
Submitted:
14 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
Autophagy is a highly conserved catabolic process in eukaryotic cells. Reactive nitrogen species play roles as inductors and signaling molecules of autophagy. A key mechanism of NO-mediated signaling is S-nitrosylation, a posttranslational modification (PTM) of proteins at cysteine resi-dues. In the present work we analyzed the patterns of protein S-nitrosylation during the induc-tion of autophagy in Triticum aestivum roots. The accumulation of S-nitrosylated proteins in the cells during autophagy induced by KNO2 and antimycin A was visualized using monoclonal an-tibodies by Western blot analysis, and proteins were identified using a standard bottom-up pro-teomics approach. Protein S-nitrosylation is a labile and reversible PTM, and therefore SNO group can be lost during experimental procedures. Subsequent bioinformatic analysis using predictive algorithms and protein-ligand docking showed that identified proteins possess hypothetical S-nitrosylation sites. Analyzing protein-protein interaction networks enabled us to discover the targets that can directly interact with autophagic proteins, and those that can interact with them indirectly via key multifunctional regulatory proteins. In this study, we show that S-nitrosylation is a key mechanism of NO-mediated regulation of autophagy in wheat roots. A combination of in silico predictive algorithms with a mass spectrometry analysis provides a targeted approach for the identification of S-nitrosylated proteins.
Keywords:
Triticum aestivum
; autophagy
; nitric oxide
; protein S-nitrosylation
1. Introduction
Autophagy, a highly conserved catabolic process, is of great importance in eukaryotes. In plants, the role of autophagy has been shown in onto- and organogenesis, for example, in the formation of aerenchyma and xylem vessels [1], and in the processes of aging and programmed cell death (PCD) [2]. Autophagy is also considered as a defence reaction for survival of an organism in stressful environments [3]. Activation of autophagic processes under stress is necessary for the efficient breakdown of macromolecules to provide cells with construction blocks and energy substrates [4], as well as the timely removal of oxidized or spent macromolecules and damaged structures [5]. Autophagy is characterized by the appearance of double membrane vesicles (autophagosomes), whose formation is controlled by the sequential activity of multiple autophagy-related (ATG) proteins [6]. The involvement of reactive oxygen species (ROS) and nitrogen species (RNS), including nitric oxide (NO), in the regulation of autophagy has been most extensively studied in animal cells where evidence exists for both the induction [3,7] and the suppression [8,9] of autophagy by RNS. In photosynthetic organisms, little information is available about the effects of NO on autophagic processes. Induction of autophagosome formation by increasing NO levels has been shown in the roots of wheat seedlings [10]. In the unicellular alga Chlamidomonas reinhardii, exposure to high light induced NO emission and cell death via autophagy, which was confirmed by an increase in ATG8 protein content and activation of other ATG genes [11]. Autophagy was suppressed in the presence of the NO acceptor (cPTIO, 2-4-carboxyphenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide). Interestingly, co-treatment of cells with H2O2 and NO donors induced autophagy and resulted in cell death after 24 h, and this effect was also eliminated by cPTIO. These data suggest that the regulatory role of NO in autophagic processes is largely due to the synergistic effect of ROS and RNS [12].
Nitric oxide regulates protein activity through various posttranslational modifications (PTMs), such as S-nitrosylation of sulfhydryl (thiol) groups of proteins and oxidative nitrosylation of iron and other metal-containing proteins. S-nitrosylation of proteins is a mechanism involving the covalent attachment of the -NO group to the cysteine thiol in a protein resulting in the formation of S-nitrosothiol (SNO). Despite the importance of this PTM for cellular biology, in plants our understanding of the molecular mechanisms underlying S-nitrosylation of proteins remains limited. Current estimates suggest that approximately 70% of the proteome is subject to S-nitrosylation, and most of the S-nitrosylation sites are conserved [13]. This particular protein PTM is known to be a key element of the NO-mediated signal transduction mechanism in many processes, including autophagy [14]. Another NO-mediated modification of proteins is tyrosine nitration, which is an irreversible reaction of a nitrating agent with the tyrosine residue of a target protein [15]. Nitration of oxidoreductase enzymes, such as catalase, ascorbate peroxidase, monodehydroascorbate reductase, superoxide dismutase, and nitrate reductase, has been shown to reduce their activity [16]. It seems likely that an increase in protein tyrosine nitration could be considered as a consequence of nitrosative and oxidative stresses.
