of this compound compared to its non-methylated derivative, resveratrol, partly explained the protective effect exerted by BTH against subsequent contaminations by P. viticola and E. necator [73]. These experiments thus clearly confirmed that the protection provided by BTH treatment involved an activation of SAR (Table 2).
Very interesting experiments were conducted in the vineyard for two consecutive years (2014 and 2015) comparing two protection treatments, one with a classic fungicide, pyrimethanil, and the other with the chemical elicitor BTH on grapevine cv Sémillon, which is very susceptible to gray mold (B. cinerea) [76]. In vitro activity tests have shown that pyrimethanil had an IC50, i.e., 51-times lower than BTH (67.5 mg/L vs. 3450 mg/L). In the vineyard, the differences observed regarding the protective effects of these two compounds were considerably reduced: 35% reduction of disease incidence for pyrimethanil and 20% for BTH in 2014; 29% and 25% respectively in 2015, demonstrating the relative efficacy of the two treatments. The correlation between the reported protection activities with an overexpression of the grapevine defense genes was even trickier. If the genes coding for several PR proteins were up-regulated as well as those encoding flavonoid biosynthesis, VvCHS and VvF3H resulting in a 42% increase in the total polyphenol content 48 h after the treatments’ application, an up-regulation of the genes encoding stilbene biosynthetic pathways, VvSTS and VvROMT, was delayed seven days after the treatment. Overexpression of these two genes was not accompanied by a significant production of stilbenes (piceid and pterostilbene) making it difficult to correlate the protective activity of BTH in the vineyard and the activation of phytoalexin biosynthesis [76].
The eliciting effect of methyljasmonate, a derivative of jasmonic acid, a phytohormone implicated in the regulation of plant defense responses as well as possessing regulatory functions in plant growth and development, has been the subject of numerous studies on grapevine cell cultures which have been reviewed in other articles [5,77]. We will limit ourselves here to works carried out on grapevine plants. V. vinifera cv Cabernet Sauvignon grapevine plants placed in a confined atmosphere and subjected to methyljasmonate (MeJa) vapors at a concentration of 400 nmol/L, 15 and 30 days after veraison, saw the piceid concentration increasing in the leaves for the two phenological stages (1200 and 990 nmol/g DW) but without any effect on resveratrol concentration which remained at very low levels (21 and 74 nmol/DW) [78]. In the berries, stilbenes were almost undetectable or in very low amounts whatever the phenological stage. Interestingly, MeJa showed a potentiating effect on the production of resveratrol in the leaves in response to UV stress (1632 and 2658 nmol/g DW) but without any significant effect on the resveratrol berry content. Treatments with airborne MeJa could have an interest in the vineyard for the protection of the leaves against the attacks of pathogenic agents at the last stages of ripening but would not produce notable effects on that of the berries. This type of treatment would find, on the other hand, an application for the protection of flowers and young berries whose synthesis of stilbene phytoalexins is very active [79]. Vezzuli et al. [80] reported similar experiments by spraying MeJa at different phenological stages in potted grapevine of the Barbera variety (fruit set, veraison and maturity or by combining these treatments over the entire ripening period). The activating effect of MeJa on stilbene production (resveratrol + ε-viniferin) was mainly observed in ripe berries, but the levels of phytoalexins attained were not high enough (< μg/g DW) to inhibit further pathogens’ development (Table 2).
Pre-treatment of foliar cuttings (cv Cabernet Sauvignon) or in the vineyard (cv Merlot) with MeJa used at doses of 5 mM (in a single application for potted plants or in several applications in the vineyard every 7–10 days from bloom to veraison) conferred a 75 and 73% reduction in the symptom incidence of powdery mildew (E. nector), respectively, while treatment with fosetyl-Al provided with an 83% protection [81]. However, no protective effect against the development of downy mildew (14%) was reported. Rise in the resistance to powdery mildew was correlated with a preinfectional increase in the transcript levels of the class 4 acid chitinase (25 times) as well as PAL and STS (44 and 11 times, respectively). Activation of the PAL and STS genes was accompanied by an increase in the stilbene content of the leaves (244 nmol/g DW for piceid; 176 nmol/g DW for resveratrol; 10 nmol/g DW for pterostilbene; 80 and 8 nmol/g DW for the dimers ε- and δ-viniferin, respectively). These data are very interesting insofar as they showed a correlation between pre-treatment with MeJa, stimulation of grapevine defense mechanisms including phytoalexin response, and protection against powdery mildew in the vineyard [81].
Three different elicitors [82] were tested as regard to the protection of potted grapevines (cv Cabernet Sauvignon) against contamination by downy mildew (P. viticola), MeJa, BTH and phosphonates (PHOS), a constituent of fosetyl-Al [83], registered as a fungicide for the treatment of downy mildew in grapevine. Pretreatment with the three elicitors at the respective doses of 2 g/L, 1.5 g/L and 1.09 g/L (5 mM) applied six days before inoculation provided a protective effect of 98.5, 97 and 85.8%, respectively towards contamination by downy mildew. At the gene level, BTH induced the overexpression of many PR proteins unlike leaves treated with MeJa and PHOS. The three treatments caused an increase in the concentration of flavanols, which are antifungal compounds (MeJa>BTH>PHOS) as well as a significant induction of stilbene biosynthesis (piceid up to 8 μg/mg DW; ε-viniferin: 9 μg/mg DW) and, to a lesser extent, of resveratrol and pterostilbene (< 1 μg/mg DW) for MeJa. This strong stilbene accumulation appeared, however, to be decorrelated from the expression of the VvPAL and VvSTS genes (Table 2).
Ethylene is a gaseous phytohormone that plays a crucial role in plant development and growth, plant stress responses, flowering, maturation as well as senescence and gemination [84]. Ethephon (2-chloroethylphosphonic acid) is an ethylene-releasing releasing compound. Used at a dose of 0.5 g/L (Sierra, Bayer, Cropscience), it showed a protective effect against E. necator both on leaf discs and grapevine cuttings of Cabernet Sauvignon with disease control rates ranging respectively from 64 to 70% [85]. Alongside providing protection against powdery mildew, there was a significant induction in the expression levels of PR proteins as well as those of PAL and STS genes by a factor of 10 and 67, respectively associated with a correlative increase in the synthesis of piceid (312 nmoles/g DW), resveratrol (133 nmoles/g DW), ε-viniferin (58 nmoles/g DW) as well as δ-viniferin and pterostilbene (12 and 10 nmoles/g DW). While MeJa or ethephon applied individually to foliar cuttings (cv Cabernet Sauvignon) at the respective doses of 5 nM and 6.94 nM reduced powdery mildew (E. necator) colonization by 60%, fosetyl-Al showing an 85% effectiveness under the same conditions, the MeJa/ethephon combination only reduced the incidence of the disease by 30% [86]. The effect of the combination of the two chemicals on the stimulation of defense genes (GLU, β-1,3-glucanase, PAL1 and STS) and the production of stilbenes was difficult to interpret since gene expression analysis was carried out on cell suspensions, a model very different from that used for the protection tests. The MeJa/ethephon co-treatment, although not conferring a similar protection level compared to each of the treatments, led to an overexpression of the PAL1, STS and GLU genes (respectively 120, 125 and 4500 times) as well as an increase in the contents of resveratrol (10.8 nmol/mg DW) and piceid (18.3 nmol/mg DW). The antagonistic effect observed during the application of the two products in combination at the plant protection level, could be due to their different actions on signaling pathways and probably linked to suppression of the MeJa-induced expression of some PR proteins and LOX genes, by ethephon [86].
Table 2.
Induction of grapevine disease resistance linked with stilbene phytoalexin synthesis by phytohormone derivatives.
Table 2.
Induction of grapevine disease resistance linked with stilbene phytoalexin synthesis by phytohormone derivatives.
| Compounds | Plant Material | Biological Inputs | References |
|---|---|---|---|
| Benzathiadiazole | Detached grape bunches (cv Merlot) | Significant decrease in B. cinerea incidence of the grape clusters (87% of them exhibited between 0 and 10% infection). Low stilbene accumulation | [71] |
| Benzathiadiazole | Potted grapevine (cv Grïuner Veltiner) | Reduction of downy mildew disease incidence to 15.8%. Strong activation of PAL and STS genes as well as peroxidases and PR-proteins | [72] |
| Benzathiadiazole | Leaves of grapevine plants (cv Cabernet Sauvignon) | Growth inhibition of P. viticola (60–98% and E. necator (65–75%). Overexpression of the STS gene with high piceid accumulation and significant accumulation of pterostilbene. Overexpression of several genes coding for PR proteins, LOX and GST | [73] |
| Benzathiadiazole vs. Pyrimethanil | Two-year period experiments in the vineyard (cv Semillon) | Decreases of grey mold incidence of respectively 35 and 20% for pyrimethanil and benzathiadiazole in 2014; −29 and −25% in 2015. Upregulation of VvSTS and VvROMT genes with no correlated accumulation of stilbenes | [76] |
| Methyljasmonate | Grapevine plants placed in a confined atmosphere with methyljasmonate (cv Cabernet Sauvignon) | High increase in leaf piceid content and little or no effect on the resveratrol content of grape berries. No test of disease pathogen control |
[78] |
| Methyljasmonate | Potted grapevine of the Barbera variety | Low increase in stilbene phytoalexin potential | [80] |
| Methyljasmonate | Plant cuttings (one application); multiple applications in the vineyard (from floraison to veraison) (cv Cabernet Sauvignon) |
Decreases of powdery mildew incidence of 75 and 73% in plant cuttings or in the vineyard, but not for downy mildew. Up-regulation of genes coding for PR-proteins as well as PAL and STS along with increase in stilbene accumulation | [81] |
| Methyljasmonate (MeJa), Benzathiadiazole (BTH), Phosphonates (PHOS) | Plant cuttings (cv Cabernet Sauvignon) |
Respectively 98.5, 97.3 and 85.8% protection towards downy mildew infection with MeJa, BTH and PHOS. High induction of stilbene production decorrelated from VvPAL and VvSTS expression. High overexpression of PR proteins with BTH, lower with MeJa and PHOS | [82] |
| Ethephon (ethylene) | Plant cuttings and leaf discs (cv Cabernet Sauvignon) | Increases in protection (64–70%) against powdery mildew. Up-regulation of defense genes (PAL and STS) along with strong stilbene accumulation (piceid, resveratrol and dimers) | [85] |
| Methyljasmonate/Ethephon | Plant cuttings and cell suspensions (cv Cabernet Sauvignon) | Decreases of 60% of the colonization by downy mildew by methyljasmonate or ethephon alone (only 30% when applied in combination; 85% with fosetyl-Al). Increases in PAL1 and STS along with increase in piceid and resveratrol production in cell suspensions. Inhibition of some PR proteins and LOX gene-induced expression by ethephon | [86] |
3.3. Bio-Elicitors
In this section will be treated what we have chosen to arbitrarily qualify as bio-elicitors such as, among others, laminarin, chitosan, fungal sterols, bacterial rhamnolipids, β-amino-butyric acid and tagatose, which are able to induce stilbene synthesis [87,88] (Table 3).
