
Submitted:
14 August 2023
Posted:
15 August 2023
You are already at the latest version
Abstract
Phosphorus (P) is an essential plant macronutrient; however, its availability is often limited in soils. Plants have evolved complex mechanisms for efficient phosphate (Pi) absorption, which are responsive to changes in external and internal Pi concentration, and orchestrated through local and systemic responses. To explore these systemic Pi responses, here, we identified AtMYB44 as a phloem-mobile mRNA, an Arabidopsis homolog of Cucumis sativus MYB44, that is responsive to the Pi-starvation stress. qRT-PCR assays revealed that AtMYB44 was up-regulated and expressed in both shoot and root in response to Pi-starvation stress. The atmyb44 mutant displayed higher shoot and root biomass, compared to wild-type plants, under Pi-starvation conditions. Interestingly, the expression of PHOSPHATE TRANSPORTER1;2 (PHT1;2) and PHT1;4 was enhanced in atmyb44 in response to a Pi-starvation treatment. A split-root assay showed that AtMYB44 expression was systemically regulated, under Pi-starvation conditions and, in atmyb44, systemic controls on PHT1;2 and PHT1;4 expression were moderately disrupted. Heterografting assays confirmed graft transmission of AtMYB44 transcripts, and PHT1;2 and PHT1;4 expression was decreased in heterografted atmyb44 rootstocks. Taken together, our findings support the hypothesis that mobile AtMYB44 mRNA serves as a long-distance Pi response signal, which negatively regulates Pi transport and utilization in Arabidopsis.
Keywords:
Pi-starvation stress
; mobile mRNA
; phloem
; systemic signaling
1. Introduction
Phosphorus (P) is an essential element that is involved in various functionalities in living organisms, including plants. Therefore, it is critical to ensure proper phosphorus nutrition for optimal plant growth and development, by supplying phosphate (Pi) fertilizers to maximize crop productivity in modern agriculture systems [1,2]. However, soil-applied Pi fertilizer becomes rapidly immobilized, due to chemical reactions with cations (e.g., magnesium, calcium, aluminum, iron) in both alkaline and acidic soils. This can limit the availability to the plant of applied Pi to around 20-30% for plant use. Furthermore, Pi is a finite and non-renewable resource, therefore, it has led to increased interest in understanding the mechanisms by which plants absorb and use Pi, under limiting Pi conditions. Here, the goal is to develop elite plant lines having enhanced P acquisition and utilization efficiency traits [3,4,5,6,7].
Plants have evolved sophisticated adaptative mechanisms, which involve a range of physiological, morphological, biochemical, and molecular processes, to respond to low Pi conditions for efficient Pi foraging in soils [4,5,8,9,10,11]. These adaptative processes, known as phosphate starvation responses (PSRs), are categorized into local and systemic Pi responses as Pi sensing and long-distance signaling, and the adaptative efficiency can be determined by the PSR regulatory capacity in plants [6,10,12,13].
The local Pi responses are largely associated with traits for root growth, to perceive the Pi level in the soil and determine the root developmental fate, in response to the imposed Pi-starvation stress, whereas internal Pi concentration activates the systemic responses to integrate the Pi level information in distantly located plant tissues/organs [6,10,12,13]. Systemic Pi responses play an important role in global P homeostasis within plants, and microRNA399 (miR399) was the first identified long-distance regulatory component in Pi-stress signaling [14,15]. Under Pi-starvation conditions, miR399 is expressed in shoots and delivered into roots, via the phloem, to mediate in the degradation of PHO2 mRNA, which encodes an E2 ubiquitin-conjugating enzyme and regulates level of high-affinity PHOSTPHATE TRANSPORTER1 (PHT1) protein in roots. Therefore, this systemic regulatory mechanism serves in control over Pi acquisition under Pi-starvation stress conditions [6,14,15,16,17,18,19,20].
2. Results
2.1. Identification of mobile CsMYB44 orthologs in Arabidopsis.
In a previous study, cucumber was used as a model plant to examine the role of the phloem in systemic signaling under Pi-stress conditions [29]. The phloem mRNA profiles were observed to change, dynamically, in response to an imposed Pi-stress treatment, and a range of graft-transmissible mRNAs were shown to target specific sink organs/tissues, under Pi-stress conditions [29]. The Cucumis sativus MYB44 (CsMYB44, Csa6G491690), encoding a R2R3 MYB transcription factor (TF), was among these Pi-stress-induced graft-transmissible mRNAs and was delivered into sink tissue [29]. Thus, we selected CsMYB44 to further assess its role as a long-distance signaling mRNA in response to Pi-stress.
First, to facilitate testing our hypothesis that MYB44 plays a role in phloem-mediated systemic signaling, under Pi-starvation stress conditions, we conducted a phylogenetic analysis to identify putative CsMYB44 orthologs in Arabidopsis. A total of 132 Arabidopsis R2R3 MYB family members were analyzed (Figure S1) and, here, the phylogenetic tree analysis revealed the presence of CsMYB44 in a specific MYB subfamily subclade (Figure 1A).
To gain insight into the regulation of the genes in this MYB subclade, we used the Arabidopsis cis-regulatory element database (https://agris-knowledgebase.org/AtcisDB) to analyze the promoter regions of AtMYB44, AtMYB70, AtMYB73 and AtMYB77 with the aim to identify potential cis-elements that might serve to control their expression under Pi-stress conditions. Numerous Pi-starvation responsive (PSR) genes are regulated by the PHOSPHATE STARVATION RESPONSE1 (PHR1) TF, a key regulator of PSR genes, and the PHR1 binding site (P1BS) is enriched in the promoter regions of many Arabidopsis Pi-responsive genes [30,31,32].
Our cis-element analysis indeed revealed that P1BS motifs were located within the promoters of AtMYB44, AtMYB70 and AtMYB73, but not AtMYB77 (Figure 1B). Furthermore, qRT-PCR analysis established that the transcript levels of AtMYB44, AtMYB70, AtMYB73 and AtMYB77 were responsive to a Pi- stress treatment, in both shoots and roots (Figure 1C-D). For AtMYB44 and AtMYB77, at least a two-fold increase and decrease in transcript levels, respectively, were detected in shoots, under Pi-stress conditions (Figure 1C). By contrast, AtMYB70 and AtMYB73 transcript levels in the shoots were only slightly changed in response to Pi-stress (10 µM Pi) (Figure 1C). In roots, the AtMYB44, AtMYB70, AtMYB73 and AtMYB77 transcript levels were significantly increased under Pi-stressed conditions (10 µM Pi) (Figure 1D). Taken together, our results demonstrate that the putative CsMYB44 homologs, AtMYB44, AtMYB70, AtMYB73 and AtMYB77, are regulated in response to the imposed Pi-stress treatment. As the AtMYB44 transcript levels increased, in both shoot and root, we focused on investigation on the role of AtMYB44 in Pi-stress signaling.
2.2. AtMYB44 is expressed in leaf and root vascular tissues under Pi-stress conditions.
To further understand the mechanism of AtMYB44 regulation in Arabidopsis, we generated transgenic plants carrying the GFP-GUS reporter, under the control of the native AtMYB44 promoter and analyzed the AtMYB44 expression pattern (Figure 2). Consistent with our qRT-PCR results (Figure 1C,D), the β-glucuronidase (GUS) staining assays showed signal in both shoots and roots, and revealed stronger staining under Pi-stress (10 µM), compared with Pi-sufficient conditions (500 µM) (Figure 2A,B), consistent with induction of AtMYB44 expression in response to Pi-stress. Here, AtMYB44 expression was detected in leaf lamina, leaf vasculature, root vasculature, root tips of primary and lateral roots, and in lateral root primordia (Figure 2B–H). These findings were consistent with the notion that AtMYB44 plays a role in transcriptional regulation of adaptive shoot and root development, in response to Pi-stress treatment.
2.3. AtMYB70, AtMYB73 and AtMYB77 transcript levels were elevated in atmyb44 mutant, compared with Wild-type.
The atmyb44 mutant exhibited slightly longer primary roots and a higher number of lateral roots, compared with the WT, under Pi-sufficient conditions (Figure S2A–C). However, under Pi-stress conditions, no significant difference in primary root growth and lateral root numbers were observed between WT and atmyb44. As expression of other MYB family genes, close to CsMYB44, was also responsive to imposed Pi-stress (Figure 1D), we hypothesized that other redundant MYBs could compensate in the absence of AtMYB44 under Pi-stress conditions. To test this notion, we examined the expression level of AtMYB70, AtMYB73 and AtMYB77 in the atmyb44 mutant background, under Pi-stress conditions, using qRT-PCR (Figure 3). Here, we confirmed that expression of AtMYB70, AtMYB73 and AtMYB77 increased (Figure 3). Enhanced transcript levels of AtMYB70, AtMYB73 and AtMYB77 were detected in atmyb44 plants under both Pi-sufficient and stress conditions, compared with WT (Figure 3). These assays supported a role for AtMYB70, AtMYB73 and AtMYB77, in the functional compensation of AtMYB44 in its absence.
2.4. The atmyb44 root had an elevated Pi concentration under Pi-stress conditions.
Under the Pi-stress conditions, the atmyb44 plants did not exhibit a clear phenotypic difference in primary root growth and lateral root number, compared with WT (Figure S2). As both plant lines were grown on solid medium, which limited observation of plant developmental changes, to apply a prolonged Pi-stress, we next employed a hydroponic system to conduct phenotypic analysis of atmyb44 and AtMYB44 overexpression (OX) lines [33,34,35].
Compared to WT, higher and lower shoot biomass was observed in the atmyb44 and AtMYB44 OX plants, respectively, under Pi-sufficient conditions, although no significant difference in shoot biomass was detected under Pi-stress conditions (Figure 4A, Figure S3A). However, the root biomass was obviously higher in atmyb44 than WT and AtMYB44 OXs, under both Pi-sufficient and stress conditions (Figure 4B, Figure S3B). In atmyb44, the Pi concentration was higher in shoots and lower in roots, compared to the level in WT, under Pi-sufficient conditions. In contrast, the root Pi concentration in atmyb44 was higher than WT, under Pi-stress conditions, even though no significant difference in shoot Pi concentration was detected between WT and atmyb44 (Figure 4C,D). Taken together, these results suggested that AtMYB44 functions as a negative regulator in both shoot and root development, under Pi-sufficient conditions and that an absence of AtMYB44 might increase Pi uptake into Arabidopsis roots.
2.5. Expression of PHT1;2 and PHT1;4 is negatively regulated by AtMYB44 in roots.
The in-silico analysis, using ConnecTF (https://connectf.org) [36], revealed that AtMYB44 could be involved in many gene regulatory pathways and, interestingly, bound to the promoter regions of PHOSPAHTE TRANSPORTER 1;2 (PHT1;2) and PHT1;4 as potential AtMYB44-target sites (Table S2). As we observed an enhanced level of Pi concentration in the root of atmyb44 plants, under Pi-stress conditions (Figure 4D), and PHT1 regulates the initial uptake of Pi [6,37,38,39,40,41,42,43,44], we hypothesized that AtMYB44 regulates PHT1 expression in roots for Pi acquisition under Pi-stress conditions.
To test this hypothesis, we examined the expression level of PHT1;2 and PHT1;4 in WT, atmyb44 and AtMYB44 OX-1 roots, using qRT-PCR (Figure 5). The expression levels of PHT1;2 and PHT1;4 were much higher in atmyb44, under Pi-stress conditions, compared to WT (two- to four-fold higher expression; Figure 5) and the increased levels of PHT1;2 and PHT1;4 in atmyb44 were diminished in AtMYB44 OX-1 (Figure 5). We did not detect any significant differences in the expression levels of PHT1;2 and PHT1;4 in AtMYB44 OX-1, compared with WT (Figure 5). This result suggested that AtMYB44 targets the promoter of PHT1;2 and PHT1;4, where it acts as a negative regulator.
