
Submitted:
10 August 2023
Posted:
11 August 2023
You are already at the latest version
Abstract
Researchers have been exploring seaweed to reduce methane (CH4) emissions from livestock. This study aimed to investigate the potential of a red alga, Bonnemaisonia hamifera, as an alternative to mitigate CH4 emissions. B. hamifera, harvested from the West coast of Sweden, was used in an in vitro experiment using a fully automated gas production system. The experiment was a ran-domized complete block design consisting of a 48-h incubation that included a control (grass si-lage) and B. hamifera inclusions at 2.5%, 5.0% and 7.5% of grass silage OM mixed with buffered rumen fluid. Predicted in vivo CH4 production and total gas production were estimated by ap-plying a set of models to the gas production data and in vitro fermentation characteristics were evaluated. The results demonstrated that the inclusion of B. hamifera reduced (P = 0.01) predicted in vivo CH4 and total gas production, and total gas production linearly decreased (P = 0.03) with higher inclusion of B. hamifera. The molar proportion of propionate increased (P = 0.03) while isovalerate decreased (P = 0.04) with inclusion of B. hamifera. There was a tendency for increased (0.06 ≤ P ≤ 0.10) total volatile fatty acid production, as well as lower proportions of butyrate, isobutyrate, and 2-methylbutyrate. Chemical analyses revealed that B. hamifera had moderate concentrations of polyphenols. The iodine content was low and there was no detectable bromo-form, suggesting quality advantages over Asparagopsis taxiformis. Additionally, B. hamifera exhib-ited antioxidant activity comparable to the positive control Resveratrol. The findings of this study indicated that B. hamifera harvested from temperate waters in Sweden possesses capacity to mitigate CH4 in vitro.
Keywords:
dairy cow
; greenhouse gas
; macroalga
; methane
1. Introduction
Over the past decade, the discussion on the negative impact of meat and dairy production on the environment has gained a considerable momentum due to methane (CH4) emissions and global warming. Globally, as much as 44% of the total CH4 emissions can be attributed to agriculture [1]. Approximately 40% of these emissions can be attributed to the fermentation of feed by cattle [2]. Research has demonstrated that the macroalga Asparagopsis taxiformis is among the most effective feed additives for mitigating enteric CH4 emissions from ruminants [3,4]. The mechanism of reduction is largely attributed to halogenated secondary metabolites, particularly bromoform [3], which acts by directly inhibiting methanogenesis [5]. Researchers concluded that commercial production of A. taxiformis could create new economies due to the fact that small quantities of this seaweed in the diet of ruminant animals reduced CH4 emissions by up to 98% when included at 0.05% of organic matter (OM) intake [5]. However, bromoform is a known carcinogen, and there has been elevated concentrations bromide and iodine in the milk of dairy cows fed with A. taxiformis [6,7]. A. taxiformis is native to South Australia and it is currently not cultivated in large quantities in the Northern hemisphere.
Bonnemaisonia hamifera is also a type of red alga of the same order Bonnemaisoniales and family Bonnemaisoniaceae as A. taxiformis. In New Zealand B. hamifera was shown to have a strong CH4 inhibitory effect in vitro of 95.4%, and 98.8% relative to the basal feed substrate at inclusion levels of 6% and 10% on OM basis [8]. Furthermore, Mihaila et al. [8] showed that the primary bioactive compound bromoform in A. taxiformis was not detected in B. hamifera. We hypothesized that native B. hamifera wild-harvested from the West coast of Sweden was going to display a CH4 inhibitory effect in vitro and be a temperate seaweed alternative to cultivate, and less susceptible to the loss of harmful volatile bioactives during processing and handling. The objectives of this study was to measure the CH4 inhibitory effect in vitro of B. hamifera harvested in temperate water in Sweden.
2. Materials and Methods
The macroalga B. hamifera was harvested from Kristineberg Center for Marine Research and Innovation in Fiskebäckskil (58°14′N 11°27′E) on the West coast of Sweden. The seaweed was harvested from the shore in accordance with the Nagoya protocol guidelines (https://www.cbd.int/abs/doc/protocol/nagoya-protocol-en.pdf), packed in cool boxes, and transported via overnight courier to Swedish University of Agricultural Sciences in Umeå on dry ice. Samples were washed to remove sand and epiphytes and stored at -18°C. All samples were freeze-dried using a laboratory-scale Labconco FreeZone freeze dryer equipped with tray dryers (Labconco, Kansas city, Missouri, USA) operating at -84°C.
