
Submitted:
08 August 2023
Posted:
10 August 2023
You are already at the latest version
Abstract
Many redox diseases, including diabetes, are associated with an imbalance of calcium homeostasis, regulated mainly by a Ca2+-ATPase pump (SERCA). Dysfunction of this enzyme may be prevented by natural polyphenolic compounds, representing a possible supporting treatment. Compounds that increase SERCA activity/expression may be useful for the treatment of diabetic complications. Stimulation of SERCA1 activity was analyzed experimentally and by molecular modeling. SERCA1 activity under methylglyoxal- and palmitate-induced oxidative stress was evaluated. The viability of INS-1E cells and insulin secretion were determined. [6]-Gingerol, resveratrol, and ellagic acid increased SERCA1 activity and exerted a protective effect under oxidative stress in the noncellular system. We found for the first time that the binding of polyphenols ([6]-gingerol, resveratrol, ellagic acid) to Glu439 in the SERCA1 P-domain may be critical for the stimulation of its activity. Moreover, this binding may also be important in the protective effects against oxidative stress. Direct stimulation of SERCA1 activity by ellagic acid was observed for the first time. In INS-1E cells, these compounds increased insulin secretion.
Keywords:
SERCA activity
; insulin release
; pancreatic beta-cells
; polyphenolic compounds
1. Introduction
Intracellular calcium is a critical coordinator for many cellular events. The calcium concentration of 1 mM in the extracellular space and around 100 nM in the intracellular space is tightly regulated by several enzymes, channels, and transporters, resulting in precise calcium signals [1,2]. The main player in maintaining calcium homeostasis by accumulating released Ca2+ back into the sarco/endoplasmic reticulum (SR/ER) is the Ca2+-ATPase pump enzyme (SERCA). Many redox diseases, including diabetes, are associated with dysfunction of SERCA caused by its modified activity and/or expression [2,3].
SERCA isoforms are highly conserved in structure, with 75% or more homology between proteins from the SERCA1, SERCA2, and SERCA3 families [4]. In pancreatic beta-cells, three different isoforms (SERCA2a, SERCA2b, SERCA3) are expressed, with SERCA2b being the most prevalent one and representing the key regulator of ER Ca2+ transport in these cells [5].
Drugs maintaining correct calcium levels in SR/ER represent one of the novel therapeutic targets for the treatment of redox diseases. Natural phenolic compounds can bind to SERCA, thus modulating its activity [6]. Most of the phenolic compounds are known as SERCA inhibitors, and only a few are activators.
While small molecules able to lower SERCA activity or expression [7,8,9,10,11] have been proposed and used as anticancer drugs [12], SERCA activators, which increase enzyme activity and expression, may be used in the treatment of heart diseases and diabetes and are summarized in a review by Chemaly et al. [2].
Until now, no clear explanation exists regarding a structure-related molecular mechanism of SERCA allosteric activation. However, certain indications appeared in mutation studies concerning the modulatory function of ATP. Clausen et al. [13] found that the mutation Glu439Ala resulted in a significant increase in the SERCA dephosphorylation rate (E2P→E2 transition) when measured as a function of ATP concentration. Thus, the interaction of Glu439 with active ligands could contribute to an increase in SERCA activity, as dephosphorylation is the rate-limiting step (or set of steps) for the second period of the SERCA1a activity cycle [14].
In conditions of oxidative stress in vitro, natural polyphenolic compounds may regulate SERCA activity by their antioxidant ability, trapping of oxidants, and/or binding to SERCA. More complex conditions occur in the cellular system or in vivo, where also other factors contribute to SERCA regulation. SIRTs, nicotinamide adenine dinucleotide (NAD+)-dependent histone deacetylases, and the peroxisome proliferator-activated receptors (PPARs) are important regulators of SERCAs subjected to loss of function in conditions of diabetes. Another way of regulating SERCAs is the modulation of the activity of phosphodiesterases (PDEs), which degrade cAMP. PDEs belong to a class of enzymes that hydrolyze the phosphodiester bonds of cAMP and cGMP, thereby modulating various cellular signaling pathways [15].
SERCA2b is specifically downregulated in INS-1E cells in the presence of high glucose [16] and is directly regulated by the nuclear receptor PPARγ. Loss of beta-cell SERCA2b expression was observed in several models of type 2 diabetes [17,18]. SERCA2b likely plays a significant role in the regulation of insulin secretion [17,19].
Protection against beta-cell dysfunction may also be realized via the modulation of PDE activities associated with SERCA regulation; SERCA2 may be regulated by PDE3A in the human myocardium [20].
Overproduction of free fatty acids critically enhances the generation of reactive oxygen species (ROS), inducing lipotoxicity associated with endoplasmic reticulum (ER) stress, calcium dysregulation, mitochondrial dysfunction, and cell death [21]. Palmitate is one of the most abundant free fatty acids (FFA) in plasma [22,23] and a potential inductor of ROS generation in different types of cells [24,25,26]. Palmitate oxidation produces excessive electrons in the respiratory chain, resulting in the overproduction of superoxide dismutase. The molecular mechanisms of ROS production in cells induced by palmitate are still unexplained. The formation of ROS induced by palmitate results in an incorrect redox status of ER and the accumulation of abnormal clustered proteins [27].
A significant increase in FFA concentration was found in patients suffering from type 2 diabetes mellitus (T2DM) in comparison with control healthy people. The release of Ca2+, caused by palmitate-induced oxidative stress, resulted in calcium overloading in the cytosol and mitochondria, thus accelerating additional ROS generation in mitochondria. Calcium channel activation induced by the depletion of Ca2+ from ER increases the impairment of calcium homeostasis. Interruption of this vicious circle of ROS generation, as well as dysregulation of calcium homeostasis, may be a promising therapeutic aim for the prevention and treatment of metabolic diseases associated with lipotoxicity [27]. Oxidative stress induced by palmitate represents the main mechanism of the uncoupling of insulin signaling.
Palmitate is often used to examine the direct effects of lipotoxicity, and it may cause alterations by activating protein acetylation. Prolonged exposure to palmitate induces changes in beta-cell protein lysine acetylation, which leads to cell damage [28]. Acetylation dysregulated by sirtuins may be a target to counteract palmitate-induced beta-cell lipotoxicity.
MGX is a highly reactive compound derived mainly from glucose and fructose metabolism. It is implicated in diabetic complications as it is a strong advanced glycation end product (AGE) precursor involved in insulin resistance and beta-cell dysfunction [29,30], and it is a potent inducer of oxidative stress [31,32]. MGX reacts with DNA, lipids, and proteins mainly via arginine, lysine, and cysteine, producing AGEs [33]. Membrane permeable MGX may reach SERCA, which is sensitive to MGX-induced injury.
Our research is focused on activation of SERCA by polyphenolic compounds (ellagic acid, [6]-gingerol, resveratrol). In addition, resveratrol and [6]-gingerol derivatives were studied due to their better solubility, bioavailability, and higher antioxidant properties. The effects of the compounds tested were evaluated in the noncellular system of the rabbit sarcoplasmic reticulum from fast-twitch skeletal muscles. The aim of this study was to determine whether the scavenging/antioxidant effects of the polyphenols tested or the special binding of compounds to SERCA1 is responsible for the enzyme activity increase important in antidiabetic effects. The pancreatic INS-1E beta-cell line was used as a cellular system to analyze insulin release regulated by SERCA2b.
2. Results
Studies of natural SERCA activators were performed in the noncellular system of the sarcoplasmic reticulum of skeletal muscles, as well as in the cellular system of INS-1E cells. Oxidative stress was induced by palmitate or methylglyoxal.
In introductory experiments, we analyzed the dependence of SERCA1 activity on MGX and palmitate and the dependence of INS-1E cell viability on the concentrations of the same oxidants. Figure 1A shows the dependence of SERCA1 activity under control conditions and in the presence of MGX (IC50 = 2.98 mM). Likewise, Figure 1B depicts the dependence of SERCA1 activity in the absence and presence of palmitate (IC50 = 0.65 mM). Figure 1C shows the dependence of INS-1E viability on MGX (IC50 = 2.54 mM). Figure 1D presents the dependence of INS-1E viability on palmitate (IC50 = 0.41 mM).
2.1. [6]-Gingerol and [6]-shogaol
2.1.1. [6]-Gingerol and [6]-shogaol in the noncellular system
Figure 2A shows the effect of [6]-gingerol on SERCA1 activity in the absence of oxidants, with [6]-gingerol stimulating SERCA1 activity in the concentration range of 30 – 100 µM (EC50 = 36.94 µM). In contrast, its derivative [6]-shogaol decreased SERCA1 activity at concentrations of 75 and 100 µM (Figure 3A). As indicated in Figure 2B, [6]-gingerol protected MGX-induced injury in the concentration range of 100 – 200 µM; on the other hand, [6]-shogaol induced a significant decrease in SERCA1 activity (Figure 3B). In the presence of palmitate, [6]-gingerol (100 µM) induced only a partial increase in SERCA1 activity at various times of incubation (1 and 3 h) (Figure 2C). [6]-Shogaol induced a decrease in SERCA1 activity at concentrations of 20 µM (incubation times of 3 and 6 h) and 100 µM (incubation times of 1, 3, and 6 h) (Figure 3C). According to calculations of antioxidant/prooxidant activity and lipophilicity (Table 1), SHO appeared to be a better antioxidant and weaker prooxidant than GIN. As indicated by partition and distribution coefficients, SHO is a significantly more lipophilic compound.
2.1.2. [6]-Gingerol and [6]-shogaol in the cellular system
Concerning the effect of [6]-gingerol on the viability of INS-1E cells, a decrease in viability was observed in the concentration range of 50 – 150 µM (Figure 2D), and an additional viability decrease was found in the presence of MGX (2.5 mM) in the [6]-gingerol concentration range of 20 - 100 µM (Figure 2E). [6]-Shogaol induced a decrease in cell viability in the concentration range of 50 – 200 µM (Figure 3D), and an additional decrease in cell viability was found in the presence of MGX (2.5 mM) in the [6]-Shogaol concentration range of 100 – 200 µM (Figure 3E). In the presence of palmitate (0.4 mM), too, no protection of beta-cells was observed. In addition, [6]-shogaol significantly and time-dependently reduced the viability of damaged beta-cells at a concentration of 50 μM (Figure 3F).
2.2. Resveratrol and oxyresveratrol
2.2.1. Resveratrol and oxyresveratrol in the noncellular system
Resveratrol significantly increased SERCA1 activity in the concentration range of 100 – 200 µM (Figure 4A), and oxyresveratrol significantly decreased enzyme activity only at a concentration of 100 μM (Figure 5A). Resveratrol exerted a protective effect against MGX oxidation, as indicated by the concentration-dependent increase in SERCA1 activity in the concentration range of 50 – 300 µM (Figure 4B). In contrast, oxyresveratrol induced a decrease in SERCA1 activity in the concentration range of 10 – 100 µM under the same conditions (Figure 5B). Resveratrol protected SERCA1 activity against impairment induced by palmitate at a concentration of 100 µM during the whole time period of incubation, and a lower concentration was without any effect (Figure 4C). Oxyresveratrol exerted no protective effect; even a decrease in SERCA1 activity was observed at a concentration of 20 µM and with an incubation time of 6 h (Figure 5C). As theoretically predicted (Table 1), RES may be the most significant antioxidant of all agents tested. ORES exhibited lower antioxidant activity but higher prooxidant activity. RES is a more lipophilic compound than ORES.
2.2.2. Resveratrol and oxyresveratrol in the cellular system
Resveratrol decreased the viability of INS-1E cells in the concentration range of 50 – 150 µM (Figure 4D), as does oxyresveratrol in the same concentration range (Figure 5D). Under conditions of oxidative stress in the presence of MGX, no significant changes in cell viability were observed (Figure 4E). Under the same conditions, oxyresveratrol exerted a decrease in cell viability at concentrations of 100 and 150 µM (Figure 5E). Resveratrol induced no significant changes in cell viability in the presence of palmitate (0.4 mM), except at a concentration of 10 µM after 6 h of incubation, which induced a cell viability decrease (Figure 4F). However, oxyresveratrol increased the viability of cells after 15 h of incubation at the concentrations tested (10 and 50 µM). A concentration of 50 µM increased viability also after 3 h of incubation (Figure 5F).