2.6. Mobile AtMYB44 mRNA functions as a negative regulator of PHT1;2 and PHT1;4 expression.
We demonstrated that AtMYB44 expression responds to Pi-stress (Figure 1C,D) and AtMYB44 appears to play a role in regulating PHT1;2 and PHT1;4 expression, under these conditions (Figure 5). As AtMYB44 was identified as a mobile CsMYB44 ortholog in Arabidopsis (Figure 1A), it is plausible that AtMYB44 could serve as a long-distance signal to exert control over Pi-starvation responses in Arabidopsis. To test whether AtMYB44 acts as a systemic signal, in response to Pi-stress, we employed a split-root system to test whether AtMYB44-mediated gene expression is a systemic or local response, under Pi-stress conditions. The Arabidopsis root systems grown in the hydroponic culture system, were separated into two parts with each half of the root system placed into a vessel container nutrient solution with Pi (200 µM) or with no Pi (0 µM). As the control, both vessels for each half of the root system on a plant contained the same nutrient solution (CP200 for 200 µM Pi, CP0 for 0 µM) to mimic plant growth under homogenous Pi-sufficient or deficient conditions [12].
Total RNA was extracted from these roots for qRT-PCR analysis. First, we investigated regulation of AtMYB44 expression, to see if it is part of a systemic response, after Pi-sufficient and deficient treatments were applied to different halves of the root system on individual plants area (Figure 6). As expected, no AtMYB44 transcript was detected in atmyb44 tissues and AtMYB44 expression in WT was enhanced in response to the imposed Pi-stress (C0), compared with the mRNA level under Pi-sufficient conditions (CP200) (Figure 6A). In WT, a higher level of AtMYB44 mRNA was detected in that half of a plant’s root system placed in the compartment with 200 µM Pi (SP200), relative to the control (CP200). In addition, lower AtMYB44 mRNA levels were observed for the split root system in 0 µM Pi (SP0) compared to the transcript level in the homogenously Pi-deficient control (C0) (Figure 6A). These findings support our hypothesis that AtMYB44 serves as a systemic regulator.
Next, we examined systemic regulation of PHT1;2 and PHT1;4 expression in AtMYB44 knockout mutants again using this split-root system to test whether AtMYB44 serves as a factor in systemic PHT1;2 and PHT1;4 regulation. Here, the qRT-PCR analysis revealed that, consistent with the previous report [12], expression of both PHT1;2 and PHT1;4 in WT plants was downregulated in roots within the SP0 container, compared to PHT1;2 and PHT1;4 expression in CP0 in WT (Figure 6B,C). However, in atmyb44 plants, the enhanced transcript levels of PHT1;2 and PHT1;4 were decreased in SP0, compared to CP0 (Figure 6B,C). Taken together, these results suggest that AtMYB44 expression is regulated, in a systemic manner, and that other long-distance regulators might be involved in the systemic regulation of PHT1;2 and PHT1;4 regulation in the atmyb44 background.
To investigate whether AtMYB44 mRNA acts as a long-distance regulator for control over PHT1;2 and PHT1;4 expression, we performed micrografting assays between atmyb44 and AtMYB44 OX-1 lines (Figure 7A). Here, our RT-PCR analyses revealed that, as controls, AtMYB44 expression was increased in AtMYB44 OX-1, but not in atmyb44, and AtMYB44 OX-1, and atmyb44 were subsequently used as shoot scions or root stocks (Figure 7B). Analysis of these grafted tissues revealed that the transgene AtMYB44 was detected in rootstocks of heterografted AtMYB44 OX-1 (scion)/atmyb44 (rootstock), but not in the atmyb44 (scion)/ AtMYB44 OX-1 (rootstock), consistent with AtMYB44 mRNA being phloem-mobile from source to sink tissues (Figure 7B). Interestingly, the level of PHT1;2 and PHT1;4 expression was decreased in the rootstocks of these heterografted AtMYB44 OX-1 (scion)/atmyb44 (rootstock) plants, compared to the atmyb44 homograft (Figure 7C). These data support the hypothesis that AtMYB44 mRNA acts as a mobile, negatively acting, regulator of PHT1;2 and PHT1;4 expression in Arabidopsis roots.
3. Discussion
The local and systemic regulatory mechanisms acting in Pi acquisition and distribution, under Pi-stress conditions, have been extensively studied and many important genes have been identified as crucial components that regulate Pi homeostasis in plants, such as Arabidopsis and rice [4,5,6,34,44,45,46,47]. However, limited information is available on the nature of the systemic Pi signaling agents that control Pi homeostasis, at the whole-plant level [6,14,15,17,29,48,49,50,51]. In this study, we identified AtMYB44 mRNA as a potential mobile systemic Pi signaling component in Arabidopsis.
3.1. AtMYB44 expression responds to an imposed Pi-stress.
In a previous study, we identified CsMYB44 as mobile mRNA that accumulated in sink tissues, under an applied Pi-stress condition [29]. As mobile CsMYB44 was detected in sink tissues, during the early stage of Pi-stress treatment, we hypothesized that this mobile CsMYB44 mRNA serves as an early long-distance signaling factor for systemic Pi responses. Four homologs of CsMYB44 were identified, as potential CsMYB44 orthologs in Arabidopsis (Figure 1A, Figure S1), and their expression patterns were also responsive to an imposed Pi-stress (Figure 1B–D). As the promoter regions of AtMYB44, AtMYB70 and AtMYB73, but not AtMYB77, included PIBS motifs, this suggested that AtMYB44, AtMYB70 and AtMYB73 expression could be regulated in a PHR1-dependent manners under Pi-stress conditions.
An increase in the level of AtMYB44 expression during Pi-starvation treatment, and detection of GUS signals within the vascular tissues of both shoot and root (Figure 2), suggested that AtMYB44 could function within the vasculature in response to Pi-starvation stress. In addition, we also observed that AtMYB44 expression was strongly detected in the root tips and lateral root primordia (Figure 2E,F). Based on our in silico analysis, the AtMYB44 TF could be associated with the promoter regions of many AUXIN RESPONSE FACTORs (ARFs), which contribute to control over PHR1 expression in Arabidopsis roots under Pi-starvation conditions [52,53] (Table S2). As AtMYB44 expression is responsive to auxin levels in the root [54], AtMYB44 may participate in auxin-dependent root development in response to Pi-stress.
AtMYB44, AtMYB70, AtMYB73 and AtMYB77 share high structural similarity and participate in root system development [24,55,56]. Interestingly, we detected enhanced expression of AtMYB70, AtMYB73, and AtMYB77 in atmyb44 plants, compared with WT (Figure 3). It was earlier proposed that AtMYB44 is a functional paralog of AtMYB73 and AtMYB77 for auxin-mediated lateral root growth and development [55]. Even though the functional redundancy of AtMYB70 with AtMYB44, in root growth has not yet been examined, it is implicit that AtMYB70, AtMYB73 and AtMYB77 are redundant with AtMYB44 to coordinate adaptive root development, under Pi-stress conditions.
Like a previous report [25], the atmyb44 root appears to have slightly enhanced root growth performance under both Pi-sufficient and -stress conditions, compared to WT (Figures 4A,B, S2 and S3). One proposed AtMYB44 function is as a negative regulator in abscisic acid (ABA) responses, which are involved in root growth inhibition [25,57,58]. AtMYB44 interacts with PYRABACTIN RESISTANCE 1-LIKE 8 (PYL8), identified as an ABA receptor, to regulate early ABA signaling and promote lateral root growth [25,55]. ABA signaling can play a role in enhancing the promoter activity of various auxin-responsive genes [58]. Although limited information is available regarding the role of ABA in Pi-starvation responses [59], it is plausible that AtMYB44 might inhibit ABA responses and activate auxin signaling to enhance root development in response to the imposed Pi-stress. However, AtMYB44 OX lines did not show a clearly opposite phenotype with the atmyb44 line, under Pi-stress (Figures 4A,B, S2 and S3). Although this result could not be explained with the role of AtMYB44 in Pi-starvation signaling, as AtMYB44 has been proposed to be involved in multiple signaling pathways, ectopic expression of AtMYB44 might result in aberrant ABA and auxin signaling to establish unidentified molecular responses, under these Pi-stress conditions.
3.2. AtMYB44 mRNA acts as a systemic Pi signaling factor to negatively regulate root Pi transport systems.
It was earlier reported that MYB TFs are involved in the control over Pi-starvation responses in plants [28,45,60,61,62]. Arabidopsis MYB62, induced in response to Pi-stress, appears to function as a negative regulator for PSR gene expression and be involved in gibberellic acid biosynthesis [61]. Another MYB-like TF, REGULATOR OF LEAF INCLINATION 1 (RLI1), interacts with SPX1 (for Syg1/Pho81/XPR1) and the SPX1-RLI1 complex appears to block RLI1 locating to the promoter regions of RLI1 target genes, which are involved in elongation of lamina join cells in rice [63,64]. Additionally, potato MYB44 functions as a negative regulator for PHOSPHATE1 (PHO1) expression, whose protein is engaged in Pi transport, from root to shoot, through Pi loading into the xylem [28].
In this study, we provide insight into the function of AtMYB44 as a negative regulator in systemic responses to Pi-stress. Enhanced PHT1;2 and PHT1;4 expression and elevated soluble Pi concentration were detected in atmyb44 plants, compared to WT (Figures 4C,D and 5). It is noticeable that the soluble Pi concentration was higher in shoots and lower in roots of the atmyb44 mutant, under Pi-sufficient conditions, compared with WT (Figure 4C,D). This suggests that, in Arabidopsis, AtMYB44 plays a negative role in root-to-shoot Pi transport under Pi-sufficient condition. Hence, Pi translocation through the xylem might be enhanced in atmyb44 plants.
Interestingly, under Pi-stress conditions, although a similar level of soluble Pi concentration was detected in the shoots of WT and atmyb44, its level was higher in atmyb44 roots, and this result seems to be correlated with increased root biomass (Figure 4 and S2). Additionally, shoot biomass of atmyb44 was significantly elevated under sufficient Pi, but not Pi-limiting conditions (Figure 4). It is possible that AtMYB44 functions in roots as a negative regulator of PHT1;2 and PHT1;4 (Figure 5), thus, enhanced levels of PHT1;2 and PHT1;4 would increase the efficiency of Pi uptake and plant growth performance in atmyb44 plants under Pi-stress conditions. Taken together, it appears that AtMYB44 might play a negative role in Pi transport from root to shoot, under Pi-sufficient conditions, but during a Pi-stress treatment, it might act as a negative regulator on Pi uptake in Arabidopsis.
The split-root assays demonstrated that AtMYB44 expression is systemically regulated by Pi-stress (Figure 6A). Consistent with a previous study [12], systemic responses of PHT1;2 and PHT1;4 were observed and, interestingly, such long-distance regulation was partially disrupted in the atmyb44 mutant background (Figure 6B,C). The mobility of AtMYB44 mRNA appeared to reduce the level of PHT1;2 and PHT1;4 transcript abundance in roots (Figure 7). In this regard, various RNA species, including mRNAs, small interfering RNAs, non-coding RNAs, mRNAs etc., have been shown to be translocated through the phloem and some mobile RNA molecules can act as signaling agents in plant development and physiology [14,29,49,50,65,66,67,68,69,70,71]. For example, GIBBERELLIC ACID-INSENSITIVE mRNA is transported from the source to sink tissues and contributes to regulating leaf development [72,73]. Additionally, potato tuberization is regulated by transport of SP6A mRNA from potato leaves to the underground stolon [74,75]. The shoot-derived INDOLEACETIC ACID18 (IAA18), IAA28 and TRANSLATIONALLY CONTROLLED TUMOUR PROTEIN (TCTP) traffic into the roots to regulate the lateral root development [76,77,78].