The donor animals of rumen inoculum, equipment used and procedures of the in vitro followed the recent work reported by Krizsan et al. [9]. In brief, rumen fluid was directly transported to the laboratory after collection and filtered through cheesecloth into Thermos flasks. The samples were in total repeated across two water baths to get one bottle with blank (i.e. bottles with 60 mL of buffered rumen fluid with no sample or treatment in), duplicate bottles with control sample consisting of grass silage, and three replicates of treatment samples containing grass silage and B. hamifera in each bath. The B. hamifera was added at inclusion levels of 2.5%, 5% and 7.5% on OM basis. All samples were randomly distributed among the in vitro bottles in each bath. Gas production was measured with a fully automated system (Gas Production Recorder, GPR-2, Version 1.0 2015, Wageningen UR). Measurement of CH4 was performed by withdrawing gas samples (0.2 mL) at 2, 4, 8, 24, 32, and 48 h from all in vitro bottles. The concentration of CH4 was determined immediately after collection by injecting the gas sample in a Trace 1300 gas chromatograph (Thermo Fisher Scientific, Waltham, MA, USA). The CH4 concentration (mL/g sample) of all samples were used in model simulations to achieve in vivo predicted CH4 according to Ramin and Huhtanen [10].
For the alga, the N percentage in the sample was determined using the LECO FP628 (LECO Corp., MI, USA) protein analyser applying the Dumas AOAC method 992.15 (1990) [11] and protein content was obtained using a conversion factor of 5.0 [12]. The NDF concentration was determined free of residual ash following the protocol outlined by Van Soest et al. [13], using a 1020 hot and 1021 cold extractor (Tecator Fibertec System; Foss Analytical) with addition of heat-stable α-amylase and sodium sulphite. The percentage lipid in each sample was assessed using the Oracle NMR Smart Trac rapid Fat analyser (CEM Corporation, USA) using AOAC official methods 985.14. The ash and moisture contents were determined according to [11].
As detailed in Krizsan et al. [9], the total polyphenol concentration (TPC) of the macroalga was estimated using the Folin Ciocalteau reagent; the iodine content was determined using the Iodine Colorimetric Assay Kit (BioVision, Milpitas, California, USA), and the antioxidant capacity was determined using the 2,2-diphenyl-1-picrylhydrazine (DPPH) Antioxidant Assay Kit (AbCam, The Netherlands -ab289847, K2078). Bromoform concentration in Macroalga extract was carried out as described in Krizsan et al. [9].
Individual volatile fatty acid (VFA) concentrations in in vitro rumen fluid samples were determined using a Waters Alliance 2795 UPLC system as described by Puhakka et al. [14].
All statistical analyses were performed using SAS version 9.4 (SAS Institute Inc.). Data were subjected to ANOVA using the MIXED procedure in SAS with treatment, water bath, and their interaction as fixed effects and bottle position in water bath as a random effect.
Treatments were compared using orthogonal contrasts; contrasts were constructed to evaluate the effects of inclusion of B. hamifera and the linear and quadratic effects of inclusion levels.
3. Results
The macroalga had DM, OM, CP, NDF and crude fat concentrations of 152±1.3 g/kg of fresh weight, and 505±6.7, 97±3.1 and 4.3±0.26 g/kg of DM.
The inclusion of B. hamifera decreased (P = 0.01) predicted in vivo CH4 (Figure 1a) and total gas production (Figure 1b). There was a quadratic effect (P = 0.01) of increased levels of B. hamifera on predicted in vivo CH4 production. The predicted in vivo total gas production linearly decreased (P = 0.03).
