
Submitted:
11 August 2023
Posted:
11 August 2023
You are already at the latest version
Abstract
The high-mountain and arctic plants are considered especially sensitive to the climate changes because of close adaptations to the cold environment. Kalmia procumbens, the typical arctic-alpine species reaches southernmost European localities in the Pyrenees and Carpathians. The aim of the study was the assessment and comparison of K. procumbens current potential niche areas in the Pyrenees and Carpathians and their possible reduction due to climate change, depending on the scenario. Realized niches of K. procumbens in the Pyrenees are compact while in the Carpathians dispersed. In both mountain chains, the species occurs in the alpine and subalpine vegetation belts, going down to elevations of about 1500-1600 m, while the most elevated localities in the Pyrenees are at ca 3000 m, about 500 m higher than in the Carpathians. The localities of K. procumbens in the Carpathians have a more continental climate than in the Pyrenees, with lower precipitation and temperatures but higher seasonality of temperature and of precipitation. The species covered a larger area of geographic distribution during Last Glacial, and reduced area during mid Holocene. Due to the climate warming, the strong reduction of potential area of occurrence to 2100 is expected in the Carpathians and moderate reduction in in the Pyrenees.
Keywords:
biogeography
; climate change
; ecological niche modeling
; geographic range
; relict plant
1. Introduction
The origin of cold-adapted plant species and the formation of tundra in northern Europe, Asia and North America took place in turn of the Pliocene/Pleistocene [1,2,3] as a reaction of the plant cover to the climate cooling. The plants adapted to the low temperatures and short vegetation period in the Arctic zone and in the high mountains in southern-more regions evolved during approximately similar periods [3]. The Arctic plants reached mountains in the central Europe, Asia and North America escaping to the south before Pleistocene glaciers [1]. Inversely, the alpine plants could migrate to the North in the periglacial zone during deglaciations [4], as in the case of retreating glaciers, observed during the last centuries in the Alps [5]. During interglacial periods of Pleistocene (Holocene including), the plant species connected with the cold climate could survive only in the Arctic and in the high mountains above the timberline, sometimes also on mires at lower altitudes [6,7,8], being the glacial relicts [9,10].
The high-mountain and arctic plant species are considered to be especially sensitive to the climate changes because of close adaptations to the cold environment [11].The high temperatures and low humidity are expected to be the most important threats [12,13,14,15,16,17,18,19]. High risk results also from the forest line shift and expansion of trees, high shrubs and herbs, which could colonize or at least shade the sites of light-demanding tundra and alpine plant species [20,21,22,23,24].
Kalmia procumbens (L.) Gift & Kron & P.F.Stevens is an arctic-alpine, circum-polar, amphi-Atlantic plant [25,26,27]. It is an evergreen, dwarf, prostrate shrub, frequently creeping to the ground, especially at high elevations. In northern Europe and in the mountains it prefers the rocky ridges [24,28]. In Europe, it reaches its southernmost localities in the Pyrenees, Alps, northern Dinaric Alps and Carpathians. The species is one of the typical cold-adapted, glacial relicts in the Central-European mountains [29,30]. Despite, it can survive in temperatures above 50 °C with very high diurnal amplitude of temperatures [31].
In the Pyrenees and in the Carpathians, K. procumbens localities are confined predominantly to the alpine and sometimes to the subalpine vegetation belts, at lower locations connected mainly with the north-facing slopes [32,33,34], with relatively shorter vegetation period, lower temperatures and temperature amplitudes comparing to the south expositions [35,36,37,38]. It is stress-tolerant, adapted to extreme conditions of existence, characterized by slow growth in a relatively long growing season [39]. The formation of the photosynthetic apparatus is aimed at the minimum return of energy, which contributes to the conservation of resources. The population structure is dominated by plants of mature (generative) stages [40]. It prefers rocky ridges with small snow deposition during winter [28,38]. Such characteristics indicate that the species, although resistant to external influences, recovers rather slowly when it loses its position. The sites of K. procumbens are susceptible to influence of the global climate change [28,38,41]. Despite that, the last observations indicated its more intense growth in the mountains of the Japan islands [42] and even expansion in the Ukrainian Carpathians resulting from the reduction of snow cover [21]. The species had a broader potential ecological niche area at the global scale during Last Glacial Maximum (LGM) than at present [41].
The study aimed to assess and compare K. procumbens current potential niche areas in the Pyrenees and Carpathians and their possible reduction due to climate change depending on the scenario. Additionally, the retrospection of its’ local, Carpathians’ and Pyrenean geographic ranges during Last Glacial Maximum (LGM) was analysed, based on the most probable high level of ecological niche conservatism in plants [43,44,45]. We expected the Carpathian populations would be more prone to the temperature rise and extension of the vegetation period than Pyrenean ones.
2. Results
2.1. Realized geographic niches
The distribution of K. procumbens in the Pyrenees and in the Carpathians is connected with the most elevated mountain massifs (Figure 1). The species has a more compact distribution of their localities in the Pyrenees than in the Carpathians. The altitudinal range in the Pyrenees is broader than in the Carpathians. The altitudinal minima in both mountain systems are similar, but the most elevated localities in the Pyrenees have reached about 400-500 m higher elevations than in the Carpathians (Figure 2). Current range of species differ from the potential range in LGM (Figure S1), when the Carpathian arc was more suitable for the species, whereas conditions in the Pyrenees were less suitable.
Current potential niches of K. procumbens in the Pyrenees and Carpathians are determined first of all by elevation, which influences to the current distribution of the species attain nearly 80% in the Pyrenees and 60% in the Carpathians (Table 1). From the climatic variables, only precipitation of the driest month (bio 14) influences at a relatively high degree in both mountain chains (Table 1). Additional important bioclimatic factor, which influences the potential niche of K. procumbens reaching 1% or more is the temperature mean diurnal range (bio2), temperature annual range (bio7), and precipitation of the driest quarter (bio17) in the Pyrenees, and annual mean temperature (bio1), temperature seasonality (bio4) precipitation seasonality (bio15) and precipitation of warmest quarter (bio18) in the Carpathians (Table 1).
The average values of every bioclimatic factor differed at a statistically significant level (p<0.05) between localities of K. procumbens in the South and East Carpathians (the latter include also one locality from the West Carpathians) (Table 2). The East Carpathian region of K. procumbens occurrence has generally higher precipitation (bio12 – bio19) and is characterized by slightly but statistically significantly lower temperature factors (bio1, bio5, bio6, bio8 – bio 11) than South-Carpathian (Table 2). Populations from Carphatians occur most often in the North-eastern exposition, whereas in the Pyrenees in the North exposition (Figure S2).
Principal Component Analysis (PCA) of the bioclimatic factors for K. procumbens realized niches indicated separate grouping of Carpathian and Pyrenean localities. The East Carpathian localities are well separated from the South Carpathian ones, while Central Pyrenean localities in the great part intermixed East Pyrenean ones (Figure 3). The potential ecological niches estimated on the basis of the species localities in Central and East Pyrenees recognized the realized niches in both Pyrenean regions (Figure 4). Inversely, the potential niche of K. procumbens estimated on the East Carpathian localities recognized realized niche in the East Carpathians, but not in the South Carpathians. Additionally, this combination indicated also highly suitable conditions in the Tatra Mts. in the West Carpathians, where only one natural locality of the species currently exists. The potential ecological niche of K. procumbens estimated on the South Carpathian localities did not detect suitable environmental conditions for the species occurrence neither in the East, nor in the West Carpathians (Figure 4).
2.2. Future geographic niches
The potential niches highly suitable for K. procumbens in the Carpathians completely disappear in 2100, independently of the scenario of climate warming. Only not many South Carpathian populations could persist, mainly in the Fagarash, while the East Carpathian ones would not find suitable conditions (Figure 5). The potential niches in 2100 in the Carpathians would be determined in ca. 50% by elevation in the East, and in ca. 70% in the South Carpathians. The next restrictive factor in the East Carpathians would be precipitation of the warmest quarter (bio18), attaining 30% and more (Table 3). The other bioclimate factors influencing the potential niches in 2100 are the same as current (compare Table 1 and Table 3), but values are slightly smaller.
In the Pyrenees, the situation of K. procumbens populations in 2100 would not be so drastically worse than at present (Figure 6). The environmental conditions in the East Pyrenees allow persist the species populations and even attain broader area of potential niches distribution in the more optimistic scenario (Table 4). In the Central Pyrenees, the potential niche area suitable for K. procumbens in the high and very high level would be restricted (Table 4). As currently, the most influential would be the elevation, reaching about 76 and 80% in the Central and East Pyrenees, respectively (Table 3). The remaining bioclimate factors are the same, as determining current ecological niches of the species in the Pyrenees (see Table 1 and Table 3).
The phytoindication method revealed potential threat to K. procumbens occurrence in the East Carpathians in Ucraine. It results mainly from the changes of hydrological regime of the specias sites. The growth of average yearly temperature by 2 °C (pessimistic scenario) put K. procumbens at moderate, while by 3 °C at catastrophic risk of extinction (Figure 7).
3. Discussion
3.1. Realized potential niches in the Carpathians and Pyrenees
During LGM, the potential range of K. procumbens was broader in the Carpathians, and for East Carpathians populations the model predict suitable areas also at the lower elevation, closer to the ice sheet. In the Pyrenees during LGM, potential range was located in lower elevations, and conditions for Central Pyrenees populations were almost unsuitable. In current conditions, the species attain in the Pyrenees their southern-most, and in the Carpathians close to southern-most localities in Europe [24,27]. Kalmia procumbens survived in both mountain chains in the subalpine and alpine vegetation belts due to high-mountain climate with low temperatures and relatively high precipitations, mostly in the places with restricted snow cover during winters [21,22,28,38].
The low altitudinal border of K. procumbens occurrence in the Carpathians and in the Pyrenees are at similar elevations, as a rule in the habits orographically or edaphically inaccessible for the shrubs, tall herbs, and grasses. The specific site conditions are mostly on the slopes exposed to the North, in the rocky places with very thin layers of soil or on the rocks completely without soil, and in the places open to the winds. Sometimes such conditions can be anthropogenic, for example, could effect from over-pasturing. This kind of pressure stopped last decades and could be one of the reasons for the disappearance of the lowest localities of K. procumbens in the East Carpathians, reported from 1455 m in the Chornokhora [46], not found later [22,33]. The reduction of pastoralism in the Pyrenees during the last decades could also cause the disappearance of the species' lowest localities due to the expansion of the tall herbs and shrubs.
The maximal altitudes of occurrence of K. procumbens in the Pyrenees are more elevated than in the Carpathians. Such a rule was also observed in other subalpine and alpine plants, common for the Pyrenees and Carpathians, as for example Juniperus communis L. var. saxatilis Pall., Salix reticulata L., Salix herbacea L., Salix hastata L., Dryas octopetala L., Vaccinium gaultherioides Bigelow (V. uliginosum L.) (Table S1). The differences in the altitudinal maxima of the subalpine and alpine plants between the Carpathians and Pyrenees surely result from the higher elevations of the latter. The Carpathians arc is composed predominantly of medium-sized mountain ridges, with only three or four massifs revealing sufficiently well, and several other with only fragmentary developed alpine vegetation belt [47,48]. Inversely, in the Pyrenees, this type of vegetation is more frequent and cover a broader area [28,49,50,51].
Kalmia procumbens is well adapted to micro-habitats with continental climatic conditions, to the extremely high diary amplitude of temperatures during vegetation season [52,53] and early snowmelt [54], but the species could suffer from the frost during the beginning of the vegetation season, when development of the generative structures starts [55]. On the other hand, the late frost disturbances and high temperatures in the exposed places of K. procumbens occurrence are reducing quite all other plant species, promoting Kalmia successful regeneration [55]. In relation to the surrounding grass plant communities, for which the Index of Continentality is 19.7 or 16.5, after Gorchynsky and Rivas-Martinez, respectively, in Loiseleurio-Cetrarietum of the East Carpathians it attains 21.6 and/or 17.5 [40]. Additionally, the average annual temperatures on the elevation of 1000 m in grass plant communities reach 2.6 °C, but in the rock coenoses dominated by K. procumbens attains 3.6 °C [19]. Nevertheless, K. procumbens does not reach its potential altitudinal maximum in most of the Carpathian ridges, as it was concluded for some other alpine shrubby plants in the East Carpathians [48].
3.2. Environmental conditions of K. procumbens realized niches
Kalmia procumbens is a calcifuge species occurring in the mountains composed of metamorphic siliceous rocks pH [28,56,57,58]. The plant communities with dominance of this species are classified as association Cetrario nivalis-Loiseleurietum procumbentis Br.-Bl. in Br.-Bl. et Jenny 1926 [59], from alliance Rhododendro-Vaccinion Br.-Bl. 1926). In the Pyrenees, the plant community Cetrario nivalis-Loiseleurietum is developed on the North-exposed, acid sites where the winds blow-out snow reducing snow cover [49,50,57,58,60]. Similar plant community is formed in the South Carpathians [61,62], and fragmentary in the East Carpathians [38,56]. In the latter mountains and in the South Carpathians, K. procumbens occurs in the grassland communities on the siliceous rocks [56,63,64].
