
Submitted:
11 August 2023
Posted:
14 August 2023
You are already at the latest version
Abstract
Keywords: Porcine reproductive and respiratory syndrome virus, baculovirus surface display vectors, neutralizing antibody, IFN-γ, IL-4.
Keywords:
Porcine reproductive and respiratory syndrome virus
; baculovirus surface display vectors
; neutralizing antibody
; IFN-γ
; IL-4
1. Introduction
Porcine reproductive and respiratory syndrome virus (PRRSV) belongs to the family Arteriviridae and is a single-stranded forward RNA virus with an envelope [1]. PRRSV contains at least 10 open reading frames (ORF) [3,4,5,6], of which ORFs 1a and 1b can translate non-structural proteins) [7,8,9], and ORF2-7 can translate viral structural proteins) [10,11]. Translation of ORF1a yields the pp1a polyprotein. ORF1b is expressed through a ribosomal frame shift, leading to the formation of a large pp1ab polyprotein [7,8,9]. The pp1a and pp1ab polyproteins are processed by viral proteases to release 14 non-structural proteins, which include four proteases (NSP1α, NSP1β, NSP2 and NSP4), the RNA-dependent RNA polymerase (NSP9), a helicase (NSP10) and an endonuclease (NSP11)[7,8,9]. ORFs 2-5 encode glycosylated membrane proteins GP2-GP5, ORF6 encodes a non-glycosylated membrane protein (M), and ORF7 encodes the nucleocapsid (N) protein [10,11]. ORF2b is enclosed fully within ORF2 and encodes the small, non-glycosylated E or 2b protein [4]. PRRSV emerged in Europe and in the US in the early 1990s and has since become a problem to the swine industry worldwide [12,13,14]. PRRSV is divided into two major genotypes, genotype 1 (European type, EU-type) and genotype 2 (North American type, NA-type) [15]. These two genotypes share only approximately 70% nucleotide sequence identity [16,17]. PRRSV causes a persistent and severe diseases that are characterized by severe reproductive failure in sows and respiratory disease in young pigs as well as weight loss and poor growth [18,19,20,21,22]. Other signs such as fever, blue ear, weight loss, poor growth, diarrhea, stillbirth, dyspnoea, and pneumonia could be observed as well. In late pregnancy, sows have clinical symptoms such as abortion, stillbirth, and mummified fetuses. [18,19,20,21]. The mortality rate of infected piglets is extremely high, causing serious economic losses to the global pig industry [20,21]. The PRRSV was diagnosed in Illinois, USA in 1987 [22], and the invasion of PRRSV was confirmed in Taiwan in 1993 [23]. The current PRRSV strains in the fields in Taiwan belong to the North American type [24].
The baculovirus expression system was established by Smith and Summer in 1983 [25] and this system has potential as a system for producing vaccines. The baculovirus gene has a strong promoter, which can produce a large amount of exogenous proteins and can carry out protein processing and modification, such as glycosylation and phosphorylation, so as to make the protein folding more correct. Thus, the baculovirus expression system is used to produce the recombinant protein that is closer to the original protein in antigenicity, immunity and biological activity. Baculovirus does not infect humans and is safe to operate. It has been used as vaccine vectors, RNA interference mediators, gene delivery vectors, and gene therapy [26,27,28,29,30]. The high lethality and widespread distribution of porcine reproductive and respiratory syndrome indicate the importance of PRRSV vaccination. According to previous studies, GP2, GP3, GP4, GP5 and M proteins are structural proteins of PRRSV, which are related to virus infection and cell entry, and can induce neutralizing antibodies to fight the virus [31,32,33,34,35,36,37]. Additionally, it has been investigated that GP3, GP4, and GP5 of PRRSV offer important antigenic properties for inducing neutralizing antibodies [32,33,34,35,36,37]. Therefore, this study was designed to construct novel baculovirus surface display vectors with multiple expression cassettes to increase expression levels of viral proteins and simultaneously express multiple viral proteins of PRSSV. In this work, two recombinant baculoviruses BacDual 4Display(BacDD)-2GP2-2GP4 (with 2 sets of GP2 and 2 sets of GP4) and BacDD-4GP5 (with 4 sets of GP5) were constructed for preparation of subunit vaccines of PRRSV. To test different combinations of protein co-expression levels and its immunogenicity, another two recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2GP5-2M were further created. This study also tested the optimum conditions for expressing PRRSV proteins by the novel baculovirus surface display vectors. The yield of recombinant proteins was analyzed under different infection days, different MOIs, different cell numbers and different serum concentrations of recombinant baculovirus. Our results revealed that high levels of PRRSV proteins-specific ELISA titters, neutralizing antibodies, IFN-γ, and IL-4 were elicited in SPF pigs immunized by the developed PRRSV subunit vaccine.
2. Materials and Methods
2.1. Cells and Viruses
To propagate PRRSV, MARC-145 cell line was cultured in Dulbecco's Modified Ea gle's Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin and 10 mM 4-(2-hydroxyethyl) piperazine-1-ethanesulphonic acid (HEPES) (pH 7. 2) at 37°C in a 5% CO2 humidified incubator. Cells were seeded one day before each experiment in 10-cm culture dishes and cultured until they reached about 75% confluence. Cells were infected with PRRSV at 37°C in a 5% CO2 humidified incubator for 3 days. In this work, Spodoptera frugiperda (Sf-9) cells were grown as monolayers in TNM-FH medium (Sigma, St. Louis, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco-BRL, Gaithersburg, USA), 100 U/mL of penicillin and 100 μg/mL of streptomycin. Recombinant viruses were propagated and titrated in Sf-9 cells.
2.2. Reverse Transcription (RT) and Polymerase Chain Reaction (PCR) and cDNA Cloning
Total RNA was isolated from PRRSV-infected cells using Trizol and Rneasy Mini Kit (Qiagen,Venlo, Netherlands) according to the manufacturer’s protocols. Total RNAs were subjected to RT-PCR assay. Transcription was carried out at 42°C for 1h using Oligo dT primer (20 M) (Promega, Fitchburg, USA ). After inactivation of reverse transcriptase at 95°C for 2min, the cDNA was subjected to 35 cycles of denaturation at 94°C for 1 min, annealing at 58°C for 1min, and extension at 72°C for 1 min and a final extension of 72°C for 7 min. PCR reaction was carried out in 50 μl containing 1 μl of PRRSV cDNA, 2 μl 2.5 mM dNTP, 2 μl 10 μM forward primer, 2 μl 10 μM reverse primer, 5 μl 10X pfu buffer, and 1 μl pfu DNA polymerase. The primers for amplification of the PRRSV genes are shown in Table 1. PCR products were purified and the products were digested with the respective restriction enzymes. After digestion, the viral genes were inserted into the corresponding restriction sites of pBacSC, BacDual Display (BacDD)-EGFP, and pET32a, respectively. All plasmids were transformed separately into E. coli strain DH5α and the white colonies were selected from Luria-Bertani (LB) agar plates containing ampicillin. The alkaline lysis method was used for mini-preparations of plasmid DNA. All plasmids were checked by both restriction enzyme digestions and DNA sequencing.
2.3. Construction of Baculovirus Surface Display Vectors Carrying Multiple Expression
2.3.1. Cassettes.
The detailed procedures for creating baculovirus surface display vectors with multiple expression cassettes was described previously [38,39,40]. Briefly, to construct the baculovirus surface display vector pBacSC [29], plasmid pBacCE was designed using pFast-Bac DUAL[39]. Sequences coding gp64 SS, His6, multiple cloning sites (Xho I, Xba I, PstI, and EcoRI) localized between His6 and baculovirus gp64 CTD, and baculovirus gp64 TM were inserted into pBacCE and the resultant plasmid was named pBacSC. The baculovirus surface display vector BacDD-EGFP was constructed previously [38,39] and used to create pBacSC-mtGP5, pBacDD-2GP2-2GP4 and pBacDD-4mtGP5, pBacDD-GP2-GP3-2GP4, and pBacDD-2mtGP5-2M constructs. To construct the pBacSC-mtGP5 plasmid, the PRRSSV GP5 gene was amplified by PCR (Table 1) and subcloned into the XhoI/PstI sites of the pBacSC plasmid.
2.4. Preparation of Recombinant Bacmid DNA and Construction of Recombinant Baculoviruses
The detailed procedures for preparation of recombinant Bacmid DNA and construction of recombinant Baculoviruses were described previously [38,39,40]. The resultant constructs pBacSC-mtGP5, pBacDD-2GP2-2GP4, pBacDD-4mtGP5, BacDD-GP2-GP3-2GP4, and BacDD-2mtGP5-2M were verified by enzyme digestion and DNA sequencing. Competent DH10Bac E. coli were then transformed with these recombinant plasmids and the non-recombinant plasmid pBacCE, respectively. The non-recombinant pBacCE was used as a negative control. After two rounds of blue/white selection, recombinant bacmids were isolated from white colonies according to the manufacturer’s instructions (Invitrogen, Carlsbad, USA). The recombinant clones were checked for the presence of the insert by PCR (Table 1). Positive colonies were cultured in order to isolate the bacmid DNA. Sf-9 cells were cultured at 27°C in Sf-900 II SFM. 9×105 cells were seeded in 35-mm wells of a six-well plate and allowed to attach for 1 h before transfection. Transfected Sf-9 cells were incubated for 5 h at 27°C and replaced with fresh medium. After incubation for 48 h at 27°C, recombinant viruses were selected based on GFP expression and purified by three rounds of plaque isolations. Individual recombinant viruses were titrated by plaque assay and high titer stocks were used for infection of the cells.
2.5. Confocal Microscopy
Confocal microscopy was performed as described previously [29,38]. Sf-9 cells were cultured on sterile cover slips (placed in six well plates) and infected at an MOI of 10. Two days post infection, the cells were fixed by methanol/acetone (v:v = 1:1) for 5 min at −20°C, rinsed with PBS, and blocked with 2% bovine serum albumin for 30 min at 37°C. The cells were then incubated with the primary antibody (anti-mouse IgG, 1:3000 dilution) for 1 h at 37°C, followed by three PBS washes. The cells were subsequently incubated with the secondary antibody (FITC-conjugated goat anti-mouse IgG, 1:50 dilution) for 1 h at 37 °C, followed by three PBS washes. Negative control cells were treated the same way. Protein localization was visualized using a confocal microscope (LSM 510 META, Zeiss, Germany).
2.6. Optimum Conditions for Production of Viral Proteins
To increase the expression/display levels of PRRSV proteins, optimum conditions for recombinant baculovirus transfection were examined. In the present study, various MOIs (10, 20, 30, 40 and 50 MOI), infection times (2 and 3 days), and different percentages (6-10 %) of heat inactivated FBS were tested.
