
Submitted:
09 August 2023
Posted:
10 August 2023
You are already at the latest version
Abstract
Unnatural diet composition and frequent feeding regimes may play an aetiological role in the multiple diseases prevalent in captive cheetahs. This study investigated the responses of captive-born (hand-reared) cheetahs (n = 6) to a reduced feeding frequency schedule. During the 3-week treatment period, the study cheetahs were fed four once-daily meals per week and two daily rations six days a week, subsequently, during the control period of equal duration. Total weekly food intake was maintained throughout the study. Variations in body temperature (Tb), heart rate (HR), locomotor activity (LA), behaviour, faecal glucocorticoid metabolite concentration, and faecal consistency score (FCS) were measured. Less frequent feeding resulted in higher FCS (p < 0.01) and LA (p < 0.0001) and lower HR (p < 0.0001) among the studied cheetahs. The study found that their HR was higher when the cheetahs were fed (p < 0.0001) than on days when they were not. Additionally, Tb (p < 0.0001) and HR (p < 0.05) were higher during feeding, suggesting a psychological excitation to food consumption. The results of the FCS analysis indicate that the more natural feeding pattern could have benefited the studied cheetahs’ GI health without a significant behavioural or physiological stress response to the change in feeding frequency.
Keywords:
cheetah
; captive diet
; wildlife husbandry
; gastrointestinal health
; stress
1. Introduction
Cheetahs (Acinonyx jubatus) in the wild typically prey on small to medium-sized antelope species with a body mass of between 23 and 56 kg [1] and, if left undisturbed, can consume a large proportion of the carcass [2]. They seldom eat daily or at a fixed interval but are opportunistic hunters [3]. However, captive cheetahs are routinely fed a non-varying diet of skinned muscle meat from livestock species, commercially prepared carnivore diets, carcass parts, or a combination [4,5] offered fixed intervals once or twice daily, with only one fast day per week. A whole carcass diet of the cheetah’s natural prey species and the associated feeding habits are challenging to accommodate in captivity [6]. Some facilities argue that frequent feeding allows for daily monitoring of the animals' appetites as an indicator of health and reduces boredom-induced stress [7]. However, unnatural diet composition and frequent feeding regimes may play an aetiological role in the prevalence of gastrointestinal (GI) and metabolic diseases in captive cheetahs [5,8].
One of these diseases, Helicobacter-associated gastritis, causes significant morbidity and mortality in captive cheetahs worldwide [9,10,11]. Helicobacter species, spiral bacteria colonising the stomach, infect most captive and wild cheetahs [12]. Captive cheetahs typically have some degree of inflammation (gastritis) that can be asymptomatic or associated with regurgitation, vomiting, the passage of undigested food, and weight loss [13,14]. However, in free-ranging cheetahs, there is colonisation by abundant spiral bacteria but little to no associated inflammation [10,15], demonstrating the likely multifactorial aetiopathogenesis of gastritis.
Diet, as a potential risk factor for GI pathology in captive cheetahs, was previously dismissed for the most part [10]. More recently, Whitehouse-Tedd et al. [5] found that feeding horsemeat has a significant (detrimental) relationship with gastritis risk in captive cheetahs. They attributed this to its high protein content [5] and/or digestibility [16] relative to other meat types fed to captive cheetahs. Crude protein is anticipated to be high in all carnivorous diets, including that of free-ranging cheetahs. However, the frequency with which it is fed in captivity and its quality could affect GI health by changing the amount of protein reaching the large intestine. Colonic fermentation of poorly digested dietary protein modifies microbiota composition in favour of proteolytic bacteria, some of which can be pathogenic in high concentrations [17,18] and produce putrefactive compounds (e.g., ammonia, indoles, phenols) associated with various disease states [19,20,21]. Moreover, horse (in particular) is commonly fed as muscle meat without low to non-digestible collagen-rich matter (e.g., bone, tendons, cartilage); therefore, its relative lack of ‘animal fibre’ may further increase putrefaction of digesta in the intestine [19,20,22,23].
Transforming gut bacteria-derived putrefactants into toxic metabolites negatively affects multiple other organ systems and metabolic pathways. Uraemic toxicity of indoxyl sulphate is associated with the progression of chronic renal failure [24], a significant cause of death in captive cheetahs [25,26,27,28]. The renal lesions in captive cheetahs resemble diabetic glomerulopathy in humans and chronic progressive nephropathy in rats [25]. High-protein diets, particularly when fed ad-libitum and continually, accelerate glomerulosclerosis in rats [29,30,31] and could be a comparable dietary risk factor for kidney damage in captive cheetahs.
A growing body of evidence suggests that feeding restrictions shape the gut ecosystem, function, and interaction with the host. Intermittent fasting has beneficial regulatory effects on immune homeostasis and intestinal microbiota composition in human and rodent models [32,33,34,35]. Furthermore, intermittent fasting attenuates the colon tissue inflammatory response and oxidative stress [32,36]. Following the adaption of captive lions (Panthera leo) from a conventional zoo feeding programme of predictable, fixed, small daily meals to a more natural gorge and fast feeding schedule of larger, more infrequent meals, Altman et al. [37] reported improved digestibility of a horsemeat-based diet. Considering similarities in the species’ natural feeding ecology, fasting conditions could have digestive health benefits for the cheetah as they did for the lion.
This study’s overall aim was to investigate the responses of captive-born (hand-reared) cheetahs to a reduced feeding frequency schedule distinguished by offering larger quantities of food less frequently. We hypothesised that a more natural feeding pattern would beneficially impact the GI ecosystem, including the microbial fermentation process. In previous studies, faecal consistency scoring has been used as a non-invasive method of measuring GI health in cheetahs, other exotic felids [5,38], and domestic carnivore species [39,40]. We also hypothesised that changing the feeding frequency may result in a behavioural and/or physiological stress response. Poor faecal consistency has been linked to GI stress in captive carnivores [38]. Behavioural observations [41] and faecal glucocorticoid metabolite (fGCM) analysis [42] were used as more established stress-related markers in the cheetah. In addition, we explored the use of biologging technology to record body temperature (Tb), heart rate (HR), and locomotor activity (LA) simultaneously. We predicted that higher fGCM concentrations, Tb, and HR would indicate a physiological stress response.
2. Materials and Methods
2.1. Study site and animals
The experimental trials took place between April and September 2019 at the Cango Wildlife Ranch and Conservation Centre (33°33’S, 22°12’E), 4 km north of Oudtshoorn, a semi-arid region in the Western Cape of South Africa. Study months (autumn and mainly winter) were distinguished by short photoperiods and cold air temperatures (Ta), ranging from 13 to 17 °C.
Three male (CH-2205, -2206, and -2271) and three female (CH-2207, -2276 and -2277) adult cheetahs (Table 1) habituated to human presence and interacting daily with the facility’s caretakers in the absence of restraint were assigned to this study. The study cheetahs were housed off-exhibit at the Jill Bryden-Fayers Reserve, neighbouring the Cango Wildlife Ranch. They were held in outdoor enclosures ranging from 400 to 1350 m2, adjoining conspecifics. The enclosures’ topography was varied and naturalistic, consisting of a dirt substrate, vantage points and marking areas (e.g., rocks, tree stumps), sufficient vegetation to hide, and a wooden shed for shelter. Enclosures were cleaned once or twice daily.
2.2. Body temperature, heart rate, and locomotor activity recordings
Each cheetah in the study had a single cardiac-, temperature- and movement-sensitive biologging unit (DST centi-HRT ACT, Star-Oddi, Gardabaer, Iceland) implanted, measuring 46 mm x 15 mm x 15 mm, and weighing approximately 19 g. The biologgers were calibrated against a high-accuracy thermometer (Hart 1504, Fluke, Utah, US) to within 0.1 °C. They were set to record tri-axial LA, i.e., heave, surge, sway, for the overall dynamic body acceleration (ODBA) [43] every minute, 24 hours a day, and Tb (°C) and leadless single-channel electrocardiogram (ECG)-derived HR (beats per minute [bpm]) at 200 Hz every 5 minutes. Representative traces of raw ECG recordings were saved every 24 hours for validating the HR measurements' quality index (QI) (where QI0 was the highest quality and QI1, QI2, and QI3 were progressively reduced).
2.3. Surgical procedures
Before surgical implantation, the biologgers were sterilised using ethylene oxide. Each study cheetah received a combination of 0.03 mg/kg of medetomidine (Medetomidine 20-mg/mL, Kyron Laboratories, Johannesburg, South Africa) and 0.8 mg/kg of tiletamine-zolazepam (Zoletil, Virbac Animal Health, Johannesburg, South Africa), administered intramuscularly (IM) by hand injection while in their enclosures. Once recumbent, the study cheetahs were placed in crates to which they were habituated and transported by vehicle (±500 m) to an onsite clinic at the Cango Wildlife Ranch. There, they were intubated with an endotracheal tube of appropriate size and maintained under anaesthesia with 2–3% isoflurane (Forane, Abbott, Weltevreden Park, South Africa), administered in 100% oxygen. The depth of anaesthesia and standard anaesthetic parameters were monitored throughout the procedure. Intravenous lactated Ringer’s solution (B Braun, Johannesburg, South Africa) was administered to maintain intraoperative normovolemia.
At the commencement of surgical procedures, 5 mL of blood was collected from the jugular vein of each study cheetah. Blood samples were divided between lithium heparin and EDTA tubes. Following this, a comprehensive haematological evaluation (using an onsite HM5 analyser; Abaxis Veterinary Diagnostics, California, US) and plasma biochemistry profile (using an onsite VetScan VS2 analyser; Abaxis Veterinary Diagnostics, California, US) were conducted to determine the health status of the study cheetahs before the onset of the field research. Urine samples were collected via a 6 FG dog urinary catheter and evaluated using a standard multiparameter dipstick and refractometer (Jorgensen Laboratories, Colorado, US).
The study cheetahs were placed in a left lateral recumbent position, and a thoracic incision site was shaved and sterilised with chlorhexidine gluconate (Hibitane, Zeneca, Johannesburg, South Africa). A 50 mm cranial-caudal incision was made in the skin, and a single biologging unit was implanted IM between the deep and superficial pectoralis major muscles without tethering. Wounds were closed with continuous subcutaneous and intradermal sutures (5/0 Monosyn, B-Braun, Barcelona, Spain) and treated with a topical antiseptic and ectoparasiticide spray (F10 Germicidal Wound Spray, Health and Hygiene Pty Ltd, Roodepoort, South Africa).
The study cheetahs were returned to the crates, where the effects of the anaesthesia were reversed with 0.1 mg/kg atipamezole (Antisedan, Zoetis, Johannesburg, South Africa) IM. Once recovered from the anaesthetic, they were transported back to their enclosures. The caretakers conducted daily observations to monitor for signs of surgical site infection or other postoperative complications.
Following the completion of the experimental trials, the same anaesthesia-surgical protocols were used to remove the biologgers.
2.4. Experimental design
The experimental trials commenced following a ≥12-day washout period after surgical implantation of the biologgers. In a pilot study conducted on the study cheetahs, Tb, HR, LA, faecal consistency score (FCS), and fGCM concentration data demonstrated temporal recovery by postoperative day 10 [44]; therefore, the authors felt ≥12 days after surgery to be sufficiently long to commence the experimental trials. They were conducted using a within-subject experimental design, where each cheetah in the study served as their own control. The study cheetahs received the treatment, i.e., a reduced feeding frequency schedule in an initial 3-week period followed by a control period of equal duration.
At the Cango Wildlife Ranch, cheetahs are fed a supplemented (Predator powder, V-Tech Pty Ltd, Midrand, South Africa, and Glycine, WildCat Nutrition Pty Ltd, Pretoria, South Africa) horsemeat-based diet prepared onsite. Meals are offered at variable intervals to prevent food anticipatory behavioural activity [45]. Once weekly at random, cheetahs are fed horse shank or rib bones with some meat intact equivalent to their daily ration in place of the day’s meals to maintain variety and provide periodontal stimulation. The bones are not consumed by the cheetahs and are removed and discarded. Regarding paired housing, the facility’s caretakers separate individuals to reduce competition for food. Leftover food is removed, weighed, recorded, and discarded. Water was available ad-libitum.
During the treatment period, the study cheetahs were fed on a reduced feeding frequency schedule, where meals were offered once daily between 0800 and 1700 hours, Monday, Tuesday, Thursday, and Friday (Figure S1). Weekly starve days were assigned to Wednesday, Saturday, and Sunday. Following the introduction of additional starve days, larger than regular meals were offered on feed days to maintain total weekly food intake. The three-year-old study cheetahs (CH-2205, -2206, and -2207) were fed 2.7 kg per day four days a week, and the two-year-old study cheetahs (CH-2271, -2276, and -27) were fed 2.5 kg per day four days a week. During the control period, the study cheetahs were fed on a feeding schedule routinely used at the Cango Wildlife Ranch, where meals were offered twice daily at 0800–1200 and 1500–1700 hours, Monday to Saturday. Weekly starve days are assigned to Sundays. The three-year-old study cheetahs were fed 1.8 kg per day portioned into two rations six days a week, and the two-year-old study cheetahs were fed 1.6 kg per day portioned into two daily rations six days a week.
There was a ≥1-day washout period between the treatment and control periods, during which the study cheetahs were fed on the routine feeding schedule. The duration of each period, i.e., treatment, control, and washout, was selected to accommodate the inverse relationship between the number of Tb, HR, and LA recordings made by the biologgers and the lifespan of their batteries.
Other than the specific intervention being investigated, i.e., a reduced feeding frequency schedule, the study cheetahs’ environment, housing, and management (refer to section 2.1. Study site and animals for more information) were maintained across the treatment and control, including bones offered randomly once weekly in place of the day’s meals.
Throughout the study, i.e., in the treatment and control, each cheetah was monitored regarding (i) behaviour; (ii) fGCM concentration; (iii) FCS; and (iv) Tb, HR, and LA simultaneously recorded by implanted biologgers.
2.5. Behavioural data collection
Each cheetah in the study was observed 15 times during the treatment and control periods, respectively. These observations were conducted between 0700 and 1700 hours, Monday to Sunday, within the operating hours of the Cango Wildlife Ranch. During five weekly 60-minute observation sessions, the principal investigator (KLB) carried out 12 instantaneous scan samples [46] with a 5-minute inter-scan interval per enclosure. Regarding paired housing (Table 1), the study cheetahs were observed together using physical identifiers to assign individual behaviour. Sampling was conducted on a variable day-and-time basis between enclosures, randomly selected to prevent time-of-day effects.
The study recorded 15 behaviours categorised as ‘inactive’, ‘active’, and ‘not observed’ (Table 2). The ethogram used for this study was informed by previous literature on felid behaviour (specifically cheetahs) [37,47,48,49] and initial observations of the cheetahs being studied. Time spent out of sight (hiding or staying away from the human observer) was noted as its performance has been linked to a psychological stress response in felids [50,51,52,53,54].