The climate in the regions of K. procumbens occurrence in the mountains of the Central Europe is of oceanic to sub-continental type, cryo-oro-temperate termotype and sub-humid to hyper-humid ombrotype [65,66]. Despite this, the average bioclimatic data retrieved from World Clim for K. procumbens localities in the Pyrenees revealed slightly milder conditions, than in the Carpathians (Table 2). The climate of the Carpathian localities of. K. procumbens appeared more continental with lower factors of temperature and precipitation and higher temperature seasonality. The continental climate of the steppes easterly and southerly from the Carpathians and closer distance to the continental climate of the central Euro-Asiatic continent could play some role in lowering positions of K. procumbens there, and alpine and sub-alpine vegetation belt, comparing to the Pyrenees.
The average bioclimatic factors of K. procumbens localities presented a low level of differences between the Central and Eastern Pyrenees, revealed mostly in the lower values of factors connected with precipitation in the central, more continental parts of this mountain chain (Figure 3). Nevertheless, the more humid climatic conditions in the Eastern Pyrenees can be a reason of more dispersed and not so abundant localities of the species due to prolonged snow cover [28]. Similarly, the more Atlantic climate conditions in the Northern than in the Southern Pyrenees [65,66] can explain a lower abundance and frequency of occurrence of K. procumbens on the southern macro-slopes [28]. On the other hand, the higher and longer lasting of snow observed in the Eastern Pyrenees could reduce the potential habitats accessible for K. procumbens, as the species occurrence is connected mostly with the places with thin snow deposits [21,22,28,38,54]. Kalmia procumbens occurs mostly on specific microhabitats, mainly the rocky ridges and rocks, where the snow is being blown away and temperatures reveal high diurnal amplitudes.
The bioclimatic differences between localities of K. procumbens in the East versus South Carpathians appeared a higher than between East and Central Pyrenees (Figure 3). This finding could result from the higher elevations of the South Carpathians mountain massifs than those in the East Carpathians, and consequently, the greater number of K. procumbens populations reported from the higher elevations, which are characterized with lower temperatures and higher mean diurnal amplitude and isotermality.
The snow-free period appeared important for micro-sites inhabited by small ericaceous shrubs in the high mountains and in the arctic zone [67]. The longer snow-free period positively influenced the wood-ring increment in the Empetrum hermaphroditum Hagerup [68], the species frequently occurring with K. procumbens. At the same time, less snowfall and shorter duration of snow cover observed during the last decade were a reason for more abundant K. procumbens growth [21,69].
The differences of climatic conditions of K. procumbens current localities in the Pyrenees and Carpathians could result from (1) adaptation to different climates during the Holocene (2), origin of the Pyrenean and Carpathian populations from two different regions of the Arctic, the Atlantic in the Pyrenees and more continental in the Carpathians, or simultaneous action of both processes.
3.3. Possible influence of climate differences
It shall be expected, that detected differences between average climatic conditions of K. procumbens localities in the Carpathians and Pyrenees influenced genetic structure of the species. The isolation of K. procumbens populations between Pyrenees, Alps and Carpathians lasting at least during Holocene [4,70,71], should be a reason of genetic and morphological differences. The gene exchange in Ericaceae is limited due to the low rate of seed dispersal [72] and restricted pollen transport, especially between populations from distant mountain chains [73]. These limitations could be a reasons for genetic and morphological differences, as described for other subalpine/alpine plant species, Salix herbacea L. [74], Ranunculus glacialis L. [75], Saxifraga oppositifolia L. [76], Soldanella alpina L. [77] or Rhododendron ferrugineum L. [78,79], and even a reason of speciacion, as in case of Rhododendron ferrugineum and R. myrtifolim Schott & Kotschy [80].
The world-wide genetic structure of K. procumbens detected using sequences of multiple nuclear loci revealed that southernmost European and East-Asiatic populations are genetically similar but different from the Arctic ones [81,82]. The southern populations of K. procumbens in the mountains diverged from the Arctic during the LG [82,83]. This is in contrast with the results of the amplified fragment length polymorphism (AFLP) analyses, which indicated the isolation of Central-European mountain clade [84]. The number of verified populations and individuals used in these papers [82,84] was sufficient for description of K. procumbens general pattern of geographic differentiation but was rather too small to its genetic differentiation in the Central-European mountains. It could be expected, the species reveal differences between Pyrenean, Alpine and Carpathian localities, this hypothesis, however, should be verified in a specific study.
3.4. Future ecological niches in the Carpathians and Pyrenees
The climate change caused reduction of the potential niches of K. procumbens at their southern limit of realized ecological niches in the mountains of Central Europe could be predicted and even expected, similarly as in case of other subalpine and alpine plants [10,28,29,38,41,48]. In that context, the drastic reduction of potential niches in the Carpathians in general and quite complete disappearance of suitable niches in the East Carpathians is not surprising. The process of reduction of the geographic ranges of cold adapted, high-mountain plants and their shifts to the higher elevations first of all is restricted by mountain highest elevations. The extinction could also results from rather slow uppermost colonization by the plants [85]. The East Carpathians have not the massifs crossing an elevation of 2500 m, which are the main centres of occurrence of alpine and subalpine plants [47,86]. It shall be stressed, that process of disappearance of area of potential niches detected in case of K. procumbens, in fact concerns more or less the entire alpine and subalpine flora in the East Carpathians [24,48].
In the Pyrenees, the reduction of potential niches area suitable for K. procumbens is less drastic than in the Carpathians. It results from the more ‘alpine’ character of the Pyrenees and the higher elevations of their highest peaks, which form the conditions for the occurrence of alpine flora [28,50]. The more intense reduction of potential niches suitable for K. procumbens in the Central than in the East Pyrenees could be explained by more continental climate of the Central Pyrenees, influenced by the close distance to the very dry and warm central part of the Ebro Basin from the South. This could reduce the influence of the Atlantic climate [58,66] (66-Rivas-Martínez et al. 2017; 58-Ninot et al. 2017). In fact, the westernmost localities of the species were detected on the northern macroslopes of the Pyrenees, with the more prominent impact of the Atlantic climate (Figure 1). This could also corroborate the amphi-Atlantic biogeographic character of K. procumbens proposed by Hultén [25] (1973).
The influence of the reduction of the area of potential niches to the currently realized niches of K. procumbens could be diminished by presence of the microrefugia suitable for the species within the area of its occurrence [87] (87-Hülber et al. 2016). The species in fact settled in this kind of microsites at their lowermost localities. The current localities of K. procumbens in the East Carpathians (Figure 1), however, exist in the area of very low level of the potential niches, especially at low elevations, which are quite exclusively on the North-facing slopes. It could be expected, that K. procumbens can exist for a long time outside the optimal environmental conditions on the northern slopes, which make the extinction following the climate change more prolonged, especially for the plants moderately sensitive to the lack of humidity [88] (88-Maclean et al., 2015) and to heat [31] (31-Buchner et al., 2013). Thus, the persistence of K. procumbens in the environments at a very low level of suitability could result from the specific site conditions of the particular localities eliminating competition of the other plants, but it also could be caused by mykorrhiza. The symbiosis of K. procumbens with fungi [89,90] (89-Treu et al., 1996; 90-Koizumi & Nara, 2017) could mitigate the extinction rate, as the plants with the fungal symbiosis are less prone to the harsh environmental conditions [91,92] (91-Haselwandter, 1979; 92-Cripps & Eddington, 2005).
The extinction rate of K. procumbens due to the climate change could also be restricted by the shrub longevity, found as more than 100 years [93] (93-Schweingruber & Poschlod 2005). The longevity with tolerance to the harsh environment, in particular relatively high temperatures, the high diurnal temperature amplitudes, winter frosts [24,52,53,94,95,96] (52-53-Larcher & Wagner, 2009, 2011; 94-Scherrer & Körner, 2011; 95-Zeidler & Banaš, 2013; 96-García et al. 2020; 24-Löffler & Pape, 2020), episodic summer frosts [97] (97-Kuprian et al. 2014), allow moderate the climate change influence. The K. procumbens plants covered with shallow snow stratum are less vulnerable to the spring frosts [98] (98-Palacio et al., 2015). From the other hand, the speed of shifting of K. procumbens in the mountains after glacier regression could be rather moderate. In the Alps, it exists together with other ericaceous shrubs, in the areas with relatively stabilized plant cover, and colonize new terrain about a century after the glacier regression [5] (5-Fischer et al. 2019). Our results suggest that cooler slopes may act as microrefugia, buffering the effects on plant communities of increases in temperature by delaying the extinctions of species with low temperature requirements [88] (88-Maclean et al., 2015).
The precipitation of the driest month (bio 14) and precipitation of the warmest quarter (bio 18) are the most influential to the present realized niche of K. procumbens in the East Carpathians in Ukraine. These bioclimatic factors acts as limitation by lack of water in the vegetation period, which determine position of K. procumbens out of the zone of acceptable risk when the average annual temperature rises by 2 °C, and deep into the zone of catastrophic risk when temperature rises by 3 °C. The role of other ecofactors is much lower and concerns only the acidity and salinity of soils, which are stable in the rocks on the localities of K. procumbens.
4. Materials and Methods
4.1. Study areas
The Pyrenees and Carpathians have been elevated during alpine orogenesis and currently conserve the alpine floras with an abundance of endemic species [4,48,86,99,100]. The Pyrenees includes several massifs reaching elevation of more than 3000 m, with subalpine and alpine vegetation belts harbouring several arctic-alpine plants [28,50,51]. Kalmia procumbens occupies elevated parts of the eastern and central Pyrenees [28,34].
In comparison to the Pyrenees, the Carpathians cover a broader area, are more fragmented and divided into several mountain chains, but only a few of them sufficiently high the subalpine and alpine vegetation zones could be developed [47,48,86,101,102]. Localities of K. procumbens are occupying the most elevated sites in the South and East Carpathians [33,103], with only one natural locality in the West Carpathians [104,105,106].
4.2. Data sampling and geographic analyses
Data on the natural localities of the species were extracted from Global Biodiversity Information Facility (GBIF) database, the literature, herbaria, and authors’ field notes. The geographic coordinates of localities were determined using Google Earth when not reported in the original data. Totally, we gathered more than 2000 data but after verification and exclusion of duplicates we analysed 641 georeferenced data, 140 for the Carpathians and 501 for the Pyrenees. Maps of the distribution of K. procumbens in the Pyrenees and Carpathians were prepared using QGIS 3.16.4 “Hannover” [107]. Altitudinal ranges of the species in both mountains system were presented on the graphs.
4.3. Environmental variables
The temperature, precipitation and elevation determined at the highest degree current (realized) ecological niches of species [108]. In spite of that, we used nineteen bioclimatic variables [109] and altitude (Table 1) to find factors, which determine current potential niche of K. procumbens. Usage of all these data could shed new light on the adaptation of the species to specific climatic data, retrieved from WorldClim (WC) database (http://worldclim.org/) [110]. For LGM (21 ka BP) we used PaleoClim (PC) data (http://www.paleoclim.org/), which is based on the CHELSA algorithm on PMIP3 data [111,112]. The spatial resolution of 30 arc-seconds (~1 km) of climate variables was applied. For the current climate (average for the years 1970-2000), we used WorldClim 2.1 database [110] bioclimatic data. For the future climate, we based on the Community Climate System Model (CCSM) [113] and used two representative concentration pathways (RCPs), RCP 2.6 and RCP 8.5 [114]. The RCP 2.6 provided increase of radiation forcing by 2.6 W/m2 and an increase of temperature by 1 °C before 2070 (average for 2061-2080), the RCP 8.5 by 8.5 W/m2 and 2 °C during the same period. Both are climate projections from GCMs, that were downscaled and calibrated using WorldClim 1.4 as baseline climate.
The PC used climate data of CAPE project [115] and of Community Climate System Model (CCSM) [113] for delineation of potential niches during Eemian Interglacial (LIG, 120-140 ka BP). These climatic data are based on geomorphological and geographical characteristics and do not take into account edaphic features and the structure of the substrate, which somewhat changes the microclimatic conditions.
For retrospective analyses of climate of EM (LGM, 21 ka BP), the CHELSA algorithm on PMIP3 data were used. For Mid-Holocene (MH) climate (ca 6 ka BP) the CCSM4 were used. For the current climate (average for the years 1970-2000), we used WorldClim 2.1 database [110] bioclimatic data. The previsions of the future climate changes were utilized scenarios of two representative concentration pathways (RCPs), RCP 2.6 and RCP 8.5 [114]. The RCP 2.6 provided increase of radiation forcing by 2.6 W/m2 and increase of temperature by 1 °C before 2070 (average for 2061-2080), the RCP 8.5 by 8.5 W/m2 and 2 °C during the same period. Both are climate projections from GCMs, that were downscaled and calibrated using WorldClim 1.4 as baseline climate.
