
Submitted:
11 August 2023
Posted:
11 August 2023
You are already at the latest version
Abstract
The present study investigated the insulin-like growth factors (IGFs) and their receptors and binding proteins among different pig breeds during weaning. Sixty Duroc (DR), Taoyuan black (TB), and Xiangcun black (XB) piglets (20 piglets per breed) were selected at 21 and 24 days of age (three days of post-weaning) to analyze organ indices, plasma concentrations of IGF and IGF-binding proteins (IGFBPs) using ELISA kits, and gene expression of IGF system-related components in different tissues. The plasma IGFBP-3 level in TB piglets was higher (P > 0.05) than in the XB and DR piglets at 21 days of age. At 21 days of age, compared with the DR piglets, the IGF-1 expression was lower (P < 0.05) in the kidney but was higher (P < 0.05) in the spleen of XB and TB piglets. At 24 days of age, the IGF-1 expression was higher (P < 0.05) in the kidney of TB piglets than in the XB and DR piglets, while IGFBP-3 in the stomach and IGFBP-4 in the liver of XB and TB piglets were lower (P < 0.05) compared with the DR piglets. Weaning down-regulated (P < 0.05) the IGF-1 expression in the jejunum, spleen, and liver of piglets, while up-regulated (P < 0.05) IGFBP-3 expression in the stomach, IGFBP-4 in the liver, IGFBP-5 in the ileum, and IGFBP-6 in the jejunum of DR piglets. Spearman’s correlation analysis showed a negative correlation (P < 0.05) between plasma IGFBP-2 and IGFBP-5 and the organ index of piglets. Collectively, there were significant differences in IGF system components among different pig breeds. The IGF system components were altered during weaning, which might be involved in weaning stress to decrease the growth of piglets.
Keywords:
Chinese indigenous pig
; insulin-like growth factors
; receptors
; binding protein
; weaning
1. Introduction
Growth rate is one of the most important traits in domestic animal breeding programs and is also considered an important factor in determining meat production [1]. Pork is a good and comparatively economical source of animal protein and is the most consumed meat in China [2]. Over the past decades, antibiotics have been used as growth promoters to prevent diseases and improve pork production in the pig industry [3]. Excessive use of antibiotics leads to increased antibiotic resistance in pathogenic bacteria and food residues in the pig industry [4]. In response, many European countries and China have banned antibiotics for animal growth promotion [5-6]. Therefore, understanding the growth and development profiles of pigs is of great significance to the improvement of pork production and pig breeding, and developing alternatives to antibiotic growth promoters.
The growth and development of animals is regulated by numerous physiological networks [7]. Hormones are primarily responsible for the integrated communication of several physiological systems associated with growth and development regulation. Among these hormones, the insulin-like growth factor (IGF) signaling system, including IGF, IGF receptors (IGF-Rs), and IGF-binding proteins (IGFBPs), is considered to be one of the key hormones of the growth axis [8]. In particular, the IGF system is vital part in initiating the connection between nutrition and postnatal growth and development of various tissues and organs through secretory, paracrine, and endocrine actions [9-10]. Numerous environmental factors, such as nutritional status, stress, and temperature, significantly affect the growth hormone-IGF system [11]. In the commercial swine industry, suckling to weaning is the most stressful period for piglets due to the rapid changes in diets, physiology, and social environments, leading to weaning stress [12]. Research evidence indicated that weaning stress can limit the growth rate of piglets [13] and decrease serum IGF-1 and IGF-2 concentrations [14]. Hence, the changes in serum insulin-like growth factors (IGFs) concentrations may affect the growth rate of weaned piglets. Previously, it has been found that piglets exhibited stress damage on the 3rd day after weaning [15]. However, the changes in the IGF system of piglets during this weaning period (3rd day after weaning) are still unclear.
It is well known that Duroc (DR) pig, a commercial pig, has the characteristics of faster growth, higher ratio of lean meat, and lower fat content [16]. The Chinese indigenous pig breeds are distinguished by presenting high prolificacy, better meat quality, and roughage feeding tolerance; however, the growth rate of these breeds is slower than that of the DR pigs [17]. Different pig breeds have been crossed to obtain heterosis and complementarity [18]. Xiangcun black (XB) pig is derived from a crossbreed of DR pig as the paternal lines with TB pig as the maternal lines. The XB pig has the characteristics of strong adaptability to roughage feeding, higher intramuscular fat content, and a faster growth rate [19]. The availability of this breed provides a model for exploring the effects of the IGF system on the growth of pigs. Therefore, we hypothesized that the difference in growth and development of different pig breeds might be related to the regulation of IGFs, IGF-Rs, and IGFBPs. Gene expression profiling studies have focused more attention on such factors as breed, age, and tissue specificity [9]. Previously, we found that weaning could decrease the body weight (WB) of DR, TB, and XB piglets [20]. Therefore, the present study focused on weaned DR, TB, and XB piglets to determine the changes in plasma IGF component concentrations and the expression levels of IGF, IGF-Rs, and IGFBPs in different tissues of piglets. The study will offer an informative perspective on the growth and development of different pig breeds and avenues to alleviate the adverse effects of weaning stress on the growth and development of piglets.
2. Materials and Methods
2.1. Animals and Experimental Design
Thirty litters of newborn DR, TB, and XB piglets (10 litters from each breed) were obtained from sows with similar parity (2−3) and litter size (9−11). Two piglets close to the average BW of litter were chosen from each litter, and each pig breed consisted of 20 piglets (TB and XB piglets were half female and half castrated male, and DR piglets were all castrated male). The piglets only received sows’ milk during lactation. After weaning at 21 days of age, the piglets were fed four times (8:00, 12:00, 14:00, and 17:00) daily with a creep feed. The piglets had ad libitum access to feed and water during the trial. Piglets were kept in individual pens at a controlled temperature (23−26°C) and forced-air ventilation. The piglets were not vaccinated with any vaccine during the trial.
2.2. Sample Collection
At 21 and 24 days of age, ten piglets from each breed were weighed and slaughtered for sampling 12 h after the last feeding. The average BW of DR, TB, and XB piglets were 6.01±1.91, 5.06±1.12, and 3.42±1.12 kg at 21 days of age, and 5.74±1.59, 4.37±1.78, and 3.22±0.99 kg at 24 days of age, respectively. In order to obtain organ index and IGF system expressions in organs, piglets were exsanguinated after an intravenous injection of 4% sodium pentobarbital (40 mg/kg). Blood (10 mL) samples were taken from the anterior vena cava into heparin sodium tubes (Aosaite, Shangdong, China) and centrifuged at 3500 × g for 15 min at 4°C to obtain plasma for IGFs indices analysis. The kidney, spleen, liver, LD muscle, stomach, duodenum, jejunum, and ileum were separated and weighed to calculate organ indices: organ weight (g)/body weight (kg). A 5-cm section of posterior segments of the duodenum, jejunum, and ileum was taken, placed in liquid nitrogen, and kept at –80°C for total RNA purification.
2.3. Measurement of IGF System Component in Plasma
Plasma levels of IGF-1, IGF-2, IGFBP-1, IGFBP-2, IGFBP-3, and IGFBP-5 were determined by the porcine ELISA kits (Shanghai Huyu, Shanghai, China). The absorbance values were read on the Multiscan Specturum Spectrophotometer (Tecan, Infinite M200 Pro, Männedorf, Switzerland).
2.4. Expression Analysis of IGFs System Component Genes
Total RNA was extracted from the stomach, duodenum, jejunum, ileum, liver, kidney, spleen, and LD muscle tissues using the TRIzol reagent (Accurate Biotechnology Co., Ltd. Hunan, China). The concentration and purity of the RNA were determined using a NanoDrop 2000 spectrophotometer (Thermo Fischer Scientific, Waltham, MA, USA). RNA was converted into cDNA by reverse transcription with PrimeScript RT Reagent Kit plus gDNA Eraser (Accurate Biotechnology Co., Ltd. Hunan, China). Real-time PCR assays were performed on the Light Cycler R 480 II Real-Time PCR System (Roche, Basel, Switzerland). The PCR conditions were as follows: 95°C for 5 min, then 40 cycles of denaturation at 95°C for 5 s and annealing at 60°C for 30 s, with a final extension at 72°C for 30 s. Primers for porcine IGF system-related genes and housekeeping gene β-actin were synthesized by Tsingke Biotechnology Co., Ltd. (Beijing, China) and are shown in Table 1. Gene expression was normalized against β-actin and determined using the 2−ΔΔCt method [21].
2.5. Statistical Analysis
The data were analyzed by ANOVA using a 2×2 factorial treatment arrangement with SPSS 26.0 (SPSS Inc., Chicago, IL, USA). The model included weaning, and breed, and their interactions. Multiple comparisons of means among different groups were performed with Tukey’s post-hoc test for valid interaction with a significance value at P < 0.05. Data are expressed as mean ± standard error of the mean (SEM). Spearman’s correlation analysis was performed to determine the relationships among plasma IGF parameters, BW, and organ indices, as well as IGF system component gene expressions and organ indices in the three pig breeds.