2.6. Faecal sample collection and consistency scoring
Regarding paired housing (Table 1), 1 tbsp of uncooked rice was thoroughly mixed into the diet of study cheetahs CH-2206 and CH-2277 once daily to assign individual faecal samples. The facility’s caretakers monitored the study cheetahs during feeding times to ensure sufficient rice consumption and prevent meal sharing. Only faeces found to have uncooked rice were considered to have been excreted by those individuals fed rice.
During the operating hours of the Cango Wildlife Ranch (0800–1700 hours), faeces were collected within one hour after defecation. Faeces excreted between 1700–0800 hours, when the study cheetahs’ enclosures could not be entered, were collected within 16 hours after defecation (4–10 °C Ta) [55].
Following sample collection, the principal investigator (KLB) assigned FCS by a five-point faecal scoring system adapted from that developed by Whitehouse-Tedd et al. [5]. In this study, the five-point faecal scoring system used (grades ranging from 1 to 5, where grade 1 was the lowest and grades 2–5 were progressively higher) included two points (grade 4: firm and dry and grade 5: firm) considered to be ‘normal,’ and three points (grades 1–3: liquid, soft without shape, and soft with shape) considered to be ‘suboptimal’ according to free-ranging cheetah scat. As a species inhabiting semi-arid regions (such as Oudtshoorn), a dry faecal consistency was not considered dissimilar to faeces found in free-ranging cheetahs [56].
Afterwards, the samples were deposited into appropriately labelled (sample collection date, study cheetah, and sample identification number) 50 mL polypropylene specimen containers and frozen at –20 °C.
2.7. Faecal steroid extraction and quantification
Following completion of the experimental trials, faecal samples were transported frozen to the Endocrine Research Laboratory, University of Pretoria, South Africa. Faecal steroids were extracted and subsequently analysed for fGCM concentration.
Frozen faecal samples were lyophilised, and the resultant dry faeces were pulverised and sieved through a mesh strainer to remove fibrous material [57]. Between 0.050–0.055 g of faecal powder was weighed per sample and extracted using 3 mL of 80% ethanol. The suspensions were vortexed for 15 minutes and centrifuged at 1500 x g for 10 minutes [58]. Supernatants were decanted into 1.5 mL safe-lock microcentrifuge tubes, labelled, and frozen at –20 °C until further analysis.
Immunoreactive fGCM concentrations were quantified using a corticosterone-3-CMO enzyme immunoassay (EIA) [55,59] according to procedures described by Ganswindt et al. [60]. Detailed assay characteristics, including full descriptions of the assay components and antibody cross-reactivities, are provided by Palme and Möstl [59]. The sensitivity of the EIA used at 90% binding was 3.6 ng/g faecal dry weight (DW). Inter-assay coefficients of variation (CV), determined by repeated measurements of low- and high-quality controls, were 11,74% and 12.91%, respectively, and intra-assay CV were 5.59% and 6.61%, respectively. Faecal steroid concentrations are presented as µg/g faecal DW.
2.8. Data preparation
2.8.1. Body temperature, heart rate, and locomotor activity recordings
Battery malfunction of the biologgers implanted in four of the six study cheetahs (CH-2207, -2271, -2276, and -2277) prevented those units from recording partial Tb, HR, and LA data for CH-2276 during the control period, and whole data sets for CH-2207, CH-2271, and CH-2277 (Table S1).
The initial examination of the raw HR data for the study cheetahs with functional biologgers (CH-2205 and -2206), and partial data for CH-2276, revealed values ranging from 0 to 1005 bpm, the extremes of which were likely due to incomplete, low-quality readings or implant movement within the pectoral muscle when the study cheetahs were active. To remove erroneous measurements, ensuring only plausible values were included in the analyses, upper and lower thresholds were created. At the low end of the range, all readings of 0 bpm were removed, establishing a new minimum of 30 bpm. For the upper end, where most unlikely measurements occurred, EGC data available in the literature on the cheetah [61,62] informed the HR values to be filtered. Those values to which QI3 was assigned (indicating the lowest quality) were also removed. Once filtered, all HR recordings (n = 30225/33443, which represented over 90% of the raw data initially captured) fell within the 30- and 200-bpm thresholds set. As Tb and LA recordings were not sensitive to implant movement within the pectoral muscle, they were not filtered, in addition to the lack of extreme Tb values.
To investigate the circadian patterns of Tb, HR, and LA, data were assigned to six part-of-the-day categories based on the hour they were recorded during the day, namely (i) early morning (0000–0400 hours), (ii) morning (0400–0800 hours), (iii) late morning (0800–1200 hours), (iv) afternoon (1200–1600 hours), (v) evening (1600–2000 hours), and (vi) night (2000–0000 hours).
2.8.2. Behavioural observations
The frequency with which each cheetah in the study performed each behaviour during each observation session was calculated as a proportion of the total number of scan samples carried out during that observation session per study cheetah [47,48]. The resulting data highlighted the proportion of scan samples in which each behaviour was observed during the treatment and control periods and on feed and starve days for each study cheetah.
2.8.3. Faecal consistency scores and glucocorticoid metabolite concentrations
Cheetahs typically defecate once daily, attenuating diurnal and pulsatory glucocorticoid (GC) secretion variations in the faeces [63]. However, differences in species and individual traits can affect hormone concentrations and GI transit time [64]. In a study by Terio et al. [42], peak concentrations of GC metabolites were found in the first faecal sample collected from cheetahs after administering adrenocorticotropic hormone. This was comparable to domestic cats' faecal cortisol excretion rate [65]. This study assumed FCS and fGCM concentrations to reflect the previous day’s intervention to account for cheetahs’ specific 24-hour gut passage rate and excretory pattern. The within-subject experimental design, where each cheetah served as their own control, eliminated interindividual variability [66].
2.9. Statistical analysis
Statistical analysis was performed using Microsoft Excel (version 16.0) and JMP Pro software (version 16.0) for Windows, developed by SAS Institute Inc (North Carolina, US). Body temperature, HR, LA, behaviour, FCS, and fGCM concentration data were explored for univariate outliers greater than three interquartile ranges (IQR) away from the 99.5th or 0.05th percentiles. Outlying values were detected for LA (n = 40) and subsequently excluded from descriptive statistics and analyses. Normal distribution and homogeneity of variance were explored using Anderson-Darling [67] and Levene’s tests [68], respectively. The variables measured were Box-Cox transformed [69] to satisfy the assumption of normality and homogeneity due to their departure. The data were back-transformed for descriptive statistics and visual representation to maintain statistical integrity. In this study, a mixed model for repeated measures (MMRM) analysis [70] was used to investigate the independent fixed effects of part of the day and the hour within part of the day on (i) Tb, (ii) HR, and (iii) LA. An MMRM analysis investigated the independent fixed effects of the study period and feed versus starve day on (i) Tb, (ii) HR, (iii) LA, (iv) behavioural, (v) FCS, and (vi) fGCM concentration, and their interaction to test the moderator effect of the study period on feed versus starve day. An MMRM analysis investigated the independent fixed effects of when the study cheetahs were feeding versus when they were not (i.e., feeding time [yes versus no]) and part of the day during which they were offered meals (late morning, afternoon, and evening) on (i) Tb, (ii) HR, and (iii) LA, and their interaction to test the moderator effect of part of the day on feeding time. The study cheetah was included as the random effect in the analyses.
Multiple pairwise comparisons were explored using Tukey’s honestly significant difference (HSD) post hoc tests [71]. Effect sizes of pairwise comparisons were calculated using the following formula:
where d = Cohen’s d effect size; x̅1 and x̅2 = means of the two groups; and SD1 and SD2 = standard deviation of the two groups [72]. Root mean square standardised effects (RMSSE) were interpreted as small (d = 0.2), medium (d = 0.5), and large (d = 0.8) based on Cohen’s d effect size criteria. Descriptive statistics were reported as median (IQR), and the significance level, alpha, was set at 0.05.
d = x̅1 − x̅2 / √SD12 + SD22 / 2,
3. Results
3.1. Body temperature, heart rate, and locomotor activity recordings
The MMRM analysis revealed that the circadian rhythm of Tb fluctuated significantly during the 24-hour day (for part of the day: F5,33417 = 2886.93, p < 0.0001 and for the hour nested within part of the day: F18,33417 = 171.90, p < 0.0001) (Figure 1i). Throughout the study, the overall median Tb was higher during the evening (38.0 [0.4] °C) between 1600–1659 hours (38.1 [0.2] °C) than the other parts of the day and hours.
The MMRM analysis revealed that the fixed effects of the study period and feed versus starve day on Tb failed to achieve statistical significance; therefore, post hoc testing was not performed (Figure S2).
The MMRM analysis revealed that Tb was significantly higher (F1,16698 = 30.41, p < 0.0001) when the study cheetahs were feeding (37.7 [0.5] °C) than when they were not (37.9 [0.3] °C). Post hoc comparisons using Tukey’s HSD test revealed that Tb was significantly higher at feeding time during the afternoon (yes: 38.0 [0.2] °C versus no: 37.9 [0.3] °C; t16697 = 3.26, p = 0.0141) and during the evening (yes: 38.2 [0.3] °C versus no: 38.0 [0.4] °C; t16697 = 4.53, p < 0.0001) (Figure 2).
Effect size calculations using Cohen’s d revealed a small RMSSE (d = 0.41; t16698 = 9.46, p < 0.0001) of feeding time on Tb. The RMSSE of feeding time on Tb by part of the day was medium during the afternoon (d = 0.54; t16698 = 6.22, p < 0.0001) and during the evening (d = 0.65; t16698 = 7.24, p < 0.0001) (Table 3).
The MMRM analysis revealed that the circadian rhythm of HR fluctuated significantly during the 24-hour day (for part of the day: F5,30199 = 300.96, p < 0.0001 and for the hour nested within part of the day: F18,30199 = 81.77, p < 0.0001) (Figure 1ii). Throughout the study, the overall median HR was higher during the morning (85 [55] bpm) between 0700–0759 hours (108 [39] bpm) than the other parts of the day and hours.
The MMRM analysis revealed that HR was significantly lower (F1,30210 = -56.85, p < 0.0001) during the treatment period (76 [39] bpm) than during the control period (78 [34] bpm). The MMRM analysis revealed that HR was significantly higher (F1,30221 = 170.83, p < 0.0001) on feed days (79 [35] bpm) than on starve days (69 [41] bpm). Post hoc comparisons using Tukey’s HSD test revealed that HR was significantly higher on treatment feed days (80 [37] bpm) than on treatment starve days (67 [44] bpm; t30219 = 17.94, p < 0.0001) and control starve days (76 [36] bpm; t30219 = 3.24, p = 0.0065) and significantly higher on control feed days (78 [33] bpm) than on treatment starve days (t30219 = 19.26, p < 0.0001) and control starve days (t30219 = 3.89, p = 0.0006) (Figure 3).
The MMRM analysis revealed that HR was significantly higher (F1,14832 = 6.56, p = 0.0104) when the study cheetahs were feeding (81 [39] bpm) than when they were not (80 [35] bpm). Post hoc comparisons using Tukey’s HSD test revealed that HR was significantly lower (t14833 = -3.69, p = 0.0031) at feeding time during the late morning (yes: 80 [44] bpm versus no: 84 [46] bpm) (Figure 4).
Effect size calculations using Cohen’s d revealed a small RMSSE of the study period (d = 0.11; t30221 = 7.11, p < 0.0001) and feed versus starve day (d = 0.20; t30221 = 13.26, p < 0.0001) on HR (Table 3). The RMSSE of feed versus starve day on HR by the study period was small between treatment feed days and treatment starve days (d = 0.30; t30221 = 18.05, p < 0.0001) and control starve days (d = 0.09; t30221 = 3.64, p = 0.0003) and between control feed days and treatment starve days (d = 0.31; t30221 = 18.97, p < 0.0001) and control starve days (d = 0.10; t30221 = 4.03, p < 0.0001). Effect size calculations using Cohen’s d revealed a small RMSSE of feeding time (d = 0.09; t14834 = 1.97, p = 0.0484) on HR. The RMSSE of feeding time on HR by part of the day was small during the late morning (d = 0.14; t14834 = 3.69, p = 0.0002).
The MMRM analysis revealed that the circadian rhythm of LA fluctuated significantly during the 24-hour day (for part of the day: F5,33377 = 723.70, p < 0.0001 and for the hour nested within part of the day: F18,33377 = 138.37, p < 0.0001) (Figure 1iii). Throughout the study, the overall median LA was higher during the morning (55.0 [106.4] ODBA) between 0700–0759 hours (117.0 [121.6] ODBA) than the other parts of the day and hours.
The MMRM analysis revealed that LA was significantly higher (F1,33397 = 115.50, p < 0.0001) during the treatment period (40.5 [52.0] ODBA) than during the control period (33.2 [43.6] ODBA). The MMRM analysis revealed that LA was significantly higher (F1,33397 = 10.79, p = 0.0010) on starve days (38.6 [49.6] ODBA) than on feed days (36.6 [47.6] ODBA). Post hoc comparisons using Tukey’s HSD test revealed that LA was significantly higher on treatment starve days (39.8 [55.3] ODBA) than on treatment feed days (40.8 [50.6] ODBA; t33397 = 5.99, p < 0.0001) and control feed days (33.0 [44.2] ODBA; t33397 = 13.18, p < 0.0001) (Figure 5).
The MMRM analysis revealed that LA was significantly higher (F1,16685 = 9.38, p = 0.0022) when the study cheetahs were feeding (52.6 [70.4] ODBA) than when they were not (39.8 [49.0] ODBA). Post hoc comparisons using Tukey’s HSD test revealed that the interaction effect of feeding time on LA by part of the day failed to achieve statistical significance during the late morning, the afternoon, and the evening (Figure S5).
Effect size calculations using Cohen’s d revealed a small RMSSE of the study period (d = 0.22; t33399 = 15.48, p < 0.0001) and feed versus starve day (d = 0.01; t33399 = 0.38, p = 0.7070) on LA (Table 3). The RMSSE of feed versus starve day on LA by the study period was small between treatment starve days and treatment feed days (d = 0.03; t33399 = 2.11, p = 0.0347) and control feed days (d = 0.22; t33399 = 14.11, p < 0.0001). Effect size calculations using Cohen’s d revealed a small RMSSE of feeding time (d = 0.33; t16686 = 7.74, p < 0.0001) on LA.
3.2. Behavioural observations
Three hundred and sixty scan samples per study cheetah were collected during the treatment (n = 180) and control periods (n = 180). The study cheetahs spent most of their time inactive (for the treatment: 40.37% and control periods: 43.72%, and for feed: 40.11% and starve days: 51.30%) (Table S3). The MMRM analysis revealed that the fixed effects of the study period and feed versus starve day on each behaviour failed to achieve statistical significance; therefore, post hoc testing was not performed.