Resveratrol exerted no effect on insulin release both under a normal glucose level (3 mM) and under a higher level of glucose (16.7 mM); a concentration of resveratrol of 100 µM even caused a decrease in insulin release (Figure 4G). Oxyresveratrol caused no changes in insulin release under a normal glucose level (3 mM). At the higher level of glucose (16.7 mM), a decrease in insulin release was observed at oxyresveratrol concentrations of 50 and 100 µM (Figure 5G).
2.3. Ellagic acid
2.3.1. Ellagic acid in the noncellular system
EA-induced SERCA1 activity increases in the concentration range of 30 – 100 µM (Figure 6A). EA (50 – 100 µM) also induced a SERCA1 activity increase under conditions of MGX-induced oxidative stress (Figure 6B). Protective effects of EA were also observed against palmitate-induced oxidative stress. EA (10 µM) increased SERCA1 activity for 2 min – 3 h, except for 6 h of incubation. EA (20 µM) increased SERCA1 activity during the whole incubation period (2 min – 6 h) (Figure 6C). According to our calculations (Table 1), EA was the weakest antioxidant and strongest prooxidant of all agents tested. This compound also exerted the weakest lipophilicity.
2.3.2. Ellagic acid in the cellular system
EA significantly decreased the viability of INS-1E beta-cells in the concentration range of 30 - 100 µM (Figure 6D). In the presence of MGX, an additional significant decrease in cell viability was observed at concentrations of 70 and 100 µM (Figure 6E).
EA (10 μM) increased cell viability with incubation periods of 3 - 24 h under palmitate-induced oxidative stress. EA (50 uM) increased viability only at longer incubation times (15 and 24 h) (Figure 6F). Insulin release in INS-1E beta-cells increased at both EA concentrations tested (10 and 50 µM) under a normal glucose level.
Under a higher glucose level, only a concentration of 50 µM induced an increase in insulin release (Figure 6G).
2.4. In silico study
Gingerol, resveratrol, and ellagic acid, the compounds able to increase SERCA activity, were bound to Glu439 either via a hydrogen bond ([6]-gingerol in Figure 7A and resveratrol in Figure 7B) or with a π-H bond (ellagic acid in Figure 7C). Moreover, ellagic acid also created an H-bond with Arg174 (Figure 7C).
We used theoretical calculations to compare the antioxidant and prooxidant properties of the agents tested (Table 1). EHOMO is the energy of the highest occupied molecular orbital and represents the electron-donating properties of the molecule—in other words, the antioxidant potential of the polyphenolic molecule. Conversely, ELUMO is the energy of the lowest unoccupied molecular orbital and represents the electron-accepting properties of the molecule—in other words, the prooxidant potential of the polyphenolic molecule. The higher EHOMO, the easier the donation of a polyphenolic electron. The lower EHOMO, the easier the acceptance of an external electron.
According to EHOMO, the antioxidant ability decreased in the following order: RES (- 507.4 > ORES (- 516.9) > SHO (- 530.4) > GIN (- 542.2) > EA (- 581.3). According to ELUMO, the prooxidant ability decreased in the following order: EA (- 196.4) > ORES (- 119.1) > RES (- 115.2) > GIN (- 78.4) > SHO (- 38.1). According to the partition and distribution coefficients, the lipophilicity decreased in the following order: SHO > GIN > RES > ORES > EA.
Taking the values of EHOMO and ELUMO (Table 1) as approximate measures of the antioxidant and prooxidant potentials, respectively, we find that resveratrol provides the best antioxidant effects. Ellagic acid, is the strongest prooxidant. The predicted values of the distribution coefficient indicate that gingerol and shogaol are the best at penetrating through a cellular membrane.
3. Discussion
Elevated levels of palmitate or methylglyoxal play a role in the oxidative impairment associated with diabetic complications. Therefore, we analyzed the effects of these oxidants in vitro in the sarcoplasmic reticulum (SR) of skeletal muscles, as well as in the cell system of pancreatic INS-1E beta-cells. MGX decreased Ca2+-ATPase activity in the SR of skeletal muscles (SERCA1) with an IC50 of 2.98 mM, and palmitate with an IC50 of 0.65 mM, indicating that palmitate is a more effective inhibitor of SERCA1. Concerning the effects of the above-mentioned oxidants in a model of INS-1E beta-cells, in which cell viability was evaluated, palmitate similarly induced a stronger impairment, with an IC50 of 0.41 mM, compared with an IC50 of 2.54 mM for MGX. This may probably be caused by different mechanisms of injury used by the different oxidants, in addition to the fact that both are inductors of ROS generation. MGX can cross the membrane and bind to specific amino acids (arginine, lysine, and cysteine) of SERCA [34]. Impairment induced by palmitate is characterized by its incorporation into the membrane [27]. Protective effects of natural polyphenolic compounds, diminishing the impairment induced by methylglyoxal and palmitate, were evaluated in noncellular as well as cellular systems to better understand their mechanism of action.
We suppose that the direct interaction of the compounds tested with crucial amino acids of SERCA1 induces an increase or decrease in SERCA1 activity. In the presence of oxidants, protection against SERCA impairment may be caused by the competition of polyphenolic compounds with oxidants or their oxidative products in binding to important binding sites, thereby influencing SERCA activity. Trapping of oxidants (MGX) may also occur. Additional mechanisms, such as the regulation of SERCA by sirtuins, PPARs, and PDEs, may be included in the cellular system.
3.1. [6]-Gingerol (GIN) and [6]-shogaol (SHO)
We found a concentration-dependent increase in SERCA1 activity, induced by [6]-gingerol (EC50 = 36.94 µM). Stimulation of SERCA from both skeletal and cardiac muscles was also found previously, suggesting direct SERCA protein activation [35].
Thermally labile gingerols easily undergo dehydration to form the corresponding shogaols, both exhibiting several identical biological activities, ranging from anticancer, antioxidant, antimicrobial, anti-inflammatory, and antiallergic activity [36]. In addition, ginger, ginger extracts, and shogaol could provide valuable protection against diabetes and cardiac and hepatic disorders [37]. Although [6]-gingerol and [6]-shogaol exerted identical biological effects, their mechanisms of action are probably different. Whereas [6]-gingerol increased SERCA1 activity, [6]-shogaol exerted no SERCA1 stimulation, and an inhibitory effect was observed at higher concentrations (75 and 100 μM). These differences may be caused by distinct interactions of these compounds with SERCA1. According to molecular modeling, we found that [6]-gingerol can bind to Glu439 of SERCA1, a typical binding site for SERCA activators, while [6]-shogaol was bound without direct interaction with this amino acid. Other authors tested the effects of 160 gingerol derivatives on SERCA. Their experimental results correlated with our molecular modeling. Derivatives able to stimulate SERCA1 possess at least 6 carbon chains, an o-methoxy phenolic group, and OH or NH groups situated behind the oxygen group. In this respect, [6]-gingerol can stimulate SERCA; however, [6]-shogaol is not able to do this [38]. Thus, it may be concluded that the structure of polyphenolic compounds important for SERCA stimulation may play a key role in the activation of SERCA and the protective effects in conditions of oxidative stress, as indicated below.
Opposite effects of these compounds were also observed in conditions of oxidative stress, in the presence of MGX. Whereas [6]-gingerol significantly and concentration-dependently protected SERCA1 activity against MGX impairment, [6]-shogaol was without any preventive effect. At higher concentrations (75 and 100 µM), shogaol exerted even inhibition, although [6]-shogaol was found to be a stronger antioxidant and scavenger than [6]-gingerol [39]. Also, according to our theoretical calculations using EHOMO and ELUMO values, SHO is a stronger antioxidant with higher lipophilicity than GIN (Table 1).
Antioxidant properties of [6]-gingerol were previously demonstrated by stimulation of superoxide dismutase, catalase, glutathione peroxidase, and GSH activities [40], while [6]-shogaol decreased activities/levels of antioxidant enzymes [41]. However, this is not the case in the sarcoplasmic reticulum, where we do not expect the presence of antioxidant enzymes. Both agents were able to effectively inhibit the formation of MGX-induced AGEs by trapping MGX in a time-dependent manner in the human serum albumin (HSA)-MGX system [42].
[6]-Gingerol (100 µM, incubation times of 1 and 3 h) partially increased SERCA1 activity in the presence of palmitate. [6]-Shogaol did not activate SERCA. Higher concentrations (75 – 100 µM) and prolonged incubation times even decreased SERCA1 activity.
We can suggest that SERCA activation by [6]-gingerol or its protective effect in oxidative stress conditions is not preferentially associated with its antioxidant/scavenging properties. Its ability to specifically interact with SERCA plays a significant role in enzyme activation or protection, as indicated experimentally and by molecular modeling. Direct interaction of [6]-gingerol and [6]-shogaol with SERCA is most important for SERCA1 activation/inhibition or protection against oxidants [38].
Although [6]-gingerol can activate SERCA1 and SERCA2a in microsome preparations, this compound does not appear to activate the SERCA2b pump isoform, based on experiments using intact NG115–401L neuronal cells that express only the SERCA2b isoform. The SERCA2b isoform is also known to be the largest of the SERCA proteins, containing an additional 49 C-terminal amino acid residues. These residues could alter the conformational states of the protein in such a way that [6]-gingerol binding may exert an inhibitory influence on one or more of the key partial reactions of the enzymatic cycle [43]. The authors concluded that there exists the possibility that [6]-gingerol, as opposed to acting like a pump stimulator in SR skeletal and cardiac microsomes, appears more likely to be a SERCA inhibitor when tested in the context of an intact cell assay. [6]-Gingerol probably exerts its effect indirectly via binding abilities and alterations on SERCA-associated regulatory proteins. Thus, it would be possible that [6]-gingerol could produce a very different effect on the SERCA2b pump expressed in NG115-401L cells because it can regulate a separate protein associated with the regulation of SERCA enzymatic function, as has been well documented for phospholamban, working in accordance with SERCA2a in cardiac muscles.
This agrees with our results on INS-1E pancreatic beta-cells, in which SERCA2b is the dominant SERCA. The viability of INS-1E pancreatic cells in our experiments was affected neither by the agents tested nor by conditions of oxidative stress.
Cell viability inhibition by [6]-gingerol was contrary to the increased insulin release at a higher concentration of glucose (16.7 mM), regulated by SERCA2b, which may suggest an adaptive mechanism. Increased plasma insulin levels after [6]-gingerol treatment were also observed in mice with type 2 diabetes induced by sodium arsenide, which improved impaired insulin signaling. In these experiments, [6]-gingerol was reported to act as a SERCA2a activator by improving diabetes-induced myocardial diastolic dysfunction and by enhancing the relaxation and the Ca2+ transient decay rate [35]. On the other hand, we found that [6]-shogaol did not affect insulin release in INS-1E cells.
We can conclude that [6]-gingerol and [6]-shogaol, agents with significant antidiabetic properties, act via different mechanisms. SERCA1 activation and the increase in insulin secretion in INS-1E cells may at least partially be involved in the antidiabetic effect of [6]-gingerol. On the other hand, the inhibitory effect of [6]-shogaol on adipogenesis and lipid accumulation may be one of its most significant protective effects. As previously reported, both [6]-gingerol and [6]-shogaol could assist in preventing diabetes by improving adipocyte dysfunction [44]. However, of all ginger components tested, [6]-shogaol had the most potent inhibitory effect on adipogenesis and lipid accumulation in 3T3-LI cells at a concentration of 40 μM [45,46]. [6]-Shogaol also exhibited an antidiabetic effect by decreasing the level of blood glucose, reducing body weight, and attenuating pathological changes to normal levels in diabetic mice. Furthermore, it has a beneficial effect on pancreas, kidney, and liver damage in diabetic mice. [6]-Shogaol also prevented damage from STZ-induced stress [37].
3.2. Resveratrol (RES) and oxyresveratrol (ORES)
We tested the effects of RES and its derivative ORES on SERCA1 activity in the noncellular system of the skeletal muscle sarcoplasmic reticulum and insulin secretion in pancreatic INS-1E beta-cells. Oxyresveratrol is a tetrahydroxy stilbene derivative, which is metabolically more stable and more widely distributed in the body than resveratrol. This derivative should be considered a potential lead compound for further drug development [47].