The first article on the regulation of autophagy mediated by protein S-nitrosylation in animal cells was published only in 2011 [8]. Currently, the study of the effects of protein S-nitrosylation on autophagic processes is a hot topic in medical research [7,17,18]. Targeting the pro-death and pro-survival functions of protein S-nitrosylation in autophagy has become a novel therapeutic strategy for treatments of Parkinson's disease [19], cellular tumorigenesis [20,21], myocardial ischemia [22], and cocaine dependence [23]. One of the targets of S-nitrosylation is the family of cysteine proteases ATG4, which occurs universally in eukaryotic cells. For example, ATG4B undergoes S-nitrosylation at Cys189 and Cys292 when glucose levels increase in human neuronal cells [18]. This modification reduces the efficiency of the proteolytic and delipidative activities of ATG4, resulting in the inhibition of the ability of ATG4B to process ATG8 family precursors and deconjugate ATG8 to PE, and as a consequence in the reduction of autophagic flux.
For plants, only fragmentary information is available about the regulation of autophagy mediated by protein S-nitrosylation. A recent study demonstrated that in Arabidopsis thaliana hypoxic conditions induce S-nitrosylation of the protein AtGSNOR1 (S-nitrosoglutathione reductase 1) at the conserved Cys10 residue [24]. This, in turn, changes the conformational structure of the protein and promotes its interaction with AtATG8 via the ATG8-interacting motif (AIM). These protein-protein interactions (PPIs) result in the degradation of AtGSNOR1 via selective autophagy. These data provide direct evidence of a functional link between protein S-nitrosylation and autophagy in plants during stress, particularly in response to hypoxia. Unfortunately, proteins that undergo S-nitrosylation have not been identified in plants exposed to conditions that induce autophagy. Therefore, the aim of the present study was to identify potential protein targets of S-nitrosylation in the cells of wheat roots, in which autophagy was induced by the application of antimycin A, a mitochondrial inhibitor, and KNO2, a donor of NO. We have previously showed that antimycin A [25] and KNO2 [26] efficiently induce autophagy in wheat roots. To identify S-nitrosylated proteins, we applied a complex approach including polyacrylamide gel electrophoresis (PAGE), immunoblotting with monoclonal antibodies followed by protein identification in visualized electrophoretic zones (bands) using the bottom-up proteomics approach, i.e. tryptic digestion of individual bands with subsequent nanoflow reversed phase high-performance liquid chromatography - (tandem) mass spectrometry (nanoRP-HPLC-MS and MS/MS). Furthermore, this classical bottom-up proteomics approach was combined with algorithms for prediction of hypothetical S-nitrosylation sites and modelling of the protein-ligand molecular interactions. In silico analysis of PPIs clearly demonstrated that S-nitrosylation of proteins, as a result of NO signaling, provides an important link between the key metabolic pathways and autophagy.
2. Materials and Methods
2.1. Plant material
Seeds of bread wheat (Triticum aestivum L.) variety Kazanskaya Yubileinaya were purchased from the Tatar Research Institute of Agriculture, FRC Kazan Scientific Center, Russian Academy of Sciences, Kazan, Russia. Seedlings were grown hydroponically in distilled water for 4 d at 22°C in a growth chamber with a 12 h-light/dark photoperiod at a light intensity of 150 μmol photons m-² s-1. Intact roots were incubated for 3 h in distilled water (control); 1 μM antimycin A (AA); 1 mM KNO2; and for 15 min in 5 mM S-nitrosoglutathione (GSNO).