3.3. Faecal consistency scores
Two hundred and thirteen faecal samples were collected from the study cheetahs during the treatment (n = 105) and control periods (n = 108). The soft with shape faecal grade was the most frequently recorded in the study cheetahs (for the treatment: 45.71% and the control periods: 48.15%, and for feed: 48.81% and starve days: 40.00%) (Table S4). The MMRM analysis revealed that FCS was significantly higher (F1,205.2 = 10.22, p = 0.0016) during the treatment period (3 [2]) than during the control period (3 [1]). The MMRM analysis revealed that the fixed effect of feed versus starve day on FCS failed to achieve statistical significance. Post hoc comparisons using Tukey’s HSD test revealed that FCS was significantly lower on control starve days (2.5 [1.25]) than on treatment feed days (3 [1.25]; t205.4 = -2.96, p = 0.0180), treatment starve days (4 [2]; t205.4 = -3.31, p = 0.0060), and control feed days (3 [2]; t205.4 = -2.83, p = 0.0265) (Figure 6).
Effect size calculations using Cohen’s d revealed a medium RMSSE (d = 0.66; t209 = 3.31, p = 0.0011) on FCS between the treatment and control periods (Table 3). The RMSSE of feed versus starve day on FCS by the study period was large between control starve days and treatment feed days (d = 1.07; t209 = 3.17, p = 0.0017), treatment starve days (d = 1.23; t209 = 3.43, p = 0.0007), and control feed days (d = 1.01; t209 = 3.04, p = 0.0027).
3.4. Faecal glucocorticoid metabolite concentrations
The MMRM analysis revealed that the fixed effects of the study period and feed versus starve day on fGCM concentration failed to achieve statistical significance; therefore, post hoc testing was not performed (Figure S6).
4. Discussion
This study aimed to investigate the responses of captive-born (hand-reared) cheetahs to a reduced feeding frequency. The results of the FCS analysis support, to some extent, the researchers’ hypothesis that the more natural feeding pattern would beneficially impact the GI ecosystem, including the microbial fermentation process. Overall, the findings indicate that the change in feeding frequency did not result in a significant behavioural or physiological stress response, contrary to what was predicted.
Animals’ GI tracts harbour essential gut microbes serving various functions [18]. However, disturbance-related deviation in the microbial diversity and abundance pattern beyond a natural range, i.e., gut dysbiosis, can advance pathophysiology and affect host health [73]. Considering the purported link with intestinal microbiota composition [74,75], higher FCS indicates that less frequent feeding could have benefited the studied cheetahs’ GI health. The data present here is consistent with Altman et al.’s [37] work concerning the impact of a random gorge and fast feeding schedule on the digestion of a horsemeat-based diet in captive lions.
Studies have shown that chronic or repeated exposure to stressors can disrupt gut homeostasis [76,77]; therefore, an alternative interpretation of this result may be the stress-reducing effects of less frequent feeding as a potential form of environmental enrichment (EE). In animal husbandry, the principle of EE is widely used to provide species-appropriate challenges to captive animals that lack adequate stimuli. This encourages them to engage actively with their environments, reducing stress and stereotypical behaviour [78,79]. One common type of enrichment is food-based, which is also applicable to the cheetah [41,47,48,80,81].
Absent hunting opportunities, offering carnivores predictable, fixed, and small daily meals can worsen their tendency to be inactive in captivity. This can result in obesity and affect their well-being [22]. In this study, the cheetahs spent most of their time inactive, consistent with previous research on captive cheetahs [81] and other felids [82,83,84,85,86,87,88]. The reduced feeding frequency schedule resulted in numerically lower inactivity and higher locomotion, and significantly higher LA. Increased activity has similarly been reported in captive lions following the adaption from a conventional zoo feeding programme to a randomised feeding schedule [37]. It is also possible to interpret the observed increase in activity among the studied cheetahs as behavioural agitation. Cheetahs and other carnivores in the wild spend a significant proportion of their activity budget resting [89]. While reducing the risk of obesity, the motivation for increased activity levels, possibly hunger, could have negative welfare implications. A lack of satiety is further supported by higher LA on treatment starve days (in particular). These results highlight the importance of offering sufficiently large meals if additional starve days are introduced not to be contrary to animals’ well-being.
Stress reduction by EE extends to sympathoadrenal responses. The two branches: the sympathetic nervous system (SNS) and the hypothalamic-pituitary-adrenal (HPA) axis, work together to maintain or re-establish homeostasis by orchestrating behavioural and physiological adaptations to the stressor [90]. The SNS provides the immediate first wave of the stress response, mediating the rapid release of the catecholamine hormones epinephrine (E) and norepinephrine from the adrenal medulla [91]. The second wave is more gradual and involves GCs, the end-product of the hormonal cascade along the HPA axis [92]. Environmental enrichment blunts E release [93] and, as a result, reduces stress-related increases in HR [94,95,96,97,98]. When the studied cheetahs were fed less frequently, they had a lower HR, indicating stress reduction [99] to a greater degree as it co-occurred with increased activity. During physical activity, requirements for oxygen and gluconeogenetic substrates in skeletal muscle are increased, as are the removal of metabolites and carbon dioxide [100]. The cardiac output is increased to meet the demand for blood flow by contracting muscles, attributed to sympathetically-mediated increases in HR and stroke volume. Increased Tb follows an increase in HR due to heat generated during nutrient conversion to muscular work.
Multiple hypotheses have been proposed to explain the stress-reducing effects of EE, some of which are based on the contention that EE itself acts as a mild stressor [101]. By providing challenges appropriate to an animal’s sensory, physical, and cognitive capacities [102], EE is thought to enable arousal and activation of the physiological stress response without pushing the animal into high, maladaptive stress levels [103,104]. In this manner, EE is adaptive, improving animals’ capacity to cope with stressors; that is, stress resilience [105,106,107].
Physiologically, enrichment-induced arousal was attended by higher HR (a salient feature of the SNS-mediated reaction to stress) on days when the studied cheetahs were fed. Additionally, higher Tb and HR were observed when they were feeding than when they were not. Stress-induced hyperthermia has been shown to some extent by a study in which the increase in Tb of free-ranging cheetahs following a successful hunt was double that of an unsuccessful chase (mean ± standard deviation [SD]; 1.38 ± 0.28 °C versus 0.58 ± 0.18 °C), despite comparable levels of physical activity [108,109]. The authors attributed the hyperthermia experienced by hunting cheetahs to the stress associated with vulnerability to attack and kleptoparasitism by more dominant intraguild predators [108,109]. In this study, the cheetahs were fed in a space free of interference competition; therefore, the thermal and cardiac response related to feeding suggests psychological excitation to food consumption rather than a predator-induced stress response. Alternatively, as higher Tb and HR at feeding time co-occurred with higher LA, these results could be interpreted as the demands of increased physical activity.
More work remains to be done before fully understanding the optimal feeding regime(s) to beneficially modulate cheetahs’ intestinal microbiota composition. This study has at least five potential limitations that should be considered. Firstly, faecal consistency scoring lacks specificity as a discrete measure of GI health and should not be interpreted as providing empirical evidence of diet suitability for maintaining cheetahs in captivity. Further research is needed to validate FCS against other measurements of microbiome health effects, such as digestibility, pH, the incidence of vomiting or diarrhoea, veterinary diagnosis of GI disease, fermentation by-products, faecal frequency, dry weight percentage, and short-chain fatty acids.
It must also be acknowledged that the present study was conducted on hand-reared cheetahs. In mammals, there is evidence that the high microbial diversity of infants’ gut communities may be inherited from their biological mothers [110]. Studies have shown that the rearing method (hand- versus mother-reared) can affect animal gut microbiota composition [111,112]. Therefore, the findings should be generalised to only some captive cheetahs.
Another significant limitation was the battery malfunction of the biologgers implanted in four of the six study cheetahs, preventing those units from recording partial Tb, HR, and LA data or whole data sets. As with any emerging technological advance, there is a need for further development, including more reliable and affordable devices, novel attachment or implantation and retrieval methods, and remote data transmission, before the routine application of biologgers in stress and animal welfare studies.
Thirdly, the experimental design of this study was constructed based on the inverse relationship between the number of Tb, HR, and LA recordings made by the biologgers and the lifespan of their batteries. The 3-week treatment and control periods may have needed longer to produce a definitive response in the studied cheetahs, limiting the conclusions that can be made from the results. For example, in mice, stress-related alterations in the composition and function of faecal microbiota were described after eight days, while they appeared after ten days in rats [113,114]. To quantify the treatment effects better, it would be recommendable for future research to lengthen the current study period.
A fourth potential limitation to consider is GI transit time. Measuring fGCM concentrations provides an integrated measure of adrenocortical activity. It reflects the cumulative secretion of plasma GCs over several hours (6–24 hours, depending on species-specific defecation rate), attenuating fluctuations due to secretory patterns [115,116,117,118]. However, dietary intake could affect the faecal excretion of steroid hormone metabolites independent of a stress response [119]. Due to accelerated GI transit time, larger quantities of food may decrease the accumulation time of faeces in the intestine and increase metabolite concentration variability [120].
By design, there were fewer starve days during the routine feeding schedule than during the reduced feeding frequency schedule. The days on which the studied cheetahs’ behaviour was observed were not adapted to maintain starve-day observation sessions equivalent between the treatment and control periods; therefore, it is possible that starve days were over- and under-represented during the treatment and control periods, respectively.
Finally, it must be acknowledged that the treatment effects on the measured variables, though at times statistically significant, were likely irrelevant to the biology of the cheetahs studied. The magnitude of the differences in numeric values for Tb, HR, and LA may not have been sufficient for biological significance. For example, Hetem et al. [108,109] reported a significant difference in Tb increase between successful and unsuccessful hunts (mean ± SD; 1.38 ± 0.28 °C versus 0.58 ± 0.18 °C; p < 0.0001), while the difference in Tb between when the study cheetahs were feeding and when they were not (median [IQR]; 37.7 [0.5] °C versus 37.9 [0.3] °C; p < 0.0001) significant here was comparatively less. This study’s reliance on only six study subjects, made worse by the battery malfunction of four of the six biologgers, is likely to have reduced our power to detect a biologically relevant effect. Cheetahs and other large carnivores are typically kept in low population densities at facilities like the Cango Wildlife Ranch. Future research should increase sample size through multi-institutional studies to improve the findings' quality, as Swaisgood and Shepherdson previously recommended [79].
5. Conclusions
Despite these limitations, the results of this research suggest practical and theoretical welfare implications for cheetahs in captivity. Though the validity of FCS as a measure of GI health must be established by further research, these data have provided sufficient support for a reduced feeding frequency schedule to mediate the unnatural composition of horsemeat-based diets routinely fed to captive cheetahs. While previous studies have mainly examined the epidemiological relationship between diet composition and GI disease [5,19,20,22,23], the findings presented here indicate that feeding regimes may also play a significant role. This study expands on existing research by Whitehouse-Tedd et al. [5] in developing a global standard by which captive facilities can score their cheetahs’ faecal consistency. We further demonstrated that biologging technology can be used to record simultaneous Tb, HR, and LA and, together with behavioural and physiological metrics, has tremendous potential to measure stress in the cheetah.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Biologger recordings for the study cheetahs during the treatment and control periods; Table S2. Root Mean Square Standardised Effects on body temperature (°C), heart rate (beats per minute [bpm]), locomotor activity (overall dynamic body acceleration [ODBA]), behaviour, faecal consistency score (grade), and faecal glucocorticoid metabolite concentration (µg/g dry weight [DW]) for the study cheetahs (CH-2205, -2206, -2207, -2271, -2276, and -2277). Effect sizes calculated using Cohen’s d; Table S3. The proportion of scan samples in which each behaviour was observed for the study cheetahs (CH-2205, -2206, -2207, -2271, - 2276, and -2277) during the treatment and control periods and on feed and starve days; Table S4. The proportion of faecal samples collected of each faecal consistency score for the study cheetahs (CH-2205, -2206, -2207, -2271, - 2276, and -2277) during the treatment and control periods and on feed and starve days; Figure S1. The experimental design used in the study; Figure S2. Box and whisker plot of body temperature (°C) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feed versus starve day by the study period; Figure S3. Box and whisker plot of body temperature (°C) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feeding time (yes [Y] versus no [N]) by part of the day (late morning); Figure S4. Box and whisker plot of heart rate (beats per minute [bpm]) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feeding time (yes [Y] versus no [N]) by part of the day (afternoon, evening); Figure S5. Box and whisker plot of locomotor activity (overall dynamic body acceleration [ODBA]) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feeding time (yes [Y] versus no [N]) by part of the day (late morning, afternoon, evening); Figure S6. Box and whisker plot of faecal glucocorticoid metabolite concentration (µg/g dry weight [DW]) for the study cheetahs (CH-2205, -2206, -2207, -2271, -2276, and -2277). Effect of feed versus starve day by the study period.
Author Contributions
Conceptualization, K.L.B. and A.S.W.T.; methodology, K.L.B. and A.S.W.T.; validation, K.L.B. and A.S.W.T.; formal analysis, K.L.B.; investigation, K.L.B., G.S., and A.S.W.T.; resources, A.G. and A.S.W.T.; data curation, K.L.B; writing—original draft preparation, K.L.B.; writing—review and editing, A.G., G.S., and A.S.W.T.; visualization, K.L.B.; supervision, A.G., G.S., and A.S.W.T.; project administration, K.L.B. and A.S.W.T.; funding acquisition, K.L.B. and A.S.W.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by THE DOCTORAL RESEARCH BURSARY of the UNIVERSITY OF PRETORIA, grant number SFH170622243855 and THE NATIONAL RESEARCH FOUNDATION, grant number MND200714543782. THE KEVIN RICHARDSON FOUNDATION provided additional financial support. The funders had no role in study design, data collection and analysis, the decision to publish, or the presentation of the manuscript.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Ethics Committee of the UNIVERSITY OF PRETORIA (protocol code V075-18, approved on August 28, 2019) and the Research Ethics Committee of the UNIVERSITY OF PRETORIA (protocol code REC069-18, approved on September 04, 2018). Surgical procedures were performed under medetomidine-tiletamine-zolazepam anaesthesia, and all efforts were made to minimise the study cheetahs’ suffering (refer to section 2.3. Surgical procedures for more information).
Data Availability Statement
The data presented in this study are available in supplementary material.