We found that RES increased SERCA1 activity; however, ORES exerted no stimulatory effect. This may be caused by different interactions between the tested compounds and SERCA1, as determined by molecular modeling. RES was able to interact with Glu439 in SERCA1, while OXYRES was not able to bind to this amino acid. RES also protected SERCA1 enzyme activity in conditions of oxidative stress induced by both MGX and palmitate. Protection of RES against MGX oxidation was concentration dependent (EC50 = 111.77 µM). ORES induced SERCA1 inhibition, although both compounds possess the ability to scavenge MGX [48,49]. Controversial results were found after oxidation with palmitate; a protective effect of RES and inhibition of ORES were observed. Antioxidant effects of RES and ORES may play a role against oxidative stress in SR. Our theoretical calculations, using EHOMO and ELUMO values, indicated that RES was only a slightly more potent antioxidant than ORES. On the other hand, it seems that ORES has significantly stronger prooxidant properties. However, according to the experimental results of Lorenz et al. [50], ORES, known for its lower cytotoxicity toward microglia and good water solubility, was found to be a better scavenger and antioxidant than RES. According to our calculations of the distribution and partition coefficients, RES is more lipophilic than ORES, so it can easier penetrate the SR membrane and, consequently, better reach SERCA.
We can conclude that the antioxidant and scavenging effects do not play a key role in the protective effects of RES against SERCA1 oxidative inhibition. We suppose that the protective effect of RES may be associated with its specific way of binding to SERCA1. The inhibitory effect of ORES may be related to its prooxidant and lipophilic properties.
In the cellular system, RES can regulate diabetes by several mechanisms, most of which are at least partially associated with SERCA. The first mechanism is its ability to activate the NAD+-dependent deacetylase of SIRT1, which increased 8-fold in the presence of RES [51]. Drastically reduced expression of SERCA2a was found in both types of diabetes [52,53,54]. It was also observed that SIRT1 acts as a transcriptional activator of SERCA2a gene expression in high-glucose conditions. These results demonstrate that RES acting via SIRT1 could regulate SERCA2a expression in diabetic mice [55].
Most studies on the protective effect of RES in conditions of diabetes focused on the expression of SERCA2a in the heart. However, some investigations focused on SERCA2b in beta-cells, playing a dominant role among SERCA isoforms in these cells. SERCA2b is specifically downregulated in the presence of a high glucose concentration. Numerous clinical and preclinical studies clearly demonstrate that agents that act as agonists of PPARγ have direct effects on the improvement of pancreatic beta-cell function and survival in T2DM [56]. SERCA2b from INS-1 pancreatic beta-cells is directly regulated by the nuclear receptor PPARγ [17]. Under both normal and stressed conditions, transient overexpression of SERCA2b significantly increased insulin release in INS-1 cells [17].
PPARs represent the second way of diabetic disorder improvement regulated by SERCA and are important targets in diabetes therapy. PPAR𝛾 is expressed in a variety of tissues, including the islet beta-cell [57,58]. It may play a role in beta-cell insulin release, is able to improve islet function, and may directly activate genes encoding SERCAs [59]. Resveratrol was demonstrated to act as a partial agonist of the PPARγ receptor, which opens the possibility for its use in the treatment of metabolic disorders.
Of the 56 resveratrol derivatives, ORES was one of the two tetrahydroxy stilbene derivatives with better solubility and affinity toward PPARγ [47]. RES (10 and 50 µM) exerted no protection on the viability of INS-1E pancreatic beta-cells in the presence of palmitate. On the other hand, ORES possessed a protective effect, which may be due to its more significant scavenging and antioxidant properties [50]; its better affinity toward PPARγ may, however, also play a role. Neither RES nor ORES influenced the viability of INS-1E cells in the presence of MGX.
RES has been reported to act as a PDE inhibitor in various cell types, including beta-cell lines and human islets. RES enhances beta-cell function by regulating PDE expression and activity, thus increasing intracellular cAMP levels and subsequent insulin release [60]. In our experiments, we found no increase in insulin secretion. This may be caused by the fact that other types of pancreatic beta-cells (β-Min6 cells) were used by Rouse et al. [60].
3.4. Ellagic acid (EA)
Ellagic acid (EA) is a natural polyphenolic antioxidant with potent anti-inflammatory, anticancerogenic, antidiabetic, cardioprotective, and other health-protective properties. Antioxidant and antiapoptotic effects of EA were found to be associated with a significant overexpression of SIRT1 in renal tissues, which is the key factor in cell survival [61].
Ellagic acid also increased the deacetylase activity of SIRT6 up to 50-fold. SIRT6 regulates distinct cellular functions, genome stability, DNA repair, and inflammation of related diseases. SIRT6 indirectly inhibits gluconeogenesis by inactivating a key transcriptional regulator of gluconeogenesis, PPARγ [62,63].
According to our knowledge, the effect of EA on skeletal muscle SERCA1 has never been studied. However, EA increased the affinity for Ca2+ of cardiac SERCA2a, most likely by direct interaction with PLB [64,65]. It has also been previously concluded that EA is the activator of myocardial SERCA2a [35]. Diabetes mellitus-induced myocardial diastolic dysfunction is partly caused by a reduction in SERCA2a function and can be ameliorated by SERCA activators, including ellagic acid.
However, we found for the first time a direct stimulating effect of EA on SERCA1. According to our results, EA concentration-dependently increased SERCA1 activity with EC50 = 33.19 µM. The experimental results were in agreement with the molecular modeling, suggesting that EA interacts with Glu439 and Arg174 via hydrogen bonds. In the presence of MGX and palmitate, EA exerted protective effects on SERCA1 activity under conditions of oxidative stress. The protective effects of EA in SR against oxidative stress may be associated either with its antioxidant effect (which is relatively low) or with its more feasible interaction with SERCA1. However, of all the agents tested by molecular modeling, EA exerted the lowest antioxidant effect and exhibited the weakest lipophilicity (Table 1).
In the system of pancreatic INS-1E beta-cells, which contain mostly SERCA2b, EA (10 µM) decreased cell viability, while an increase in insulin secretion was observed at normal as well as elevated concentrations of glucose. We suppose that there exists a time delay between the effects of EA on viability and the insulin release in cells, which may be an adaptive reaction to a decrease in cell viability. Despite a decrease in cell viability induced by EA alone, a protective effect of EA in conditions of palmitate-induced oxidative stress was observed. In the presence of palmitate, EA (10 µM) exerted a time-dependent increase in beta-cell viability. EA (50 µM) was effective only at 15 and 24 h of incubation. The protective effect of EA may be caused by its antioxidant effects induced via the PPARγ signaling pathway [66]. However, no protective effect of EA on the viability of INS-1E cells in MGX-induced oxidative stress was observed.
Regulation of the mRNA and proteins of p38-MAPK, MEKK1, and c-Jun by EA, which in turn upregulates PPARγ expression, immediately followed by caspase-3 activation, finally leads to necrosis, apoptosis, and cell death [67].
This is probably a dual effect, with, on the one hand, the presence of EA leading to apoptosis through caspase-3 activation and, on the other hand, EA protecting cells from MGX via antioxidant activity and against palmitate damage through sirtuin activation. Other authors concluded that EA lowered glucose and lipid levels in type 2 diabetic animals by inhibiting beta-cell apoptosis and stimulating insulin production [68]. Our results are in line with the conclusions of Qiu et al. [67], who performed experiments at the cell level. On the other hand, Harakeh et al. [68] used diabetic animals for their experiments. According to Harakeh et al. [68], STZ causes a notable reduction in insulin release by the destruction of pancreatic beta-cells, which may lead to an increase in glucose levels. A significant decrease in glucose level was observed in EA-treated diabetic rats compared with non-treated rats [68]. One possible mechanism of glucose decrease may be the potentiation of the pancreatic release of insulin from beta-cells. Similarly, we also observed an increase in insulin in beta-cells as a protective effect of EA against oxidative stress induced by palmitate.
4. Materials and methods
4.1. Compounds evaluated
Resveratrol was purchased from Career Henan Chemical Co. (Zhengzhou, People’s Republic of China) and had >98% purity. Oxyresveratrol was isolated from Morus nigra roots by using a multistep chromatographic purification technique as published before [69]. [6]-Gingerol was purified from a commercial ginger extract purchased from Xi’an Pincredit Bio-Tech Co., Ltd. (Xi’an, People’s Republic of China) by flash chromatography using a CombiFlash Rf+ Lumen instrument equipped with an integrated evaporative light-scattering detector (Teledyne Isco, Lincoln, NE, USA), a RediSep Gold silica column, and a gradient elution of acetone (from 0% to 15%) in n-hexane. An aliquot of 4 g of extract was separated, and a 36.8% yield of [6]-gingerol was obtained. [6]-Shogaol was subsequently semi-synthesized from [6]-gingerol using the method reported by Wei et al. [70] with slight modifications. Briefly, p-toluenesulphonic acid (TsOH) was used as a dehydrating agent and toluene as a solvent under reflux for 15 min. The product was purified by solid-phase extraction over silica using n-hexane–acetone (8:2, v/v), followed by flash chromatography on the above-mentioned instrument, using a gradient elution of 0%–5% of acetone in n-hexane and achieving a yield of 46.5%.
4.2. SERCA1 activity measurement
The activity of Ca2+-ATPase from the sarcoplasmic reticulum (SERCA1) isolated from the fast-twitch skeletal muscle of a New Zealand female rabbit was measured by two spectrophotometric methods modified by us. The first method was the NADH-coupled enzyme assay outlined by Ortiz et al. [71]. The second method was based on measurements of p-nitrophenyl phosphate (pNPP) activity, with the release of p-nitrophenyl (pNP) from pNPP being detected as outlined by Rossi et al. [72]. SR vesicles (1 mg prot./mL) were incubated for different times at 37 °C, pH 7.2.
4.2.1. NADH-coupled enzyme assay
SERCA1 activity was measured spectrophotometrically by the NADH-coupled enzyme assay, as outlined by Ortiz et al. [71] and modified by us. An enzyme-coupled NADH-linked ATPase assay was used to measure SERCA1 ATPase activity in 96-well microplates. The final amount of SR vesicles was 1.25 μg/well. Each well contained an assay mix (50 mM MOPS, pH 7.0), 100 mM KCl, 5 mM MgCl2, 1 mM EGTA, 0.2 mM NADH, 1 mM phosphoenolpyruvate, 10 IU/mL of pyruvate kinase, 10 IU/mL of lactate dehydrogenase, and 1 µM of the calcium ionophore A23187 (Sigma-Aldrich, St. Louis, MO, USA).
The total volume of the reaction mixture was 0.25 mL. The reaction was started by the addition of CaCl2 (10 µM). The reaction rate was determined by measuring the decrease in NADH absorbance at 340 nm, at 37 °C, on a microplate reader (Infinite M200, Tecan, Switzerland).
4.2.2. pNPP hydrolysis assay
SERCA1 activity was measured spectrophotometrically by the pNPP hydrolysis enzyme assay, as outlined by Rossi et al. [72]. The p-nitrophenyl phosphatase activity was measured at 37 ˚C in a medium containing 26 mM Tris-HCl (pH 7.4), 0.05 mM CaCl2, 120 mM KCl, 10 mM MgCl2, and 10–20 µg/mL Ca2+-ATPase, as well as the individual compound concentrations specified for each experiment, in a final volume of 0.23 mL. After different incubation periods, the reactions were started with 0.03 mL of pNPP to a final concentration of 3 mM. The pNP produced was estimated by measuring the absorbance at 400 nm recorded with a microplate reader (Infinite M200, Tecan, Switzerland).
4.3. Cell culture
The INS-1E insulinoma pancreatic beta-cell line (kindly provided by Prof. Claes Wollheim, University of Geneva) was cultured in RPMI 1640 (11 mM glucose, Sigma Aldrich). RPMI 1640 was supplemented with 100 U/mL penicillin, 100 µg/mL streptomycin, 2 mM L-glutamine, 1 mM Na-pyruvate, 55 µM 2-mercaptoethanol, 10 mM HEPES, 1% nonessential amino acids, and 10% fetal bovine serum (pH 7.0–7.4). The cells were grown in a humidified incubator containing 5% CO2 at 37 °C. Cells were incubated with various concentrations of the individual compounds for different incubation periods and afterward measured using a cytotoxicity assay (MTT) or an insulin release assay.
4.4. Cytotoxicity assay (MTT)
The MTT (Sigma Aldrich) reduction assay was used as an indicator of cell damage and performed according to a standard protocol. INS-1E cells (5 × 104 cells per well) were seeded into 96-microwell plates. The cells were preincubated for different time periods with or without various concentrations of individual compounds (5% CO2, at 37 °C). MTT was added to a final concentration of 0.5 mg/mL. After incubation for a period of 4 h, the MTT medium was removed before the addition of DMSO and was let to stand for 15 min to solubilize the formazan formed. The absorbance was recorded at 500 nm with a microplate reader (Infinite M200, Tecan, Switzerland).
