
Submitted:
10 August 2023
Posted:
14 August 2023
You are already at the latest version
Abstract
Inflammatory diseases are common pathological processes caused by various acute and chronic factors and some of them are autoimmune diseases. Exosomes are fundamental extracellular vesicles secreted by almost all cells, which contain a series of constituents, i.e. cytoskeletal and cytosolic proteins (actin, tubulin, histones), nucleic acids (mRNA, miRNA, DNA), lipids (diacylglycerophosphates, cholesterol, sphingomyelin, ceramide), and other bioactive components (cytokines, signal transduction proteins, enzymes, antigen presentation and membrane transport/fusion molecules, adhesion molecules). This review would be a synopsis of the actual knowledge on the contribution of exosomes from different cell sources as possible therapeutic agents against inflammation, focusing on several inflammatory diseases, neurological diseases, rheumatoid arthritis and osteoarthritis, intestinal bowel disease, asthma, liver and kidney injuries. Current knowledge indicates that the role of exosomes in therapy of inflammation and in inflammatory diseases could be distinctive. Main limitations to their clinical translation are still production, isolation, and storage. Additionally, there is an urgent need to personalize the treatments in terms of selection of exosomes, their dosages and routes of administration, and a deeper knowledge about their biodistribution, type and incidence of adverse events and long-term effects of exosomes. In conclusion, exosomes can very promising next generation therapeutic option, superior to synthetic nanocarriers and cell therapy, and can represent a new strategy of effective, safe, versatile and selective delivery systems in the future.
Keywords:
extracellular vesicles
; exosomes
; chemical composition
; miRNA
; nanocarriers
; inflammation
; neurological diseases
; liver
; kidney and lung injuries
; rheumatoid arthritis and osteoarthritis
; intestinal bowel diseases
1. Introduction
In the last decades, nanotechnology has substantially posed the basis to the development of smart nanocarriers, which are currently on the market or under clinical evaluation [1]. These nano-therapeutics represent a unique opportunity to improve the safety and efficacy of conventional drug delivery systems because they can improve drug solubility, enhance its bioavailability, decreasing toxicity, and give a superior dose response with a consequent greater patient compliance [2]. In particular, vesicles are the most versatile nano-platforms, as reflected by the clinical translation and marketed authorization of several formulations [3]. However, the major limitations of these synthetic nanodelivery systems are low physical stability and moderate ability to proficiently cross the physiological barriers, or penetrate the tissues, as well as cellular membranes [4]. In the last years, research has focused on other vesicles, the cell-derived extracellular-vesicle (EV), as smart nanocarriers for different possible applications because of their unique ability to strongly cross the physiological and pathological barriers and the resulting modifications induced in the targeted tissues and cells [5].
Indeed, EVs are able to cross the blood brain barrier and other physiological barriers, even if the mechanism is still not completely identified, and they can interact with cell membrane through a variety of ligands/receptors due to lipid and protein composition. Crossing and targeting properties result in a superior internalization via endosomes with respect to synthetic nanocarriers [6]. According to these data, there are currently several clinical trials ongoing with the aim to evaluate safety and efficacy of these innovative EV treatments and potentially recompense for the shortcomings of artificial delivery systems, mainly displaying inferior toxicity in spleen and liver, and having a lower immunogenicity [7]. Finally, in order to utilize EVs as drug carriers, a prerequisite is to find a strategy for efficient cargo loading and two different approaches can be distinguished: an exogenous approach after EV isolation, and an endogenous loading can be obtained during EVs biogenesis [8].
1.1. Classification and characteristics of the different EVs
EVs have in part a similar architecture to that of synthetic vesicles such as liposomes, mainly represented by a basic structure made of phospholipids. Proteins represent the main non-lipid molecules, localized in the intraluminal space or associated to the membrane, as unique protein surface decoration, conferring natural targeting properties. Protein composition is related to the different biogenesis pathways, but until now, it is not possible to discriminate between the different EV populations [9]. Their inner content also includes sugars, growth factors, protease inhibitors, adhesion integrins, and different kinds of genetic material including double stranded DNA, and numerous coding and non-coding RNAs [10].
EVs are produced from all types of cells, including prokaryotic cells and mammalian, as evidenced by the analysis of tissue culture supernatants. EXs can be isolated from biological fluids including plasma, saliva, breast milk, cerebrospinal fluids and malignant ascites. EVs can be distinguished in several subtypes according to the different biogenesis and the substantially different size distribution. Generally, three different populations are reported in the literature, namely apoptotic bodies, microvesicles (MVs) and exosomes (EXs), a classification based mainly on their biogenesis, release pathways, function and size (Figure 1) [5].
Apoptotic bodies have a size from 500 nm up to 5000 nm in diameter, generally tending to the larger side and they are released into the extracellular space by dying cells. Their production is due to the improved hydrostatic pressure during the final phase of apoptotic death. Apoptotic bodies are variable in structure and composition and may contain an extensive variety of cellular components: cytosol portions, chromatin remnants, micronuclei, DNA fragments, degraded proteins, or even intact organelles [11].
Macrophages, parenchymal cells, or neoplastic cells degrade apoptotic bodies after their release in the extracellular space by phagocytosis. Even if there are many reports concerning apoptotic body formation, studies on their role and function are still very limited [12].
MVs have a size typically in the range from 100 nm up to 1μm in diameter and include cytoplasmic material. Their formation is not well understood, and it seems to be related to the donor cell’s physiological state and the microenvironment (Figure 1). Protein and lipid constituents from the cytosolic and plasma membrane are main constituents of MVs. In particular, protein can be 100-fold higher concentration in MVs compared to the cell lysate, and these proteins also include cytoskeletal proteins, proteins containing post translational alterations, such as glycosylation and phosphorylation and integrins, typically adhesion molecules. Integrins could affect the vesicle trafficking and uptake, while other proteins such as the glycan binding proteins, generally present on the surface of MVs, could be related to targeting effects [13]. Nucleic acids, and several bioactive lipids represent other constituents of MVs. Recent studies have demonstrated that the proteomic profiles of MVs are profoundly related to the isolation technique and cell origin. MVs are released within the extracellular space like apoptotic bodies, and once entered into the circulation they can transfer their cargo to adjacent or distant cells to induce changes in the phenotype or function, which are important for numerous physiological and pathological conditions [14].
Finally, EXs have a diameter ranging from 30 to 150 nm and peculiar characteristics, which can differentiate these EVs from the MVs and apoptotic bodies. EXs are released into the extracellular space by different cells including adipocytes, fibroblasts, erythrocytes, lymphocytes, platelets, dendritic cells, brain and stem cells, and cancer cells. Their biogenesis of EXs occurs by inner budding of the plasma membrane, generating an endosomal vesicle and multivesicular bodies, which are fused with lysosomes, and generate the EXs into extracellular space (Figure 1). EXs can interact with the extracellular matrix, or provoke a reaction within adjacent or distant cells [15].
Most body fluids contain EXs, including plasma and serum, cerebral spinal fluid, synovial fluid, bronchial fluid, saliva, urine, breast milk, semen, amniotic fluid, tears, gastric medium, lymph, and bile. Their content is strongly related to physiological or pathological conditions, including cancer, neurodegenerative diseases and viral infections, for this reason EXs are impressively studied as a source of novel biomarkers (Figure 2). Similarly, to MVs, the protein and lipid composition of EXs is related to their origin. The exosome membrane is made of glycerophospholipids of containing long and saturated and unsaturated fatty-acyl chains (i.e., phosphatidylserine), and numerous lipid-rafts (sphingolipids, cholesterol, ceramides, Figure 2). The membrane is decorated with different proteins, including proteins that are essential for membrane transport and fusion (annexins, flotillin, Rab proteins), harbor tetraspanins (CD63, CD81, CD82, and CD9), adhesion integrins, and major histocompatibility complexes (MHC I and II), which are involved in antigen presentation (Figure 2). Inside EXs contain heat-shock proteins (Hsp60, Hsp70, and Hsp90), proteins associated with MVB biogenesis i.e., ALG2-interacting protein X , (Alix, also known as PDCD6IP) and tumour susceptibility gene 101 protein (TSG101), cytoskeletal proteins (tubulin, actin, cofilin, profilin, and so on) and metabolic enzymes (pyruvate kinase and GAPDH) (Figure 2). In addition, lipid layering molecules like glycosylphosphatidylinositol-anchored protein (LBPA), signal transduction molecules (G protein and protein kinases), and nucleic acids, represented by mRNAs, miRNAs, non-coding RNAs (ncRNAs), and mitochondrial DNA, which are involved in receive and send signals in intercellular communications are also present in Exs (Figure 2) [16,17,18].
2. Possible roles of exosomes in therapy
EXs have a crucial function in cell–cell interaction by transferring bioactive molecules from the donor cells to the recipient cells thanks to the surface receptor molecules and ligands. For these reasons, EXs possess a unique therapeutic potential for numerous diseases due to this ability and specificity for intracellular transporting ]. Hence, EXs have been investigated as vectors exerting biological activities in the targeted sites or nanocarriers loaded with specific molecules to produce a biological reaction. [15]. In both circumstances, their isolation and characterization from biological fluids or cell cultures are the critical precondition to establish their therapeutic potential. Internalization of EXs arises via membrane incorporation route mediated by lipid raft or via caveolae or clathrin-dependent endocytosis. Phagocytosis and micropinocytosis are also reported as possible technique of EXs internalization [19]. These processes represent a therapeutic potential as targeted delivery systems for efficiently performing therapeutic functions in the management of numerous infectious diseases, musculoskeletal and cardiovascular disorders, neurodegenerative disorders, and cancer [7]. Their possible use in different therapeutic application is also enforced by their very low immunogenicity and the capacity of crossing physiological barriers including blood-brain-barrier [20]. About 30 clinical trials have been completed till now and have proved the positive diagnostic and therapeutic effects of exosome-based treatment in various cancers, orthopedic conditions, neurodegenerative and hematological illnesses, auto-immune and cardiovascular diseases [21].
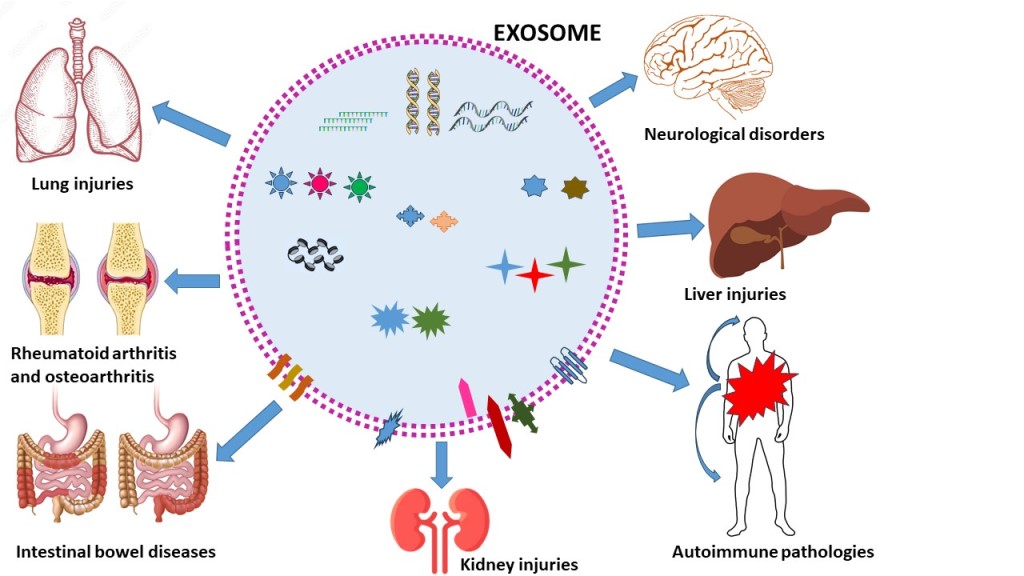
3. Role of EXs in Inflammatory diseases
Inflammation represents an essential factor in the pathogenesis of numerous diseases and there is a direct correlation between inflammation and the levels of specific EXs suggesting that EXs play a significant role in inflammation and immune responses. Various studies have demonstrated that EXs have potent immunomodulatory activities, mostly through the high expression of miRNAs, which can target the immune system and modulate angiogenesis [22,23,24].
The aim of the present review is to highlight the possible role of EXs as nanotherapeutic agents in inflammation, focusing on several different inflammatory diseases, neurological diseases, rheumatoid arthritis and osteoarthritis, intestinal bowel disease, asthma, liver and kidney injuries. The review would like to the state of art of the current knowledge of exosome-based systems with an emphasis on the contribution of exosomes as therapeutic agents, both as autologous treatment or as carriers of drugs.
3.1. Neurological disorders
Neurological disorders are diseases of the central and peripheral nervous systems and include brain, spinal cord, cranial nerves, peripheral nerves, nerve roots, autonomic nervous system, neuromuscular junction, and muscles. These disorders comprise Parkinson's disease (PD), epilepsy, Alzheimer’ s disease (AD), and other dementias, and cerebrovascular diseases represented by stroke, migraine and other headache disorders, multiple sclerosis, neuroinfections, brain tumors, and traumatic disorders of the nervous system due to head trauma and many others. According to WHO, neurological disorders affect Hundreds of millions of people worldwide are affected.
It is well known that cells of the central nervous system namely oligodendrocytes, astrocytes, neurons, and microglia produce EXs, which have an important role as messengers among neural cells both under normal and pathological conditions. These EXs enhance neuronal survival and improve neurite growth by stimulating synaptic activity, increasing ceramide and sphingosine metabolism in the recipient neurons and releasing synapsin, under cell stress or during high neuronal activity [25,26]. EXs are able to mediate neuroprotection, indeed astrocytes increase the secretion of high levels of heat shock protein 70 (Hsp70) under oxidative stress and hyperthermia helping the survival of adjacent neurons. Microglia also produce and release EXs similar to those of B lymphocytes, having a role in the immune system [27,28]. Schwann cells myelinate axons in the peripheral nervous system produce and release EXs to help axonal regeneration after injuries, which are able to suppress of the action of a GTPase (RhoA) activated in response to injury [29].