2.7. Expression of PRRSV Proteins in E.coli and Establishment of PRRSV ELISA
To establish the PRRSV ELISA, the PRRSV proteins (GP2, GP3, GP4, GP5, and M) were expressed and used as the ELISA coating antigens. The amplified PCR products of the respective PRRSV genes were cloned into the responding sites of pET32a expressing vectors. The procedures for PRRSV protein expression in E.coli have been described previously [41,42]. Briefly, after induction for 3 h with isopropylthin-B-D-galactoside (IPTG) at a final concentration of 0.4 mM in culture medium, E. coli cells containing the constructs were induced, and the final pellets were separated on 12% acrylamide gels. For the His-tagged fusion proteins, cells were harvested by centrifugation, followed by resuspension in pET system lysis buffer (20 mM Tris-HCl, pH 8.0, 300 mM NaCl, 0.2 mM PMSF, 10% glycerol, 5 mM imidazole) and sonicated. The cell suspension was centrifuged at 12,000 xg for 20 min at 4 °C. After centrifugation, soluble proteins were collected for use in the ELISA. The supernatant was applied to a nickel column. After washing beads with 150 ml of washing buffer, the His fusion protein was eluted from the affinity column with elution buffer (20 mM Tris-HCl, pH 8.0, 300 mM NaCl, 0.2 mM PMSF, 10% glycerol, 200 mM imidazole). The protein concentration was determined according to the Lowry method [43]. To determine the optimum dilution of PRRSV antigens in ELISA, checkerboard titrations were carried out as described previously [41]. Purified viral proteins were used at a twofold dilution (0.25-16 ug) in 0.05 M carbonate buffer (pH 9.6) and 50 ul of antigen was added to each well of the ELISA plate. Antigen was coated onto the wells by incubation at 4 °C overnight. Standardization of the ELISA procedures were described previously [42].
2.8. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDSP-AGE) and Western Blot Analysis
The infected cell lysates were subjected to 12% SDS-PAGE and then transferred to PVDF membrane (GE Healthcare Life Sciences, Chicago, USA). Protein expression levels were examined using the appropriate primary antibody and horseradish peroxidase secondary antibody conjugate. The results were detected on X-ray films (Kodak, Rochester, USA) after membrane incubation with the enhanced chemiluminescence (ECL plus) reagent (Amersham Biosciences, Little Chalfont, England). The quantity of target proteins was calculated using ImageJ.
2.9. Immunization of SPF Pigs and Measurement of Viral Proteins Titers
Recombinant baculoviruses were propagated in Sf-9 cells at an MOI of 20 and har vested 3 days after infection. In this study, BacDD-2GP2-2GP4- and BacDD-4mtGP5- infected cells (1x 108 cells) were used as one dose to immunize SPF pigs. The recombinant protein forms a fusion protein with gp64, so that the target protein is expressed in a large amount on the cell membrane and stably hangs on the cell membrane. The Sf-9 cells infected with the recombinant virus were collected, and the cells were lysed by freezing and thawing three times. The cell debris contained PRRSV recombinant proteins GP2, GP4, and mtGP5. The cells were directly taken as vaccines without purification, which could save time and cost of vaccine preparation. In this work, ISA201 (Montanide) is a water-in-oil-in-water adjuvant which was used as an adjuvant in the pig vaccine administration. Its outer water layer antigens can be more easily recognized by the immune system, and the inner water layer antigens can be granulated and slowly released to continuously induce immune responses. Freund's adjuvant (Thermo) was used in the mouse immunization test. In the present study, BacDD-2GP2-2GP4- and BacDD-4mtGP5-infected cells (cell lysates) formulated with ISA70 were prepared. SPF pigs were divided into three vaccine groups (n=3, each); BacDD-2GP2-2GP4- and BacDD-4mtGP5-infected cell lysates, MLV commercial vaccine, and negative control group (BacCE). 5-week-old SPF pigs were immunized at the base of the ear by the intramuscular route with the above immunogens, respectively. In this work, Freund's adjuvant (Thermo) was used in the mouse immunization test.
2.10. Virus Neutralization Test and Analysis of IFN-γ and IL-4 in Swine Immunized with Different Antigens
A previous work revealed that IFN-γ and IL-4 is an important factor in the vaccine-induced cellular immune response[44,45,46]. Therefore, IFN-γ was used as an indicator of cellular immunity. Sera samples were collected at 4 and 11 weeks after primary immunization for cytokine and virus neutralization tests. Analysis of IFN-γ and IL-4 by ELISA was performed using the IFN-γ and IL-4 ELISA Kit (Uscn, Houston, USA). The serum source of this experiment is the 11th week serum of the pig immunization experiment. Firstly, the MARC-145 cell line was cultured in a 96-well plate, and 104 cells were cultured in each well. A 2-fold serial dilutions was performed and then 70 μl of 200 TCID50 PRRSV virus solution was added to each well and let it stand at 37°C for 1 hour. The culture medium in the 96-well plate was blotted dry, and 100 μl of PBS was added to each well to wash the cells. The serum and PRRSV virus were mixed evenly and added to a 96-well plate, cultured for 4 to 6 days, and the CPE was observed. Finally, the neutralizing antibody titer was calculated by the Reed-Muench method [47] . Sera samples were also collected every week after primary immunization for detection of the PRRSV proteins-specific ELISA titters. All swine was confirmed without antibodies against PRRSV before used.
2.11. Ethics Statement
The animal experimental protocols used in this study were approved by the Research Ethics Com mittee of National Chung Hsing University. All animal experimental procedures were carried out according to the Regulations for the Administration of Affairs Concerning Experimental Animals approved by Council of Agriculture, Taiwan.
3. Results
3.1. PCR Amplification of the PRRSV GP2, GP4, and mtGP5 Genes and Construction of Baculovirus Surface Display Vectors pBacSC-mtGP5, pBacDD-2GP2-2GP4, and pBacDD-4mtGP5
The GP2, GP4 and mtGP5 genes of PRRSV were amplified by RT-PCR with the respective primer pairs. The PCR products were analyzed by electrophoresis on a 1.2% agarose gel and the expected sizes of the PCR products are shown in (Figure 1A,B). A previous study has confirmed that the ability of PRRS GP5 protein to induce neutralizing antibody is superior to other viral proteins [48]. GP5 protein is a highly glycosylated protein, and the glycosylation structure causes the epitope to be shielded, which seriously affects the ability to induce neutralizing antibodies. A eariler report suggested that when the GP5 N-linked glycosylation sites (N-linked glycosylation sites) N34 and N51 of the PRRS undergo point mutation [49], not only the virus replication ability is weakened, but neutralizing antibody titers induced is reduced. In this work, site-directed mutagenesis was used to create GP5N34A/N51A mutant (mtGP5). The amplified PCR products were digested with the respective restriction enzymes, followed by subcloning into the pBacDD vector. The plasmid vectors were confirmed by restriction enzyme digestion and DNA sequencing. The resultant pBacDD-2GP2-2GP4 plasmid was digested with SalI and BamHI enzymes respectively to confirm the correctness of the recombinant fragment. After electrophoresis analysis with 1.2% agarose gel, the results showed that the target gene fragments were all in line with the expected size of 444 bp (Figure 1C). The constructed pBacDD-4mtGP5 vector was digested with XhoI and KpnI enzymes and electrophoresis analysis was performed on a 1% agarose gel. The results showed that the target gene fragments were all in line with the expected size of 220 bp and 650 bp (Figure 1D). Schematic illustration of the resultant novel baculovirus surface display vectors with multiple expression cassettes (pBacDD-2GP2-2GP4 and pBacDD-4mtGP5) are shown in Figure 1E,F. To improve the recombinant virus selection, enhanced green fluorescent protein (EGFP) coding sequences were subcloned into the corresponding site in the pBacDD vector under the strong viral polyhedron (polh) promoter (Figure 1E,F).
3.2. Preparation of Genetically Recombinant Baculoviruses BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4GP5
In this work, baculovirus surface display vectors pBacDD-EGFP, pGem-T-easy-DD, and pBacSC were constructed as described previously [20]. These vectors were used to create pBacSC-mtGP5, pBacDD-2GP2-2GP4, and pBacDD-4GP5 constructs, respectively. To create recombinant baculoviruses (BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4mtGP5), the pBacSC-mtGP5, pBacDD-2GP2-2GP4, and pBacDD-4mtGP5 plasmids were transformed into DH10BacTM E. coli, and homologous recombination was performed with the help of the help vector, and the successful recombinant bacmid would destroy the lacZ gene. The medium containing X-gal was used for screening blue and white colonies. The recombined baculoviruses bacmid mtGP5, GP2-2GP4, and 4mtGP5 were transfected into Sf-9 insect cells to construct the recombinant baculoviruses BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4GP5. The EGFP gene constructed in the vectors could be used to rapid screening the genetic recombinant baculoviruses [38]. In this study, EGFP and PRRSV proteins were expressed from different promoters in the same vector, allowing rapid identification of the recombinant baculoviruses in Sf-9 insect cells, and thus eliminating cumbersome and time-consuming assays. As shown in Figure 2A, three days after transfection, fluorescence signals were observed in BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4mtGP5 baculoviruses-infected Sf-9 cells by fluorescence microscope. It could be seen that the EGFP was expressed in transfected-Sf-9 insect cells and green fluorescence was seen except for un-transfected Sf-9 cells. In order to confirm that the target protein is successfully expressed in insect cells, the recombinant baculoviruses BacDD-2GP2-2GP4, BacDD-4GP5, BacSC-mtGP5, and BacCE (empty vector) were used to infect insect cells at an MOI of 20 for three days, and the recombinant baculovirus-transfected Sf-9 and Sf-9 alone cells (negative control) were collected for Western blotting. To analyze whether both mtGP5 and EGFP genes are successfully expressed in insect cells, the recombinant baculoviruses BacSC-mtGP5, BacDD-4mtGP5, and BacCE (empty vector) were used to infect insect cells at an MOI of 20 for three days, and the transfected Sf-9 and Sf-9 alone cells were collected for Western blotting. The results revealed that expression of the mtGP5 protein of PRRSV with the expected size of 15 kDa could be detected by the PRRSV polyclonal antibodies and His monoclonal antibodies (Figure 2B). The higher expression levels of mtGP5 by the pBacDD-4GP5 construct with four expression cassettes were seen (Figure 2B). The Western blotting analysis showed that the expression level of mtGP5 by the pBacDD-4GP5 vector (with four sets of mtGP5) is about 3-4 times higher than that of the BacSC-mtGP5 (with single set of mtGP5). To examine whether the mtGP5 mutant of PRRSV fused with gp64 signal sequence and gp64CTD were successfully displayed on insect cell membranes, mtGP5 protein was visualized by confocal microscopy using anti-His antibodies. Recombinant baculovirus BacSC-mtGP5 displaying the His6-tagged mtGP5-GP64TM-gp64CTD protein anchored on the plasma membrane of Sf-9 cells were observed under confocal microscopy (Figure 2C). No fluorescence signals were detected in Sf-9 cells (negative control). In this work, co-expression of the GP2 and GP4 protein of PRRSV with the expected size of 26 kDa and 20 kDa, respectively was detected using the PRRSV polyclonal antibody and His monoclonal antibody, respectively (Figure 2D). By comparing the expression levels of the GP2 and GP4 proteins of PRSSV by the recombinant baculovirus BacDD-2GP2-2GP4, we found that the expression level of GP2 was slightly higher than that of GP4 protein. No proteins were detectable in the mock groups (Bac CE and Sf-9).