4.5. Insulin release
The insulin release as the response to glucose was measured in INS-1E beta-cells between passages 10 and 30. Briefly, after 24 h of preincubation of beta-cells with individual compounds, the cells were washed with a glucose-free KRBH buffer. After that, they were incubated for 30 min at 37 °C in a glucose-free KRBH buffer and then incubated for 30 min at 37 °C in a KRBH buffer supplemented with 3 mM glucose (normal level) or 16.7 mM glucose (pathological level). Afterward, the supernatants were collected and detected by an RIA kit (Mercodia, Sweden) for insulin release using rat insulin as standard.
4.6. In silico study
The optimal structures of the ligands and the corresponding values of the highest occupied molecular orbital (EHOMO) and the lowest unoccupied molecular orbital (ELUMO) were obtained by Spartan software (Spartan’20, Version 1.1.4) using the conformer search method and the MMFF94 force field and subsequent optimization using the DFT B3LYP 6-31G* method [73]. The docking study was performed by the Molecular Operating Environment (MOE 2020.0901) modeling program [74], using a triangle matcher and the London dG score for basic docking and the GBVI/WSA DG score for the induced-fit refinement of the geometry. Ligands were observed for protonation under a physiological pH. Ellagic acid was treated as dianion [75]. We used the pdb structure 2zbe for the E2P state. Proteins were treated with a QuickProp protocol to correct the bonds and protonation state. The values of the distribution coefficient (h logD, pH = 7.4) and partition coefficient (h logP) were estimated using the menu Descriptors-Calculate in the program MOE.
4.7. Statistical analysis
Statistical analysis was performed using the Bonferroni test. Values are given as the mean ± SD of at least three or more independent experiments, with each sample being measured in duplicate or triplicate. Statistical significance was set as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Corresponding graphs were created using the GraphPad Prism 8 program.
5. Conclusions
Natural polyphenolic compounds and related derivatives were tested in the noncellular system of the skeletal muscle sarcoplasmic reticulum and in the INS-1E cell line, with respect to SERCA1 enzyme activation and insulin secretion induced by SERCA2b. For [6]-gingerol, resveratrol, and ellagic acid, activation of skeletal muscle SERCA1 may be associated with binding to amino acid Glu439 of SERCA1. Derivatives of the above-mentioned compounds, [6]-shogaol and oxyresveratrol, which cannot bind to Glu439, exerted no SERCA1 activation. We can conclude that not the scavenging/antioxidant properties but the mode of interaction with SERCA is crucial for protection against methylglyoxal and palmitate oxidative stress. We found experimentally and confirmed by molecular modeling, for the first time, the direct activation of SERCA1 activity by ellagic acid.
In INS-1E cells, [6]-gingerol, resveratrol, ellagic acid (SERCA1 activators) increased insulin release regulated by SERCA2b. SERCA activation/expression may be one of the possible regulators of the antidiabetic effect of natural polyphenolic compounds.
Author Contributions
Vladimir Heger, Mgr., Ph.D.: experimental and validation work, visualization, collection of resources, editing. Barbora Benesova, Ph.D. student: experimental work. Magdalena Majekova, Dr., Ph.D.: molecular modeling. Petronela Rezbarikova, Mgr., Ph.D.: collection of resources and editing. Attila Huynadi, Dr., Ph.D.: conceptualization and supervised Ph.D. students preparing and analyzing curcumin, gingerol, and their derivatives. Gábor Girst, Ph.D. student: preparing curcumin and THCU, evaluation of NMR spectra. Sara Hassan Hassan Ahmed, Ph.D. student: all work concerning [6]-gingerol and [6]-shogaol. Tímea Gonda, postdoc: conceptualization and started all studies on ginger and co-supervised Sara Ahmed’s work on this. Lubica Horakova, Dr., Ph.D., corresponding author: conceptualization and writing of the manuscript, supervised Barbora Benesova.
Funding
This research was funded by Slovak National grant APVV-20-0543, VEGA 2/0113/21, VEGA 2/0063/22, VEGA 2/0103/22, National Research, Development, and Innovation Office (NKFIH), Hungary (K- 134704), and project no. TKP2021-EGA-32, implemented with the support provided by the Ministry of Innovation and Technology of Hungary from the NKFIH, financed under the TKP2021-EGA funding scheme.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giorgi, C.; Danese, A.; Missiroli, S.; Patergnani, S.; Pinton, P. Calcium Dynamics as a Machine for Decoding Signals. Trends Cell Biol 2018, 28, 258–273. [Google Scholar] [CrossRef] [PubMed]
- Chemaly, E.R.; Troncone, L.; Lebeche, D. SERCA Control of Cell Death and Survival. Cell Calcium 2018, 69, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Urano, F. Wolfram Syndrome: Diagnosis, Management, and Treatment. Curr Diab Rep 2016, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Remmen, H. Van The SarcoEndoplasmic Reticulum Calcium ATPase (SERCA) Pump: A Potential Target for Intervention in Aging and Skeletal Muscle Pathologies. Skelet Muscle 2021, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, I.X.; Raghavan, M.; Satin, L.S. The Endoplasmic Reticulum and Calcium Homeostasis in Pancreatic Beta Cells. Endocrinology 2020, 161. [Google Scholar] [CrossRef]
- Ogunbayo, O.A.; Harris, R.M.; Waring, R.H.; Kirk, C.J.; Michelangeli, F. Inhibition of the Sarcoplasmic/Endoplasmic Reticulum Ca2+-ATPase by Flavonoids: A Quantitative Structure-Activity Relationship Study. IUBMB Life 2008, 60, 853–858. [Google Scholar] [CrossRef]
- Kranias, E.G.; Hajjar, R.J. Modulation of Cardiac Contractility by the Phopholamban/SERCA2a Regulatome. Circ Res 2012, 110, 1646–1660. [Google Scholar] [CrossRef]
- Bhupathy, P.; Babu, G.J.; Periasamy, M. Sarcolipin and Phospholamban as Regulators of Cardiac Sarcoplasmic Reticulum Ca2+ ATPase. J Mol Cell Cardiol 2007, 42, 903–911. [Google Scholar] [CrossRef]
- Anderson, D.M.; Anderson, K.M.; Chang, C.-L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R.; et al. A Micropeptide Encoded by a Putative Long Noncoding RNA Regulates Muscle Performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef]
- Kho, C.; Lee, A.; Jeong, D.; Oh, J.G.; Chaanine, A.H.; Kizana, E.; Park, W.J.; Hajjar, R.J. SUMO1-Dependent Modulation of SERCA2a in Heart Failure. Nature 2011, 477, 601–605. [Google Scholar] [CrossRef]
- Bidasee, K.R.; Zhang, Y.; Shao, C.H.; Wang, M.; Patel, K.P.; Dincer, U.D.; Besch, H.R. Diabetes Increases Formation of Advanced Glycation End Products on Sarco(Endo)Plasmic Reticulum Ca2+-ATPase. Diabetes 2004, 53, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Bakhshi, J.; Weinstein, L.; Poksay, K.S.; Nishinaga, B.; Bredesen, D.E.; Rao, R. V Coupling Endoplasmic Reticulum Stress to the Cell Death Program in Mouse Melanoma Cells: Effect of Curcumin. Apoptosis 2008, 13, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Clausen, J.D.; McIntosh, D.B.; Woolley, D.G.; Andersen, J.P. Modulatory ATP Binding Affinity in Intermediate States of E2P Dephosphorylation of Sarcoplasmic Reticulum Ca2+-ATPase. Journal of Biological Chemistry 2011, 286, 11792–11802. [Google Scholar] [CrossRef]
- Clausen, J.D.; Andersen, J.P. Glutamate 90 at the Luminal Ion Gate of Sarcoplasmic Reticulum Ca2+-ATPase Is Critical for Ca2+ Binding on Both Sides of the Membrane. Journal of Biological Chemistry 2010, 285, 20780–20792. [Google Scholar] [CrossRef] [PubMed]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, U.; Elumalai, S.; Moon, J.S.; Jeon, J.H.; Kim, N.D.; Park, K.G.; Won, K.C.; Leem, J.; Lee, I.K. Myricetin Protects against High Glucose-Induced β-Cell Apoptosis by Attenuating Endoplasmic Reticulum Stress via Inactivation of Cyclin-Dependent Kinase 5. Diabetes Metab J 2019, 43, 192–205. [Google Scholar] [CrossRef]
- Kono, T.; Ahn, G.; Moss, D.R.; Gann, L.; Zarain-Herzberg, A.; Nishiki, Y.; Fueger, P.T.; Ogihara, T.; Evans-Molina, C. PPAR-γ Activation Restores Pancreatic Islet SERCA2 Levels and Prevents β-Cell Dysfunction under Conditions of Hyperglycemic and Cytokine Stress. Molecular Endocrinology 2012, 26, 257–271. [Google Scholar] [CrossRef]
- Yamamoto, W.R.; Bone, R.N.; Sohn, P.; Syed, F.; Reissaus, C.A.; Mosley, A.L.; Wijeratne, A.B.; True, J.D.; Tong, X.; Kono, T.; et al. Endoplasmic Reticulum Stress Alters Ryanodine Receptor Function in the Murine Pancreatic β Cell. Journal of Biological Chemistry 2019, 294, 168–181. [Google Scholar] [CrossRef]
- Lockridge, A.; Jo, S.; Gustafson, E.; Damberg, N.; Mohan, R.; Olson, M.; Abrahante, J.E.; Alejandro, E.U. Islet O-GlcNAcylation Is Required for Lipid Potentiation of Insulin Secretion through SERCA2. Cell Rep 2020, 31, 107609. [Google Scholar] [CrossRef]
- Ahmad, F.; Shen, W.; Vandeput, F.; Szabo-Fresnais, N.; Krall, J.; Degerman, E.; Goetz, F.; Klussmann, E.; Movsesian, M.; Manganiello, V. Regulation of Sarcoplasmic Reticulum Ca2+ ATPase 2 (SERCA2) Activity by Phosphodiesterase 3A (PDE3A) in Human Myocardium. Journal of Biological Chemistry 2015, 290, 6763–6776. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Effect of High-Fat Diets on Oxidative Stress, Cellular Inflammatory Response and Cognitive Function. Nutrients 2019, 11, 2579. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, A.I.S.; Pitt, S.J.; Smith, T.K.; Ajjan, R.A.; Stewart, A.J. Lipidomic Profiling of Plasma Free Fatty Acids in Type-1 Diabetes Highlights Specific Changes in Lipid Metabolism. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2021, 1866, 158823. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, A.I.S.; Blindauer, C.A.; Stewart, A.J. Changes in Plasma Free Fatty Acids Associated with Type-2 Diabetes. Nutrients 2019, 11, 2022. [Google Scholar] [CrossRef] [PubMed]
- Bagherieh, M.; Kheirollahi, A.; Zamani-Garmsiri, F.; Emamgholipour, S.; Meshkani, R. Folic Acid Ameliorates Palmitate-Induced Inflammation through Decreasing Homocysteine and Inhibiting NF-ΚB Pathway in HepG2 Cells. Arch Physiol Biochem 2021, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Villanueva, A.H.; Oun, A.; Buist-Homan, M.; Blokzijl, H.; Faber, K.N.; Dolga, A.; Moshage, H. Protective Effect of Metformin against Palmitate-Induced Hepatic Cell Death. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2020, 1866, 165621. [Google Scholar] [CrossRef]
- Sadeghi, A.; Rostamirad, A.; Seyyedebrahimi, S.; Meshkani, R. Curcumin Ameliorates Palmitate-Induced Inflammation in Skeletal Muscle Cells by Regulating JNK/NF-KB Pathway and ROS Production. Inflammopharmacology 2018, 26, 1265–1272. [Google Scholar] [CrossRef]
- Ly, L.D.; Xu, S.; Choi, S.-K.; Ha, C.-M.; Thoudam, T.; Cha, S.-K.; Wiederkehr, A.; Wollheim, C.B.; Lee, I.-K.; Park, K.-S. Oxidative Stress and Calcium Dysregulation by Palmitate in Type 2 Diabetes. Exp Mol Med 2017, 49, e291–e291. [Google Scholar] [CrossRef]
- Ciregia, F.; Bugliani, M.; Ronci, M.; Giusti, L.; Boldrini, C.; Mazzoni, M.R.; Mossuto, S.; Grano, F.; Cnop, M.; Marselli, L.; et al. Palmitate-Induced Lipotoxicity Alters Acetylation of Multiple Proteins in Clonal β Cells and Human Pancreatic Islets. Sci Rep 2017, 7, 13445. [Google Scholar] [CrossRef]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol Rev 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Matafome, P.; Sena, C.; Seiça, R. Methylglyoxal, Obesity, and Diabetes. Endocrine 2013, 43, 472–484. [Google Scholar] [CrossRef]
- Desai, K.M.; Wu, L. FREE RADICAL GENERATION BY METHYLGLYOXAL IN TISSUES. Drug Metabol Drug Interact 2008, 23. [Google Scholar] [CrossRef]