Among polysaccharides, laminarin, a β-1,3-glucan extracted from a brown alga, Laminaria digitata, has shown protective effects against B. cinerea and P. viticola by reducing the symptoms of these two diseases by respectively 55% and 75% on leaves of grapevine in vitro plantlets of the Chardonnay and Gamay varieties, both susceptible to gray mold and downy mildew [89]. Although carried out on a different plant model, parallel experiments realized with cell suspensions, showed an induction of responsive defense genes (preceded by an oxidative burst), particularly the genes coding for PR proteins as well as a strong overexpression (20 times higher than that of the controls) of the PAL and STS1 genes with a concomitant accumulation of the phytoalexins resveratrol and ε-viniferin (Table 3).
Chitosan is also a polysaccharide composed of D-glucosamine and of N-acetyl-D-glucosamine units partially deacetylated and obtained from the chitin of crabs, shrimps and other crustaceans, known for its bio-stimulant and elicitor properties [90]. The protective properties of chitosan alone or in combination with copper sulfate have been studied towards gray mold and downy mildew as well as its action on the stimulation of grapevine defense mechanisms [50,91]. Pretreatment of grapevine plantlets (cv Chardonnay) with chitosan at a dose of 150 mg/L resulted in the total suppression of B. cinerea lesions on leaves, demonstrating the protection effect of chitosan regarding gray mold [91]. Interestingly, this protective effect was observed after removal of the elicitor, by washing the leaves, and before inoculation by B. cinerea, seeming to indicate that the action of this elicitor was not only linked to its direct antifungal activity but to the activation of some plant defense mechanisms (overexpression of chitinase, PAL and LOX activities), even if parallel works seemed to question the direct antifungal activity of chitosan [50]. Similarly, pretreatment with chitosan with a molecular mass of 1500 Da and partially deacetylated (20%) at a dose of 200 μg/mL induced 60 to 71% protection towards, respectively, the contamination of grapevine in vitro plantlets or potted plants by gray mold and downy mildew [50]. This protective effect was associated with an induction of plant chitinase and β-1,3-glucanase activities and an increase in stilbene contents (resveratrol: 18 μg/g FW; ε-viniferin: 50 μg/g FW and piceid: 40 μg/g FW). The protection rate increased to 85% in the case of downy mildew upon combination of chitosan (200 μg/mL) and copper sulfate (50 μg/mL) with potentiation of the phytoalexin response [50] (Table 3).
Ergosterol is a major sterol found in fungi that plays a role in the fluidity and permeability of cell membranes. Pretreatment with ergosterol administered at a concentration of 200 μM on grapevine plants (cv Ugni Blanc) conferred a protective effect of 75% against contamination by B. cinerea [92]. Conversely, only 55% of protection were achieved in case of pretreatment with BTH. The resistance provided by ergosterol was accompanied by an induction, mediated by the mobilization of the trans-acting elements of the transcription factor WRKY, of stilbene synthase gene (VST1) expression and incidentally by the overexpression of the VvLTP1 gene coding for the lipid transfer protein 1. Overexpression of the VST1 gene was correlated with low resveratrol synthesis in the plants treated with ergosterol (227 ng/g FW) (Table 3).
β-amino-butyric acid (BABA) is a non-protein amino acid known to induce resistance against oomycetes in plants [93,94]. Pretreatment with aqueous solutions of BABA (1 mM) prior to contamination by downy mildew (P. viticola) on two cultivars (plant cuttings), one susceptible (Chasselas), the other resistant (Solaris), induced a strong reduction in sporulation as well as the growth of P. viticola hyphae [95]. The priming effect exerted by BABA resulted in an increase in the overall amounts of stilbenes (resveratrol, piceid and ε-viniferin) alongside stilbenes displaying strongly fungicidal properties such as δ-viniferin and pterostilbene. Priming activity of BABA on the expression of PAL and C4H was mainly observed in the downy mildew-resistant variety, Solaris, whereas STS was up-regulated in both the BABA-primed cultivars (Table 3).
Works have reported that natural rhamnolipids can potentially be used for the protection of plants against bacterial and fungal diseases, protecting for example, oilseed rape and tomato from B. cinerea attacks [96,97]. In grapevine (V. vinifera), pretreatment with a mixture of rhamnolipids (RLs) extracted from the bacterium Pseudomonas aeruginosa at concentrations of 0.1 and 1 mg/L showed a protective effect on in vitro whole plantlets against contamination by B. cinerea [98]. RL activity resulted in a strong increase in the transcript accumulation of genes encoding various PR proteins, LOX as well as PAL and STS genes. Because of their lipo-solubility, RLs exerted a direct antifungal action and were capable to potentiate the action of other bio-elicitors such as chitosan.
The role of sugars in the interactions between plants and microbes and in the resistance of the former to numerous pathogens in connection with the concept of sweet immunity is well established [99,100]. In this context, rare sugars have been used to stimulate the defenses of plants and thus increase their resistance to phytopathogenic agents [101]. Among these, D-tagatose has shown protective activity especially against members of the class of Oomycetes [102,103]. Root or aerial pretreatments of grapevine in vitro plantlets (cv Chardonnay) by IFP48, an 80% D-tagatose-based product currently under registration on the European market and used at a dose of 5 g/L, showed an anti-oomycete activity, reducing sporangia density of P. viticola by 35% [104]. This protective effect was correlated with an overexpression of genes coding for PR proteins, LOX9 as well as PAL and STS genes (with a respective induction of 10 and 8 times). Overexpression of these responsive defense genes in grapevine explained, at least in part, the protection conferred by IFP48 as the level of overexpression of those genes remained high after inoculation with downy mildew. It should also be noted that IFP48 exhibited a direct anti-oomycete activity, reducing by 100% sporangia density of P. viticola. A similar experiment carried out by the same group confirmed the protective role of IFP48 with a 50% reduction of P. viticola sporangia density but showed no effect, on the other hand, of this product on the control of gray mold. (B. cinerea) [105]. The protection mechanisms of IFP48 resulted in modulation of the expression of some responsive defense genes controlled by the salicylic acid pathway (PR1 and PR2 genes) and that of jasmonic acid (ERF1, Ethylene Response Factor 1 and PR3c genes) in the absence of pathogen contamination. In both studies, only a low accumulation of stilbenes was reported (3–5 μg/g FW for resveratrol and a few micrograms per grams of FW for viniferins ε and δ).
Table 3.
Use of bio-elicitors for grapevine protection and stimulation of the phytoalexin response.
| Elicitor | Plant Material | Biological Inputs | References | |
|---|---|---|---|---|
| Laminarin | Grapevine in vitro plantlets (cv Chardonnay and Gamay) | 55 to 75% protection against respectively gray mold and downy mildew. Overexpression of genes coding for PR proteins as well as PAL and STS1 genes with concomitant accumulation of phytoalexins (cell suspensions) | [89] | |
| Chitosan partially deacetylated | Grapevine in vitro plantlets (cv Chardonnay) | Total suppression of B. cinerea necrosis on leaves. Overexpression of chitinases, PAL and LOX activities. Direct antifungal activity. No mention of stilbene production | [91] | |
| Chitosan partially deacetylated (20%) | Grapevine in vitro plantlets and potted plants (cv Chardonnay) | 60 to 70% protection against respectively gray mold and downy mildew linked with an increase in chitinase, β-1,3-glucanase activities and augmentation of stilbene content (resveratrol, piceid and ε-viniferin) | [50] | |
| Chitosan + copper sulfate | Potted plants (cv Chardonnay) | 83% protection against downy mildew associated with a potentiation of the phytoalexin response | [50] | |
| Ergosterol | Grapevine plants(cv Ugni Blanc) | 75% protection towards B. cinerea contamination (55% with benzothiadiazole). Overexpression of the WRKY transcription factor, VvST1 and VvLTP1 with correlated resveratrol production | [92] | |
| β-amino-butyric acid | Plant cuttings of cvs Chasselas andSolaris | Strong inhibition of P. viticola sporulation and hyphal growth linked with a priming effect on stilbene accumulation correlated with overexpression of PAL and C4H genes in Solaris and STS in both cultivars | [95] | |
| Bacterial rhamnolipidss | Grapevine in vitro plantlets |
Protection of leaves against B. cinerea infection. Overexpression of PR proteins, LOX, PAL and STS genes | [98] | |
| D-Tagatose (IFP48) | Grapevine in vitro plantlets (cv Chardonnay) | Reduction by 35% of sporangia density of P. viticola associated with a direct anti-oomycete activity. Overexpression of genes encoding PR proteins, LOX9, PAL and STS along with low stilbene production (resveratrol, piceid and dimers) | [104] | |
| D-Tagatose (IFP48) | Grapevine in vitro plantlets (cv Chardonnay) | Reduction by 50% of sporangia density of P. viticola but no effect towards B. cinerea. Overexpression of PR1 and PR2 genes (SA pathway), ERF1 and PR3c genes (JA pathway). Low stilbene production (resveratrol, piceid and dimers) | [105]) | |
1 SA, salicylic acid; JA, jasmonic acid.
4. Control of grapevine diseases by beneficial organisms involving stimulation of phytoalexin synthesis
The biological control of plant diseases can be defined as the partial or total, direct or indirect, inhibition of the growth and development of a pathogenic agent responsible for a given disease by another living organism (or group of organisms), often antagonistic and considered as a beneficial organism. The living organisms (or group of organisms) responsible for controlling a disease are then called biological control agents (BCAs) [106,107,108,109,110,111]. The fungus, Trichoderma harzanium strain T39 has been reported, for example, as a biocontrol agent for downy mildew in grapevine showing an 86% reduction in disease symptoms on potted grapevine, cv Pinot Noir. Among the differentially-induced defense responsive genes analyzed, PAL and STS genes were found to be up-regulated without mention regarding stilbene phytoalexin synthesis [112]. There are works describing induction of phytoalexin biosynthesis by BCAs, for example, to name just a few, the accumulation of scoparone and scopoletin in the control of Penicillium digitatum, P. italicum and B. cinerea by the yeast Rhodotorula glutinis in orange fruit [113], the synthesis of dianthranilide-type phytoalexins during the biocontrol of fusarium wilt by a Pseudomonas strain in carnation [114] or camalexin priming in the induced resistance of Arabidopsis by beneficial bacteria in the control of B. cinerea and P. syringae [115]. To our knowledge, the first study reporting the biocontrol of a grapevine disease, gray mold, by a soil bacterium causing an elicitation of the biosynthesis of the phytoalexin resveratrol dates back 1998 [116]. Co-inoculation of in vitro grapevine plantlets of V. vinifera (susceptible) and V. rupestris (mid-tolerant) with B. cinerea and the uncharacterized soil bacterium B-781 led to a complete suppression of the disease symptoms. The biocontrol of B. cinerea could have been linked to an increase in the production of resveratrol in V. vinifera (6 μg/g FW) and in greater amounts in V. rupestris (13 μg/g FW). A potentiating effect of the fungus/bacterium co-inoculation was observed on resveratrol production (78 μg/g FW) in V. vinifera and to a lesser extent in V. rupestris (31 μg/g FW) [115] (Table 4).