3.3. Optimal Conditions for Expressing Recombinant PRRSV Protein
To increase the expression/display levels of the PRRSV proteins and to reduce the production cost of recombinant vaccines, the optimum conditions for recombinant baculovirus transfection and cell growth were studied. The conditions of an MOI of 20 (BacD4D-2GP2-2GP4) and 3 day post infection have higher expression levels of the GP2 and GP4 protein of PRRSV (Figure 3A). Furthermore, production of the GP2 and GP4 proteins of PRRSV in different cell numbers infected with recombinant baculovirus BacD4D-2GP2-2GP4 at an MOI of 20 was examined. Our results revealed that higher levels of the PRRSV GP2 and GP4 protein were seen in both BacD4D-2GP2-2GP4 and 4x105 cell numbers (Figure 3B). Furthermore, the PRRSV GP2, GP3, and GP4 protein expression in different serum concentration culture conditions were tested. Cells were infected with baculovirus BacD4D-GP2-GP3-2GP4 with at an MOI of 20 supplemented with different FBS concentrations (10%, 9%, 8%, 7% , and 6%). Our results reveal that higher expression levels of the GP2, GP3, and GP4 proteins of PRRSV were seen in both 7% and 3 day post infection (Figure 4).
3.4. Expression of GP2, GP4, and mtGP5 Proteins of PRRSV in E. coli
After confirming the gene sequences of pET32a-GP2, pET32a-GP4, and pET32a-GP5, the plasmids were transformed into E. coli BL21 (DE3) strain, and a single colony was selected for amplification and culture. Expression of GP2 and GP4 proteins was induced by IPTG for 0 to 5 hours. Our results show that the expected size of GP2 protein (insoluble protein) of 36 kDa and the expected size of GP4 (insoluble protein) of 30 kDa have been successfully expressed (Figure 5A,B). Both GP2 and GP4 proteins were detected by Western blotting using His monoclonal antibodies and PRRSV polyclonal antibodies (Figure 5C). The results showed that the expression levels of GP2 and GP4 proteins increased with the increase of induction times. As shown in Figure 5D,E, both the supernatant and the pellet had the mtGP5 protein with the expected size of 24 kDa. In addition, in order to confirm the antigenicity of the mtGP5 protein expressed by E. coli, His monoclonal antibodies and PRRSV polyclonal antibodies were used to detect the mtGP5 protein (Figure 5F), and the results showed that the protein was in line with the expected size.
3.5. Analysis of the Levels of Serum Antibodies and Neutralizing Antibodies in SPF Pigs Immunized with the Subunit Vaccine (BacD4D-2GP2-2GP4 and BacD4D-4mtGP5-Infected Sf-9 Cell Lysates)
In order to prepare an ELISA kit for detecting antibodies against viral proteins GP2, GP4, and mtGP5, the GP2, GP4, and mtGP5 proteins expressed by E. coli were used as antigens. The optimal serum dilution factor is 128 folds. ELISA-specific antibody analysis was performed under these conditions. In the group vaccinated with (BacD4D-2GP2-2GP4 and BacD4D-4mtGP5) subunit vaccine, the antibody levels in the sixteen weeks after the first and second immunization gradually increased to reach a peak of about OD405 nm of 1-1.2, which was significantlydifferent from the BacCE negative control group (Figure 6A). This result shows that the subunit vaccine (BacD4D-2GP2-2GP4 and BacD4D-4mtGP5) can indeed induce immune responses in pigs.
Diluted serum from pigs was mixed with virus in equal proportions, and after 1 hour of neutralization reaction, it was added to a 96-well plate containing MARC-145 cells, and cultured for 3-5 days to observe cytopathic effect (CPE). Without neutralizing antibodies, PRRSV cannot be neutralized, the virus will infect MARC-145 cells, and CPE can be observed, which is non-neutralization. On the other hand, if there is a phenomenon of neutralizing the virus, no CPE will be generated, which is regarded as neutralization. Our results confirmed that the BacD4D-2GP2-2GP4 and BacD4D-4mtGP5 subunit vaccines developed in this study can induce the higher levels of neutralizing antibodies, which in turn inhibit PRRSV infection. This result showed that the neutralizing antibody titers of the developed subunit vaccine and commercial vaccine (MLV) were 32 folds, which was significantly higher than that of the BacCE negative group (Figure 6B).
3.6. Analysis of IFN-γ and IL-4 in Swine Immunized with the BacD4D-2GP2-2GP4 and BacD4D-4GP5 Subunit Vaccines
A previous study showed that IFN-γ is an important factor in the vaccine-induced cellular immune response [44,45,46]. Therefore, IFN-γ was used as an indicator of cellular immunity in this study. It was confirmed by IFN-γ sandwich enzyme-linked immunosorbent assay (Sandwich ELISA). Our results revealed that the subunit vaccine (BacD4D-2GP2-2GP4 and BacD4D-4GP5) can stimulate immune cells to secrete IFN-γ and IL-4 and induce cellular immune responses.The results showed that the pigs immunized with the vaccine prepared by infecting cells with the recombinant baculoviruses BacD4D-2GP2-2GP4 and BacD4D-4GP5 could induce the production of IFN-γ and IL-4. The amounts of IFN-γ and IL-4 could be measured in the serum at 400 pg/ml and 270 pg/ml at the fourth week after immunization, respectively (Figure 6C). There was a significant difference in BacCE in the negative group. The level of IFN-γ was gradually declined at the eleventh week after immunization while the level of IL-4 was not altered.
3.7. Preparation of Genetically Recombinant Baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M
To compare the expression levels and efficacy of subunit vaccines prepared with different combinations of co-expression of viral proteins, two genetic recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M were further generated for examining simultaneous expression of GP2, GP3, GP4, GP5, and M proteins of PRRSV.To create recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M, the GP3 and M genes of PRRSV were amplified by RT-PCR with the respective primer pairs. The PCR products were analyzed by electrophoresis on 1.2% agarose gel, and the results showed that the target gene fragments were 318 bp and 48 bp in size as expected (Figure 7A,B). In this work, pBacDD-GP2-GP3-2GP4 and pBacDD-2mtGP5-2M vectors were created. The plasmid vectors were confirmed by restriction enzyme digestion and DNA sequencing. The resultant pBacDD-GP2-GP3-2GP4 and pBacDD-2mtGP5-2M vectors were digested with enzymes respectively to confirm the correctness of the recombinant fragment. After electrophoresis analysis with 1.2% agarose gel, the results showed that the target gene fragments were all in line with the expected sizes of 318 bp (Figure 7C) as well as 800 bp and 200bp (Figure 7D). To create recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacD4-2mtGP5-2M, the pBacDD-GP2-GP3-2GP4 and pBacDD-2mtGP5-2M plasmids were transformed into DH10BacTM E. coli, and homologous recombination was performed with the help of the help vector. The successful recombinant bacmid would destroy the lacZ gene. The medium containing X-gal was used for blue and white screening. The recombined baculovirus bacmid GP2-GP3-2GP4 and 2mtGP5-2M were transfected into Sf-9 insect cells to construct the recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacD4D-2mtGP5-2M. Schematic illustration of the resultant novel baculovirus surface display vectors with multiple expression cassettes (pBacD4-GP2-GP3-2GP4 and pBacDD-2mtGP5-2M plasmids) are shown in Figure 7E,F. As shown in Figure 7G, three days after transfection, fluorescence signals were observed in BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M baculoviruses-infected Sf-9 cells by fluorescence microscope. It could be seen that the EGFP was expressed in transfected-Sf-9 insect cells and green fluorescence was seen except for un-transfected Sf-9 cells. In order to confirm that the target protein is successfully expressed in insect cells, the recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M, and BacCE (empty vector) were used to infect insect cells at an MOI of 20 for three days, and the recombinant baculovirus-transfected Sf-9 and Sf-9 alone cells were collected for Western blotting. To analyze whether the GP2, GP3, GP4, mtGP5, and M proteins were successfully expressed in insect cells, the recombinant baculoviruses BacDD-GP2-GP3-2GP4, BacDD-2mtGP5-2M, and BacCE (empty vector) were used to infect insect cells at an MOI of 20 for three days, and the transfected Sf-9 and Sf-9 alone cells were collected for Western blotting. The results revealed that co-expression of the GP2, GP3, GP4, GP5, and M proteins of PRRSV with the expected size of 26, 22, 20, 15, and 11 kDa, respectively, could be detected by the PRRSV polyclonal antibodies and His monoclonal antibodies (Figure 7H,I).
3.9. Analysis of the Serum Antibody Levels of Mice Immunized with the Subunit Vaccine (BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M)
As mentioned above, the PRRSV GP2, GP4, and mtGP5 proteins were expressed by E. coli and were used as ELISA antigens (Figure 5A–E). In order to prepare GP3- and M-ELISA kits for detecting antibodies against viral proteins GP3 and M proteins, these proteins were also expressed by E. coli and used as ELISA antigens. After confirming the gene sequences of pET32a-GP3 and pET32a-M, the plasmids were transformed into E. coli BL21 (DE3) strain, and a single colony was selected for amplification and culture. Expression of the GP3 and M fusion proteins was induced by IPTG for 0 to5 hours. The results show that the expected size of GP3 protein (soluble protein) of 32 kDa and the expected size of M (soluble protein) of 21 kDa have been successfully expressed (Figure S1A). GP3 and M fusion proteins were detected by Western blotting using His monoclonal antibody and PRRSV polyclonal antibody (Figure S1B). In the group vaccinated with (BacD4D-GP2-GP3-2GP4 and BacD4D-2mtGP5-2M) subunit vaccine, the antibody levels in the thirteen weeks after immunization reach a peak of about OD405 nm of 1.3-1.8, which was significantly different from the BacCE negative control group (Figure S1C). This result shows that the subunit vaccine (BacD4D-GP2-GP3-2GP4 and BacD4D-2mtGP5-2M) can indeed induce immune responses in mice. Furthermore, the subunit vaccine developed in this study can induce the production of neutralizing antibodies in mice. This result showed that the neutralizing antibody titer of the subunit vaccine and commercial vaccine was 8 times, which was significantly higher than that of the BacCE and PBS negative groups (Figure S1D). In addition, in this study, to test whether the developed recombinant vaccine can increase the humoral and cellular immune responses and enhance its protection against PRRSV, the genetic recombinant baculoviruses BacDD-GP2-GP3-2GP4 (expressing GP2, GP3 and GP4) and BacDD-2mtGP5-2M (expressing 2 sets of GP5 and 2 sets of M proteins) were created. The recombinant vaccine developed in this study has higher antibody titers than the negative control group and has a significant difference. We found that the antibody titers of mice immunized with the commercially available attenuated vaccine in the positive control group were higher than those in the 5x107 cell number vaccine group. We have previously immunized mice with the same dose of BacDD-4mtGP5 and its immune response is better than that of the positive control group. We think it may be because the original BacDD-4mtGP5 was changed to BacDD-2mtGP5-2M in this experiment. The combination of which GP5 is the antigen that mainly induces the production of neutralizing antibodies, and the M protein is more related to cellular immunity, resulting in lower antibody titers. In addition, the antibody titer of mice decreased 4 weeks after the second immunization, and this result was similar to that of a previous report in mice immunized with the PCV Cap subunit vaccinet [39]. The sensitivity of the protein is different, and the immune effect induced is also different, so that the antibody titer cannot be maintained for a long time. In the future, we will use the immunization results as the basis for different protein combinations of PRRSV subunit vaccines.