3. Results
3.1. Weaning Effects on Organ Indices in Different Breeds of Piglets
Organ indices of different breeds of piglets are listed in Table 2. The stomach and duodenum indices of the XB piglets were higher (P < 0.05) compared with the TB and DR piglets, regardless of weaning. Weaning increased (P < 0.05) the stomach index of piglets, regardless of pig breed. There was no interaction (P > 0.05) between the weaning and pig breed on the stomach, duodenum, jejunum, ileum, kidney, spleen, and liver indices.
3.2. Weaning Effects on Plasma IGFs and IGFBPs Concentration in Different Breeds of Piglets
Plasma IGFs and IGFBPs concentration in different breeds of piglets are presented in Table 3. The plasma IGF-1 concentration was higher (P < 0.05) in the XB piglets compared with the TB and DR piglets, regardless of weaning. At 21 days of age, plasma IGFBP-3 concentration was elevated (P < 0.05) in the TB piglets compared with the XB and DR piglets. Weaning reduced (P < 0.05) the plasma IGFBP-3 concentration in the TB piglets. There was an interaction (P < 0.05) between weaning and pig breed on plasma IGFBP-3 concentration.
3.3. Weaning Effects on IGF-Related Gene Expressions in Different Breeds of Piglets
Gene expressions related to IGFs in the stomach, duodenum, jejunum, ileum, kidney, liver, spleen, and LD muscle of different breeds of piglets are listed in Table 4. At 21 days of age, IGF-1 expression was down-regulated in the kidney and up-regulated in the spleen of the XB and TB piglets compared with the DR piglets (P < 0.05). The IGF-1 expression was down-regulated (P < 0.05) in the liver of the XB piglets compared with the TB and DR piglets. At 24 days of age, IGF-1 expression was up-regulated (P < 0.05) in the kidney of the TB piglets compared with the XB and DR piglets. At 21 days of age, IGF-1 expression was down-regulated (P < 0.05) in the jejunum, spleen, and liver of piglets, regardless of pig breed, as well as IGF-2 expression in the stomach, kidney, and spleen. The IGF-1 expression in the jejunum, kidney, spleen, and liver showed weaning-breed interactions (P < 0.05). However, there was no interaction (P > 0.05) between weaning and pig breed on IGF-2 expression.
3.4. Weaning Effects on IGF-Receptors (IGF-Rs) Gene Expressions in Different Breeds of Piglets
The expressions of IGF-Rs genes in different breeds of piglets are presented in Table 5. The IGF-1R expression was up-regulated (P < 0.05) in the liver of the XB and TB piglets compared with the DR piglets at 21 days of age, as well as in the XB piglets compared with the TB and DR piglets at 24 days of age. The IGF-1R expression was up-regulated (P < 0.05) in the ileum of the XB piglets compared with the TB and DR piglets regardless of weaning, whereas it was down-regulated (P < 0.05) in the spleen (three breeds of piglets) and liver (TB piglets) at 21 days of age compared with the 24 days of age. There was an interaction (P < 0.05) between weaning and pig breed on IGF-1R expression in the liver.
In comparison to the TB and DR piglets, XB piglets exhibited down-regulation (P < 0.05) of the IGF-2R expression in the kidney at 21 days of age; however, IGF-2R expression in the spleen and liver of XB piglets was up-regulated (P < 0.05) regardless of weaning. The IGF-2R expression was up-regulated (P < 0.05) in the stomach and kidney of different breeds of piglets. There was an interaction (P < 0.05) between weaning and pig breeds on IGF-2R expression in the ileum and liver.
3.5. Weaning Effects on IGF Binding Proteins (IGFBPs) Expression in Different Breeds of Piglets
The IGFBPs expressions in piglets are shown in Table 6. The IGFBP-1 expression in the liver was up-regulated, whereas it was very low in other tissues in the three breeds of piglets. Therefore, only the data for IGFBP-1 expression in the liver are presented in Table 6. The IGFBP-1 expression in the liver of the XB and TB piglets was up-regulated (P < 0.05) compared with the DR piglets regardless of weaning. There was no interaction (P > 0.05) between weaning and pig breed for IGFBP-1 expression in all tested tissues.
At 21 days of age, IGFBP-2 expression was up-regulated (P < 0.05) in the duodenum, ileum, kidney, and spleen of the TB piglets compared with the XB and DR piglets. The IGFBP-2 expression was up-regulated (P < 0.05) in the liver of the XB and TB piglets compared with the DR piglets. At 24 days of age, IGFBP-2 expression was down-regulated (P < 0.05) in the LD muscle of the XB piglets compared with the DR piglets. Weaning down-regulated the IGFBP-2 expression in the ileum of the XB piglets, as well as in the duodenum, ileum, and spleen of the TB piglets, but it was up-regulated in the kidney of the XB and DR piglets and the LD muscle of DR piglets (P < 0.05). There were interactions (P < 0.05) between weaning and breed for IGFBP-2 expressions in the duodenum, ileum, kidney, spleen, liver, and LD muscle.
At 21 days of age, IGFBP-3 expression was up-regulated (P < 0.05) in the duodenum and liver of the TB piglets compared with the XB and DR piglets. At 24 days of age, IGFBP-3 expression was down-regulated (P < 0.05) in the stomach of the XB and TB piglets compared with the DR piglets. Weaning down-regulated the IGFBP-3 expression in the duodenum and liver of the TB piglets, but it was up-regulated in the stomach of the DR piglets (P < 0.05). There were interactions (P < 0.05) between weaning and breed for IGFBP-3 expressions in the stomach, duodenum, and liver.
The IGFBP-4 expression was down-regulated in the ileum, whereas it was up-regulated in the liver of the XB and TB piglets compared with the DR piglets at 21 days of age (P < 0.05). At 24 days of age, IGFBP-4 expression was down-regulated (P < 0.05) in the liver of the XB and TB piglets compared with the DR piglets. Weaning down-regulated (P < 0.05) the IGFBP-4 expression in the ileum of three pig breeds and the liver of XB and TB piglets, whereas it was up-regulated (P < 0.05) in the duodenum and jejunum, regardless of breed, and the liver of the DR piglets. There were interactions (P < 0.05) between weaning and breed for IGFBP-4 expressions in the ileum and liver.
In comparison to the DR piglets, IGFBP-5 expression was down-regulated in the ileum but up-regulated in the kidney and liver of the XB and TB piglets at 21 days of age (P < 0.05). The IGFBP-5 expression was up-regulated (P < 0.05) in the duodenum and spleen of the XB piglets compared with the TB and DR piglets, regardless of weaning. Weaning up-regulated the IGFBP-5 expression in the ileum of the DR piglets, as well as in the kidney and liver of the XB and TB piglets, while it was up-regulated in the stomach, spleen, and LD muscle of piglets, regardless of breed. There were interactions (P < 0.05) between weaning and breed for IGFBP-5 expressions in the ileum, kidney, and liver.
At 21 days of age, IGFBP-6 expression was up-regulated (P < 0.05) in the liver of the TB piglets compared with the XB and DR piglets, as well as in the LD muscle of the XB and TB piglets compared with the DR piglets. At 24 days of age, IGFBP-6 expression was down-regulated (P < 0.05) in the stomach, jejunum, and ileum of the XB piglets compared with the DR piglets. Weaning down-regulated the IGFBP-6 expression in the ileum and LD muscle of the XB and TB piglets, while it was up-regulated in the jejunum of the TB and DR piglets (P < 0.05). The IGFBP-6 expression in the stomach, jejunum, ileum, liver, and LD muscle showed weaning-breed interaction (P < 0.05).
3.6. Weaning Effects on IGFs Expressions in Different Tissues of the Same Breed of Piglets
IGFs expressions in different tissues of the same breed of piglets at 21 and 24 days of age are shown in Figure 1. At 21 days of age, IGF-1 expression in the liver of different breeds of piglets was up-regulated (P < 0.05) in comparison to the other tissues. At 24 days of age, IGF-1 expression in the liver of DR piglets was up-regulated (P < 0.05) compared with the other tissues, as well as in the liver, stomach, and jejunum of TB piglets and the stomach of XB piglets.
At 21 days of age, IGF-2 expression was up-regulated (P < 0.05) in the liver and kidney of the DR piglets, as well as in the liver of the XB and TB piglets compared to the other tissues. At 24 days of age, IGF-2 expression was up-regulated (P < 0.05) in the liver of the DR piglets and in the liver and kidney of the XB and TB piglets compared to the other tissues.
The IGF-1R expression in the kidney of different breeds of piglets was up-regulated (P < 0.05), as well as IGF-2R expression in the kidney of the DR piglets compared with the other tissues at 21 and 24 days of age. At 21 days of age, IGF-2R expression was up-regulated (P < 0.05) in the kidney and liver of the TB piglets, as well as in the liver of the XB piglets in comparison to the other tissues. At 24 days of age, IGF-2R expression was up-regulated (P < 0.05) in the kidney of the TB and XB piglets in comparison to the other tissues.