Two rhizobacteria of the genus Pseudomonas, Pseudomonas fluorescens CHA0 and Pseudomonas aeruginosa 7NSK2 induced resistance in leaves of in vitro grapevine plantlets of the Chardonnay variety (susceptible) to B. cinerea [117], the protection conferred towards grey mold infection being greater than 20% with P. fluorescens CHA0 and about 35% with and P. aeruginosa 7NSK2 (Table 4). The authors ruled out the hypothesis of a direct inhibitory activity of the bacteria towards B. cinerea. In both cases, induction of the systemic resistance to B. cinerea could be correlated with a priming effect on stilbene phytoalexin accumulation in the leaves, this effect being initially relatively weak at the pre-contamination stage (before B. cinerea inoculation), but significant compared to the control (10–25 μg/g FW for resveratrol and 15 to 30 μg/g FW for ε-viniferin). At 3 days post-inoculation with B. cinerea, these concentrations reached very high values (800 μg/g FW for resveratrol and 250 μg/g FW for ε-viniferin with P. fluorescens CHA0 and > 700 μg/g FW for resveratrol and 180 μg/g FW for ε-viniferin with P. aeruginosa 7NSK2). Phytoalexin production was preceded by an early oxidative burst. Very interestingly, two mutants of the 7NSK2 strain, one deficient in pyochelin (Pch) and pyoverdin (Pvd), two bacterial compounds inducers of resistance, the mutant KMPCH, and the other one deficient in Pch, Pvd and SA, the mutant KMPCH-567, showed differential induction profiles in the resistance to B. cinerea. The KMPCH mutant (Pch- and Pvd-negative) showed a protective effect of 35% against B. cinerea, that is, an effect comparable to that of the parental strain 7NSK2, the KMPCH-567 mutant (Pch-, Pvd- and SA negative) inducing little or no protection against gray mold infection (< 10%). The protective activity linked to the KMPCH mutant was found to be correlated with a potentiation of the production of resveratrol in the leaves, the KMPCH-567 mutant only inducing weak resveratrol amounts at two days post-inoculation. These data therefore reinforced the role played by phytoalexins in the bacteria-induced disease resistance to B. cinerea [117].
Several bacteria isolated in the Champagne vineyard, Bacillus subtilis, Pantoea agglomerens, Acinetobacter lwoffii and Pseudomonas fluorescens induced a protective effect against B. cinerea on in vitro grapevine plantlets (cv Chardonnay), B. subtilis showing a 35% reduction in gray mold symptoms, with P. fluorescens showing the highest protection rate (70%), P. agglomerens and A. lwoffii resulting in a 60% reduction in leaf symptoms [118]. All the bacteria triggered an early oxidative burst preceding an induction of the biosynthesis of the phytoalexins resveratrol and ε-viniferin of the order of 10 to 20 μg/g FW. A priming effect on phytoalexin accumulation was observed with P. fluorescens and A. lwoffii but not in the case of P. agglomerens and B. subtilis, where pretreatment with these bacteria had no effect on subsequent B. cinerea-induced phytoalexin production. Later studies focused on the mechanisms of action of the bacterium P. fluorescens PTA-CT2 on grapevine [119,120]. Induction of the systemic resistance in grapevine against B. cinerea resulted in a 60% reduction in the symptoms of this fungus on in vitro grapevine plantlets cv Chardonnay, through the differential expression of defensive response genes at the root level (organ colonized by the bacterium) and at the leaf level (lack of colonization). Genes of the phenylpropanoid pathway and resveratrol synthesis (PAL and STS) as well as those of the anthocyanin biosynthetic route (CHS, chalcone synthase, CHI, chalcone isomerase, ANS, anthocyanidin synthase) displayed a much higher overexpression level in the leaves compared to the roots [119]. The PTA-CT2 bacterium was able to prime phytoalexin synthesis (resveratrol, piceid and ε-viniferin) in response to B. cinerea inoculation, the amounts of stilbenes accumulated in the leaves in response to P. fluorescens (3 days after the beginning of the infection), being higher in the leaves (piceid: 20 μg/g FW; resveratrol: 4 μg/g FW; ε-viniferin 10 μg/g FW) than in the roots ((piceid: ~ 0 μg/g FW; resveratrol: 1 μg/g FW ε-viniferin 5μg/g FW). Stilbene accumulation was fully consistent with the up-regulation levels of the PAL and STS genes, high in the leaves and low in the roots. Other defense genes, particularly those encoding PR proteins, showed a differential response depending on the organs analyzed, leaves or roots (Table 4).
Although the effectiveness of the PTA-CT2 strain of the bacterium P. fluorescens has already been demonstrated for the biocontrol of gray mold (B. cinerea) [117,118,119,121,122,123], that of this bacterium against the oomycete P. viticola (downy mildew), on the other hand, had not been the subject of any study. In the work of Lakkis et al. [120], the effectiveness of the PTA-CT2 strain was evaluated on two-year-old potted grapevines of the varieties Pinot Noir (susceptible) and Solaris (tolerant) by soil drenching with the bacterial solution. Application of the bacterium led to a reduction in the growth development of P. viticola of 80% in Pinot Noir and only 55% in Solaris; this reduction was 73–80% for Pinot Noir and 43% for Solaris following contamination by B. cinerea. In the absence of any contamination, PTA-CT2 did not induce changes in the basal defenses of the plant, but induced changes at the hormonal level and an improvement in the photosynthetic capacities for the two varieties. In contrast, PTA-CT2 primed defensive pathways including PAL and STS gene overexpression, which was correlated with increased phytoalexin levels in both varieties. The two varieties showed quite similar profiles of phytoalexin accumulation after PTA-CT2/P. viticola or PTA-CT2/B. cinerea co-inoculation, respectively, resveratrol: 15 μg/g FW, ε-viniferin: 6–8 μg/g FW and δ-viniferin: 12–17 μg/g FW; resveratrol: 40 μg/g FW, ε-viniferin: 12–25 μg/g FW and δ-viniferin: 5–10 μg/g FW. These results clearly demonstrated that the effectiveness of the biocontrol exerted by the bacterium P. fluorescens was mediated by the upregulation of genes involved in the stilbene biosynthetic pathway resulting in priming of the phytoalexin response and reinforced the role played by these compounds in grapevine/plant pathogens interactions.
There are few experiments describing the use of beneficial bacteria as biocontrol agents of fungal diseases in the vineyard and linking a possible protective effect of these bacteria on the stimulation of grapevine defense systems [121,122,123]. Tests carried out in the vineyard for four consecutive years from 2002 to 2005 regarding the biological control of B. cinerea by grapevine-associated bacteria, reported interesting results towards the reduction on leaves and berries (reaching 50%) of the symptoms of B. cinerea by application of bacteria by the soil drenching method or by spray [121]. This resistance to B. cinerea was correlated with a stimulation of β-1,3-glucanase and chitinase activities in grape leaves and berries, but no analysis of phytoalexins was described at this time. Very similar work by the same group reported a 22 to 75% decrease in gray mold development in the vineyard following drenching of the grapevine soil with bacterial mixtures, this protection effect being linked to increases in β-1,3-glucanase and chitinase activities without any notification regarding phytoalexin production [122].
Table 4.
Control of grapevine diseases by beneficial organisms involving stimulation of phytoalexin synthesis.
Table 4.
Control of grapevine diseases by beneficial organisms involving stimulation of phytoalexin synthesis.
| Biocontrol Agent | Plant Material | Biological Inputs | References |
|---|---|---|---|
| Uncharacterized soil bacterium B-781 | In vitro grapevine plantlets of V. vinifera (susceptible) and V. rupestris (mid-tolerant) | Complete suppression of gray mold symptoms with increase in resveratrol accumulation | [116] |
| Pseudomonas fluorescens CHA0 and Pseudomonas aeruginosa 7NSK2 | In vitro grapevine plantlets cv Chardonnay | Protection towards gray mold > 20% with P. fluorescens CHA0 and about 35% with and P. aeruginosa 7NSK2. At 3 days post-inoculation with B. cinerea, stilbene concentrations reached very high values in the order of several hundred μg/g FW | [117] |
| Bacillus sutilis, Pantoea agglomerens, Acinetobacter lwoffii and Pseudomonas fluorescens | In vitro grapevine plantlets cv Chardonnay | 35% reduction in gray mold symptoms with B. subtilis, 70% with P. fluorescens and 60% with P. agglomerens and A. lwoffii. Priming effect on phytoalexin accumulation with P. fluorescens and A. lwoffii but not with P. agglomerens and B. subtilis | [118] |
| Trichoderma harzanium strain T39 | Potted grapevine, cv Pinot Noir | 86% reduction in disease symptoms towards downy mildew; induction of PAL and STS genes, no mention of stilbene production | [112] |
| P. fluorescens PTA-CT2 | In vitro grapevine plantlets cv Chardonnay | 60% reduction in gray mold symptoms. Differential expression of PAL and STS genes (higher in leaves than in roots) correlating with accumulation of stilbenes in the two organs | [119] |
| P. fluorescens PTA-CT2 | Two-year-old potted grapevins of the varieties Pinot Noir (susceptible) and Solaris (tolerant) | Reduction in the growth development of P. viticola of 80% in Pinot Noir and only 55% in Solaris, 73–80% for Pinot Noir and 43% for Solaris towards B. cinerea. No induced changes in the basal defenses of the plant with PTA-CT2 alone. PTA-CT2 primed defensive pathways including PAL and STS gene overexpression, which was correlated with increased phytoalexin levels in both varieties | [120] |
| B. subtilis (PTA-271), P. fluorescens (PTA-CT2) and P. agglomerens (PTA-AF2) alone or as binary mixtures | One-year experiment on grapevine plants (cv Chardonnay) in the vineyard including leaves and berries | 80 to 90% reduction in symptoms towards B. cinerea) with PTA-AF2 + PTA-271 on leaves. 93% reduction of B. cinerea symptoms with CT2 + AF2 on berries well correlating with phytoalexin accumulation | [123] |
To our knowledge, the only experiment describing use of endophytic and rhizospheric bacteria for the biocontrol of gray mold in vineyard conditions and linking this protective effect to the induced production of phytoalexins both in the leaves and the berries, was that conducted by Aziz et al. [123] (Table 4). Three living bacteria isolated from grapevines in the Champagne vineyards, one rhizospheric, B. subtilis (PTA-271), the other two being endophytic bacteria obtained from tissues of healthy grapevine plants, P. fluorescens (PTA-CT2) and P. agglomerens (PTA-AF2), previously studied on grapevine in vitro plantlets [118], were brought to grapevine plants (cv Chardonnay) in the vineyard by drenching the soil, either individually or in the form of binary mixtures, in two treatments spaced one month apart in June and July. On grape leaves, it was interesting to note that it was the combination PTA-AF2 + PTA-271, which triggered the highest level of systemic resistance (80 to 90% reduction in symptoms towards B. cinerea), 75 days after the first bacterial application and which also displayed the highest level of total phytoalexins (35 to 40 μg/g FW) as well as the highest values for resveratrol (25 μg/g FW) and ε-viniferin (7 μg/g FW) confirming the good correlation between protection towards contamination by B. cinerea and rate of accumulation of antifungal phytoalexins. The results were more difficult to interpret regarding bacteria alone, since PTA-271 which provided the best protection level on leaves (75% reduction in B. cinerea symptoms) was not the one which induced the highest phytoalexin response compared to the PTA-CT2 + PTA-AF2 mixture or PTA-CT2 alone.