4. Discussion
Within a few years of its emergence in the late 1980s, the PRRSV had spread globally to become the foremost infectious disease concern for the pork industry. Since 1994, modified live-attenuated vaccines against PRRSV have been widely used, but have failed to provide complete protection against emerging and heterologous field strains of the virus. Moreover, like many other MLVs, PRRSV-MLVs have safety concerns including vertical and horizontal transmission of the vaccine virus and several documented incidences of reversion to virulence. Previous studies have pointed out that when the live virus vaccine virus strain remains in the animal, it undergoes homologous recombination with the field strain virus, which will cause the vaccine strain to restore the original pathogencity of the virus [51]. Attenuated vaccines are still virulent and may infect weak piglets or pregnant sows and transmit the virus through body fluids and semen. Thus, the development of efficacious inactivated vaccines is warranted for the control and eradication of PRRSS. Since the early 1990s, scientists have been attempting to develop inactivated PRRSV vaccines, but most of the candidates have failed to elicit protective immunity even against homologous virus challenge. Recent reports relating to both inactivated and subunit candidate PRRSV vaccines have shown promise, but they need to be pursued further to improve their heterologous efficacy and cost-effectiveness before considering commercialization. In this study, we provide information on attempts to develop PRRSV inactivated and subunit vaccines. These include various virus inactivation strategies, adjuvants, nanoparticle-based vaccine delivery systems, DNA vaccines, and recombinant subunit vaccines produced using baculovirus, plant, and replication-deficient viruses as vector vaccines.
In this study, the pBacDual 4Display vector, the baculovirus surface display system developed in this study, is characterized by including the signal protein (SP), transmembrane region (TM) and cytoplasmic region (CTD), transcription is initiated by pPH promoter and p10 promoter, the target protein will form a fusion protein with gp64, gp64 can help the target protein to be presented on the Sf-9 cell membrane, and Sf-9 cells are collected as antigens to prepare gene recombinant subunit vaccines. In addition, the pPH promoter alone expresses EGFP, which can quickly observe the virus production under a fluorescent microscope, shorten the time for measuring the titer, and quickly determine whether the genetically recombined baculovirus is successfully obtained. The vector constructed in this study contains four expression cassettes that can express different proteins respectively, so as to improve the expression quantity and diversity of proteins. The baculovirus surface display vectors have also successfully applied to prepare subunit vaccines [29,30,38,39,40], further supporting that baculovirus surface display system has the value for developing subunit vaccines. The GP4 and GP5 proteins of PRRSV are important antigens for inducing neutralizing antibodies [32,33,34], so this study constructed two recombinant baculoviruses BacDD-2GP2-2GP4 and BacDD-4mtGP5 (with 4 sets of mtGP5). The Western blotting analysis showed that the expression level of mtGP5 by the pBacDD-4mtGP5 vector is about 3-4 times higher than that of the the BacSC-mtGP5 vector (with single set of mtGP5). Our results revealed that the pBacDD-2GP2-2GP4-EGFP plasmid does express two proteins of different sizes, GP2 and GP4, which confirms that multiple target proteins could be co-expressed, and the expression level of the target protein (mtGP5) could be elevated while using the baculovirus surface display vector with multiple expression cassetes. To further test the expression levels of different combinations of protein co-expression and their immunogenicity, another two baculovirus surface display vectors (pBacDD-GP2-GP3-2GP4 and pBacDD-2mtGP5-2M) that co-expressed/displayed different combinations of viral proteins were designed. Mice were immunized with the vaccine prepared from the above recombinant baculoviruses-infected Sf-9 cells and boosted at 14 days post-inoculation, showing that this subunit vaccine could induce high levels of specific anti-PRRSV antibodies and neutralizing antibodies, suggesting that vaccines prepared from the recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M-infected cells have the potential to serve as subunit vaccines against PRRSV. In this work, the neutralizing antibody titers of induced by BacDD-2GP2-2GP4-and BacDD-2mtGP5-infected cell lysates and commercial vaccine (MLV) were 32 folds, which was significantly higher than that of baculovirues BacDD-GP2-GP3-2GP4- and BacDD-2GP5-2M-infected cell lysates as well as the BacCE negative group. In this work, the pBacDD-4mtGP5 construct was changed to pBacDD-2mtGP5-2M. The combination of which GP5 is the antigen that mainly induces the production of neutralizing antibodies [32,33,34,36,48] , and the M protein is more related to cellular immunity, resulting in lower antibody titers.
Furthermore, to reduce the cost of the vaccine production and to increase expression level of viral proteins, different conditions such as cell numbers, MOI, infected days were tested. Our results suggest that two days post infection under MOI 30 show higher expression levels of viral proteins. After infecting Sf-9 cells with genetically recombinant baculovirus at different MOIs for 2 or 3 days, it was confirmed by Western blotting that the expression of PRRSV protein was higher after infecting cells with MOI 30, and observed under the microscope. The cell death rate of 3 days of infection is higher than that of 2 days of infection. In the future, we will use MOI 30 to infect cells for 2 days as the mass production condition. Cells are rapidly infected with high doses of virus, and live cells are harvested as a vaccine before the cells burst to death. A previous study suggested that the effect of using live cells as a vaccine is better than that of immunizing all cells (including dead cells), which may be related to the release of protease decomposition proteins after cell rupture [39]. In order to reduce the cost, we tested the challenge under different serum concentration conditions, and the results showed that when the virus was challenged, replacing the 10% FBS medium with 9% FBS medium could reduce the cost and have better protein expression. The preparation will rapidly infect cells with a high dose of MOI 30, and prepare antigens at this serum concentration to reduce the cost of vaccine production.
There are two main mechanisms to induce an immune response after vaccine injection. One is that antigens are taken up by antigen presenting cells (APCs) after immunization, and antigen fragments are presented to T cells by major histocompatibility complexes (MHCs). CD8+ T cells are mainly killer T cells, which can recognize the first class of MHC molecules and induce cytotoxicity and cytokine production. CD4+ T cells are dominated by TH cells and can recognize the second class of MHC molecules. TH1 cells induce phagocytosis of cytokine-activated macrophages, and TH2 cells activate B cells to produce antibodies. Another is the direct binding of antigens to B cells to produce specific antibodies [52],. A previous report has pointed out that IFN-γ is an indicator of vaccine-induced cellular immune response. CD4+ T cells secrete IFN-γ to activate CD8+ T cells and promote cytotoxicity, while activated CD8+ T cells continue to secrete IFN-γ, regulating cellular immunity [44,45,46], A good vaccine must be able to induce both humoral and cellular immune responses. In this work, the SPF pigs were immunized intramuscularly at day 0 and boosted at day 14 with 1×108 recombinant baculovirus-infected cells. Our findings revealed that the PRRSV subunit vaccine prepared from BacDD-2GP2-2GP4 and BacDD-4GP5-infected cells could induce high level of neutralizing antibody titers against PRRSV as well as the high levels of IFN-γ and IL-4. The antibody titers of the vaccine developed in this study can last for more than 15 weeks. Previous studies have confirmed that neutralizing antibody titers up to 8 times can effectively inhibit viremia, and neutralizing antibody titers up to 16 times can avoid placental vertical infection of PRRSV [50]. The vaccine developed in this study can induce high levels of neutralizing antibody titers and IFN-γ and IL-4 in pigs after immunization. Based on the above results, it is confirmed that the developed subunit vaccine can induce both humoral and cellular immune responses in pigs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figure S1: Coomassie blue-stained SDS-PAGE of E. coli expressing the PRRSV proteins and detection of ELISA titers and serum neutralization titers.
Author Contributions
All authors made substantive intellectual contributions to the present study and approved the final manuscript. HJL conceived of the study and generated the original hypothesis, wrote the paper, and supervised the project; CYH and YJ performed most of the experiments. CYH,YJ, WRH, CYW, HWW, PCT, and CYY analyzed data; MM and HJL revised and edited the manuscript.
Funding
This work was financially supported by Ministry of Science and Technology of Taiwan (109-2313-B-005-006-MY3& 112-2313-B-005-050-MY3), The iEGG and Animal Biotechnology Center from The Feature Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education (MOE) in Taiwan (111S0023A).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
We declare that we have no competing interests.