Acknowledgments
Narinda Beukes and the rest of the staff at the Cango Wildlife Ranch and Conservation Centre, Oudtshoorn, South Africa, are gratefully acknowledged for their cooperation during the experimental trials. Special thanks to Craig Gouws, Tamblyn Williams, and the facility’s caretakers for their support of the research and assistance during the sample and data collection. Dr Glen A. P. Carlisle is acknowledged for his tenured experience in the management and veterinary treatment of the study cheetahs. Dr Nicole Hagenah and the technicians at the Endocrine Research Laboratory, University of Pretoria, South Africa, are thanked for analysing steroid concentrations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hayward, M.W., Hofmeyr, M., O’Brien, J., Kerley, G.I.H. Prey preferences of the cheetah Acinonyx jubatus: morphological limitations or the need to captive rapidly consumable prey before kleptoparasites arrive? J. Zool. 2006, 270(4), 615-27. https://doi.org/10.1111/j.1469-7998.2006.00184.x. [CrossRef]
- Phillips, J.A. Bone Consumption by Cheetahs at Undisturbed Kills: Evidence for a Lack of Focal-Palatine Erosion. J. Mammal. 1993, 74(2), 487-92. https://doi.org/10.2307/1382408. [CrossRef]
- Mills, M.G.L., Harvey, M. African predators. Struik Publishers: Cape Town, South Africa, 2001.
- Tordiffe, A.S., Wachter, B., Heinrich, S.K., Reyers, F., Mienie, L.J. Comparative Serum Fatty Acid Profiles of Captive and Free-Ranging Cheetahs (Acinonyx jubatus) in Namibia. PLoS One 2016, 11(12), e0167608. https://doi.org/10.1371/journal.pone.0167608. Pubmed PMID: 27992457; Pubmed Central PMCID: PMC5167222. [CrossRef]
- Whitehouse-Tedd, K.M., Lefebvre, S.L., Janssens, G.P. Dietary factors associated with faecal consistency and other indicators of gastrointestinal health in the captive cheetah (Acinonyx jubatus). PLoS One 2015, 10(4), e0120903. https://doi.org/10.1371/journal.pone.0120903. Pubmed PMID: 25830636; Pubmed Central PMCID: PMC4382097. [CrossRef]
- Young, R.J. The importance of food presentation for animal welfare and conservation. Proc. Nutr. Soc. 1997, 56(3), 1095-104. https://doi.org/10.1079/pns19970113. PMID: 9483674. [CrossRef] [PubMed]
- Tordiffe, A.S.W. (University of Pretoria, Pretoria, Gauteng, South Africa). Personal communication, 2023a.
- Lane, E.P., Miller, S., Lobetti, R., Caldwell, P., Bertschinger, H.J., Burroughs, R., Kotze, A., van Dyk, A. Effect of diet on the incidence of and mortality owing to gastritis and renal disease in captive cheetahs (Acinonyx jubatus) in South Africa. Zoo Biol. 2012, 31(6), 669-82. https://doi.org/10.1002/zoo.20431. Epub 2011 Nov 14. Pubmed PMID: 22083933. [CrossRef] [PubMed]
- Kotsch, V., Kübber-Heiss, A., Url, A., Walzer, C., Schmidt, P. Diseases of captive cheetahs (Acinonyx jubatus) within the European endangered species program (EEP)—A 22-year retrospective histopathological study. Wien. Tierarztl. Monatsschr. 2002, 89(12), 341-50.
- Munson, L., Terio, K.A., Worley, M., Jago, M., Bagot-Smith, A., Marker, L. Extrinsic factors significantly affect patterns of disease in free-ranging and captive cheetah (Acinonyx jubatus) populations. J. Wildl. Dis. 2005, 41(3), 542-8. https://doi.org/10.7589/0090-3558-41.3.542. PMID: 16244064. [CrossRef] [PubMed]
- Une, Y., Uchida, C., Konishi, M., Ui, T., Kawakami, S., Ito, S., Nomura, Y. Pathological study of cheetahs (Acinonyx jubatus) dying in captivity in Japan. Vet. Pathol. 2001, 38, 585.
- Terio, K.A., Mitchell, E., Walzer, C., Schmidt-Küntzel, A., Marker, L., Citino, S. Diseases Impacting Captive and Free-Ranging Cheetahs. In: Biodiversity of the World—Cheetahs: Biology and Conservation, 1st ed.; Marker, L., Boast, L.K., Schmidt-Küntzel, A., Eds.; Elsevier: San Diego, California, 2018; pp. 349-60. https://doi.org/10.1016/B978-0-12-804088-1.00028-9. [CrossRef]
- Eaton, K.A., Radin, M.J., Kramer, L., Wack, R., Sherding, R., Krakowka, S., Fox, J.G., Morgan, D.R. Epizootic gastritis associated with gastric spiral bacilli in cheetahs (Acinonyx jubatus). Vet. Pathol. 1993, 30(1), 55-63. https://doi.org/10.1177/030098589303000107. Pubmed PMID: 8442328. [CrossRef] [PubMed]
- Terio, K.A., Munson, L., Moore, P.F. Characterization of the gastric immune response in cheetahs (Acinonyx jubatus) with Helicobacter-associated gastritis. Vet. Pathol. 2012, 49(5), 824-33. https://doi.org/10.1177/0300985811412620. Epub 2011 Jul 5. PMID: 21730348. [CrossRef] [PubMed]
- Terio, K.A., Munson, L., Marker, L., Aldridge, B.M., Solnick, J.V. Comparison of Helicobacter spp. in Cheetahs (Acinonyx jubatus) with and without gastritis. J. Clin. Microbiol. 2005, 43(1), 229-34. https://doi.org/10.1128/JCM.43.1.229-234.2005. Pubmed PMID: 15634976; Pubmed Central PMCID: PMC540127. [CrossRef]
- Vester, B.M., Beloshapka, A.N., Middelbos, I.S., Burke, S.L., Dikeman, C.L., Simmons, L.G., Swanson, K.S. Evaluation of nutrient digestibility and fecal characteristics of exotic felids fed horse- or beef-based diets: use of the domestic cat as a model for exotic felids. Zoo Biol. 2010, 29(4), 432-48. https://doi.org/10.1002/zoo.20275. Pubmed PMID: 19830746. [CrossRef] [PubMed]
- Becker, A.A.M.J., Hesta, M., Hollants, J., Janssens, G.P.J., Huys, G. Phylogenetic analysis of faecal microbiota from captive cheetahs reveals underrepresentation of Bacteroidetes and Bifidobacteriaceae. BMC Microbiol. 2014, 14(1), 43. https://doi.org/10.1186/1471-2180-14-43. [CrossRef]
- Wasimuddin, Menke, S., Melzheimer, J., Thalwitzer, S., Heinrich, S., Wachter, B., Sommer, S. Gut microbiomes of free-ranging and captive Namibian cheetahs: Diversity, putative functions and occurrence of potential pathogens. Mol. Ecol. 2017, 26(20), 5515-27. https://doi.org/10.1111/mec.14278. Epub 2017 Sep 08. Pubmed PMID: 28782134. [CrossRef] [PubMed]
- Depauw, S., Bosch, G., Hesta, M., Whitehouse-Tedd, K., Hendriks, W.H., Kaandorp, J., Janssens, G.P.J. Fermentation of animal components in strict carnivores: a comparative study with cheetah fecal inoculum. J. Anim. Sci. 2012a, 90(8), 2540-8. https://doi.org/10.2527/jas.2011-4377. Epub 2012 Jan 27. Pubmed PMID: 22287677. [CrossRef] [PubMed]
- Depauw, S., Hesta, M., Whitehouse-Tedd, K., Vanhaecke, L., Verbrugghe, A., Janssens, G.P.J. Animal fibre: the forgotten nutrient in strict carnivores? First insights in the cheetah. J. Anim. Physiol. Anim. Nutr. (Berl.). 2013, 97(1), 146-54. https://doi.org/10.1111/j.1439-0396.2011.01252.x. Epub 2011 Nov 10. Pubmed PMID: 22074361. [CrossRef] [PubMed]
- Rochus, K., Bosch, G., Vanhaecke, L., van de Velde, H., Depauw, S., Xu, J., Fievez, V., van de Wiele, T., Hendriks, W.H., Janssens, G.P.J., et al. Incubation of selected fermentable fibres with feline faecal inoculum: correlations between in vitro fermentation characteristics and end products. Arch. Anim. Nutr. 2013, 67(5), 416-31. https://doi.org/10.1080/1745039X.2013.830519. Epub 2013 Aug 19. Pubmed PMID: 23952674. [CrossRef] [PubMed]
- Depauw, S., Hesta, M., Whitehouse-Tedd, K., Stagegaard, J., Buyse, J., Janssens, G.P.J. Blood values of adult captive cheetahs (Acinonyx jubatus) fed either supplemented beef or whole rabbit carcasses. Zoo Biol. 2012b, 31(6), 629-41. https://doi.org/10.1002/zoo.20427. Epub 2011 Nov 03. Pubmed PMID: 22052742. [CrossRef] [PubMed]
- Depauw, S., Heilmann, R.M., Whitehouse-Tedd, K., Hesta, M., Steiner, J.M., Suchodolski, J.S., Janssens, G.P.J. Effect of diet type on serum and faecal concentration of S100/calgranulins in the captive cheetah. J. Zoo Aquar. Res. 2014, 2(2), 33-8. https://doi.org/10.19227/jzar.v2i2.81. [CrossRef]
- Niwa, T. Uremic toxicity of indoxyl sulfate. Nagoya J. Med. Sci. 2010, 72(1-2), 1-11. Pubmed PMID: 20229698. [PubMed]
- Bolton, L.A., Munson, L. Glomerulosclerosis in captive cheetahs (Acinonyx jubatus). Vet. Pathol. 1999, 36(1), 14-22. https://doi.org/10.1354/vp.36-1-14. Pubmed PMID: 9921751. [CrossRef] [PubMed]
- Munson, L. Diseases of captive cheetahs (Acinonyx jubatus): Results of the cheetah research council pathology survey, 1989-1992. Zoo Biol. 1993, 12(1), 105-24. https://doi.org/10.1002/zoo.1430120110. [CrossRef]
- Munson, L., Nesbit, J.W., Meltzer, D.G., Colly, L.P., Bolton, L., Kriek, N.P. Diseases of captive cheetahs (Acinonyx jubatus jubatus) in South Africa: a 20-year retrospective survey. J. Zoo Wildl. Med. 1999, 30(3), 342-7. Pubmed PMID: 10572855. [PubMed]
- Papendick, R.E., Munson, L., O'Brien, T.D., Johnson, K.H. Systemic AA amyloidosis in captive cheetahs (Acinonyx jubatus). Vet. Pathol. 1997, 34(6), 549-56. https://doi.org/10.1177/030098589703400602. Pubmed PMID: 9396135. [CrossRef] [PubMed]
- Bertani, T., Zoja, C., Abbate, M., Rossini, M., Remuzzi, G. Age-related nephropathy and proteinuria in rats with intact kidneys exposed to diets with different protein content. Lab. Invest. 1989, 60(2), 196-204. Pubmed PMID: 2915514. [PubMed]
- Brenner, B.M., Meyer, T.W., Hostetter, T.H. Dietary protein intake and the progressive nature of kidney disease: the role of hemodynamically mediated glomerular injury in the pathogenesis of progressive glomerular sclerosis in aging, renal ablation, and intrinsic renal disease. N. Engl. J. Med. 1982, 307(11), 652-9. https://doi.org/10.1056/NEJM198209093071104. Pubmed PMID: 7050706. [CrossRef] [PubMed]
- Olson, J.L., Heptinstall, R.H. Nonimmunologic mechanisms of glomerular injury. Lab. Invest. 1988, 59(5), 564-78. Pubmed PMID: 3054312. [PubMed]
- Li, L., Su, Y., Li, F., Wang, Y., Ma, Z., Li, Z., Su, J. The effects of daily fasting hours on shaping gut microbiota in mice. BMC Microbiol. 2020, 20(1), 65. https://doi.org/10.1186/s12866-020-01754-2. Pubmed PMID: 32209070; Pubmed Central PMCID: PMC7092480. [CrossRef]
- Li, G., Xie, C., Lu, S., Nichols, R.G., Tian, Y., Li, L., Patel, D., Ma, Y., Brocker, C.N., Yan, T., et al. Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota. Cell. Metab. 2017, 26(4), 672-85.e4. https://doi.org/10.1016/j.cmet.2017.08.019. Epub 2017 Sep 14. Erratum in: Cell Metab. 2017 Nov 07;26(5):801. Pubmed PMID: 28918936; Pubmed Central PMCID: PMC5668683. [CrossRef]
- Maifeld, A., Bartolomaeus, H., Löber, U., Avery, E.G., Steckhan, N., Markó, L., Wilck, N., Hamad, I., Šušnjar, U., Mähler, A., et al. Fasting alters the gut microbiome reducing blood pressure and body weight in metabolic syndrome patients. Nat. Commun. 2021, 12(1):1970. https://doi.org/10.1038/s41467-021-22097-0. Pubmed PMID: 33785752; Pubmed Central PMCID: PMC8010079. [CrossRef]
- Patterson, R.E., Sears, D.D. Metabolic Effects of Intermittent Fasting. Annu. Rev. Nutr. 2017, 37, 371-93. https://doi.org/10.1146/annurev-nutr-071816-064634. Epub 2017 Jul 17. Pubmed PMID: 28715993. [CrossRef] [PubMed]
- Zhang, X., Zou, Q., Zhao, B., Zhang, J., Zhao, W., Li, Y., Liu, R., Liu, X., Liu, Z. Effects of alternate-day fasting, time-restricted fasting and intermittent energy restriction DSS-induced on colitis and behavioral disorders. Redox Biol. 2020, 32, 101535. https://doi.org/10.1016/j.redox.2020.101535. Epub 2020 Apr 10. Erratum in: Redox Biol. 2021 Aug;44:101955. Pubmed PMID: 32305005; Pubmed Central PMCID: PMC7162980. [CrossRef]
- Altman, J.D., Gross, K.L., Lowry, S.R. Nutritional and behavioral effects of gorge and fast feeding in captive lions. J. Appl. Anim. Welf. Sci. 2005, 8(1), 47-57. https://doi.org/10.1207/s15327604jaws0801_4. Pubmed PMID: 16004544. [CrossRef] [PubMed]
- Vester, B.M., Burke, S.L., Dikeman, C.L., Simmons, L.G., Swanson, K.S. Nutrient digestibility and fecal characteristics are different among captive exotic felids fed a beef-based raw diet. Zoo Biol. 2008, 27(2), 126-36. https://doi.org/10.1002/zoo.20172. PMID: 19360610. [CrossRef] [PubMed]
- 39. Nery, J., Biourge, V., Tournier, C., Leray, V., Martin, L., Dumon, H, Nguyen, P. Influence of dietary protein content and source on fecal quality, electrolyte concentrations, and osmolarity, and digestibility in dogs differing in body size. J. Anim. Sci. 2010, 88(1), 159-69. https://doi.org/10.2527/jas.2008-1666. Epub 2009 Oct 23. PMID: 19854997. [CrossRef] [PubMed]
- 40. Nery, J., Goudez, R., Biourge, V., Tournier, C., Leray, V., Martin, L., Thorin, C., Nguyen, P., Dumon, H. Influence of dietary protein content and source on colonic fermentative activity in dogs differing in body size and digestive tolerance. J. Anim. Sci. 2012, 90(8), 2570-80. https://doi.org/10.2527/jas.2011-4112. Epub 2012 Feb 10. PMID: 22328724. [CrossRef] [PubMed]
- Quirke, T., O’Riordan, R.M., Zuur, A. Factors influencing the prevalence of stereotypical behaviour in captive cheetahs (Acinonyx jubatus). Appl. Anim. Behav. Sci. 2012, 142(3-4), 189-97. https://doi.org/10.1016/j.applanim.2012.09.007. [CrossRef]
- Terio, K.A., Citino, S.B., Brown, J.L. Fecal cortisol metabolite analysis for noninvasive monitoring of adrenocortical function in the cheetah (Acinonyx jubatus). J. Zoo Wildl. Med. 1999, 30(4), 484-91. Pubmed PMID: 10749432. [PubMed]
- Wilson, R.P., Börger, L., Holton, M.D., Scantlebury, D.M., Gómez-Laich, A., Quintana, F., Rosell, F., Graf, P.M., Williams, H., Gunner, R., et al. Estimates for energy expenditure in free-living animals using acceleration proxies: A reappraisal. J. Anim. Ecol. 2020, 89(1), 161-72. https://doi.org/10.1111/1365-2656.13040. Epub 2019 Jun 27. Pubmed PMID: 31173339; Pubmed Central PMCID: PMC7030956. [CrossRef]
- Brown, K.L. Responses to potential captivity-induced stressors in captive-born cheetah (Acinonyx jubatus): implications for behavioural and physiological stress and gastrointestinal health. Doctoral thesis, University of Pretoria, Pretoria, January 2023.