In our study, we propose that mobile AtMYB44 mRNA serves as a long-distance phloem-based signal in the Pi-stress response (Figure 7). Although AtMYB44 lacked modified base 5-methylcytosine (m5C), CU- or tRNA-like motifs, which have been characterized as contributing to phloem mRNA mobility [79,80,81], our heterografting assay established the mobility of AtMYB44 mRNA in Arabidopsis (Figure 7and S4). As AtMYB44 and CsMYB44 protein have not been detected in the phloem exudate [82,83], it is likely that AtMYB44 mRNA and not protein serves as the signaling agent in Pi homeostasis. Although mobile AtMYB44 mRNA is involved in regulating PHT1 expression, in roots, the regulatory mechanism underlying its phloem mobility, in Pi acquisition and utilization, remains to be elucidated. Based on previous studies [80,81], the shoot derived AtMYB44 mRNA could be transported through the phloem and then translated in the targeted root tissues to regulate Pi transport.
Our study showed enhanced AtMYB44 expression, under Pi-stress conditions, and a negative role for AtMYB44 in PHT1 expression in Arabidopsis roots (Figure 8). As an increase in both AtMYB44 and PHT1 expression was detected under Pi-stress, this raises the question as to why plants would enhance AtMYB44 expression, to repress the Pi transport system under Pi-stress conditions, even though one would expect these plants should increase their Pi uptake capacity, to adapt under limited Pi input. Mobile AtMYB44 mRNA might play a role in fine-tuning the regulation of Pi homeostasis, in response to Pi-stress, for control over energy balance between adaptive plant development and Pi uptake (Figure 8). Our current findings provide insight into plant Pi-stress regulatory pathways and further studies will be required to reveal the molecular mechanism by which such mobile AtMYB44 mRNA effect the ability of the plant to acquire and utilize Pi during Pi-stress conditions. Such information would be of value in genetic engineering of crops for improved yield performance under reduced Pi fertilizer applications.
4. Materials and Methods
4.1. Plant materials
Arabidopsis plants were grown in controlled environment chambers under long-day conditions (16 h light/8 h dark, 120-150 μmol m-2 s-1 of photosynthetically active radiation), at temperatures of 22° C day/18°C night [84,85]. The T-DNA knock-out line (SALK_039074 [atmyb44]) for AT5G67300 was obtained from the Arabidopsis Biological Resource Center (ABRC). The genotypes were confirmed using PCR analysis with appropriate primer sets (Table S1).
4.2. Growth conditions and Pi-stress treatments
Arabidopsis plant lines were grown on solid medium (pH 5.7), in sand, or in a hydroponic culture system, as described previously [29,84,86,87]. Briefly, Arabidopsis seeds were sterilized, germinated on solid medium with 500 µM NH4H2PO4 and then, at 7 days after germination (DAG), they were transferred onto fresh medium or medium with (NH4)2SO4 that was partially, or completely replaced, NH4H2PO4. For hydroponically grown Arabidopsis plants, the tip of a 10 µL pipette was cut and then filled with 0.7% (w/v) agar, upon which sterilized seeds were sown, prior to setting them up on a floating board system. Hydroponic solution was replaced every 5 days. The solution for Pi-sufficient and starvation treatment contained 200 µM and 5 µM of NH4H2PO4, respectively. For growing Arabidopsis plants in sand, seeds were germinated on solid medium with 500 µM (Pi-sufficient), or 10 µM (Pi-starvation) of NH4H2PO4 for 14 days, followed by transplanting to silica sand. Arabidopsis plants were further grown in silica sand for 14 days with nutrient solution, which contained 500 µM (Pi-sufficient), or 10 µM (Pi-starvation) of NH4H2PO4, supplied to the roots. Images of root system architecture were captured with a Nikon D7200 (Nikon) digital camera. Acquired images were processed and analyzed using ImageJ software, as described previously [88].
Shoot and root tissues were collected from hydroponically cultured Arabidopsis plants and were used to measure biomass and Pi concentration, as described previously [34,87]. Briefly, collected shoot and root tissues were frozen in liquid nitrogen after measurement of fresh weight and then homogenized using a bead beater. Ground samples were digested with 5M H2SO4 and then Pi concentrations were determined using a continuous flow analyzer (Skalar), following the manufacturer’s instructions.
Four-week-old Arabidopsis plants were used for the split-root assays, as described previously [12,89], with modifications. Briefly, Arabidopsis roots were divided into two parts and then immersed in two separate compartments, which contained either the same nutrient solution with 200 µM Pi (C200) or 0 µM Pi (C0), or a different nutrient solution with 200 µM Pi (SP200) and 0 µM Pi (SP0), for 7 days [12,89].
4.3. Phylogenetic analysis
The amino acid sequences of AtMYBs were obtained from the Arabidopsis Information Resource Database (https://www.arabidopsis.org/) and aligned using MEGA 11 software. The phylogenetic tree was constructed using a neighbor-Joining (NJ) method with 1,000 bootstrap replicates.
4.4. β-Glucuronidase (GUS) histochemical analysis
The 2,916 bp upstream region, which was from the ATG start codon of AtMYB44, was amplified from genomic DNA of Arabidopsis WT (Col-0), using the primer set listed in Table S1. The fragment was then inserted into the TOPO-D vector (Invitrogen), followed by LR clonase reaction (Invitrogen), to subclone the AtMYB44 promoter into pBGWFS7. The construct was introduced into Agrobacterium tumefaciens strain GV3101 to generate transgenic Arabidopsis plants. The GUS staining of transgenic plants, carrying a GUS reporter gene under the control of the AtMYB44 native promoter, was conducted, as described previously [84]. Briefly, 12-day-old transgenic plants were immersed in 50 mM sodium phosphate buffer (pH 7.0) with 1 mM K3Fe(CN)6, 1 mM K4Fe(CN)6, 2 mM 5-bromo-4-chloro-3-indolyl β-D-glucuronide sodium salt and 10 mM EDTA (pH 8.0) and incubated at 37 °C. The chlorophyll was eliminated using 70% ethanol. T3 plants collected from three independent transgenic lines were used for GUS assays and imaged, using a THUNDER stereomicroscope (Leica) and Zeiss Axioskop 2 Plus microscope.
4.5. Micrografting
Arabidopsis micrografting was performed, as described previously [76,90]. Briefly, hypocotyls of 5-day-old Arabidopsis seedlings, grown on 1/2 × MS medium with 0.8% agar, were cut on nylon membranes using a surgical blade. The scion and stock were placed together to connect the shoot and root. Grafted Arabidopsis plants were grown on 1/2 × MS medium with 1.5% agar for 5 days. Successfully grafted plants were transferred to 1/2 × MS medium with 0.8% agar for further studies.
4.6. RNA extraction and qRT-PCR
Total RNA was extracted from Arabidopsis leaves and roots, using the TRIzol® Reagent (Invitrogen, Life Technologies), following the manufacturer’s instructions. Total RNA of 1 µg was used for cDNA synthesis with the SuperScript IV first-strand synthesis system (Invitrogen). The qRT-PCR analysis was conducted, as described previously [91]. Briefly, the qRT-PCR was performed with PowerUp™ SYBR™ Green Master Mix (Thermo Fisher), using the QuantStudio™ 6 Flex Real-Time PCR Systems (Life Technologies), to detect AtMYB44, AtMYB70, AtMYB73, AtMYB77, PHT1;2 and PHT1;4, with the primer sets listed in Table S1. AtActin was used as a reference gene for normalization of transcript levels. Transcript levels and ratios were calculated using the 2−∆Ct or the 2−∆∆Ct method, respectively. Statistical analyses were performed with the Student’s t-test and Tukey’s HSD (honestly significant difference) test. At least three biological and three technical replicates were used for qRT-PCR analyses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Phylogenetic analysis of the A. thaliana MYB family; Figure S2: AtMYB44 is involved in primary and lateral root development; Figure S3: AtMYB44 serves as a potential negative regulator in shoot and root development; Figure S4: Mobility of AtMYB44 mRNA Table S1: List of PCR primers used in this study; Table S2: Potential AtMYB44-mediated gene regulation in Arabidopsis.
Author Contributions
TO: JC and BH conceived the project. TO, JC and BH designed the experiments. TO, JC, YZ and CQ conducted the experiments; BH and LK provided technical advice. All authors contributed to perform the data analysis and interpretation. JC and TO prepared the draft and BH revised the manuscript. All authors read and approved this article.
Funding
Financial support for this article was provided by the Canada Excellence Research Chair for Food Systems and Security (to L.V.K.), the Global Institute for Food Security (to L.V.K and B.-K.H.), Canada Foundation for Innovation – John R. Evans Leaders Fund (CFI#38103 to B.-K.H.), Natural Sciences and Engineering Research Council of Canada (#RGPIN-2019-04421 to BKH) and the New Frontiers in Research Fund NFRFE-2020-01108 (to B.-K.H.).
Data Availability Statement
The data in this study are available within the article.
Acknowledgments
Transgenic lines, overexpressing AtMYB44 (42-6 and 31-2 were referred to as AtMYB44 OX-1 and OX-2 in this study, respectively), were kindly provided by Dr. Huang Daiqing at the National Research Council of Canada, Saskatoon.
Conflicts of Interest
All authors declare no conflict of interest.