The average values of bioclimatic variables were compared between the Carpathians and Pyrenees, as well as between regions within these mountain ranges. The Mann-Whitney U test conducted in the R environment was used for this purpose [116]. The influence of particular climatic variables on the current potential niches of K. procumbens in the Carpathians and in the Pyrenees was verified by Principal component analysis (PCA). In PCA, the data for localities in the North Carpathians and South Carpathians, as well as for Central and East Pyrenees were treated as separate groups.
4.4. Niche modeling
For the prediction of the potential range of K. procumbens, bioclimatic data related to their localities were used. The MaxEnt 3.4.1. [117,118,119] was applied in analyses with maximum entropy modelling for the estimation of a probable distribution of the species outside their realized niche. The model with ENMeval R software [120] for the current climate was evaluated at first. The procedure of evaluation follows those described by Salva-Catarineu et al. [121]. For evaluation of the results of modelling, the Receiver Operating Characteristic (ROC) curves were used [122,123], assessing the values of Area Under the Curve (AUC) below 0.6 as nearly random.
The QGIS 3.16.4 “Hannover” [107] was applied for mapping the current and predicted potential niches on the climate variables. The potential distribution of K. procumbens species was calculated for the different classes of suitability [45,121].
The phytoindication method was used for the assessment of econiches, indicators of the leading climatic and edaphic ecofactors, forecasting their changes depending on the climate for the Eastern Carpathians [40,124]. Modeling of econiche change and assessment of habitat loss threats for populations of the Eastern Carpathians was performed on the basis of phytoindication data. For this purpose, the point values of the amplitude (x±2σ) were calculated for the leading ecofactors, and their changes were evaluated depending on the increase in average annual temperatures by 1, 2, and 3 °C. The acceptable risk zone is, when the average values of the obtained data are intside the confidence intervals of the ±2σ, and a catastrophic risk zone when the amplitudes did not overlap, which means the complete disappearance of the species from this place [40].
5. Conclusions
Kalmia procumbens occurs in the alpine and subalpine vegetation belts, going down to the elevations of about 1500-1600 m, while the most elevated localities in the Pyrenees are at ca 3000 m, about 500 m higher than in the Carpathians. The localities of K. procumbens in the Carpathians have the more continental climate than in the Pyrenees, with lower precipitation and temperatures but higher seasonality of temperature and precipitation. Due to climate warming, the strong reduction of potential area of occurrence to 2100 is expected in the Carpathians and moderate reduction in in the Pyrenees.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Provided potential range of Kalmia procumbens during Last Glacial Maximum in the Carpathians (a and b) estimated using: a – environmental conditions from the East Carpathians, b – from the South Carpathians; in the Pyrenees (c and d) using: c – conditions from the East Pyrenees, d – from the Central Pyrenees; Figure S2: Occurene of populations of Kalmia procumbens in different exposition in the Carpathians (a and b): a – East Carpathians, b –South Carpathians; and in the Pyrenees (c and d): c –East Pyrenees, d –Central Pyrenees; Table S1: Altitudinal maxima of subalpine and alpine species in the Carpathians [48,103,104] and Pyrenees [28]; CEUR – Central European mountain, ARALP – Arctic-Alpine; EUROS – Euro-Siberian mountain.
Author Contributions
Conceptualization, Ł.W. and A.B.; methodology, Ł.W., M.P. and A.B.; validation, M.M., Á.R. and L.T.; formal analysis, A.B; investigation, M.M., Ł.W. and Y.P.; resources, M.M. and A.B.; data curation, M.M. and A.B.; writing—original draft preparation, A.B; writing—review and editing, Ł.W., A.B, L.T. and Y.P; visualization, Ł.W.; supervision, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by Institute of Dendrology Polish Academy of Sciences under statutory activity.
Acknowledgments
The Institute of Dendrology Polish Academy of Sciences in Kórnik, Kazimierz Wielki University in Bydgoszcz, Botanical Institute of Spanish Research Council in Barcelona, Ivan Franko National University in Lviv and M.G. Kholodny Institute of Botany National Academy of Ukraine in Kyiv are acknowledged for help in providing authors in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hultén, E. Outline of the history of arctic and boreal biota during the quarternary period. Bokförlags aktiebolaget Thule: Stockholm, 1937.Author 1, A.B.; Author 2, C.D. Title of the article. Abbreviated Journal Name Year, Volume, page range.
- Billings, W.D. Adaptations and origins of alpine plants. Arctic Alpine Res. 1974, 6, 129–142. [Google Scholar] [CrossRef]
- Hagen, O.; Vaterlaus, L.; Albouy, C.; Brown, A.; Leugger, F.; Onstein, R.E.; Novaes de Santana, C.; Scotese, C.R.; Pellissier, L. Mountain building, climate cooling and the richness of cold-adapted plants in the Northern Hemisphere. J. Biogeogr. 2019, 46, 1792–1807. [Google Scholar] [CrossRef]
- Schmitt, T. Molecular biogeography of the high mountain systems of Europe: An overview. In: . In High mountain conservation in a changing world; Catalan, J., Ninot, J.M., Mercè Aniz, M., Eds.; Adv. Glob. Change Res. 2017, 62, 63–74.
- Fischer, A.; Fickert, T.; Schwaizer, G.; Patzelt, G.; Groß, G. 2019. Vegetation dynamics in Alpine glacier forelands tackled from space. Sci. Rep. 2019, 9, 13918. [Google Scholar] [CrossRef] [PubMed]
- Huntley, B.; Birks, H.J.B. An atlas of past and present pollen maps for Europe: 0-13000 years ago. Cambridge, Cambridge University Press: Cambridge, U.K., 1983.
- Ralska-Jasiewiczowa, M.; Latałowa, M.; Wasylikowa, K.; Tobolski, K.; Madeyska, E.; Wright, H.E.; Turner C. Late Glacial and Holocene history of vegetation in Poland based on isopollen maps. W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2004.
- Stivrins, N.; Soininen, J.; Amon, L.; Fontana, S.L.; Gryguc, G.; Heikkilä, M.; Heiri, O.; Kisielienè, D.; Reitalu, T.; Stančikaitè, M.; Veski, S.; Seppä, H. Biotic turnover rates during the Pleistocene-Holocene transition. Quat. Sci. Rev. 2016, 151: 100–110.
- Kornaś, J., Medwecka-Kornaś, A.. Geografia roślin. Państwowe Wydawnictwo Naukowe: Warszawa, 2002.
- Habel, J.C.; Assmann, T. Relict Species. Phylogeography and Conservation Biology. Springer: Berlin, Heidelberg, Germany, 2010.
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Nat. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef]
- Lesica, P. Arctic-Alpine plants decline over two decades in Glacier National Park, Montana, U. S.A., AAAR 2014, 46, 327–332. [Google Scholar] [CrossRef]
- Rixen, C, Wipf S. ; Frei, E.; Stöckli, V. Faster, higher, more? Past, present and future dynamics of alpine and arctic flora under climate change. Alpine Bot. 2014, 124, 77–79. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; García-Calvo, L.; García, P.; Acebes, J.L. Anticipating extinctions of glacial relict populations in mountain refugia. Biol. Conserv. 2016, 201, 243–251. [Google Scholar] [CrossRef]
- Lesica, P.; Crone, E.E. Arctic and boreal plant species decline at their southern range limits in the Rocky Mountains. Ecol. Let. 2017, 20, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Rixen, C.; Wipf, S. Non-equilibrium in alpine plant assemblages: shifts in Europe’s summit floras. In High mountain conservation in a changing world; Catalan, J., Ninot, J.M., Mercè Aniz, M., Eds.; Adv. Glob. Change Res. 2017, 62, 285–303.
- Pauli, A.; Halloy, S.R.P. High mountain ecosystems under climate change. Oxford Research Encyclopedia fo Climate science. 2019. [CrossRef]
- Fragnière, Y.; Pittet, L.; Clément, B.; Bétrisey, S.; Gerber, E.; Ronikier, M.; Parisod, C.; Kozlowski, G. Climate change and alpine screes: No future for glacial relict Papaver occidentale (Papaveraceae) in Western Prealps. Diversity 2020, 12, 346. [Google Scholar] [CrossRef]
- Didukh, Y.P.; Chorney, I.I.; Budzhak, V.V.; Tokaryuk, A.I.; Kish, R.YA.; Protopopova, V.V.; Shevera, M.V.; Kozak, O.M.; Kontar, I.S.; Rozenblit, Yu.V.; Norenko, K.V. Climatogenic changes of plant life of the Ukrainian Carpathians. DrukArt: Chernivtsi, Ukraine, 2016.
- Trivedi, M.R.; Morecroft, M.D.; Berry, P.M.; Dawson, T.P. Potential effects of climate change on plant communities in three montane nature reserves in Scotland, UK. Biol. Conserv. 2008, 141, 1665–1675. [Google Scholar] [CrossRef]
- Kobiv, Y. Response of rare alpine plant species to climate change in the Ukrainian Carpathians. Folia Geobot. 2017, 52, 217–226. [Google Scholar] [CrossRef]
- Kobiv, Y. Trends in population size of rare plant species in the alpine habitats of the Ukrainian Carpathians under climate change. Diversity 2018, 10, 62. [Google Scholar] [CrossRef]
- Leunda, M.; González-Sampériz, P.; Gil-Romera, G.; Bartolomé, M.; Belmonte-Ribas, Á.; Gómez-García, D.; Kaltenrieder, P.; Rubiales, J.M.; Schwörer, C.; Tinner, W.; Morales-Molino, C.; Sancho, C. Ice cave reveals environmental forcing of long-term Pyrenean tree line dynamics. J. Ecol. 2018, 107, 814–828. [Google Scholar] [CrossRef]
- Löffler, J.; Pape, R. Thermal niche predictors of alpine plant species. Ecology 2020, 101, e02891. [Google Scholar] [CrossRef]
- Hultén, E. The amphi-Atlantic plants and their phytogeographic connections. Koeltz: Königstein, Germany, 1973.
- Meusel, H.; Jäger, E.; Rauschert, S.; Weinert, E. 1978. Vergleichende chorologie der zentraleuropäischen flora. Fischer: Jena, Germany, 1978, Volume 2.
- Hultén, E.; Fries, M. Atlas of North European vascular plants, 1-3. Koelz: Königstein, Germany, 1986.
- Gómez García, D.; Ferrández Palacio, J.V.; Bernal Gálvez, M.; Campo González, A.; López Retamero, J.R.; Ezquerra Rivas, V. Plantas de las cumbres del Pirineo. Instituto Pirenaicode Ecologia: Jaca, Spain; Prames: Zaragoza, Spain, 2020. [Google Scholar]
- Abeli, T.; Vamosi, J.C.; Orsenigo, S. The importance of marginal population hotspots of cold-adapted species for research on climate change and conservation. J. Biogeogr. 2018, 45, 977–985. [Google Scholar] [CrossRef]
- Dítě, D.; Hájek, M.; Svitková, I.; Košuthová, A.; Šoltés, R.; Kliment, J. Glacial-relict symptoms in the Western Carpathian flora. Folia Geobot. 2018, 53, 277–300. [Google Scholar] [CrossRef]
- Buchner, O.; Karadar, M.; Bauer, I.; Neuner, G. A novel system for in situ determination of heat tolerance of plants: first results on alpine dwarf shrubs. Plant Methods 2013, 9, 7. [Google Scholar] [CrossRef]
- Chopik, V.I. Vysokohirna flora Ukrains’kykh Karpat. Naukova Dumka: Kyiv, Ukraine, 1976.
- Boratyński, A.; Didukh, Ya.P. Loiseleuria procumbens (Ericaceae) in the Ukrainian Carpathians. Dendrobiology 2002, 47, 3–8. [Google Scholar]
- Boratyński, A.; Romo, A. Loiseleuria procumbens (Ericaceae) in the Spanish Pyrenees. Acta Soc. Bot. Pol. 2003, 72, 125–133. [Google Scholar] [CrossRef]
- Graebherr, G.; Gottfried, M.; Pauli, H. Climate effects on mountain plants. Nature 1994, 369, 448. [Google Scholar] [CrossRef]
- Körner, C. Alpine plant life: functional plant ecology of high mountain ecosystems. Springer: Berlin 1999.
- Körner, C. The green cover of mountains in a changing environment. In Global change and mountain regions; Huber, U.M., Bugmann, K.M., Reasoner, M.A., Eds.; Springer, Dordrecht, The Netherlands, 2005, Advences in global change research 23, 367–376.
- Cherepanyn, R.M. Effect of climate changes on the habitat of rare arctic-alpine plant species in the high mountain part of the Ukrainian Carpathians. Biol. Stud. 2018, 12, 73–86. [Google Scholar] [CrossRef]
- Grime, J.P.; Price, S. The evolutionary strategies that shape ecosystems. John Wiley & Sons: West Sussex, UK, 2012.
- Didukh, Y.P. World of plants of Ukraine in aspect of the climate change. Naukova Dumka: Kyiv, Ukraine, 2023.