EXs produced by stem cells can decrease the expression of TNF-α and IL-6 inflammatory cytokines and enhance that of IL-4 and IL-10, reducing brain injury. Furthermore, they can also increase the stimulation of CD4+ and CD8+ lymphocytes, and reduce dendritic cells number [30]. In a neuroblastoma tauopathy model, EXs can regulate misfolded protein accumulation and can reduce Aβ deposition in AD experimental mouse models, reestablish the function of microglia, relieving memory deficits. EXs can also transfer phosphorylated tau protein associated with neurodegeneration, as detected in the cerebrospinal fluid of AD patients [25]. EXs derived from human mesenchymal stem cells (MSCs) give an improvement of the disease in a mouse model of autistic-like behaviour in mice, even if the exact mechanism of action it is not known [31], while in a swine model of traumatic brain injury exosomes originated from human MSC sustain neuroprotection [32]. An overview of the literature has significantly demonstrated that EXs, particularly those derived from MSC, neurons, stem cells, microglia and astrocytes can have a possible role in many neurological diseases, i.e., AD, PD, amyotrophic lateral sclerosis, multiple sclerosis, acute ischemic stroke, and spinal cord injury [33]. Indeed, MSCs can differentiate into different types of cells and produces EXs with potent antinflammatory and immunomodulatory activities [34], as well as neuronal stem cells (NSCs) which can differentiate as neurons, astrocytes, and oligodendrocytes [35] release exosomes, having strong neuroproliferative and neuroregenerative potential, and improving recovery in models of neurological injury [36].
Recently, it has been demonstrated that EXs derived from the oligodendrocyte precursor cells evolving into mature oligodendrocytes, showed a substantial efficacy on re-myelination and improvement of motor coordination in model mice [37].
Other cells able to generate exosomes with interest in anti-inflammatory and neurogenic properties are pluripotent stem cells [38]. Furthermore, EXs secreted by astrocytes and microglia, have an important role in regeneration and repair of CNS tissues. Indeed, it is reported that EXs from astrocytes stimulate axonal growth and re-myelination and can also restore the spinal cord, while EXs from microglial control inflammation and stimulate neuronal survival [39,40]. Finally, EXs can also be produced by oligodendrocytes and Schwann cells, the first produce myelin and support axons [41], while Schwann cells can restore connections of muscle nerve and tissues [42]. It has been also reported that EXs derived from MSCs have a role in the regulation of both antigen- specific and antigen-nonspecific immunity, thus modulating inflammatory and autoimmune diseases [43].
3.1.1. Alzheimer’s Disease
AD represent the most common type of dementia (accounts for 60-80% of dementia cases) involving regions of the brain controlling memory, behavior and language. WHO has estimated that globally 47.5 million people with dementia with 7.7 million new cases every year. Two kinds of anomalous structures, plaques (deposits of a protein fragment called beta-amyloid) and tangles (twisted fibers of tau protein) are responsible for damaging and killing nerve cells.
A recent review has reported on the possible role of exosomes from MSCs in AD addressed by four principal actions: release of anti-inflammatory constituents (proteins/lipids/microRNAs/enzymes), immunomodulation, decrease of Aβ plaques by internalization and degradation of Aβ oligomers, and regenerative effects [43]. In vitro EXs derived from MSCs protect neurons from neuronal oxidative stress, suggesting they can have a role in the preservation of synapse integrity when neurons are exposed to soluble oligomers of the Aβ peptide, having a leading role in the cognitive decline of AD. Authors of the study related this activity to the EXs content of anti-inflammatory and trophic constituents and antioxidant enzymes [44].
An interesting in vivo study with APP transgenic mice (an AD model) demonstrated that intracerebral injection of neuroblastoma-derived EXs reduced Aβ1–42 by binding to glycosphingolipids of the EXs surface and carrying Aβ peptides to microglia determining their phagocytosis and as a consequence their clearance [45]. Using animal models of AD, EXs from MSCs can decrease glial reactivity due to increased levels of the anti-inflammatory cytokines (IL-4, IL-10) and reduction of the pro-inflammatory ones (IL-1β, TNF-α). Indeed, it is reported that cognitive decline, typical of AD, is related to high levels of TNF-α and IL-1β. [46]. From the above studies it seems that the regulation of inflammatory processes can be related to the behavioral enhancements observed in animal models of AD, suggesting the contribution of iNOS.
In further in vivo studies, using AD mouse models, EXs derived from stem cell decrease Aβ deposition, reestablish microglial function and relieve memory impairments [47,48,49]. It is reported that the effects of EXs are also related to the regulation of gene expression mainly represented by miRNA (principally miRNA- 21), mRNAs, transcription factors, and other noncoding RNAs [43,50].
3.1.2. Parkinson’s Disease
PD is a brain disorder affecting the nervous system and the parts of the body controlled by the nerves that causes unintentional or uncontainable movements, causing deep difficulties of coordination and leading to progressive problems to walk and talk. A study reported the exploitability of EXs released from Raw 264.7 macrophages as successful carriers with antioxidant activity. EXs were loaded with catalase obtaining high loading efficiency, sustained release, and catalase preservation against proteases degradation. Exosomes were readily uptaken by neuronal cells in vitro. EXs after intranasal administration proficiently reduced oxidative stress and improved neuronal survival in an in vivo animal model of PD [52]. A further study reported that EXs produced by intracerebrally implanted cells in a mice model of PD, decreased neurotoxicity, and neuroinflammation through their mRNA cargo [53]. Finally, EXs derived from MSCs and loaded with curcumin improved significantly movement and motor coordination in a PD mice model after intranasal administration. Reported mechanisms include the decreasing α-synuclein aggregates, the promotion of neuron function recovery, and alleviation of the neuroinflammation. After treatment with theses drug deliveries, the movement and coordination ability of PD model mice were significantly enhanced [54].
3.1.3. Ischemic stroke
Ischemic stroke precludes brain cells from getting oxygen and nutrients, which start to die in a few minutes giving brain damage and other complications. WHO reported that more than 6 millions of people die because of stroke each year. A recent review addressed the possible regulatory effects of EXs obtained from stem cells on the inflammatory response after ischemic stroke. EXs derived from MSCs can regulate microglial polarization through various pathways such as NLRP3, NF-κB, and STATs, demonstrating that microglial M1 and M2 phenotypes have a relationship with the inflammatory response after ischemic stroke. In particular, the paper describes micro-RNAs from different origins possibly involved in the effects. It is reported that miRNA21a-5p induces microglial M2 polarization by targeting STAT3, miRNA-138-5p promotes astrocyte proliferation and inhibits inflammatory response, miRNA-542-3p suppresses inflammation and prevents cerebral infarction, miRNA-146a-5p is anti-inflammatory (IRAK1/TRAF6), and miRNA-150-5p reduces neuronal apoptosis and inflammation (TLR5) [30]. A further study demonstrated that EXs originating from adipose-derived stem cell can attenuate stroke by The study showed that exosomes from hypoxic pre-treated ADSCs attenuated acute ischemic stroke-induced brain injury via delivery of circ-Rps5 and modifying microglia from M1 to M2 phenotype in the hippocampus [55].
3.1.4. Multiple sclerosis and autoimmune encephalomyelitis
Multiple sclerosis (MS) is an autoimmune lifelong chronic inflammatory disease that can affect the brain and spinal cord, causing serious disability and a wide range of potential symptoms, including problems with vision, arm or leg movement, sensation or balance. In MS, the immune system attacks the layer that surrounds and protects the nerves called the myelin sheath, meaning that messages travelling along the nerves become decreased or disrupted. Exactly what causes the immune system to act in this way is unclear, but most experts think a combination of genetic and environmental factors is involved.
A very recent study demonstrated that EXs originated by RAW macrophages and loaded with resveratrol, significantly decreased inflammatory responses in both the central and peripheral nervous system in a mouse model of multiple sclerosis, successfully improving the clinical progression [56]. EXs when administered intranasally significantly improved disease severity and decreased inflammatory responses in central nervous system (TGF-β, IFN-γ, IL-1β, IL-6 and IL-17), in brain/spinal cord (IL-10 in spinal cord), in periphery (TGF-β, IL-1β, IL-6, IL-17 and IL-10) and in spleen/blood (IFN-γ). These data were confirmed by histopathology with a positive effect on the inflammatory infiltration and myelin recovery [56].
Autoimmune encephalomyelitis is a brain inflammation originated by the body’s immune system, which attacks healthy cells and tissues in the brain or spinal cord, sometimes misdiagnosed as a psychiatric or neurological disorder. It is a rare but complex disease, which can cause quick alterations of physical and mental health. Certain forms may be associated with tumours, as the ovarian tumours. In a mouse model of autoimmune encephalomyelitis, EXs derived from human MSCs and produced after stimulation with IFNγ were able to reduce neuroinflammation, decrease demyelination, and upregulate the amount of T cells characterized by CD4, CD25 surface antigens, and transcription factor forkhead box P3 within the spinal cords of treated mice [57]. Co-culture of the EXs with activated peripheral blood mononuclear cells in vitro reduced the cells proliferation and decreased pro-inflammatory cytokines levels, while immunosuppressive cytokine levels were increased. Anti-inflammatory RNAs were detected in the EXs [57].
3.1.5. Spinal cord injury
Spinal cord injury is a damage to any part of the spinal cord or nerves that sends and receives signals from the brain to and from the rest of the body. It can be caused by direct injury or from damage to the tissue and vertebrae of the spinal cord and it can be temporary or permanent loss of sensori-motor function. A study evaluated the efficacy of miRNAs isolated from EXs in a spinal cord injury experimental animal model. Furthermore, studies on EXs produced in the presence of insulin growth factor-1 (IGF) was evaluated both in vitro and in vivo. High levels of miR-219a-2-3p were found in EXs stimulated with IGF which led to the inhibition of the NF-κB pathway, a partial decrease of neuroinflammation and a promotion of neuroprotective effects in the animal model. The study evidenced that miR-219a-2-3p could be useful to decrease apoptosis and neuro-inflammation [58].
In a further study using EXs derived from MSCs, it was found that miRNA-216a-5p represents the most important miRNA to restore traumatic spinal cord injury by acting on microglial M1/M2 phenotypes. TLR4 was identified as the target downstream gene of miR-216a-5p together with NF-κB/PI3K/AKT signaling cascades [59].
EXs derived from human placental stem cells have proven to increase the expression of nerve trunk/progenitor cell markers in the spinal cord and to endorse recovery of both motor and autonomic functions after spinal cord injury experimental animal model with complete transection of the thoracic segment when intravenously administered. EXs also endorsed the stimulation of proliferating endogenous neural stem/progenitor cells and exhibited a significant high neurogenesis. In vitro, EXs promoted the production of neural stem cells and the increase of the phosphorylated levels of CREB, ERK, and MEK [60].
3.1.6. Traumatic brain injury
Traumatic brain injury occurs when a unexpected, exterior, physical attack injuries the brain and it represent one of the most common causes of disability and death in adults, with a vast array of injuries that happen to the brain which can be focal or diffuse originating mild to severe injuries. EXs obtained from astrocyte, enriched with miRNA-873a-5p decreased neuroinflammation, and reduced neurological deficits in post-traumatic brain injury models. Promotion of microglial polarization into the M2 phenotype was evidenced and the mechanism was related to the inhibition of NF-κB p65 and ERK phosphorylation [61]. In addition, EXs derived from astrocytes significantly decreased the mitochondrial levels of H2O2 and oxidative stress by enhancement of the activity of antioxidant enzymes (principally SOD and CAT) in the neurons of the hippocampus of rats with traumatic brain injury [61]. In an in vitro study, EXs derived from bone marrow mesenchymal stem cells stimulated the polarization of activated BV2 microglia to induce the phenotype with anti-inflammatory activity. Studies in animals demonstrated the reduction of cell apoptosis in cortical tissue, the inhibition of neuroinflammation, and the promotion of synthesis of the microglia phenotype with anti-inflammatory activity [62]. Similar results have also been found for EXs derived from human adipose MSCs. These EXs enhanced functional recovery, decreased neuronal apoptosis, suppressed neuroinflammation, and enhanced neurogenesis in a rat model of traumatic brain injury. In addition, the inhibition of P38 mitogen-activated protein kinase signaling and NF-κB was shown [63].