References
- MacDonald, G.K.; Bennett, E.M.; Potter, P.A.; Ramankutty, N. Agronomic phosphorus imbalances across the world’s croplands. Proc Natl Acad Sci U S A 2011, 108, 3086–3091. [Google Scholar] [CrossRef] [PubMed]
- Withers, P.J.; Sylvester-Bradley, R.; Jones, D.L.; Healey, J.R.; Talboys, P.J. Feed the crop not the soil: rethinking phosphorus management in the food chain. Environ Sci Technol 2014, 48, 6523–6530. [Google Scholar] [CrossRef] [PubMed]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.R.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol 2012, 195, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Arredondo, D.L.; Leyva-Gonzalez, M.A.; Gonzalez-Morales, S.I.; Lopez-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: improving low-phosphate tolerance in crops. Annu Rev Plant Biol 2014, 65, 95–123. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhaes, J.V. Plant Adaptation to Acid Soils: The Molecular Basis for Crop Aluminum Resistance. Annu Rev Plant Biol 2015, 66, 571–598. [Google Scholar] [CrossRef] [PubMed]
- Ham, B.K.; Chen, J.Y.; Yan, Y.; Lucas, W.J. Insights into plant phosphate sensing and signaling. Curr Opin Biotechnol 2018, 49, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Root phenotypes for improved nutrient capture: an underexploited opportunity for global agriculture. New Phytol 2019, 223, 548–564. [Google Scholar] [CrossRef]
- Morcuende, R.; Bari, R.; Gibon, Y.; Zheng, W.; Pant, B.D.; Blasing, O.; Usadel, B.; Czechowski, T.; Udvardi, M.K.; Stitt, M.; et al. Genome-wide reprogramming of metabolism and regulatory networks of Arabidopsis in response to phosphorus. Plant Cell Environ 2007, 30, 85–112. [Google Scholar] [CrossRef]
- Lei, M.G.; Zhu, C.M.; Liu, Y.D.; Karthikeyan, A.S.; Bressan, R.A.; Raghothama, K.G.; Liu, D. Ethylene signalling is involved in regulation of phosphate starvation-induced gene expression and production of acid phosphatases and anthocyanin in Arabidopsis. New Phytologist 2011, 189, 1084–1095. [Google Scholar] [CrossRef]
- Puga, M.I.; Rojas-Triana, M.; de Lorenzo, L.; Leyva, A.; Rubio, V.; Paz-Ares, J. Novel signals in the regulation of Pi starvation responses in plants: facts and promises. Curr Opin Plant Biol 2017, 39, 40–49. [Google Scholar] [CrossRef]
- Li, L.; Liu, K.H.; Sheen, J. Dynamic Nutrient Signaling Networks in Plants. Annu Rev of Cell Dev Biol 2021, 37, 341–367. [Google Scholar] [CrossRef]
- Thibaud, M.C.; Arrighi, J.F.; Bayle, V.; Chiarenza, S.; Creff, A.; Bustos, R.; Paz-Ares, J.; Poirier, Y.; Nussaume, L. Dissection of local and systemic transcriptional responses to phosphate starvation in Arabidopsis. Plant J 2010, 64, 775–789. [Google Scholar] [CrossRef]
- Chen, J.; Ham, B.K. Systemic Signaling: A Role in Propelling Crop Yield. Plants 2022, 11. [Google Scholar] [CrossRef]
- Aung, K.; Lin, S.I.; Wu, C.C.; Huang, Y.T.; Su, C.L.; Chiou, T.J. pho2, a phosphate overaccumulator, is caused by a nonsense mutation in a MicroRNA399 target gene. Plant Physiol 2006, 141, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Pant, B.D.; Buhtz, A.; Kehr, J.; Scheible, W.R. MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis. Plant J 2008, 53, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Mudge, S.R.; Rae, A.L.; Diatloff, E.; Smith, F.W. Expression analysis suggests novel roles for members of the Pht1 family of phosphate transporters in Arabidopsis. Plant J 2002, 31, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.I.; Chiang, S.F.; Lin, W.Y.; Chen, J.W.; Tseng, C.Y.; Wu, P.C.; Chiou, T.J. Regulatory network of microRNA399 and PHO2 by systemic signaling. Plant Physiol 2008, 147, 732–746. [Google Scholar] [CrossRef]
- Nagarajan, V.K.; Jain, A.; Poling, M.D.; Lewis, A.J.; Raghothama, K.G.; Smith, A.P. Arabidopsis Pht1;5 Mobilizes Phosphate between Source and Sink Organs and Influences the Interaction between Phosphate Homeostasis and Ethylene Signaling. Plant Physiol 2011, 156, 1149–1163. [Google Scholar] [CrossRef]
- Huang, T.K.; Han, C.L.; Lin, S.I.; Chen, Y.J.; Tsai, Y.C.; Chen, Y.R.; Chen, J.W.; Lin, W.Y.; Chen, P.M.; Liu, T.Y.; et al. Identification of downstream components of ubiquitin-conjugating enzyme PHOSPHATE2 by quantitative membrane proteomics in Arabidopsis roots. Plant Cell 2013, 25, 4044–4060. [Google Scholar] [CrossRef]
- Park, B.S.; Seo, J.S.; Chua, N.H. NITROGEN LIMITATION ADAPTATION recruits PHOSPHATE2 to target the phosphate transporter PT2 for degradation during the regulation of Arabidopsis phosphate homeostasis. Plant Cell 2014, 26, 454–464. [Google Scholar] [CrossRef]
- Martin, C.; Paz-Ares, J. MYB transcription factors in plants. Trends Genet 1997, 13, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr Opin Plant Biol 2001, 4, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Seo, J.S.; Han, S.W.; Koo, Y.J.; Kim, C.H.; Song, S.I.; Nahm, B.H.; Choi, Y.D.; Cheong, J.J. Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis. Plant Physiol 2008, 146, 623–635. [Google Scholar] [CrossRef]
- Jaradat, M.R.; Feurtado, J.A.; Huang, D.; Lu, Y.; Cutler, A.J. Multiple roles of the transcription factor AtMYBR1/AtMYB44 in ABA signaling, stress responses, and leaf senescence. BMC Plant Biol 2013, 13, 192. [Google Scholar] [CrossRef] [PubMed]
- Persak, H.; Pitzschke, A. Dominant repression by Arabidopsis transcription factor MYB44 causes oxidative damage and hypersensitivity to abiotic stress. Int J Mol Sci 2014, 15, 2517–2537. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, M.; Jia, Z.; Liu, F.; Ma, H.; Huang, Y.; Song, S. AtMYB44 Positively Regulates the Enhanced Elongation of Primary Roots Induced by N-3-Oxo-Hexanoyl-Homoserine Lactone in Arabidopsis thaliana. Mol Plant Microbe Interact 2016, 29, 774–785. [Google Scholar] [CrossRef]
- Zhou, X.; Zha, M.; Huang, J.; Li, L.; Imran, M.; Zhang, C. StMYB44 negatively regulates phosphate transport by suppressing expression of PHOSPHATE1 in potato. J Exp Bot 2017, 68, 1265–1281. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Zheng, Y.; Ham, B.K.; Chen, J.Y.; Yoshida, A.; Kochian, L.V.; Fei, Z.J.; Lucas, W.J. Vascular-mediated signalling involved in early phosphate stress response in plants. Nature Plants 2016, 2. [Google Scholar] [CrossRef]
- Müller, R.; Morant, M.; Jarmer, H.; Nilsson, L.; Nielsen, T.H. Genome-wide analysis of the Arabidopsis leaf transcriptome reveals interaction of phosphate and sugar metabolism. Plant Physiol 2007, 143, 156–171. [Google Scholar] [CrossRef]
- Bustos, R.; Castrillo, G.; Linhares, F.; Puga, M.I.; Rubio, V.; Perez-Perez, J.; Solano, R.; Leyva, A.; Paz-Ares, J. A central regulatory system largely controls transcriptional activation and repression responses to phosphate starvation in Arabidopsis. PLoS Genet 2010, 6, e1001102. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Song, L.; Zhang, Y.; Zheng, Z.; Liu, D. Arabidopsis PHL2 and PHR1 Act Redundantly as the Key Components of the Central Regulatory System Controlling Transcriptional Responses to Phosphate Starvation. Plant Physiol 2016, 170, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Jaradat, M.R.; Wu, W.; Ambrose, S.J.; Ross, A.R.; Abrams, S.R.; Cutler, A.J. Structural analogs of ABA reveal novel features of ABA perception and signaling in Arabidopsis. Plant J 2007, 50, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, Y.; Ni, J.; Wang, Y.; Bai, Y.; Shi, J.; Gan, J.; Wu, Z.; Wu, P. OsPHF1 regulates the plasma membrane localization of low- and high-affinity inorganic phosphate transporters and determines inorganic phosphate uptake and translocation in rice. Plant Physiol 2011, 157, 269–278. [Google Scholar] [CrossRef]
- Yan, J.; Chia, J.C.; Sheng, H.; Jung, H.I.; Zavodna, T.O.; Zhang, L.; Huang, R.; Jiao, C.; Craft, E.J.; Fei, Z.; et al. Arabidopsis Pollen Fertility Requires the Transcription Factors CITF1 and SPL7 That Regulate Copper Delivery to Anthers and Jasmonic Acid Synthesis. Plant Cell 2017, 29, 3012–3029. [Google Scholar] [CrossRef]
- Brooks, M.D.; Juang, C.L.; Katari, M.S.; Alvarez, J.M.; Pasquino, A.; Shih, H.J.; Huang, J.; Shanks, C.; Cirrone, J.; Coruzzi, G.M. ConnecTF: A platform to integrate transcription factor-gene interactions and validate regulatory networks. Plant Physiol 2021, 185, 49–66. [Google Scholar] [CrossRef]
- Daram, P.; Brunner, S.; Persson, B.L.; Amrhein, N.; Bucher, M. Functional analysis and cell-specific expression of a phosphate transporter from tomato. Planta 1998, 206, 225–233. [Google Scholar] [CrossRef]
- Muchhal, U.S.; Pardo, J.M.; Raghothama, K.G. Phosphate transporters from the higher plant Arabidopsis thaliana. Proc Natl Acad Sci U S A 1996, 93, 10519–10523. [Google Scholar] [CrossRef]
- Okumura, S.; Mitsukawa, N.; Shirano, Y.; Shibata, D. Phosphate transporter gene family of Arabidopsis thaliana. DNA Res 1998, 5, 261–269. [Google Scholar] [CrossRef]
- Raghothama, K.G. Phosphate transport and signaling. Curr Opin Plant Biol 2000, 3, 182–187. [Google Scholar] [CrossRef]
- Nussaume, L.; Kanno, S.; Javot, H.; Marin, E.; Pochon, N.; Ayadi, A.; Nakanishi, T.M.; Thibaud, M.C. Phosphate Import in Plants: Focus on the PHT1 Transporters. Front Plant Sci 2011, 2, 83. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C.; Tran, H.T. Metabolic adaptations of phosphate-starved plants. Plant Physiol 2011, 156, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Bayle, V.; Arrighi, J.F.; Creff, A.; Nespoulous, C.; Vialaret, J.; Rossignol, M.; Gonzalez, E.; Paz-Ares, J.; Nussaume, L. Arabidopsis thaliana high-affinity phosphate transporters exhibit multiple levels of posttranslational regulation. Plant Cell 2011, 23, 1523–1535. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Y.; Wang, F.; Yang, J.; Gao, M.; Li, C.; Liu, Y.; Liu, Y.; Yamaji, N.; Ma, J.F.; et al. The rice CK2 kinase regulates trafficking of phosphate transporters in response to phosphate levels. Plant Cell 2015, 27, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V.; Linhares, F.; Solano, R.; Martin, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Dev 2001, 15, 2122–2133. [Google Scholar] [CrossRef]
- Hamburger, D.; Rezzonico, E.; Petetot, J.M.C.; Somerville, C.; Poirier, Y. Identification and characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem. Plant Cell 2002, 14, 889–902. [Google Scholar] [CrossRef]
- Rouached, H.; Arpat, A.B.; Poirier, Y. Regulation of phosphate starvation responses in plants: signaling players and cross-talks. Mol Plant 2010, 3, 288–299. [Google Scholar] [CrossRef]
- Liu, T.Y.; Chang, C.Y.; Chiou, T.J. The long-distance signaling of mineral macronutrients. Curr Opin Plant Biol 2009, 12, 312–319. [Google Scholar] [CrossRef]
- Ham, B.K.; Lucas, W.J. Phloem-Mobile RNAs as Systemic Signaling Agents. Annu Rev Plant Biol 2017, 68, 173–195. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Zheng, Y.; Ham, B.K.; Zhang, S.P.; Fei, Z.J.; Lucas, W.J. Plant lncRNAs are enriched in and move systemically through the phloem in response to phosphate deficiency. J Integr Plant Biol 2019, 61, 492–508. [Google Scholar] [CrossRef]