B. cinerea is a particularly redoutable parasite that damages the harvested grape quality because of its late attacks on grape berries and due to the fact that the amounts of inducible phytoalexins during stress at this stage of maturity are low [79,124]. Specific attention must therefore be paid to the level of protection of the grape clusters towards this fungus. The effectiveness of bacteria alone in terms of reduction of the B. cinerea symptoms on grape berries was very good, varying from 78% (PTA-CT2) to 87% (PTA-271), which correlated well with the accumulation of total phytoalexins (15 μg/g FW for PTA-271 and around 40 μg/g FW for PTA-CT2) along with a significant production of the antifungal dimer, ε-viniferin (>12 μg/g FW), 75 days after application of the first bacterial treatment. The CT2 + AF2 combination displayed the highest level of protection on grape berries (93% reduction of the B. cinerea symptoms) correlating with a significant accumulation of total phytoalexins (> 20 μg/g FW). It was interesting to note that, in almost all cases, accumulation levels of phytoalexins in grape berries remained high, even 91 and 99 days after application of the first bacterial treatment, i.e., at dates close to maturity, stages where they are most vulnerable to gray mold attacks [123].
5. Conclusions
Most of the treatments described in this work (chemicals, phytohormone-derivatives, bio-elicitors, biocontrol agents) led to variable protective effects against various pathogens (powdery mildew, downy mildew, gray mold, esca) which number among the phytopathogenic agents responsible for grapevine major fungal diseases, this protection being most often correlated with overexpression of genes of the phenylpropanoid pathway (PAL and C4H) and of the synthesis of resveratrol (STS) alongside an up-regulation of the expression of other responsive defense genes (LOX, GST, PER, ERF1 and genes encoding PR proteins). Induction of PAL and STS genes was generally accompanied by a notable increase in the content of stilbenes (resveratrol, piceid, pterostilbene and dimers) which may explain the protection observed towards pathogens. However, in some cases, the level of accumulation of these compounds regarding the doses required for them to exert their antifungal activity, remained insufficient to support the protective effects observed [71,80,104,105] and, sometimes, a decorrelation was described between PAL and STS genes’ up-regulation and stilbene production [76,82,104,105]. Even if some data exposed in this review were difficult to interpret, there were compelling facts to support the role of stilbene phytoalexin inducers in controlling various pathogens thereby providing grapevine with relative protection against diseases. Although phytoalexins represent only one component of the plant response to stress, it appeared that their production and the overexpression levels of the genes involved in their biosynthesis (PAL, C4H, STS), were often correlated with those of other responsive defense genes (PR proteins, LOX, GST, PER, etc...).
Metallic salts possess a significant activity on the stimulation of stilbene synthesis, particularly, aluminum chloride and copper sulfate associated with a certain level of protection of grapevine against gray mold [50,63,64] and, to a lesser extent, towards downy mildew [50]. The effect of copper sulfate is to be linked to the use since the beginning of the 19th century of the bouillie bordelaise in the vineyard. The aluminum contained in AlCl3 was a very effective inducer of phytoalexin synthesis leading to the accumulation of huge amounts of resveratrol in the leaves (> 500 μg/g fresh weight (FW)) [51]. The mechanisms by which this metal cation triggers the hyperproduction of this stilbene go through an overexpression of two STS gene sub-families and the transcription factor MYB14 controlling the STS gene promoter, have recently been deciphered [66]. Initiation of the overexpression of these genes, which depends on Al-induced remodeling of intracellular actin, involved the ROBh protein on which elicitor molecules other than aluminum salts whose toxicity is recognized, could be sought representing an interesting path for further technological development.
The different elicitation methods used were able to act either on the stimulation of the basal levels of phytoalexin biosynthesis (action of the treatment at the preinfectional stage), or by a priming effect (synergy of the effect linked to the pre-treatment combined with the phytoalexin response following contamination by the pathogen). The elicitor alone may also have no effect on the basal defenses of the plant, its action being exerted essentially by priming of the phytoalexin response. The level of accumulation of the latter in the plant treated with the elicitor and co-infected with the pathogen is then greater than that of the plant only infected with the pathogen.
Fosetyl-Al, for example, can significantly increase the basal phytoalexin response when applied alone or displays no effect on this response, its activity being mainly due to a priming effect [61,62]. Aluminum chloride used alone as well as copper sulfate, chitosan or a combination of both, were capable of inducing the synthesis of high amounts of resveratrol [50,51]. Pretreatment with numerous elicitors such as benzothiadiazole, phosphonates, methyljasmonate, ethephon, chitosan or ergosterol also triggered significant changes in the basal levels of stilbene phytoalexins [50,73,78,81,82,85,92]. Other experiments have shown that benzothiadiazole induced at the same time a rise in the basal syntheses of phytoalexins as well as a phytoalexin priming activity [73]. β-amino-butyric acid displayed a priming effect on the phytoalexin response but no action on the stimulation of phytoalexin synthesis when applied alone [95].
Biocontrol agents can stimulate phytoalexin synthesis during preinfectional treatments [50,116] or may be able to induce both a basal phytoalexin response and to prime this response [116,117]. Finally, inoculation with beneficial bacteria often showed low or no effect on basal phytoalexin levels but, above all, a priming effect [118,119,120].
The tests carried out in the vineyard faced the versatility of the experimental conditions (climate, disease pressure, way of applying the treatments and frequency of applications). Because of these constraints, few trials have been carried out in the vineyard [64,76,81,123]. Experiments conducted with Synermix (AlCl3 + seaweed extract) obtained good results in terms of grapevine protection towards Botrytis cinerea, application trials with Synermix achieving results close to those obtained with Iprodione, the Synermix + Iprodione combination showing a real efficacy, better than the theoretical one [63,64]. The protective effects observed with Synermix had then been correlated with the strong eliciting activity of AlCl3 on resveratrol synthesis [51]. Surprisingly, this product was withdrawn from the market, probably due to the presence of aluminum in the formulation. Experiments including applications of methyljasmonate in the vineyard every 7–10 days, from bloom to veraison conferred a 73% reduction in the symptom incidence of powdery mildew [81]. This protective effect was correlated with a large increase in preinfectional accumulation of various stilbene phytoalexins (resveratrol, piceid and dimers).
Input of endophytic or rhizophytic bacteria alone or as mixture to grapevine plants grown in the vineyard by drenching the soil in two treatments spaced one month apart in June and July, showed very promising results in terms of protection of this plant against contamination by gray mold with reduction rates of the disease symptoms reaching 90% in the leaves and 93% in grape berries [123]. This protection effect was accompanied by a significant increase in the basal levels of phytoalexins (resveratrol and ε-viniferin). Most importantly, phytoalexin amounts in the grape berries remained high, even 91 and 99 days after application of the first bacterial treatment, i.e., at stages close to fruit maturity when the fruits are particularly susceptible to B. cinerea attacks.
In sum, all the works presented and discussed in this review, showed that the protective effect observed towards phytopathogenic agents, by the application of different elicitors was correlated with an increase in the biosynthetic basal levels of stilbene phytoalexins as well as phytoalexin priming in grapevine under various conditions, making it possible to validate the concept of using phytoalexin induction as a means for crop protection.