- Mistlberger, R.E. Circadian food-anticipatory activity: formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 18(2), 171-95. https://doi.org/10.1016/0149-7634(94)90023-x. PMID: 8058212. [CrossRef] [PubMed]
- Altmann, J. Observational study of behavior: sampling methods. Behaviour 1974, 49(3), 227-67. https://doi.org/10.1163/156853974x00534. Pubmed PMID: 4597405. [CrossRef] [PubMed]
- Quirke, T., O’Riordan, R.M. The effect of different types of enrichment on the behaviour of cheetahs (Acinonyx jubatus) in captivity. Appl. Anim. Behav. Sci. 2011a, 133(1), 87-94. https://doi.org/10.1016/j.applanim.2011.05.004. [CrossRef]
- Quirke, T., O’Riordan, R.M. The effect of a randomised enrichment treatment schedule on the behaviour of cheetahs (Acinonyx jubatus). Appl. Anim. Behav. Sci. 2011b, 135(1-2), 103-9. https://doi.org/10.1016/j.applanim.2011.10.006. [CrossRef]
- Regaiolli, B., Rizzo, A., Ottolini, G., Miletto Petrazzini, M.E., Spiezio, C., Agrillo, C. Motion Illusions as Environmental Enrichment for Zoo Animals: A Preliminary Investigation on Lions (Panthera leo). Front. Psychol. 2019, 10, 2220. https://doi.org/10.3389/fpsyg.2019.02220. Pubmed PMID: 31636583; Pubmed Central PMCID: PMC6788361. [CrossRef]
- Carlstead, K., Brown, J.L., Strawn, W. Behavioral and physiological correlates of stress in laboratory cats. Appl. Anim. Behav. Sci. 1993, 38(2), 143-58. https://doi.org/10.1016/0168-1591(93)90062-T. [CrossRef]
- Davey, G. Relationships between exhibit naturalism, animal visibility and visitor interest in a Chinese Zoo. Appl. Anim. Behav. Sci. 2006, 96(1), 93-102. https://doi.org/10.1016/j.applanim.2005.04.018. [CrossRef]
- Hosey, G. Hediger revisited: how do zoo animals see us? J. Appl. Anim. Welf. Sci. 2013, 16(4), 338-59. https://doi.org/10.1080/10888705.2013.827916. Pubmed PMID: 24079488. [CrossRef] [PubMed]
- Morgan, K.N., Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102(3-4), 262-302. https://doi.org/10.1016/j.applanim.2006.05.032. [CrossRef]
- Sellinger, R.L., Ha, J.C. The effects of visitor density and intensity on the behavior of two captive jaguars (Panthera onca). J. Appl. Anim. Welf. Sci. 2005, 8(4), 233-44. https://doi.org/10.1207/s15327604jaws0804_1. Pubmed PMID: 16436028. [CrossRef] [PubMed]
- Ludwig, C., Wachter, B., Silinski-Mehr, S., Ganswindt, A., Bertschinger, H., Hofer, H., Dehnhard, M. Characterisation and validation of an enzyme-immunoassay for the non-invasive assessment of faecal glucocorticoid metabolites in cheetahs (Acinonyx jubatus). Gen. Comp. Endocrinol. 2013, 180, 15-23. https://doi.org/10.1016/j.ygcen.2012.10.005. Epub 2012 Oct 26. Pubmed PMID: 23108105. [CrossRef] [PubMed]
- Tordiffe, A.S.W. (University of Pretoria, Pretoria, Gauteng, South Africa). Personal communication, 2023b.
- Fieß, M., Heistermann, M., Hodges, J.K. Patterns of urinary and fecal steroid excretion during the ovarian cycle and pregnancy in the African elephant (Loxodonta africana). Gen. Comp. Endocrinol. 1999, 115(1), 76-89. https://doi.org/10.1006/gcen.1999.7287. Pubmed PMID: 10375466. [CrossRef] [PubMed]
- Ganswindt, A., Muenscher, S., Henley, M., Henley, S., Heistermann, M., Palme, R., Thompson, P., Bertschinger, H. Endocrine correlates of musth and the impact of ecological and social factors in free-ranging African elephants (Loxodonta africana). Horm. Behav. 2010, 57(4-5), 506-14. https://doi.org/10.1016/j.yhbeh.2010.02.009. Epub 2010 Feb 24. Pubmed PMID: 20188104. [CrossRef] [PubMed]
- Palme, R., Möstl, E. Measurement of cortisol metabolites in faeces of sheep as a parameter of cortisol concentration in blood. Int. J. Mammal Biol. 1997, 62 Suppl II, 192-7.
- Ganswindt, A., Heistermann, M., Borragan, S., Hodges, J.K. Assessment of testicular endocrine function in captive African elephants by measurement of urinary and fecal androgens. Zoo Biol. 2002, 21, 27-36. https://doi.org/10.1002/zoo.10034. [CrossRef]
- Button, C., Meltzer, D.G., Mülders, M.S. The electrocardiogram of the cheetah (Acinonyx jubatus). J. S. Afr. Vet. Assoc. 1981, 52(3), 233-5. Pubmed PMID: 7310794. [PubMed]
- Schumacher, J., Snyder, P., Citino, S.B., Bennett, R.A., Dvorak, L.D. Radiographic and electrocardiographic evaluation of cardiac morphology and function in captive cheetahs (Acinonyx jubatus). J. Zoo Wildl. Med. 2003, 34(4), 357-63. https://doi.org/10.1638/01-008. Pubmed PMID: 15077711. [CrossRef] [PubMed]
- Keay, J.M., Singh, J., Gaunt, M.C., Kaur, T. Fecal glucocorticoids and their metabolites as indicators of stress in various mammalian species: a literature review. J. Zoo Wildl. Med. 2006, 37(3), 234-44. https://doi.org/10.1638/05-050.1. Pubmed PMID: 17319120. [CrossRef] [PubMed]
- Palme, R. Measuring fecal steroids: guidelines for practical application. Ann. N. Y. Acad. Sci. 2005, 1046, 75-80. https://doi.org/10.1196/annals.1343.007. Pubmed PMID: 16055844. [CrossRef] [PubMed]
- Graham, L.H., Brown, J.L. Cortisol metabolism in the domestic cat and implications for non-invasive monitoring of adrenocortical function in endangered felids. Zoo Biol. 1996, 15(1), 71-82. https://doi.org/10.1002/(SICI)1098-2361(1996)15:1<71::AID-ZOO7>3.0.CO;2-9. [CrossRef]
- Maclure, M. The case-crossover design: a method for studying transient effects on the risk of acute events. Am. J. Epidemiol. 1991, 133(2), 144-53. https://doi.org/10.1093/oxfordjournals.aje.a115853. Pubmed PMID: 1985444. [CrossRef] [PubMed]
- Anderson, T.W., Darling, D.A. Asymptotic theory of certain “goodness-of-fit” criteria based on stochastic processes. Ann. Math. Stat. 1952, 23(2), 193-212. https://doi.org/10.1214/aoms/1177729437. [CrossRef]
- Levene, H. Robust Tests for Equality of Variances. In: Contributions to Probability and Statistics; Olkin, I., Ed.; Stanford University Press: Palo Alto, California, 1960; pp. 278-92.
- Box, G.E.P., Cox, D.R. An Analysis of Transformations. J. R. Stat. Soc. Ser. B Methodol. 1964, 26(2), 211-52. https://doi.org/10.111/j.2517-6161.1964.tb00553.x. [CrossRef]
- Fisher, R.A. The correlation between relatives on the supposition of Mendelian inheritance. Trans. R. Soc. Edinb. 1918, 53, 399-433.
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5(2), 99-114. https://doi.org/10.2307/3001913. [CrossRef]
- Cohen, J. In: Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates: Hillsdale, New Jersey, 1988. https://doi.org/10.4324/9780203771587. [CrossRef]
- Shreiner, A.B., Kao, J.Y., Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31(1), 69-75. https://doi.org/10.1097/MOG.0000000000000139. PMID: 25394236; PMCID: PMC4290017. [CrossRef]
- Tigchelaar, E.F., Bonder, M.J., Jankipersadsing, S.A., Fu, J., Wijmenga, C., Zhernakova, A. Gut microbiota composition associated with stool consistency. Gut 2016, 65(3), 540-2. https://doi.org/10.1136/gutjnl-2015-310328. Epub 2015 Aug 14. Pubmed PMID: 26276682. [CrossRef] [PubMed]
- Vandeputte, D., Falony, G., Vieira-Silva, S., Tito, R.Y., Joossens, M., Raes, J. Stool consistency is strongly associated with gut microbiota richness and composition, enterotypes and bacterial growth rates. Gut 2016, 65(1), 57-62. https://doi.org/10.1136/gutjnl-2015-309618. Epub 2015 Jun 11. Pubmed PMID: 26069274; Pubmed Central PMCID: PMC4717365. [CrossRef]
- Accarie, A., Vanuytsel, T. Animal Models for Functional Gastrointestinal Disorders. Front. Psychiatry 2020, 11, 11:509681. https://doi.org/10.3389/fpsyt.2020.509681. Pubmed PMID: 33262709; Pubmed Central PMCID: PMC7685985. [CrossRef]
- Farzi, A., Fröhlich, E.E., Holzer, P. Gut Microbiota and the Neuroendocrine System. Neurotherapeutics 2018, 15(1), 5-22. https://doi.org/10.1007/s13311-017-0600-5. Pubmed PMID: 29380303; Pubmed Central PMCID: PMC5794709. [CrossRef]
- Carlstead, K., Shepherdson, D. Alleviating stress in zoo animals with environmental enrichment. In: The Biology of Animal Stress - Basic Principles and Implications for Animal Welfare; Moberg, G.P., Mench, J.A., Eds.; CAB International: New York, 2000; pp. 337-54.
- Swaisgood, R.R., Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: what’s been done and where should we go next? Zoo Biol. 2005, 24(6), 499-518. https://doi.org/10.1002/zoo.20066. [CrossRef]
- Bond, J.C., Lindburg, D.G. Carcass feeding of captive cheetahs (Acinonyx jubatus): the effects of a naturalistic feeding program on oral health and psychological well-being. Appl. Anim. Behav. Sci. 1990, 26(4), 373-82. https://doi.org/10.1016/0168-1591(90)90036-D. [CrossRef]
- Skibiel, A.L., Trevino, H.S., Naugher, K. Comparison of several types of enrichment for captive felids. Zoo Biol. 2007, 26(5), 371-81. https://doi.org/10.1002/zoo.20147. Pubmed PMID: 19360587. [CrossRef] [PubMed]
- Acaralp-Rehnberg, L.K., Coleman, G.J., Magrath, M.J.L., Melfi, V., Fanson, K.V., Bland, I.M. The Effect of Behind-The-Scenes Encounters and Interactive Presentations on the Welfare of Captive Servals (Leptailurus serval). Animals (Basel) 2020, 10(4), 743. https://doi.org/10.3390/ani10040743. Pubmed PMID: 32344609; Pubmed Central PMCID: PMC7222754. [CrossRef]
- Bashaw, M.J., Kelling, A.S., Bloomsmith, M.A., Maple, T.L. Environmental effects on the behavior of zoo-housed lions and tigers, with a case study of the effects of a visual barrier on pacing. J. Appl. Anim. Welf. Sci. 2007, 10(2), 95-109. https://doi.org/10.1080/10888700701313116. Pubmed PMID: 17559318. [CrossRef] [PubMed]
- De Souza Resende, L., Lima e Neto, G., Gonçalves Duarte Carvalho, P., Landau-Remy, G., de Almeida Ramos-Júnior, V., Andriolo, A., Genaro, G. Time budget and activity patterns of oncilla cats (Leopardus tigrinus) in captivity. J. Appl. Anim. Welf. Sci. 2014, 17(1), 73-81. https://doi.org/10.1080/10888705.2014.856253. Pubmed PMID: 24484312. [CrossRef] [PubMed]
- Macri, A.M., Patterson-Kane, E. Behavioural analysis of solitary versus socially housed snow leopards (Panthera uncia), with the provision of simulated social contact. Appl. Anim. Behav. Sci. 2011, 130(3-4), 115-23. https://doi.org/10.1016/j.applanim.2010.12.005. [CrossRef]
- Moreira, N., Brown, J.L., Moraes, W., Swanson, W.F., Monteiro-Filho, E.L. Effect of housing and environmental enrichment on adrenocortical activity, behavior and reproductive cyclicity in the female tigrina (Leopardus tigrinus) and margay (Leopardus wiedii). Zoo Biol. 2007, 26(6), 441-60. https://doi.org/10.1002/zoo.20139. Pubmed PMID: 19360593. [CrossRef] [PubMed]
- Shepherdson, D.J., Carlstead, K., Mellen, J.D., Seiden-sticker, J. The influence of food presentation on the behavior of small cats in confined environments. Zoo Biol. 1993, 12(2), 203-16. https://doi.org/10.1002/zoo.1430120206. [CrossRef]
- Weller, S.H., Bennett, C.L. Twenty-four hour activity budgets and patterns of behavior in captive ocelots (Leopardus pardalis). Appl. Anim. Behav. Sci. 2001, 71(1), 67-79. https://doi.org/10.1016/s0168-1591(00)00169-6. Pubmed PMID: 11179560. [CrossRef] [PubMed]
- Siegel, J.M. Clues to the functions of mammalian sleep. Nature 2005, 437(7063), 1264-71. https://doi.org/10.1038/nature04285. Pubmed PMID: 16251951; Pubmed Central PMCID: PMC8760626. [CrossRef]
- Coffman, J. Chronic stress, physiological adaptation and developmental programming of the neuroendocrine stress system. Future Neurol. 2020, 15(8), FNL39. https://doi.org/10.2217/fnl-2019-0014. [CrossRef]
- Romero, L.M., Wingfield, J.C. Mediators of Stress. In: Tempests, Poxes, Predators, and People: Stress in Wild Animals and How They Cope, 1st edn; Oxford University Press: New York, 2015; pp. 23-68.