- Yan, Y.; Ham, B.K. The Mobile Small RNAs: Important Messengers for Long-Distance Communication in Plants. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Perez-Torres, C.A.; Lopez-Bucio, J.; Cruz-Ramirez, A.; Ibarra-Laclette, E.; Dharmasiri, S.; Estelle, M.; Herrera-Estrella, L. Phosphate availability alters lateral root development in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor. Plant Cell 2008, 20, 3258–3272. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.L.; Ma, G.J.; Zhang, M.L.; Xiong, H.; Wu, H.; Zhao, C.Z.; Liu, C.S.; Jia, H.X.; Chen, L.; Kjorven, J.O.; et al. The ARF7 and ARF19 Transcription Factors Positively Regulate PHOSPHATE STARVATION RESPONSE1 in Arabidopsis Roots. Plant Physiol 2018, 178, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Kranz HD, Denekamp M, Greco R, Jin H, Leyva A, Meissner RC, Petroni K, Urzainqui A, Bevan M, Martin C, Smeekens S, Tonelli C, Paz-Ares J, Weisshaar B. Towards functional characterisation of the members of the R2R3-MYB gene family from Arabidopsis thaliana. Plant J 1998, 16, 263–276. [CrossRef]
- Zhao, Y.; Xing, L.; Wang, X.; Hou, Y.J.; Gao, J.; Wang, P.; Duan, C.G.; Zhu, X.; Zhu, J.K. The ABA receptor PYL8 promotes lateral root growth by enhancing MYB77-dependent transcription of auxin-responsive genes. Sci Signal 2014, 7, ra53. [Google Scholar] [CrossRef]
- Wan, J.; Wang, R.; Zhang, P.; Sun, L.; Ju, Q.; Huang, H.; Lu, S.; Tran, L.S.; Xu, J. MYB70 modulates seed germination and root system development in Arabidopsis. iScience 2021, 24, 103228. [Google Scholar] [CrossRef]
- Petricka, J.J.; Winter, C.M.; Benfey, P.N. Control of Arabidopsis root development. Annu Rev Plant Biol 2012, 63, 563–590. [Google Scholar] [CrossRef]
- Belin, C.; Megies, C.; Hauserova, E.; Lopez-Molina, L. Abscisic Acid Represses Growth of the Arabidopsis Embryonic Axis after Germination by Enhancing Auxin Signaling. Plant Cell 2009, 21, 2253–2268. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.T.; Wang, L.F.; Guo, J.X.; Lu, K.K.; Song, R.F.; Zuo, J.X.; Chen, H.H.; Liu, W.C. Abscisic acid facilitates phosphate acquisition through the transcription factor ABA INSENSITIVE5 in Arabidopsis. Plant J 2022, 111, 269–281. [Google Scholar] [CrossRef]
- Nilsson, L.; Muller, R.; Nielsen, T.H. Increased expression of the MYB-related transcription factor, PHR1, leads to enhanced phosphate uptake in Arabidopsis thaliana. Plant Cell Envir 2007, 30, 1499–1512. [Google Scholar] [CrossRef]
- Devaiah, B.N.; Madhuvanthi, R.; Karthikeyan, A.S.; Raghothama, K.G. Phosphate starvation responses and gibberellic acid biosynthesis are regulated by the MYB62 transcription factor in Arabidopsis. Mol Plant 2009, 2, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.Y.; Wang, Y.Y.; Yang, A.; Zhang, W.H. OsMYB2P-1, an R2R3 MYB Transcription Factor, Is Involved in the Regulation of Phosphate-Starvation Responses and Root Architecture in Rice. Plant Physiol 2012, 159, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Guo, M.; Xu, L.; Wang, X.; Zhao, H.; Wang, J.; Yi, K. An SPX-RLI1 Module Regulates Leaf Inclination in Response to Phosphate Availability in Rice. Plant Cell 2018, 30, 853–870. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, Y.; Jia, X.; Wang, X.; Zhang, Y.; Liu, J.; Yang, Q.; Ruan, W.; Yi, K. Alternative splicing of REGULATOR OF LEAF INCLINATION 1 modulates phosphate starvation signaling and growth in plants. Plant Cell 2022, 34, 3319–3338. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, L.C.; Lin, S.I.; Shih, A.C.; Chen, J.W.; Lin, W.Y.; Tseng, C.Y.; Li, W.H.; Chiou, T.J. Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing. Plant Physiol 2009, 151, 2120–2132. [Google Scholar] [CrossRef]
- Buhtz, A.; Pieritz, J.; Springer, F.; Kehr, J. Phloem small RNAs, nutrient stress responses, and systemic mobility. BMC Plant Biol 2010, 10, 64. [Google Scholar] [CrossRef] [PubMed]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.; Yang, L.; Minambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat Plants 2015, 1, 15025. [Google Scholar] [CrossRef]
- Hao, L.; Zhang, Y.; Wang, S.; Zhang, W.; Wang, S.; Xu, C.; Yu, Y.; Li, T.; Jiang, F.; Li, W. A constitutive and drought-responsive mRNA undergoes long-distance transport in pear (Pyrus betulaefolia) phloem. Plant Sci 2020, 293, 110419. [Google Scholar] [CrossRef]
- Xia, C.; Huang, J.; Lan, H.; Zhang, C. Long-Distance Movement of Mineral Deficiency-Responsive mRNAs in Nicotiana Benthamiana/Tomato Heterografts. Plants 2020, 9. [Google Scholar] [CrossRef]
- Kitagawa, M.; Wu, P.; Balkunde, R.; Cunniff, P.; Jackson, D. An RNA exosome subunit mediates cell-to-cell trafficking of a homeobox mRNA via plasmodesmata. Science 2022, 375, 177–182. [Google Scholar] [CrossRef]
- Kehr, J.; Morris, R.J.; Kragler, F. Long-Distance Transported RNAs: From Identity to Function. Annu Rev Plant Biol 2022, 73, 457–474. [Google Scholar] [CrossRef]
- Haywood, V.; Yu, T.S.; Huang, N.C.; Lucas, W.J. Phloem long-distance trafficking of GIBBERELLIC ACID-INSENSITIVE RNA regulates leaf development. Plant J 2005, 42, 49–68. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.C.; Yu, T.S. The sequences of Arabidopsis GA-INSENSITIVE RNA constitute the motifs that are necessary and sufficient for RNA long-distance trafficking. Plant J 2009, 59, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.K.; Chatterjee, M.; Yu, Y.; Suh, S.G.; Miller, W.A.; Hannapel, D.J. Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell 2006, 18, 3443–3457. [Google Scholar] [CrossRef] [PubMed]
- Hannapel, D.J.; Banerjee, A.K. Multiple Mobile mRNA Signals Regulate Tuber Development in Potato. Plants 2017, 6. [Google Scholar] [CrossRef]
- Notaguchi, M.; Wolf, S.; Lucas, W.J. Phloem-mobile Aux/IAA transcripts target to the root tip and modify root architecture. J Integr Plant Biol 2012, 54, 760–772. [Google Scholar] [CrossRef]
- Toscano-Morales, R.; Xoconostle-Cazares, B.; Martinez-Navarro, A.C.; Ruiz-Medrano, R. Long distance movement of an Arabidopsis Translationally Controlled Tumor Protein (AtTCTP2) mRNA and protein in tobacco. Front Plant Sci 2014, 5, 705. [Google Scholar] [CrossRef]
- Branco, R.; Masle, J. Systemic signalling through translationally controlled tumour protein controls lateral root formation in Arabidopsis. J Exp Bot 2019, 70, 3927–3940. [Google Scholar] [CrossRef]
- Ham, B.K.; Brandom, J.L.; Xoconostle-Cazares, B.; Ringgold, V.; Lough, T.J.; Lucas, W.J. A polypyrimidine tract binding protein, pumpkin RBP50, forms the basis of a phloem-mobile ribonucleoprotein complex. Plant Cell 2009, 21, 197–215. [Google Scholar] [CrossRef]
- Zhang, W.N.; Thieme, C.J.; Kollwig, G.; Apelt, F.; Yang, L.; Winter, N.; Andresen, N.; Walther, D.; Kragler, F. tRNA-Related Sequences Trigger Systemic mRNA Transport in Plants. Plant Cell 2016, 28, 1237–1249. [Google Scholar] [CrossRef]
- Yang, L.; Perrera, V.; Saplaoura, E.; Apelt, F.; Bahin, M.; Kramdi, A.; Olas, J.; Mueller-Roeber, B.; Sokolowska, E.; Zhang, W.; et al. m(5)C Methylation Guides Systemic Transport of Messenger RNA over Graft Junctions in Plants. Curr Biol 2019, 29, 2465–2476. [Google Scholar] [CrossRef] [PubMed]
- Guelette, B.S.; Benning, U.F.; Hoffmann-Benning, S. Identification of lipids and lipid-binding proteins in phloem exudates from Arabidopsis thaliana. J Exp Bot 2012, 63, 3603–3616. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.Y.; Ham, B.K.; El-Shabrawi, H.M.; Alexander, D.; Zhang, D.B.; Ryals, J.; Lucas, W.J. Proteomics and metabolomics analyses reveal the cucurbit sieve tube system as a complex metabolic space. Plant J 2016, 87, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Ham, B.K.; Li, G.; Kang, B.H.; Zeng, F.; Lucas, W.J. Overexpression of Arabidopsis plasmodesmata germin-like proteins disrupts root growth and development. Plant Cell 2012, 24, 3630–3648. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Ham, B.K.; Chong, Y.H.; Yeh, S.D.; Lucas, W.J. A Plant SMALL RNA-BINDING PROTEIN 1 Family Mediates Cell-to-Cell Trafficking of RNAi Signals. Mol Plant 2020, 13, 321–335. [Google Scholar] [CrossRef]
- Duan, K.; Yi, K.; Dang, L.; Huang, H.; Wu, W.; Wu, P. Characterization of a sub-family of Arabidopsis genes with the SPX domain reveals their diverse functions in plant tolerance to phosphorus starvation. Plant J 2008, 54, 965–975. [Google Scholar] [CrossRef]
- Yan, J.; Chia, J.C.; Sheng, H.; Jung, H.I.; Zavodna, T.O.; Zhang, L.; Huang, R.; Jiao, C.; Craft, E.J.; Fei, Z.; et al. Arabidopsis Pollen Fertility Requires the Transcription Factors CITF1 and SPL7 That Regulate Copper Delivery to Anthers and Jasmonic Acid Synthesis. Plant Cell 2017, 29, 3012–3029. [Google Scholar] [CrossRef]
- Hoekenga, O.A.; Maron, L.G.; Pineros, M.A.; Cancado, G.M.; Shaff, J.; Kobayashi, Y.; Ryan, P.R.; Dong, B.; Delhaize, E.; Sasaki, T.; et al. AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proc Natl Acad Sci U S A 2006, 103, 9738–9743. [Google Scholar] [CrossRef]
- Gansel, X.; Munos, S.; Tillard, P.; Gojon, A. Differential regulation of the NO3- and NH4+ transporter genes AtNrt2.1 and AtAmt1.1 in Arabidopsis: relation with long-distance and local controls by N status of the plant. Plant J 2001, 26, 143–155. [Google Scholar] [CrossRef]
- Melnyk, C.W. Grafting with Arabidopsis thaliana. Methods Mol Biol 2017, 1497, 9–18. [Google Scholar] [CrossRef]
- Ham, B.K.; Wang, X.; Toscano-Morales, R.; Lin, J.; Lucas, W.J. Plasmodesmal endoplasmic reticulum proteins regulate intercellular trafficking of Cucumber mosaic virus in Arabidopsis. J Exp Bot 2023. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
AtMYB44 is a potential functional homolog of CsMYB44. (A) The clade, which includes the closest MYB homologs of CsMYB44, in the phylogenic tree of the Arabidopsis MYB family, are shown in Figure S1. Numbers above the phylogenetic tree indicate posterior probabilities. (B) P1BSs, which are PHR1 binding motifs, predicted in the promoter regions of AtMYB44, AtMYB70 and AtMYB73, but not AtMYB77. Red bars indicate the P1BS motifs. (C) and (D) Relative expression of AtMYB44, AtMYB70, AtMYB73, and AtMYB77 in the shoot (C) and the root (D), under Pi-sufficient (500 µM) and -starvation (10 µM) conditions. Arabidopsis seedlings were transplanted onto the medium with 500 µM, or 10 µM Pi, 5 days after germination on solid medium with 500 µM Pi. Shoot and root samples of Arabidopsis plants were harvested 7 days after Pi-sufficient (500 µM), or -starvation (10 µM) treatment. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values between Pi-sufficient and -starvation conditions (Student’s t-test, p<0.05).
Figure 1.
AtMYB44 is a potential functional homolog of CsMYB44. (A) The clade, which includes the closest MYB homologs of CsMYB44, in the phylogenic tree of the Arabidopsis MYB family, are shown in Figure S1. Numbers above the phylogenetic tree indicate posterior probabilities. (B) P1BSs, which are PHR1 binding motifs, predicted in the promoter regions of AtMYB44, AtMYB70 and AtMYB73, but not AtMYB77. Red bars indicate the P1BS motifs. (C) and (D) Relative expression of AtMYB44, AtMYB70, AtMYB73, and AtMYB77 in the shoot (C) and the root (D), under Pi-sufficient (500 µM) and -starvation (10 µM) conditions. Arabidopsis seedlings were transplanted onto the medium with 500 µM, or 10 µM Pi, 5 days after germination on solid medium with 500 µM Pi. Shoot and root samples of Arabidopsis plants were harvested 7 days after Pi-sufficient (500 µM), or -starvation (10 µM) treatment. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values between Pi-sufficient and -starvation conditions (Student’s t-test, p<0.05).