Propionate was greater (P = 0.03) and isovalerate, lower (P = 0.04) with the inclusion of B. hamifera compared to the control (Table 1). A tendency of increased (0.06 ≤ P ≤ 0.10) total VFA production and proportions of butyrate, isobutyrate, and 2-methylbutyrate were observed with the inclusion of B. hamifera compared to the control. We also found a quadratic effect (P ≤ 0.05) on proportions of isobutyrate, 2-methylbutyrate, and isovalerate as well as a quadratic tendency (0.08 ≤ P ≤ 0.10) on total VFA production and the proportion of butyrate with an increase in B. hamifera inclusion levels.
The respective average polyphenol and iodine contents of B. hamifera sample were 0.165 mg Gallic acid equivalents and 71.1 µg/L iodine. The value obtained for total antioxidant activity of B. hamifera was 0.395 µM Trolox equivalents mg/ml sample. This is comparable to the positive control Resveratrol which had a DPPH value of 0.409 µM Trolox equivalents mg/ml (n=3). There was no bromoform detected in the B. hamifera used in this study.
4. Discussion and Conclusion
The potential of feeding red algae to reduce CH4 emissions from ruminants is a promising solution for a more sustainable production of food from cattle. However, there needs to be a system for use, i.e. cultivating, distributing and storing red algae on the farm without a change in the active substances occurring and assuring safety. Primarily, in order to assure an efficient CH4 mitigation, but of no less importance to minimize the harmful risk of substances like bromofrom. Poor mixing and an accidently large dose of A. taxiformis could cause damage to the rumen wall of single cows [15] and reduced feed intake [6].
It is worth noting that a high concentration of bromoform in red algae have lead to greater CH4 reduction [5]. In our study, the inclusion of B. hamifera resulted in a modest 12.3% reduction in predicted in vivo CH4 production compared to an earlier in vitro study conducted in New Zealand that reported CH4 reductions of most 98.8% at an inclusion of 10% on an OM basis [8]. The inhibitory effect seemed to be mediated by longer-chained halogenated hydrocarbons, likely by the same inhibitory mechanism as A. taxiformis [8]. Enge et al. [16] found that B. hamifera was chemically defended by producing 1,1,3,3-tetrabromo-2-heptanone (an halogenated secondary metabolites) as the main feeding deterrent compound. This compound could be a prospective candidate for exhibiting anti-methanogenic effect in the rumen.
In terms of ruminal fermentation patterns, most in vivo experiments with red algae have demonstrated a shift towards increased propionate production, confirming its role in CH4 inhibition [5,6,7,17]. Several CH4 inhibitory mechanisms could have been the reason for the effect observed in the present study, but most likely the bioactive substances in Swedish B. hamifera affected a broader spectrum of the microbiome since total gas was decreased in supplemented treatments in vitro. Depending on where they grow and when they are harvested, algae will contain different levels of bioactive substances [12], which likely can explain the observed differences between B. hamifera harvested in Sweden and New Zealand. Ruminal branched-chain VFA (BCVFA; isobutyrate, isovalerate, and 2-methylbutyrate) are derived mainly from the deamination of branched-chain amino acids in the diet. Branched-chain VFA supplementation has been shown to improve digestibility and production in ruminants by providing an additional energy source and promoting the proliferation of cellulolytic bacteria [18]. In our study, the reduction in BCVFA proportions may indicate less microbial activity, contributing to the overall reduction in CH4 emissions.
In many ways, red algae open up the possibility of producing organic food from dairy cows with reduced CH4 emissions. B. hamifera harvested on the West coast of Sweden does not give a satisfactory reduction of CH4 compared to other more readily available dietary mitigation strategies that could be suitable also in organic cattle production. On the other hand, the low iodine content and absence of bromoform in B. hamifera make it a potentially safer and more environmentally friendly option compared to A. taxiformis for CH4 mitigation in ruminants. These characteristics reduce the risk of negative health effects on animals and minimize potential ecological concerns. However, further research is necessary to fully understand the specific bioactive substances present in B. hamifera and their effects on CH4 production to optimize its utilization as a sustainable solution for reducing greenhouse gas emissions in livestock production. To further understand the differences observed in CH4 inhibition, it is important to investigate the conditions specific to New Zealand, where more significant reductions in CH4 emissions were reported in previous studies.