2.2. Extraction and immunodetection of S-nitrosylated wheat proteins
Water-soluble proteins were isolated from the root tips by fixing them in liquid nitrogen and homogenizing in a medium containing 50 mM HEPES, pH 7.5, 1 mM EDTA, and a protease inhibitor cocktail (Sigma, Germany). Homogenate was then centrifuged at 12,000 g, 4oC for 10 min. Protein electrophoresis was performed in a 4% polyacrylamide concentrating gel at 40 V and in a 10% separating gel at 120 V in a Mini-PROTEAN Tetra Cell chamber (Bio-Rad, USA) [27]. The content of water-soluble proteins was determined using Qubit™ (Invitrogen, USA), according to the manufacturer's protocol. In all samples, 30 µg of protein were applied to the lane. Proteins separated electrophoretically were transferred onto polyvinylidene difluoride (PVDF) membranes by semi-dry blotting using SDS-PAGE Transfer Buffer at 150 mA for 1 h. Membranes were incubated with mouse monoclonal antibodies HY8E12 (ab268288, Abcam, USA), which specifically recognize bound forms of S-nitroso-L-cysteine, at a dilution of 1:1000. To visualize S-nitrosylated proteins, blots were incubated with secondary anti-mouse IgG antibodies conjugated with horseradish peroxidase (ab205719, Abcam, USA), then with chemiluminescent substrate (0.1 M Tris-HCl, luminol, p-coumaric acid, 30% H2O2), and finally scanned using an imaging system (ChemiDoc MP, BioRad, USA). The intensity of chemiluminescence was analyzed using ImageLab software (BioRad, USA).
2.3. In-gel trypsin digestion
To identify S-nitrosylated proteins, electrophoretically separated protein bands, which corresponded to those visualized by immune blots, were subjected to in-gel trypsin digestion according to Bassal et al. [28] with minor modifications. A protein band from the gel was cut into small 1-2 mm sized pieces and after rinsing with LC-Grade H2O, sample was destained with 30% (v/v) acetonitrile (ACN) in 100 mmol/L NH4HCO3 and dried under reduced pressure in the vacuum concentrator (CentriVap Vacuum Concentrator, Labconco, USA) at 4°C. For reduction of disulfides, 10 mM dithiothreitol (DTT) in 100 mM NH4HCO3 was supplemented and the samples were incubated for 15 min at 22°C under continuous shaking in dark. Afterwards, the alkylation solution (54 mM iodoacetamide in 100 mM NH4HCO3) was added, and the samples were incubated under the same conditions. Further, the liquid phase was discarded, 100 µL of destaining solution (30% (v/v) ACN in 100 mmol/L NH4HCO3) were added, and gel pieces were shaken for 10 min (450 rpm, 22°C). This step was repeated twice. The stock solution of trypsin (0.5 µg/µL) was freshly prepared in 50 mmol/L NH4HCO3 buffer, added to the solutions (with an enzyme-to protein-ratio w/w of 1:20), and then incubated using continuous shaking (450 rpm, 37°C, 4 h). After that, an additional portion of trypsin stock solution (with an enzyme-to protein-ratio w/w of 1:50) was added and the samples were left shaken overnight (450 rpm, 37°C). Next day 70 µL of the extraction solution (4 % v/v trifluoroacetic acid in 30 % v/v ACN) were added to the digest mixture and shaken at 22°C for 40 min, and then the supernatants collected. This step was repeated twice, and then samples were dried under the reduced pressure in the vacuum concentrator at 4°C.
2.4. Solid-phase extraction
2.5. Nano LC-MS/MS
Protein hydrolysates were loaded onto an Acclaim PepMap 5 mm Trap Cartridge (Thermo Fisher Scientific) and separated on a Bruker FORTY separation column (C18 ReproSil AQ, 40 cm 75 m, 1.9 m, 120 A; Bruker Daltonics, Bremen, Germany) using a nanoElute UHPLC chromatography system (Bruker Daltonics) coupled on-line to a TimsToF Pro quadrupole time-of-flight mass spectrometer (QqTOF-MS) via a CaptiveSpray ion source (Bruker Daltonics). Details of the chromatographic separation method are summarized in Supplementary Information S1 (Table S1-3). The UHPLC-QqTOF-MS/MS analysis relied on data-dependent acquisition experiments performed in the positive ion mode, comprising a survey TOF-MS scans and dependent MS/MS scans for the most abundant signals with the charge states ranging from 2 to 5 at the acquisition rate of 8 to 32 Hz in the cycle time of 3 s. The mass spectrometer settings are summarized in Table S2.