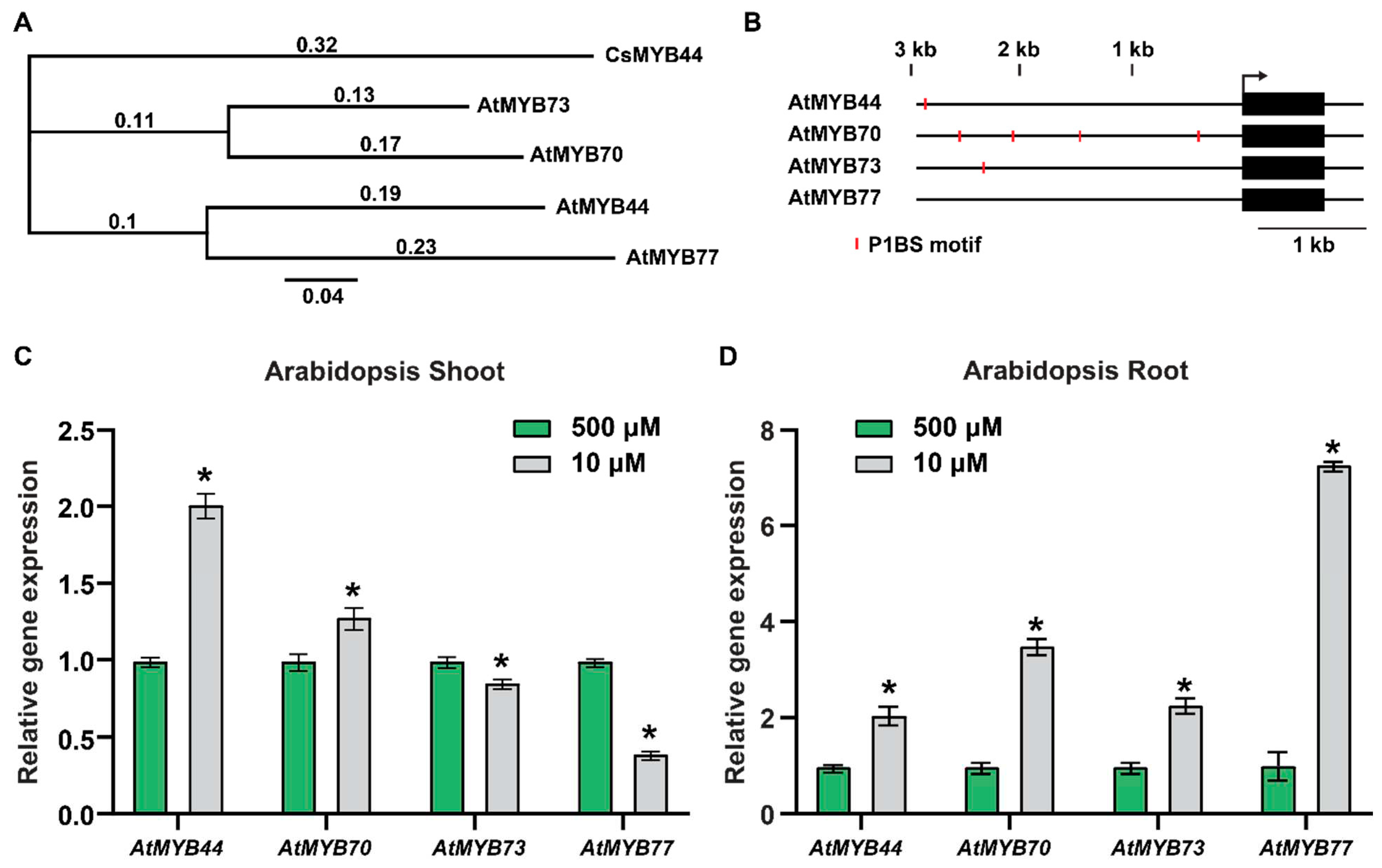
Figure 2.
AtMYB44 is expressed in response to Pi-starvation stress. Histochemical staining in 12-day-old transgenic Arabidopsis plants expressing the GFP-GUS reporter under the control of the AtMYB44 native promoter. Transgenic Arabidopsis seeds were germinated on solid medium with 500 µM Pi (Pi-sufficient medium) for 5 days and then transplanted to medium with 500 µM, or 10 µM Pi. Arabidopsis seedlings were collected 7 days after Pi-sufficient (500 µM), or starvation (10 µM) treatment. (A) GUS expression was barely detectable in Arabidopsis seedlings, grown under Pi-sufficient (500 µM) conditions. (B) Under Pi-starvation (10 µM) conditions, GUS expression was observed in cotyledons (C), developing true leaves (D), lateral roots (E-F), and developing lateral root primordia (G-H). Dotted boxes indicate magnified regions for (C-H). Bars: 5 mm in (A-B), 500 µm in (C-D), and 50 µm in (E-H).
Figure 2.
AtMYB44 is expressed in response to Pi-starvation stress. Histochemical staining in 12-day-old transgenic Arabidopsis plants expressing the GFP-GUS reporter under the control of the AtMYB44 native promoter. Transgenic Arabidopsis seeds were germinated on solid medium with 500 µM Pi (Pi-sufficient medium) for 5 days and then transplanted to medium with 500 µM, or 10 µM Pi. Arabidopsis seedlings were collected 7 days after Pi-sufficient (500 µM), or starvation (10 µM) treatment. (A) GUS expression was barely detectable in Arabidopsis seedlings, grown under Pi-sufficient (500 µM) conditions. (B) Under Pi-starvation (10 µM) conditions, GUS expression was observed in cotyledons (C), developing true leaves (D), lateral roots (E-F), and developing lateral root primordia (G-H). Dotted boxes indicate magnified regions for (C-H). Bars: 5 mm in (A-B), 500 µm in (C-D), and 50 µm in (E-H).
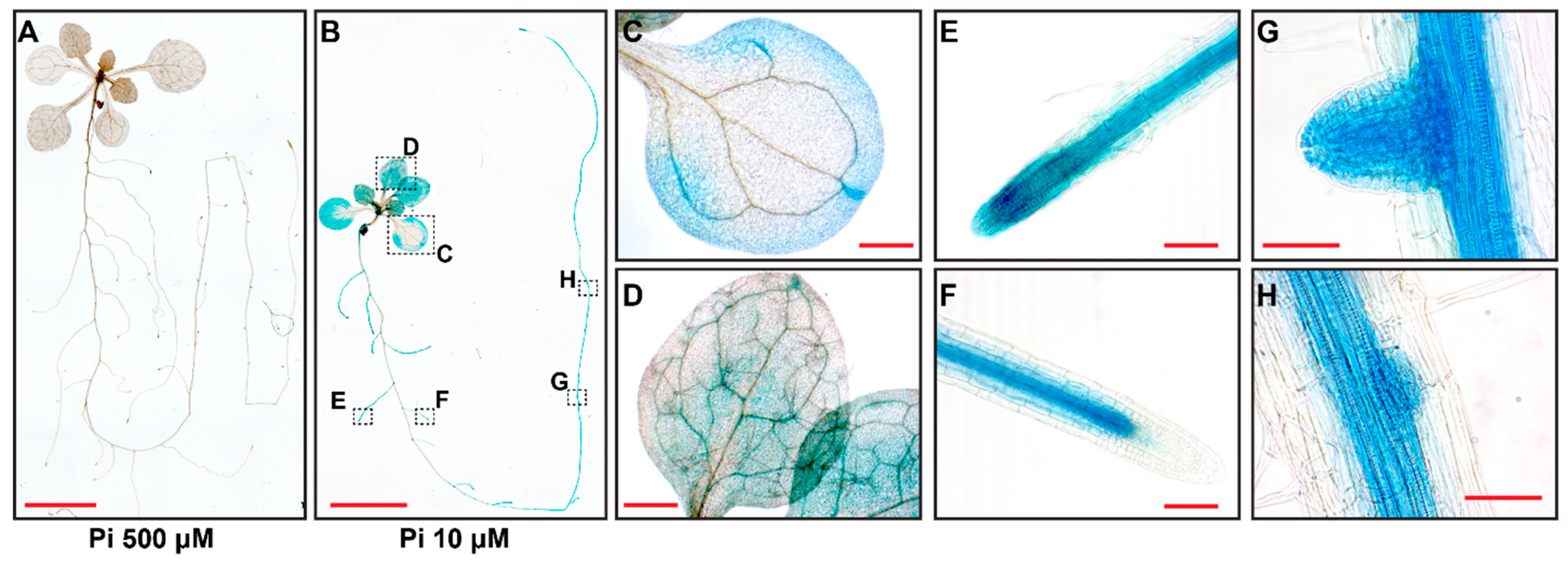
Figure 3.
Transcript levels of AtMYB70, AtMYB73, and AtMYB77 are enhanced in atmyb44 knock-out mutants. Relative expression levels of AtMYB70 (A), AtMYB73 (B), and AtMYB77 (C) in WT (Col-0) and the atmyb44 mutant, under Pi-sufficient (200 µM) and -starvation (10 µM) conditions. Arabidopsis seedlings were transplanted onto medium with 500 µM, or 10 µM Pi, 5 days after germination on solid medium with 500 µM Pi. Arabidopsis seedlings were harvested 7 days after Pi-sufficient (500 µM), or -starvation (10 µM) treatment. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values between WT (Col-0) and atmyb44 mutant (Student’s t-test, p<0.05).
Figure 3.
Transcript levels of AtMYB70, AtMYB73, and AtMYB77 are enhanced in atmyb44 knock-out mutants. Relative expression levels of AtMYB70 (A), AtMYB73 (B), and AtMYB77 (C) in WT (Col-0) and the atmyb44 mutant, under Pi-sufficient (200 µM) and -starvation (10 µM) conditions. Arabidopsis seedlings were transplanted onto medium with 500 µM, or 10 µM Pi, 5 days after germination on solid medium with 500 µM Pi. Arabidopsis seedlings were harvested 7 days after Pi-sufficient (500 µM), or -starvation (10 µM) treatment. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values between WT (Col-0) and atmyb44 mutant (Student’s t-test, p<0.05).