Author Contributions
Conceptualization, A. G., F. G., and S. K.; methodology, A. G., M.H., and S. K.; software, A.G.; validation, A. G. and S. K.; formal analysis, A. G.; investigation, A. G. and S. K.; resources, A. G., M. H., F. G., and S. K.; data curation, A. G.; writing—original draft preparation, A. G., M. H., and S. K.; writing—review and editing, A. G. and S. K., visualization, A.G.; supervision, S.K.; project administration A. G. and S. K.; funding acquisition, A. G., M.H., F. G., and S. K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Union’s Horizon2020 Research & Innovation Programme under grant agreement No 696356 and from ICA Sverige AB, Solna, Sweden.
Institutional Review Board Statement
The animal study protocol was approved by the Swedish Ethics Committee on Animal Research (Dnr A 32-16), represented by the Court of Appeal for Northern Norrland in Umeå, and the experiment was carried out in accordance with laws and regulations governing experiments performed with live animals in Sweden. .
Conflicts of Interest
The authors declare no conflict of interest.
References
- UNEP. Partnership and United Nations Environment Programme. Reducing consumer food waste using green and digital technologies. 2021. Available online: https://www.unep.org/resources/publication/reducing-consumer-food-waste-using-green-and-digital-technologies (accessed on 14 July 2023).
- Gerber, P.J.; Hristov, A.N.; Henderson, B.; Makkar, H.; Oh, J.; Lee, C.; Meinen, R.; Montes, F.; Ott, T.; Firkins, J.; Rotz, A. Technical options for the mitigation of direct methane and nitrous oxide emissions from livestock: a review. Animal 2013, 7, 220–234. [Google Scholar] [CrossRef]
- Machado, L.; Kinley, R.D.; Magnusson, M.; de Nys, R.; Tomkins, N.W. The potential of macroalgae for beef production systems in Northern Australia. J. Applied Phycology 2015, 27, 2001–2005. [Google Scholar] [CrossRef]
- Kinley, R.D.; de Nys, R.; Vucko, M.J.; Machado, L.; Tomkins, N.W. The red macroalgae Asparagopsis taxiformis is a potent natural antimethanogenic that reduces methane production during in vitro fermentation with rumen fluid. Anim. Prod. Sci. 2016, 56, 282–289. [Google Scholar] [CrossRef]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J. Cleaner Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Stefenoni, H.A.; Räisänen, S.E.; Cueva, S.F.; Wasson, D.E.; Lage, C.F.A.; Melgar, A.; Fetter, M.E.; Smith, P.; Hennessy, M.; Vecchiarelli, B.; Bender, J. Effects of the macroalga Asparagopsis taxiformis and oregano leaves on methane emission, rumen fermentation, and lactational performance of dairy cows. J. Dairy Sci 2021, 104, 4157–4173. [Google Scholar] [CrossRef] [PubMed]