- Sapolsky, R.M., Romero, L.M., Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21(1), 55-89. https://doi.org/10.1210/edrv.21.1.0389. Pubmed PMID: 10696570. [CrossRef] [PubMed]
- Moncek, F., Duncko, R., Johansson, B.B., Jezova, D. Effect of environmental enrichment on stress related systems in rats. J. Neuroendocrinol. 2004, 16(5), 423-31. https://doi.org/10.1111/j.1365-2826.2004.01173.x. Pubmed PMID: 15117335. [CrossRef] [PubMed]
- Azar, T.A., Sharp, J.L., Lawson, D.M. Effects of cage enrichment on heart rate, blood pressure, and activity of female Sprague-Dawley and spontaneously hypertensive rats at rest and after acute challenges. J. Am. Assoc. Lab. Anim. Sci. 2012, 51(3), 339-44. Pubmed PMID: 22776192; Pubmed Central PMCID: PMC3358983.
- Marashi, V., Barnekow, A., Ossendorf, E., Sachser, N. Effects of different forms of environmental enrichment on behavioral, endocrinological, and immunological parameters in male mice. Horm. Behav. 2003, 43(2), 281-92. https://doi.org/10.1016/s0018-506x(03)00002-3. Pubmed PMID: 12694638. [CrossRef] [PubMed]
- Ravenelle, R., Santolucito, H.B., Byrnes, E.M., Byrnes, J.J., Donaldson, S.T. Housing environment modulates physiological and behavioral responses to anxiogenic stimuli in trait anxiety male rats. Neuroscience 2014, 270, 76-87. https://doi.org/10.1016/j.neuroscience.2014.03.060. Epub 2014 Apr 05. Pubmed PMID: 24713371; Pubmed Central PMCID: PMC4047719. [CrossRef]
- Sharp, J., Azar, T., Lawson, D. Effects of a complex housing environment on heart rate and blood pressure of rats at rest and after stressful challenges. J. Am. Assoc. Lab. Anim. Sci. 2014, 53(1), 52-60. Pubmed PMID: 24411780; Pubmed Central PMCID: PMC3894648.
- Sharp, J.L., Zammit, T.G., Azar, T.A., Lawson, D.M. Stress-like responses to common procedures in male rats housed alone or with other rats. Contemp. Top. Lab. Anim. Sci. 2002, 41(4), 8-14. Pubmed PMID: 12109891. [PubMed]
- Wascher, C.A.F. Heart rate as a measure of emotional arousal in evolutionary biology. Phil. Trans. R. Soc. 2021, B376, 20200479. https://doi.org/10.1098/rstb.2020.0479. [CrossRef]
- Burton, D.A., Stokes, K.A., Hall, G.M. Physiological effects of exercise. CEACCP 2004, 4(6), 185-8. https://doi.org/10.1093/bjaceaccp/mkh050. [CrossRef]
- Smail, M.A., Smith, B.L., Nawreen, N., Herman, J.P. Differential impact of stress and environmental enrichment on corticolimbic circuits. Pharmacol. Biochem. Behav. 2020, 197, 172993. https://doi.org/10.1016/j.pbb.2020.172993. Epub 2020 Jul 11. Pubmed PMID: 32659243; Pubmed Central PMCID: PMC7484282. [CrossRef]
- Meehan, C.L., Mench, J.A. The challenge of challenge: Can problem solving opportunities enhance animal welfare? Appl. Anim. Behav. Sci. 2007, 102(3-4), 246-61. https://doi.org/10.1016/j.applanim.2006.05.031. [CrossRef]
- Ashokan, A., Sivasubramanian, M., Mitra, R. Seeding Stress Resilience through Inoculation. Neural. Plast. 2016, 2016, 4928081. https://doi.org/10.1155/2016/4928081. Epub 2016 Jan 05. Pubmed PMID: 26881112; Pubmed Central PMCID: PMC4736400. [CrossRef]
- Liu, H., Zhang, C., Ji, Y., Yang, L. Biological and Psychological Perspectives of Resilience: Is It Possible to Improve Stress Resistance? Front. Hum. Neurosci. 2018, 12, 326. https://doi.org/10.3389/fnhum.2018.00326. Pubmed PMID: 30186127; Pubmed Central PMCID: PMC6110926. [CrossRef]
- Crofton, E.J., Zhang, Y., Green, T.A. Inoculation stress hypothesis of environmental enrichment. Neurosci. Biobehav. Rev. 2015, 49, 19-31. https://doi.org/10.1016/j.neubiorev.2014.11.017. Epub 2014 Nov 29. Pubmed PMID: 25449533; Pubmed Central PMCID: PMC4305384. [CrossRef]
- Lyons, D.M., Parker, K.J., Katz, M., Schatzberg, A.F. Developmental cascades linking stress inoculation, arousal regulation, and resilience. Front. Behav. Neurosci. 2009, 3, 32. https://doi.org/10.3389/neuro.08.032.2009. Pubmed PMID: 19826626; Pubmed Central PMCID: PMC2759374. [CrossRef]
- Parker, K.J., Maestripieri, D. Identifying key features of early stressful experiences that produce stress vulnerability and resilience in primates. Neurosci. Biobehav. Rev. 2011, 35(7), 1466-83. https://doi.org/10.1016/j.neubiorev.2010.09.003. Epub 2010 Sep 17. Pubmed PMID: 20851145; Pubmed Central PMCID: PMC3023826. [CrossRef]
- Hetem, R.S., Mitchell, D., De Witt, B.A., Fick, L.G., Maloney, S.K., Meyer, L.C.R., Fuller, A. Body temperature, activity patterns and hunting in free-living cheetah: biologging reveals new insights. Integr. Zool. 2019, 14(1), 30-47. https://doi.org/10.1111/1749-4877.12341. Pubmed PMID: 29851240. [CrossRef] [PubMed]
- Hetem, R.S., Mitchell, D., De Witt, B.A., Fick, L.G., Meyer, L.C.R., Maloney, S.K., Fuller, A. Cheetahs do not abandon hunts because they overheat. Biol. Lett. 2013, 9(5), 20130472. https://doi.org/10.1098/rsbl.2013.0472. [CrossRef]
- Sprockett, D., Fukami, T., Relman, D.A. Role of priority effects in the early-life assembly of the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2018, 15(4):197-205. https://doi.org/10.1038/nrgastro.2017.173. Epub 2018 Jan 24. PMID:29362469; PMCID: PMC6813786. [CrossRef]
- Li, X., Bi, R., Xiao, K., Roy, A., Zhang, Z., Chen, X., Peng, J., Wang, R., Yang, R., Shen, X., et al. Hen raising helps chicks establish gut microbiota in their early life and improve microbiota stability after H9N2 challenge. Microbiome 2022, 10(1), 14. https://doi.org/10.1186/s40168-021-01200-z. PMID: 35074015; PMCID: PMC8785444. [CrossRef]
- Chen, C.Y., Chen, C.K., Chen, Y.Y. et al. Maternal gut microbes shape the early-life assembly of gut microbiota in passerine chicks via nests. Microbiome 2020, 8, 129. https://doi.org/10.1186/s40168-020-00896-9. [CrossRef]
- Yoshikawa, K., Kurihara, C., Furuhashi, H., Takajo, T., Maruta, K., Yasutake, Y., Sato, H., Narimatsu, K., Okada, Y., Higashiyama, M., et al. Psychological stress exacerbates NSAID-induced small bowel injury by inducing changes in intestinal microbiota and permeability via glucocorticoid receptor signaling. J. Gastroenterol. 2017, 52(1), 61-71. https://doi.org/10.1007/s00535-016-1205-1. Epub 2016 Apr 13. Pubmed PMID: 27075753. [CrossRef] [PubMed]
- Zhang, J., Song, L., Wang, Y., Liu, C., Zhang, L., Zhu, S., Liu, S., Duan, L. Beneficial effect of butyrate-producing Lachnospiraceae on stress-induced visceral hypersensitivity in rats. J. Gastroenterol. Hepatol. 2019, 34(8), 1368-76. https://doi.org/10.1111/jgh.14536. Epub 2018 Dec 16. Pubmed PMID: 30402954; Pubmed Central PMCID: PMC7379616. [CrossRef]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229-243. https://doi.org/10.1016/j.physbeh.2018.11.021. Epub 2018 Nov 20. PMID: 30468744. [CrossRef] [PubMed]
- Palme, R., Rettenbacher, S., Touma, C., El-Bahr, S.M., Möstl, E. Stress hormones in mammals and birds: comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Ann. N. Y. Acad. Sci. 2005, 1040, 162-71. https://doi.org/10.1196/annals.1327.021. PMID: 15891021. [CrossRef] [PubMed]
- Shepherdson, D., Lewis, K.D., Carlstead, K., Bauman, J., Perrin, N. Individual and environmental factors associated with stereotypic behavior and fecal glucocorticoid metabolite levels in zoo housed polar bears. Appl. Anim. Behav. Sci. 2013, 147(3-4), 268-77. https://doi.org/10.1016/j.applanim.2013.01.001. [CrossRef]
- Sheriff, M.J., Dantzer, B., Delehanty, B., Palme, R., Boonstra, R. Measuring stress in wildlife: techniques for quantifying glucocorticoids. Oecologia 2011, 166(4), 869-87. https://doi.org/10.1007/s00442-011-1943-y. Epub 2011 Feb 23. PMID: 21344254. [CrossRef] [PubMed]
- von der Ohe, C.G., Servheen, C. Measuring Stress in Mammals Using Fecal Glucocorticoids: Opportunities and Challenges. Wildl. Soc. Bull. 2002, 30(4), 1215-25. https://doi.org/10.2307/3784291. [CrossRef]
- Palme, R., Fischer, P., Schildorfer, H., Ismail, M.N. Excretion of infused 14C-steroid hormones via faeces and urine in domestic livestock. Anim. Reprod. Sci. 1996, 43(1), 43-63. https://doi.org/10.1016/0378-4320(95)01458-6. [CrossRef]
Figure 1.
Circadian rhythm of (i) body temperature (°C), (ii) heart rate (beats per minute [bpm]), and (iii) locomotor activity (overall dynamic body acceleration [ODBA]) (median, interquartile range) for the study cheetahs (CH-2205, -2206, and -2276) throughout the study, i.e., in the treatment and control. Statistics were performed using a mixed model for repeated measures.
Figure 1.
Circadian rhythm of (i) body temperature (°C), (ii) heart rate (beats per minute [bpm]), and (iii) locomotor activity (overall dynamic body acceleration [ODBA]) (median, interquartile range) for the study cheetahs (CH-2205, -2206, and -2276) throughout the study, i.e., in the treatment and control. Statistics were performed using a mixed model for repeated measures.
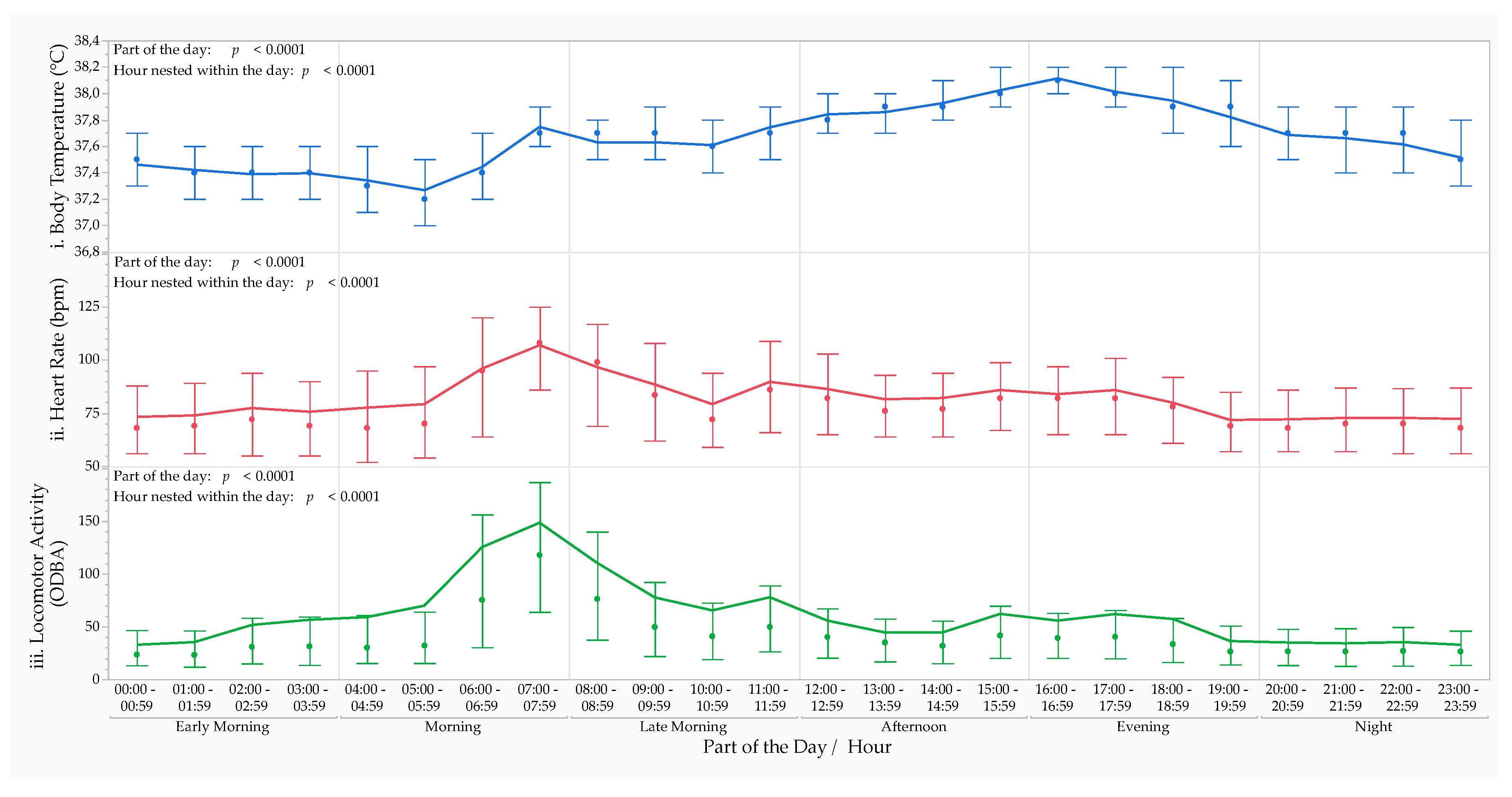
Figure 2.