- Dhar, A.; Desai, K.; Kazachmov, M.; Yu, P.; Wu, L. Methylglyoxal Production in Vascular Smooth Muscle Cells from Different Metabolic Precursors. Metabolism 2008, 57, 1211–1220. [Google Scholar] [CrossRef]
- Chang, T.; Wu, L. Methylglyoxal, Oxidative Stress, and Hypertension. Can J Physiol Pharmacol 2006, 84, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Zizkova, P.; Viskupicova, J.; Heger, V.; Rackova, L.; Majekova, M.; Horakova, L. Dysfunction of SERCA Pumps as Novel Mechanism of Methylglyoxal Cytotoxicity. Cell Calcium 2018, 74. [Google Scholar] [CrossRef] [PubMed]
- Namekata, I.; Hamaguchi, S.; Wakasugi, Y.; Ohhara, M.; Hirota, Y.; Tanaka, H. Ellagic Acid and Gingerol, Activators of the Sarco-Endoplasmic Reticulum Ca2+-ATPase, Ameliorate Diabetes Mellitus-Induced Diastolic Dysfunction in Isolated Murine Ventricular Myocardia. Eur J Pharmacol 2013, 706, 48–55. [Google Scholar] [CrossRef]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A.M. Gingerols and Shogaols: Important Nutraceutical Principles from Ginger. Phytochemistry 2015, 117, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.K.; Ryoo, Z.Y.; Ha, J.J.; Oh, D.Y.; Kim, M.O.; Kim, S.H. Beneficial Effects of 6-Shogaol on Hyperglycemia, Islet Morphology and Apoptosis in Some Tissues of Streptozotocin-Induced Diabetic Mice. Diabetol Metab Syndr 2019, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.H.H.; Gonda, T.; Hunyadi, A. Medicinal Chemistry Inspired by Ginger: Exploring the Chemical Space around 6-Gingerol. RSC Adv 2021, 11, 26687–26699. [Google Scholar] [CrossRef]
- Dugasani, S.; Pichika, M.R.; Nadarajah, V.D.; Balijepalli, M.K.; Tandra, S.; Korlakunta, J.N. Comparative Antioxidant and Anti-Inflammatory Effects of [6]-Gingerol, [8]-Gingerol, [10]-Gingerol and [6]-Shogaol. J Ethnopharmacol 2010, 127, 515–520. [Google Scholar] [CrossRef]
- Chakraborty, D.; Mukherjee, A.; Sikdar, S.; Paul, A.; Ghosh, S.; Khuda-Bukhsh, A.R. [6]-Gingerol Isolated from Ginger Attenuates Sodium Arsenite Induced Oxidative Stress and Plays a Corrective Role in Improving Insulin Signaling in Mice. Toxicol Lett 2012, 210, 34–43. [Google Scholar] [CrossRef]
- Annamalai, G.; Kathiresan, S.; Kannappan, N. [6]-Shogaol, a Dietary Phenolic Compound, Induces Oxidative Stress Mediated Mitochondrial Dependant Apoptosis through Activation of Proapoptotic Factors in Hep-2 Cells. Biomedicine & Pharmacotherapy 2016, 82, 226–236. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, Y.; Wang, P.; Ahmedna, M.; Sang, S. Bioactive Ginger Constituents Alleviate Protein Glycation by Trapping Methylglyoxal. Chem Res Toxicol 2015, 28, 1842–1849. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bose, D.D.; Thomas, D.W. Paradoxical Effects of Sarco/Endoplasmic Reticulum Ca2+-ATPase (SERCA) Activator Gingerol on NG115-401L Neuronal Cells: Failure to Augment ER Ca2+ Uptake and Protect against ER Stress-Induced Cell Death. Eur J Pharmacol 2015, 762, 165–173. [Google Scholar] [CrossRef]
- Isa, Y.; Miyakawa, Y.; Yanagisawa, M.; Goto, T.; Kang, M.-S.; Kawada, T.; Morimitsu, Y.; Kubota, K.; Tsuda, T. 6-Shogaol and 6-Gingerol, the Pungent of Ginger, Inhibit TNF-α Mediated Downregulation of Adiponectin Expression via Different Mechanisms in 3T3-L1 Adipocytes. Biochem Biophys Res Commun 2008, 373, 429–434. [Google Scholar] [CrossRef]
- Tzeng, T.-F.; Liu, I.-M. 6-Gingerol Prevents Adipogenesis and the Accumulation of Cytoplasmic Lipid Droplets in 3T3-L1 Cells. Phytomedicine 2013, 20, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. The Chemical and Pharmacological Basis of Ginger (Zingiber Officinale Roscoe) as Potential Therapy for Diabetes and Metabolic Syndrome. In Medicinal Foods as Potential Therapies for Type-2 Diabetes and Associated Diseases; 2019; pp. 639–687. [Google Scholar]
- Pavlović, N.; Đanić, M.; Stanimirov, B.; Goločorbin-Kon, S.; Stankov, K.; Lalić-Popović, M.; Mikov, M. In Silico Discovery of Resveratrol Analogues as Potential Agents in Treatment of Metabolic Disorders. Curr Pharm Des 2019, 25, 3776–3783. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, Z.; Sheng, Z. Ability of Resveratrol to Inhibit Advanced Glycation End Product Formation and Carbohydrate-Hydrolyzing Enzyme Activity, and to Conjugate Methylglyoxal. Food Chem 2017, 216, 153–160. [Google Scholar] [CrossRef]
- Wang, W.; Yang, R.; Yao, H.; Wu, Y.; Pan, W.; Jia, A.-Q. Inhibiting the Formation of Advanced Glycation End-Products by Three Stilbenes and the Identification of Their Adducts. Food Chem 2019, 295, 10–15. [Google Scholar] [CrossRef]
- Lorenz, P.; Roychowdhury, S.; Engelmann, M.; Wolf, G.; Horn, T.F.W. Oxyresveratrol and Resveratrol Are Potent Antioxidants and Free Radical Scavengers: Effect on Nitrosative and Oxidative Stress Derived from Microglial Cells. Nitric Oxide 2003, 9, 64–76. [Google Scholar] [CrossRef]
- Sonnett, T.E.; Levien, T.L.; Gates, B.J.; Robinson, J.D.; Campbell, R.K. Diabetes Mellitus, Inflammation, Obesity: Proposed Treatment Pathways for Current and Future Therapies. Annals of Pharmacotherapy 2010, 44, 701–711. [Google Scholar] [CrossRef]
- Pereira, L.; Matthes, J.; Schuster, I.; Valdivia, H.H.; Herzig, S.; Richard, S.; Gómez, A.M. Mechanisms of [Ca2+]i Transient Decrease in Cardiomyopathy of Db / Db Type 2 Diabetic Mice. Diabetes 2006, 55, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Ahmed, S.; Grupp, I.L.; Matlib, M.A. Altered SR Protein Expression Associated with Contractile Dysfunction in Diabetic Rat Hearts. American Journal of Physiology-Heart and Circulatory Physiology 2001, 281, H1137–H1147. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.M.; Zhong, Y.; Hoit, B.D.; Grupp, I.L.; Hahn, H.; Dilly, K.W.; Guatimosim, S.; Lederer, W.J.; Matlib, M.A. Defective Intracellular Ca 2+ Signaling Contributes to Cardiomyopathy in Type 1 Diabetic Rats. American Journal of Physiology-Heart and Circulatory Physiology 2002, 283, H1398–H1408. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, M.; Matta, M.J.; Sunderesan, N.R.; Gupta, M.P.; Periasamy, M.; Gupta, M. Resveratrol, an Activator of SIRT1, Upregulates Sarcoplasmic Calcium ATPase and Improves Cardiac Function in Diabetic Cardiomyopathy. American Journal of Physiology-Heart and Circulatory Physiology 2010, 298, H833–H843. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Kono, T.; Evans-Molina, C. The Role of Peroxisome Proliferator-Activated Receptor γ in Pancreatic β Cell Function and Survival: Therapeutic Implications for the Treatment of Type 2 Diabetes Mellitus. Diabetes Obes Metab 2010, 12, 1036–1047. [Google Scholar] [CrossRef]
- Dubois, M.; Pattou, F.; Kerr-Conte, J.; Gmyr, V.; Vandewalle, B.; Desreumaux, P.; Auwerx, J.; Schoonjans, K.; Lefebvre, J. Expression of Peroxisome Proliferator-Activated Receptor γ (PPARγ) in Normal Human Pancreatic Islet Cells. Diabetologia 2000, 43, 1165–1169. [Google Scholar] [CrossRef]
- Rosen, E.D.; Kulkarni, R.N.; Sarraf, P.; Ozcan, U.; Okada, T.; Hsu, C.-H.; Eisenman, D.; Magnuson, M.A.; Gonzalez, F.J.; Kahn, C.R.; et al. Targeted Elimination of Peroxisome Proliferator-Activated Receptor γ in β Cells Leads to Abnormalities in Islet Mass without Compromising Glucose Homeostasis. Mol Cell Biol 2003, 23, 7222–7229. [Google Scholar] [CrossRef]
- Zarain-Herzberg, A.; Alvarez-Fernandez, G. Sarco(Endo)Plasmic Reticulum Ca 2+ -ATPase-2 Gene: Structure and Transcriptional Regulation of the Human Gene. The Scientific World JOURNAL 2002, 2, 1469–1483. [Google Scholar] [CrossRef]
- Rouse, M.; Younès, A.; Egan, J.M. Resveratrol and Curcumin Enhance Pancreatic β-Cell Function by Inhibiting Phosphodiesterase Activity. Journal of Endocrinology 2014, 223, 107–117. [Google Scholar] [CrossRef]
- Mohammed, E.T.; Hashem, K.S.; Abdelazem, A.Z.; Foda, F.A.M.A. Prospective Protective Effect of Ellagic Acid as a SIRT1 Activator in Iron Oxide Nanoparticle-Induced Renal Damage in Rats. Biol Trace Elem Res 2020, 198, 177–188. [Google Scholar] [CrossRef]
- Dominy, J.E.; Lee, Y.; Jedrychowski, M.P.; Chim, H.; Jurczak, M.J.; Camporez, J.P.; Ruan, H.-B.; Feldman, J.; Pierce, K.; Mostoslavsky, R.; et al. The Deacetylase Sirt6 Activates the Acetyltransferase GCN5 and Suppresses Hepatic Gluconeogenesis. Mol Cell 2012, 48, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Rahnasto-Rilla, M.; Järvenpää, J.; Huovinen, M.; Schroderus, A.-M.; Ihantola, E.-L.; Küblbeck, J.; Khadeer, M.; Moaddel, R.; Lahtela-Kakkonen, M. Effects of Galloflavin and Ellagic Acid on Sirtuin 6 and Its Anti-Tumorigenic Activities. Biomedicine & Pharmacotherapy 2020, 131, 110701. [Google Scholar] [CrossRef]
- Prestle, J.; Quinn, F.; Smith, G. Ca2+-Handling Proteins and Heart Failure: Novel Molecular Targets? Curr Med Chem 2003, 10, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Coll, K.E.; Johnson, R.G.; McKenna, E. Relationship between Phospholamban and Nucleotide Activation of Cardiac Sarcoplasmic Reticulum Ca 2+ Adenosinetriphosphatase. Biochemistry 1999, 38, 2444–2451. [Google Scholar] [CrossRef]
- Chen, J.; Yang, H.; Sheng, Z. Ellagic Acid Activated PPAR Signaling Pathway to Protect Ileums Against Castor Oil-Induced Diarrhea in Mice: Application of Transcriptome Analysis in Drug Screening. Front Pharmacol 2020, 10. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhou, B.; Jin, L.; Yu, H.; Liu, L.; Liu, Y.; Qin, C.; Xie, S.; Zhu, F. In Vitro Antioxidant and Antiproliferative Effects of Ellagic Acid and Its Colonic Metabolite, Urolithins, on Human Bladder Cancer T24 Cells. Food and Chemical Toxicology 2013, 59, 428–437. [Google Scholar] [CrossRef]
- Harakeh, S.; Almuhayawi, M.; Jaouni, S. Al; Almasaudi, S.; Hassan, S.; Amri, T. Al; Azhar, N.; Abd-Allah, E.; Ali, S.; El-Shitany, N.; et al. Antidiabetic Effects of Novel Ellagic Acid Nanoformulation: Insulin-Secreting and Anti-Apoptosis Effects. Saudi J Biol Sci 2020, 27, 3474–3480. [Google Scholar] [CrossRef]
- Supervisor, Z.Z. Antispasmodic Activity of Prenylated Phenolic Compounds from the Root Bark of Morus Nigra. Molecules 2019, 24, 2497. [Google Scholar]
- Wei, C.-K.; Tsai, Y.-H.; Korinek, M.; Hung, P.-H.; El-Shazly, M.; Cheng, Y.-B.; Wu, Y.-C.; Hsieh, T.-J.; Chang, F.-R. 6-Paradol and 6-Shogaol, the Pungent Compounds of Ginger, Promote Glucose Utilization in Adipocytes and Myotubes, and 6-Paradol Reduces Blood Glucose in High-Fat Diet-Fed Mice. Int J Mol Sci 2017, 18, 168. [Google Scholar] [CrossRef]
- Aguayo-Ortiz, R.; Creech, J.; Jiménez-Vázquez, E.N.; Guerrero-Serna, G.; Wang, N.; da Rocha, A.M.; Herron, T.J.; Espinoza-Fonseca, L.M. A Multiscale Approach for Bridging the Gap between Potency, Efficacy, and Safety of Small Molecules Directed at Membrane Proteins. Sci Rep 2021, 11, 16580. [Google Scholar] [CrossRef]
- Rossi, J.P.F.C.; Garrahan, P.J.; Rega, A.F. The Activation of Phosphatase Activity of the Ca2+-ATPase from Human Red Cell Membranes by Calmodulin, ATP and Partial Proteolysis. Biochimica et Biophysica Acta (BBA) - Biomembranes 1986, 858, 21–30. [Google Scholar] [CrossRef]
- Shao, Y.; Molnar, L.F.; Jung, Y.; Kussmann, J.; Ochsenfeld, C.; Brown, S.T.; Gilbert, A.T.B.; Slipchenko, L. V; Levchenko, S. V; O’Neill, D.P.; et al. Advances in Methods and Algorithms in a Modern Quantum Chemistry Program Package. Phys Chem Chem Phys 2006, 8, 3172–3191. [Google Scholar] [CrossRef] [PubMed]
- Vilar, S.; Cozza, G.; Moro, S. Medicinal Chemistry and the Molecular Operating Environment (MOE): Application of QSAR and Molecular Docking to Drug Discovery. Curr Top Med Chem 2008, 8, 1555–1572. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Terauchi, M.; Kikuchi, Y.; Nakao, A.; Okubo, J.; Yoshinaga, T.; Hiratsuka, H.; Kobayashi, M.; Hoshi, T. Deprotonation Processes of Ellagic Acid in Solution and Solid States. Monatshefte für Chemie /Chemical Monthly 2003, 134, 811–821. [Google Scholar] [CrossRef]
Figure 1.