Author Contributions
Conceptualization, P.J., P.T.-A. and A.A.; resources, C.J.; writing—original draft preparation, P.J.; review and editing, C.J., C.C., I.M. C.M., H.K.; project administration, C.J.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Müller, K.O.; Börger, H. Experimentelle Untersuchungen über die Phytophthora Resistenz der Kartoffel. Arbeit. Biol. Reichsant. Land Forstwirtsch. 1940, 23, 189–231. [Google Scholar]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Jeandet, P.; Hébrard, C.; Deville, M.A.; Cordelier, S.; Dorey, S.; Aziz, A.; Crouzet, J. Deciphering the role of phytoalexins in plant-microorganism interactions and human health. Molecules 2014, 19, 18033–18056. [Google Scholar] [CrossRef] [PubMed]
- Keylor, M.H.; Matsuura, B.S.; Stephenson, C.R.J. Chemistry and biology of resveratrol-derived natural products. Chem. Rev. 2015, 115, 8976–9027. [Google Scholar] [CrossRef]
- Jeandet, P.; Vannozzi, A.; Sobarzo-Sanchez, E.; Uddin, M.S.; Bru, R.; Martinez-Marquez, A.; Clément, C.; Cordelier, S.; Manayi, A.; Nabavi, S.F.; et al. Phytostilbenes as agrochemicals: biosynthesis, bioactivity, metabolic engineering and biotechnology. Nat. Prod. Rep. 2021, 28, 1282–1329. [Google Scholar] [CrossRef]
- Jeandet, P.; Uddin, M.S.; Clément, C.; Aziz, A.; Jacquard, C; Khan, H.; Shah, M.A.; Ait-Barka, E.; Koffas, M.; Nabavi, S.M.; et al. Production of high molecular-ordered stilbene oligomers: Total synthesis, bio-catalyzed synthesis and production by plant systems. Nat. Prod. Rep. 2023, 30, 1045–1057. [CrossRef]
- Delaunois, B.; Cordelier, S.; Conreux, A.; Clément, C.; Jeandet, P. Molecular engineering of resveratrol in plants. Plant Biotechnol. J. 2009, 7, 2–12. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clément, C.; Courot, E. Biosynthesis, metabolism, molecular engineering and biological functions of stilbene phytoalexins in plants. BioFactors 2010, 36, 331–341. [Google Scholar] [CrossRef]
- Jeandet, P.; Sobarzo-Sánchez, E.; Clément, C.; Nabavi, S.F.; Habtemariam, S.; Nabavi, S.M.; Cordelier, S. Engineering stilbene metabolic pathways in microbial cells. Biotechnol. Adv. 2018, 36, 2264–2283. [Google Scholar] [CrossRef]
- Cartwright, D.W.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Chemical activation of host defence mechanisms as a basis for crop protection. Nature 1977, 267, 511–513. [Google Scholar] [CrossRef]
- Langcake, P.; Pryce, R.J. The production of resveratrol by Vitis vinifera and other members of the Vitaceae as a response to infection or injury. Physiol. Plant Pathol. 1976, 9, 77–86. [Google Scholar] [CrossRef]
- Langcake, P.; Pryce, R.J. The production of resveratrol and the viniferins by grapevines in response to ultraviolet irradiation. Phytochemistry 1977, 16, 1193–1196. [Google Scholar] [CrossRef]
- Langcake, P. Disease resistance of Vitis spp. and the production of the stress metabolites resveratrol, ε-viniferin, α-viniferin and pterostilbene. Physiol. Plant Pathol. 1981, 18, 213–226. [Google Scholar] [CrossRef]
- Nabavi, S.; Samec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid biosynthetic pathways in plants: versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 39, 107461. [Google Scholar] [CrossRef] [PubMed]
- Rivière, C.; Pawlus, A.D.; Mérillon, J.-M. Natural stilbenoids: Distribution in the plant kingdom and chemotaxonomic interest in Vitaceae. Nat. Prod. Rep. 2012, 29, 1317–1333. [Google Scholar] [CrossRef]
- Parage, C.; Tavares, R.; Réty, S.; Baltenweck-Guyot, R.; Poutaraud, A.; Renault, L.; Heintz, D.; Lugan, R.; Marais, G.A.B.; Aubourg, S.; et al. Structural, functional, and evolutionary analysis of the unusually large stilbene synthase gene family in grapevine. Plant Physiol. 2012, 160, 1407–1419. [Google Scholar] [CrossRef]
- Vannozzi, A.; Dry, I.B.; Fasoli, M.; Zenoni, S.; Lucchin, M. Genome-wide analysis of the grapevine stilbene synthase multigenic family: genomic organization and expression profiles upon biotic and abiotic stresses. BMC Plant Biol. 2012, 12, 130. [Google Scholar] [CrossRef]
- Qian, Y.; Lynch, J.H.; Guo, L.; Rhodes, D.; Morgan, J.A.; Dudareva, N. Completion of the cytosolic post-chorismate phenylalanine biosynthetic pathway in plants. Nat. Comm. 2019, 10, 15. [Google Scholar] [CrossRef]
- Bavaresco, L.; Fregoni, M.; Trevisan, M.; Mattivi, F.; Vrhovsek, U.; Falchetti, R. The occurrence of the stilbene piceatannol in grapes. Vitis 2002, 41, 133–136. [Google Scholar]
- Langcake, P.; Cornford, C.A.; Pryce, R.J. Identification of pterostilbene as a phytoalexin from Vitis vinifera leaves. Phytochemistry, 1979, 18, 1025–1027. [Google Scholar] [CrossRef]
- Schmidlin, L.; Poutaraud, A.; Claudel, P.; Mestre, P.; Prado, E.; Santos-Rosa, M.; Wiedemann-Merdinoglu, S.; Karst, F.; Merdinoglu, D.; Hugueney, P. A stress-inducible resveratrol-O-methyltransferase involved in the biosynthesis of pterostilbene in grapevine. Plant Physiol. 2008, 148, 1630–1639. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.-M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [PubMed]
- Hall, D.; De Luca, V. Mesocarp localization of a bi-functional resveratrol/hydroxycinnamic acid glucosyltransferase of Concord grape (Vitis labrusca). Plant J. 2007, 49, 579–591. [Google Scholar] [CrossRef]
- Härtl, K.; Huang, F. C.; Giri, A. P.; Franz-Oberdorf, K.; Frotscher, J.; Shao, Y.; Hoffmann, T.; Schwab, W. Glucosylation of smoke-derived volatiles in grapevine (Vitis vinifera) is catalyzed by a promiscuous resveratrol/guaiacol glucosyltransferase. J. Agric. Food Chem. 2017, 65, 5681–5689. [Google Scholar] [CrossRef] [PubMed]
- Calderon, A.A.; Pedreno, M.A.; Munoz, R.; Ros Barcelo, A. Zymographic screening of plant peroxidase isoenzymes oxidizing 4-hydroxystilbenes. Electrophoresis 1990, 11, 507–508. [Google Scholar] [CrossRef]
- Calderon, A.A.; Zapata, J.M.; Pedreno, M.A.; Munoz, R.; Ros Barcelo, A. Levels of 4-hydroxystilbene-oxidizing isoperoxidases related to constitutive disease resistance in in vitro-cultured grapevine. Plant Cell, Tissue Organ Cult. 1992, 29, 63–70. [Google Scholar] [CrossRef]
- Morales, M.; Alcantara, J.; Ros Barcelo, A. Oxidation of trans-resveratrol by a hypodermal peroxidase isoenzyme from gamay rouge grape (Vitis vinifera) berries. Am. J. Enol. Vitic. 1997, 48, 33–38. [Google Scholar] [CrossRef]
- Langcake, P.; Pryce, R.J. Oxidative dimerisation of 4-hydroxystilbenes in vitro: production of a grapevine phytoalexin mimic. J.C.S. Chem. Comm. 1977, 7, 208–210. [Google Scholar] [CrossRef]
- Jeandet, P.; Sobarzo-Sánchez, E.; Sanches Silva, A.; Clément, C.; Nabavi, S.F.; Battino, M.; Rasekhian, M.; Belwal, T.; Habtemariam, S.; Koffas, M.; Nabavi, S.M. Whole-cell biocatalytic, enzymatic and green chemistry methods for the production of resveratrol and its derivatives. Biotechnol. Adv. 2020, 38, 107461. [Google Scholar] [CrossRef]
- Jeandet, P.; Bessis, R.; Sbaghi, M.; Meunier, P. Production of the phytoalexin resveratrol by grapes as a response to Botrytis attack under natural conditions. J. Phytopathol. 1995, 143, 135–139. [Google Scholar] [CrossRef]
- Bavaresco, L.; Petegolli, D.; Cantu, E.; Fregoni, M.; Chiusa, G.; Trevisan, M. Elicitation and accumulation of stilbene phytoalexins in grapevine berries infected by Botrytis cinerea. Vitis 1997, 36, 77–83. [Google Scholar]
- Dercks, W.; Creasy, L.L. The significance of phytoalexins in the Plasmopara viticola-grapevine interaction. Physiol. Mol. Plant Pathol. 1989, 34, 189–202. [Google Scholar] [CrossRef]
- Malacarne, G.; Vrhovsek, U.; Zulini, L.; Cestaro, A.; Stefanini, M.; Mattivi, F.; Delledonne, M.; Velasco, R.; Moser, C. Resistance to Plasmopara viticola in a grapevine segregating population is associated with stilbenoid accumulation and with specific host transcriptional responses. BMC Plant Biol. 2011, 11, 114. [Google Scholar] [CrossRef] [PubMed]
- Flamini, R.; Zanzotto, A.; de Rosso, M.; Lucchetta, G.; Dalla Vedova, A.; Bavaresco, L. Stilbene oligomer phytoalexins in grape as a response to Aspergillus carbonarius infection. Physiol. Mol. Plant Pathol. 2016, 93, 112–118. [Google Scholar] [CrossRef]
- Calzarano, F.; D’Agostino, V.; Pepe, A.; Osti, F.; Della Pelle, F.; De Rosso, M.; Flamini, R.; Di Marco, S. Patterns of phytoalexins in the grapevine leaf stripe disease symptoms (esca complex)/grapevine pathosystem. Phytopathol. Mediterr., 2016, 55, 410–426. [Google Scholar]
- Calzarano, F.; Osti, F.; D’Agostino, V.; Pepe, A.; Della Pelle, F.; De Rosso, M.; Flamini, R.; Di Marco, S. Levels of phytoalexins in vine leaves with different degrees of grapevine leaf stripe disease symptoms (Esca complex of diseases). Phytopathol. Mediterr., 2017, 56, 494–501. [Google Scholar]
- Adrian, M.; Jeandet, P.; Veneau, J.; Weston, L.A.; Bessis, R. Biological activity of resveratrol, a stilbenic compound from grapevines, against Botrytis cinerea, the causal agent for gray mold. J. Chem. Ecol. 1997, 23, 1689–1702. [Google Scholar] [CrossRef]
- Pezet, R.; Gindro, K.; Viret, O.; Richter, H. Effects of resveratrol, viniferins and pterostilbene on Plasmopara viticola zoospore mobility and disease development. Vitis 2004, 43, 143–145. [Google Scholar]
- Chalal, M.; Klinguer, A.; Echairi, A.; Meunier, P.; Vervandier-Fasseur, D.; Adrian, M. Antimicrobial activity of resveratrol analogues. Molecules 2014, 19, 7679–7688. [Google Scholar] [CrossRef]
- Pezet, R.; Pont, V. Mise en evidence de ptérostilbène dans les grappes de Vitis vinifera. Plant Physiol. Biochem. 1988, 26, 603–607. [Google Scholar]
- Alessandro, M.; Di Marco, S.; Osti, F. Cesari, A. bioassays on the activity of resveratrol, pterostilbene and phosphorous acid towards fungi associated with esca of grapevine. Phytoapathol. Mediterr. 2000, 39, 357–365. [Google Scholar]
- Gabaston, J.; Richard, T.; Biais, B.; Waffo-Teguo, P.; Pedrot, E.; Jourdes, M.; Corio-Costet, M.F.; Mérillon, J.M. Stilbenes from common spruce (Picea abies) bark as natural antifungal agent against downy mildew (Plasmopara viticola). Ind. Crop Prod. 2017, 103, 267–273. [Google Scholar] [CrossRef]
- Gabaston, J.; Cantos-Villar, E.; Biais, B.; Waffo-Teguo, P.; Renouf, E.; Corio-Costet, M.F.; Richard, T.; Mérillon, J.M. Stilbenes from Vitis vinifera L. waste: A sustainable tool for controlling Plasmopara viticola. J. Agric. Food Chem. 2017, 65, 2711–2718. [Google Scholar] [CrossRef] [PubMed]
- Elmer, P.A.G.; Reglinski, T. Biosupression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Jamiołkowska, A. Natural compounds as elicitors of plant resistance against diseases and new biocontrol strategies. Agronomy 2020, 10, 173. [Google Scholar] [CrossRef]
- Monteiro, E.; Gonçalves, B.; Cortez, I.; Castro, I. The Role of biostimulants as alleviators of biotic and abiotic stresses in grapevine: A review. Plants 2022, 11, 396. [Google Scholar] [CrossRef]
- Perrin, D.R.; Cruickshank, I.A.M. Studies on phytoalexins. VII. Chemical stimulation of pisatin formation in Pisum sativum. Aust. J. Biol. Sci. 1965, 18, 803–816. [Google Scholar] [CrossRef]
- Sweigard, J.A.; Matthews, D.E.; VanEtten, H.D. Synthesis of the phytoalexin pisatin by a methyltransferase from pea. Plant Physiol. 1986, 80, 277–279. [Google Scholar] [CrossRef]
- Hanawa, F.; Tahara, S.; Mizutani, J. Antifungal stress compounds from Veratrum grandiflorum leaves treated with cupric chloride. Phytochemistry 1992, 31, 3005–3007. [Google Scholar] [CrossRef]
- Aziz, A.; Trotel-Aziz, P.; Dhuicq, L.; Jeandet, P.; Couderchet, M.; Vernet, G. Chitosan oligomers and copper sulfate induce grapevine defense reactions and resistance to fungal pathogens. Phytopathology 2006, 96, 1188–1194. [Google Scholar] [CrossRef]
- Adrian, M.; Jeandet, P.; Bessis, R.; Joubert, J.M. Induction of phytoalexin (resveratrol) synthesis in grapevine leaves treated with aluminium chloride (AlCl3). J. Agric. Food Chem. 1996, 44, 1979–1981. [Google Scholar] [CrossRef]
- Magarey, P. A.; Wachtel, M. F.; Newton, M. R. Evaluation of phosphonate, fosetyl-Al and several phenylamide fungicides for post-infection control of grapevine downy mildew caused by Plasmopara viticola. Australas. Plant Pathol. 1991, 20, 34–40. [Google Scholar] [CrossRef]
- Gisi, U. Chemical control of downy mildews. In Advances in Downy Mildew Research, Spencer-Phillips, P.T.N.; Gisi, U.; Lebeda, A., Eds, Springer, The Netherlands, 2002, pp. 119–159.