3.2. Rheumatoid Arthritis and Osteoarthritis
Rheumatoid Arthritis (RA) is a chronic autoimmune disease characterized by inflammation of synovial tissue, leukocyte infiltration into joints, liberation of inflammatory mediators and proteases, and degradation of the extracellular matrix mediated by matrix metalloproteinases (MMPs) [64]. A study demonstrated that EXs produced by MSCs which overexpress miR-150-5p (Exos-150) can inhibit angiogenesis, migration and invasion of Fibroblast-Like Synoviocytes, which produce MMPs that degrade type II collagen, and relieve the symptoms of RA by downregulating MMP14, considered a key enzyme mediating cartilage invasion and vascular endothelial growth factor (VEGF) expression [65]. Another strategy involves the modification of EXs surface by the metabolic glycoengineering of EXs produced by Adipose-Derived Stem Cells to target activated macrophages in the inflamed joints of RA. These EXs can reproduce natural inflammation resolution in the lesion via M1-M2 polarization, which defines a tissue remodeling, and repair improvement through the release of anti-inflammatory cytokines let-7b-5p and miR-24-3p contained in EXs were responsible for M1-M2 macrophage polarization via the JAK-STAT signaling pathway [66]. Compared with the unloaded EXs, the loaded EXs reduced the level of IFN-γ and promoted macrophage polarization toward the M1 phenotype together with the secretion of TNF-α and IL-6 targeting the activated macrophages. Contemporary, the IL-4 levels have been up-regulated, promoting the macrophage polarization toward the M2 type, implying the depletion of the M1 macrophages [66]. Indeed, the imbalance between the activities of pro-inflammatory M1 and anti-inflammatory M2 macrophages in RA induces synovial inflammation and autoimmunity, leading to joint damage. During inflammation, M1-type macrophages produce pro-inflammatory factors, interleukin-1β (IL-1β), and IL-6, which promote RA progression. In contrast, M2-type macrophages secrete IL-10, a powerful anti-inflammatory agent, which inhibits the production of pro-inflammatory factors and blocks the inflammatory response, while promoting healthy tissue remodeling. Therefore, increasing IL-10 secretion and decreasing pro-inflammatory factor levels can promote M1-to-M2 macrophage polarization, which can mitigate RA [67]. A study has evidenced that miR-204 synthetized by EXs plays an essential role in maintaining joint homeostasis to protect against RA pathogenesis. Indeed, exosomal miR-204-5p was present in mice with collagen-induced arthritis, while in humans exosomal miR-204-5p expression was inversely associated with disease parameters of RA (rheumatoid factor, erythrocyte sedimentation rate, and C-reactive protein). The in vitro study demonstrated that human T lymphocytes produced EXs containing large amounts of miR-204-5p able to inhibit cell proliferation. In vivo tests demonstrated that administration of lentiviruses expressing miR-204-5p noticeably relieved the collagen-induced arthritis in mice [68].
Osteoarthritis (OA) is a common age-related degenerative disease of joints that causes disability in elderly individuals. It is characterized by synovial inflammation that affects all tissues of the joint and lead to a dysfunction of the whole joint. However, OA pathogenesis is not fully understood, and no regenerative treatment has been approved to prevent or slow down the disease progression [69]. Numerous are the in vitro studies to evaluate the effects of EXs from different origin in regulating repair and regeneration of chondrocytes. Among the principal mechanisms, EXs are able to promote chondrocyte proliferation probably by miRNAs, regulating gene expression and modulating various signaling pathways responsible of cell proliferation through the regulation of CDH11, NF-kB, ROCK1, TLR9, and Wnt5a [70,71,72,73]. Moreover, miRNAs from EXs produced by MSCs can inhibit chondrocyte apoptosis by preventing the expression of pro-apoptotic proteins (IL-1β, HMGB1, and RUNX2) [74,75,76]. Additionally, EXs produced by MSCs are able to stimulate the production of extracellular matrix components in chondrocytes, such as proteoglycans and collagen (principally aggrecan and collagen II) [165]. EXs produced by MSCs control the action of enzymes (aggrecanases and MMPs) involved in extracellular matrix synthesis and degradation, stimulating extracellular matrix synthesis and preventing degradation in chondrocytes [77,78].
An interesting study investigated EXs derived from MSC can also be used as carriers to encapsulate curcumin, an anti-inflammatory drug with poor bioavailability. The in vitro modulatory effects of curcumin-primed human (h)BMSC-derived EXs (Cur-EXs) on IL-1β stimulated human osteoarthritic chondrocytes was evaluated. Cur-EXs relieved IL-1β-induced catabolic effects on human osteoarthritic chondrocytes by endorsing viability and migration, decreasing apoptosis and phosphorylation of Erk1/2, PI3K/Akt, and p38 MAPK thus modulating pro-inflammatory signaling pathways. Treatment of human osteoarthritic chondrocytes with Cur-EXs gives upregulation of expression of hsa-miR-126-3p which is involved in modulation of anabolic response of human osteoarthritic chondrocytes [69].
Another study tested the efficacy of EXs obtained from curcumin-treated bone marrow-derived MSCs in a surgically-induced OA mouse model by intra-articular injection of The EXs attenuated the progression of osteoarthritis and reduced the DNA methylation of miR-143 and miR-124 promoters. As a result, miR-143 and miR-124 were up-regulated to further inhibit the expression of their target genes, ROCK1 and NF-kB, which were closely related to the development of OA [79].
A further strategy to treat osteoarthritis is intra-articular injection of Platelet-Rich Plasma (PRP) in the joint because they are rich in EXs. Furthermore, to prolong EXs retention in the joints and obtain therapeutic effects, EXs were incorporated in injectable thermosensitive hydrogel (Poloxamer-407 and 188). In vivo studies demonstrated that EXs formulated in the gel improved their local retention, decreased the apoptosis and hypertrophy of chondrocytes, increased their production, and delayed the development of OA [80].
Intra-articular injection of EXs produced by human embryonic cells-derived MSCs improved gross appearance, upgraded histological scores, and gave the complete restoration of cartilage and subchondral bone [81]. In a similar study, intra-articular injection of EXs produced by miR-140-5p-overexpressing human synovial mesenchymal stem cells was tested in surgically-induced OA rat model. EXs improved the proliferation and migration of articular chondrocytes without harming extracellular matrix secretion and prevented OA in the rat model [82].
A rat model of osteoporosis has been investigated to further characterize the beneficial effects of EXs derived from MSC and in particular evaluate their safety profile. Indeed, the EXs from MSC improved bone restoration and angiogenesis in a dose-dependent way [83].
The effect of EXs from bone marrow MSCs on OA was evidenced by an in vivo study using an OA rat model obtained by surgery. Intra-articular injection of both bone marrow MSCs or their EXs similarly protected rat from joint damage. It was found that EXs preserved the chondrocyte matrix by enhancing collagen type II synthesis and reducing ADAMTS5, MMP13 and Col II expression in the presence of IL-1β in vitro. In addition, EXs protect chondrocytes from apoptosis and senescence, playing a particular role in regulating cell proliferation, in repair and regeneration challenge [84].
3.4. Lung injuries
Asthma is the principal disease among lung injuries affecting 300 million people worldwide. It is a heterogeneous chronic inflammatory airway disease, involving many cells of the innate and adaptive immune systems. It acts on airway epithelial cells triggering bronchial hyper-reactivity and airway remodeling in response to environmental stimuli such as allergens, infections, or air pollutants, which stimulate pro-inflammatory cytokines interleukins IL-4, IL-5, and IL-13 released by activated Th2 cells increasing numbers and airway mast cells and eosinophils. Additionally, mucus is observed [85]. A few studies have evidenced the association between circulating EXs and EXs-mediated miRNAs, proposed as asthma biomarkers. Indeed, as a consequence of asthma, EXs are released from dendritic cells, eosinophils, mast cells, and bronchial epithelial cells and can modulate the chronic inflammatory processes involved in asthma [85]. A study evaluated the activity of EXs produced by M2 macrophages from semisynthetic origin, whose membrane was coated PLGA@Dnmt3aossmart silencer (EM-PLGA@Dnmt3aossmart silencer) and administered intravenously to Der f1-induced asthma mice, a model of allergic asthma. EXs improved the pathology with a strong reduction of lung inflammation. EXs after the administration were distributed in various organs (including the lungs) and were able to target M2 macrophages. The treatment decreased the proportion of M2 macrophages and inflammatory cytokines in the airway, without suppress the overall immune function [86]. Acute lung injury (ALI) and the acute respiratory distress syndrome (ARDS) are further lung injuries characterized by acute respiratory failure with considerable morbidity and mortality. The incidence of these injuries is difficult to assess and ALI/ARDS represent an unmet medical need. Both syndromes are characterized by improved permeability of the membrane of alveoli and capillaries, interstitial and intra-alveolar edema, severe hypoxemia, and a reduction of pulmonary compliance [87].
A recent review has evaluated the use of EXs to treat ALI/ ARDS. The possible role of EXs can be related to their anti-inflammatory effects; modulation of apoptosis and cell regeneration, particularly through the microRNA contained in exosomes can participate in intercellular communication and play an immunomodulatory role in ALI/ARDS disease models [87]. The effectiveness of EXs produced by bone marrow MSCs in reducing inflammatory factor expression and the infiltration of neutrophils and macrophages was due to the miR-150. The EXs role was assessed by the regulation of the MAPK pathway [87]. In addition, EXs produced by MSCs decreased NF-kB activation and decreased the expressions of pro-inflammatory cytokines. Furthermore, EXs produced by MSCs decreased the expressions of proteins related to the TLR4/NF-kB pathway and this effect can be reversed when the miR-451 expression in the EXs was inhibited. Additionally, EXs derived from MSCs improved the reduction of PI3K and AKT phosphorylation produced by oxidative damage in rat lung tissue due to the activation of PI3K/AKT signaling pathway [87]. Additionally, EXs derived from bone marrow MSCs stimulated the expressions of anti-apoptotic proteins and ligand proteins by activating the Hippo-YAP pathway and JAK1-STAT3 signaling pathway, while EXs produced by airway myeloid-derived regulatory cells enhanced the T cell reaction in chronic inflammation by transferring mitochondria to T cells [87]. EXs produced by MSCs improved the production of reactive oxygen species and mitochondrial DNA damage and relieved mitochondrial dysfunction, restoring the barrier integrity in alveolar epithelial cells [87].
A very interesting study investigated the use of EXs produced by MSCs in preclinical models of ALI/ARDS. EXs produced by human umbilical cord MSCs reduced the levels of inflammatory factors and inhibited lung inflammation and oxidative stress in LPS-induced ALI by inducing autophagy in vivo. The study proved that miR-377-3p in the EXs played a crucial role in regulating autophagy. In particular, EXs released by human fetal lung fibroblast cells overexpressing miR-377-3p very efficiently inhibited the inflammatory factors and gave autophagy, giving recuperation of ALI. MiR-377-3p targeted regulatory-associated protein of mTOR , which induce autophagy both in vivo and in vitro. [88]. Additionally, EXs produced from adipose-derived MSCs were evaluated in rat with septic lung injury with the aim to evaluate the effects of microRNA 16-5p. EXs decreased lung injury and endorsed M2 polarization due to the miR-16-5p. In particular, silencing TLR4 enhanced lung inflammation and an overexpression of TLR4 reduced the influence of miR-16-5p in the mice. Accordingly, miR-16-5p mechanism was via suppressing TLR4 [89].
EXs produced by endothelial progenitor cell were investigated to an ALI model of mice. EXs were able to reduce cytokines/chemokines and protein concentration in the bronchoalveolar lavage fluid, myeloperoxidase activity, pulmonary edema and lung injury score due to the presence of miRNA-126 [90]. A further study confirmed that the EXs produced by endothelial progenitor cell improved the endothelial cell function through miRNA-126 via RAK/ERK signaling pathway and thus improving [91]. EXs obtained by alveolar progenitor type II cells promoted the production of ATII cells and improved epithelial regeneration in damaged alveolar cells in bleomycin-induced lung injury models. In particular, miRNA-371b-5p found in the EXs was responsible of the activity via Pi3k/Akt signaling [92].
A study evidenced that EXs produced by endothelial cell were able to inhibit topoisomerase II α expression, with a consequent protective role in a mouse model of sepsis-induced ALI. Activity was related to miRNA-125b-5p [93].
EXs produced by adipose tissue reduced the permeability of the pulmonary endothelial barrier and decreased the inflammatory reaction, by the inhibition of the transient receptor potential vanilloid 4 /Ca2+ signaling pathway. Therefore, a reduction of the pro-inflammatory cytokines induced by mechanical ventilation, resulting in a protective effect in a ventilator-induced lung injury [94].
A very interesting study investigated EXs engineered with receptor for advanced glycation end product-binding peptide, an anti-inflammatory peptide. The efficacy of these EXs was assessed by cytokine assays in lipolissacharide-activated macrophage cells. EXs were loaded with curcumin which demonstrated had greater anti-inflammatory effects than curcumin alone or unloaded EXs. In the in vivo study, the EXs were administrated by intra-tracheal instillation into the lungs of ALI model. A powerful reduction of pro-inflammatory cytokines and inhibition of inflammation was stated [95].
EXs have been also evaluated in patients with severe Coronavirus disease (COVID-19) that experienced acute respiratory distress syndrome (ARDS), as reported by a very recent review [96]. COVID-19 is an infectious disease caused by the SARS-CoV-2 virus which can evolving to severe symptoms and in some cases death. Severe form of COVID-19 is characterized by lymphopenia and thromboembolic complications, disorders of the nervous system, kidney, and acute cardiac and liver injuries. Signs of inflammation and increasing level of C-reactive protein and interleukin-6 are characteristic of severe patients [96]. WHO has calculated that about 800.000.000 patients experienced COVID-19, and about 7.000.000 patients died. Several studies have been reported to evaluate the activity of EXs produced from immune cells in reducing the inflammatory response and inhibit the proliferation and activity of T helper cell 17, which have a significant role in inducing respiratory failure through the delivery of regulatory miRNAs [96]. One of the first studies involved three COVID-19 patients with severe ARDS and multi-organ complications, which have been treated with Zofin (also known as Organicell Flow). The Zofin treatment resulted a safe and well tolerated therapy. Zofin was based on EVs, naturally produced by human amniotic fluid. The patients showed improvements of the disease with a strong improvement of the inflammatory biomarkers [97].
In a further clinical trial, EXs produced by bone marrow MSCs were given intravenously for the treatment of patients with ARDS caused by severe COVID-19. The study demonstrated an improvement of oxygenation status and clinical symptoms of the improved respiratory function. In particular, the expressions of the inflammatory biomarkers (C-reactive protein and IL-6) reduced, as well as decreased the Sequential Organ Failure Assessment scores. Significant was the decrease of ferritin and D-dimer, and the growth of neutrophil and lymphocyte cells without any adverse events [98].
In a very recent clinical study, ExoFlo (an extracellular signal product isolated from human bone marrow MSCs that contains growth factors and EVs including EXs) has been used to treat moderate to severe ARDS in patients with severe COVID-19. No adverse events were reported during the clinical trial and the treatment was useful to decrease mortality compared with placebo. Ventilation-free days improved after ExoFlo treatment [99].