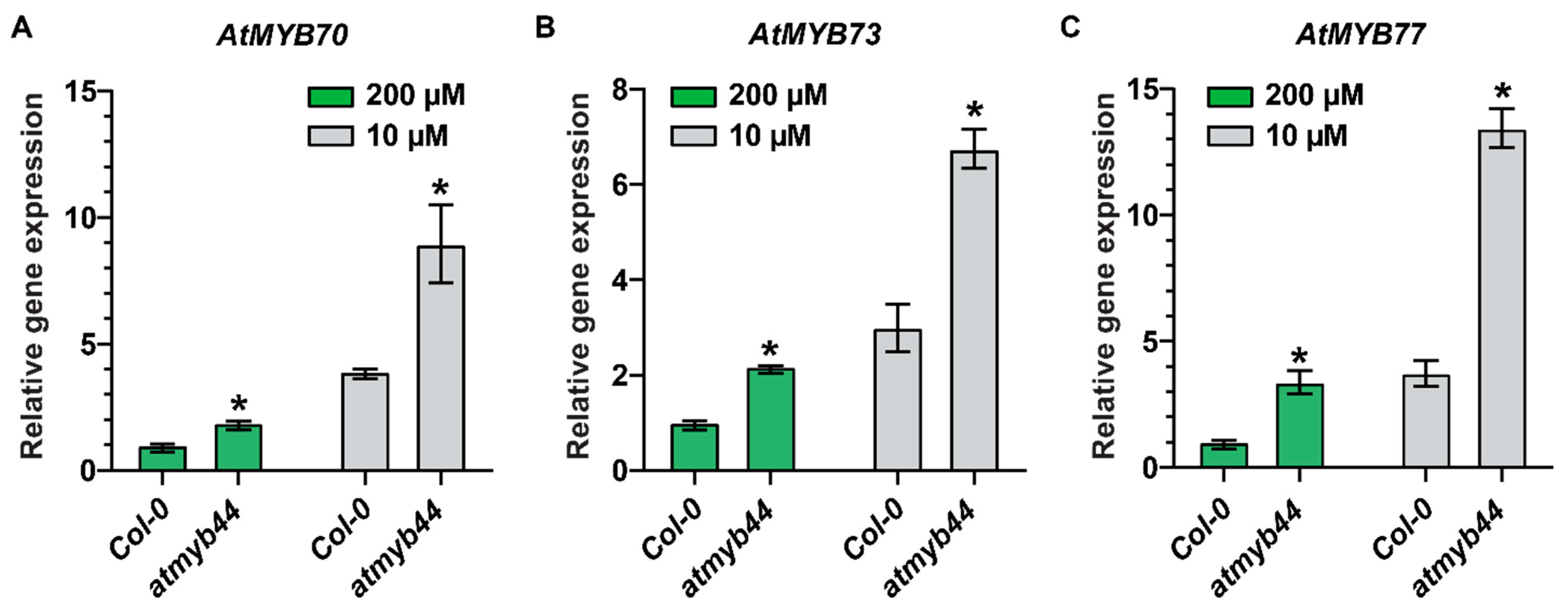
Figure 4.
AtMYB44 functions in plant growth under Pi-starvation conditions. (A) Shoot and (B) root biomass measurements on 4-week-old Col-0, atmyb44, AtMYB44 OX-1 and OX2 lines, grown in a hydroponic culture system with high (200 µM) and low (5 µM) Pi concentrations. Shoot and root biomass was examined as fresh weight per Arabidopsis plant. Soluble Pi concentration in (C) shoot and (D) root of 4-week-old Col-0, atmyb44, AtMYB44 OX-1 and OX2 plants grown in a hydroponic culture system with high (200 µM) and low (5 µM) Pi concentrations. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values, compared with WT (Col-0) (Student’s t-test, p<0.05).
Figure 4.
AtMYB44 functions in plant growth under Pi-starvation conditions. (A) Shoot and (B) root biomass measurements on 4-week-old Col-0, atmyb44, AtMYB44 OX-1 and OX2 lines, grown in a hydroponic culture system with high (200 µM) and low (5 µM) Pi concentrations. Shoot and root biomass was examined as fresh weight per Arabidopsis plant. Soluble Pi concentration in (C) shoot and (D) root of 4-week-old Col-0, atmyb44, AtMYB44 OX-1 and OX2 plants grown in a hydroponic culture system with high (200 µM) and low (5 µM) Pi concentrations. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values, compared with WT (Col-0) (Student’s t-test, p<0.05).
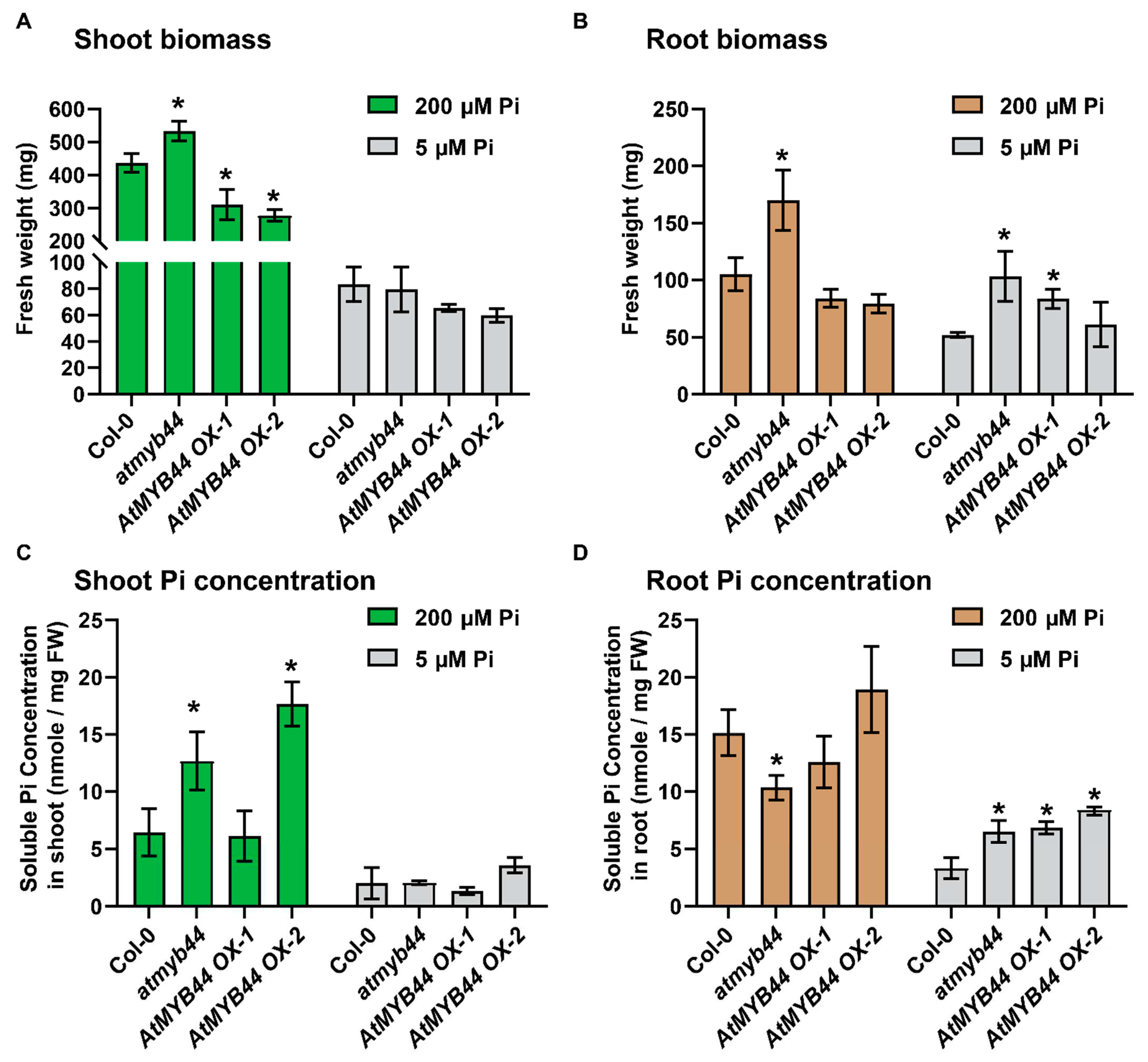
Figure 5.
Expression of PHT1 is enhanced in atmyb44 roots. Relative expression level of PHT1;2 (A) and PHT1;4 (B) in WT (Col-0), atmyb44 and AtMYB44 OX-1 roots, under Pi-sufficient (200 µM) and -starvation (10 µM) conditions. Arabidopsis seedlings were transplanted onto medium with 500 µM, or 10 µM Pi, 5 days after germination on solid medium with 500 µM Pi. Arabidopsis seedlings were harvested 7 days after Pi-sufficient (500 µM), or -starvation (10 µM) treatment. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). The different lowercase letters indicate significant differences in relative gene expression levels, determined by Tukey’s test (p<0.05).
Figure 5.
Expression of PHT1 is enhanced in atmyb44 roots. Relative expression level of PHT1;2 (A) and PHT1;4 (B) in WT (Col-0), atmyb44 and AtMYB44 OX-1 roots, under Pi-sufficient (200 µM) and -starvation (10 µM) conditions. Arabidopsis seedlings were transplanted onto medium with 500 µM, or 10 µM Pi, 5 days after germination on solid medium with 500 µM Pi. Arabidopsis seedlings were harvested 7 days after Pi-sufficient (500 µM), or -starvation (10 µM) treatment. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). The different lowercase letters indicate significant differences in relative gene expression levels, determined by Tukey’s test (p<0.05).
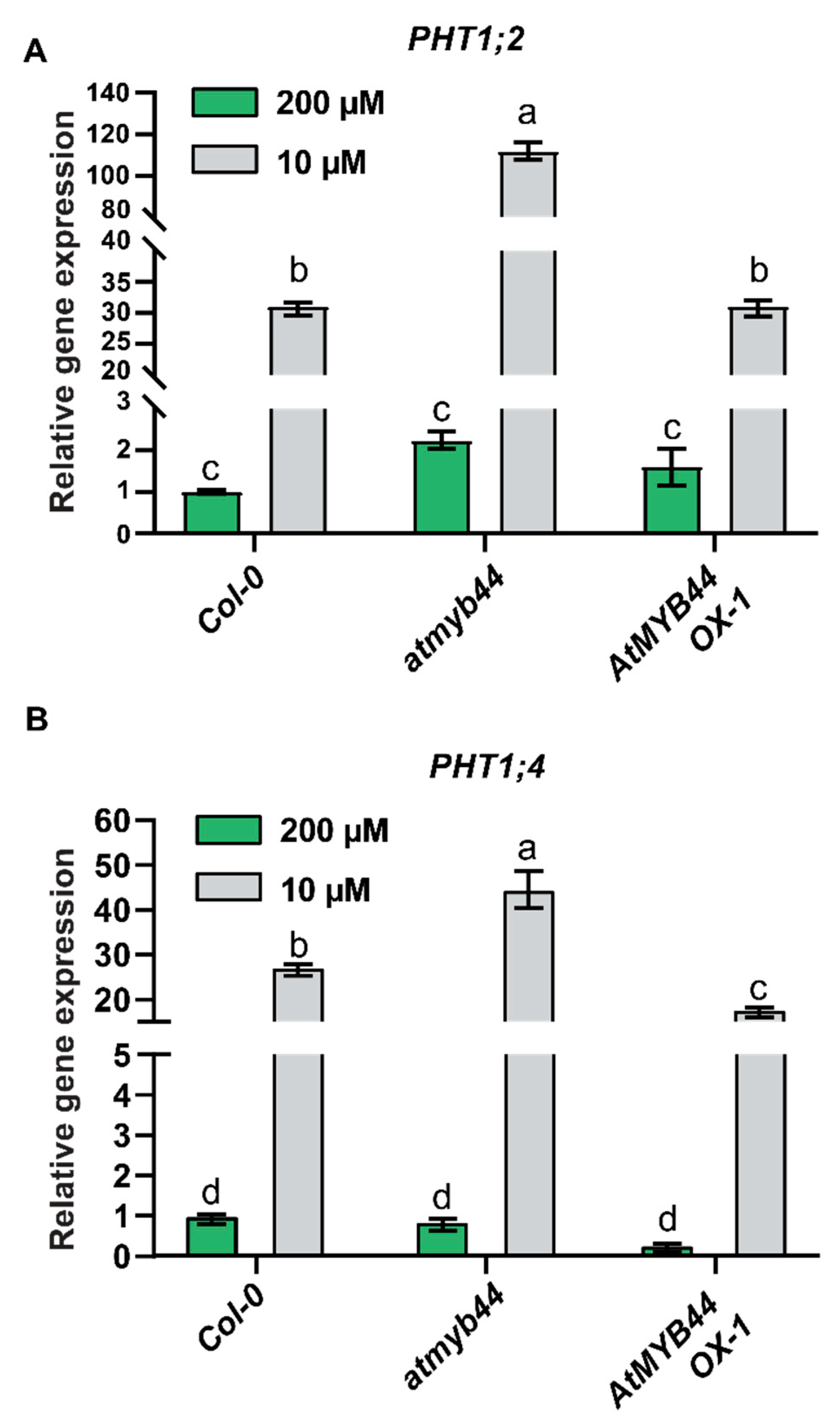
Figure 6.