References
- Cavanagh, D. Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1997, 142, 629–633. [Google Scholar] [PubMed]
- Meulenberg, J.J. PRRSV, the virus. Vet. Res. 2000, 31, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Meulenberg, J.J.; Hulst, M.M.; de Meijer, E.J.; Moonen, P.L.; den Besten, A.; de Kluyver, E.P.; Wensvoort, G.; Moormann, R.J. Lelystad virus, the causative agent of porcine epidemic abortion and respiratory syndrome (PEARS), is related to LDV and EAV. Virology 1993, 19, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Meulenberg, J.J.; Petersen den Besten, A.; de Kluyver, E.; van Nieuwstadt, A.; Wensvoort, G.; Moormann, R.J. Molecular characterization of Lelystad virus. Vet. Microbiol. 1997, 55, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Meulenberg, J.J. The molecular biology of arteriviruses. J Gen Virol. 1998, 79 Pt 5, 961–979. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.I.; Lee, Y.M. Overview: replication of porcine reproductive and respiratory syndrome virus. J. Microbiol. 2013, 51, 711–723. [Google Scholar] [CrossRef]
- Snijder, E.J.; Wassenaar, A.L.; Spaan, W.J. Proteolytic processing of the replicase ORF1a protein of equine arteritis virus. J. Virol. 1994, 68, 5755–564. [Google Scholar] [CrossRef]
- Chen, Z.; Lawson, S.; Sun, Z.; Zhou, X.; Guan, X.; Christopher-Hennings, J.; et al. Identification of two auto-cleavage products of nonstructural protein 1 (nsp1) in porcine reproductive and respiratory syndrome virus infected cells: nsp1 function as interferon antagonist. Virology. 2010, 398, 87–97. [Google Scholar] [CrossRef]
- Li, Y.; Tas, A.; Snijder, E.J.; Fang, Y. Identification of porcine reproductive and respiratory syndrome virus ORF1a-encoded non-structural proteins in virus infected cells. J. Gen. Virol. 2012, 93 Pt 4, 829–839. [Google Scholar] [CrossRef]
- Firth, A.E.; Zevenhoven-Dobbe, J.C.; Wills, N.M.; Go, Y.Y.; Balasuriya, U.B.; Atkins, J.F.; et al. Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production. J. Gen. Virol. 2011, 92 Pt 5, 1097–1106. [Google Scholar]
- Johnson, C.R.; Griggs, T.F.; Gnanandarajah, J.; Murtaugh, M.P. Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses. J. Gen. Virol. 2011, 92 Pt 5, 1107–1116. [Google Scholar] [CrossRef]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; et al. Mystery swine disease in The Netherlands: the isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; et al. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J Vet Diagn Invest. 1992, 4, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Paton, D.J.; Brown, I.H.; Edwards, S.; Wensvoort, G. 'Blue ear' disease of pigs. Vet. Rec. 1991, 128, 617. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P.; Stadejek, T.; Abrahante, J.E.; Lam, T.T.; Leung, F.C. The ever-expanding diversity of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154, 18–30. [Google Scholar] [CrossRef]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; et al. Mystery swine disease in The Netherlands: the isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; et al. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Invest. 1992, 4, 127–133. [Google Scholar] [CrossRef]
- Chung, W.B.; Lin, M.W.; Chang, W.F.; Hsu, M.; Yang, P.C. Persistence of porcine reproductive and respiratory syndrome virus in intensive farrow-to-finish pig herds. Can. J. Vet. Res. 1997, 61, 292–8. [Google Scholar]
- Cho, J.G.; Dee, S.A. Porcine reproductive and respiratory syndrome virus. Theriogenology. 2006, 66, 655–662. [Google Scholar] [CrossRef]
- Neumann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.J.; Seitzinger, A.H.; et al. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J Am Vet Med Assoc. 2005, 227, 385–392. [Google Scholar] [CrossRef]
- Nieuwenhuis, N.; Duinhof, T.F.; van Nes, A. Economic analysis of outbreaks of porcine reproductive and respiratory syndrome virus in nine sow herds. Vet. Rec. 2012, 170, 225. [Google Scholar] [CrossRef] [PubMed]
- Rossow, K.D. Porcine reproductive and respiratory syndrome. Vet. Pathol. 1998, 35, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chung, W.B.; Lin, M.W.; Weng, C.N.; Yang, P.C.; Chiu, Y.T.; Chang, W.F.; Chu, R.M. Porcine reproductive and respiratory syndrome (PRRS) in Taiwan I. Viral isolation. J. Chin. Soc. Vet. Sci. 1993, 19, 268–276. [Google Scholar]
- Chueh, L.L.; Lee, K.; Wang, F.I.; Pang, V.F.; Weng, C.N. Sequence analysis of the nucleocapsid protein gene of the porcine reproductive and respiratory syndrome virus Taiwan MD-001 strain. Adv. Exp. Med. Biol. 1998, 440, 795–799. [Google Scholar]
- Smith, G.E.; Vlak, J.M.; Summers, M.D. Physical Analysis of Autographa californica Nuclear Polyhedrosis Virus Transcripts for Polyhedrin and 10,000-Molecular-Weight Protein. J. Virol. 1983, 45, 215–225. [Google Scholar] [CrossRef]
- Mäkelä, A.R.; Oker-Blom, C. Baculovirus display: a multifunctional technology for gene delivery and eukaryotic library development. Adv. Virus Res. 2006, 68, 91–112. [Google Scholar]
- Borg, J.; Nevsten, P.; Wallenberg, R.; Stenstrom, M.; Cardell, S.; Falkenberg, C.; et al. Amino-terminal anchored surface display in insect cells and budded baculovirus using the amino-terminal end of neuraminidase. J. Biotechnol. 2004, 114, 21–30. [Google Scholar] [CrossRef]
- Wang, S.; Fang, L.; Fan, H.; Jiang, Y.; Pan, Y.; Luo, R.; et al. Construction and immunogenicity of pseudotype baculovirus expressing GP5 and M protein of porcine reproductive and respiratory syndrome virus. Vaccine 2007, 25, 8220–8227. [Google Scholar] [CrossRef]
- Lin, Y.H.; Lee, L.H.; Shih, W.L.; Hu, Y.C.; Liu, H.J. Baculovirus surface display of sigma C and sigma B proteins of avian reovirus and immunogenicity of the displayed proteins in a mouse model. Vaccine. 2008, 26, 6361–6367. [Google Scholar] [CrossRef]
- Lu, H.Y.; Chen, Y.H.; Liu, H.J. Baculovirus as a vaccine vector. Bioengineered 2012, 3, 1–4. [Google Scholar] [CrossRef]
- Balasuriya, U.B.; Timoney, P.J.; McCollum, W.H.; MacLachlan, N.J. Phylogenetic analysis of open reading frame 5 of field isolates of equine arteritis virus and identification of conserved and nonconserved regions in the GL envelope glycoprotein. Virology 1995, 214, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Frey, M.L.; Yoon, K.J.; Zimmerman, J.J.; Platt, K.B. Categorization of North Ameri Can porcine reproductive and respiratory syndrome viruses: epitopic profiles of the N, M, GP5 and GP3 proteins and susceptibility to neutralization. Arch Virol. 2000, 145, 1599–619. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Jiang, P.; Wang, X.; Li, Y.; Du, Y. Enhanced immune responses of mice inoculated recombinant adenoviruses expressing GP5 by fusion with GP3 and/or GP4 of PRRS virus. Virus Res. 2008, 136, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Plagemann, P.G. The primary GP5 neutralization epitope of North American isolates of porcine reproductive and respiratory syndrome virus. Vet Immunol Immunopathol. 2004, 102, 263–75. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Jiang, P.; Li, Y.; Wang, X.; Du, Y. Analysis of immunogenicity of minor envelope protein GP3 of porcine reproductive and respiratory syndrome virus in mice. Virus Genes. 2007, 35, 695–704. [Google Scholar] [CrossRef]
- Cao, J.; Wang, X.; Du, Y.; Li, Y.; Wang, X.; Jiang, P. CD40 ligand expressed in adenovirus can improve the immunogenicity of the GP3 and GP5 of porcine reproductive and respiratory syndrome virus in swine. Vaccine 2010, 28, 7514–7522. [Google Scholar] [CrossRef]
- Das, P.B.; Vu, H.L.X.; Dinh, P.X.; Cooney, J.L.; Kwon, B.; Osorio, F.A.; Pattnaik, A.K. Glycosylation of minor envelope glycoproteins of porcine reproductive and respiratory syndrome virus in infectious virus recovery, receptor interaction, and immune response. Virology. 2011, 410, 385–394. [Google Scholar] [CrossRef]
- Tung, M.C.; Lu, H.Y.; Chang, Y.K.; Huang, W.R.; Liao, T.L.; Wu, H.Y.; Chang, C.D.; Nielsen, B.L.; Liu, H.J. Baculovirus surface display of the HA protein of H5N2 avian influenza virus and its immunogenicity against a lethal challenge with H5N1 virus in chickens. Vet, Microbiol. 2020, 243, 108640. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Yang, W.C.; Chang, Y.K.; Wang, C.Y.; Huang, W.R.; Li, J.Y.; Wu, H.Y.; Chang, C.D.; Nielsen, B.L.; Liu, H.J. Construction of polycistronic baculovirus surface display vectors to express the PCV2 Cap(d41) protein and its immunogenicity of the expressed proteins in mice and swine. Vet. Res. 2020, 51, 112. [Google Scholar] [CrossRef]
- Yang, D.G.; Chung, Y.C.; Lai, Y.K.; Lai, C.W.; Liu, H.J.; Hu, Y.C. Avian influenza virus hemagglutinin display on baculovirus envelope: cytoplasmic domain affects virus properties and vaccine potential. Mol. Ther. 2007, 15, 989–996. [Google Scholar] [CrossRef]
- Chiu, H.C.; Huang, W.R.; Liu, J.H.; Chi, P.I.; Nielsen, B.L.; Liao, T.L.; Liu, H.J. Mechanistic insights into avian reovirus p17-modulated suppression of cell-cycle CDK/cyclin complexes and enhancement of p53 and cyclin H interaction. J. Biol. Chem. 2018, 293, 12542–12562. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.J.; Kuo, L.C.; Hu, Y.C.; Liao, M.H.; Lien, Y.Y. Development of an ELISA for the detection of antibodies to avian reovirus in chickens. J. Virol. Methods 2002, 102, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with folin phenol reagent. J. Biol. Chem. 1951, 193, 265. [Google Scholar] [CrossRef]
- Fort, M.; Fernandes, L.T.; Nofrarias, M.; Diaz, I.; Sibila, M.; Pujols, J.; Mateu, E.; Segales, J.J. Development of cell-mediated immunity to porcine circovirus type 2 (PCV2) in caesarean-derived, colostrum-deprived piglets. Vet Immunol Immunopathol. 2009, 129, 101–107. [Google Scholar] [CrossRef]
- Carter, Q.L.; Curiel, R.E. Interleukin-12 (IL-12) ameliorates the effects of porcine respiratory and reproduc-tive syndrome virus (PRRSV) infection. Vet. Immunol. Immunopathol. 2005, 107, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Thanawongnuwech, R.; Thacker, E.L. Interleukin-10, interleukin-12, and interferon-gamma levels in the respiratory tract following mycoplasma hyopneumoniae and PRRSV infection in pigs. Viral Immunol. 2003, 16, 357–367. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Weiland, E.; Wieczorek-Krohmer, M.; Kohl, D.; Conzelmann, K.K.; Weiland, F. Monoclonal antibodies to the GP5 of porcine reproductive and respiratory syndrome virus are more effective in virus neutralization than monoclonal antibodies to the GP4. Vet. Microbiol. 1999, 66, 171–186. [Google Scholar] [CrossRef]
- Israrul, H.; Ansari, I.H.; Kwon, B.; Osorio, F.A.; Pattnaik, A.K. Influence of N-linked glycosylation of porcine reproductive and respiratory syndrome virus GP5 on virus infectivity, antigenicity, and ability to induce neutralizing antibodies. J. Virol. 2006, 80, 3994–4004. [Google Scholar]
- Lopez, O.J.; Oliveira, M.F.; Garcia, A.; Kwon, B.J.; Doster, A.; Osorio, F.A. Protection against porcine reproductive and respiratory syndrome virus (PRRSV) infection through passive transfer of PRRSV-neutralizing antibodies is dose dependent. Clin. Vaccine Immunol. 2007, 14, 269–275. [Google Scholar] [CrossRef]
- Botner, A. Diagnosis of PRRS. Vet. Microbiol. 1997, 55, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; O'Hagan, D. Advances in vaccine adjuvants. Nat. Biotechnol. 1999, 17, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PCR amplification of the GP2, GP4, and mtGP5 genes of PRRSV and schematic illustration of novel baculovirus surface display vectors carrying multiple expression cassettes for display of PRRSV proteins on the cell membrane. (A, B) DNA electrophoresis analysis of PCR products of the GP2, GP4, and mtGP5 genes of PRSSV. Lane M represents DNA marker (Bio-100 bp DNA ladder). (C, D) The resultant pBacDD-2GP2-2GP4 and pBacDD-4mtGP5 plasmid DNA was digested with SalI and BamHI enzymes as well as XhoI and KpnI, respectively to confirm the correctness of the recombinant fragments. After electrophoresis analysis with 1.2 % agarose gel, the results showed that the target gene fragments were all in line with the expected size of 444 bp (panel C) and the expected size of 220 bp and 650 bp (panel D). (E, F) Schematic illustration of baculovirus surface display vectors carrying multiple expression cassettes for higher level display of GP2, GP4, and mtGP5 proteins of PRRSV on the cell membrane . To simultaneously express and increase expression levels of multiple viral proteins of PRRSV, two novel baculoviral vectors pBacDD-2GP2-2GP4 and pBacDD-4mtGP5, which carry multiple expression cassettes for higher level display of proteins on cell membrane were constructed. The transmembrane domain and cytoplasmic domain of gp64 and the signal peptide sequence of gp64 are shown. The GP2, GP4, and mtGP5 genes of PRRSV was inserted between the gp64 signal peptide and gp64 cytoplasmic domains and CTD of the surface display vectors under the control of the p10 or Pph promoter. The resultant pBacDD-2GP2-2GP4 and pBacDD-4mtGP5 clones were used to create recombinant baculoviruses BacDD-2GP2-2GP4 and BacDD-4mtGP5.