The IGFBP-2 expression in the liver was up-regulated (P < 0.05) in different breeds of piglets, as well as IGFBP-3 expression in the kidney of the DR piglets compared with the other tissues at 21 and 24 days of age. At 21 days of age, IGFBP-3 expression was up-regulated (P < 0.05) in the liver of the TB and XB piglets in comparison to the other tissues. At 24 days of age, IGFBP-3 expression was up-regulated (P < 0.05) in the kidney of the TB piglets, as well as in the liver and kidney of the XB piglets compared with the other tissues.
The IGFBP-4 expression was up-regulated (P < 0.05) in the kidney of the DR piglets at 21 days of age, as well as in the liver at 24 days of age. At 21 and 24 days of age, IGFBP-4 expression was up-regulated (P < 0.05) in the liver of the TB and XB piglets, as well as IGFBP-5 and IGFBP-6 expressions in the kidney of the three breeds of piglets, when compared with the other tissues.
3.7. Correlations between Plasma Concentrations of IGFs System Components, BW and Organ Indices, as Well as IGFs System Components Gene Expression and Organ Indices of Different Piglet Breeds
Spearman’s correlation analysis was performed to assess the correlations between plasma concentrations of IGFs system components, BW, and organ indices of different breeds of piglets (Figure 2). In DR piglets, a positive correlation was included between plasma IGF-1 concentration with BW(R = 0.479, P < 0.05), whereas a negative correlation was found between plasma IGFBP-2 with duodenum index (R = −0.530, P < 0.05) and plasma IGFBP-5 with stomach index (R = −0.461, P < 0.05). In TB piglets, the positive correlation (R = −0.530, P < 0.05) included between plasma IGFBP-1 with stomach index (R = 0.444, P < 0.05), and the negative correlation included between IGFBP-1 with spleen index (R = −0.457, P < 0.05). In addition, the negative correlation included between plasma IGFBP-2 with kidney index (R = −0.481, P < 0.05) and IGFBP-5 with jejunum (R = −0.654, P < 0.05) and ileum indices (R = −0.506, P < 0.05) in the TB piglets. In XB piglets, the negative correlation included plasma IGFBP-1 with BW (R = −0.553, P < 0.05) and plasma IGFBP-2 with jejunum index (R = −0.449, P < 0.05).
The correlations between IGF system components gene expressions and organ indices are shown in Figure 3. In DR piglets, the IGF-1R expression was negatively correlated with the duodenum index (R = −0.454, P < 0.05). In TB piglets, the IGF-1 gene expression was negatively correlated with the duodenum index (R = −0.513, P < 0.05), as well as IGF-1R gene expression and kidney index (R = −0.446, P < 0.05). IGFBP-4 and IGFBP-6 gene expressions were positively correlation with the ileum index (R = −0.494, −0.503, P < 0.05). In XB piglets, IGF-2 gene expression was positively correlated with the duodenum index (R = 0.603, P < 0.05), whereas IGF-2 gene expression was negatively correlated with the stomach index (R = −0.547, P < 0.05) and IGF-2R gene expression with the ileum index (R = −0.543, P < 0.05).
4. Discussion
Weaning is one of the most stressful events of the pig’s life and is often associated with growth retardation [23]. The IGFs system is essential for the growth and development of animals [24]. It motivates interest in evaluating the plasma concentrations of IGF system components and their gene expressions in specific tissues of the TB, XB, and DR piglets during weaning. We found several significant differences in the IGF system components and their gene expressions among different breeds of piglets. Weaning altered the IGF system components expression in various tissues, including the stomach, duodenum, jejunum, ileum, kidney, liver, and LD muscle of piglets.
IGF-1 and IGF-2 are IGF ligands with various functions in animals [25]. The IGF-1 circulates in the blood to regulate the overall growth rate, while IGF-1 within tissues enhances cellular growth, differentiation, and survival. The IGF-2 concentration during the prenatal period affects fetal growth and development. In addition, IGF-2 has important tissue-specific functions in the finishing pigs, such as maintaining the stem cell population [26]. In the present study, plasma IGF-1 level in the XB pigs was higher than in the TB and DR piglets. Our previous study showed that XB and TB piglets have lower BW while having higher nitrogen metabolism ability than the DR piglets [20]. A previous study reported that IGF-1 promotes protein synthesis [27]. Therefore, we speculated that the higher IGF-1 of XB piglets is used for protein synthesis, while IGF-1 of DR pigs may be contributing to growth regulation. However, further studies are necessary to explore the exact mechanism. In the present study, the effects of weaning on plasma IGF-1 and IGF-2 concentrations in different breeds of piglets were not significant. However, Matteri et al. [14] reported that weaning reduced serum IGF-1 and IGF-2 concentrations in pigs. These discrepancies might be explained by the pig breed, physiological stage, and experimental conditions.
IGF-1 and IGF-2 are essential for whole-body growth, physiology, and metabolism [26]. IGF-1 and IGF-2 were mainly upregulated in the liver of piglets in the present study, which is in agreement with a previous study [28]. Weaning decreased the IGF-1 expression in the jejunum, spleen, and liver, as well as IGF-2 expression in the kidney and spleen of piglets regardless of breed. These findings indicated IGF expressions were reduced in organs with substance metabolism or immune function. Previously, it has been found that weaning adversely affects the nutritional metabolism and immune function of piglets [29]. Moreover, Zhong et al. [30] found that IGF-1 injection promotes nutrient utilization and immune function in the stomach of aquatic animals. These results suggest that lower IGFs level may also affect the digestion, absorption, and immunity in weaned piglets. The IGF-1 expression was up-regulated in the stomach of the XB piglets, as well as in the stomach and jejunum of the TB piglets at 24 days of age in this study, suggesting that the XB and TB piglets may be more capable of resisting weaning stress than the DR piglets. Feng et al. [31] also reported that the proportions of anti-stress sensitive genes genotype (Hal-NN) in Chinese indigenous pig breeds were significantly elevated compared with commercial pig breeds.
IGFs bind to IGFRs to active mammalian target of rapamycin or mitogen-activated protein kinase signaling pathways, leading to metabolic or mitogenic outcomes [32]. The IGF-1R exists in various cells and tissues and possesses a high binding affinity for IGF-1 and IGF-2 [33], while IGF-2R has no catalytic function and therefore does not activate intracellular signaling processes [34]. The combination of IGFs and IGF-1R is essential for regulating the growth, endocrine, and metabolism of animals [35]. Reindl et al. [11] reported that IGFRs were expressed in the muscle, spleen, liver, kidney, intestine, and heart. In the present study, IGF-1R expression was mainly up-regulated in the kidney of different breeds of piglets, and IGF-1R expression was up-regulated in the liver of the XB piglets compared with the DR piglets at 21 and 24 days of age. A previous study reported that the up-regulated IGF-1R expression markedly contributed to liver regeneration [36]. Those results are consistent with our previous findings, which indicated a higher liver index of the XB piglets than the DR piglets at 21 days of age [22]. Overall, these findings support the hypothesis that up-regulated IGF-1R expression is one of the reasons for the strong anti-stress capability of Chinese indigenous pigs during weaning.
The activity of IGFs is mediated by the high affinity of IGFBPs that temporal-spatially maintain normal metabolism [37]. To prolong their half-lives and modulate tissue access, IGFBPs bind to IGF-1 and IGF-2, thereby regulating the IGF function. Six IGFBPs isoforms (IGFBP-1−6) have distinct functions to activate or suppress the actions of IGFs [38]. IGFBP-3, one of the most abundant IGFBP in the blood, binding approximately 90% of IGF-1 [39], can block IGF action and inhibit cell growth [40]. In the present study, plasma IGFBP-3 concentration in the TB piglets was higher compared with the DR and XB piglets at 21 days of age, which may be one of the reasons for the slow growth rate of TB piglets. In addition, weaning reduced the plasma IGFBP-3 concentration in the TB piglets, suggesting that TB piglets can resist the adverse effects of weaning stress on growth.
IGFBPs predominantly inhibit IGFs action and are expressed in several tissues. In the present study, IGFBPs expressions in different tissues differed with pig breeds, which may result in selection pressure on obtaining different phenotypes related to these pig breeds [9]. A previous study also reported that the changes in IGFBPs of calves were related to nutrient intake [41]. In the present study, weaning down-regulated IGFBP-2, IGFBP-3, IGFBP-4, and IGFBP-6 expressions in specific tissues of the XB and TB piglets but up-regulated in the DR piglets, suggesting that TB and XB piglets have higher anti-stress ability. Nishihara et al. [42] also indicated that down-regulated IGFBP-2, IGFBP-3, and IGFBP-6 expressions could activate IGFs in rumen epithelial cells and promote papillae growth in weaned calves. It is worth noting that weaning up-regulated the IGFBP-5 expression in several tissues of piglets in the present study. Previously, it has been reported that IGFBP-5 expression was controlled by weaning in Japanese black calves [43]. However, IGFBP-5 not only inhibits IGFs action, but also has the potential for IGF action [44]. Hence, the function of IGFBP-5 needs to be further explored.