- Krizsan, S.J.; Ramin, M.; Chagas, J.C.; Halmemies-Beauchet-Filleau, A.; Singh, A.; Schnürer, A.; Danielsson, R. Effects on rumen microbiome and milk quality of dairy cows fed a grass silage-based diet supplemented with the macroalga Asparagopsis taxiformis. Front. Anim. Sci. 2023, 4, 1112969. [Google Scholar] [CrossRef]
- Mihaila, A.A.; Glasson, C.R.; Lawton, R.; Muetzel, S.; Molano, G.; Magnusson, M. New temperate seaweed targets for mitigation of ruminant methane emissions: an in vitro assessment. Applied Phycology 2022, 3, 274–284. [Google Scholar] [CrossRef]
- Krizsan, S.J.; Hayes, M.; Gröndahl, F.; Ramin, M.; O’Hara, P.; Kenny, O. Characterization and in vitro assessment of seaweed bioactives with potential to reduce methane production. Front. Anim. Sci. 2022, 3, 1062324. [Google Scholar] [CrossRef]
- Ramin, M.; & Huhtanen, P. Development of an in vitromethod for determination of methane production kinetics usinga fully automated in vitro gas system—A modelling approach. Anim. Feed Sci. Technol. 2012, 174, 190–200. [Google Scholar] [CrossRef]
- AOAC International. AOAC International Official Methods of Analysis, 9th ed.; AOAC Int.,: Gaithersburg, MD, 2012. [Google Scholar]
- Abbott, D.W.; Aasen, I.M.; Beauchemin, K.A.; Gröndahl, F.; Gruninger, R.; Hayes, M.; Huws, S.; Kenny, D.A.; Kirwan, S.; Krizsan, S.J.; Lind, V.; Meyer, U.; Ramin, M.; Theodoridou, K.; von Soosten, D.; Walsh, P.J.; Waters, S.; Xing, M. Seaweed and seaweed bioactives for mitigation of enteric methane: Challenges and opportunities. Animals. 2020, 10, 2432. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J.; Robertson, J.B.; B., A. Lewis. Methods for dietary fiber, neutral detergent fiber and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Puhakka, L.; Jaakkola, S.; Simpura, I.; Kokkonen, T.; Vanhatalo, A. Effects of replacing rapeseed meal with fava bean at two concentrate crude protein levels on feed intake, nutrient digestion, and milk production in cows fed grass silage-based diets. J. Dairy Sci. 2016, 99, 7993–8006. [Google Scholar] [CrossRef]
- Muizelaar, W.; Groot, M.; van Duinkerken, G.; Peters, R.; Dijkstra, J. Safety and transfer study: Transfer of bromoform present in Asparagopsis taxiformis to milk and urine of lactating dairy cows. Foods 2021, 10, 584. [Google Scholar] [CrossRef] [PubMed]