Figure 1.
PCR amplification of the GP2, GP4, and mtGP5 genes of PRRSV and schematic illustration of novel baculovirus surface display vectors carrying multiple expression cassettes for display of PRRSV proteins on the cell membrane. (A, B) DNA electrophoresis analysis of PCR products of the GP2, GP4, and mtGP5 genes of PRSSV. Lane M represents DNA marker (Bio-100 bp DNA ladder). (C, D) The resultant pBacDD-2GP2-2GP4 and pBacDD-4mtGP5 plasmid DNA was digested with SalI and BamHI enzymes as well as XhoI and KpnI, respectively to confirm the correctness of the recombinant fragments. After electrophoresis analysis with 1.2 % agarose gel, the results showed that the target gene fragments were all in line with the expected size of 444 bp (panel C) and the expected size of 220 bp and 650 bp (panel D). (E, F) Schematic illustration of baculovirus surface display vectors carrying multiple expression cassettes for higher level display of GP2, GP4, and mtGP5 proteins of PRRSV on the cell membrane . To simultaneously express and increase expression levels of multiple viral proteins of PRRSV, two novel baculoviral vectors pBacDD-2GP2-2GP4 and pBacDD-4mtGP5, which carry multiple expression cassettes for higher level display of proteins on cell membrane were constructed. The transmembrane domain and cytoplasmic domain of gp64 and the signal peptide sequence of gp64 are shown. The GP2, GP4, and mtGP5 genes of PRRSV was inserted between the gp64 signal peptide and gp64 cytoplasmic domains and CTD of the surface display vectors under the control of the p10 or Pph promoter. The resultant pBacDD-2GP2-2GP4 and pBacDD-4mtGP5 clones were used to create recombinant baculoviruses BacDD-2GP2-2GP4 and BacDD-4mtGP5.
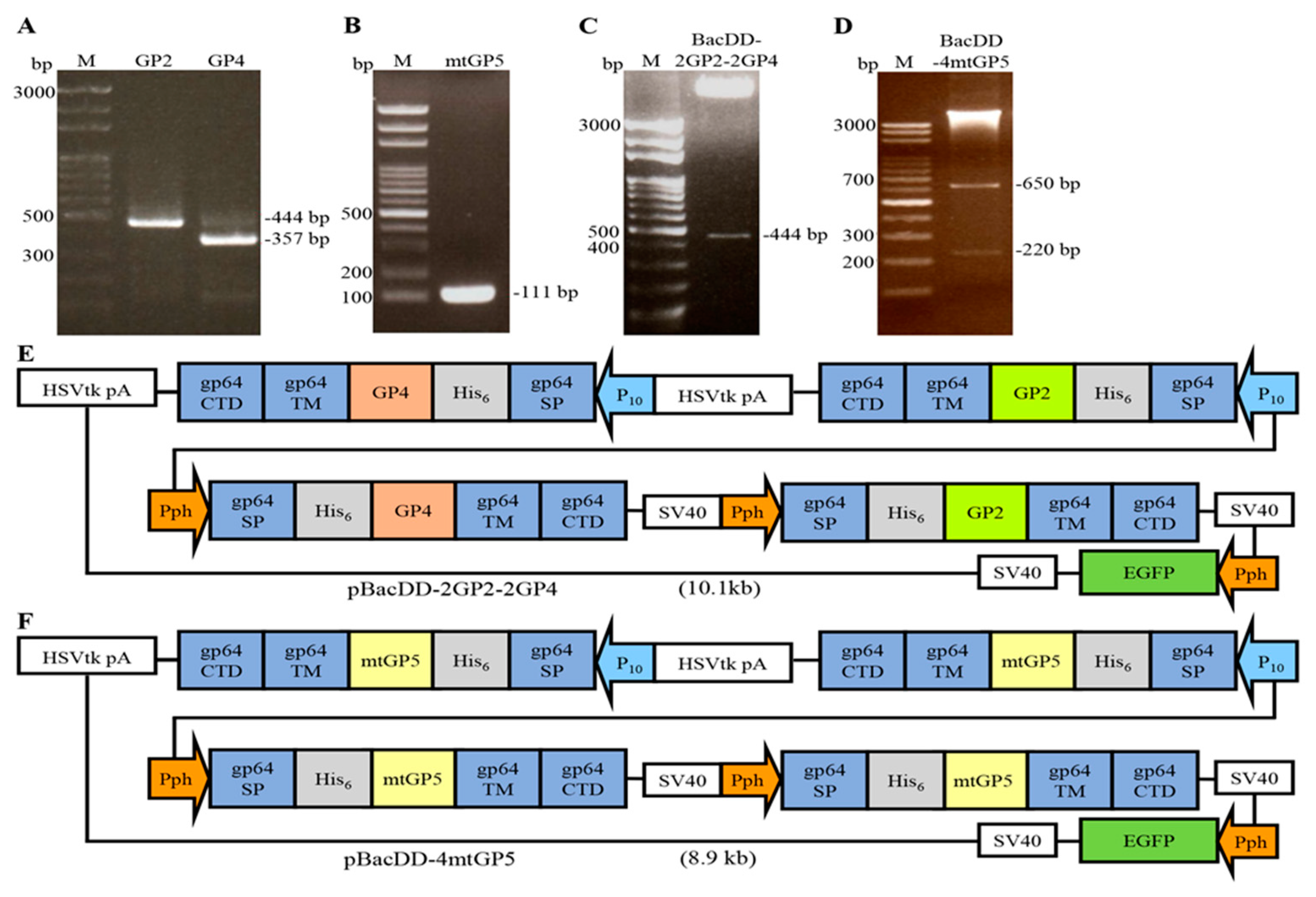
Figure 2.
Construction of BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4mtGP5 recombinant baculoviruses. (A) Images of BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4mtGP5 virus-infected SF-9 cells at 3 days post-infection. Phase contrast images (upper panel) and fluorescence images (lower panel). All images were magnified at 200X. (B) BacSC-mtGP5 and BacDD-4mtGP5 genetic recombinant baculoviruses, respectively, at an MOI of 20, harvested 3 days post infection, and subjected to Western blot assay using anti-His monoclonal antibody and PRRSV polyclonal antibodies, respectively. The 15 kDa mtGP5 protein was detected. No proteins were detectable in the negative controls (BacCE and Sf-9 cell alone). Signals in Western blots were quantified using Image J software and shown in the right panel. (C) Anchoring of mtGP5 on the plasma membrane of Sf-9 cells as revealed by immunofluorescence/confocal microscopy. (D) BacDD-2GP2-2GP4 virus at an MOI of 20, harvested 3 days post infection, and subjected to Western blot assay using anti-His monoclonal antibody and PRRSV polyclonal antibodies, respectively. Th 26 kDa GP2 and 20 kDa GP4 proteins were detected. No proteins were detectable in the negative controls (BacCE and Sf-9 cell alone).
Figure 2.
Construction of BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4mtGP5 recombinant baculoviruses. (A) Images of BacSC-mtGP5, BacDD-2GP2-2GP4, and BacDD-4mtGP5 virus-infected SF-9 cells at 3 days post-infection. Phase contrast images (upper panel) and fluorescence images (lower panel). All images were magnified at 200X. (B) BacSC-mtGP5 and BacDD-4mtGP5 genetic recombinant baculoviruses, respectively, at an MOI of 20, harvested 3 days post infection, and subjected to Western blot assay using anti-His monoclonal antibody and PRRSV polyclonal antibodies, respectively. The 15 kDa mtGP5 protein was detected. No proteins were detectable in the negative controls (BacCE and Sf-9 cell alone). Signals in Western blots were quantified using Image J software and shown in the right panel. (C) Anchoring of mtGP5 on the plasma membrane of Sf-9 cells as revealed by immunofluorescence/confocal microscopy. (D) BacDD-2GP2-2GP4 virus at an MOI of 20, harvested 3 days post infection, and subjected to Western blot assay using anti-His monoclonal antibody and PRRSV polyclonal antibodies, respectively. Th 26 kDa GP2 and 20 kDa GP4 proteins were detected. No proteins were detectable in the negative controls (BacCE and Sf-9 cell alone).