Dependence of SERCA1 activity in SR and viability of INS-1E pancreatic beta-cells on oxidative stress induced by MGX and palmitate. SERCA1 activity in SR (A) in the absence or presence of increasing concentrations of MGX and (B) in the absence or presence of increasing concentrations of palmitate. Viability of INS-1E cells (C) in the absence or presence of increasing concentrations of MGX and (D) in the absence or presence of increasing concentrations of palmitate. Individual values are averages of 3 independent measurements.
Figure 1.
Dependence of SERCA1 activity in SR and viability of INS-1E pancreatic beta-cells on oxidative stress induced by MGX and palmitate. SERCA1 activity in SR (A) in the absence or presence of increasing concentrations of MGX and (B) in the absence or presence of increasing concentrations of palmitate. Viability of INS-1E cells (C) in the absence or presence of increasing concentrations of MGX and (D) in the absence or presence of increasing concentrations of palmitate. Individual values are averages of 3 independent measurements.
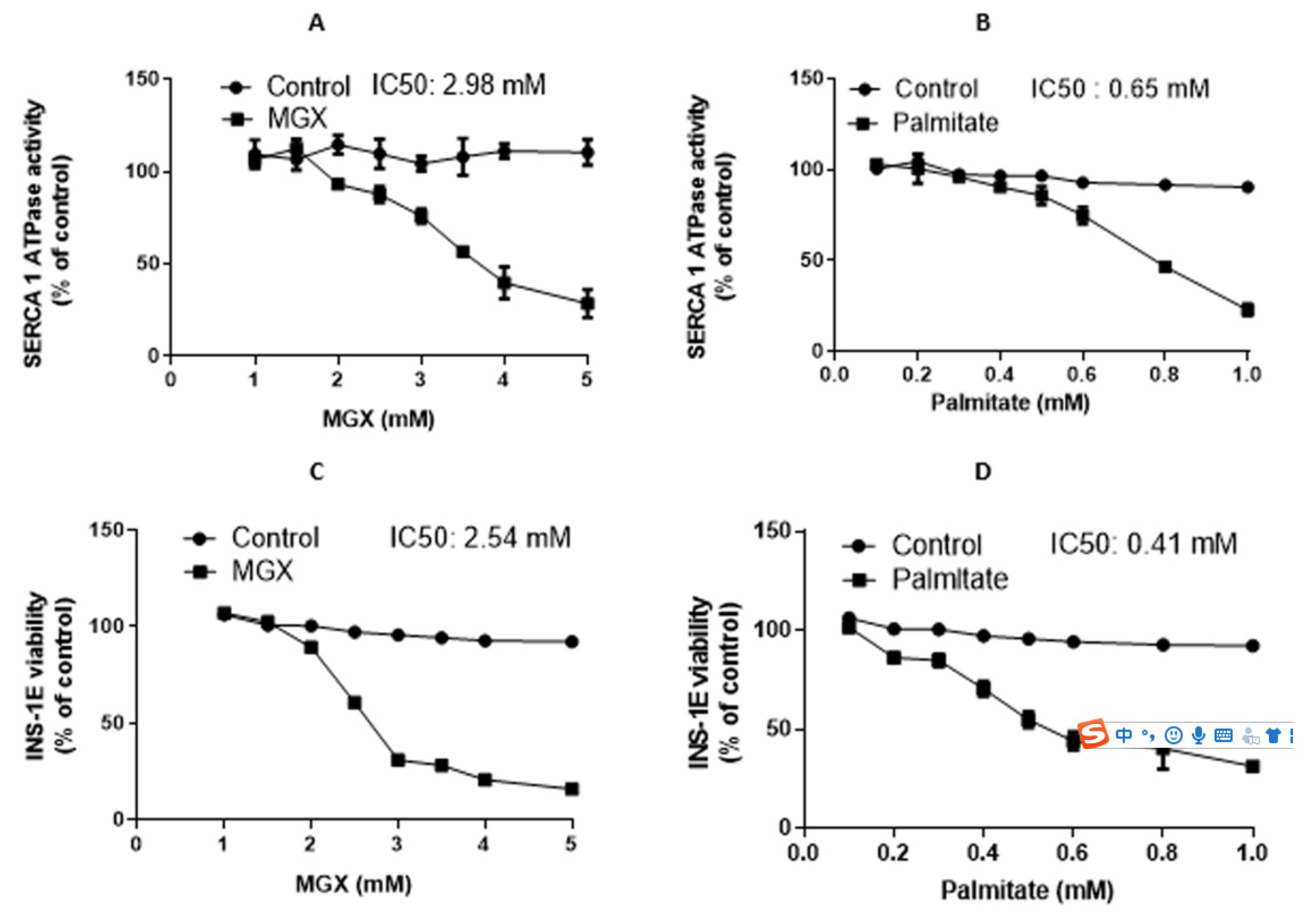
Figure 2.
Effect of [6]-gingerol (GIN) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of GIN. (B) Effect of GIN on MGX pre-treated SR vesicles. (C) GIN mediated time-dependent effect on SERCA1 activity impaired by palmitate. ns - nonsignificant changes. (D) Viability of INS-1E beta-cells in the presence of GIN. (E) Viability of INS-1E beta-cells in the presence of MGX and GIN (10 – 100 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and GIN (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
Figure 2.
Effect of [6]-gingerol (GIN) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of GIN. (B) Effect of GIN on MGX pre-treated SR vesicles. (C) GIN mediated time-dependent effect on SERCA1 activity impaired by palmitate. ns - nonsignificant changes. (D) Viability of INS-1E beta-cells in the presence of GIN. (E) Viability of INS-1E beta-cells in the presence of MGX and GIN (10 – 100 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and GIN (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
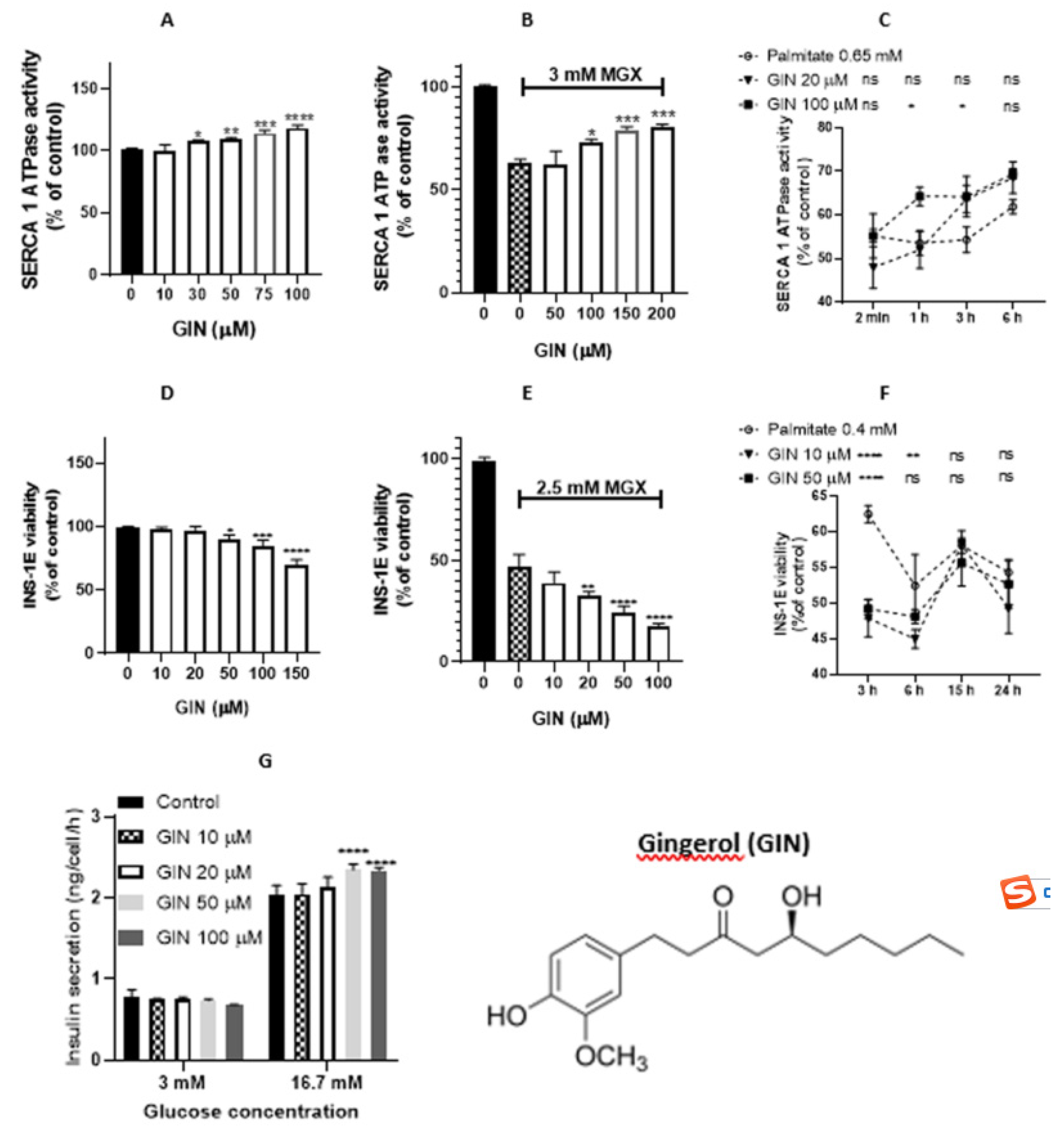
Figure 3.