Box and whisker plot of body temperature (°C) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feeding time (yes [Y] versus no [N]) by part of the day (afternoon, evening). Statistics were performed using Tukey’s HSD post hoc test.
Figure 2.
Box and whisker plot of body temperature (°C) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feeding time (yes [Y] versus no [N]) by part of the day (afternoon, evening). Statistics were performed using Tukey’s HSD post hoc test.
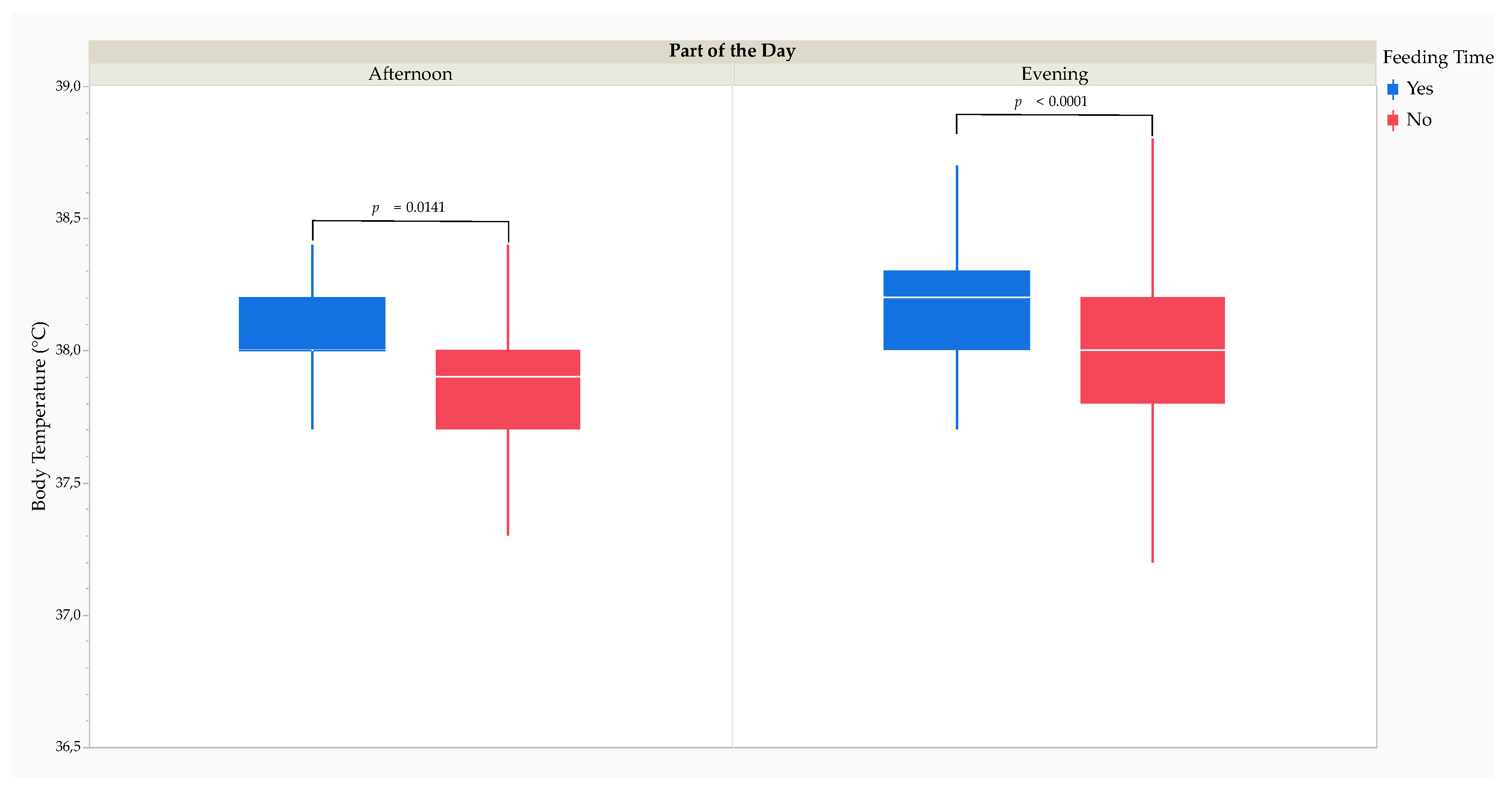
Figure 3.
Box and whisker plot of heart rate (beats per minute [bpm]) for study cheetahs (CH-2205, -2206, and -2276). Effect of feed versus starve day by the study period. Statistics were performed using Tukey’s HSD post hoc test.
Figure 3.
Box and whisker plot of heart rate (beats per minute [bpm]) for study cheetahs (CH-2205, -2206, and -2276). Effect of feed versus starve day by the study period. Statistics were performed using Tukey’s HSD post hoc test.
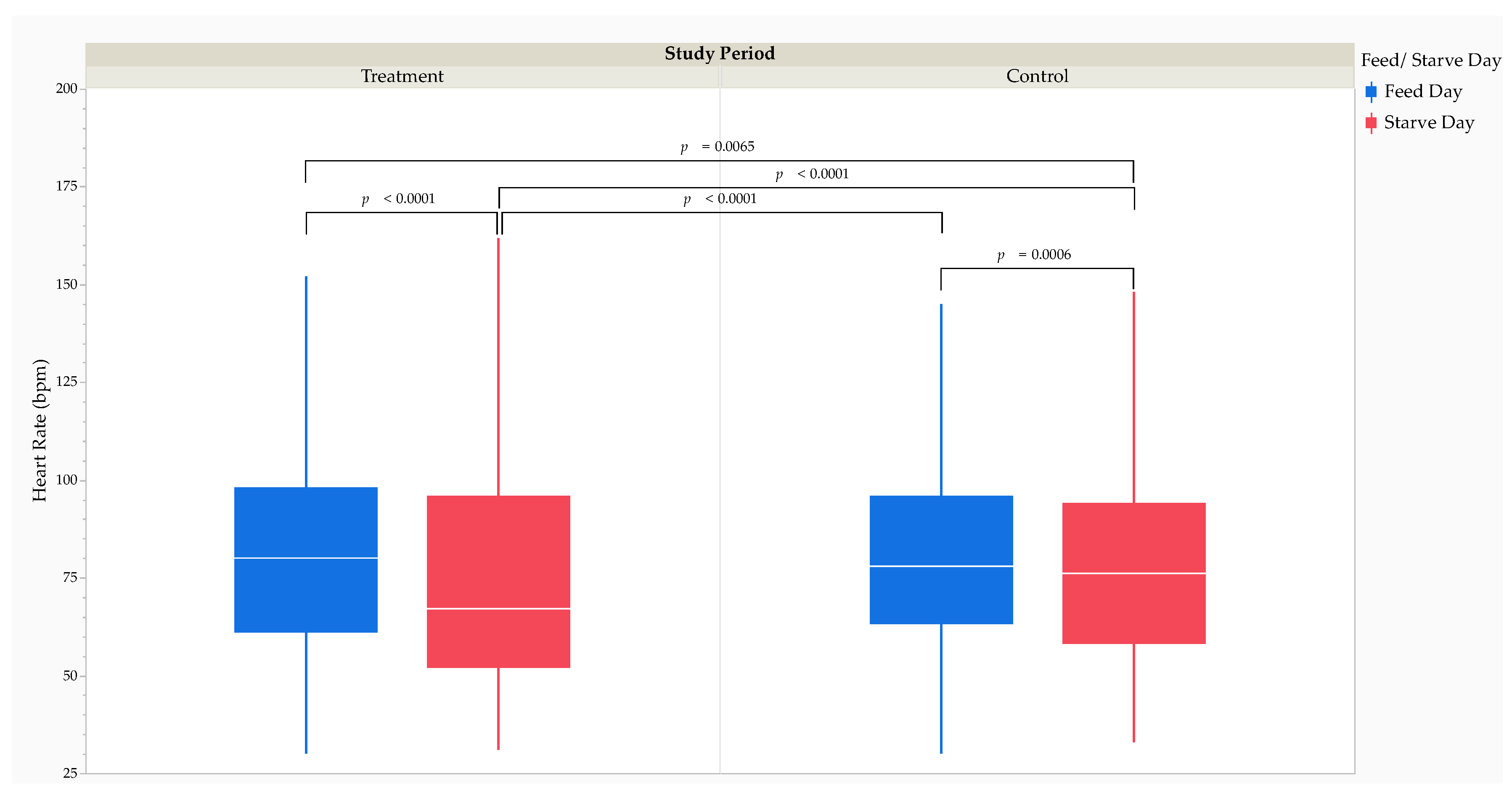
Figure 4.
Box and whisker plot of heart rate (bpm) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feeding time (yes [Y] versus no [N]) by part of the day (late morning). Statistics were performed using Tukey’s HSD post hoc test.
Figure 4.
Box and whisker plot of heart rate (bpm) for the study cheetahs (CH-2205, -2206, and -2276). Effect of feeding time (yes [Y] versus no [N]) by part of the day (late morning). Statistics were performed using Tukey’s HSD post hoc test.
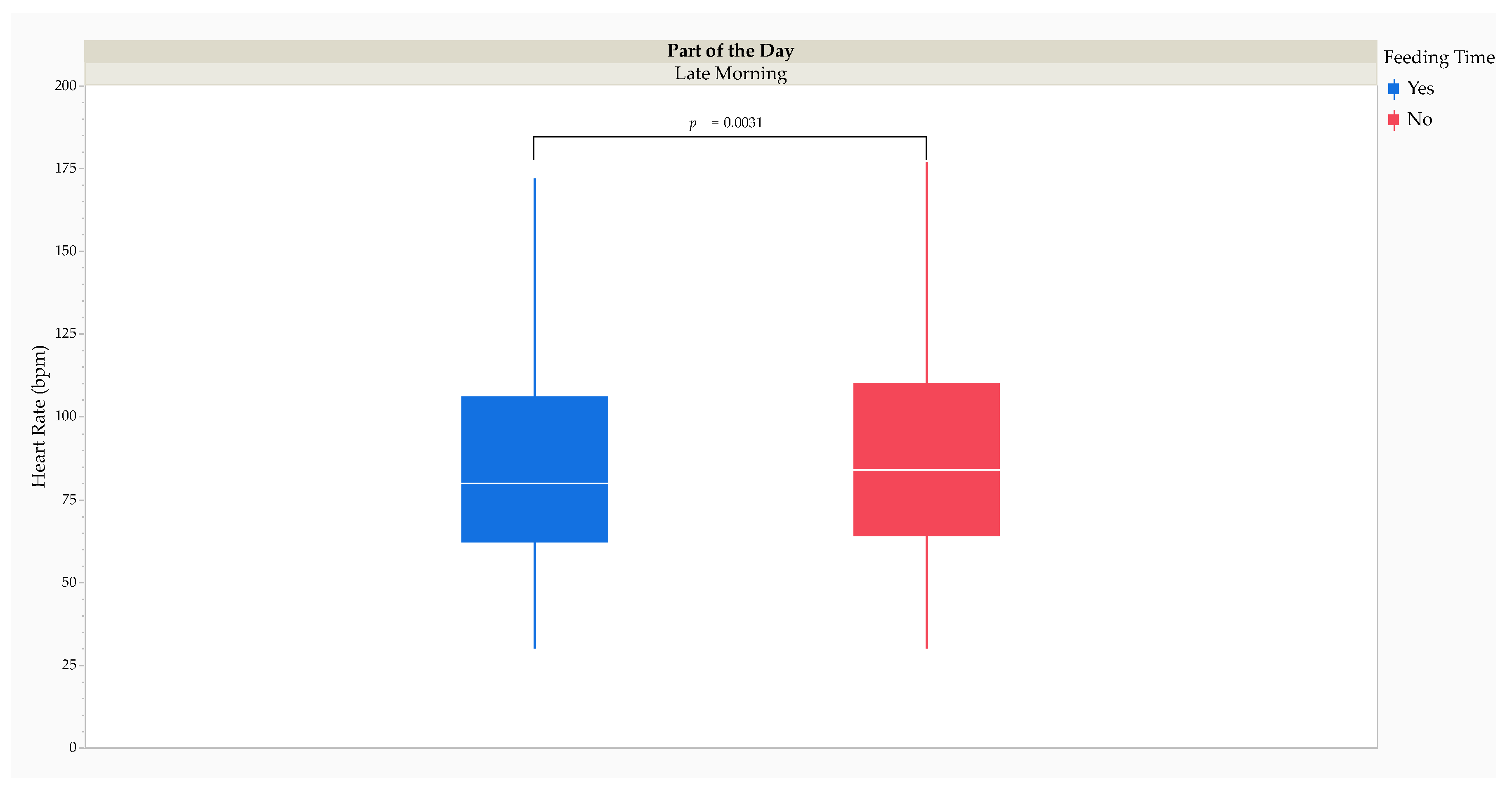
Figure 5.
Box and whisker plot of locomotor activity (overall dynamic body acceleration [ODBA]) for study cheetahs (CH-2205, -2206, and -2276). Effect of feed versus starve day by the study period. Statistics were performed using Tukey’s HSD post hoc test.
Figure 5.
Box and whisker plot of locomotor activity (overall dynamic body acceleration [ODBA]) for study cheetahs (CH-2205, -2206, and -2276). Effect of feed versus starve day by the study period. Statistics were performed using Tukey’s HSD post hoc test.