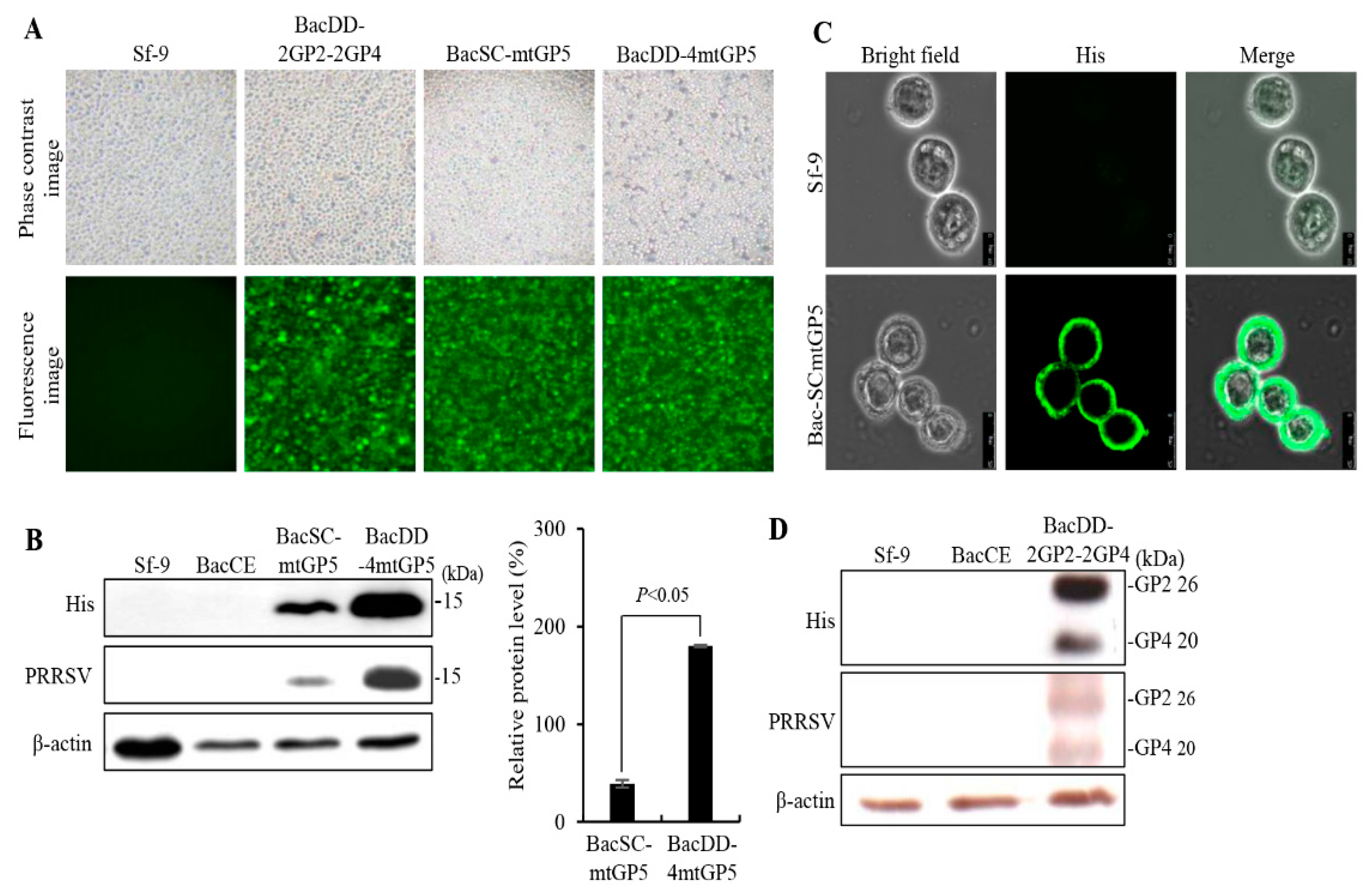
Figure 3.
Optimum conditions for production of the PRRSV proteins. Effects of different infection times, MOI, and cell numbers for expressing the the PRRSV proteins (GP2 and GP4) were tested. (A-B) Sf-9 cells were infected with BacDD-2GP2-2GP4 at different MOIs, different infection times or cell numbers. The Sf-9 cell alone and CE were used as negative controls. The expressed proteins were probed by anti-His monoclonal antibodies. Signals in all western blots were quantified using Image J software. β-actin was used as an internal control for normalization. Relative abundance (%) are shown. The results were calculated from the data shown in the upper portion of each panel. All data shown represent the mean ± SE calculated from three independent experiments. A student t-test was conducted for analysis.
Figure 3.
Optimum conditions for production of the PRRSV proteins. Effects of different infection times, MOI, and cell numbers for expressing the the PRRSV proteins (GP2 and GP4) were tested. (A-B) Sf-9 cells were infected with BacDD-2GP2-2GP4 at different MOIs, different infection times or cell numbers. The Sf-9 cell alone and CE were used as negative controls. The expressed proteins were probed by anti-His monoclonal antibodies. Signals in all western blots were quantified using Image J software. β-actin was used as an internal control for normalization. Relative abundance (%) are shown. The results were calculated from the data shown in the upper portion of each panel. All data shown represent the mean ± SE calculated from three independent experiments. A student t-test was conducted for analysis.
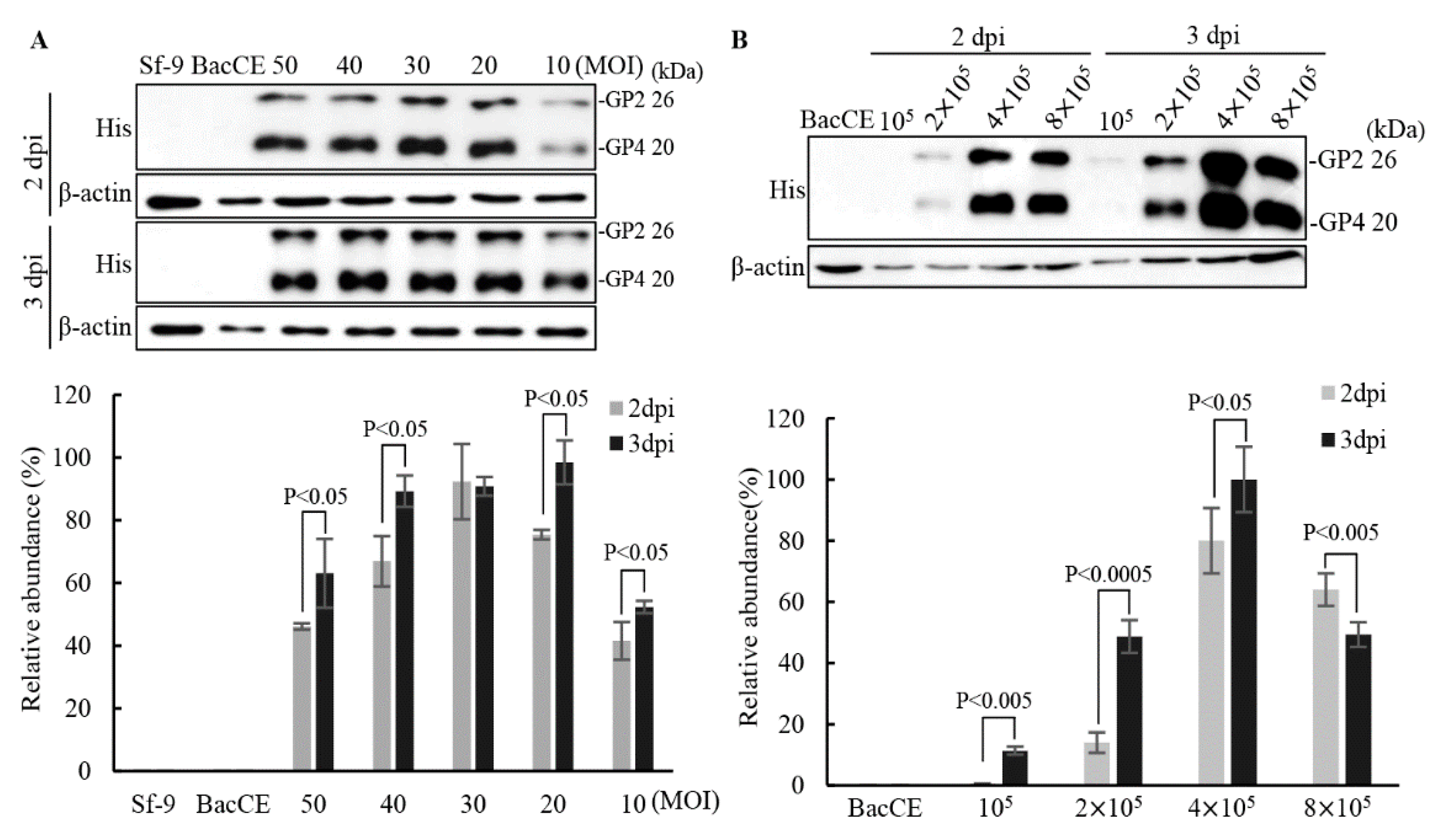
Figure 4.
Optimum conditions for production of the PRRSV proteins. Sf-9 cells were infected with baculovirus BacD4D-GP2-GP3-2GP4 with at an MOI of 20 supplemented with different FBS concentrations. The Sf-9 cell alone and CE were used as negative controls. The expressed proteins were probed by anti-His monoclonal antibodies. Signals in all western blots were quantified using Image J software. β-actin was used as an internal control for normalization. Relative abundance (%) are shown. The results were calculated from the data shown in the left panel. All data shown represent the mean± SE calculated from three independent experiments. A student t-test was conducted for analysis.
Figure 4.
Optimum conditions for production of the PRRSV proteins. Sf-9 cells were infected with baculovirus BacD4D-GP2-GP3-2GP4 with at an MOI of 20 supplemented with different FBS concentrations. The Sf-9 cell alone and CE were used as negative controls. The expressed proteins were probed by anti-His monoclonal antibodies. Signals in all western blots were quantified using Image J software. β-actin was used as an internal control for normalization. Relative abundance (%) are shown. The results were calculated from the data shown in the left panel. All data shown represent the mean± SE calculated from three independent experiments. A student t-test was conducted for analysis.
Figure 5.
Coomassie blue-stained SDS-PAGE of E. coli DE3 expressing the PRRSV proteins. The PRRSV proteins were expressed as shown in panel (A, B, D, E). (A)GP2, (B) GP4, (D) mtGP5 (suppernatant), and (E)mtGP5 (pellet). E. coli cells containing the indicated constructs were induced for different time points with IPTG at a final concentration of 0.4 mM in culture medium. The pET32a (empty vector) was used as the negative control. The expressed proteins were marked by red frame. (C, F) The expressed proteins GP2, GP4, and mtGP5 were probed by anti-His monoclonal antibodies or PRRSV polyclonal antibodies, respectively.
Figure 5.
Coomassie blue-stained SDS-PAGE of E. coli DE3 expressing the PRRSV proteins. The PRRSV proteins were expressed as shown in panel (A, B, D, E). (A)GP2, (B) GP4, (D) mtGP5 (suppernatant), and (E)mtGP5 (pellet). E. coli cells containing the indicated constructs were induced for different time points with IPTG at a final concentration of 0.4 mM in culture medium. The pET32a (empty vector) was used as the negative control. The expressed proteins were marked by red frame. (C, F) The expressed proteins GP2, GP4, and mtGP5 were probed by anti-His monoclonal antibodies or PRRSV polyclonal antibodies, respectively.