The IGF system regulates the growth and development of animals. In this study, Spearman’s correlation analysis revealed that plasma IGF-1 concentration was positively correlated with the BW of the DR piglets, which was confirmed by the fact that DR pigs have a faster growth rate than Chinese indigenous pigs. In addition, plasma IGFBP-2 concentration was negatively correlated with the duodenum index of the DR and XB piglets and the kidney index of the TB piglets, as well as plasma IGFBP-5 concentration with the stomach index of the DR piglets and the jejunum and ileum indexes of the TB piglets. These findings suggest that IGFBPs may regulate the growth and development of organs. In the present study, there was also a negative correlation between IGF-Rs and organ indexes, which might be related to the compensatory response of piglets to weaning stress. The specific mechanism needs to be further explored.
5. Conclusions
The tissue expression profiles of IGF system components varied among different breeds of pigs, and IGFBPs have inhibitory effects on organ development. Weaning stress slowed down pigs’ growth by down-regulating the IGF-1 expression. The expressions of IGF system components in response to weaning stress differed among breeds; particularly, IGFBPs expressions were down-regulated in the XB and TB piglets during weaning, which might be associated with the resistance response to the effects of weaning stress on slowing down the growth of piglets. Further studies are needed to confirm it. These findings have important implications for understanding the growth of different breeds of pigs during weaning and also provide a new perspective to alleviate the adverse effects of weaning stress on piglet growth.
Author Contributions
X.K. designed the experiments. S.D. and Y.C. conducted animal feeding experiments. M.D., Y.L., and B.S. analyzed data and M.D. wrote the manuscript. X.K., M.A., and M.D. revised the manuscript. All authors read and approved the final manuscript.
Funding
This study was supported by the Key Project of Regional Innovation and Development Joint Fund of National Natural Science Foundation of China (U20A2056).
Institutional Review Board Statement
The animal study was reviewed and approved by the Animal Care and Use Committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences (Approval no.20200018).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in the study are included in the article, and further inquiries can be directed to the corresponding author.
Acknowledgments
We thank staff members and postgraduate students from Hunan Provincial Key Laboratory of Animal Nutritional Physiology and Metabolic Process for collecting samples and technicians from the CAS Key Laboratory of Agro-ecological Process in Subtropical Region for providing technical assistance.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Pragna, P.; Sejian, V.; Bagath, M.; Krishnan, G.; Archana, P.R.; Soren, N.M.; Beena, V.; Bhatta, R. Comparative assessment of growth performance of three different indigenous goat breeds exposed to summer heat stress. J. Anim. Physiol. Anim. Nutr. 2018, 102, 825–836. [Google Scholar] [CrossRef]
- Huang, Y.; Li, J.; Zhang, J.; Jin, Z. Dynamical analysis of the spread of African swine fever with the live pig price in China. Math. Biosci. Eng. 2021, 18, 8123–8148. [Google Scholar] [CrossRef]
- Dibner, J.J.; Richards, J.D. Antibiotic growth promoters in agriculture: history and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef]
- Li, L.; Sun, X.; Zhao, D.; Dai, H. Pharmacological applications and action mechanisms of phytochemicals as alternatives to antibiotics in pig production. Front. Immunol. 2021, 12, 798553. [Google Scholar] [CrossRef]
- Van, T.P.; Glennon, E.E.; Chen, D.; Gilbert, M.; Robinson, T.P.; Grenfell, B.T.; Levin, S.A.; Bonhoeffer, S.; Laxminarayan, R. Reducing antimicrobial use in food animals. Science 2017, 357, 1350–1352. [Google Scholar]
- Hu, Y.; Cowling, B.J. Reducing antibiotic use in livestock, China. Bull World Health Organ 2020, 98, 360–361. [Google Scholar] [CrossRef]
- Ables, E.T.; Armstrong, A.R. Nutritional, hormonal, and metabolic drivers of development. Dev. Biol. 2021, 476, 171–172. [Google Scholar] [CrossRef]
- Kraemer, W.J.; Ratamess, N.A.; Hymer, W.C.; Nindl, B.C.; Fragala, M.S. Growth hormone(s), testosterone, insulin-like growth factors, and cortisol: Roles and integration for cellular development and growth with exercise. Front. Endocrinol. 2020, 11, 33. [Google Scholar] [CrossRef]
- Pierzchała, M.; Pareek, C.S.; Urbański, P.; Goluch, D.; Kamyczek, M.; Różycki, M.; Smoczynski, R.; Horbańczuk, J.O.; Kurył, J. Study of the differential transcription in liver of growth hormone receptor (GHR), insulin-like growth factors (IGF1, IGF2) and insulin-like growth factor receptor (IGF1R) genes at different postnatal developmental ages in pig breeds. Mol. Biol. Rep. 2012, 39, 3055–3066. [Google Scholar] [CrossRef]
- Kasprzak, A.; Adamek, A. The insulin-like growth factor (IGF) signaling axis and hepatitis C virus-associated carcinogenesis (review). Int. J. Oncol. 2012, 41, 1919–1931. [Google Scholar] [CrossRef]
- Reindl, K.M. , Sheridan, M.A. Peripheral regulation of the growth hormone-insulin-like growth factor system in fish and other vertebrates. Comp. Biochem. Physiol. 2012; 163, 231–245. [Google Scholar]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Furbeyre, H.; van Milgen, J.; Mener, T.; Gloaguen, M.; Labussière, E. Effects of oral supplementation with Spirulina and Chlorella on growth and digestive health in piglets around weaning. Animal, 2018; 12, 2264–2273. [Google Scholar]
- Matteri, R.L.; Dyer, C.J.; Touchette, K.J.; Carroll, J.A.; Allee, G.L. Effects of weaning on somatotrophic gene expression and circulating levels of insulin-like growth factor-1 (IGF-1) and IGF-2 in pigs. Domest. Anim. Endocrinol. 2000, 19, 247–259. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, N.; Ji, Y.; Tso, P.; Wu, Z. Weaning stress in piglets alters the expression of intestinal proteins involved in fat absorption. J. Nutr. 2022, 152, 2387–2395. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhao, P.; Zheng, X.; Zhou, L.; Wang, C.; Liu, J. Genome-wide detection of selection signatures in Duroc revealed candidate genes relating to growth and meat quality. Genes Genomes Genetics 2020, 10, 3765–3773. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, W.; Cai, J.; Ni, Y.; Xiao, L.; Zhang, J. Transcriptome analysis in comparing carcass and meat quality traits of Jiaxing black pig and Duroc × Duroc × Berkshire × Jiaxing black pig crosses. Gene 2022, 808, 145978. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chu, H.; Jiang, Y.; Lin, C.; Li, S.; Li, K.; Weng, G.; Cheng, C.; Lu, D.; Ju, Y. Empirical selection of informative microsatellite markers within co-ancestry pig populations is required for improving the individual assignment efficiency. Asian-Australas. J. Anim. Sci. 2014, 27, 616–627. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, H.; Liu, L.; Jiang, B.; Yu, Q.; He, J. Study on nutrient requirement of Xiangcun black pig at different growth stage. Acta Vet. et Zootechnica Sinica, 2013, 44, 1400–1410. [Google Scholar]