- Rekanovic, E.; Potocnik, I.; Stepanovic, M.; Milijasevic, S.; Todorovic, B. Field efficacy of fluopicolide and fosetyl-Al fungicide combination (Profiler R©) for control of Plasmopara viticola (Berk. & Curt.) Berl. & Toni. in grapevine. Pestic. i Fitomedicina 2008, 23, 183–187. [Google Scholar]
- Koledenkova, K.; Esmaeel, Q.; Jacquard, C.; Nowak, J.; Clément, C.; Ait-Barka, E. Plasmopara viticola the causal agent of downy mildew of grapevine: From its taxonomy to disease management. Front. Microbiol. 2022, 13, 889472. [Google Scholar] [CrossRef]
- Di Marco, S.; Osti, F.; Calzarano, F.; Roberti, R.; Veronesi, A.; Amalfitano, C. Effects of grapevine applications of fosetyl-aluminium formulations for downy mildew control on “esca” and associated fungi. Phytopathol. Mediterr., 2011, 50, S285–S299. [Google Scholar] [CrossRef]
- Guest, D.I. Modification of defense responses in tobacco and capsicum following treatment with fosetyl-Al [aluminium tris (o-ethylphosphonate]. Physiol. Plant Pathol. 1984, 25, 125–134. [Google Scholar] [CrossRef]
- Guest, D.I. Evidence from light microscopy of living tissues that fosetyl-Al modifies the defense response in tobacco seedlings following inoculations by Phytphthora nicotianae var. nicotianae. Physiol. Plant Pathol. 1986, 29, 251–261. [Google Scholar] [CrossRef]
- Nemesthoty, G.S.; Guest, D.I. Phytoalexin accumulation, phenylalanine ammonia lyase activity and ethylene biosynthesis in fosetyl-Al treated resistant and susceptible tobacco cultivars infected by Phytophtora nicotianae var nicotianae. Physiol. Mol. Plant Pathol. 1990, 37, 207–219. [Google Scholar] [CrossRef]
- Afek, U.; Sztejnberg, A. Effects of fosetyl-Al and phosphorus acid on scoparone, a phytoalexin associated with resistance of Citrus to Phytophthora citrophthora. Phytopathology 1989, 79, 736–739. [Google Scholar] [CrossRef]
- Raynal, G.; Ravisé, A.; Bompeix, G. Action du tris-O-éthylphosphonate d’aluminium (phosétyl d’aluminium) sur la pathogénie de Plasmopara viticola et sur la stimulation des réactions de défense de la vigne. Ann. Phytopathol. 1980, 12, 163–175. [Google Scholar]
- Dercks, W.; Creasy, L.L. Influence of fosetyl-Al on phytoalexin accumulation in the Plasmopara viticola-grapevine interaction. Physiol. Mol. Plant Pathol. 1989, 34, 203–213. [Google Scholar] [CrossRef]
- Jeandet, P.; Bessis, R.; Adrian, M.; Joubert, J.M.; Yvin, J.C. Use of aluminium chloride as a resveratrol formation elicitor in plants. French Patent N° 95 13 642, PCT Int. Appl. WO 97 18,715, 1997. [Google Scholar]
- Jeandet, P.; Adrian, M.; Breuil, A.C.; Sbaghi, M.; Debord, S.; Bessis, R.; Weston, L.A.; Harmon, R. Chemical induction of phytoalexin synthesis in grapevines: Application to the control of grey mould in the vineyard. Acta Hortic. 2000, 528, 591–596. [Google Scholar] [CrossRef]
- Colby, S.R. Calculating synergistic and antagonistic responses of herbicide combinations. Weeds 1967, 15, 20–22. [Google Scholar] [CrossRef]
- Wang, R.; Duan, D.; Metzger, C.; Zhu, X.; Riemann, M.; Pla, M.; Nick, P. Aluminium can activate grapevine defense through actin remodeling. Hort. Res. 2022, 9, uhab016. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Marin, M.I.; Guerrero, R.F.; Puertas, B.; Garcia-Parrilla, M.C.; Collado, I.G.; Cantos-Villar, E. Impact of preharvest and postharvest treatment combinations on increase of stilbene content in grape. J. Int. Sci. Vigne Vin 2013, 47, 203–212. [Google Scholar] [CrossRef]
- Miliordos, D.-E.; Alatzas, A.; Kontoudakis, N.; Unlubayir, M.; Hatzopoulos, P.; Lanoue, A.; Kotseridis, Y. Benzothiadiazole affects grape polyphenol metabolism and wine quality in two Greek cultivars: Effects during ripening periods over two years. Plants 2023, 12, 1179. [Google Scholar] [CrossRef]
- Friedrich, L.; Lawton, K.; Ruess, W.; Masner, P.; Specker, N.; Gut Rella, M.; Meier, B. Dincher, S.; Staub, T.; Uknes, S. et al. A benzothiadiazole derivative induces systemic acquired resistance in tobacco. Plant J. 1996, 10, 61–70. [Google Scholar] [CrossRef]
- Görlach, J.; Volrath, S.; Knauf-Beiter, G.; Hengy, G.; Beckhove, U.; Kogel, K.H.; Oostendorp, M.; Staub, T.; Ward, E.; Kessmann, H.; et al. Benzothiadiazole, a novel class of inducers of systemic acquired resistance, activates gene expression and disease resistance in wheat. Plant Cell 1996, 8, 629–643. [Google Scholar]
- Iriti, M.; Rossoni, M.; Borgo, M.; Faoro, F. Benzothiadiazole enhances resveratrol and anthocyanin biosynthesis in grapevine, meanwhile improving resistance to Botrytis cinerea. J. Agric. Food Chem. 2004, 52, 4406–4413. [Google Scholar] [CrossRef]
- Harm, A.; Kassemeyer, H.H.; Seibicke, T.; Regner, F. Evaluation of chemical and natural resistance inducers against downy mildew (Plasmopara viticola) in grapevine. Am. J. Enol. Vitic. 2011, 62, 184–192. [Google Scholar] [CrossRef]
- Dufour, M.C.; Lambert, C.; Bouscaut, J.; Mérillon, J.M.; Coriot-Costet, M.F. Benzothiadiazole-primed defence responses and enhanced differential expression of defence genes in Vitis vinifera infected with biotrophic pathogens Erysiphe necator and Plasmopara viticola. Plant Pathol. 2013, 62, 370–382. [Google Scholar] [CrossRef]
- Jeandet, P.; Breuil, A.C.; Adrian, M.; Weston, L.A.; Debord, S.; Meunier, P.; Maume, G.; Bessis, R. HPLC analysis of grapevine phytoalexins coupling photodiode array detection and fluorometry. Anal. Chem. 1997, 69, 5172–5177. [Google Scholar] [CrossRef]
- Duke, S.O. Benefits of resveratrol and pterostilbene to crops and their potential nutraceutical value to mammals. Agriculture 2022, 12, 368. [Google Scholar] [CrossRef]
- Bellée, A.; Cluzet, S.; Dufour, M.-C.; Mérillon, J.-M.; Corio-Costet, M.-F. Comparison of the impact of two molecules on plant defense and on efficacy against Botrytis cinerea in the vineyard: A plant defense inducer (benzothiadiazole) and a fungicide (pyrimethanil). J. Agric. Food Chem. 2018, 66, 3338–3350. [Google Scholar] [CrossRef] [PubMed]
- Jeandet, P.; Clément, C.; Tisserant, L.P.; Crouzet, J.; Courot, E. Use of grapevine cell cultures for the production of phytostilbenes of cosmetic interest. C.R. Chimie 2016, 19, 1062–1070. [Google Scholar] [CrossRef]
- Larronde, F.; Gaudillière, J.P.; Krisa, S.; Descendit, A.; Deffieux, G.; Mérillon, J.M. Airborne methyl jasmonate induces stilbene accumulation in leaves and berries of grapevine plants. Am. J. Enol. Vitic. 2003, 54, 63–66. [Google Scholar] [CrossRef]
- Creasy, L.L.; Coffee, M. Phytoalexin production potential of grape berries. J. Am. Soc. Hortic. Sci. 1988, 113, 230–234. [Google Scholar] [CrossRef]
- Vezzulli, S.; Civardi, S.; Ferrari, F.; Bavaresco, L. Methyljasmonate treatment as a trigger of resveratrol synthesis in cultivated grapevine. Am. J. Enol. Vitic. 2007, 58, 530–533. [Google Scholar] [CrossRef]
- Belhadj, A.; Saigne, C.; Telef, N.; Cluzet, S.; Bouscaut, J.; Corio-Costet, M.-F.; Mérillon, J.-M. Methyl jasmonate induces defense responses in grapevine and triggers protection against Erysiphe necator, J. Agric. Food Chem. 2006, 2006 54, 9119–9125. [Google Scholar] [CrossRef]
- Burdziej, A.; Bellée, A.; Bodin, E.; Fonayet, J.V.; Magnin, N.; Szakiel, A.; Richard, T.; Cluzet, S.; Corio-Costet, M.-F. Three types of elicitors induce grapevine resistance against downy mildew via common and specific immune responses. J. Agric. Food Chem. 2021, 69, 1781–1795. [Google Scholar] [CrossRef]
- Smillie, R.; Grant, B.R.; Guest, D.I. The mode of action of phosphite: Evidence for both direct and indirect modes of action on three Phytophthora spp. in plants. Phytopathology 1989, 79, 921–926. [Google Scholar] [CrossRef]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The pivotal role of ethylene in plant growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Belhadj, A.; Telef, N.; Cluzet, S.; Bouscaut, J.; Corio-Costet, M.-F.; Mérillon, J.-M. Ethephon elicits protection against Erysiphe necator in grapevine. J. Agric. Food Chem. 2008, 56, 5781–5787. [Google Scholar] [CrossRef] [PubMed]