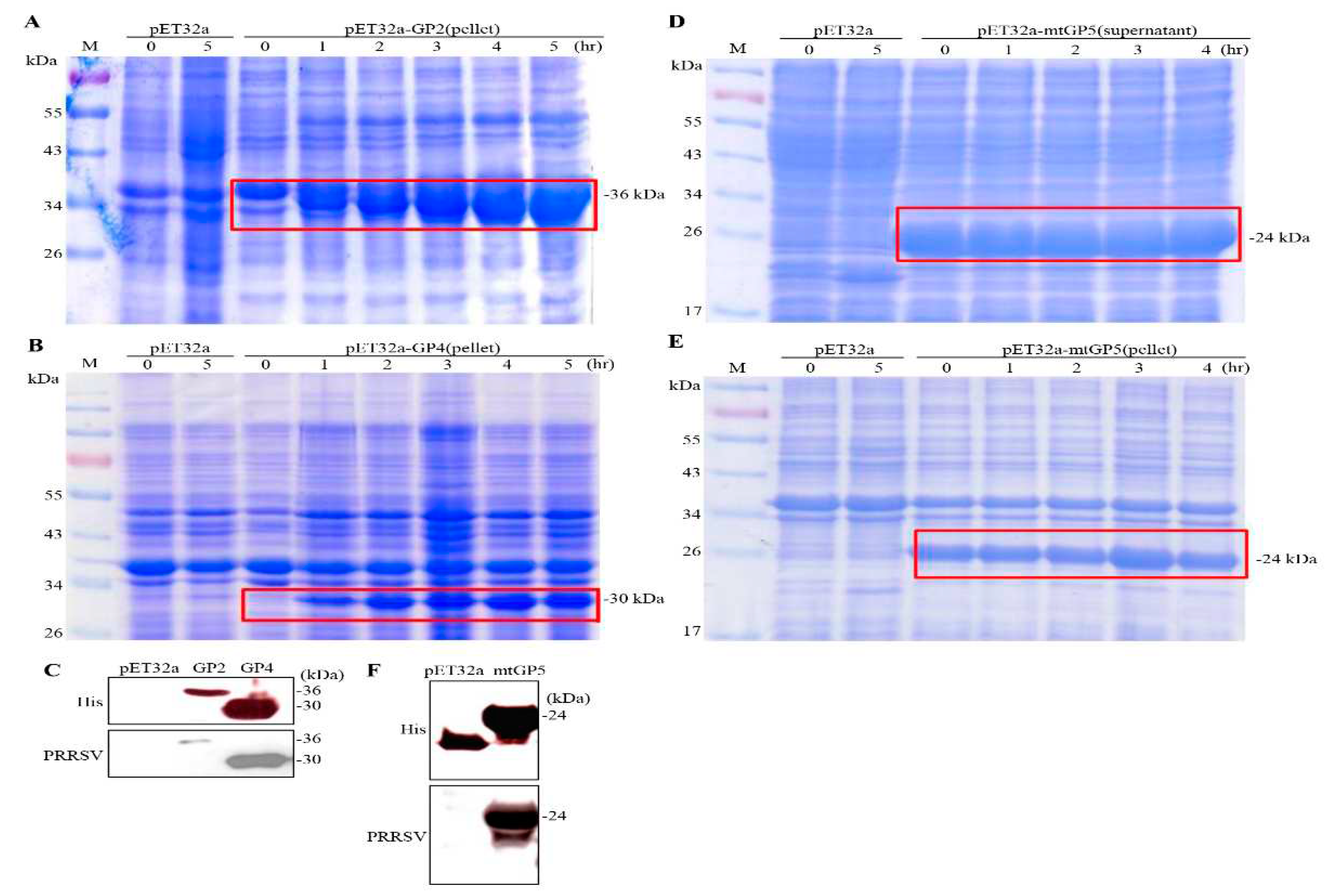
Figure 6.
Detection of ELISA titers, serum neutralization titers, and the levels of IFN-γand IL-4 in 5-week-old SPF pigs immunized with various immunogens. (A) Detection of anti-PRRSV protein titers by ELISA. 2-month-old SPF pigs were immunized at the base of the ear by the intramuscular route with BacCE and PRRSV vaccine (BacD4D-2GP2-2GP4 and BacD4D-4mtGP5), respectively. Serum samples were collected every week after the first immunization to determine the PRRThe PCR products were analyzed by electrophoresis on a 1.2% agarose gel andSV proteins-specific ELISA antibodies. (B) Serum neutralization (SN) titers in SPF pigs immunized with various immunogens (BacD4D-2GP2-2GP4 and BacD4D-4mtGP5; commcercial MLV vaccine). BacCE was used as the negative control. Sera were from 11 week post vaccination. (C) Analysis of IFN-γ by ELISA was performed using the IFN-γ ELISA Kit. The value of OD405 nm was measured with an ELISA reader. All data shown represent the mean± SE calculated from three independent experiments. A student t-test was conducted for analysis.
Figure 6.
Detection of ELISA titers, serum neutralization titers, and the levels of IFN-γand IL-4 in 5-week-old SPF pigs immunized with various immunogens. (A) Detection of anti-PRRSV protein titers by ELISA. 2-month-old SPF pigs were immunized at the base of the ear by the intramuscular route with BacCE and PRRSV vaccine (BacD4D-2GP2-2GP4 and BacD4D-4mtGP5), respectively. Serum samples were collected every week after the first immunization to determine the PRRThe PCR products were analyzed by electrophoresis on a 1.2% agarose gel andSV proteins-specific ELISA antibodies. (B) Serum neutralization (SN) titers in SPF pigs immunized with various immunogens (BacD4D-2GP2-2GP4 and BacD4D-4mtGP5; commcercial MLV vaccine). BacCE was used as the negative control. Sera were from 11 week post vaccination. (C) Analysis of IFN-γ by ELISA was performed using the IFN-γ ELISA Kit. The value of OD405 nm was measured with an ELISA reader. All data shown represent the mean± SE calculated from three independent experiments. A student t-test was conducted for analysis.

Figure 7.
PCR amplification of the GP3 and M genes of PRRSV and schematic illustration of baculovirus surface display vectors carrying multiple expression cassettes for display of PRRSV proteins on the cell membrane. (A, B) DNA electrophoresis analysis of PCR product of the GP3 and M genes of PRRSV. (C, D) The resultant pBacDD-GP2-GP3-2GP4- and pBacDD-2mtGP5-2M plasmid was digested with the respective restriction enzymes to confirm the correctness of the recombinant fragments. After electrophoresis analysis with 1.2% agarose gel, the results showed that the target gene fragments were all in line with the expected size of 318 bp (panel C) and the expected size of 800 bp and 200 bp (panel D). (E, F) Schematic illustration of baculovirus surface display vectors carrying multiple expression cassettes for higher level display of GP2, GP3, GP4, mtGP5, and M proteins of PRRSV on the cell membrane. To simultaneously express and increase expression levels of multiple viral proteins of PRRSV, two novel baculoviral vectors pBacDD-GP2-GP3-2GP4 (panel E) and pBacDD-2mtGP5-2M (panel F), which carry multiple expression cassettes for higher level display of proteins on cell membrane were constructed. The transmembrane domain and cytoplasmic domain of gp64 and the signal peptide sequence of gp64 are shown. The GP2, GP3, GP4, mtGP5, and M genes of PRRSV was inserted between the gp64 signal peptide and gp64 cytoplasmic domains and CTD of the surface display vectors under the control of the p10 or Pph promoter. The resultant pBacDD-2GP2-2GP4 and pBacDD-4mtGP5 clones were used to create recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M. (G) Images of BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M recombinant baculoviruses-infected Sf-9 cells at 3 days post-infection. All images were magnified at 200×.
Figure 7.
PCR amplification of the GP3 and M genes of PRRSV and schematic illustration of baculovirus surface display vectors carrying multiple expression cassettes for display of PRRSV proteins on the cell membrane. (A, B) DNA electrophoresis analysis of PCR product of the GP3 and M genes of PRRSV. (C, D) The resultant pBacDD-GP2-GP3-2GP4- and pBacDD-2mtGP5-2M plasmid was digested with the respective restriction enzymes to confirm the correctness of the recombinant fragments. After electrophoresis analysis with 1.2% agarose gel, the results showed that the target gene fragments were all in line with the expected size of 318 bp (panel C) and the expected size of 800 bp and 200 bp (panel D). (E, F) Schematic illustration of baculovirus surface display vectors carrying multiple expression cassettes for higher level display of GP2, GP3, GP4, mtGP5, and M proteins of PRRSV on the cell membrane. To simultaneously express and increase expression levels of multiple viral proteins of PRRSV, two novel baculoviral vectors pBacDD-GP2-GP3-2GP4 (panel E) and pBacDD-2mtGP5-2M (panel F), which carry multiple expression cassettes for higher level display of proteins on cell membrane were constructed. The transmembrane domain and cytoplasmic domain of gp64 and the signal peptide sequence of gp64 are shown. The GP2, GP3, GP4, mtGP5, and M genes of PRRSV was inserted between the gp64 signal peptide and gp64 cytoplasmic domains and CTD of the surface display vectors under the control of the p10 or Pph promoter. The resultant pBacDD-2GP2-2GP4 and pBacDD-4mtGP5 clones were used to create recombinant baculoviruses BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M. (G) Images of BacDD-GP2-GP3-2GP4 and BacDD-2mtGP5-2M recombinant baculoviruses-infected Sf-9 cells at 3 days post-infection. All images were magnified at 200×.


Table 1.
PRRSV primers used in this study.
| Gene | Accession no. | Sequence (5’-3’)* | Location | Expectedsize (bp) |
|---|---|---|---|---|
| GP2 | NC001961.1 | F:GTCGACTCACCATCGCCGGTT(SalI) | 12193-12207 | 444 |
| R:GGATCCACTCAAAGTACTATTATACAC(BamHI) | 14282-14303 | |||
| GP2 | NC001961.1 | F:GCTAGCTCACCATCGCCGGTTGGCTG(NheI) | 12193-12210 | 444 |
| R:ATCGATACTCAAAGTACTATTATACAC(ClaI) | 14282-14303 | |||
| GP3 | NC001961.1 | F:GTCGACTGCCTCACCCGGCAAGC(SalI) | 12864-12880 | 318 |
| R:GGATCCTGAAATGTTGTCATGGCG(BamHI) | 5443-5461 | |||
| GP4 | NC001961.1 | F:GCATGCCCATGTTTCAGTTCGAG(SphI) | 13313-13330 | 267 |
| R:ACCGGT TCCCTTTTCGCTCATCTC(AgeI) | 4430-4448 | |||
| GP4 | NC001961.1 | F:CTCGAGCCATGTTTCAGTTCGAG(XhoI) | 13313-13329 | 267 |
| R:GAATTC TCCCTTTTCGCTCATCTC(EcoRI) | 4430-4448 | |||
| GP5 | NC001961.1 | F:CTACTCGAGATGTTGGGGAAATGCTTGACC(Xho I) | 5441-5465 | 111 |
| R:ATCTCTAGACTCAGGACGACCCCATTGTTC(Xba I) | 6055-6079 | |||
| GP5 | NC001961.1 | F:GCGGCCGCATGTTGGGGAAATGCTTGACC(Not I) | 12354-12375 | 111 |
| R:GTCGACCTCAGGACGACCCCATTGTTC(Sal I) | 2407-2428 | |||
| GP5 | NC001961.1 | F:GCATGCATGTTGGGGAAATGCTTGACC(SphI) | 12354-12375 | 111 |
| R:ACCGGT CTCAGGACGACCCCATTGTTC(AgeI) | 2407-2428 | |||
| GP5 | NC001961.1 | F:CATATGATGTTGGGGAAATGCTTGACC(NdeI) | 12354-12375 | 111 |
| R:GCTAGCCTCAGGACGACCCCATTGTTC(NheI) | 2407-2428 | |||
| M | NC001961.1 | F:CTCGAGATGGGGTCGTCCCTAGAC(Xho I) | 14377-14395 | 48 |
| R:GAATTCCTGCGGAGCCGTGCT(EcoRI) | 6767-6782 | |||
| M | NC001961.1 | F:CATATGATGGGGTCGTCCCT(NdeI) | 14377-14391 | 48 |
| R:GCTAGCCTGCGGAGCCGTGC(Nhe I) | 6767-6781 |
*Underlines indicate the location of restriction sizes. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