- Pellissier, L.; Eidesen, P.B.; Ehrich, D.; Descombes, P.; Schönswetter, P.; Tribsch, A.; Westergaard, K.B.; Alvarez, N.; Guisan, A.; Zimmermann, N.E.; Normand, S.; Vittoz, P.; Luoto, M.; Damgaard, C.; Brochmann, C.; Wisz, M.S.; Alsos, I.G. 2015. Past climate-driven range shifts and population genetic diversity in arctic plants. J. Biogeogr. 2015, 43, 461–470. [Google Scholar] [CrossRef]
- Wada, N.; Shimmono, M.; Miyamoto, M.; Kojima, S. Warming effects on shoot developmental growth production in sympatric evergreen alpine dwarf shrubs Empetrum nigrum and Loiseleuria procumbens. Ecol. Res. 2002, 17, 125–132. [Google Scholar] [CrossRef]
- Rodríguez-Sanchéz, F.; Arroyo, J. Reconstructing the demise of Tethyan plants: Climate-driven range dynamics of Laurus since the Pliocene. Glob. Ecol. Biogeogr. 2008, 17, 685–695. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Fløjgaard, C.; Marske, K.A.; Nógues-Bravo, D.; Normand, S. Applications of species distribution modelling to paleobiology. Quat. Sci. Rev. 2011, 30, 2930–2947. [Google Scholar] [CrossRef]
- Walas, Ł.; Sobierajska, K.; Ok, T.; Dönmez, A.A.; Kanoğlu, S.S.; Bou Dagher-Kharrat, M.; Douaihy, B.; Romo, A.; Stephan, J.; Jasińska, A.K.; Boratyński, A. Past, present and future geographic range of the oro-Mediterranean Tertiary relict: Juniperus drupacea case study. Reg. Environ. Change 2019, 19, 5, SI: 1507–1520. [Google Scholar] [CrossRef]
- Zapałowicz, H. Roślinna szata gór Pokucko-Marmaroskich. Sprawozdanie Komisyi Fizyjograficznej 24. Akademia Umiejętności: Kraków, Poland, 1889.
- Mráz, P.; Ronikier, M. 2016. Biogeography of the Carpathians: evolutionary and spatial facets of biodiversity. Biol. J. Linn. Soc. 2016, 119, 528–559. [Google Scholar] [CrossRef]
- Tasenkevich, L.; Boratyński, A.; Skrypec, K.; Seniv, M.; Khmil, T.; Walas, Ł. Biodiversity of high-mountain woody plants in the East Carpathians in Ukraine. Dendrobiology 2023, 89, 1–19. [Google Scholar] [CrossRef]
- Braun-Blanquet, L. La végétation alpine des Pyrénées Olrientales, étude de phytosociologie compare. Consejo Superior de Investigaciones Científicas: Barcelona, Spain, 1948.
- Villar, L.; Sesé, J.A.; Ferrández, J.V. Atlas de la flora del Pirineo Aragonés. Consejo de Protección de la Naturaleza de Aragón e Instituto de Estudios Altoaragoneses: Huesca, Spain, 2001, Volume 2.
- Gómez García, D.; Ferrández Palacio, J.V.; Tejero, P.; Font, X. Spatial distribution and environmental analysis of the alpine flora in the Pyrenees. Pirineos 2017, 172: e027.
- Larcher, W.; Wagner, J. High mountain bioclimate: temperatures near the ground recorded from the timberline to the nival zone in the Central Alps. Contributions to Natural History Museum, Bern 2009, 12, 857–874. [Google Scholar]
- Larcher, W.; Wagner, J. Temperatures in the life zones of the Tyrolean Alps. Sitzungsberichte Abteilung 1 (2010) 2011, 213, 31–51. [Google Scholar] [CrossRef]
- Wipf, S.; Stoeckli, V.; Bebi, P. Winter climate change in alpine tundra: plant responses to changes in snow depth and snowmelt timing. Clim. Change 2009, 94, 105–121. [Google Scholar] [CrossRef]
- Ladinig, U.; Hacker, J.; Neuner, G.; Wagner, J. How endangered is sexual reproduction of high-mountain plants by summer frosts? Frost resistance, frequency of frost events and risk assessment. Oecologia 2013, 171, 743–760. [Google Scholar] [CrossRef] [PubMed]
- Tasenkevich, L. Flora of the Carpathians: checklist of the native vascular plant species. State Museum of Natural History National Academy of Sciences of Ukraine: Lviv, Ukraine, 1998.
- Ninot, J.M.; Carrillo, E.; Font, X.; Carreras, J.; Ferré, A.; Masalles, R.M.; Soriano, I.; Vigo, J. Altitude zonation in the Pyrenees. A geobotanic interpretation. Phytocoenologia 2007, 37, 371–398. [Google Scholar] [CrossRef]
- Ninot, J.M.; Carrillo, E.; Ferré. A. The Pyrenees. In The vegetation of the Iberian Peninsula; Loidi, J., Ed.; Springer: Cham, Switzerland, 2017, Plant and Vegetation 12, 323–366.
- HabRef v7. Available online: https://inpn.mnhn.fr/habitat/cd_hab/23233/tab/description (accessed on 17 July 2023).
- Baudière, A.; Serve, L. Les landes rases à Loiseleuria procumbens en Pyrénées Orientales et leur intérêt phytogéographique. Coll. Phytosoc. 1975, 337–347. [Google Scholar]
- Biţă-Nicolae, C.D. The Natural Priority Habitats in the Alpine zone of Bucegi Massif Romanian Southern Carpathians). Bot. Serbica 2011, 35, 79–85. [Google Scholar]
- Stancu, D.I. The main alpine and subalpine habitats in Râiosu & Buda mountains, Fǎgǎraş massif. Muzeul Olteniei Craiova. Oltenia. Studii şi comunicǎri. Ştiinţele Naturii 2013, 29, 118–126. [Google Scholar]
- Puşcaş, M.; Gafta, D.; Cristea, V. Analyse éco-coenotique des prairies édifiées par Carex curvala All. des Carpates Roumaines, Acta Bot. Gallica 2005, 152, 497–505. [Google Scholar] [CrossRef]
- Kricsfalusy, V.V. Mountain grasslands of high conservation value in the Eastern Carpathians: syntaxonomy, biodiversity, protection and management. Thaiszia 2013, 23, 67–112. [Google Scholar]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A. World-wide bioclimatic classification system. Glob. Geobot. 2011, 1: 1-634 + maps.
- Rivas-Martínez, S.; Penas, Á.; del Rió., S.; Díaz González, T.E.; Rivas-Sáenz, S. Bioclimatology of the Iberian Peninsula and the Balearic Islands. In The vegetation of the Iberian Peninsula; Loidi J. Ed.; Springer: Cham, Switzerland, 2017, Plant and Vegetation 12, 29–80.
- Dobrowski, S.Z. A climatic basis for micro-refugia: the influence of terrain on climate. Global Change Biol. 2011, 17: 1022–1035.
- Bär, A.; Pape, R.; Bräuning, A.; Löffler, J. 2008. Growth-ring variations of dwarf shrubs reflect regional climate signals in alpine environments rather than topoclimatic differences. J. Biogeogr. 2008, 35, 625–636. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.B.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef]
- Nagy, L.; Grabherr, G.; Körner, C.; Thompson, D.B.A. Alpine biodiversity in space and time. In Alpine Biodiversity in Europe; Nagy, L., Grabherr, G., Körner, C., Thompson, D.B.A., Eds.; Springer: Berlin, Germany, 2003; pp. 453–464. [Google Scholar]
- Skrede, I.; Eidesen, P.B.; Piñeiro Portela, R.; Brochmann, C. Refugia, differentiation and postglacial migration in arctic-alpine Eurasia, exemplified by the mountain avens (Dryas octopetala L.). Mol. Ecol. 2006, 15, 1827–1840. [Google Scholar] [CrossRef]
- Stephenson, C.M.; Kohn, D.D.; Park, K.J.; Atkinson, R.; Edwards, C.; Travis, J.M. Testing mechanistic models of seed dispersal for the invasive Rhododendron ponticum (L.). Persp. Pl. Ecol., Evol. Syst. 2007, 9, 15–28. [Google Scholar] [CrossRef]
- Escaravage, N.; Wagner, J. Pollination effectiveness and pollen dispersal in a Rhododendron ferrugineum (Ericaceae) population. Pl. Biol. 2004, 6, 606–615. [Google Scholar] [CrossRef]
- Alsos, I.G.; Alm, T.; Normand, S.; Brochmann, C. Past and future range shifts and loss of diversity in dwarf willow (Salix herbacea L.) inferred from genetics, fossils and modelling. Glob. Ecol. Biogeogr. 2009, 18, 223–239. [Google Scholar] [CrossRef]
- Ronikier, M.; Schneeweiss, G.M.; Schönswetter, P. The extreme disjunction between Beringia and Europe in Ranunculus glacialis s. l. (Ranunculaceae) does not coincide with the deepest genetic split – a story of the importance of temperate mountain ranges in arctic–alpine phylogeography. Mol. Ecol. 2012, 21, 5561–5578. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.; Tribsch, A.; Schneeweiss, G.M.; Brodbeck, S.; Gugerli, F.; Holderegger, R.; Abbott, R.J.; Schönswetter, P. Tales of the unexpected: Phylogeography of the arctic-alpine model plant Saxifraga oppositifolia (Saxifragaceae) revisited. Mol. Ecol. 2012, 21, 4618–4630. [Google Scholar] [CrossRef] [PubMed]
- Kropf, M.; Comes, H.P.; Kadereit, J.W. Past, present and future of mountain species of the French Massif Central – the case of Soldanella alpina L. subsp. alpina (Primulaceae) and a review of other plant and animal studies. J. Biogeogr. 2012, 39, 799–812. [Google Scholar] [CrossRef]
- Suchan, T.; Malicki, M.; Ronikier, M. Relict populations and Central European glacial refugia: The case of Rhododendron ferrugineum (Ericaceae). J. Biogeogr. 2019, 46, 392–404. [Google Scholar] [CrossRef]
- Lewandowska, A.; Boratyńska, K.; Marcysiak, K.; Gómez, D.; Romo, A.; Malicki, M.; Iszkuło, G.; Boratyński, A. Phenotypic differentiation of Rhododendron ferrugineum populations in European mountains. Dendrobiology 2022, 87, 1–12. [Google Scholar] [CrossRef]
- Lewndowska, A.; Marcysiak, K.; Gómez, D.; Jasińska, A.K.; Romo, A.; Didukh, Ya.; Sękiewicz, K.; Boratyńska, K.; Boratyński, A. Inference of taxonomic relationships between Rhododendron ferrugineum and R. myrtifolium (Ericaceae) from leaf and fruit morphologies. Bot. J. Linn. Soc. 2023, 201, 483–497. [Google Scholar] [CrossRef]
- Ikeda, H.; Senni, K.; Fujii, N.; Setoguchi, H. High mountains of the Japanese archipelago as refugia for arctic–alpine plants: phylogeography of Loiseleuria procumbens (L.) Desvaux (Ericaceae). Biol. J. Linn. Soc. 2009, 97, 403–412. [Google Scholar] [CrossRef]
- Ikeda, H.; Eidesen, P.B.; Yakubov, V.; Barkalov, V.; Brochmann, C.; Setoguchi, H. Late Pleistocene origin of the entire circumarctic range of the arctic-alpine plant Kalmia procumbens. Mol. Ecol. 2017, 26, 5773–5783. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Carlsen, T.; Fujii, N.; Brochmann, C.; Setoguchi, H. Pleistocene climatic oscillations and the speciation history of an alpine endemic and a widespread arctic-alpine plant. New Phytol. 2012, 194, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Eidesen, P.B.; Ehrich, D.; Bakkestuen, V.; Alsos, I.G.; Gilg, O.; Taberlet, P.; Brochmann, C. Genetic roadmap of the Arctic: Plant dispersal highways, traffic barriers and capitals of diversity. New Phytol. 2013, 200, 898–910. [Google Scholar] [CrossRef]
- Pykälä, J. 2016. Relation between extinction and assisted colonization of plants in the arctic-alpine and boreal regions. Conserv. Biol. 2016, 31, 524–530. [Google Scholar] [CrossRef]
- Ronikier, M. Biogeography of high-mountain plants in the Carpathians: an emerging phylogeographical perspective. Taxon 2011, 60, 373–389. [Google Scholar] [CrossRef]
- Hülber, K.; Wessely, J.; Gattringer, A.; Moser, D.; Kuttner, M.; Essl, F.; Leitner, M.; Winkler, M.; Ertl, S.; Willner, W.; Kleinbauer, I.; Sauberer, N.; Mang, T.; Zimmermann, N.E.; Dullinger, S. Uncertainty in predicting range dynamics of endemic alpine plants under climate warming. Glob. Change Biol. 2016, 22, 2608–2619. [Google Scholar] [CrossRef]
- Maclean, I.M.D.; Hopkins, J.J.; Bennie, J.; Lawson, C.R.; Wilson, R.J. Microclimates buffer plant community response to climate change. Global Ecol. Biogeogr. 2015, 24, 1340–1350. [Google Scholar] [CrossRef]
- Treu, R.; Laursen, G.A.; Stephenson, S.L.; Landolt, J.C.; Densmore, R. Mycorrhizae from Denali National Park and Preserve, Alaska. Mycorrhiza 1996, 6, 21–29. [Google Scholar] [CrossRef]
- Koizumi, T.; Nara, K. Communities of putative Ericoid Mycorrhizal Fungi isolated from alpine dwarf shrubs in Japan: Effects of host identity and microhabitat. Microbes Envir. 2017, 32, 147–153. [Google Scholar] [CrossRef]
- Haselwandter, K. Mycorrhizal status of ericaceous plants in alpine and subalpine areas. New Phytol. 1979, 83, 427–431. [Google Scholar] [CrossRef]
- Cripps, C.L.; Eddington, L.H. Distribution of mycorrhizal types among alpine vascular plant families on the Beartooth Plateau, Rocky Mountains, U.S.A., in reference to large-scale patterns in Arctic–Alpine habitats. AAAR. 2005, 37(2), 177–188.