Actually, other clinical studies on ALI/ARDS are in progress or completed, as evidenced by the website ClinicalTrial.gov. Therapeutic approaches are represented by EXs produced by human MSCs for the treatment of ARDS and treatment of ARDS or novel Coronavirus pneumonia caused by COVID-19. Studies are related to the evaluation of the safety profile and efficacy of EXs containing CD24 (a glycosylated protein linked to the cell surface by means of a glycosyl-phosphatidylinositol anchor) in patients with COVID-19 infection, and extracellular vesicle infusion obtained from Bone Marrow MSCs for the treatment for COVID-19 associated ARDS.
3.5. Liver injury
Liver injury and liver failure are diseases characterised by necrosis and apoptosis of liver cells, preventing the liver from performing normal synthetic and metabolic functions. The main causes are viruses, drugs, lipid deposits, autoimmune reactions; encephalopathy, cerebral oedema, sepsis, renal failure, gastrointestinal bleeding, and respiratory failure are the principal complications [64]. Chronic liver inflammation can also lead to fibrosis and cirrhosis. At present, liver transplantation remains the most effective option for patients with severe conditions, but several limitations still exist, such as lack of liver donors, high surgical costs, post-transplant complications, and immune rejections. Nucleoside binding and the NLRP3 inflammasome are the newly discovered pattern recognition receptors involved in the pathogenesis of many liver diseases. NLRP3 activation induces the death of inflammatory cells and pyroptosis, and simultaneously leads to the maturation and secretion of IL-1β, suggesting an involvement in the progression of autoimmune hepatitis (AIH) a chronic inflammatory disease in the liver with potential to promote the development of liver fibrosis. EXs produced by MSCs overexpressing miRNA-233 can inhibit NLRP3 activation and consequently liver inflammation and cell death. Moreover, the use of miRNA-17 carried by EXs produced by MSCs reduces NLRP3 inflammatory bodies [64]. A recent review on mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases reported that EXs derived from human umbilical cord MSCs ameliorated liver fibrosis by inhibiting both epithelial mesenchymal transition of hepatocytes and collagen production. At the same time, EXs produced by adipose tissue-derived MSCs expressing miR-122 have shown the ability to improve therapeutic efficacy in liver fibrosis treatment. Targeting hepatic stellate cells (HSCs) this miRNA helps the regulation of genes such as P4HA1 and IGF1R, which have been shown to be involved in HSC proliferation and collagen maturation [100].
Liver fibrosis is a chronic liver disease with the presence of progressive wound healing response caused by liver injury. Currently, there are no approved therapies for liver fibrosis [101]. EXs derived from human adipose mesenchymal stem cells (hADMSCs-EXs) have been investigated for the anti-fibrotic both in vitro and in vivo. EXs inhibited the proliferation of activated hepatic stellate cells by apoptosis and by blocking mytosys in G1 phase, efficiently preventing the expression of profibrogenic proteins and epithelial-to-mesenchymal transition as demonstrated by in vitro studies. EXs was effective via PI3K/AKT/mTOR signaling pathway [102].
EXs produced by Human Wharton's jelly MSCs were evaluated in a model of liver fibrosis. In addition, EXs enriched of miRNA-124-3p were also evaluated. Both EXs efficiently reduced collagen accumulation and inhibited the inflammation. The therapeutic effect of the EXs enriched with miRNA-124-3p was significantly higher than the effects of EXs produced by Human Wharton's jelly MSCs. MiRNA-124 down-regulated STAT3, that has a critical role in liver fibrosis. Both EXs were able to switch the phenotype of splenic monocytes Ly6Chi from inflammatory to restorative [103].
A further liver injury from surgery is the hepatic ischemia-reperfusion. EXs produced by adipose-derived stem cells have been evaluated for their hepatoprotective potential in a rat model of hepatic ischemia-reperfusion injury. After EXs treatment, liver histomorphology and hepatocyte ultrastructure changes were enhanced. EXs treatment significantly downregulated IL-6, interleukin-1β (IL-1β), and TNF-α levels, and upregulated IL-10 levels. EXs displayed their hepatoprotective properties by preventing endoplasmic reticulum stress and inflammation [104].
In a recent review, the role of EXs in some liver diseases has been reported [105]. It was found that the levels of miRNA-122 were enriched in EXs after drinking alcohol, as well as the release of inflammatory mitochondrial DNA from hepatocytes. HepG2 cells treated with ethanol and overexpressing CYP2E1 (involved in alcohol metabolism), released high number of EXs able to change macrophages to the M1 type inflammatory phenotype. Nonalcoholic fatty liver disease is a liver injury with an extreme fat deposition in hepatocytes, generally connected with metabolic syndrome and represents the most widespread chronic liver disease worldwide. EXs produced by hepatocytes under lipotoxic setting stimulate NLRP3 inflammasome giving IL-1β and caspase 1 activation. Free fatty acids stimulate hepatocytes to produce EXs containing among others, miRNAs, which regulate hepatic lipid homeostasis [105].
A study investigated the association of metformin (first line treatment for diabetes), and EXs produced by Wharton's jelly MSCs in the HepG2 cells insulin resistance model. The study disclosed EXs enhance the activity of metformin without needing to change metformin by reducing inflammatory cytokines (TNF-α, IL-1, and IL-6), and apoptosis. [106].
An interesting study investigated the effects of EXs produced by MSCs and hepatocytes given in association with imipenem in a mouse model of sepsis. Sepsis was induced in C57BL/6 mice, obtained by cecal ligation and puncture. Both EXs plus imipenem controlled local and systemic inflammation and enhanced the populations of T regulatory cells. However, the mixture of EXs plus imipenem had the best performance in decreasing inflammation, conserving all T lymphocyte populations, decreasing liver injury, and finally growing the survival rate [107].
Acute liver injury is generally associated with reduced prognosis but still there is a very limited number of effective drugs. In a study animal models (cecal ligation puncture and lipopolysaccharide plus D-galactosamine) to induce sepsis-induced acute liver injury. EXs produced by MSCs significantly reduced acute liver injury and consequent death. It was found that also miRNA-26a-5p was able to protect hepatocyte death and liver injury. The study revealed that the activity was due to Metastasis Associated Lung Adenocarcinoma Transcript 1 (MALAT1), an RNA Gene [108].
EXs produced by bone marrow MSCs were investigated alone or in association with praziquantel a potential therapy for schistosomal hepatic fibrosis in experimental animals. A significant reduction of hepatic fibrosis, of hepatic granulomas (number and diameter), a decrease of NF-κB expression and an upregulation of proliferating cell nuclear antigen expression were found in animals treated with EXs and those treated with EXs plus praziquantel [109].
1.4. Kidney injuries
Among kidney injuries, Chronic Kidney Disease (CKD) is an increasing worldwide health problem, which leads to kidney failure, cardiovascular disease and premature death [110]. This disease is tied closely with diabetes and hypertension. CKD treatments are limited to slowing disease progression through blood pressure and diabetes control or avoiding drugs that can worsen renal function. In addition to CKD, acute kidney injury (AKI), also known as acute renal failure, is a sudden episode of kidney failure or kidney damage [111], or kidney transplantation and dialysis. In a further review, the use of EXs produced by MSCs to treat CKD.
Another recent review has reported the latest advances in the use of EVs as a therapeutic approach for kidney injuries. In particular, EVs produced by bone marrow-derived MSCs represent the extensively source for AKI recovery and to decrease CKD progression [112]. The reported studies used EVs and not isolated EXs, in AKI, nephrectomia, and unilateral ureteral obstruction, using intravenous administration, renal intra-capsular injection and intra-arterial injection. Enhancement of tubular cell proliferation, anti-apoptosis and reduced fibrosis in long term, support of morphologic and functional recovery, reduced tubular atrophy, improved kidney function, reduction in inflammation and cell death, decreased cell apoptosis and inflammation, represent the main effects of the EVs.
Studies with Adipose-Derived Mesenchymal Stromal Cells (ADMSCs) are mainly based on EVs used by intravenous or intra-arterial injection in AKI, DOCA-salt hypertensive model, unilateral renal stenosis. These studies evidenced reduction in apoptosis, oxidative stress and inflammation, prevention of kidney fibrosis inhibition of apoptosis, immunomodulation, recovery of intracellular ATP, preservation of mitochondria , increased cell proliferation, angiogenesis, immunomodulation. A study used Perinatal-Derived Mesenchymal Stem Cells (PDMSCs) EVs administered intravenously in Sepsis-AKI, reduced inflammation and apoptosis. Another study involved EVs produced by Kidney Progenitor Cells (KPCs). The administration of EV by intravenous injection in a model of AKI (Unilateral ischemia-reperfusion injury plus unilateral nephrectomy) improved kidney function and reduced ischemic damage. Two studies tested EVs from Human Liver Stem Cells (HLSCs) by intravenous injection. The first trial was done in a model of diabetic nephropathy and gave a prevention and reversal of the progression of glomerular and interstitial fibrosis. The second study demonstrated that intravenous injection of the EVs in a glycerol-induced AKI, improved kidney function and cell proliferation, reduced tubular necrosis. Additionally, several studies have been reported using EVs produced by Induced Pluripotent Stem Cells (iPSCs) by intravenous and intrarenal injection in unilateral and bilateral IRI. Reduced inflammation, inhibited cell apoptosis, antioxidant effects, enhanced angiogenesis and cell proliferation, inhibition of endoplasmic reticulum stress and apoptosis, improved kidney function, cell proliferation, decreased tubular injury, cell death and fibrosis, mitigation of fibrosis, reduction in necroptosis were found in the studies [112].
An interesting study investigated the efficacy of EXs originated from bone marrow-derived MSCs in a model of diabetic kidney disease, the most common CDK form. Indeed, hyperglycemia induces oxidative stress and reactive oxygen species, activate renin-angiotensin-aldosterone system and profibrotic cytokines. EXs repressed the extreme infiltration by regulating the expression of the adhesion molecule ICAM-1. EXs were able to inhibit proinflammatory cytokine expression (e.g., TNF-α) and fibrosis, down-regulated TGF-β1 expression and exerted an anti-apoptotic [113]. In a further study using a rat model of streptozotocin-induced diabetes mellitus, EXs prominently enhanced renal function and exhibited restoration of renal tissues, with substantial growth of LC3 and Beclin-1, and substantial reduction of mTOR and fibrotic marker expression in renal tissue. All previous effects were partially abolished by the autophagy inhibitors chloroquine and 3-MA [114].
EXs produced by adipose-derived MSCs decreased DKD in mice (reduced levels of serum creatinine, urine protein and blood urea nitrogen) by reversing autophagy downregulation and suppressing podocyte apoptosis [115].
EXs produced by human umbilical cord MSCs administered in an irreversible model of unilateral ureteral obstruction, alleviated kidney fibrosis and restored renal function (decreased creatinine and blood urea nitrogen) through inhibition of apoptosis, malondialdehyde, ROS, and ROS-mediated P38MAPK/ERK signaling pathways. In vitro, similar anti-fibrotic effects were also observed in TGF-β1 treated NRK-52E cells [116].
Efficacy of EXs produced by adipose-derived MSCs was also evaluated in a renal artery stenosis model. EXs stabilized the systolic blood pressure, reduce profibrotic gene collogen and TGF-β expression, downregulated hypoxia marker HIF-1a. Interestingly, both EXs, EVs and the adipose-derived MSCs treatments had a similar efficacy in decreasing the expression of the fibrotic markers TGF-β and collagen-1 [117].
More recently, a study investigated the activity of EXS produced by human induced pluripotent stem cells-derived MSCs after intravenous infusion. EXs alleviated kidney fibrosis, decreased inflammatory responses, and increased renal function in mice subjected to unilateral ureteral obstruction. The effects of EXs were via increasing SIRT6 and reducing β-catenin and its products Fsp1, PAI-1, and Axin2 [118].
EXs produced by adipose-derived-MSCs reduced levels serum creatinine, blood glucose, 24-h urinary protein, urine albumin-to-creatinine ratio, and kidney weight/body weight, suppressed kidney fibrosis in a diabetic nephropathy model of rats. EXs also inhibited levels of IL6 and promoted cell apoptosis. The results evidenced that miRNA-125a was at least partially responsible for efficacy of EXs [119].
EXs produced human urine derived MSCs were investigated prevent kidney complications from type I diabetes in rats after intravenous injection. EXs decreased urine volume and urinary albumin excretion, relieved albuminuria, and suppressed the caspase-3 overexpression [120]. In a similar study, the same EXs were tested in a model of diabetic nephropathy. The study indicated that miRNA-16-5p was evidenced to recover podocyte injury [121]. The protective function of EXs produced from adipose-derived MSCs was evaluated in a mice (C57/BL6) model of sepsis-induced acute kidney injury obtained by puncture. EXs were able to activate SIRT1 decreasing the reverse microcirculation disorders, inflammation, and apoptosis and to reduce mortality [122].
In a very interesting study, glial cell line-derived neurotrophic factor was transfected into human adipose mesenchymal stem cells via a lentiviral transfection system. EXs produced from the transfected cells were evaluated in vivo in a unilateral ureteral obstruction mouse model and in vitro in human umbilical vein endothelial cells. EXs significantly decreased renal fibrosis score and peritubular capillary in the in vivo studies. In vitro studies revealed EXs exerted cytoprotective effects on in human umbilical vein endothelial cells by enhancing SIRT1 signalling, and increased levels of phosphorylated endothelial nitric oxide synthase [123].