- Faurie, B.; Cluzet, S.; Corio-Costet, M.F.; Mérillon, J.-M. Methyl jasmonate/ethephon cotreatment synergistically induces stilbene production in Vitis vinifera cell suspensions but fails to trigger resistance to Erysiphe necator. J. Int. Sci. Vigne Vin 2009, 43, 99–110. [Google Scholar] [CrossRef]
- Delaunois, B.; Farace, G.; Jeandet, P.; Clément, C.; Bailleul, F.; Dorey, S.; Cordelier, S. Elicitors as alternative strategy to pesticides in grapevine? Current knowledge on their mode of action from controlled conditions to vineyard. Environ. Sci. Pollut. Res. 2014, 21, 4837–4846. [Google Scholar] [CrossRef] [PubMed]
- Gil-Munoz, R.; Fernandez-Fernandez, J.I.; Crespo-Villegas, O.; Garde-Cerdan, T. Elicitors used as a tool to increase stilbenes in grapes and wines. Food Res. Int. 2017, 98, 34–39. [Google Scholar] [CrossRef]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bezier, A.; Lambert, B.; Joubert, M.; Pugin, A. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microbe Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef]
- Stasinska-Jakubas, M.; Hawrylak-Nowak, B. Protective, biostimulating, and eliciting effects of chitosan and its derivatives on crop plants. Molecules 2022, 27, 2801. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Couderchet, M.; Vernet, G.; Aziz, A. Chitosan stimulates defense reactions in grapevine leaves and inhibits development of Botrytis cinerea. Eur. J. Plant Pathol. 2006, 114, 405–413. [Google Scholar] [CrossRef]
- Laquitaine, L.; Gomès, E.; François, J.; Marchive, C.; Pascal, S.; Hamdi, S.; Atanassova, R.; Delrot, S.; Coutos-Thévenot, P. Molecular basis of ergosterol-induced protection of grape against Botrytis cinerea: Induction of type I LTP promoter activity, WRKY, and stilbene synthase gene expression. Mol. Plant Microb. Interact. 2006, 19, 1103–1112. [Google Scholar] [CrossRef]
- Jakab, G.; Cottier, V.; Toquin, V.; Rigoli, G.; Zimmerly, L.; Metraux, J.P.; Mauch-Mani, B. Beta-aminobutyric acid-induced resistance in plants. Eur. J. Plant Pathol. 2001, 107, 29–37. [Google Scholar] [CrossRef]
- Ton, J.; Jakab, G.; Toquin, V.; Flors, V.; Iavicoli, A.; Maeder, M.N.; Métraux, J.P.; Mauch-Mani, B. Dissecting the β-aminobutyric acid–induced priming phenomenon in Arabidopsis. Plant Cell 2005, 17, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, A.R.; Hamiduzzaman, M. Md; Gindro, K.; Neuhaus, J.M.; Mauch-Mani, B. Beta-aminobutyric acid-induced resistance in grapevine against downy mildew: Involvement of pterostilbene. Eur. J. Plant Pathol. 2008, 122, 185–195. [Google Scholar] [CrossRef]
- Monnier, N.; Furlan, A.; Botcazon, C.; Dahi, A.; Mongelard, G.; Cordelier, S.; Clément, C.; Dorey, S.; Sarazin, C.; Rippa, S. Rhamnolipids from Pseudomonas aeruginosa are elicitors triggering Brassica napus protection against Botrytis cinerea without physiological disorders. Front. Plant Sci. 2018, 9, 1170. [Google Scholar] [CrossRef]
- Robineau, M; Le Guenic, S.; Sanchez, L.; Chaveriat, L.; Lequart, V.; Joly, N.; Calonne, M.; Jacquard, C.; Declerck, S.; Martin, P. et al. Synthetic mono-rhamnolipids display direct antifungal effects and trigger an innate immune response in tomato against Botrytis cinerea. Molecules 2020 25, 3108.
- Varnier, A. L.; Sanchez, L.; Vatsa, P.; Boudesocque, L.; Garcia-Brugger, A.; Rabenoelina, F.; Sorokin, A.; Renault, J.H.; Kauffmann, S.; Pugin, A. et al. Bacterial rhamnolipids are novel MAMPs conferring resistance to Botrytis cinerea. Plant Cell Environ. 2009, 32, 178–193. [Google Scholar] [CrossRef]
- Bolouri Moghaddam, M.R.B.; Van Den Ende, W. Sugars and plant innate immunity. J. Exp. Bot. 2012, 63, 3989–3998. [Google Scholar] [CrossRef]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, M. The role of sugars in plant responses to stress and their regulatory function during development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef]
- Mijailovic, N.; Nesler, A.; Perazzolli, M.; Ait Barka, E.; Aziz, A. Rare sugars: Recent advances and their potential role in sustainable crop protection. Molecules 2021, 26, 1720. [Google Scholar] [CrossRef]
- Chahed, A.; Lazazzara, V.; Moretto, M.; Nesler, A.; Corneo, P.E.; Ait-Barka, E.; Pertot, I.; Puopolo, G.; Perazzolli, M. The differential growth inhibition of Phytophthora spp. caused by the rare sugar tagatose is associated with species-specific metabolic and transcriptional changes. Front. Microbiol. 2021, 12, 711545. [Google Scholar] [CrossRef]
- Corneo, P.E.; Jermini, M.; Nadalini, S.; Giovannini, O.; Nesler, A.; Perazzolli, M.; Pertot, I. Foliar and root applications of the rare sugar tagatose control powdery mildew in soilless grown cucumbers. Crop Prot. 2021, 149, 105753. [Google Scholar] [CrossRef]
- Mijailovic, N.; Nesler, N.; Perazzolli, M.; Aziz, A.; Ait-Barka, E. Foliar application of a tagatose-based product reduces downy mildew symptoms through induction of grapevine resistance and anti-oomycete action. Agronomy 2022, 12, 498. [Google Scholar] [CrossRef]
- Mijailovic, N.; Richet, N.; Villaume, S.; Nesler, N.; Perazzolli, M.; Ait-Barka, E. Aziz, A. D-tagatose-based product triggers sweet immunity and resistance of grapevine to downy mildew, but not to gray mold disease. Plants 2022, 11, 296. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, J.A.; Heil, M.; Ahman, I.; Björkman, C. Optimizing crops for biocontrol of pests and disease. Trends Plant Sci. 2015, 20, 698–712. [Google Scholar] [CrossRef]
- Stenberg, J.A. A conceptual framework for integrated pest management Trends Plant Sci. 2017, 22, 759-769.
- Bakker, P.A.H.M.; Pieterse, C.M.J.; de Jonge, R.; Berendsen, R.L. The soil-borne legacy. Cell 2018, 172, 1178–1180. [Google Scholar] [CrossRef]
- Bhat, B.A.; Tariq, L.; Nissar, S.; Islam, S.T.; Islam, S.U.; Mangral, Z.; Ilyas, N.; Sayyed, R.Z.; Muthusamy, G.; Kim, W. et al. The role of plant-associated rhizobacteria in plant growth, biocontrol and abiotic stress management. J. Appl. Microbiol. 2022, 13, 2717–2741. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases –What has been achieved and what is the direction? Plant Pathol. 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Ait-Barka, E. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Perazzoli, M.; Moretto, M.; Fontana, P.; Ferrarini, A.; Velasco, R.; Moser, C.; Delledonne, M.; Pertot, I. Downy mildew resistance induced by Trichoderma harzianum T39 in susceptible grapevines partially mimics transcriptional changes of resistant genotypes. BMC Genomics 2012, 13, 660. [Google Scholar] [CrossRef] [PubMed]
- Arras, G.; d’Hallewin, G.; Molinu, M.G.; Dore, A.; Venditti, T.; Fois, M.; Lima, G.; Agabbio, M. Induction of phytoalexin biosynthesis in orange fruit by the biocontrol yeast Rhodotorula glutinis. Comm. Appl. Biol. Sci. Ghent University 2006, 71, 915–921. [Google Scholar]
- Van Peer, R.; Niemann, G.J.; Schippers, B. Induced resistance and phytoalexin accumulation in biological control of fusarium wilt of carnation by Pseudomonas sp strain WCS417r. Phytopathology 1991, 81, 728–734. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Trotel-Aziz, P.; Villaume, S.; Rabenoelina, F.; Clément, C.; Baillieul, F.; Aziz, A. Priming of camalexin accumulation in induced systemic resistance by beneficial bacteria against Botrytis cinerea and Pseudomonas syringae pv. tomato DC3000. J. Exp.Bot. 2022, 73, 3743–3757. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.; Chereyathmanjiyil, A.; Masih, I.; Chapuis, L.; Benoît, A. Biological control of Botrytis cinerea causing grey mould disease of grapevine and elicitation of stilbene phytoalexin (resveratrol) by a soil bacterium. FEMS Microbiol. Lett. 1998, 165, 65–70. [Google Scholar] [CrossRef]
- Verhagen, B.W.M. Verhagen, B.W.M., Trotel-Aziz, P.; Couderchet, M., Höfte, M.; Aziz, A. Pseudomonas spp.-induced resistance to Botrytis cinerea is associated with induction and priming of defence responses in grapevine. J. Exp. Bot. 2010, 61, 249-260.