Effect of [6]-Shogaol (SHO) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of SHO. (B) Effect of SHO on MGX pre-treated SR vesicles. (C) SHO mediated time-dependent effect on SERCA1 activity impaired by palmitate. (D) Viability of INS-1E beta-cells in the presence of SHO. (E) Viability of INS-1E beta-cells in the presence of MGX and SHO (20 – 200 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and SHO (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
Figure 3.
Effect of [6]-Shogaol (SHO) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of SHO. (B) Effect of SHO on MGX pre-treated SR vesicles. (C) SHO mediated time-dependent effect on SERCA1 activity impaired by palmitate. (D) Viability of INS-1E beta-cells in the presence of SHO. (E) Viability of INS-1E beta-cells in the presence of MGX and SHO (20 – 200 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and SHO (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
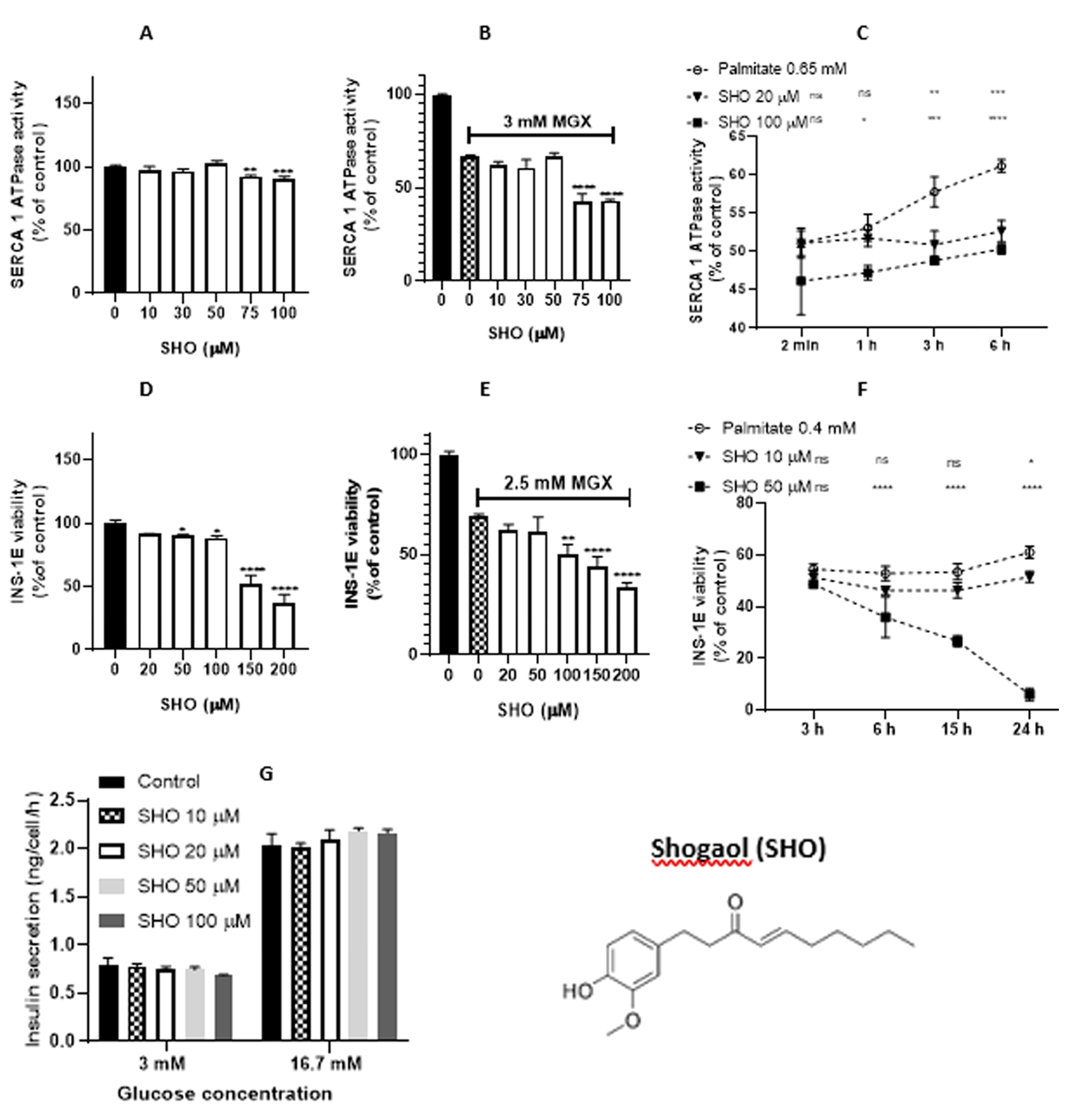
Figure 4.
Effect of resveratrol (RES) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of RES. (B) Effect of RES on MGX pre-treated SR vesicles. (C) RES mediated time-dependent effect on SERCA1 activity impaired by palmitate. (D) Viability of INS-1E beta-cells in the presence of RES. (E) Viability of INS-1E beta-cells in the presence of MGX and RES (10 – 150 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and RES (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
Figure 4.
Effect of resveratrol (RES) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of RES. (B) Effect of RES on MGX pre-treated SR vesicles. (C) RES mediated time-dependent effect on SERCA1 activity impaired by palmitate. (D) Viability of INS-1E beta-cells in the presence of RES. (E) Viability of INS-1E beta-cells in the presence of MGX and RES (10 – 150 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and RES (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
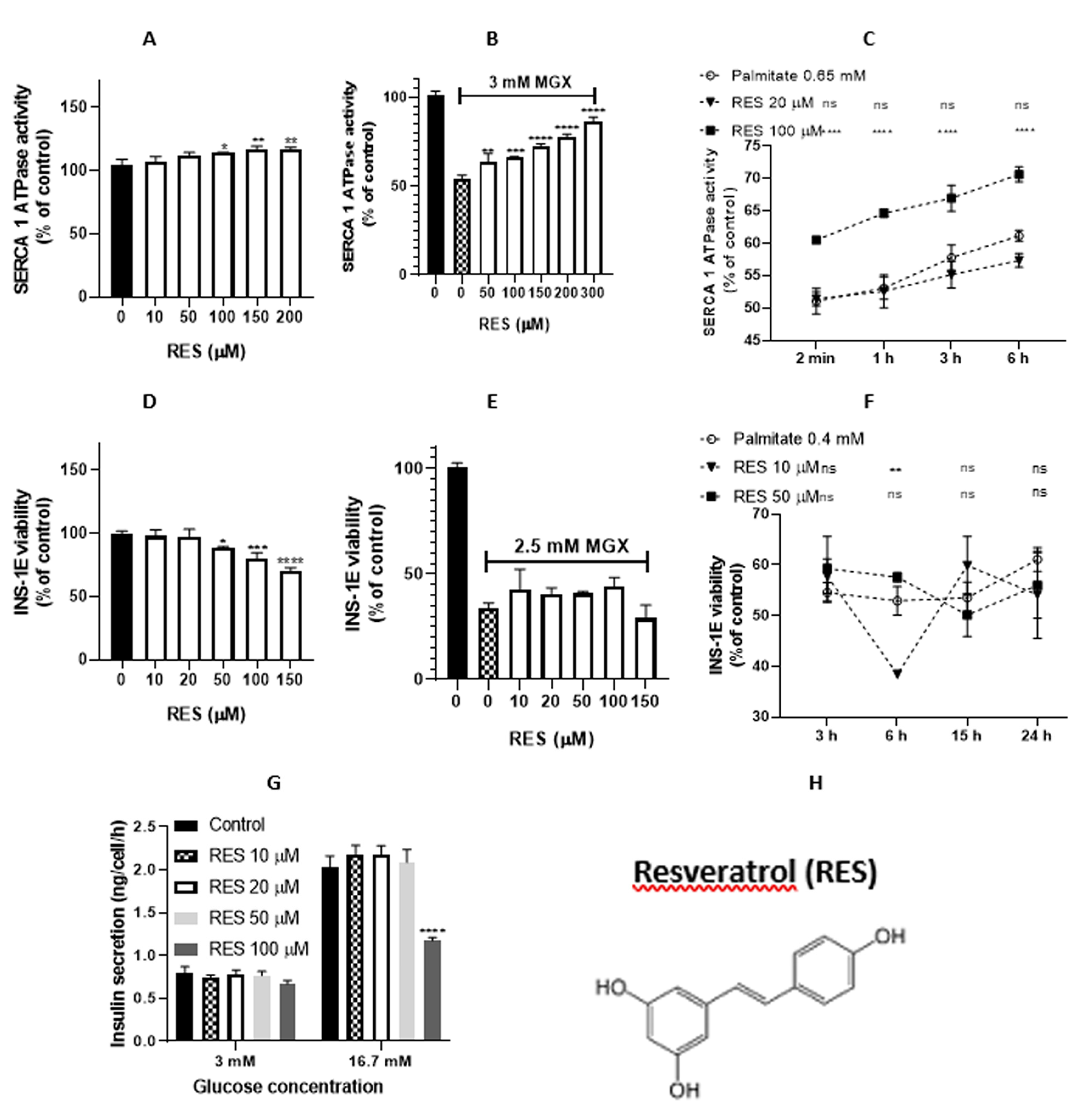
Figure 5.
Effect of oxyresveratrol (ORES) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of ORES. (B) Effect of ORES on MGX pre-treated SR vesicles. (C) ORES mediated time-dependent effect on SERCA1 activity impaired by palmitate. ns - nonsignificant changes. (D) Viability of INS-1E beta-cells in the presence of ORES. (E) Viability of INS-1E beta-cells in the presence of MGX and OXYRES (10–150 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and ORES (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
Figure 5.
Effect of oxyresveratrol (ORES) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of ORES. (B) Effect of ORES on MGX pre-treated SR vesicles. (C) ORES mediated time-dependent effect on SERCA1 activity impaired by palmitate. ns - nonsignificant changes. (D) Viability of INS-1E beta-cells in the presence of ORES. (E) Viability of INS-1E beta-cells in the presence of MGX and OXYRES (10–150 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and ORES (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
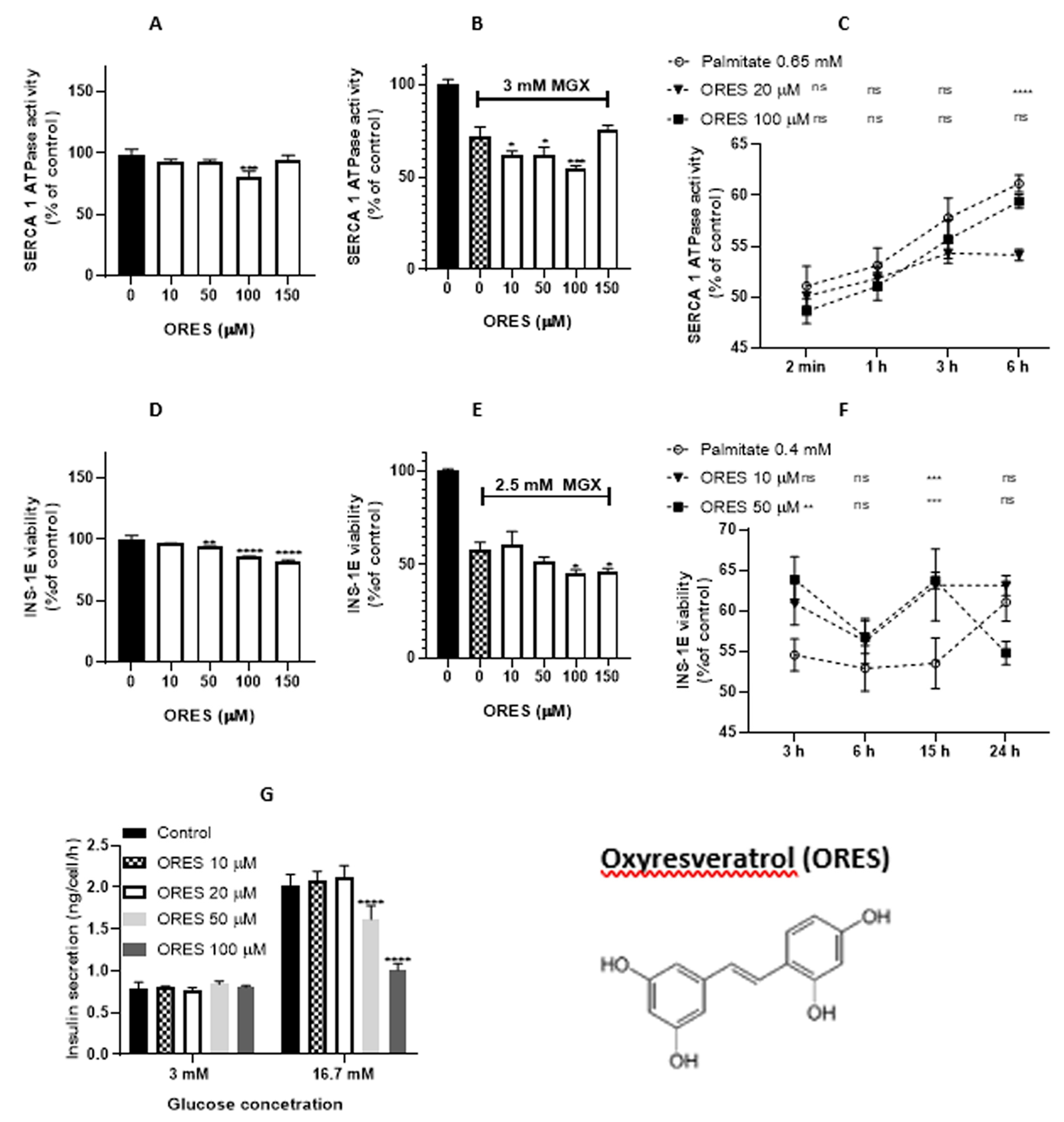
Figure 6.