3.6. Intestinal Bowel Disease (IBD)
IBD is a chronic and debilitating disease characterized by idiopathic mucosal inflammation of the gastrointestinal tract. It includes several diseases, namely Crohn’s disease (CD), which may cause harmful inflammation in any part of the gastrointestinal tract, and ulcerative colitis, which generally affects only the mucosa of the large intestine and it’s characterized by bloody diarrhoea. Recent studies have shown that genetic and environmental factors, immune response abnormalities, intestinal barrier dysfunction and intestinal flora imbalances are related to the development of IBD [124,125].
IBD disease commonly causes also intestinal epithelial cells death, not only destroying the integrity of the barrier, but also causing inflammation, increasing the levels of inflammatory cytokines, such as TNF-α. The intestinal mucosal inflammation brings to profound changes including depletion of local nutrients, imbalances in tissue oxygen supply and in demand and production of high quantities of reactive nitrogen and oxygen intermediates [126]. IBD causes are unclear but the pathology is believed to be associated with genetic, environmental, gut microbiota and immune response factors [126] and although significant progress has been made in its treatment there are still some patients who face difficulties in being cured by traditional methods.
A recent review has examined the EXs-mediated effects and applications in inflammatory diseases of the digestive system [127]. EXs produced by human umbilical cord MSCs have been investigated in a murine model of inflammatory bowel disease (dextran sulphate sodium treatment). It was found a reduction of macrophages and the inhibition of their IL-7 expression. EXs produced by dendritic cells controlled the immune response and avoided the progress of autoimmune diseases. EXs produced by Staphylococcus enterotoxin A-treated dendritic cells regulated the release of inflammatory cytokines in a mice model of colitis (dextran sulphate sodium treatment). EXs reduced expression of the pro-inflammatory cytokines interferon (IFN)-γ, TNF-α, IL-17A, and IL-22and IL-12 and increased the anti-inflammatory cytokine TGF-β [127]. EXs derived from human umbilical cord MSCs carrying miR-378a-5p were investigated in a model of colitis. EXs induced suppression of interleukin (IL)-18, IL-1β secretion, and Caspase-1 cleavage abolishing cell pyroptosis and protecting against colitis [128].
4. Future perspectives and concluding remarks
In the recent years, an incredible number of new studies have tremendously improved the knowledge of EXs’ role in inflammatory diseases elucidating also specific targets and functions. EXs are nanosized vectors produced by almost all the human cells, which are released by cells as vectors of intercellular communication (over short and long-distance), both in health and disease conditions. In vitro studies and preclinical trials have evidenced EXs can play fundamental roles in the incidence and development of inflammatory disorders of the different body organs. Current knowledge indicates that the role of EXs in inflammation and in inflammatory diseases is extensive and complex.
EXs contain numerous constituents, i.e., cytoskeletal and cytosolic proteins, nucleic acids, lipids, and many other bioactive components (cytokines, signal transduction proteins, enzymes, antigen presentation and membrane transport/fusion molecules, adhesion molecules), and their specific composition is related to their progenitor cells.
EXs activity is due to additive/synergistic effects of the different molecules they contain, and numerous are the evidences EXs can be ideal delivery vehicles for overcome physiological and pathological membranes, including blood–brain barrier. EXs activity can be related to modulation of apoptosis and cell regeneration, and anti-inflammatory effects. EXs can release selectively their cargo characterized by numerous molecules with different functional roles, involved in multiple signal pathway transduction and regulating mitochondrial function, mainly represented by a variety of growth factors and non-coding RNAs, principally miRNAs. Additionally, surface manipulation of EXs ,using molecules from genetic/biological engineering or chemical nature, is occasionally reported to optimize the interaction with targeted cells, as well as the loading of drugs or natural senolytic constituents, or simply an association of EXs and drugs given as conventional dosage forms to enhance the activity has been studied. Preliminary studies suggest that drugs (co-loaded or given in association in conventional dosage forms) and senolytic agents can increase the therapeutic activity but still their synergies/additivities with EXs is not to establish as well as appropriate doses and possible risks.
Truly, the EXs interaction with the cells and tissues modulates numerous signaling pathways, as reported in the preclinical studies. In brief, in animal models of neuroinflammation EXs provoke a decreasing of TNF-α, an increasing of IL-10, and stimulation of M2 polarization of microglia, which allow to the reduction of the expression of inflammatory proteins (IL-1β, IL-18, CXCL2, CXCL10). EXs regulate the Rheb-mTOR signaling pathway, with a significant decrease in CD3+T and CD4+T cells. In addition, in the animal models of AD, EXs reduced Aβ oligomer expression and decreased Aβ plaque deposition, via enhancing PINK1/Parkin pathway-mediated autophagy. Finally, TLR on microglia is activated.
Administration of EXs in animal models of lung injuries provoked the reduction of TNF-α, IL-1β, IL-6, and IL-8, as well as the number of inflammatory cells. Furthermore, an increase of IL-10 and superoxide dismutase activity, while the levels of H2O2, malondialdehyde, and markers of oxidative stress diminished. Furthermore, a reduction of IL-4, IL-5, and IL-13 and a growth of TGF-β expression and T regulatory cells. A regulation of the MAPK pathway and NF-kB, a decrease of proteins expression related to TLR4/NF-kB pathway, and the activation of PI3K/AKT, Hippo-YAP and JAK1-STAT3 signaling pathways are also described.
Similarly, the administration of EXs in animal models of kidney injuries increased the T regulatory cells, an upregulation of CD206, the M2 macrophage marker. A reduced oxidative stress with an increased expression levels of SOD1, AOX1, SIRT1 and SIRT2, and a reduction of expression of iNOS, TNF-α, IL-6, and IL-1β, while IL-10 increased.
Administration of EXs in animal models of liver injuries significantly reduced expression of inflammatory cytokines (TNF-α, IL-1β, IL-2, IL-6, IL-10, IL-17 MCP-1, and TGF-β) and M1 macrophage marker proteins (CD68 and CD11c). A reduced expression of fibrosis-related proteins, inflammatory chemokines and tissue inhibitor of metallopeptidase-1 was also observed.
Studies in animal models of rheumatoid arthritis and osteoarthritis evidenced that EXs regulate immunological reactivity, inhibit apoptosis and stimulate proliferation. EXs inhibit CXCL9, and downregulated VEGF and MMP-14. EXs may regulate T cell differentiation and proliferation and the generation of Tregs. EXs increase IL-10 and decrease the immune response and inhibit IL-17. EXs inactivate the NF-κB and increase IL1RN, IL6, IL10 and IL11, and osteogenic factors (ALP, RUNX2, BMP2 and BMPR). Finally EXs induce M1 macrophage polarization into M2.
In animal models of intestinal bowel diseases, the expression of pro-inflammatory cytokines (IL-1β and IL-6) was significantly reduced, while levels of IL-10 was increased. EXs also reduced JAK1 and STAT1 phosphorylation by the JAK-STAT signaling pathway. Suppression of IL-7, TNF-α, and IL-6, NF-κB is also reported.
The reported studies have evidenced that EXs could represent a novel cell-free therapeutic strategy for decreasing inflammation and may provide a novel treatment approach for the reported pathologies. Furthermore, EXs can be superior from the synthetic nano-drug delivery systems (vesicles, micelles, nanoparticles) in terms of lower immunogenicity, higher structural stability and lack of toxicity.
Although a significant number of preclinical studies has been reported in the literature, there is a scarcity of research concerning the possibility of translation to clinical applications. Actually, EXs main limitations are related to their production, isolation, and storage. Future principal challenge will be EXs standardization, which should be be easier than the whole EVs secretion, indeed EXs quality control is essential for safe and efficacious treatments. Additionally, there is an urgent need to personalize the treatments in terms of selection of type of EXs from different origins, their dosages and routes of administration. Further requirements to bring this promising next generation therapeutic option into practice are the comprehension of EXs biodistribution after administration, type and incidence of adverse events and long-term effects.
Author Contributions
Conceptualization, A.R.B., L.G., and F.M.; literature search, T.M.; data curation, F.M, and T.M.; original draft preparation, A.R.B. and T.M.; review and editing, L.G. and F.M.; supervision, A.R.B. and L.G. All authors have read and agreed to the published version of the manuscript.
Funding
This review was funded by Ministry of University and Research (MUR), National Recovery and Resilience Plan (NRRP), grant number 1553 11.10.2022.
Acknowledgments
Work supported by #NEXTGENERATIONEU (NGEU) and funded by the Ministry of University and Research (MUR), National Recovery and Resilience Plan (NRRP), project MNESYS (PE0000006)–A Multiscale integrated approach to the study of the nervous system in health and disease (DN.1553 11.10.2022).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Halwani, A.A. Development of Pharmaceutical Nanomedicines: From the Bench to the Market. Pharmaceutics 2022, 14(1), 106. [Google Scholar] [CrossRef]
- Bilia, A.R.; Piazzini, V.; Risaliti, L.; Vanti, G.; Casamonti, M.; Wang, M.; Bergonzi, M.C. Nanocarriers: A Successful Tool to Increase Solubility, Stability and Optimise Bioefficacy of Natural Constituents. Curr Med Chem 2019, 26, 4631–4656. [Google Scholar] [CrossRef]
- Bilia, A.R.; Bergonzi, M.C.; Guccione, C.; Manconi, M.; Fadda, A.M.; Sinico, C. Vesicles and micelles: Two versatile vectors for the delivery of natural products. J Drug Deliv Sci Technol 2016, 32, 241–255. [Google Scholar] [CrossRef]
- Alshawwa, S.Z.; Kassem, A.A.; Farid, R.M.; Mostafa, S.K.; Labib, G.S. Nanocarrier Drug Delivery Systems: Characterization, Limitations, Future Perspectives and Implementation of Artificial Intelligence. Pharmaceutics 2022, 14, 883. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol 2021, 16, 748–759. [Google Scholar] [CrossRef]
- Busatto, S.; Morad, G.; Guo, P.; Moses, M.A. The role of extracellular vesicles in the physiological and pathological regulation of the blood-brain barrier. FASEB BioAdvances 2021, 3(9), 665–675. [Google Scholar] [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T. A review on exosomes application in clinical trials: perspective, questions, and challenges. Cell Commun Signal CCS 2022, 20, 145. [Google Scholar] [CrossRef]
- Zeng, H.; Guo, S.; Ren, X.; Wu, Z.; Liu, S.; Yao, X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells 2023, 12, 1416. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Bonsergent, E.; Grisard, E.; Buchrieser, J.; Schwartz, O.; Théry, C.; Lavieu, G. Quantitative characterization of extracellular vesicle uptake and content delivery within mammalian cells. Nat Commun 2021, 12, 1864. [Google Scholar] [CrossRef]
- Battistelli, M.; Falcieri, E. Apoptotic Bodies: Particular Extracellular Vesicles Involved in Intercellular Communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: disease message and therapeutic target potentials. Biosci Rep 2019, 39, BSR20180992. [Google Scholar] [CrossRef]
- Clancy, J.W.; Schmidtmann, M:; D’Souza-Schorey, C. The ins and outs of microvesicles. FASEB BioAdvances 2021, 3, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Kee, L.T.; Al-Masawa, M.E.; Lee, Q.H.; Subramaniam, T.; Kok, D.; Ng, M.H.; Law, J.X. Scalable Production of Extracellular Vesicles and Its Therapeutic Values: A Review. Int J Mol Sci 2022, 23, 7986. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Skryabin, G.O.; Komelkov, A.V.; Savelyeva, E.E.; Tchevkina, E.M. Lipid Rafts in Exosome Biogenesis. Biochem Biokhimiia 2020, 85, 177–191. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci 2019, 9, 19. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal CCS 2021, 19, 47. [Google Scholar] [CrossRef]
- Gonda, A.; Kabagwira, J.; Senthil, G.N.; Wall, N.R. Internalization of Exosomes through Receptor-Mediated Endocytosis. Mol Cancer Res MCR 2019, 17, 337–347. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.H.; Jeyaraj, M.; Qasim, M.; Kim, J.H. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. ClinicalTrials.gov. Accessed July 29, 2023. https://clinicaltrials.gov/search?cond=exosomes&aggFilters=status:com.