- Enge, S.; Nylund, G.M.; Harder, T.; Pavia, H. An exotic chemical weapon explains low herbivore damage in an invasive alga. Ecology 2012, 93, 2736–2745. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Hess, P.S.; Jacobs, J.L.; Kinley, R.D.; Roque, B.M.; Neachtain, A.S.O.; Chandra, S.; Williams, S.R.O. Twice daily feeding of canola oil steeped with Asparagopsis armata reduced methane emissions of lactating dairy cows. Anim. Feed Sci. and Technol. 2023, 297, 115579. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Guo, G.; Huo, W.J.; Zhang, Y.L.; Pei, C.X.; Zhang, S.L. Effects of rumen-protected folic acid and branched-chain volatile fatty acids supplementation on lactation performance, ruminal fermentation, nutrient digestion and blood metabolites in dairy cows. Anim. Feed Sci. and Technol. 2019, 247, 157–165. [Google Scholar] [CrossRef]
Figure 1.
The effect of B. hamifera at different inclusion levels on predicted in vivo methane (a) and total gas production (b) with SEM of 1.68 and 13.1 ml/g OM, respectively.
Figure 1.
The effect of B. hamifera at different inclusion levels on predicted in vivo methane (a) and total gas production (b) with SEM of 1.68 and 13.1 ml/g OM, respectively.
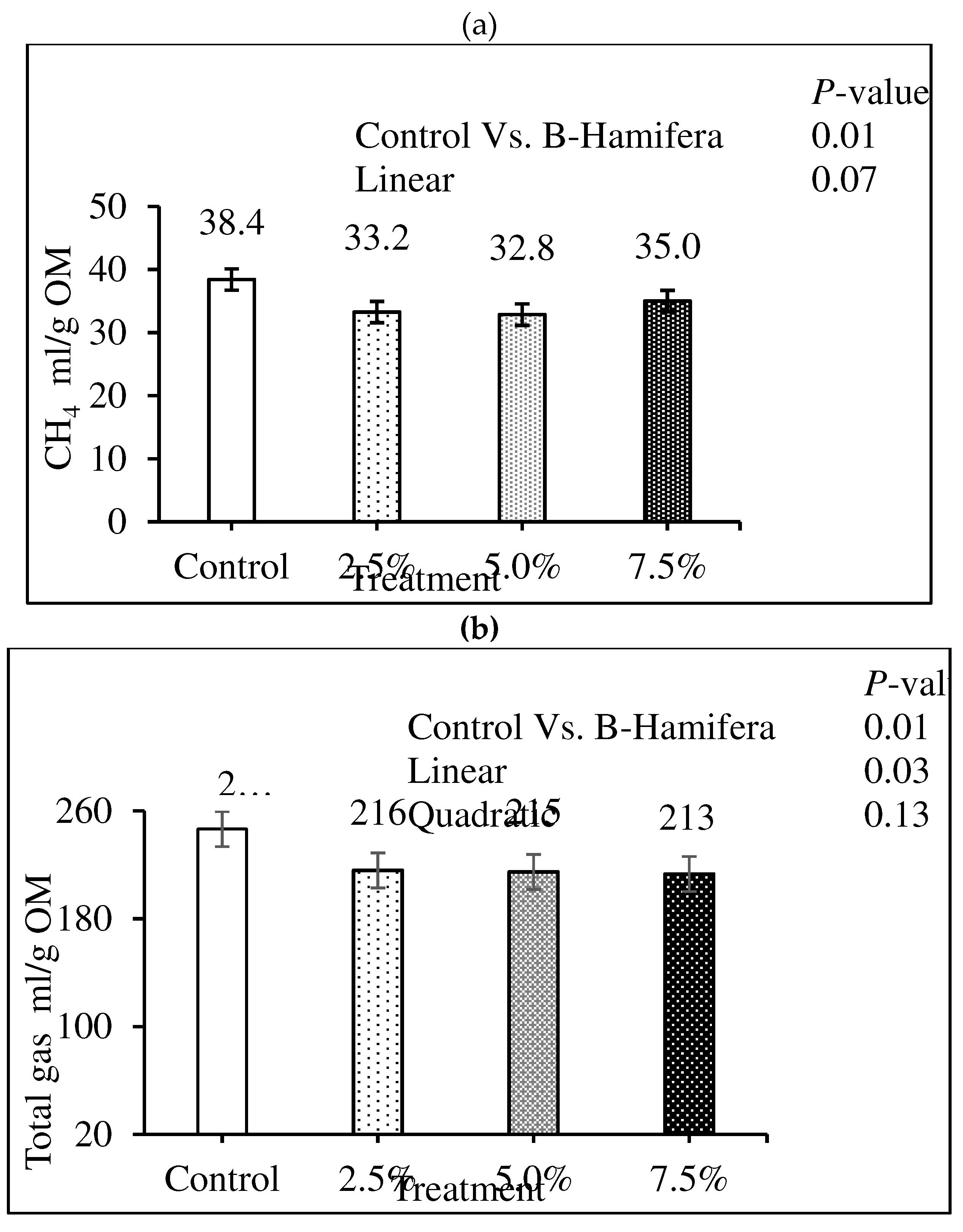

Table 1.
Effects of B. hamifera at different inclusion levels on total volatile fatty acid (VFA) and molar proportions of VFA production at 48 h of incubation in vitro.
Table 1.
Effects of B. hamifera at different inclusion levels on total volatile fatty acid (VFA) and molar proportions of VFA production at 48 h of incubation in vitro.
| Item | Treatments | SEM | P-value | |||||
| Control | B. hamifera inclusion level (% OM) | |||||||
| 2.5% | 5.0% | 7.5% | Control vs. B. hamifera | Linear | Quadratic | |||
| Total VFA, mM | 148 | 162 | 159 | 155 | 6.6 | 0.10 | 0.44 | 0.08 |
| VFA molar proportions, mmol/mol | ||||||||
| Acetate | 575 | 574 | 575 | 577 | 2.4 | 0.94 | 0.39 | 0.42 |
| Propionate | 241 | 246 | 244 | 245 | 2.2 | 0.03 | 0.11 | 0.13 |
| Butyrate | 98.4 | 96.7 | 96.9 | 97.5 | 0.90 | 0.10 | 0.39 | 0.09 |
| Isobutyrate | 14.9 | 14.2 | 14.2 | 14.6 | 0.34 | 0.06 | 0.38 | 0.04 |
| 2-Methylbutyrate | 11.6 | 10.9 | 11.0 | 11.2 | 0.31 | 0.06 | 0.36 | 0.05 |
| Isovalerate | 13.8 | 12.8 | 13.0 | 13.3 | 0.39 | 0.04 | 0.33 | 0.03 |
| Valerate | 24.5 | 23.8 | 23.8 | 19.9 | 3.49 | 0.52 | 0.24 | 0.53 |
| Caproate | 21.2 | 21.9 | 21.3 | 21.6 | 0.40 | 0.29 | 0.62 | 0.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