2.6. Data analysis
Analysis of the LC-MS and MS/MS data was accomplished with the Peaks Xpro software (Bioinformatics Solutions Inc., Canada) (For the detailed settings see Supplementary information 1, Table S3). Identification of peptide sequences and annotation of the proteins relied on the common sequence database of the plants representing Triticum aestivum and Arabidopsis thaliana species (UniProt database, downloaded on the 28.07.202). Possible sample contaminations were excluded using the CRAP database of common protein contaminations (https://www.thegpm.org/crap/). The following amino acid modifications were included in the search: carbamidomethylation of cysteines (mass increment = 57.0215); variable amino acid modifications included in the search: methionine oxidation (15.9949), N-terminal acetylation (42.0106), asparagine and glutamine deamidation (0.9840), S-nitrosylation of cysteines (28.9902). The MS1 and MS2 tolerance was 10 ppm and 0.05 Da, respectively. FDR (false discovery rate) correction was employed at the level of 0.02. The proteins were identified with at least two unique peptides. Functional annotation and protein localization relied on the UNIPROT database and employed the UNIPROT categories.
2.7. Bioinformatic analysis of the availability of S-nitrosylation sites of wheat proteins
To predict the presence of S-nitrosylation sites, amino acid sequences were selected from the UNIPROT database and analyzed using the programs iSNOPseAAC (app.aporc.org/iSNO-PseAAC/), iSNO-AAPair (app.aporc.org/iSNO-AAPair/), GPS-SNO 1.0 (high specificity - high threshold) and pLMSNOSite (github.com/KCLabMTU/pLMSNOSite). The programs have different databases of S-nitrosylated proteins and algorithms for internal evaluation of prediction efficiency. Based on the differences in the algorithms and internal bases of these programs, sites that were predicted by at least three programs were identified.
2.8. Protein-ligand docking and protein-protein interactions
Protein models were constructed in AlphaFold (alphafold.ebi.ac.uk/), and the quality of the constructed models assessed using Procheck, GSNO (ZINC3872731) was selected as a ligand from the ZINC database (zinc.docking.org/). Molecular interactions were predicted using the SwissDock web service (www.swissdock.ch/docking), and then the data were analyzed using Chimera 1.14.
To construct PPI networks, STRING v11.5 was applied [31]. The networks included a list of identified proteins, ATG proteins, and predicted functional partners, i.e. proteins that were proposed by the STRING program. We used 13 neighborhood interactors that interact with the introduced proteins. All interactions between proteins were constructed based on previous knowledge in curated databases at medium level of confidence (sources: textmining, experiments, databases, co-expression, neighborhood, gene fusion; score ≥ 0.4).
2.9. Statistical analysis
Protein immunodetection experiments were performed in six biological replicates. The experiment on the identification of S-nitrosylated proteins was performed using two technical replicates.
3. Results
3.1. Extraction and visualization of S-nitrosylated proteins during induction of autophagy
Experimental detection of S-nitrosylated proteins was performed in the water-soluble protein fractions of wheat roots after 3 h incubation in solutions of 1 µM AA and 1 mM KNO2. Roots incubated in 5 mM GSNO were used as a positive control. Determination of the protein concentrations in the obtained isolates revealed extraction yields in the range of 0.201–0.242 mg/μL. In all samples, 30 µg of protein were applied to the lane (Figure 1).