- Yu, B.; Zhang, X.; Li, X. Exosomes derived from mesenchymal stem cells. Int J Mol Sci 2014, 15, 4142–4157. [Google Scholar] [CrossRef] [PubMed]
- Leñero, C.; Kaplan, L.D.; Best, T.M.; Kouroupis, D. CD146+ Endometrial-Derived Mesenchymal Stem/Stromal Cell Subpopulation Possesses Exosomal Secretomes with Strong Immunomodulatory miRNA Attributes. Cells 2022, 11, 4002. [Google Scholar] [CrossRef]
- Kouroupis, D.; Kaplan, L.D.; Best, T.M. Human infrapatellar fat pad mesenchymal stem cells show immunomodulatory exosomal signatures. Sci Rep 2022, 12, 3609. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Pulliam, L. Exosomes as mediators of neuroinflammation. J Neuroinflammation 2014, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cesca, F.; Loers, G.; Schweizer, M.; Buck, F.; Benfenati, F.; Schachner, M.; Kleene, R. Synapsin I is an oligomannose-carrying glycoprotein, acts as an oligomannose-binding lectin, and promotes neurite outgrowth and neuronal survival when released via glia-derived exosomes. J Neurosci 2011, 31, 7275–7290. [Google Scholar] [CrossRef]
- Taylor, A.R.; Robinson, M.B.; Gifondorwa, D.J.; Tytell, M.; Milligan, C.E. Regulation of heat shock protein 70 release in astrocytes: role of signaling kinases. Dev Neurobiol 2007, 67, 1815–1829. [Google Scholar] [CrossRef]
- Tytell, M. Release of heat shock proteins (Hsps) and the effects of extracellular Hsps on neural cells and tissues. Int J Hyperth 2005, 21, 445–455. [Google Scholar] [CrossRef]
- Lopez-Verrilli, M.A.; Picou, F.; Court, F.A. Schwann cell-derived exosomes enhance axonal regeneration in the peripheral nervous system. Glia 2013, 61, 1795–1806. [Google Scholar] [CrossRef]
- Chen, N.; Wang, Y.L.; Sun, H.F.; Wang, Z.Y.; Zhang, Q.; Fan, F.Y.; Ma, Y.C.; Liu, F.X.; Zhang, Y.K. Potential regulatory effects of stem cell exosomes on inflammatory response in ischemic stroke treatment. World J Stem Cells 2023, 15, 561–575. [Google Scholar] [CrossRef]
- Perets, N.; Hertz, S.; London, M.; Offen, D. Intranasal administration of exosomes derived from mesenchymal stem cells ameliorates autistic-like behaviors of BTBR mice. Mol Autism 2018, 9, 57. [Google Scholar] [CrossRef]
- Williams, A.M.; Dennahy, I.S.; Bhatti, U.F.; Halaweish, I.; Xiong, Y.; Chang, P.; Nikolian, V.C.; Chtraklin, K.; Brown, J.; Zhang, Y.; Zhang, Z.G.; Chopp, M.; Buller, B.; Alam, H.B. Mesenchymal Stem Cell-Derived Exosomes Provide Neuroprotection and Improve Long-Term Neurologic Outcomes in a Swine Model of Traumatic Brain Injury and Hemorrhagic Shock. J Neurotrauma 2019, 36, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Giovannelli, L.; Bari, E.; Jommi, C.; Tartara, F.; Armocida, D.; Garbossa, D.; Cofano, F.; Torre, M.L.; Segale, L. Mesenchymal stem cell secretome and extracellular vesicles for neurodegenerative diseases: Risk-benefit profile and next steps for the market access. Bioact Mater 2023, 29, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Arabpour, M.; Saghazadeh, A.; Rezaei, N. Anti-inflammatory and M2 macrophage polarization-promoting effect of mesenchymal stem cell-derived exosomes. Int Immunopharmacol 2021, 97, 107823. [Google Scholar] [CrossRef] [PubMed]
- Andjus, P.; Kosanović, M.; Milićević, K.; Gautam, M.; Vainio, S.J.; Jagečić, D.; Kozlova, E.N.; Pivoriūnas, A.; Chachques, J.C.; Sakaj, M.; Brunello, G.; Mitrecic, D.; Zavan, B. Extracellular Vesicles as Innovative Tool for Diagnosis, Regeneration and Protection against Neurological Damage. Int J Mol Sci 2020, 21, 6859. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Buller, B.; Chopp, M. Exosomes - beyond stem cells for restorative therapy in stroke and neurological injury. Nat Rev Neurol 2019, 15, 193–203. [Google Scholar] [CrossRef]
- Zhai, Y.; Wang, Q.; Zhu, Z.; Hao, Y.; Han, F.; Hong, J.; Zheng, W.; Ma, S.; Yang, L.; Cheng, G. High-efficiency brain-targeted intranasal delivery of BDNF mediated by engineered exosomes to promote remyelination. Biomater Sci 2022, 10, 5707–5718. [Google Scholar] [CrossRef]
- Upadhya, R.; Madhu, L.N.; Attaluri, S.; Gitaí, D.L.G.; Pinson, M.R.; Kodali, M.; Shetty, G.; Zanirati, G.; Kumar, S.; Shuai, B.; Weintraub, S.T.; Shetty, A.K. Extracellular vesicles from human iPSC-derived neural stem cells: miRNA and protein signatures, and anti-inflammatory and neurogenic properties. J Extracell Vesicles 2020, 9, 1809064. [Google Scholar] [CrossRef] [PubMed]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-derived extracellular vesicles: Neuroreparative properties and role in the pathogenesis of neurodegenerative disorders. J Control Release 2020, 323, 225–239. [Google Scholar] [CrossRef]
- Guo, M.; Hao, Y.; Feng, Y.; Li, H.; Mao, Y.; Dong, Q.; Cui, M. Microglial Exosomes in Neurodegenerative Disease. Front Mol Neurosci 2021, 14, 630808. [Google Scholar] [CrossRef]
- Krämer-Albers, E.M.; Bretz, N.; Tenzer, S.; Winterstein, C.; Möbius, W.; Berger, H.; Nave, K.A.; Schild, H.; Trotter, J. Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: Trophic support for axons? Proteomics Clin Appl 2007, 1, 1446–1461. [Google Scholar] [CrossRef]
- Shaimardanova, A.A.; Solovyeva, V.V.; Chulpanova, D.S.; James, V.; Kitaeva, K.V.; Rizvanov, A.A. Extracellular vesicles in the diagnosis and treatment of central nervous system diseases. Neural Regen Res 2020, 15, 586–596. [Google Scholar] [PubMed]
- Elia, C.A.; Losurdo, M.; Malosio, M.L.; Coco, S. Extracellular Vesicles from Mesenchymal Stem Cells Exert Pleiotropic Effects on Amyloid-β, Inflammation, and Regeneration: A Spark of Hope for Alzheimer’s Disease from Tiny Structures? BioEssays News Rev Mol Cell Dev Biol 2019, 41, e1800199. [Google Scholar] [CrossRef] [PubMed]
- de Godoy, M.A.; Saraiva, L.M.; de Carvalho, L.R.P.; Vasconcelos-Dos-Santos, A.; Beiral, H.J.V.; Ramos, A.B.; Silva, L.R.P.; Leal, R.B.; Monteiro, V.H.S.; Braga, C.V.; de Araujo-Silva, C.A.; Sinis, L.C.; Bodart-Santos, V.; Kasai-Brunswick, T.H.; Alcantara, C.L.; Lima, A.P.C.A.; da Cunha-E Silva, N.L.; Galina, A.; Vieyra, A.; De Felice, F.G.; Mendez-Otero, R.; Ferreira, S.T. Mesenchymal stem cells and cell-derived extracellular vesicles protect hippocampal neurons from oxidative stress and synapse damage induced by amyloid-β oligomers. J Biol Chem 2018, 293, 1957–1975. [Google Scholar] [PubMed]
- Yuyama, K.; Sun, H.; Sakai, S.; Mitsutake, S.; Okada, M.; Tahara, H.; Furukawa, J.; Fujitani, N.; Shinohara, Y.; Igarashi, Y. Decreased amyloid-β pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J Biol Chem 2014, 289, 24488–24498. [Google Scholar] [CrossRef]
- Cui, G.H.; Wu, J.; Mou, F.F.; Xie, W.H.; Wang, F.B.; Wang, Q.L.; Fang, J.; Xu, Y.W.; Dong, Y.R.; Liu, J.R.; Guo, H.D. Exosomes derived from hypoxia-preconditioned mesenchymal stromal cells ameliorate cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice. FASEB J 2018, 32, 654–668. [Google Scholar] [CrossRef]
- Yang, L.; Zhai, Y.; Hao, Y.; Zhu, Z.; Cheng, G. The Regulatory Functionality of Exosomes Derived from hUMSCs in 3D Culture for Alzheimer’s Disease Therapy. Small Weinh Bergstr Ger. 2020, 16, e1906273. [Google Scholar] [CrossRef]
- Cone, A.S.; Yuan, X.; Sun, L.; Duke, L.C.; Vreones, M.P.; Carrier, A.N.; Kenyon, S.M.; Carver, S.R.; Benthem, S.D.; Stimmell, A.C.; Moseley, S.C.; Hike, D.; Grant, S.C.; Wilber, A.A.; Olcese, J.M.; Meckes, D.G. Jr. Mesenchymal stem cell-derived extracellular vesicles ameliorate Alzheimer’s disease-like phenotypes in a preclinical mouse model. Theranostics 2021, 11, 8129–8142. [Google Scholar] [CrossRef]
- Liew, L.C.; Katsuda, T.; Gailhouste, L.; Nakagama, H.; Ochiya, T. Mesenchymal stem cell-derived extracellular vesicles: a glimmer of hope in treating Alzheimer’s disease. Int Immunol 2017, 29, 11–19. [Google Scholar] [CrossRef]
- Fatima, F.; Ekstrom, K.; Nazarenko, I.; Maugeri, M.; Valadi, H.; Hill, A.F.; Camussi, G.; Nawaz, M. Non-coding RNAs in Mesenchymal Stem Cell-Derived Extracellular Vesicles: Deciphering Regulatory Roles in Stem Cell Potency, Inflammatory Resolve, and Tissue Regeneration. Front Genet. 2017, 8, 161. [Google Scholar] [CrossRef]
- Wang, S.S.; Jia, J.; Wang, Z. Mesenchymal Stem Cell-Derived Extracellular Vesicles Suppresses iNOS Expression and Ameliorates Neural Impairment in Alzheimer’s Disease Mice. J Alzheimers Dis 2018, 61, 1005–1013. [Google Scholar] [CrossRef]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; Batrakova, E.V. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release 2015, 207, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Kojima, R.; Bojar, D.; Rizzi, G.; Hamri, G.C.; El-Baba, M.D.; Saxena, P.; Ausländer, S.; Tan, K.R.; Fussenegger, M. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson’s disease treatment. Nat Commun 2018, 9, 1305. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Li, Y.; Ji, W.; Zhao, R.; Lu, Z.; Shen, J.; Wu, Y.; Wang, J.; Hao, Q.; Wang, J.; Wang, W.; Yang, J.; Zhang, X. Intranasal Administration of Self-Oriented Nanocarriers Based on Therapeutic Exosomes for Synergistic Treatment of Parkinson’s Disease. ACS Nano 2022, 16, 869–884. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Tu, Z.; Yang, D.; Hu, M.; Zhou, L.; Li, Q.; Yu, B.; Hou, S. Exosomes from hypoxic pre-treated ADSCs attenuate acute ischemic stroke-induced brain injury via delivery of circ-Rps5 and promote M2 microglia/macrophage polarization. Neurosci Lett 2022, 769, 136389. [Google Scholar] [CrossRef]
- Zheng, X.; Sun, K.; Liu, Y.; Yin, X.; Zhu, H.; Yu, F.; Zhao, W. Resveratrol-loaded macrophage exosomes alleviate multiple sclerosis through targeting microglia. J Control Release 2023, 353, 675–684. [Google Scholar] [CrossRef]