Effect of ellagic acid (EA) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of EA. (B) Effect of EA on MGX pre-treated SR vesicles. (C) EA mediated time-dependent effect on SERCA1 activity impaired by palmitate. (D) Viability of INS-1E beta-cells in the presence of EA. (E) Viability of INS-1E beta-cells in the presence of MGX and EA (10 – 100 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and EA (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
Figure 6.
Effect of ellagic acid (EA) on sarcoplasmic reticulum (SR) vesicles and INS-1E pancreatic beta-cells. (A) Concentration-dependent modulation of SERCA1 activity in the presence of EA. (B) Effect of EA on MGX pre-treated SR vesicles. (C) EA mediated time-dependent effect on SERCA1 activity impaired by palmitate. (D) Viability of INS-1E beta-cells in the presence of EA. (E) Viability of INS-1E beta-cells in the presence of MGX and EA (10 – 100 μM). (F) Viability of INS-1E beta-cells in the presence of palmitate and EA (10 and 50 μM). (G) Insulin release in INS-1E beta-cells. Statistical significances between the compared groups in this figure are indicated in Material and Methods.
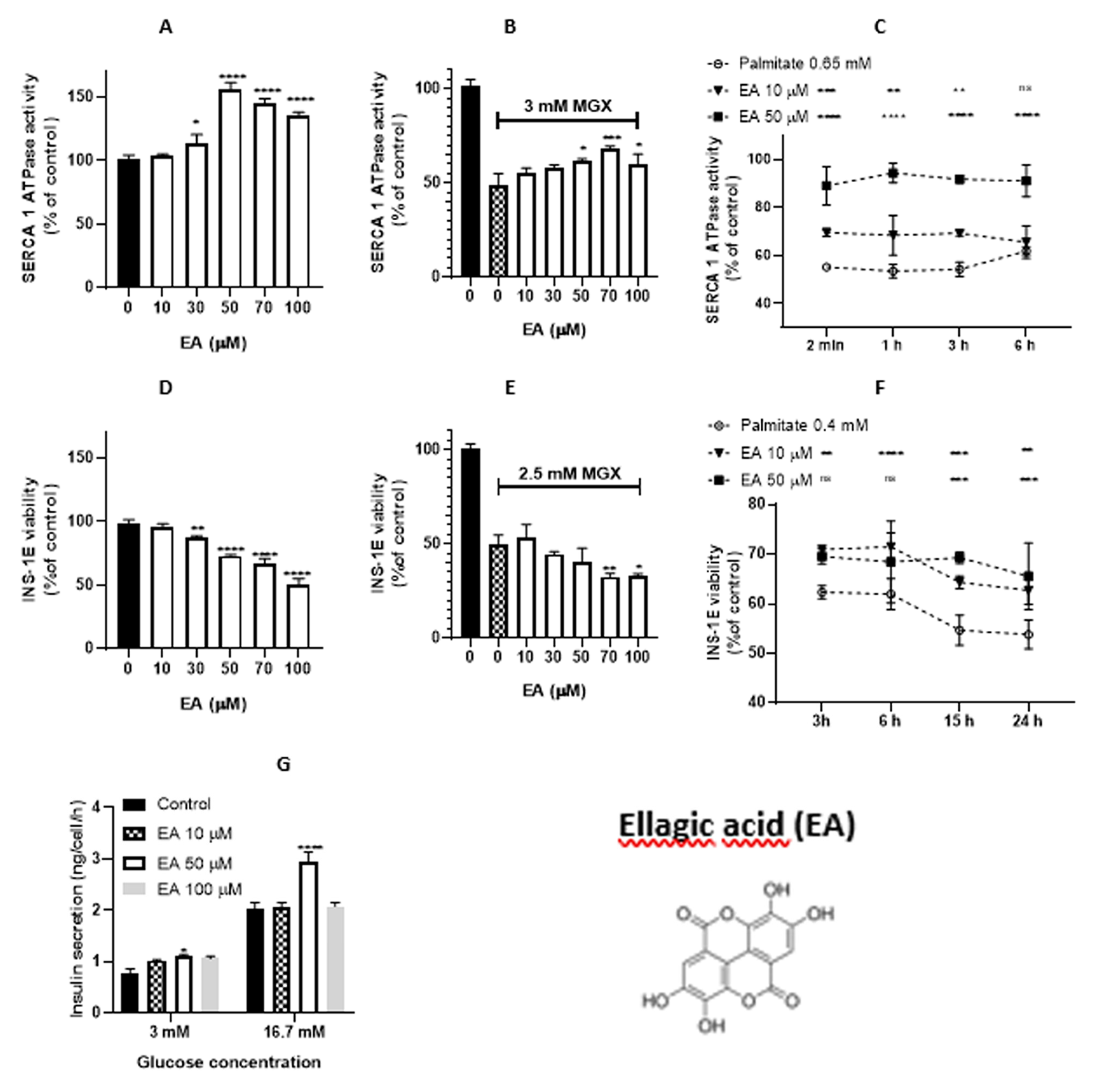
Figure 7.
In silico study of the interaction between phenolic compounds and SERCA (E2P model, pdb code 2zbe). Positions of [6]-gingerol (A), resveratrol (B) and ellagic acid (C), bound in SERCA (E2P model, pdb code 2zbe). (D) The global position of ellagic acid (magenta) bound in the cytoplasmic part of SERCA (E2P model, pdb code 2zbe).
Figure 7.
In silico study of the interaction between phenolic compounds and SERCA (E2P model, pdb code 2zbe). Positions of [6]-gingerol (A), resveratrol (B) and ellagic acid (C), bound in SERCA (E2P model, pdb code 2zbe). (D) The global position of ellagic acid (magenta) bound in the cytoplasmic part of SERCA (E2P model, pdb code 2zbe).
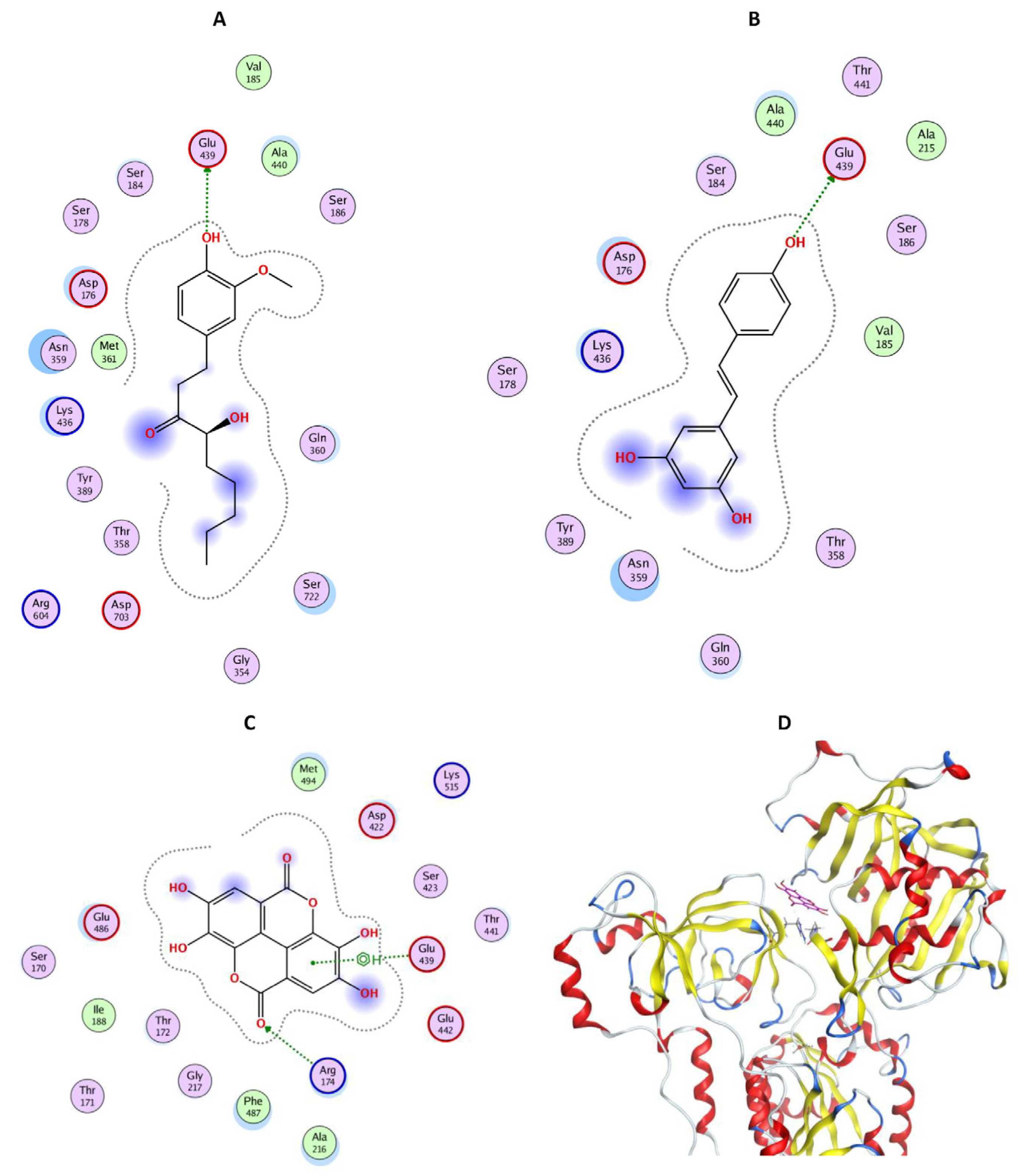

Table 1.
Values of the highest occupied molecular orbital (EHOMO) and the lowest unoccupied molecular orbital (ELUMO) obtained by Spartan software, together with the predicted values (MOE prediction) of the distribution coefficient (h logD, pH = 7.4) and the partition coefficient (h logP) for the compounds studied.
Table 1.
Values of the highest occupied molecular orbital (EHOMO) and the lowest unoccupied molecular orbital (ELUMO) obtained by Spartan software, together with the predicted values (MOE prediction) of the distribution coefficient (h logD, pH = 7.4) and the partition coefficient (h logP) for the compounds studied.
| compound | EHOMO | ELUMO | h_logD | h_logP |
|---|---|---|---|---|
| (kJ/mol) | (kJ/mol) | |||
| gingerol | -542.2 | -78.4 | 3.14 | 3.14 |
| shogaol | -530.4 | -38.1 | 4.66 | 4.66 |
| resveratrol | -507.4 | -115.2 | 2.76 | 2.75 |
| oxyresveratrol | -516.9 | -119.1 | 2.31 | 2.28 |
| ellagic acid | -581.3 | -196.4 | 1.95 | 1.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