- Cheng, Y.; Ding, S.; Azad, M.A.K.; Song, B.; Kong, X. Comparison of the pig breeds in the small intestinal morphology and digestive functions at different ages. Metabolites 2023, 13, 132. [Google Scholar] [CrossRef]
- Rocha, DJPG. ; Castro, TLP.; Aguiar, ERGR.; Pacheco, LGC. Gene expression analysis in bacteria by RT-qPCR. Methods Mol. Biol. 2020, 2065, 119–137. [Google Scholar]
- Ding, S.; Cheng, Y.; Azad, M.A.K.; Zhu, Q.; Huang, P.; Kong, X. Developmental changes of immunity and different responses to weaning stress of Chinese indigenous piglets and Duroc piglets during suckling and weaning periods. Int. J. Mol. Sci. 2022, 23, 15781. [Google Scholar] [CrossRef]
- Wang, L.; Tan, X.; Wang, H.; Wang, Q.; Huang, P.; Li, Y.; Li, J.; Huang, J.; Yang, H.; Yin, Y. Effects of varying dietary folic acid during weaning stress of piglets. Anim Nutr. 2021, 7, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wu, Z.; Ren, G.; Zhao, Y.; Liu, D. Expression patterns of insulin-like growth factor system members and their correlations with growth and carcass traits in Landrace and Lantang pigs during postnatal development. Mol. Biol. Rep. 2013, 40, 3569–3576. [Google Scholar] [CrossRef]
- O'Kusky, J.; Ye, P. Neurodevelopmental effects of insulin-like growth factor signaling. Front. Neuroendocrinol. 2012, 33, 230–251. [Google Scholar] [CrossRef] [PubMed]
- LeRoith, D.; Holly, J.M.P.; Forbes, B.E. Insulin-like growth factors: Ligands, binding proteins, and receptors. Mol. Metab. 2021, 52, 101245. [Google Scholar] [CrossRef] [PubMed]
- Iresjö, B.M.; Diep, L.; Lundholm, K. Initiation of muscle protein synthesis was unrelated to simultaneously upregulated local production of IGF-1 by amino acids in non-proliferating L6 muscle cells. PLoS One 2022, 17, e0270927. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yakar, S.; LeRoith, D. Mice deficient in liver production of insulin-like growth factor I display sexual dimorphism in growth hormone-stimulated postnatal growth. Endocrinology 2000, 141, 4436–4441. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Kim, I.H. The impact of weaning stress on gut health and the mechanistic aspects of several feed additives contributing to improved gut health function in weanling piglets-a review. Animals (Basel) 2021, 11, 2418. [Google Scholar] [CrossRef]
- Zhong, H.; Lou, C.; Ren, B.; Zhou, Y. Insulin-like growth factor 1 injection changes gene expression related to amino acid transporting, complement and coagulation cascades in the stomach of tilapia revealed by RNA-seq. Front. Immunol. 2022, 13, 959717. [Google Scholar] [CrossRef]
- Feng, W.; Long, W.; Ding, M.; Chen, X.; Chen, C. Detection of the RYR1 gene in Guizhou indigenous pigs and three-crossbred pigs. Biotechnol. Bulletin 2015, 31, 66–70. [Google Scholar]
- Taguchi, A.; White, M.F. Insulin-like signaling, nutrient homeostasis, and life span. Annu. Rev. Physiol. 2008, 70, 191–212. [Google Scholar] [CrossRef]
- Hakuno, F.; Takahashi, S.I. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef]
- Wang, Y.; MacDonald, R.G.; Thinakaran, G.; Kar, S. Insulin-like growth factor-II/cation-independent mannose 6-phosphate receptor in neurodegenerative diseases. Mol. Neurobiol. 2017, 54, 2636–2658. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, S.; Zhang, X.; Wu, Q.; Li, S.; Fu, H.; Dong, L.; Yu, H.; Hao, L. Expression profiles of IGF-1R gene and polymorphisms of its regulatory regions in different pig breeds. Protein J. 2016, 35, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Desbois-Mouthon, C.; Wendum, D.; Cadoret, A.; Rey, C.; Leneuve, P.; Blaise, A.; Housset, C.; Tronche, F.; Le Bouc, Y.; Holzenberger, M. Hepatocyte proliferation during liver regeneration is impaired in mice with liver-specific IGF-1R knockout. FASEB J. 2006, 20, 773–775. [Google Scholar] [CrossRef] [PubMed]
- Haywood, N.J.; Slater, T.A.; Matthews, C.J.; Wheatcroft, S.B. The insulin like growth factor and binding protein family: Novel therapeutic targets in obesity & diabetes. Mol. Metab. 2019, 19, 86–96. [Google Scholar] [PubMed]
- Garcia, de.la.; Serrana, D.; Macqueen, D.J. ; Insulin-like growth factor-binding proteins of teleost Fishes. Front. Endocrinol. 2018, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Ranke, M.B. Insulin-like growth factor binding-protein-3 (IGFBP-3). Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 701–711. [Google Scholar] [CrossRef]
- Rajah, R.; Valentinis, B.; Cohen, P. Insulin-like growth factor (IGF)-binding protein-3 induces apoptosis and mediates the effects of transforming growth factor-beta1 on programmed cell death through a p53- and IGF-independent mechanism. J. Biol. Chem. 1997, 272, 12181–12188. [Google Scholar] [CrossRef]
- Frieten, D.; Gerbert, C.; Koch, C.; Dusel, G.; Eder, K.; Hoeflich, A.; Mielenz, B.; Hammon, H.M. Influence of ad libitum milk replacer feeding and butyrate supplementation on the systemic and hepatic insulin-like growth factor I and its binding proteins in Holstein calves. J. Dairy Sci. 2018, 101, 1661–1672. [Google Scholar] [CrossRef]
- Nishihara, K.; Suzuki, Y.; Kim, D.; Roh, S. Growth of rumen papillae in weaned calves is associated with lower expression of insulin-like growth factor-binding proteins 2, 3, and 6. Anim. Sci. J. 2019, 90, 1287–1292. [Google Scholar] [CrossRef]
- Nishihara, K.; Kato, D.; Suzuki, Y.; Kim, D.; Nakano, M.; Yajima, Y.; Haga, S.; Nakano, M.; Ishizaki, H.; Kawahara-Miki, R.; Kono, T.; Katoh, K.; Roh, S.G. Comparative transcriptome analysis of rumen papillae in suckling and weaned Japanese black calves using RNA sequencing. J. Anim. Sci. 2018, 96, 2226–2237. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.A. Insulin-like growth factor binding proteins 4−6. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 713–722. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Expression of IGF-related genes in the LD muscle, liver, kidney, stomach, spleen, duodenum, jejunum, and ileum tissues of the same breed of piglets at 21 (a) and 24 (b) days of age. DR, Duroc; TB, Taoyuan black; XB, Xiangcun black; IGF, insulin-like growth factor; IGF-R, insulin-like growth factor receptor; IGFBP, insulin-like growth factor binding protein. The red and blue in the figure represent the richness of gene expression and the value is normalized.
Figure 1.
Expression of IGF-related genes in the LD muscle, liver, kidney, stomach, spleen, duodenum, jejunum, and ileum tissues of the same breed of piglets at 21 (a) and 24 (b) days of age. DR, Duroc; TB, Taoyuan black; XB, Xiangcun black; IGF, insulin-like growth factor; IGF-R, insulin-like growth factor receptor; IGFBP, insulin-like growth factor binding protein. The red and blue in the figure represent the richness of gene expression and the value is normalized.
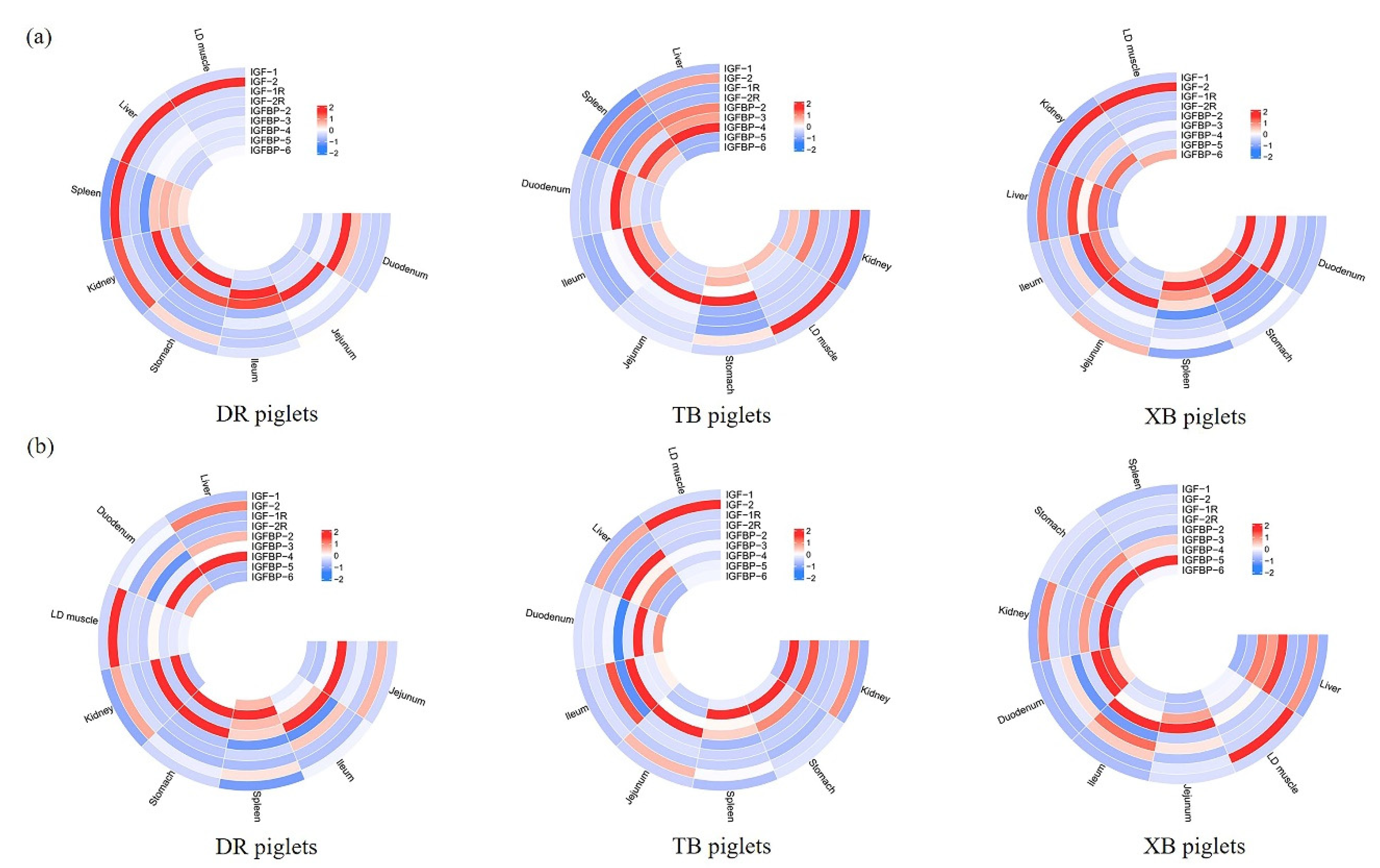
Figure 2.