- Schweingruber, F.; Poschlod, P. Growth rings in herbs and shrubs: lifespan, age determination and stem anatomy. For. Snow Landsc. Res. 2005, 79, 195–415. [Google Scholar]
- Scherrer, D. Körner, C. Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Zeidler, M.; Banaš, M. Vybrané kapitoly z ekologie horských ekosystémů. Univerzita Palackého v Olomouci, Přírodovědecká Fakulta, Olomouc, Slovakia, 2013.
- García, M.B.; Domingo, D.; Pizarro, M.; Font, X.; Gómez-García, D.; Ehrlén, J. Rocky habitats as microclimatic refuges for biodiversity. A close-up thermal spproach. Env. Exp. Bot. 2020, 170, 103886. [Google Scholar] [CrossRef]
- Kuprian, E.; Briceño, V.F.; Wagner, J.; Neuner, G. Ice barriers promote supercooling and prevent frost injury in reproductive buds, flowers and fruits of alpine dwarf shrubs throughout the summer. Environ. Exper. Bot. 2014, 106, 4–12. [Google Scholar] [CrossRef]
- Palacio, S.; Lenz, A.; Wipf, S.; Hoch, G.; Rixen, C. Bud freezing resistance in alpine shrubs across snow depth gradients. Envir. Exper. Bot. 2015, 118: 95–101.
- Tasenkevich, L. Endemism in the Carpathian flora – a chorological aspect. In Geobotanist and Taxonomist. A volume dedicated to Professor Adam Zając on the 70th anniversary of his birth; Zemanek, B., Ed.; Istitute of Botany of Jagellonian University: Kraków, Poland, 2011; pp. 157–168. [Google Scholar]
- Breman. E.; Hurdu, B.I.; Kliment, J.; Kobiv, Y.; Kučera, J.; Mráz, P.; Pușcaș, M.; Renaud, J.; Ronikier, M.; Šibík, J.; Schmotzer, A.; Štubňová, E.; Szatmari, P.M.; Tasenkevich, L.; Turis, P.; Slovák, M. Conserving the endemic for a of the Carpathian Region: an international project to increase and share knowledge of the distribution, evolution and taxonomy of Carpathian endemics and to conserve endangered species. Pl. Syst. Evol. 2020, 306, 15–59.
- Margielewski, W. Geological and geomorphological settings. In Postglacial history of vegetation in the Polish part of the Western Carpathians based on isopollen map; Obidowicz, A., Madeyska, E., Turner, C., Eds.; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2013; pp. 11–14. [Google Scholar]
- Mirek, Z. Altitudinal vegetation belts of the Western Carpathians. In Postglacial history of vegetation in the Polish part of the Western Carpathians based on isopollen maps Obidowicz, A., Madeyska, E., Turner, C. Eds.; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2013, pp. 15–21.
- Ţopa, E. Ericaceae. In Flora Republicii Popularae Romîne; Sãvulescu, T. Ed.. Academia Republicii Populare Romîne: Bucureşti, Romania, 1960, Volume 7, pp. 119–140.
- Jasičová, M. Ericaceae Juss. In Flóra Slovenska; Futák J., Bertová L. Eds.; Veda, Bratislava, Slovakia, 1982, Volume 3, 337–359.
- Turis, P.; Kliment, J.; Feráková, V.; Dítě, D.; Eliáš, P.; Hrivnák, R.; Košťál, J.; Šuvada, R.; Mráz, P.; Bernátová, D. Red List of vascular plants of the Carpathian part of Slovakia. Thaiszia 2014, 24, 35–87. [Google Scholar]
- Mirek, Z. High mountain vascular plants of the Carpathians. Atlas of distribution. W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2020.
- QGIS.org. (2022). QGIS geographic information system. QGIS Association. Available online: http://www.qgis.org.
- Bradie, L.; Leung, B. A quantitative synthesis of the importance of variables used in Maxent species distribution models. J. Biogeogr. 2017, 44, 1344–1361. [Google Scholar] [CrossRef]
- O’Donnell, M.S.; Ignizio, D.A. Bioclimatic predictors for supporting ecological applications in the conterminous United States. U.S. Geological Survey Data Series 2012, 691, 10. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Intern. J. Clim. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Brown, J.L.; Hill, D.J.; Dolan, A.M.; Carnaval, A.C.; Haywood, A.M. PaleoClim, high spatial resolution paleoclimate surfaces for global land areas. Nature, Sci. Data 2018, 5, 180–254. [Google Scholar] [CrossRef]
- Karger, D.N.; Nobis, M.P.; Normand, S.; Graham, C.H.; Zimmermann, N.E. CHELSA-TraCE21k v1. 0. Downscaled transient temperature and precipitation data since the last glacial maximum. Clim. Past. 2021, 1–27. [Google Scholar]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; Worley, P.H.; Yang, Z.-L.; Zhang, M. (2011). The Community Climate System Model Version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J.; Johns, T.; Krinner, G.; Shongwe, M.; Tebaldi, C.; Weaver, A.J.; Wehner, M. (2013). Long-term Climate Change: Projections, Commitments and Irreversibility. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, pp. 1029–1136.
- Otto-Bliesner, B.L.; Marshall, S.J.; Overpeck, J.T.; Miller, G.H.; Hu, A. Simulating arctic climate warmth and ice field retreat in the Last Interglaciation. Science 2006, 311, 1751–1753. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 20 March 2022).
- Phillips, S.J. A brief tutorial on Maxent 2017. Available online: http://biodiversi ty informatics.amnh.org/open_source/maxe t/ (accessed on 15 March 2020).
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. . Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent software for modeling species niches and distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 15 March 2020).
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. (2014). ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods in Ecology and Evolutions, 5, 1198–1205. [CrossRef]
- Salvà-Catarineu, M.; Romo, A.; Mazur, M.; Zielińska, M.; Minissale, P.; Dönmez, A.A.; Boratyńska, K.; Boratyński, A. Past, present and future geographic range of the relict Mediterranean and Macaronesian Juniperus phoenicea complex. Ecol. Evol. 2021, 11, 5075–5095. [Google Scholar] [CrossRef]
- Wang, Z.; Chang, Y.C.I.; Ying, Z.; Zhu, L.; Yang, Y. A parsimonious threshold-independent protein feature selection method through the area under receiver operating characteristic curve. Bioinformatics 2007, 23, 2788–2794. [Google Scholar] [CrossRef]
- Mas, J.-F.; Soares Filho, B.; Pontius, R.G.; Farfán Gutiérrez, M.; Rodrigues, H. A suite of tools for ROC analysis of spatial models. ISPRS Intern. J. Geo-Inform. 2013, 2, 869–887. [Google Scholar] [CrossRef]
- Didukh, Y. Climate change assessment based on synphytoindication method. In Handbook of Climate Change Mitigation and Adaptation, 3rd ed.; Lackner, M., Sajjadi, B., Chen, W.-Y., Eds.; Springer: Heidelberg, Switzerland, 2022; pp. 2759–2814. [Google Scholar]
Figure 1.
Geographical distribution of Kalmia procumbens on the basis of georeferenced data in the Carpathians (a): blue dots – East Carpathians, red dots – South Carpathians; and in the Pyrenees (b): blue dots –West Pyrenees, red dots – East Pyrenees.
Figure 1.
Geographical distribution of Kalmia procumbens on the basis of georeferenced data in the Carpathians (a): blue dots – East Carpathians, red dots – South Carpathians; and in the Pyrenees (b): blue dots –West Pyrenees, red dots – East Pyrenees.
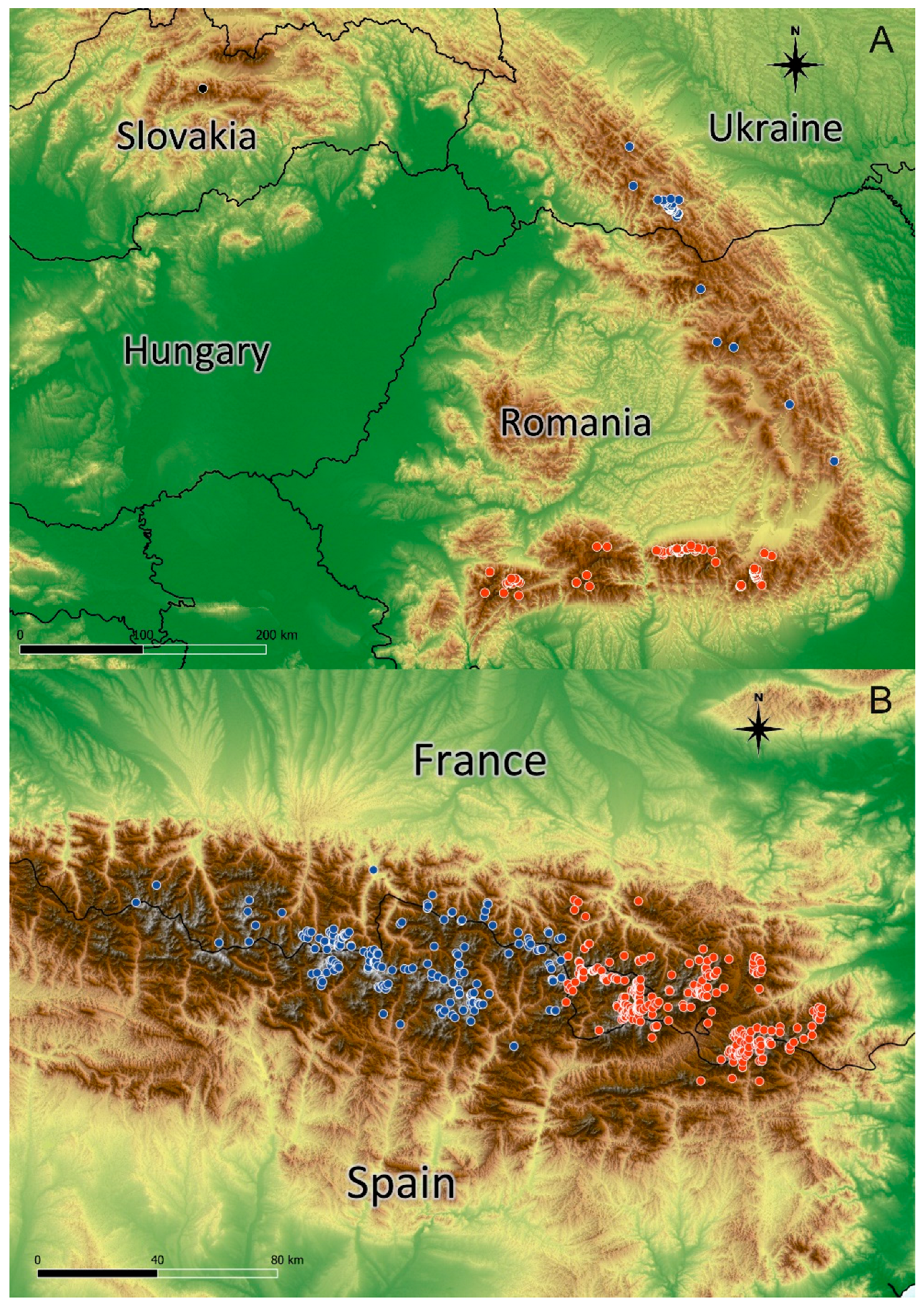
Figure 2.
Vertical distribution of Kalmia procumbens localities in the Pyrenees: CPYR – Central and EPYR – East Pyrenees and in Carpathians SCARP – South and ECARP – East Carpathians.
Figure 2.
Vertical distribution of Kalmia procumbens localities in the Pyrenees: CPYR – Central and EPYR – East Pyrenees and in Carpathians SCARP – South and ECARP – East Carpathians.