- Verhagen, B.W.M.; Trotel-Aziz, P.; Jeandet, P.; Baillieul, F. M.; Aziz, A. Improved resistance against Botrytis cinerea by grapevine-associated bacteria that induce a prime oxidative burst and phytoalexin induction. Phytopathology 2011, 101, 768–777. [Google Scholar] [CrossRef]
- Gruau, C.; Trotel-Aziz, P.; Villaume, S.; Rabenoelina, F.; Clément, C.; Baillieul, F.; Aziz, A. Pseudomonas fluorescens PTA-CT2 triggers local and systemic immune response against Botrytis cinerea in grapevine. Mol. Plant Microb. Interact. 2015, 28, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Lakkis, S.; Trotel-Aziz, P.; Rabenoelina, F.; Schwarzenberg, A.; Nguema-Ona, E.; Clément, C.; Aziz, A. Strengthening grapevine resistance by Pseudomonas fluorescens PTA-CT2 relies on distinct defense pathways in susceptible and partially resistant genotypes to downy mildew and gray mold diseases. Front. Plant Sci. 2019, 10, 1112. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Trotel-Aziz, P.; Quantinet, D.; Biagianti, S.; Aziz, A. Biological control of Botrytis cinerea by selected grapevine-associated bacteria and stimulation of chitinase and β-1,3-glucanase activities under field conditions. Eur. J. Plant Pathol. 2007, 118, 43–57. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Quantinet, D.; Couderchet, M.; Aziz, A.; Trotel-Aziz, P. Differential induction of grapevine resistance and defense reactions against Botrytis cinerea by bacterial mixtures in vineyards. BioControl 2013, 58, 117–131. [Google Scholar] [CrossRef]
- Aziz, A.; Verhagen, B.; Magnin-Robert, M.; Couderchet, M.; Clément, C.; Jeandet, P.; Trotel-Aziz, P. Effectiveness of beneficial bacteria to promote systemic resistance of grapevine to gray mold as related to phytoalexin production in vineyards. Plant Soil 2016, 405, 141–153. [Google Scholar] [CrossRef]
- Jeandet, P.; Bessis, R.; Gautheron, B. The production of resveratrol (3,5,4’-trihydroxystilbene) by grape berries in different developmental stages. Am. J. Enol. Vitic. 1991, 42, 41–46. [Google Scholar] [CrossRef]
Figure 1.
Biosynthetic pathway from phenylalanine to resveratrol and subsequent metabolism. Abbreviations: PAL, phenylalanine ammonia lyase; C4H, cinnamate-4-hydroxylase; 4CL, cinnamoyl-CoA ligase; STS, stilbene synthase; CHS, chalcone synthase; ROMT, resveratrol-O-methyltransferase; PER, peroxidases; GT; Glycosyltransferases; UDPG, uridine diphospho-glucose. Note that isoprenylated stilbenes are not present in grapevine.
Figure 1.
Biosynthetic pathway from phenylalanine to resveratrol and subsequent metabolism. Abbreviations: PAL, phenylalanine ammonia lyase; C4H, cinnamate-4-hydroxylase; 4CL, cinnamoyl-CoA ligase; STS, stilbene synthase; CHS, chalcone synthase; ROMT, resveratrol-O-methyltransferase; PER, peroxidases; GT; Glycosyltransferases; UDPG, uridine diphospho-glucose. Note that isoprenylated stilbenes are not present in grapevine.
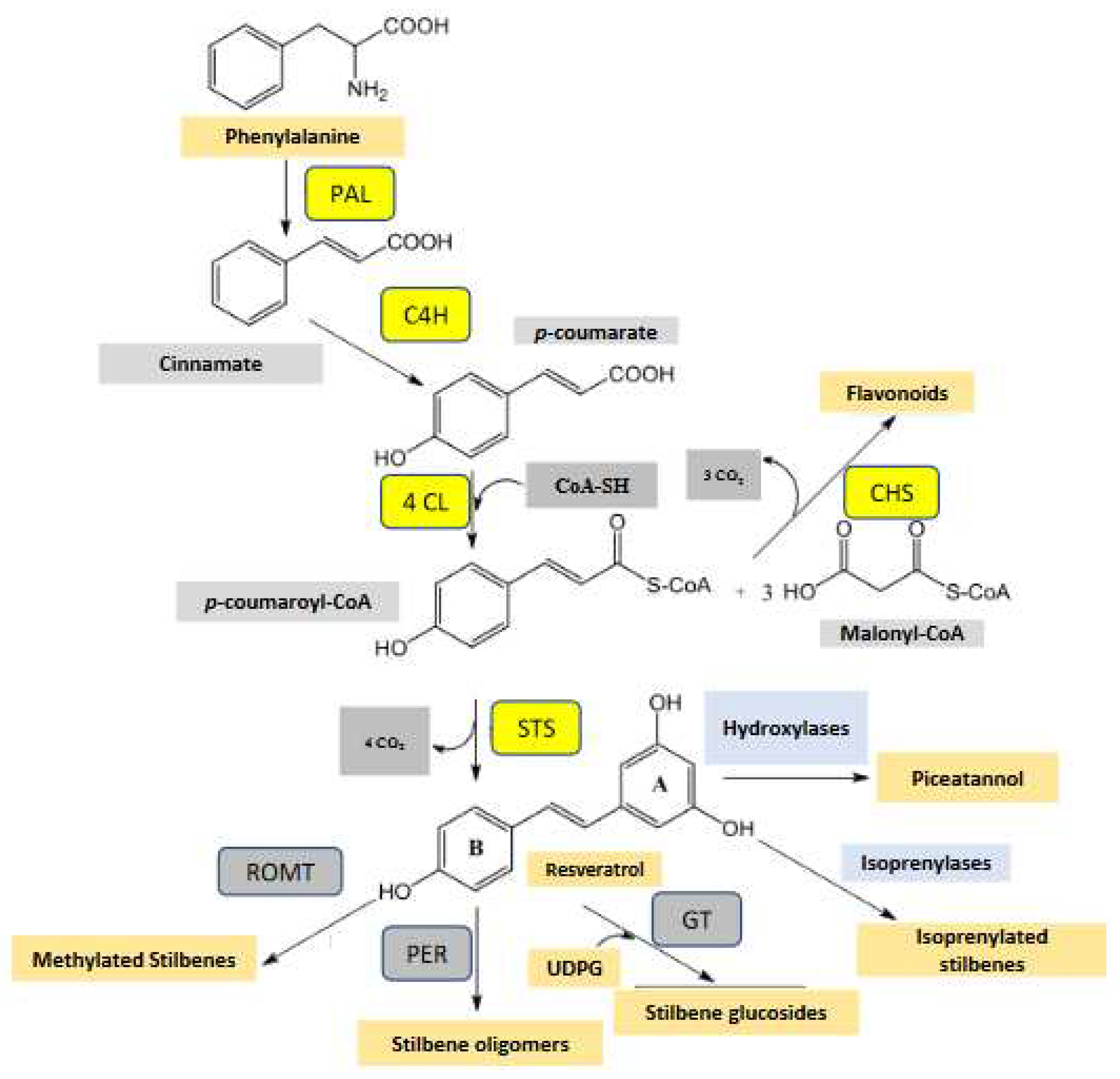
Figure 2.
Structures of the five stilbenes mainly evoked in this study. Stilbene monomers, resveratrol; pterostilbene and piceid (3-O-resveratrol-β-D-glucoside). Resveratrol dimers, δ-viniferin and ε-viniferin present in the Vitaceae. Glc= β-D-glucosyl moiety.
Figure 2.
Structures of the five stilbenes mainly evoked in this study. Stilbene monomers, resveratrol; pterostilbene and piceid (3-O-resveratrol-β-D-glucoside). Resveratrol dimers, δ-viniferin and ε-viniferin present in the Vitaceae. Glc= β-D-glucosyl moiety.
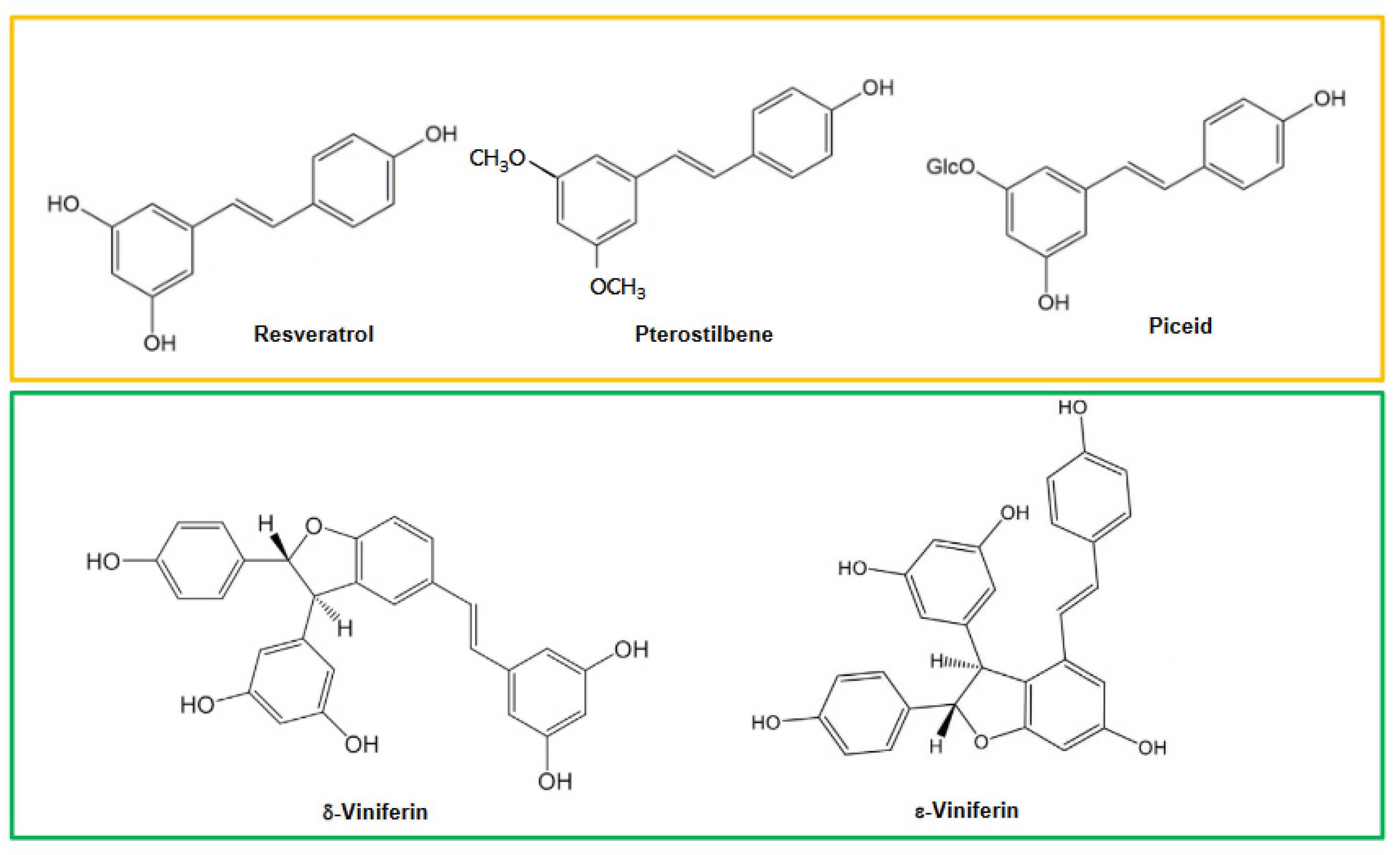
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