Spearman’s correlation analysis between body weight, organ indices, and plasma IGF system components in different breeds of piglets. DR, Duroc; TB, Taoyuan black; XB, Xiangcun black. The red and blue represent a significantly positive correlation and negative correlation, respectively. *P < 0.05, **P < 0.01.
Figure 2.
Spearman’s correlation analysis between body weight, organ indices, and plasma IGF system components in different breeds of piglets. DR, Duroc; TB, Taoyuan black; XB, Xiangcun black. The red and blue represent a significantly positive correlation and negative correlation, respectively. *P < 0.05, **P < 0.01.
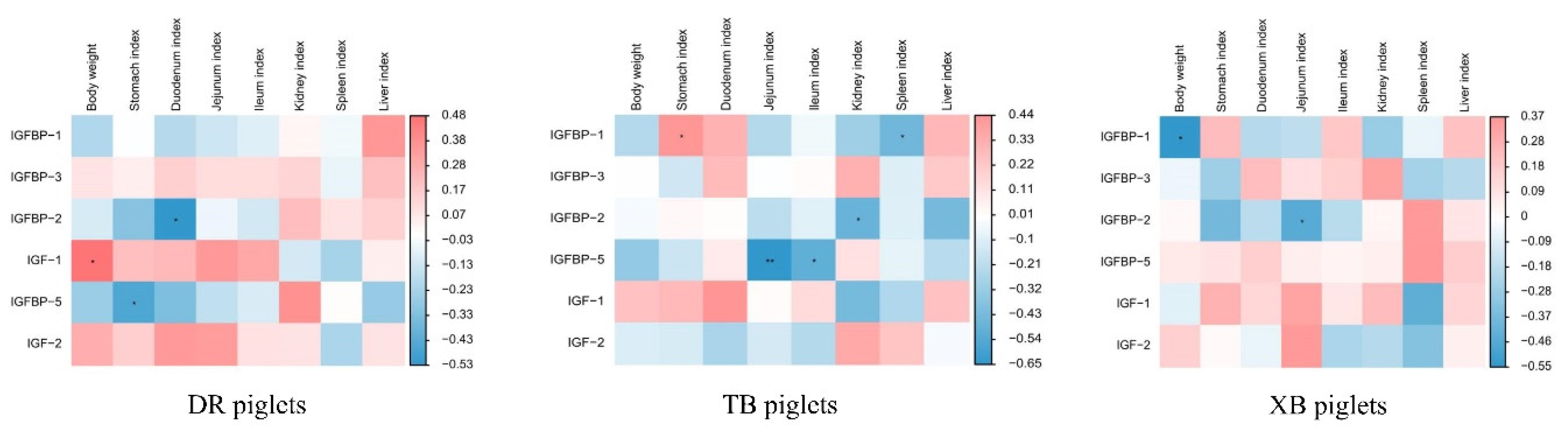
Figure 3.
Spearman’s correlation analysis between organ indices and IGF system components gene expression in different piglet breeds. DR, Duroc; TB, Taoyuan black; XB, Xiangcun black. The red and blue represent a significantly positive correlation and negative correlation, respectively. *P < 0.05, **P < 0.01.
Figure 3.
Spearman’s correlation analysis between organ indices and IGF system components gene expression in different piglet breeds. DR, Duroc; TB, Taoyuan black; XB, Xiangcun black. The red and blue represent a significantly positive correlation and negative correlation, respectively. *P < 0.05, **P < 0.01.
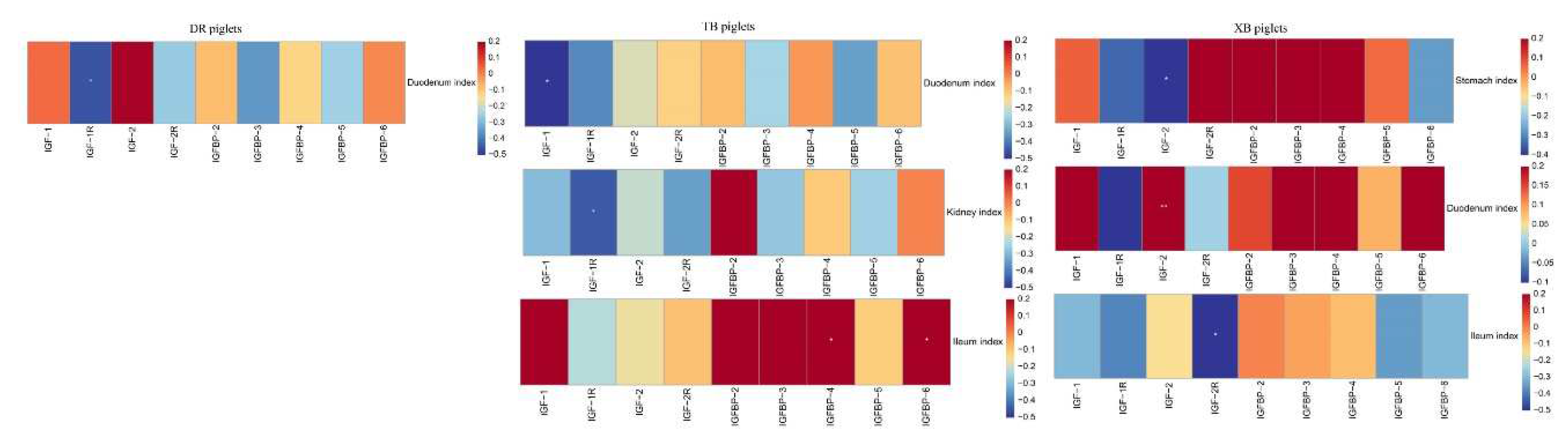

Table 1.
Primer sequences are used in the present study.
| Gene names | Gene number | Primer sequences (5’-3’) | Product size (bp) |
|---|---|---|---|
| IGF-1 | XM_005664199.3 | F: CCAAGGCTCAGAAGGAAGTACA | 137 |
| R: ACTCGTGCAGAGCAAAGGAT | |||
| IGF-2 | NM_213883.2 | F: CGGCTTCTACTTCAGCAGGC | 219 |
| R: TGCTTCCAGGTGTCATAGCG | |||
| IGF-1R | XM_021082915.1 | F: ACGAGTGGAGAAATCTGCGG | 154 |
| R: TGAGCTTGGGAAAGCGGTAG | |||
| IGF-2R | NM_001244473.1 | F: ACAGAAGCTGGACGTCATCG | 150 |
| R: CTGTCAACGTCGAACCTGCT | |||
| IGFBP-1 | NM_001195105.1 | F: CTATCACAGCAAACAGTGCGAG | 181 |
| R: CACGTGAAGGAAGAGAGCCT | |||
| IGFBP-2 | NM_214003.1 | F: CGAGCAGGTTGCAGACAATG | 288 |
| R: GTGGAGATCCGTTCCAGGAC | |||
| IGFBP-3 | NM_001005156.1 | F: AAGAAAAAGCAGTGCCGCC | 208 |
| R: GATCGTGTCCTTGGCAGTCT | |||
| IGFBP-4 | NM_001123129.1 | F: CTGCTCCGAAGAGAAGCTGG | 279 |
| R: TCACCCTCGTCCTTGTCAGA | |||
| IGFBP-5 | NM_001315595.1 | F: GCAAGCCAAGATCGAGAGAG | 102 |
| R: GTGTGCTTGGGTCGGAAGAT | |||
| IGFBP-6 | NM_001100190.1 | F: CCCTCGGGGGAGAATCCTAA | 157 |
| R: GGCAAGGGCCCATCTCAG | |||
| β-Actin | XM_0210860471 | F: GATCTGGCACCACACCTTCTACAAC | 107 |
| R: TCATCTTCTCACGGTTGGCTTTGG |
IGF, insulin-like growth factor; IGF-R, insulin-like growth factor receptor; IGFBP, insulin-like growth factor binding protein.
Table 2.