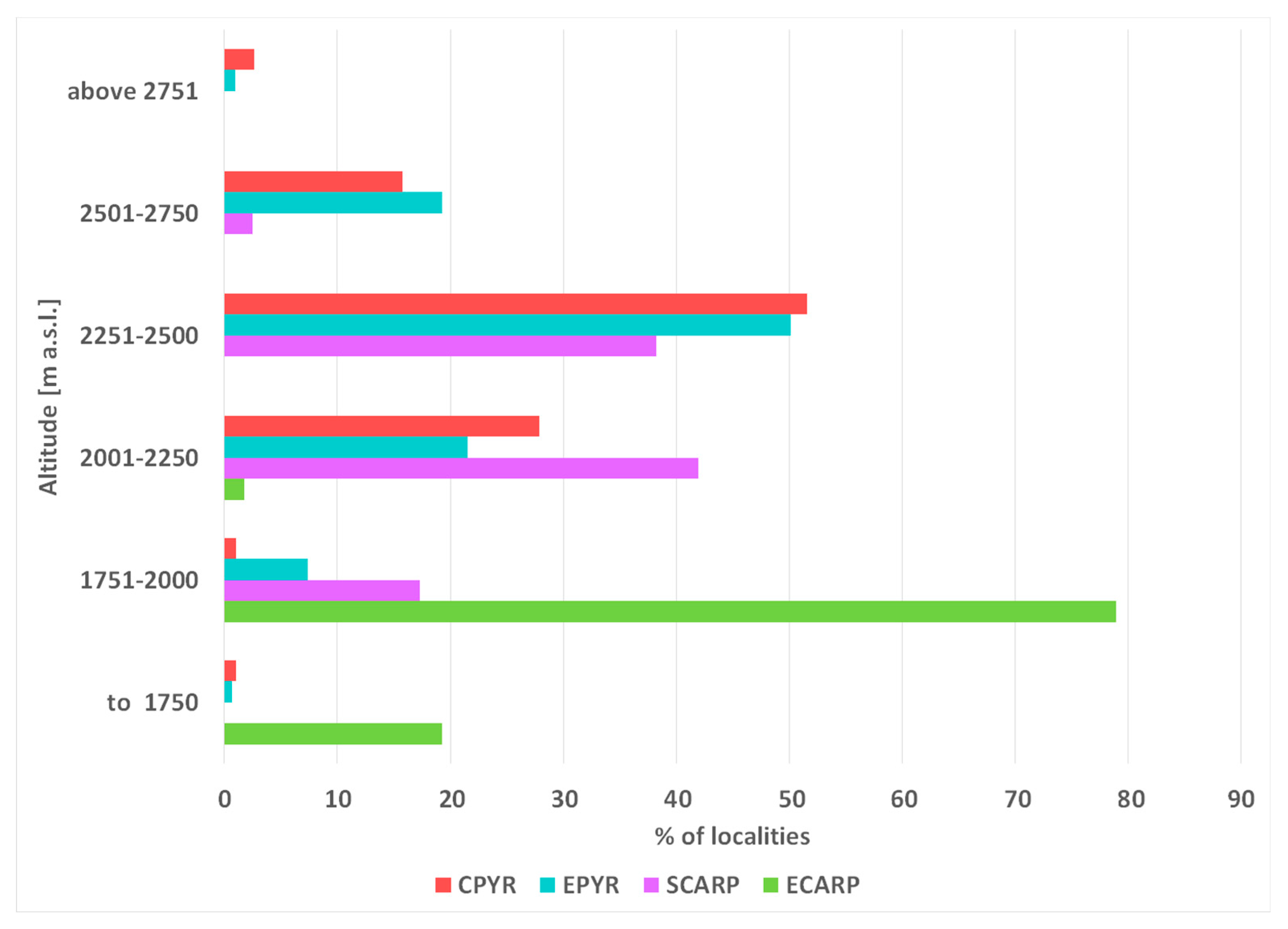
Figure 3.
Position of localities of Kalmia procumbens from Central Pyrenees (CPYR), East Pyrenees (EPYR), East Carpathians (ECARP) and South Carpatians (SCARP) in PCA on the basis of bioclimatic variables (acronyms as in Table 1); ellipses indicate the 95% confidence intervals.
Figure 3.
Position of localities of Kalmia procumbens from Central Pyrenees (CPYR), East Pyrenees (EPYR), East Carpathians (ECARP) and South Carpatians (SCARP) in PCA on the basis of bioclimatic variables (acronyms as in Table 1); ellipses indicate the 95% confidence intervals.
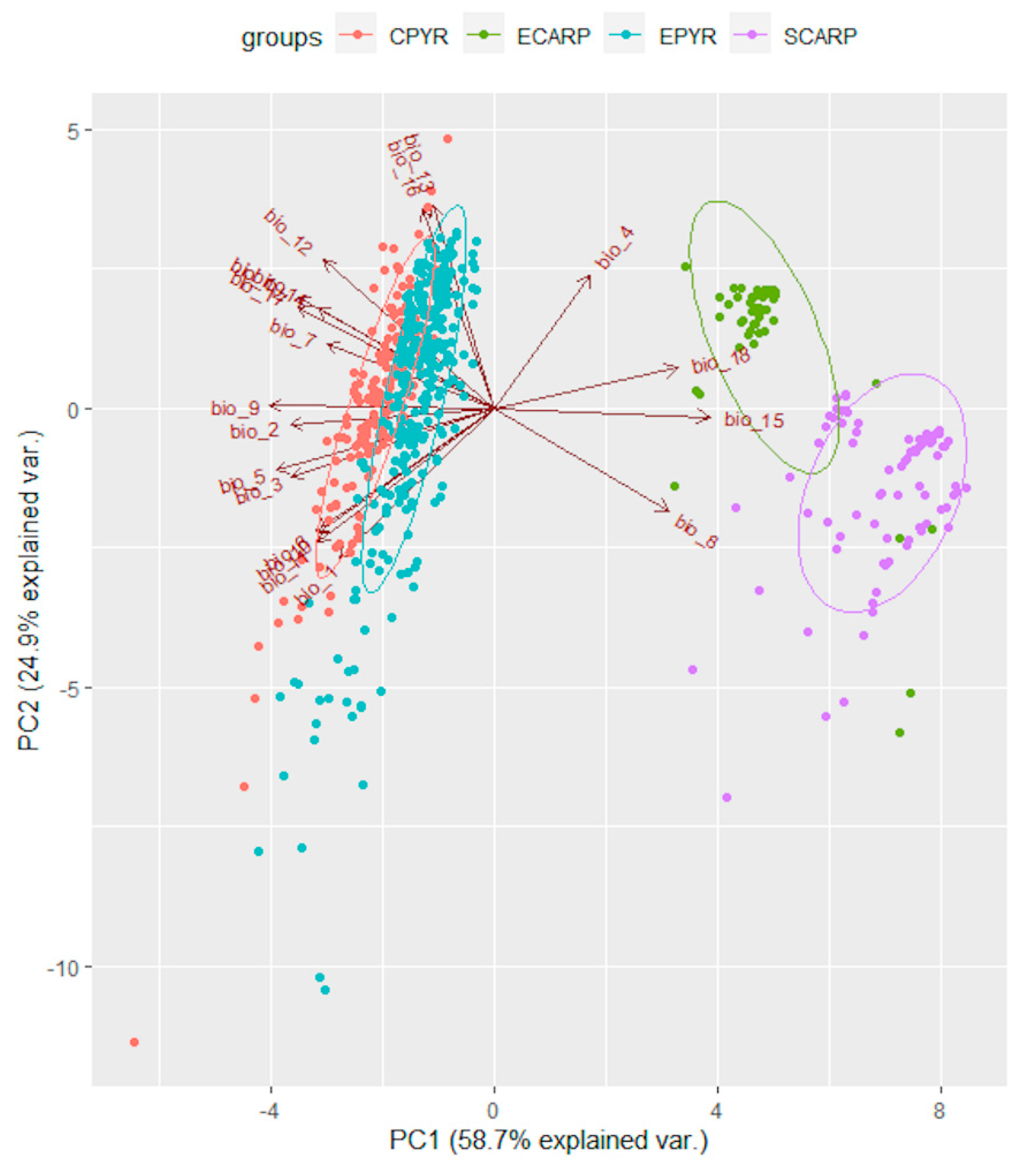
Figure 4.
Current potential range of Kalmia procumbens in the Carpathians (a and b) estimated using: a – environmental conditions from the East Carpathians, b – from the South Carpathians; in the Pyrenees (c and d) using: c – conditions from the East Pyrenees, d – from the Central Pyrenees.
Figure 4.
Current potential range of Kalmia procumbens in the Carpathians (a and b) estimated using: a – environmental conditions from the East Carpathians, b – from the South Carpathians; in the Pyrenees (c and d) using: c – conditions from the East Pyrenees, d – from the Central Pyrenees.
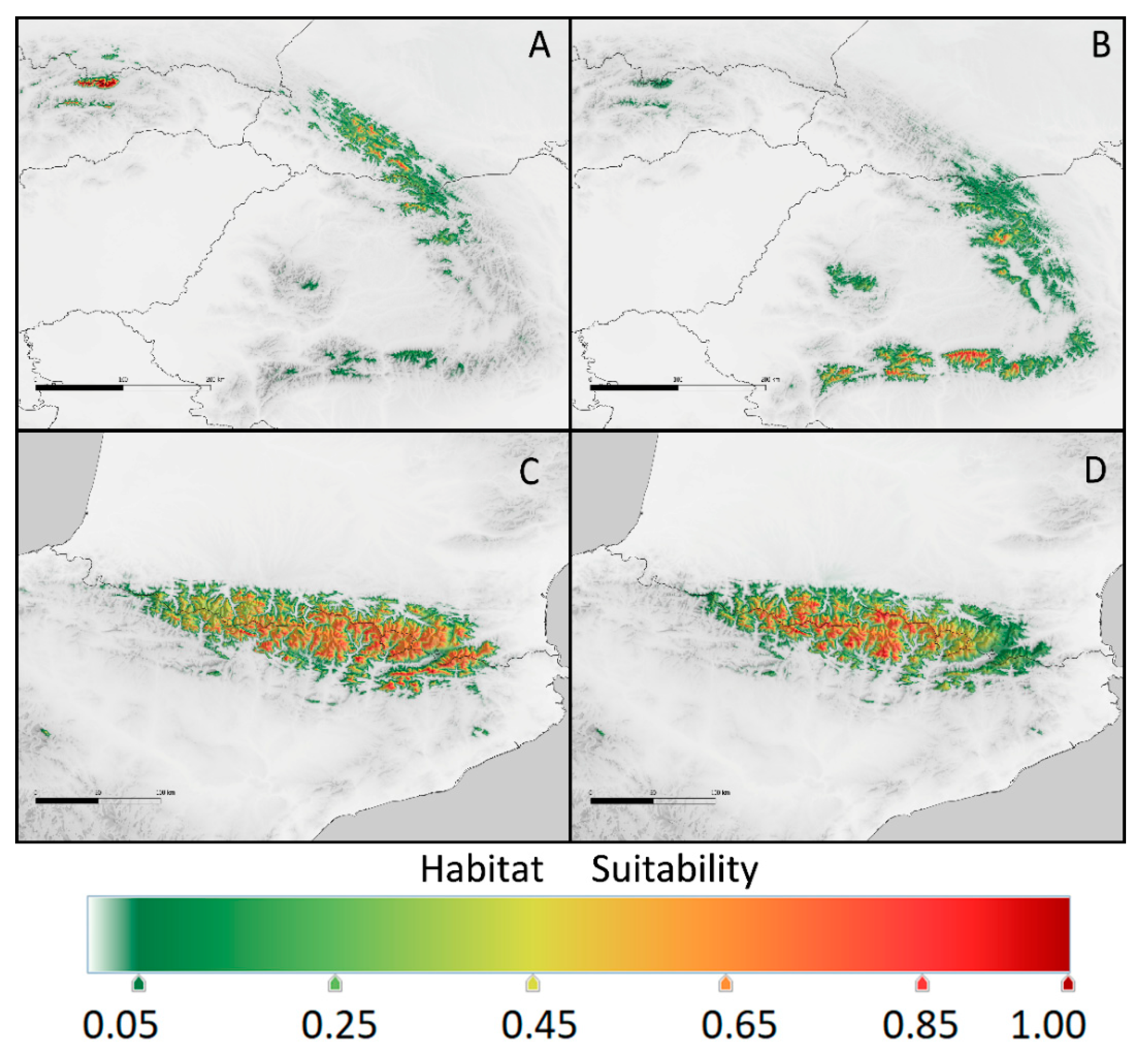
Figure 5.
Provided potential range of Kalmia procumbens in the Carpathians at 2100 estimated using current environmental conditions from East Carpathians: a – scenario RCP 2.6, b – scenario RCP 8.5, and from South Carpathians: c – scenario RCP 2.6, d – scenario RCP 8.5.
Figure 5.
Provided potential range of Kalmia procumbens in the Carpathians at 2100 estimated using current environmental conditions from East Carpathians: a – scenario RCP 2.6, b – scenario RCP 8.5, and from South Carpathians: c – scenario RCP 2.6, d – scenario RCP 8.5.