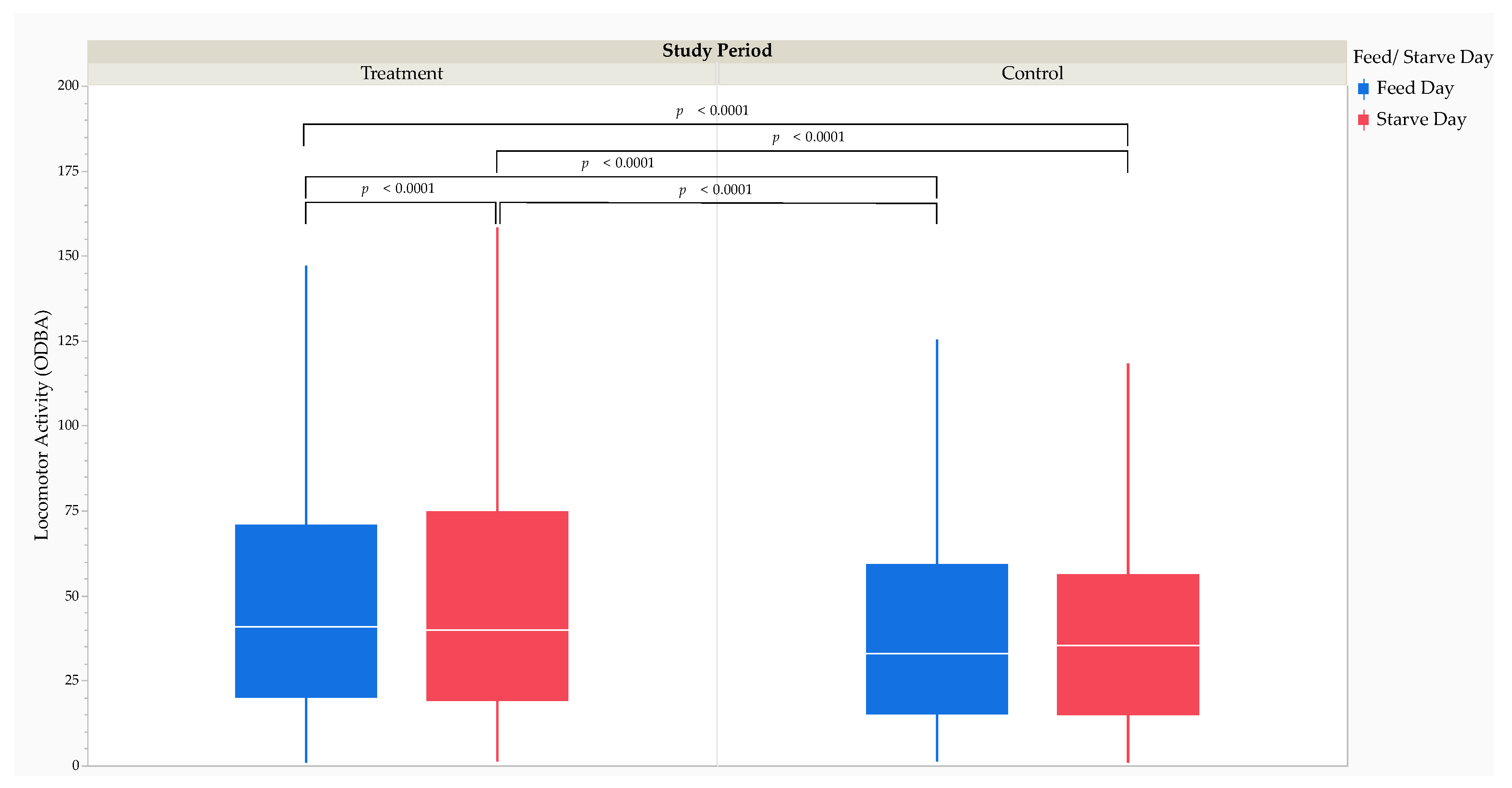
Figure 6.
Box and whisker plot of faecal consistency score (grade) for the study cheetahs (CH-2205, -2206, -2207, -2271, -2276, and -2277). Effect of feed versus starve day by the study period. Statistics were performed using Tukey’s HSD post hoc test.
Figure 6.
Box and whisker plot of faecal consistency score (grade) for the study cheetahs (CH-2205, -2206, -2207, -2271, -2276, and -2277). Effect of feed versus starve day by the study period. Statistics were performed using Tukey’s HSD post hoc test.
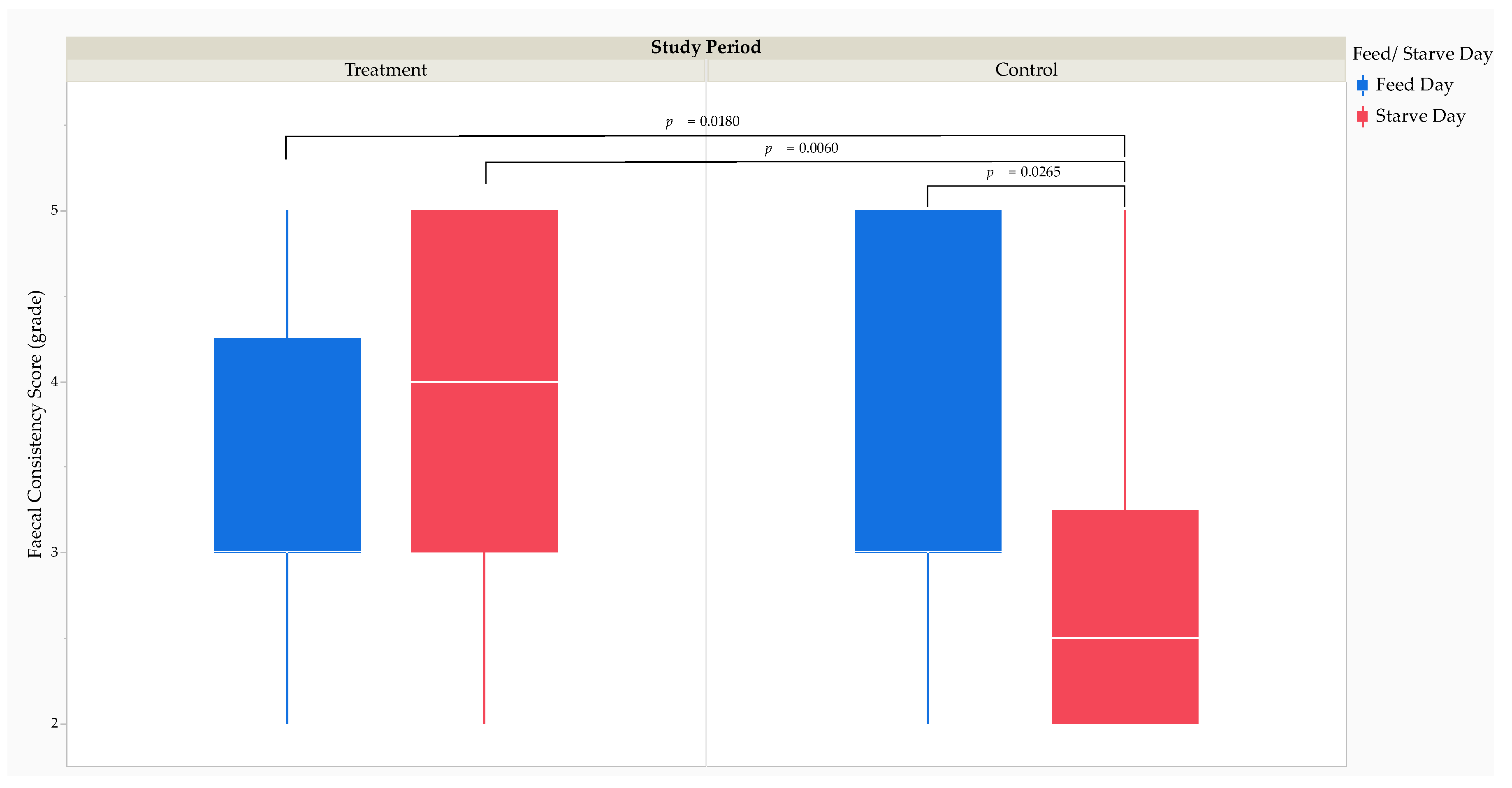

Table 1.
Demographic information of the study cheetahs.
| Group | Identification Number | Origin | Housing | DOB 1 (YY-MM-DD) | BM1 2 (kg) | BM2 3 (kg) | Sex |
|---|---|---|---|---|---|---|---|
| 1 | CH-2205 | Captive-born (hand-reared) | Paired (with CH-2206) | 2016-06-09 | 45.0 | 45.0 | M |
| CH-2206 | Captive-born (hand-reared) | Paired (with CH-2205) | 2016-06-09 | 47.15 | 47.65 | M | |
| CH-2207 | Captive-born (hand-reared) | Single | 2016-06-09 | 36.6 | 37.1 | F | |
| 2 | CH-2271 | Captive-born (hand-reared) | Single | 2017-08-28 | 37.65 | 41.75 | M |
| CH-2276 | Captive-born (hand-reared) | Paired (with CH-2277) | 2017-09-16 | 39.9 | 39.35 | F | |
| CH-2277 | Captive-born (hand-reared) | Paired (with CH-2276) | 2017-09-16 | 43.05 | 43.55 | F |
1 DOB, date of birth; 2 BM1, body mass at the start of the experimental trials; 3 BM2, body mass at the end of the experimental trials.
Table 2.
Ethogram of behaviours recorded and their definitions.
| Inactive | |
|---|---|
| Inactive | Laying/asleep, laying/awake, sitting (stationary in a bipedal position) |
| Active | |
| Individual behaviour | |
| Appetitive behaviour | Feeding, food anticipatory activity, stalking |
| Attention | Staring at one area or paying attention to any visual or auditory stimulus |
| Auto-grooming | Licking or scratching of the own body |
| Environmental enrichment | Interacting with an enrichment device by biting, dragging, scratching, or carrying it in the mouth |
| Locomotion | Jumping, running, solitary play, walking |
| Maintenance | Drinking, defecating/urinating, yawning |
| Olfactory exploration | Sniffing the air, an object, or the substrate; performing flehmen |
| Scent marking | Marking substrates or objects in the enclosure by urine-spraying (releasing urine backwards against a vertical surface or object while standing with tail raised vertically), rolling and rubbing (leaving scents on the substrate or on any object, respectively) |
| Standing | Stationary in a quadrupedal position |
| Stereotypical | Pacing (repetitive, apparently functionless locomotory movement along a given route uninterrupted by other behaviours) |
| Vocalisation | Chirping, growling, purring, stutter-barking, or yowling |
| Social behaviour | |
| Affiliative behaviour * | Social play (play-fight, chasing, or playing together with an enrichment item), pawing, or rubbing on a conspecific, social grooming (licking a conspecific or being licked), paying attention to conspecifics by observing them with interest, and interacting with human caretakers |
| Agnostic behaviour * | Aggression, dominance mount, threat display |
| Interspecific behaviour | Paying attention to another species’ presence |
| Not observed | |
| Out of sight | Focal animal is not visible from the point of observation/behaviour unknown |
Table 3.
Root Mean Square Standardised Effects on body temperature (°C), heart rate (beats per minute [bpm]), locomotor activity (overall dynamic body acceleration [ODBA]), and faecal consistency score (grade) for the study cheetahs (CH-2205, -2206, -2207, -2271, -2276, and -2277). Effect sizes were calculated using Cohen’s d.
Table 3.
Root Mean Square Standardised Effects on body temperature (°C), heart rate (beats per minute [bpm]), locomotor activity (overall dynamic body acceleration [ODBA]), and faecal consistency score (grade) for the study cheetahs (CH-2205, -2206, -2207, -2271, -2276, and -2277). Effect sizes were calculated using Cohen’s d.
| Variable | Effect | Level | - Level | t | p | df 1 | Cohen’s d | 95 % CI 2 for Cohen’s d | |
|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||||
| Body temperature (°C) | Feeding time | Yes | No | 9.46 | < 0.0001 | 16698 | 0.41 | 0.32 | 0.49 |
| Feeding time*Part of the day | Yes, evening | No, evening | 7.24 | < 0.0001 | 16698 | 0.65 | 0.48 | 0.83 | |
| Feeding time*Part of the day | Yes, afternoon | No, afternoon | 6.22 | < 0.0001 | 16698 | 0.54 | 0.37 | 0.71 | |
| Heart rate (bpm) | Study period | Control | Treatment | 7.11 | < 0.0001 | 30221 | 0.11 | 0.08 | 0.14 |
| Feed/starve day | Feed day | Starve day | 13.26 | < 0.0001 | 30221 | 0.20 | 0.17 | 0.23 | |
| Study period*FD 3/SD 4 | CFD 5 | TSD 6 | 18.97 | < 0.0001 | 30221 | 0.31 | 0.27 | 0.34 | |
| Study period*FD 3/SD 4 | TFD 7 | TSD 6 | 18.05 | < 0.0001 | 30221 | 0.30 | 0.26 | 0.33 | |
| Study period*FD 3/SD 4 | CSD 8 | TSD 6 | 7.59 | < 0.0001 | 30221 | 0.20 | 0.15 | 0.26 | |
| Study period*FD 3/SD 4 | CFD 5 | CSD 8 | 4.03 | < 0.0001 | 30221 | 0.10 | 0.05 | 0.15 | |
| Study period*FD 3/SD 4 | TFD 7 | CSD 8 | 3.64 | 0.0003 | 30221 | 0.09 | 0.04 | 0.14 | |
| Feeding Time | Yes | No | 1.97 | 0.0484 | 14834 | 0.09 | 0.00 | 0.18 | |
| Feeding time*Part of the day | No, late morning | Yes, late morning | 3.69 | 0.0002 | 14834 | 0.14 | 0.07 | 0.21 | |
| Locomotor activity (ODBA) | Study period | Treatment | Control | 15.48 | < 0.0001 | 33399 | 0.22 | 0.19 | 0.25 |
| Feed/starve day | Feed day | Starve day | 0.38 | 0.7070 | 33399 | 0.01 | -0.02 | 0.03 | |
| Study period*FD 3/SD 4 | TSD 6 | CSD 8 | 10.13 | < 0.0001 | 33399 | 0.26 | 0.21 | 0.31 | |
| Study period*FD 3/SD 4 | TFD 7 | CSD 8 | 9.39 | < 0.0001 | 33399 | 0.23 | 0.18 | 0.27 | |
| Study period*FD 3/SD 4 | TSD 6 | CFD 5 | 14.11 | < 0.0001 | 33399 | 0.22 | 0.19 | 0.25 | |
| Study period*FD 3/SD 4 | TFD 7 | CFD 5 | 14.44 | < 0.0001 | 33399 | 0.18 | 0.16 | 0.21 | |
| Study period*FD 3/SD 4 | TSD 6 | TFD 7 | 2.11 | 0.0347 | 33399 | 0.03 | 0.00 | 0.06 | |
| Feeding time | Yes | No | 7.74 | < 0.0001 | 16686 | 0.33 | 0.25 | 0.42 | |
| Faecal consistency score (grade) | Study Period | Treatment | Control | 3.31 | 0.0011 | 209 | 0.65 | 0.26 | 1.03 |
| Study period*FD 3/SD 4 | TSD 6 | CSD 8 | 3.43 | 0.0007 | 209 | 1.23 | 0.51 | 1.94 | |
| Study period*FD 3/SD 4 | TFD 7 | CSD 8 | 3.17 | 0.0017 | 209 | 1.07 | 0.40 | 1.74 | |
| Study period*FD 3/SD 4 | CFD 5 | CSD 8 | 3.04 | 0.0027 | 209 | 1.01 | 0.35 | 1.66 | |
Numbers in italics represent the medium magnitude of the effect, while bold numbers represent the large magnitude of the effects. 1 df, degrees of freedom; 2 CI, confidence interval; 3 FD, feed days; 4 SD, starve days; 5 CFD, control feed days; 6 TSD, treatment starve days; 7 TFD, treatment feed days; 8 CSD, control starve days.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