The organ indices of three breeds of piglets at 21 and 24 days of age.
| organ Indices, g/kg |
21 days of age | 24 days of age | SEM | P-values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| DR | TB | XB | DR | TB | XB | Weaning | Breed | W × B | ||
| Stomach | 6.09 | 5.55 | 6.84 | 6.40 | 6.41 | 8.15 | 0.17 | 0.003 | <0.001 | 0.322 |
| Dudenum | 0.56 | 0.61 | 0.83 | 0.56 | 0.62 | 0.66 | 0.03 | 0.262 | 0.013 | 0.270 |
| Jejunum* | 14.66 | 17.67 | 18.96 | 15.51 | 15.03 | 17.91 | 0.64 | 0.454 | 0.098 | 0.529 |
| Ileum* | 16.75 | 20.07 | 20.09 | 17.80 | 15.25 | 19.55 | 0.56 | 0.185 | 0.120 | 0.076 |
| Kidney | 6.04 | 6.21 | 6.21 | 6.01 | 5.82 | 5.94 | 0.09 | 0.230 | 0.961 | 0.729 |
| Spleen* | 2.08 | 2.04 | 2.18 | 1.93 | 1.87 | 1.75 | 0.07 | 0.074 | 0.948 | 0.646 |
| Liver* | 23.96 | 25.98 | 27.37 | 26.85 | 25.64 | 26.59 | 0.38 | 0.427 | 0.207 | 0.097 |
Data are expressed as mean ± SEM (n = 10). * Data cited from [20, 22]. DR, Duroc; TB: Taoyuan black; XB, Xiangcun black.
Table 3.
The plasma IGFs and IGFBPs concentration in different breeds of piglets at 21 and 24 days of age.
Table 3.
The plasma IGFs and IGFBPs concentration in different breeds of piglets at 21 and 24 days of age.
| Index, µg/µL | 21 days of age | 24 days of age | SEM | P-values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| DR | TB | XB | DR | TB | XB | Weaning | Breed | W × B | ||
| IGF-1 | 15.95 | 15.17 | 17.88 | 15.50 | 15.06 | 17.86 | 0.39 | 0.797 | 0.012 | 0.971 |
| IGF-2 | 1.67 | 1.61 | 1.79 | 1.79 | 1.53 | 1.83 | 0.05 | 0.799 | 0.152 | 0.740 |
| IGFBP-1 | 38.90 | 42.82 | 43.29 | 44.42 | 45.41 | 42.31 | 1.42 | 0.425 | 0.794 | 0.668 |
| IGFBP-2 | 17.82 | 22.30 | 28.76 | 18.40 | 22.57 | 22.12 | 0.99 | 0.277 | 0.006 | 0.177 |
| IGFBP-3 | 36.53b | 48.03a | 35.66b | 37.66b | 34.05b | 32.00b | 1.35 | 0.029 | 0.065 | 0.045 |
| IGFBP-5 | 12.46 | 17.27 | 23.14 | 11.52 | 17.91 | 21.81 | 0.82 | 0.588 | <0.001 | 0.694 |
Data are expressed as mean ± SEM (n = 7–10). a,b Mean with different letters within the same row indicate a significant difference (P < 0.05). DR, Duroc; TB: Taoyuan black; XB, Xiangcun black. IGF, insulin-like growth factor; IGFBP, insulin-like growth factor binding protein.
Table 4.
The gene expression of IGFs in tissues of the three breeds of piglets at 21 and 24 days of age.
Table 4.
The gene expression of IGFs in tissues of the three breeds of piglets at 21 and 24 days of age.
| Genes | Tissues | 21 days of age | 24 days of age | SEM | P-values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DR | TB | XB | DR | TB | XB | Weaning | Breed | W × B | |||
| IGF-1 | Stomach | 1.03 | 2.10 | 2.13 | 2.29 | 1.66 | 1.88 | 0.19 | 0.610 | 0.735 | 0.134 |
| Duodenum | 1.07 | 1.34 | 1.29 | 1.00 | 1.26 | 0.70 | 0.08 | 0.122 | 0.227 | 0.314 | |
| Jejunum | 1.60abc | 1.86ab | 2.52a | 1.49bc | 1.60abc | 0.78c | 0.15 | 0.011 | 0.841 | 0.030 | |
| Ileum | 0.78 | 0.61 | 0.68 | 0.78 | 0.34 | 0.33 | 0.04 | 0.003 | 0.001 | 0.101 | |
| Kidney | 0.90a | 0.58b | 0.43b | 0.60b | 0.89a | 0.62b | 0.04 | 0.308 | 0.015 | 0.002 | |
| Spleen | 0.26b | 1.25a | 0.84a | 0.17b | 0.34b | 0.21b | 0.08 | <0.001 | 0.002 | 0.028 | |
| Liver | 5.80ab | 7.66a | 3.27c | 3.77bc | 1.75c | 1.31c | 0.45 | <0.001 | 0.004 | 0.033 | |
| LD muscle | 0.28 | 0.34 | 0.33 | 0.41 | 0.43 | 0.33 | 0.03 | 0.169 | 0.729 | 0.630 | |
| IGF-2 | Stomach | 3.62 | 4.00 | 2.74 | 3.76 | 1.42 | 1.66 | 0.27 | 0.018 | 0.042 | 0.080 |
| Duodenum | 1.07 | 0.97 | 1.12 | 1.13 | 1.35 | 0.67 | 0.09 | 0.985 | 0.440 | 0.158 | |
| Jejunum | 0.82 | 0.80 | 0.70 | 4.12 | 6.71 | 0.84 | 0.67 | 0.002 | 0.045 | 0.055 | |
| Ileum | 0.46 | 0.40 | 0.48 | 0.43 | 0.43 | 0.31 | 0.03 | 0.389 | 0.817 | 0.416 | |
| Kidney | 52.33 | 65.69 | 62.44 | 31.22 | 55.00 | 49.49 | 2.76 | 0.003 | 0.007 | 0.642 | |
| Spleen | 6.82 | 6.14 | 3.21 | 3.20 | 2.81 | 1.68 | 0.38 | <0.001 | 0.001 | 0.269 | |
| Liver | 74.11 | 104.84 | 125.37 | 79.73 | 61.70 | 64.75 | 10.06 | 0.117 | 0.761 | 0.382 | |
| LD muscle | 12.82 | 11.71 | 11.81 | 16.91 | 16.86 | 9.35 | 0.99 | 0.244 | 0.152 | 0.225 | |
Data are expressed as mean ± SEM (n = 6–8). a−c Mean with different letters within the same row indicate a significant difference (P < 0.05). DR, Duroc; TB, Taoyuan black; XB, Xiangcun black; IGF, insulin-like growth factor; LD, Longissimus dorsi.
Table 5.
The gene expression of IGF receptors in tissues of the three breeds of piglets at 21 and 24 days of age.
Table 5.
The gene expression of IGF receptors in tissues of the three breeds of piglets at 21 and 24 days of age.
| Genes | Tissues | 21 days of age | 24 days of age | SEM | P-values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DR | TB | XB | DR | TB | XB | Weaning | Breed | W × B | |||
| IGF-1R | Stomach | 0.49 | 0.65 | 0.61 | 0.57 | 0.31 | 0.51 | 0.04 | 0.179 | 0.787 | 0.136 |
| Duodenum | 1.10 | 1.20 | 1.24 | 0.65 | 1.24 | 0.96 | 0.06 | 0.063 | 0.073 | 0.264 | |
| Jejunum | 1.18 | 0.99 | 0.92 | 1.16 | 1.22 | 0.51 | 0.08 | 0.671 | 0.052 | 0.271 | |
| Ileum | 0.45 | 0.40 | 0.62 | 0.50 | 0.37 | 0.63 | 0.04 | 0.927 | 0.023 | 0.866 | |
| Kidney | 11.63 | 8.27 | 10.64 | 11.74 | 12.06 | 13.71 | 0.45 | 0.006 | 0.116 | 0.154 | |
| Spleen | 1.35 | 1.77 | 2.41 | 1.05 | 1.08 | 1.27 | 0.13 | 0.003 | 0.070 | 0.312 | |
| Liver | 0.89c | 2.28a | 1.96ab | 1.25c | 1.80b | 2.32a | 0.10 | 0.521 | <0.001 | 0.013 | |
| LD muscle | 0.22 | 0.09 | 0.25 | 0.36 | 0.26 | 0.23 | 0.03 | 0.147 | 0.372 | 0.467 | |
| IGF-2R | Stomach | 0.63 | 0.75 | 0.68 | 1.18 | 0.96 | 1.41 | 0.07 | <0.001 | 0.362 | 0.167 |
| Duodenum | 1.06 | 1.81 | 1.65 | 1.41 | 1.42 | 1.23 | 0.09 | 0.387 | 0.245 | 0.159 | |
| Jejunum | 1.50 | 1.94 | 1.38 | 2.10 | 2.34 | 1.23 | 0.14 | 0.271 | 0.037 | 0.467 | |
| Ileum | 0.88 | 1.11 | 1.16 | 1.14 | 0.92 | 0.82 | 0.05 | 0.376 | 0.971 | 0.045 | |
| Kidney | 5.09b | 4.82b | 3.45c | 6.75a | 7.32a | 8.05a | 0.29 | <0.001 | 0.781 | 0.007 | |
| Spleen | 1.43 | 1.47 | 1.93 | 1.72 | 1.29 | 1.89 | 0.08 | 0.877 | 0.027 | 0.437 | |
| Liver | 3.28 | 5.29 | 5.50 | 4.28 | 4.86 | 5.03 | 0.19 | 0.914 | 0.001 | 0.111 | |
| LD muscle | 0.39 | 0.16 | 0.54 | 0.72 | 0.51 | 0.38 | 0.06 | 0.168 | 0.350 | 0.167 | |
Data are expressed as mean ± SEM (n=6-8). a−c Mean with different letters within the same row indicate a significant difference (P < 0.05). DR, Duroc; TB, Taoyuan black; XB, Xiangcun black; IGF-R, insulin-like growth factor receptor; LD, Longissimus dorsi.