- Riazifar, M.; Mohammadi, M.R.; Pone, E.J.; Yeri, A.; Lässer, C.; Segaliny, A.I.; McIntyre, L.L.; Shelke, G.V.; Hutchins, E.; Hamamoto, A.; Calle, E.N.; Crescitelli, R.; Liao, W.; Pham, V.; Yin, Y.; Jayaraman, J.; Lakey, J.R.T.; Walsh, C.M.; Van Keuren-Jensen, K.; Lotvall, J.; Zhao, W. Stem Cell-Derived Exosomes as Nanotherapeutics for Autoimmune and Neurodegenerative Disorders. ACS Nano 2019, 13, 6670–6688. [Google Scholar] [CrossRef]
- Ma, K.; Xu, H.; Zhang, J.; Zhao, F.; Liang, H.; Sun, H.; Li, P.; Zhang, S.; Wang, R.; Chen, X. Insulin-like growth factor-1 enhances neuroprotective effects of neural stem cell exosomes after spinal cord injury via an miR-219a-2-3p/YY1 mechanism. Aging 2019, 11, 12278–12294. [Google Scholar] [CrossRef]
- Liu, W.; Rong, Y.; Wang, J.; Zhou, Z.; Ge, X.; Ji, C.; Jiang, D.; Gong, F.; Li, L.; Chen, J.; Zhao, S.; Kong, F.; Gu, C.; Fan, J.; Cai, W. Exosome-shuttled miR-216a-5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization. J Neuroinflammation 2020, 17, 47. [Google Scholar] [CrossRef]
- Zhou, W.; Silva, M.; Feng, C.; Zhao, S.; Liu, L.; Li, S.; Zhong, J.; Zheng, W. Exosomes derived from human placental mesenchymal stem cells enhanced the recovery of spinal cord injury by activating endogenous neurogenesis. Stem Cell Res Ther 2021, 12, 174. [Google Scholar] [CrossRef]
- Zhang, W.; Hong, J.; Zhang, H.; Zheng, W.; Yang, Y. Astrocyte-derived exosomes protect hippocampal neurons after traumatic brain injury by suppressing mitochondrial oxidative stress and apoptosis. Aging 2021, 13, 21642–21658. [Google Scholar] [CrossRef]
- Wen, L.; Wang, Y.D.; Shen, D.F.; Zheng, P.D.; Tu, M.D.; You, W.D.; Zhu, Y.R.; Wang, H.; Feng, J.F.; Yang, X.F. Exosomes derived from bone marrow mesenchymal stem cells inhibit neuroinflammation after traumatic brain injury. Neural Regen Res 2022, 17, 2717–2724. [Google Scholar] [PubMed]
- Chen, Y.; Li, J.; Ma, B.; Li, N.; Wang, S.; Sun, Z.; Xue, C.; Han, Q.; Wei, J.; Zhao, R.C. MSC-derived exosomes promote recovery from traumatic brain injury via microglia/macrophages in rat. Aging 2020, 12, 18274–18296. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lei, B.; Zhang, E.; Gong, P.; Gu, J.; He, L.; Han, L.; Yuan, Z. Targeted Therapy for Inflammatory Diseases with Mesenchymal Stem Cells and Their Derived Exosomes: From Basic to Clinics. Int J Nanomedicine 2022, 17, 1757–1781. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic Potential of Mesenchymal Cell-Derived miRNA-150-5p-Expressing Exosomes in Rheumatoid Arthritis Mediated by the Modulation of MMP14 and VEGF. J Immunol 2018, 201, 2472–2482. [Google Scholar] [CrossRef] [PubMed]
- You, D.G.; Lim, G.T.; Kwon, S.; Um, W.; Oh, B.H.; Song, S.H.; Lee, J.; Jo, D.G.; Cho, Y.W.; Park, J.H. Metabolically engineered stem cell-derived exosomes to regulate macrophage heterogeneity in rheumatoid arthritis. Sci Adv 2021, 7, eabe0083. [Google Scholar] [CrossRef]
- Li, H.; Feng, Y.; Zheng, X.; Jia, M.; Mei, Z.; Wang, Y.; Zhang, Z.; Zhou, M.; Li, C. M2-type exosomes nanoparticles for rheumatoid arthritis therapy via macrophage re-polarization. J Control Release 2022, 341, 16–30. [Google Scholar] [CrossRef]
- Wu, L.F.; Zhang, Q.; Mo, X.B.; Lin, J.; Wu, Y.L.; Lu, X.; He, P.; Wu, J.; Guo, Y.F.; Wang, M.J.; Ren, W.Y.; Deng, H.W.; Lei, S.F.; Deng, F.Y. Identification of novel rheumatoid arthritis-associated MiRNA-204-5p from plasma exosomes. Exp Mol Med 2022, 54, 334–345. [Google Scholar] [CrossRef]
- Li, S.; Stöckl, S.; Lukas, C. Herrmann M, Brochhausen C, König MA, Johnstone B, Grässel S. Curcumin-primed human BMSC-derived extracellular vesicles reverse IL-1β-induced catabolic responses of OA chondrocytes by upregulating miR-126-3p. Stem Cell Res Ther 2021, 12, 252. [Google Scholar] [CrossRef]
- Xian Bo, S.; Chen, W.; Chang, L.; Hao Ran, Y.; Hui Hui, G.; Ya Kun, Z.; Wu Kun, X.; Hai Tao, F.; Wen Dan, C. The Research Progress of Exosomes in Osteoarthritis, With Particular Emphasis on the Therapeutic Effect. Front Pharmacol 2022, 13, 685623. [Google Scholar] [CrossRef]
- Dong, J.; Li, L.; Fang, X.; Zang, M. Exosome-Encapsulated microRNA-127-3p Released from Bone Marrow-Derived Mesenchymal Stem Cells Alleviates Osteoarthritis Through Regulating CDH11-Mediated Wnt/β-Catenin Pathway. J Pain Res 2021, 14, 297–310. [Google Scholar] [CrossRef]
- Mao, G.; Hu, S.; Zhang, Z.; Wu, P.; Zhao, X.; Lin, R.; Liao, W.; Kang, Y. Exosomal miR-95-5p regulates chondrogenesis and cartilage degradation via histone deacetylase 2/8. J Cell Mol Med 2018, 22, 5354–5366. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Liu, G.; Wu, X. The umbilical cord mesenchymal stem cell-derived exosomal lncRNA H19 improves osteochondral activity through miR-29b-3p/FoxO3 axis. Clin Transl Med 2021, 11, e255. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Liu, D.; Fu, Q. MiR-129-5p shuttled by human synovial mesenchymal stem cell-derived exosomes relieves IL-1β induced osteoarthritis via targeting HMGB1. Life Sci 2021, 269, 118987. [Google Scholar] [CrossRef]
- Qi, H.; Liu, D.-P.; Xiao, D.-W.; Tian, D.-C.; Su, Y.-W.; Jin, S.-F. Exosomes derived from mesenchymal stem cells inhibit mitochondrial dysfunction-induced apoptosis of chondrocytes via p38, ERK, and Akt pathways. Vitr. Cell. Dev. Biol. Anim. 2019, 55, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yan, K.; Ge, G.; Zhang, D.; Bai, J.; Guo, X.; Zhou, J.; Xu, T.; Xu, M.; Long, X.; et al. Exosomes derived from miR-155-5p-overexpressing synovial mesenchymal stem cells prevent osteoarthritis via enhancing proliferation and migration, attenuating apoptosis, and modulating extracellular matrix secretion in chondrocytes. Cell Biol. Toxicol. 2021, 37, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci Rep 2017, 7, 16214. [Google Scholar] [CrossRef]
- He, L.; He, T.; Xing, J.; Zhou, Q.; Fan, L.; Liu, C.; Chen, Y.; Wu, D.; Tian, Z.; Liu, B.; et al. Bone marrow mesenchymal stem cell-derived exosomes protect cartilage damage and relieve knee osteoarthritis pain in a rat model of osteoarthritis. Stem Cell Res Ther 2020, 11, 276. [Google Scholar] [CrossRef]
- Qiu, B.; Xu, X.; Yi, P.; Hao, Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J Cell Mol Med 2020, 24, 10855–10865. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Chen, J.; Qian, D.; Gao, P.; Qin, T.; Jiang, T.; Yi, J.; Xu, T.; Huang, Y.; Wang, Q.; Zhou, Z.; Bao, T.; Zhao, X.; Liu, H.; Zheng, Z.; Fan, J.; Zhao, S.; Li, Q.; Yin, G. Exosomes derived from platelet-rich plasma administration in site mediate cartilage protection in subtalar osteoarthritis. J Nanobiotechnology 2022, 20, 56. [Google Scholar]
- Zhang, S.; Chu, W.C.; Lai, R.C.; Lim, S.K.; Hui, J.H.P.; Toh, W.S. Exosomes derived from human embryonic mesenchymal stem cells promote osteochondral regeneration. Osteoarthr Cartil 2016, 24, 2135–2140. [Google Scholar] [CrossRef]
- Tao, S.-C.; Yuan, T.; Zhang, Y.-L.; Yin, W.-J.; Guo, S.-C.; Zhang, C.-Q. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics 2017, 7, 180–195. [Google Scholar] [CrossRef]
- 83 Qi, X.; Zhang, J.; Yuan, H.; Xu, Z.; Li, Q.; Niu, X.; Hu, B.; Wang, Y.; Li, X. Exosomes Secreted by Human-Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cells Repair Critical-Sized Bone Defects through Enhanced Angiogenesis and Osteogenesis in Osteoporotic Rats. Int J Biol Sci 2016, 12, 836–849. [Google Scholar] [CrossRef]
- Zhu, H.; Ji, J.; Fu, T.; Yang, J.; Gu, Z. The effect of exosomes from bone marrow mesenchymal stem cells on osteoarthritis. Ann Rheum Dis 2018, 77, 893. [Google Scholar]
- Mortaz, E.; Alipoor, S.D.; Varahram, M.; et al. Exosomes in Severe Asthma: Update in Their Roles and Potential in Therapy. BioMed Res Int 2018, 2862187. [Google Scholar] [CrossRef]
- Pei, W.; Li, X.; Bi, R.; Zhang, X.; Zhong, M.; Yang, H.; Zhang, Y.; Lv, K. Exosome membrane-modified M2 macrophages targeted nanomedicine: Treatment for allergic asthma. J Control Release 2021, 338, 253–267. [Google Scholar] [CrossRef]
- Liu, C.; Xiao, K.; Xie, L. Advances in the use of exosomes for the treatment of ALI/ARDS. Front Immunol 2022, 13, 971189. [Google Scholar] [CrossRef]
- Wei, X.; Yi, X.; Lv, H.; Sui, X.; Lu, P.; Li, L.; An, Y.; Yang, Y.; Yi, H.; Chen, G. Microrna-377-3p released by mesenchymal stem cell exosomes ameliorates lipopolysaccharide-induced acute lung injury by targeting RPTOR to induce autophagy. Cell Death Dis 2020, 11, 657. [Google Scholar]
- Tian, J.; Cui, X.; Sun, J.; Zhang, J. Exosomal microrna-16-5p from adipose mesenchymal stem cells promotes Tlr4-mediated M2 macrophage polarization in septic lung injury. Int Immunopharmacol 2021, 98, 107835. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, P.; Goodwin, A.J.; Cook, J.A.; Halushka, P.V.; Chang, E.; Zingarelli, B.; Fan, H. Exosomes from endothelial progenitor cells improve outcomes of the lipopolysaccharide-induced acute lung injury. Crit Care 2019, 23, 44. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Z.; Hu, L.; Gu, W.; Zhu, L. Exosomes derived from endothelial progenitor cells ameliorate acute lung injury by transferring mir-126. Exp Cell Res 2018, 370, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Wang, Z.; Gong, L.; Peng, X; Richard, M.A.; Zhang, J.; Fornage, M.; Alcorn, J.L.; Wang, D.A.; Zhang, J.; Fornage, M.; Alcorn, J.L.; Wang, D. Exosome mir-371b-5p promotes proliferation of lung alveolar progenitor type ii cells by using pten to orchestrate the Pi3k/Akt signaling. Stem Cell Res Ther 2017, 8, 138. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Ni, J.; Shen, G.; Xia, Z.; Zhang, L.; Xia, S.; Pan, S.; Qu, H.; Li, X. Upregulation of endothelial cell-derived exosomal microrna-125b-5p protects from sepsis-induced acute lung injury by inhibiting topoisomerase ii alpha. Inflammation Res 2021, 70, 205–16. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, D:, Wen; Tang, X.; Qi, D.; He, J.; Zhao, Y.; Deng, W.; Zhu, T. Adipose-derived exosomes protect the pulmonary endothelial barrier in ventilator-induced lung injury by inhibiting the Trpv4/Ca(2+) signaling pathway. Am J Physiol Lung Cell Mol Physiol 2020, 318, L723–l41. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Lee, Y.; Ha, J.; Han, S.; Lee, M. Engineering exosomes for pulmonary delivery of peptides and drugs to inflammatory lung cells by inhalation. J Control Release 2021, 330, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Long, J.; Meng, L.; Yang, Z.; Zhang, S.; Zhang, L. Immune cells-derived exosomes: A promising strategy for COVID-19 treatment. Clin Transl Discov 2022, 2, e138. [Google Scholar] [CrossRef]