AtMYB44 is a systemically regulated gene in response to an imposed Pi-starvation stress. Expression of endogenous AtMYB44 in roots of 4-week-old WT (Col-0) and atmyb44 plants grown in a split-root systems. CP200 and CP0 indicate homogenous Pi-sufficient (200 µM) and deficient (0 µM) treatment on roots, respectively. SP200 and SP0 indicate the half of the root system on a plant supplied with Pi-sufficient (200 µM) or deficient (0 µM) treatment, respectively. Arabidopsis roots were harvested 7 days after transferring plants into the split-root systems. (A) AtMYB44 expression is systemically regulated. AtMYB44 expression was examined in WT (Col-0) and atmyb44, using a split-root system. (B) AtPHT1;2 and (C) AtPHT1;4 expression in WT (Col-0) and atmyb44, using a split-root system. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). The different lowercase letters indicate significant differences in relative gene expression levels, determined by Tukey’s test (p<0.05).
Figure 6.
AtMYB44 is a systemically regulated gene in response to an imposed Pi-starvation stress. Expression of endogenous AtMYB44 in roots of 4-week-old WT (Col-0) and atmyb44 plants grown in a split-root systems. CP200 and CP0 indicate homogenous Pi-sufficient (200 µM) and deficient (0 µM) treatment on roots, respectively. SP200 and SP0 indicate the half of the root system on a plant supplied with Pi-sufficient (200 µM) or deficient (0 µM) treatment, respectively. Arabidopsis roots were harvested 7 days after transferring plants into the split-root systems. (A) AtMYB44 expression is systemically regulated. AtMYB44 expression was examined in WT (Col-0) and atmyb44, using a split-root system. (B) AtPHT1;2 and (C) AtPHT1;4 expression in WT (Col-0) and atmyb44, using a split-root system. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. The data are presented as mean ± SD (three technical replicates and three technical repeats). The different lowercase letters indicate significant differences in relative gene expression levels, determined by Tukey’s test (p<0.05).
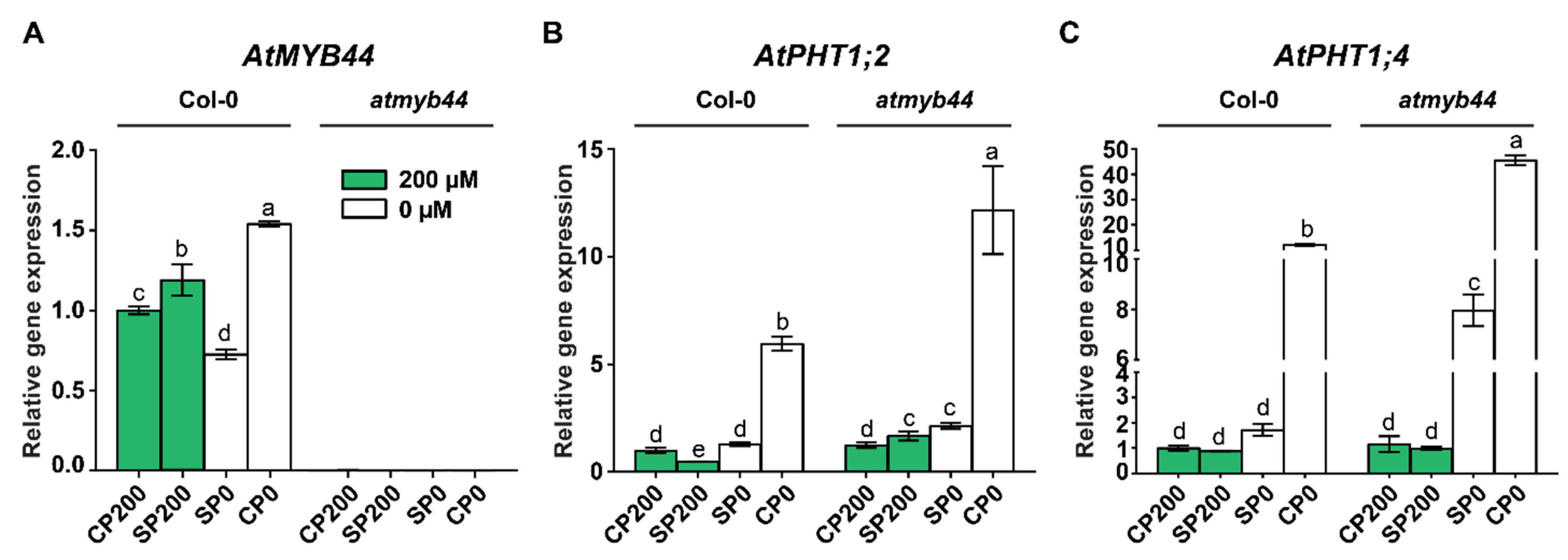
Figure 7.
AtMYB44 plays a role in systemic PHT1;2 and PHT1;4 regulation. (A) Schematic diagram of Arabidopsis micrografting between atmyb44 and AtMYB44 OX-1 seedlings. (B) AtMYB44 is graft-transmissible from a AtMYB44 OX-1 scion to an atmyb44 rootstock. (C) Expression of AtPHT1;2 and AtPHT1;4 is decreased in the heterografted atmyb44 rootstock. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. KO and OX-1 indicate atmyb44 mutant and AtMYB44 OX-1 line, respectively. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values with homografted atmyb44 plants. (Student’s t-test, p<0.05).
Figure 7.
AtMYB44 plays a role in systemic PHT1;2 and PHT1;4 regulation. (A) Schematic diagram of Arabidopsis micrografting between atmyb44 and AtMYB44 OX-1 seedlings. (B) AtMYB44 is graft-transmissible from a AtMYB44 OX-1 scion to an atmyb44 rootstock. (C) Expression of AtPHT1;2 and AtPHT1;4 is decreased in the heterografted atmyb44 rootstock. Arabidopsis Actin was used as an internal control to normalize the qRT-PCR results. KO and OX-1 indicate atmyb44 mutant and AtMYB44 OX-1 line, respectively. The data are presented as mean ± SD (three technical replicates and three technical repeats). Asterisks indicate significantly different values with homografted atmyb44 plants. (Student’s t-test, p<0.05).
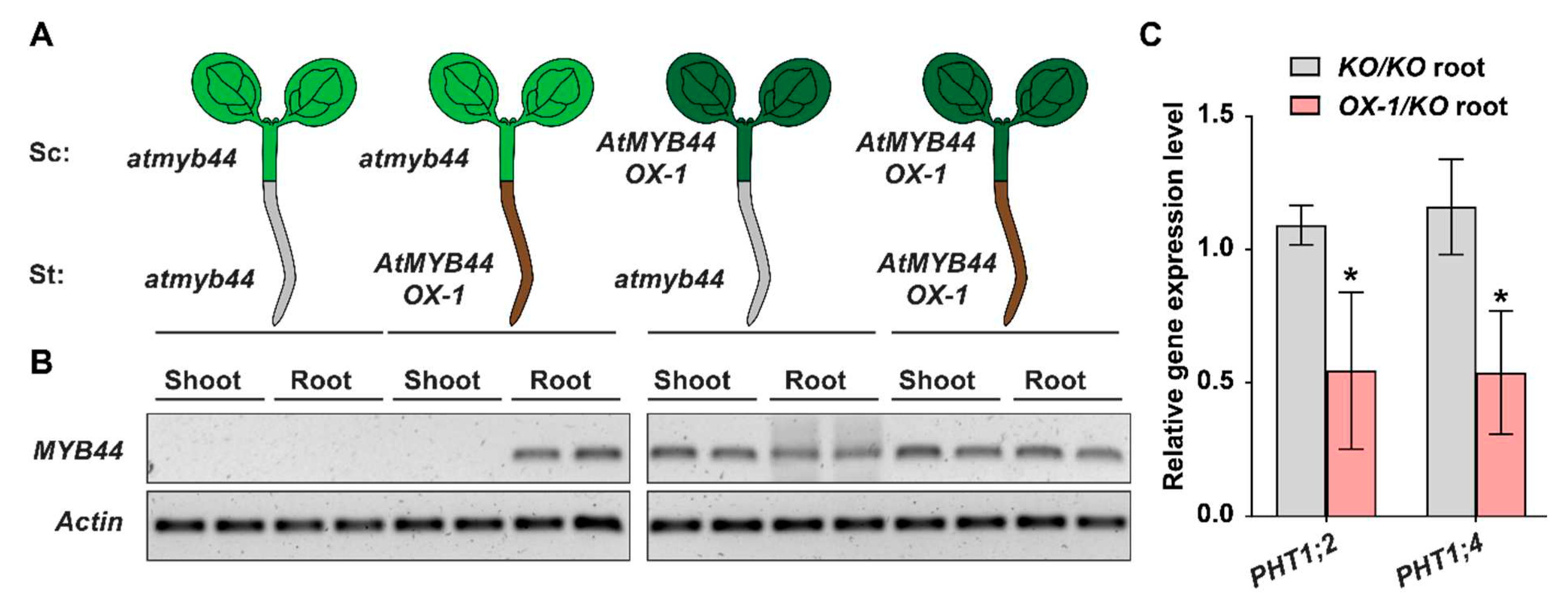
Figure 8.
Schematic model of mobile AtMYB44-mediated PHT1;2 and PHT1;4 regulation in Arabidopsis. (A) The mobile AtMYB44 transcript is long-distantly transported from source leaves to roots and serves as a systemic signal to negatively regulate the expression of Pi transporters, PHT1;2 and PHT1;4, in Arabidopsis roots. In atmyb44 mutant, AtMYB44 mRNA is absent, thereby diminishing inhibition of PHT1;2 and PHT1;4 expression in roots. It results in increases of biomass and soluble Pi concentration in both shoots and roots of atmyb44 mutant, compared to WT. (B) Under Pi-starvation conditions, PHR1 recognizes the promoter region of AtMYB44 and PHT1;2/PHT1;4 in shoots and roots, respectively, to induce those expressions. As AtMYB44 expression is abolished in atmyb44 mutant, PHT1;2 and PHT1;4 expression is more enhanced in atmyb44 roots, compared to WT, due to absence of the negative regulatory factor, mobile AtMYB44 mRNA, in those expression. Red darts and arrows indicate the increased level of designated gene expression and traits. Blue bars indicate the similar level of designated trait between WT and atmyb44 under Pi-starvation conditions.
Figure 8.
Schematic model of mobile AtMYB44-mediated PHT1;2 and PHT1;4 regulation in Arabidopsis. (A) The mobile AtMYB44 transcript is long-distantly transported from source leaves to roots and serves as a systemic signal to negatively regulate the expression of Pi transporters, PHT1;2 and PHT1;4, in Arabidopsis roots. In atmyb44 mutant, AtMYB44 mRNA is absent, thereby diminishing inhibition of PHT1;2 and PHT1;4 expression in roots. It results in increases of biomass and soluble Pi concentration in both shoots and roots of atmyb44 mutant, compared to WT. (B) Under Pi-starvation conditions, PHR1 recognizes the promoter region of AtMYB44 and PHT1;2/PHT1;4 in shoots and roots, respectively, to induce those expressions. As AtMYB44 expression is abolished in atmyb44 mutant, PHT1;2 and PHT1;4 expression is more enhanced in atmyb44 roots, compared to WT, due to absence of the negative regulatory factor, mobile AtMYB44 mRNA, in those expression. Red darts and arrows indicate the increased level of designated gene expression and traits. Blue bars indicate the similar level of designated trait between WT and atmyb44 under Pi-starvation conditions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