Table 6.
The gene expression of IGFBPs in tissues of the three breeds of piglets at 21 and 24 days of age.
Table 6.
The gene expression of IGFBPs in tissues of the three breeds of piglets at 21 and 24 days of age.
| Genes | Tissues | 21 days of age | 24 days of age | SEM | P-values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DR | TB | XB | DR | TB | XB | Weaning | Breed | W × B | |||
| IGFBP-1 | Liver | 1.39 | 18.66 | 14.45 | 11.27 | 15.90 | 16.88 | 1.58 | 0.294 | 0.014 | 0.253 |
| IGFBP-2 | Stomach | 0.66 | 0.93 | 0.54 | 1.03 | 0.65 | 0.92 | 0.06 | 0.207 | 0.753 | 0.056 |
| Duodenum | 2.63bc | 7.48a | 3.38b | 0.44c | 0.51c | 0.48c | 0.53 | <0.001 | 0.019 | 0.023 | |
| Jejunum | 0.48 | 2.23 | 0.70 | 1.39 | 2.06 | 0.58 | 0.20 | 0.568 | 0.004 | 0.406 | |
| Ileum | 0.16b | 1.05a | 0.33b | 0.09b | 0.15b | 0.22b | 0.08 | 0.016 | 0.031 | 0.039 | |
| Kidney | 0.34c | 8.31a | 5.44b | 8.05a | 6.60ab | 8.24a | 0.48 | <0.001 | <0.001 | <0.001 | |
| Spleen | 0.05b | 5.91a | 0.12b | 0.08b | 0.09b | 0.09b | 0.35 | <0.001 | <0.001 | <0.001 | |
| Liver | 10.36c | 118.90ab | 141.75a | 60.67bc | 111.30ab | 85.11ab | 9.88 | 0.775 | <0.001 | 0.034 | |
| LD muscle | 0.02b | 0.02b | 0.03b | 0.07a | 0.05ab | 0.03b | 0.00 | 0.010 | 0.270 | 0.022 | |
| IGFBP-3 | Stomach | 6.50b | 9.45ab | 6.24b | 14.92a | 5.47b | 6.72b | 1.01 | 0.385 | 0.168 | 0.032 |
| Duodenum | 1.14b | 3.73a | 1.47b | 1.05b | 1.30b | 0.87b | 0.22 | 0.004 | 0.002 | 0.018 | |
| Jejunum | 5.89 | 17.62 | 5.63 | 8.91 | 15.87 | 2.94 | 1.47 | 0.856 | 0.001 | 0.649 | |
| Ileum | 2.88 | 3.10 | 2.36 | 2.20 | 1.08 | 1.00 | 0.20 | <0.001 | 0.146 | 0.314 | |
| Kidney | 58.21 | 42.01 | 25.75 | 47.70 | 69.12 | 44.26 | 3.62 | 0.075 | 0.023 | 0.056 | |
| Spleen | 3.78 | 2.98 | 4.30 | 3.48 | 4.33 | 5.42 | 0.29 | 0.212 | 0.151 | 0.440 | |
| Liver | 11.34c | 105.14a | 58.72b | 33.50bc | 37.02bc | 62.67b | 7.10 | 0.248 | 0.005 | 0.009 | |
| LD muscle | 1.11 | 0.33 | 2.09 | 3.28 | 2.27 | 1.86 | 0.33 | 0.047 | 0.485 | 0.241 | |
| IGFBP-4 | Stomach | 1.15 | 3.30 | 1.33 | 1.66 | 1.59 | 1.46 | 0.26 | 0.476 | 0.145 | 0.161 |
| Duodenum | 1.28 | 1.66 | 1.48 | 2.47 | 3.01 | 1.87 | 0.19 | 0.008 | 0.306 | 0.505 | |
| Jejunum | 0.91 | 0.48 | 0.44 | 2.46 | 3.86 | 1.67 | 0.26 | <0.001 | 0.095 | 0.082 | |
| Ileum | 3.68a | 1.68b | 1.68b | 1.13bc | 0.47c | 0.53c | 0.19 | <0.001 | <0.001 | 0.008 | |
| Kidney | 9.40 | 8.85 | 5.44 | 8.47 | 6.30 | 8.36 | 0.49 | 0.843 | 0.209 | 0.061 | |
| Spleen | 4.10 | 6.91 | 5.60 | 3.92 | 3.25 | 2.35 | 0.46 | 0.009 | 0.493 | 0.203 | |
| Liver | 6.05c | 153.59a | 136.24a | 122.78a | 70.81b | 71.83b | 9.25 | 0.405 | 0.005 | <0.001 | |
| LD muscle | 0.66 | 0.77 | 0.53 | 1.10 | 0.62 | 0.34 | 0.08 | 0.833 | 0.074 | 0.179 | |
| IGFBP-5 | Stomach | 8.38 | 5.11 | 6.18 | 12.87 | 9.57 | 12.09 | 0.71 | <0.001 | 0.097 | 0.856 |
| Duodenum | 1.03 | 1.33 | 3.89 | 0.96 | 1.37 | 1.78 | 0.28 | 0.159 | 0.010 | 0.153 | |
| Jejunum | 1.01 | 0.85 | 0.95 | 0.86 | 0.96 | 0.53 | 0.06 | 0.156 | 0.314 | 0.161 | |
| Ileum | 0.48b | 0.52b | 0.49b | 0.83a | 0.46b | 0.45b | 0.04 | 0.224 | 0.064 | 0.030 | |
| Kidney | 47.43b | 30.93b | 42.19b | 46.07b | 81.66a | 74.67a | 3.66 | <0.001 | 0.177 | 0.001 | |
| Spleen | 3.7 | 5.15 | 8.81 | 6.74 | 11.69 | 15.23 | 0.87 | <0.001 | 0.001 | 0.508 | |
| Liver | 1.67c | 1.28c | 1.42c | 2.01c | 3.51b | 5.75a | 0.29 | <0.001 | 0.001 | <0.001 | |
| LD muscle | 0.90 | 0.88 | 1.22 | 1.59 | 1.83 | 1.35 | 0.13 | 0.020 | 0.937 | 0.377 | |
| IGFBP-6 | Stomach | 1.99bc | 4.39ab | 4.37ab | 4.78a | 1.84c | 1.55c | 0.38 | 0.220 | 0.880 | 0.002 |
| Duodenum | 1.16 | 1.48 | 1.64 | 1.57 | 2.14 | 1.21 | 0.15 | 0.495 | 0.444 | 0.350 | |
| Jejunum | 0.64b | 0.58b | 0.55b | 1.22a | 1.70a | 0.50b | 0.10 | 0.001 | 0.007 | 0.014 | |
| Ileum | 0.74b | 1.44a | 0.85b | 0.73b | 0.57bc | 0.41c | 0.06 | <0.001 | 0.001 | <0.001 | |
| Kidney | 8.10 | 8.81 | 7.69 | 6.37 | 7.09 | 6.29 | 0.31 | 0.010 | 0.410 | 0.968 | |
| Spleen | 3.26 | 3.04 | 4.38 | 3.93 | 2.17 | 2.93 | 0.23 | 0.207 | 0.098 | 0.128 | |
| Liver | 1.52 | 1.13 | 1.08 | 1.14 | 1.59 | 1.62 | 0.08 | 0.177 | 0.990 | 0.033 | |
| LD muscle | 1.67b | 3.99a | 4.93a | 2.21b | 2.07b | 1.27b | 0.29 | 0.001 | 0.082 | 0.002 | |
Data are expressed as mean ± SEM (n=6-8). a−c Mean with different letters within the same row indicate a significant difference (P < 0.05). DR, Duroc; TB, Taoyuan black; XB, Xiangcun black; IGFBP, insulin-like growth factor binding protein; LD, Longissimus dorsi.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