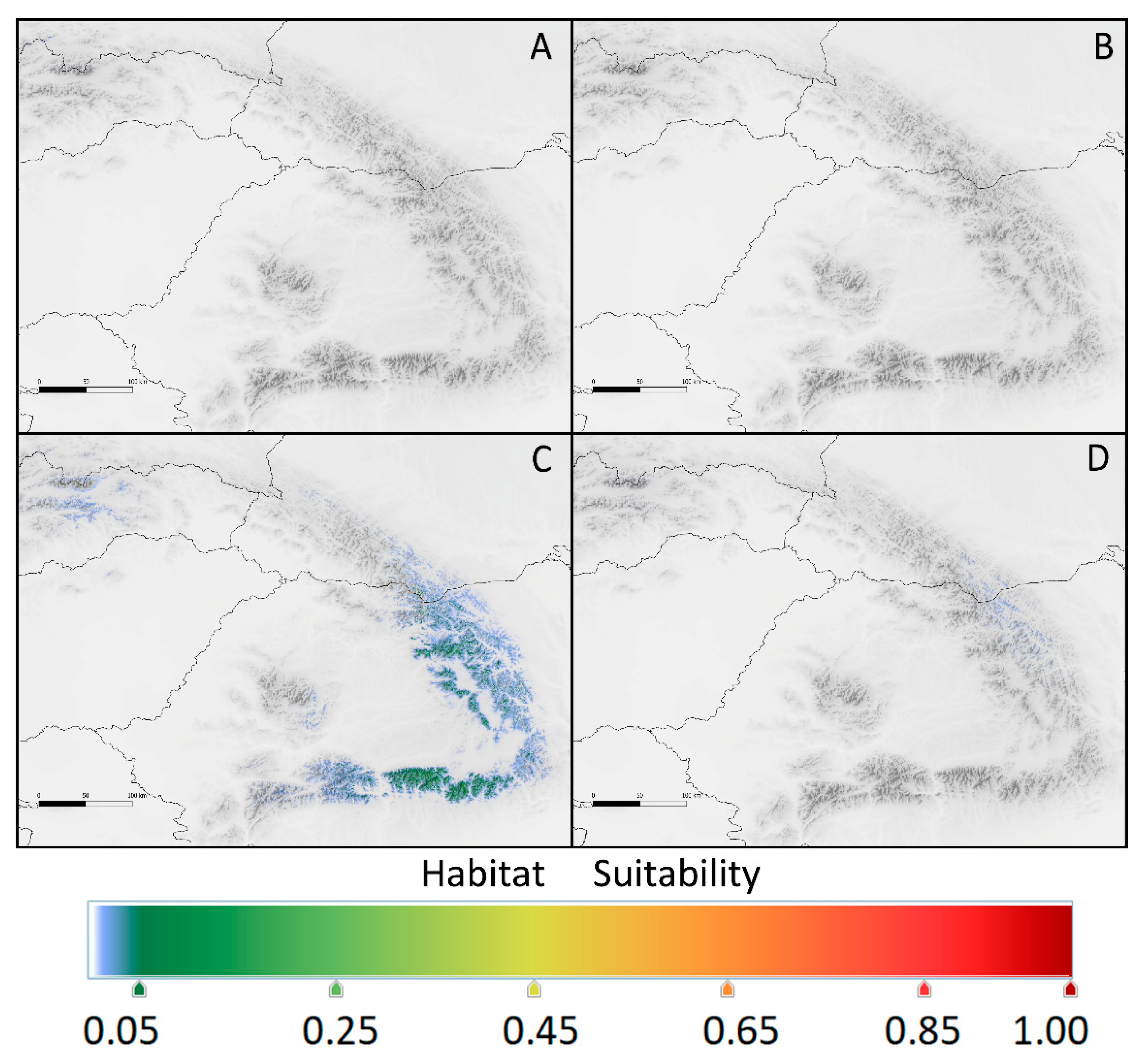
Figure 6.
Provided potential range of Kalmia procumbens in the Pyrenees at 2100 estimated using current environmental conditions from the East Pyrenees: a – scenario RCP 2.6, b – scenario RCP 8.5 and from the Central Pyrenees: c – scenario RCP 2.6, d – scenario RCP 8.5.
Figure 6.
Provided potential range of Kalmia procumbens in the Pyrenees at 2100 estimated using current environmental conditions from the East Pyrenees: a – scenario RCP 2.6, b – scenario RCP 8.5 and from the Central Pyrenees: c – scenario RCP 2.6, d – scenario RCP 8.5.
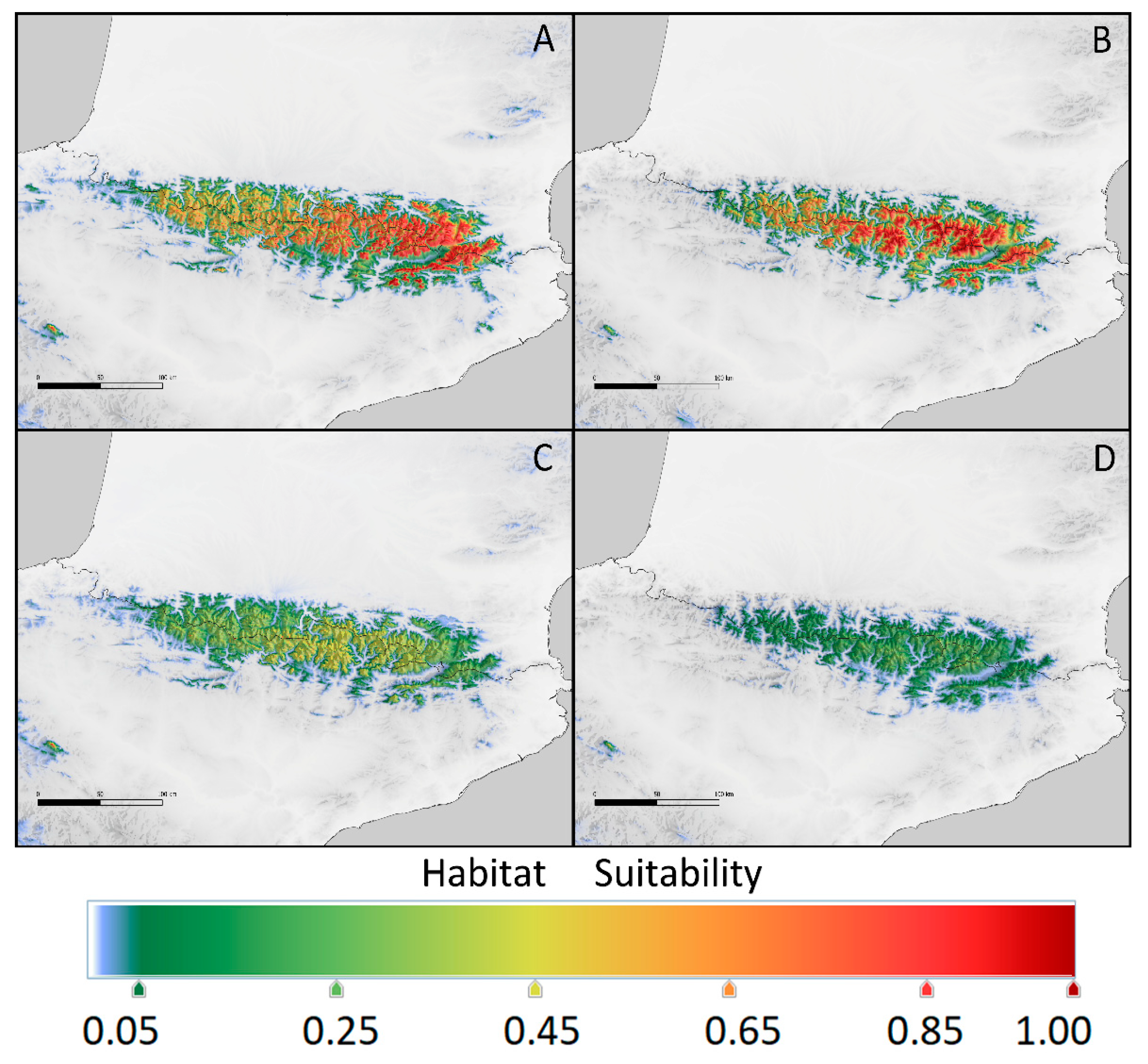
Figure 7.
Percentage values of econiche factors of Kalmia procumbens characterisation in the East Carpathians in Ukraine and potential changes depending on temperature changes: +1 °C (T+1), +2 °C (T+2) and +3 °C (T+3); Hd – soil moisture, Fh – variability of soil moisture, Rc – soil acidity, Sl – soil salinity, Ca – soil carbonate, Nt – content of mineral nitrogen available for assimilation, Ae – soil aeration, Tm – thermal regime, Om - ombro regime, Kn - continental climate, Cr - cryo regime, Lc – illumination; indicator values 2C+ - Х+2σ, 2C- Х-2σ; the range of indicators x±2σ marked in orange.
Figure 7.
Percentage values of econiche factors of Kalmia procumbens characterisation in the East Carpathians in Ukraine and potential changes depending on temperature changes: +1 °C (T+1), +2 °C (T+2) and +3 °C (T+3); Hd – soil moisture, Fh – variability of soil moisture, Rc – soil acidity, Sl – soil salinity, Ca – soil carbonate, Nt – content of mineral nitrogen available for assimilation, Ae – soil aeration, Tm – thermal regime, Om - ombro regime, Kn - continental climate, Cr - cryo regime, Lc – illumination; indicator values 2C+ - Х+2σ, 2C- Х-2σ; the range of indicators x±2σ marked in orange.
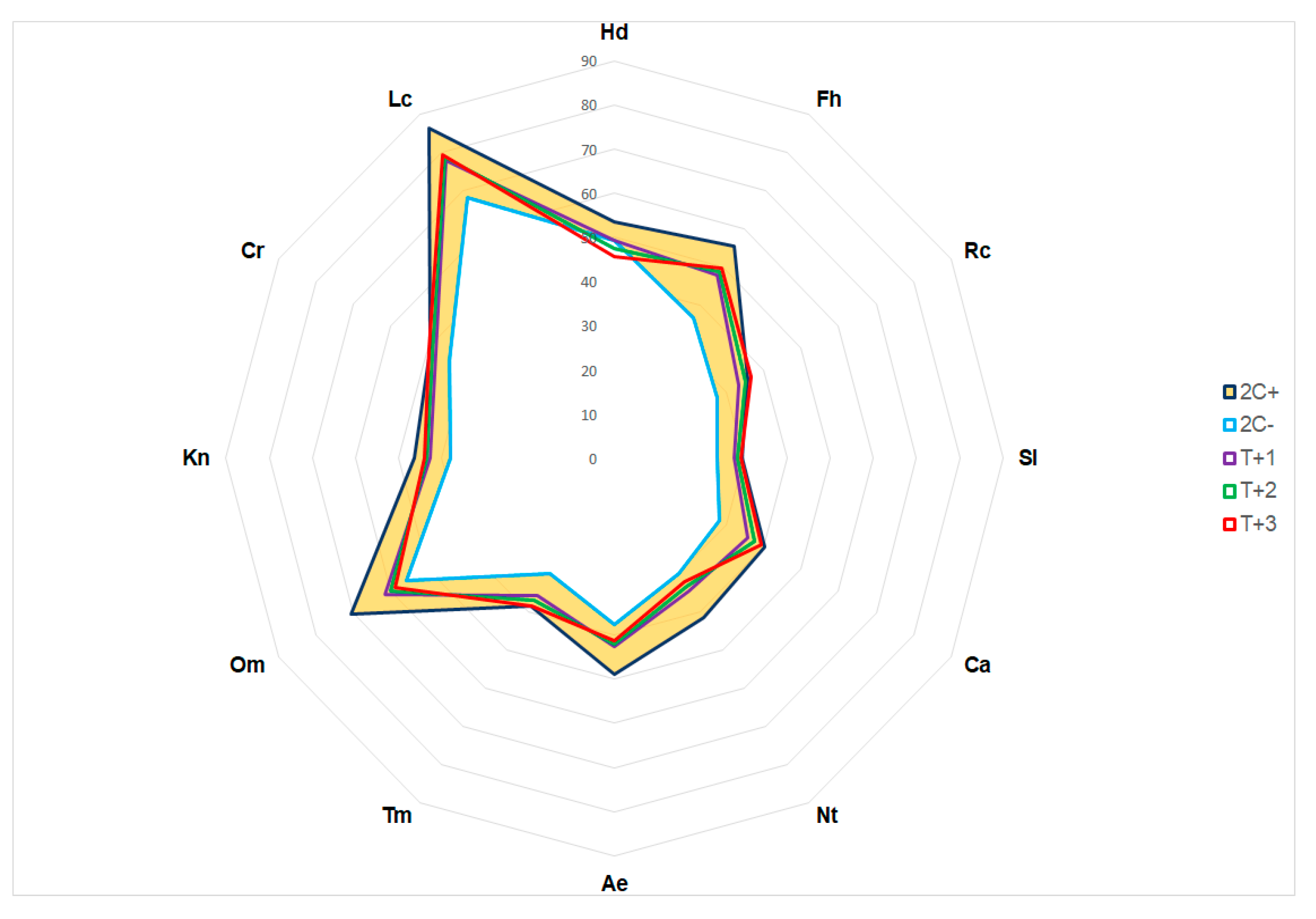

Table 1.