- Mitrani, M.I.; Bellio, M.A.; Sagel, A.; Saylor, M.; Kapp, W.; VanOsdol, K.; Haskell, G.; Stewart, D.; Abdullah, Z.; Santos, I.; Milberg, J.; Arango, A.; Mitrani, A.; Shapiro, G.C. Case report: Administration of amniotic fluid-derived nanoparticles in three severely ill covid-19 patients. Front Med 2021, 8, 583842. [Google Scholar] [CrossRef]
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19. Stem Cells Dev 2020, 29, 747–754. [Google Scholar] [CrossRef]
- Lightner, A.L.; Sengupta, V.; Qian, S.; Ransom, J.T.; Suzuki, S.; Park, D.J.; Melson, T.I.; Williams, B.P.; Walsh, J.J.; Awili, M. Bone Marrow Mesenchymal Stem Cell-Derived Extracellular Vesicle Infusion for the Treatment of Respiratory Failure From COVID-19: A Randomized, Placebo-Controlled Dosing Clinical Trial. CHEST article in press. 2023. [Google Scholar] [CrossRef]
- Lou, G.; Chen, Z.; Zheng, M.; Liu, Y. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp Mol Med 2017, 49, e346. [Google Scholar] [CrossRef]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J Clin Investig 2005, 115, 209–218. [Google Scholar] [CrossRef]
- Zhang, Z.; Shang, J.; Yang, Q.; Dai, Z.; Liang, Y.; Lai, C.; Feng, T.; Zhong, D.; Zou, H.; Sun, L.; Su, Y.; Yan, S.; Chen, J.; Yao, Y.; Shi, Y.; Huang, X. Exosomes derived from human adipose mesenchymal stem cells ameliorate hepatic fibrosis by inhibiting PI3K/Akt/mTOR pathway and remodeling choline metabolism. J Nanobiotechnology 2023, 21(1), 29. [Google Scholar] [CrossRef] [PubMed]
- Niknam, B.; Baghaei, K.; Mahmoud Hashemi, S.; Hatami, B.; Reza Zali, M.; Amani, D. Human Wharton's jelly mesenchymal stem cells derived-exosomes enriched by miR-124 promote an anti-fibrotic response in an experimental model of liver fibrosis. Int Immunopharmacol 2023, 119, 110294. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Piao, C; Xu, J.; Wang, Y.; Liu, T.; Ma, H.; Wang, H. ADSCs-exo attenuates hepatic ischemia-reperfusion injury after hepatectomy by inhibiting endoplasmic reticulum stress and inflammation. J Cell Physiol 2023, 238, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Lee, E.K.; Yim, J.; Lee, M.H.; Lee, H.; Lee, Y.S.; Seo, W. Exosomes: Nomenclature, Isolation, and Biological Roles in Liver Diseases. Biomol Ther 2023, 31, 253–263. [Google Scholar] [CrossRef]
- Malekpour, K.; Hazrati, A.; Soudi, S.; Roshangar, L.; Pourfathollah, A.A.; Ahmadi, M. Combinational administration of mesenchymal stem cell-derived exosomes and metformin reduces inflammatory responses in an in vitro model of insulin resistance in HepG2 cells. Heliyon 2023, 9(5), e15489. [Google Scholar] [CrossRef]
- Khosrojerdi, A.; Soudi, S.; Hosseini, A.Z.; Khaligh, S.G.; Hashemi, S.M. The combination of mesenchymal stem cell- and hepatocyte-derived exosomes, along with imipenem, ameliorates inflammatory responses and liver damage in a sepsis mouse model. Life Sci 2023, 326, 121813. [Google Scholar] [CrossRef]
- Cai, J.; Tang, D.; Hao, X.; Liu, E.; Li, W.; Shi, J. Mesenchymal stem cell-derived exosome alleviates sepsis- associated acute liver injury by suppressing MALAT1 through microRNA-26a-5p: an innovative immunopharmacological intervention and therapeutic approach for sepsis. Front Immunol 2023, 14, 1157793. [Google Scholar] [CrossRef]
- Ellakany, A.R.; El Baz, H.; Shoheib, Z.S.; Elzallat, M.; Ashour, D.S.; Yassen, N.A. Stem cell-derived exosomes as a potential therapy for schistosomal hepatic fibrosis in experimental animals. Pathog Glob Health 2023, 30, 1–21. [Google Scholar] [CrossRef]
- Cao, Q.; Huang, C.; Chen, X.M.; Pollock, C.A. Mesenchymal Stem Cell-Derived Exosomes: Toward Cell-Free Therapeutic Strategies in Chronic Kidney Disease. Front. Med 2022, 9, 816656. [Google Scholar] [CrossRef]
- Charles, C.; Ferris, A.H. Chronic Kidney Disease. Prim Care 2020, 47, 585–595. [Google Scholar] [CrossRef]
- Corrêa, R.R.; Juncosa, E.M.; Masereeuw, R.; Lindoso, R.S. Extracellular Vesicles as a Therapeutic Tool for Kidney Disease: Current Advances and Perspectives. Int J Mol Sci 2021, 22, 5787. [Google Scholar] [CrossRef] [PubMed]
- Nagaishi, K.; Mizue, Y.; Chikenji, T.; Otani, M.; Nakano, M.; Konari, N.; Fujimiya, M. Mesenchymal stem cell therapy ameliorates diabetic nephropathy via the paracrine effect of renal trophic factors including exosomes. Sci Rep 2016, 6, 34842. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, N.; Ahmed, I.A.; Hussien, N.I.; Dessouky, A.A.; Farid, A.S.; Elshazly, A.M.; Mostafa, O.; Gazzar, W.B.E.; Sorour, S.M.; Seleem, Y.; Hussein, A.M.; Sabry, D. Mesenchymal stem cell-derived exosomes ameliorated diabetic nephropathy by autophagy induction through the mTOR signaling pathway. Cells 2018, 7, 226. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Shi, Y.; Gong, J.; Zhao, L.; Li, Y.; He, Q.; Huang, H. Exosome secreted from adipose-derived stem cells attenuates diabetic nephropathy by promoting autophagy flux and inhibiting apoptosis in podocyte. Stem Cell Res Ther 2019, 10, 95. [Google Scholar] [CrossRef]
- Liu, B.; Hu, D.; Zhou, Y.; Yu, Y.; Shen, L.; Long, C.; Butnaru, D.; Timashev, P.; He, D.; Lin, T.; Xu, T.; Zhang, D.; Wei, G. Exosomes released by human umbilical cord mesenchymal stem cells protect against renal interstitial fibrosis through ROS-mediated P38MAPK/ERK signaling pathway. Am J Transl Res 2020, 12, 4998–5014. [Google Scholar]
- Ishiy, C.; Ormanji, M.S.; Maquigussa, E.; Ribeiro, R.S.; da Silva Novaes, A.; Boim, MA. Comparison of the effects of mesenchymal stem cells with their extracellular vesicles on the treatment of kidney damage induced by chronic renal artery stenosis. Stem Cells Int. 2020, 2020, 8814574. [Google Scholar] [CrossRef]
- Liu, L.; Wu, Y.; Wang, P.; Shi, M.; Wang, J.; Ma, H.; Sun, D. PSC-MSC-derived exosomes protect against kidney fibrosis in vivo and in vitro through the SIRT6/β-catenin signaling pathway. Int J Stem Cells 2021, 14, 310–19. [Google Scholar] [CrossRef]
- Hao, Y.; Miao, J.; Liu, W.; Cai, K.; Huang, X.; Peng, L. Mesenchymal stem cell-derived exosomes carry microRNA-125a to protect against diabetic nephropathy by targeting histone deacetylase 1 and downregulating endothelin-1. Diabetes Metab Syndr Obes 2021, 14, 1405–1418. [Google Scholar] [CrossRef]
- Jiang, Z.Z.; Liu, Y.M.; Niu, X.; Yin, J.Y.; Hu, B.; Guo, S.C.; Fan, Y.; Wang, Y.; Wang, N.S. Exosomes secreted by human urine-derived stem cells could prevent kidney complications from type I diabetes in rats. Stem Cell Res Ther 2016, 7, 24. [Google Scholar] [CrossRef]
- Duan, Y.R.; Chen, B.P.; Chen, F.; Yang, S.X.; Zhu, C.Y.; Ma, Y.L. Exosomal microRNA-16-5p from human urine-derived stem cells ameliorates diabetic nephropathy through protection of podocyte. J Cell Mol Med 2021, 25, 10798–813. [Google Scholar] [CrossRef]
- Gao, F.; Zuo, B.; Wang, Y.; Li, S.; Yang, J.; Sun, D. Protective function of exosomes from adipose tissue-derived mesenchymal stem cells in acute kidney injury through SIRT1 pathway. Life Sci 2020, 255, 117719. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, Y.; Li, S.; Zuo, B.; Zhang, X.; Wang, F.; Sun, D. Exosomes derived from GDNF-modified human adipose mesenchymal stem cells ameliorate peritubular capillary loss in tubulointerstitial fibrosis by activating the SIRT1/eNOS signaling pathway. Theranostics 2020, 10, 9425–9442. [Google Scholar] [CrossRef] [PubMed]
- Crowley, E.; Muise, A. Inflammatory bowel disease: what very early onset disease teaches us. Gastroenterol Clin 2018, 47, 755–772. [Google Scholar] [CrossRef]
- Shapiro, J.M.; Subedi, S.; LeLeiko, N.S. Inflammatory bowel disease. Pediatr Rev 2016, 37, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Ocansey, D.K.W.; Zhang, L.; Wang, Y.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. Exosome-mediated effects and applications in inflammatory bowel disease. Biological Reviews 2020, 95, 1287–1307. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, X.; Xiang, Y.; Fan, D.; An, Q.; Yue, G.; Jin, Z.; Ding, J.; Hu, Y.; Du, Q.; Xu, J.; Xie, R. Exosome-mediated effects and applications in inflammatory diseases of the digestive system. Eur J Med Res 2022, 27, 163. [Google Scholar] [CrossRef]
- Cai, X.; Zhang, Z.Y.; Yuan, J.T.; Ocansey, D.K.W.; Tu, Q.; Zhang, X.; Qian, H.; Xu, W.R.; Qiu, W.; Mao, F. hucMSC-derived exosomes colitis by regulating macrophage pyroptosis via the miR-378a-5p/ NLRP3 axis. Stem Cell Res Ther 2021, 12, 416. [Google Scholar] [CrossRef]
Figure 1.
Production of extracellular vesicles: exosomes (EXs), microvesicles (MVs) and apoptotic bodies .
Figure 1.
Production of extracellular vesicles: exosomes (EXs), microvesicles (MVs) and apoptotic bodies .
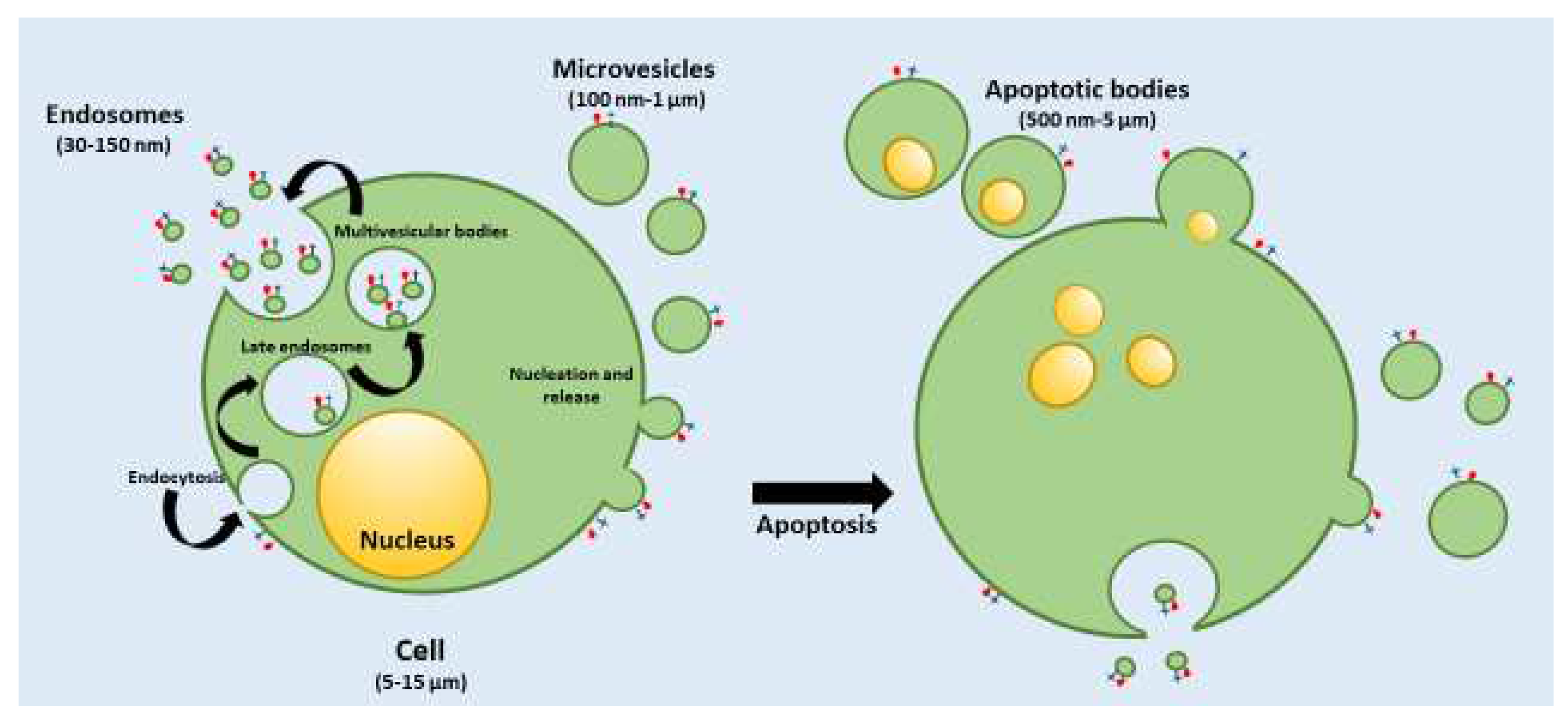
Figure 2.
EXs structure, components decorating the membrane and constituents of the inner core.
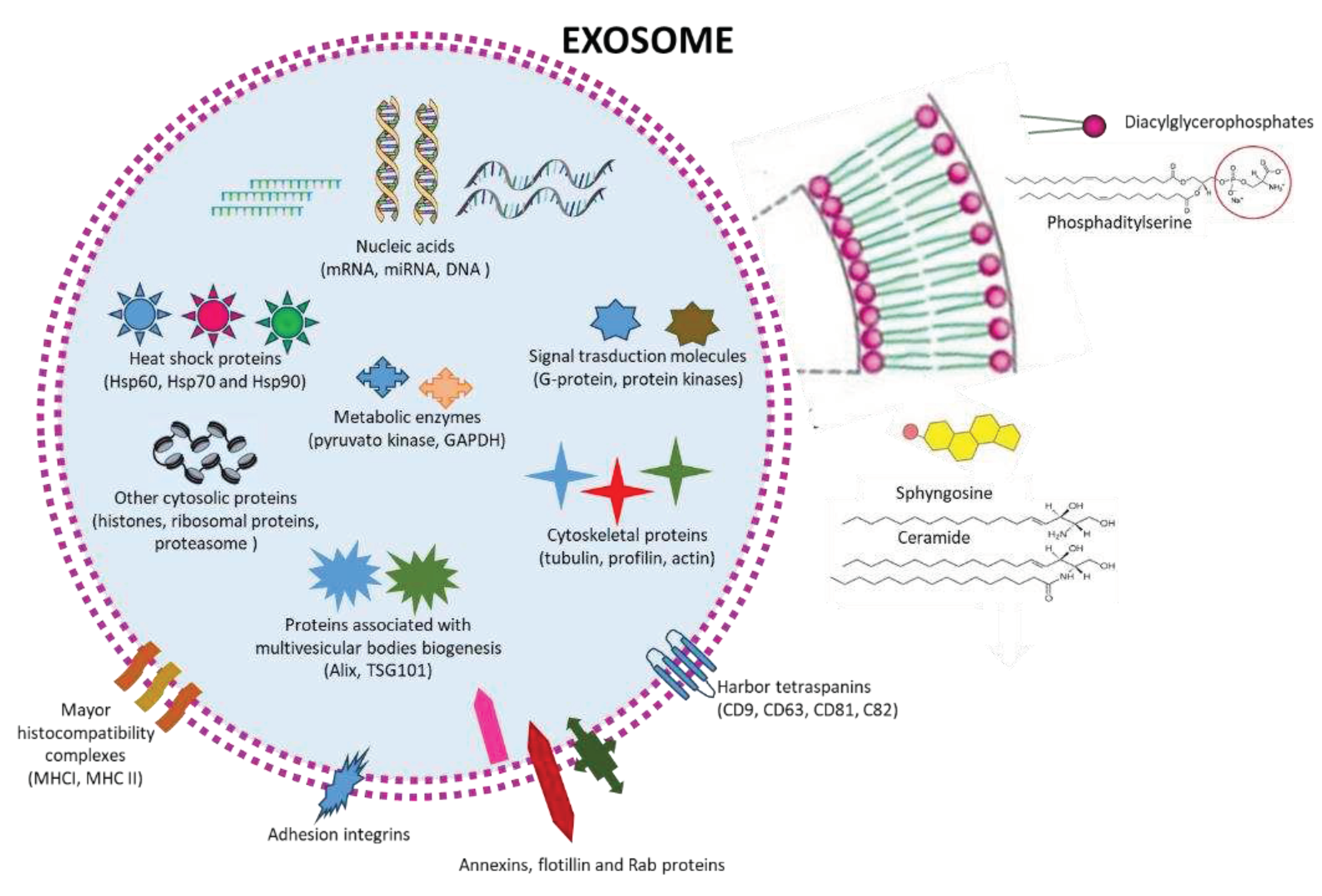
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