Contribution [%] of bioclimatic variables and altitude to the realized habitats suitable for Kalmia procumbens in the Pyrenees (PYR), Central Pyrenees (CPYR), East Pyrenees (EPYR and in the Carpathians (CARP), East Carpatians (ECARP) and South Carpathians (SCARP); values of 1.0 and higher bolded.
Table 1.
Contribution [%] of bioclimatic variables and altitude to the realized habitats suitable for Kalmia procumbens in the Pyrenees (PYR), Central Pyrenees (CPYR), East Pyrenees (EPYR and in the Carpathians (CARP), East Carpatians (ECARP) and South Carpathians (SCARP); values of 1.0 and higher bolded.
| Bioclimatic factor | CPYR | EPYR | PYR | ECARP | SCARP | CARP | |
| AUC | 0.994 | 0.991 | 0.993 | 0.999 | 0.997 | 0.998 | |
| Bio1 | Annual Mean Temperature | 0.1 | 0.1 | 0.1 | 2.0 | 0.3 | 1.3 |
| Bio2 | Mean Diurnal Range | 4.1 | 0.9 | 2.5 | 0.1 | 0.0 | 0.1 |
| Bio3 | Isothermality | 0.6 | 0.2 | 0.4 | 1.7 | 0.0 | 0.9 |
| Bio4 | Temperature Seasonality | 0.3 | 0.2 | 0.3 | 4.7 | 0.0 | 2.4 |
| Bio5 | Max Temperature of Warmest Month | 0.7 | 0.3 | 0.5 | 0.1 | 0.1 | 0.1 |
| Bio6 | Min Temperature of Coldest Month | 0.3 | 0.0 | 0.2 | 0.2 | 0.2 | 0.2 |
| Bio7 | Temperature Annual Range | 1.0 | 1.0 | 1.0 | 0.1 | 0.0 | 0.1 |
| Bio8 | Mean Temperature of Wettest Quarter | 0.4 | 2.0 | 1.2 | 0.2 | 0.2 | 0.2 |
| Bio9 | Mean Temperature of Driest Quarter | 3.8 | 6.0 | 4.9 | 5.5 | 8.2 | 6.9 |
| Bio10 | Mean Temperature of Warmest Quarter | 0.2 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 |
| Bio11 | Mean Temperature of Coldest Quarter | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 | 0.2 |
| Bio12 | Annual Precipitation | 0.6 | 0.2 | 0.4 | 0.1 | 0.0 | 0.1 |
| Bio13 | Precipitation of Wettest Month | 0.1 | 0.3 | 0.2 | 0.0 | 0.0 | 0.0 |
| Bio14 | Precipitation of Driest Month | 1.1 | 0.4 | 0.8 | 0.0 | 0.1 | 0.1 |
| Bio15 | Precipitation Seasonality | 0.3 | 0.1 | 0.2 | 3.4 | 10.0 | 6.7 |
| Bio16 | Precipitation of Wettest Quarter | 0.3 | 1.2 | 0.8 | 0.0 | 0.0 | 0.0 |
| Bio17 | Precipitation of Driest Quarter | 7.8 | 6.2 | 7.0 | 0.1 | 0.0 | 0.1 |
| Bio18 | Precipitation of Warmest Quarter | 0.1 | 0.1 | 0.1 | 31.1 | 9.9 | 20.5 |
| Bio19 | Precipitation of Coldest Quarter | 0.5 | 0.5 | 0.5 | 0.2 | 0.1 | 0.2 |
| Elevation | 77.6 | 80.2 | 78.9 | 50.1 | 70.4 | 60.3 |
Table 2.
Average values of bioclimatic variables in the studied regions. According to the Mann-Whitney test, all differences between the Pyrenees and the Carpathians (bolded) are significant (p<0.05), as are the differences between the Southern (SCARP) and Western (ECARP) Carpathians. For Eastern (EPYR) and Central (CPYR) Pyrenees, statistically significant differences (p<0.05) were observed for bio2, bio3, bio5, bio7, bio12, bio13, bio14, bio15, bio16, bio17, bio18 and bio19 (shaded).
Table 2.
Average values of bioclimatic variables in the studied regions. According to the Mann-Whitney test, all differences between the Pyrenees and the Carpathians (bolded) are significant (p<0.05), as are the differences between the Southern (SCARP) and Western (ECARP) Carpathians. For Eastern (EPYR) and Central (CPYR) Pyrenees, statistically significant differences (p<0.05) were observed for bio2, bio3, bio5, bio7, bio12, bio13, bio14, bio15, bio16, bio17, bio18 and bio19 (shaded).
| Bioclimatic factor | CPYR | EPYR | Pyrenees | ECARP | SCARP | Carpathians | |
| bio1 | Annual Mean Temperature | 2.37 | 2.40 | 2.39 | 1.07 | -0.25 | 0.30 |
| bio2 | Mean Diurnal Range | 9.27 | 8.67 | 8.90 | 6.91 | 7.16 | 7.06 |
| bio3 | Isothermality | 33.79 | 32.62 | 33.07 | 27.73 | 29.06 | 28.52 |
| bio4 | Temperature Seasonality | 649.33 | 646.56 | 647.61 | 684.48 | 657.93 | 668.81 |
| bio5 | Max Temperature of Warmest Month | 18.69 | 18.14 | 18.35 | 13.92 | 12.52 | 13.10 |
| bio6 | Min Temperature of Coldest Month | -8.73 | -8.44 | -8.55 | -10.99 | -12.08 | -11.62 |
| bio7 | Temperature Annual Range | 27.43 | 26.58 | 26.90 | 24.91 | 24.60 | 24.73 |
| bio8 | Mean Temperature of Wettest Quarter | -0.62 | -0.30 | -0.42 | 8.98 | 6.84 | 7.73 |
| bio9 | Mean Temperature of Driest Quarter | 10.80 | 10.78 | 10.79 | -6.81 | -8.02 | -7.51 |
| bio10 | Mean Temperature of Warmest Quarter | 10.93 | 10.89 | 10.90 | 9.36 | 7.83 | 8.46 |
| bio11 | Mean Temperature of Coldest Quarter | -4.71 | -4.60 | -4.64 | -7.19 | -8.14 | -7.74 |
| bio12 | Annual Precipitation | 1434.57 | 1447.54 | 1442.62 | 1274.90 | 934.12 | 1075.62 |
| bio13 | Precipitation of Wettest Month | 154.04 | 161.53 | 158.69 | 167.76 | 132.44 | 147.06 |
| bio14 | Precipitation of Driest Month | 75.71 | 68.17 | 71.03 | 65.56 | 46.75 | 54.58 |
| bio15 | Precipitation Seasonality | 20.19 | 23.53 | 22.27 | 32.36 | 39.56 | 36.57 |
| bio16 | Precipitation of Wettest Quarter | 430.82 | 457.02 | 447.08 | 462.96 | 363.02 | 404.52 |
| bio17 | Precipitation of Driest Quarter | 263.48 | 251.75 | 256.20 | 221.51 | 149.49 | 179.33 |
| bio18 | Precipitation of Warmest Quarter | 263.57 | 251.91 | 256.33 | 462.67 | 358.05 | 401.49 |
| bio19 | Precipitation of Coldest Quarter | 388.06 | 399.96 | 395.45 | 228.93 | 149.75 | 182.63 |
Table 3.
Contribution [%] of bioclimatic variables and altitude to models of future potential range of Kalmia procumbens in the Central Pyrenees (CPYR), East Pyrenees (EPYR), East Carpatians (ECARP) and South Carpathians (SCARP); values of 1.0% and higher bolded.
Table 3.
Contribution [%] of bioclimatic variables and altitude to models of future potential range of Kalmia procumbens in the Central Pyrenees (CPYR), East Pyrenees (EPYR), East Carpatians (ECARP) and South Carpathians (SCARP); values of 1.0% and higher bolded.
| Bioclimatic factor | CPYR | EPYR | ECARP | SCARP | |||||
| RCP 2.6 | RCP 8.5 | RCP 2.6 | RCP 8.5 | RCP 2.6 | RCP 8.5 | RCP 2.6 | RCP 8.5 | ||
| AUC | 0.994 | 0.994 | 0.991 | 0.991 | 0.999 | 0.999 | 0.997 | 0.997 | |
| Bio1 | Annual Mean Temperature | 0.0 | 0.1 | 0.0 | 0.0 | 1.5 | 1.1 | 0.6 | 0.4 |
| Bio2 | Mean Diurnal Range | 3.9 | 3.1 | 0.9 | 0.9 | 0.1 | 0.1 | 0.0 | 0.0 |
| Bio3 | Isothermality | 0.8 | 1.1 | 1.7 | 0.1 | 2.1 | 2.7 | 0.0 | 0.0 |
| Bio4 | Temperature Seasonality | 0.3 | 0.7 | 0.2 | 0.2 | 4.2 | 4.9 | 0.0 | 0.1 |
| Bio5 | Max Temperature of Warmest Month | 0.8 | 0.3 | 0.2 | 0.5 | 0.1 | 0.1 | 0.1 | 0.1 |
| Bio6 | Min Temperature of Coldest Month | 0.4 | 0.1 | 0.1 | 0.9 | 0.1 | 0.1 | 0.1 | 0.2 |
| Bio7 | Temperature Annual Range | 0.9 | 0.5 | 0.4 | 1.0 | 0.1 | 0.1 | 0.0 | 0.0 |
| Bio8 | Mean Temperature of Wettest Quarter | 1.2 | 0.7 | 0.4 | 4.0 | 0.0 | 0.1 | 0.1 | 0.2 |
| Bio9 | Mean Temperature of Driest Quarter | 4.3 | 5.5 | 6.4 | 4.0 | 9.5 | 3.8 | 6.6 | 5.9 |
| Bio10 | Mean Temperature of Warmest Quarter | 0.2 | 0.1 | 0.1 | 0.2 | 0.1 | 0.0 | 0.1 | 0.1 |
| Bio11 | Mean Temperature of Coldest Quarter | 0.1 | 0.0 | 0.1 | 0.1 | 0.3 | 0.3 | 0.3 | 0.2 |
| Bio12 | Annual Precipitation | 0.8 | 0.7 | 0.4 | 0.4 | 0.0 | 0.1 | 0.0 | 0.0 |
| Bio13 | Precipitation of Wettest Month | 0.3 | 0.2 | 0.1 | 1.1 | 0.0 | 0.0 | 0.1 | 0.0 |
| Bio14 | Precipitation of Driest Month | 1.0 | 1.0 | 0.4 | 0.5 | 0.0 | 0.1 | 0.0 | 0.1 |
| Bio15 | Precipitation Seasonality | 0.3 | 0.3 | 0.0 | 0.1 | 2.0 | 5.4 | 12.1 | 9.8 |
| Bio16 | Precipitation of Wettest Quarter | 0.5 | 0.3 | 0.2 | 0.3 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bio17 | Precipitation of Driest Quarter | 7.5 | 8.7 | 7.3 | 5.5 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bio18 | Precipitation of Warmest Quarter | 0.1 | 0.1 | 0.2 | 0.2 | 32.0 | 30.3 | 9.6 | 10.2 |
| Bio19 | Precipitation of Coldest Quarter | 0.4 | 0.2 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Elevation | 76.1 | 76.4 | 80.5 | 79.9 | 47.5 | 50.5 | 70.0 | 72.4 | |
Table 4.
Area of potential range according to the tested model and four sets of stands: CPYR – Central Pyrenees, EPYR – Eastern Pyrenees, ECARP – Eastern Carpathians, SCARP – Southern Carpathians.
Table 4.
Area of potential range according to the tested model and four sets of stands: CPYR – Central Pyrenees, EPYR – Eastern Pyrenees, ECARP – Eastern Carpathians, SCARP – Southern Carpathians.
| Region | Model | Area in the probability levels [hectares] | Total | |||
| Low (0.1-0.25) | Medium (0.25-0.50) | High (0.5-0.75) | Very High (>0.75) | |||
| CPYR | Current | 6 099.48 | 6 545.66 | 4 920.93 | 1 288.34 | 18 854.41 |
| RCP 2.6 | 6 344.43 | 10 857.13 | 420.55 | 16.35 | 17 638.46 | |
| RCP 8.5 | 6 698.12 | 711.52 | 4.67 | 0.00 | 7 414.31 | |
| EPYR | Current | 4 825.15 | 7 280.36 | 8 192.59 | 1 338.57 | 21 636.67 |
| RCP 2.6 | 5 756.46 | 5 986.58 | 6 773.90 | 5 156.05 | 23 672.99 | |
| RCP 8.5 | 4 482.04 | 4 812.61 | 4 664.27 | 4 470.36 | 18 429.28 | |
| ECARP | Current | 7 406.57 | 2 796.66 | 1 059.53 | 686.48 | 11 949.24 |
| RCP 2.6 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| RCP 8.5 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| SCARP | Current | 9 808.72 | 4 446.85 | 2 123.17 | 708.15 | 17 086.89 |
| RCP 2.6 | 771.79 | 0.00 | 0.00 | 0.00 | 771.79 | |
| RCP 8.5 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