After separation by denaturing PAGE of water-soluble proteins (Figure 1a) and Western blotting on PVDF membranes (Figure 1b), the electrophoretic zones containing S-nitrosylated proteins were visualized with monoclonal primary anti-Nitroso-L-cysteine antibodies and secondary peroxidase-conjugated antibodies. It was found that in all treatments, polypeptides (PP) with molecular mass less than 75 kDa are subjected to S-nitrosylation. After incubation of the roots in KNO2 and GSNO solutions, the luminescence intensity of multiple PPs with molecular mass less than 50 kDa markedly increased. In particular, PPs with molecular weights less than 25 kDa exhibited the most intensive chemiluminescence. Indeed, the luminescence intensity of PPs with a molecular mass less than 50 and 25 kDa in KNO2 and GSNO samples was 1.5-1.7 times higher than that in the control (Figure 1b). Next, the blots were matched to the gels stained with colloidal Coomassie Brilliant blue G-250, and gel bands corresponding to the electrophoretic zones matched to the areas stained with antibodies, were cut and subjected to in-gel trypsin digestion for protein identification (Figure S1).
3.2. Protein identification and search for S-nitrosylation sites
A total of 298 proteins were identified by the nanoRP-HPLC-MS/MS analysis of tryptic protein hydrolysates (Table S4). More than the half of these proteins (177 species) were present in all experimental groups. All proteins identified in control were also found in KNO2, Antimycin A and GSNO groups.
Analysis of the group-specific changes in the root proteome revealed 39 unique proteins in the roots treated with antimycin A, 51 proteins in the roots treated with KNO2, and 57 proteins in the roots treated with GSNO (Figure 2a).
Functional annotation of the total protein number identified identified in all groups with the UNIPROT database and its classification categories showed that the most of the proteins are responsible for protein biosynthesis and metabolism, transport, and carbohydrate metabolism (Figure S2, Table S5). In particular, identified the polypeptides included those involved in protein metabolism (40S and 60S ribosome subunits, ATP synthase, 26S proteasome subunits), protein biosynthesis initiation and elongation factors, heat shock protein (70 kDa), and core histones. In all samples, the proteins were predominantly localized in the cytoplasm, nucleus, mitochondria, and chloroplasts (Figure 2b, Table S6).
For the convenience of interpretation, the results of functional annotation we separately considered the proteins representing the major and the minor functional categories (i.e. those with the most and the least represented number of proteins) (See Figure 3a and b, respectively). Thus, the proteins involved in carbohydrate metabolism (aconitate hydratase, triosephosphate isomerase), stress adaptation (2-cys peroxiredoxin, L-ascorbate peroxidase), transport (ADP,ATP carrier protein 1 and V-type proton ATPase subunit) were also identified among the major functional category group (Figure 3a). The proteins representing minor functional categories included those involved in such processes as the biosynthesis of flavonoids (tricetin 3',4',5'-O-trimethyltransferase), polyamines (N-carbamoylputrescine amidase), oxylipins (putative 12-oxophytodienoate reductase), and pyrimidines (uridine 5'-monophosphate synthase) (Figure 3b). Interestingly, the proteins related to DNA replication and repair were the most abundant in the control group. To make the FDR correction less strict, a search for S-nitrosylated peptides (i.e. those containing the thionitroso group, SNO) relied on the search of the tandem-mass spectrometric data against the fasta file containing the list of only identified proteins. This analysis showed that the SNO group was lost during the sample preparation procedures. As S-nitrosylation is well-known to be a highly-labile PTM, this scenario seems to be most likely. Indeed, the reaction of thiol nitrosylation is very sensitive to experimental conditions, especially to the presence of reducing agents (e.g. DTT) used in sample preparation, light, etc. Nevertheless, the possibility of S-nitrosylation of identified proteins in planta was appreciated by predicting S-nitrosylation sites.
3.3. Prediction of S-nitrosylation sites, localization, and functional annotation of S-nitrosylated proteins
Further analysis of protein sequences using the iSNOPseAAC, iSNO-AAPair, GPS-SNO 1.0 and pLMSNOSite programs revealed 78 proteins (predicted by three programs, Table S7) and 17 proteins (predicted by all four programs, Table 1), which possess hypothetical S-nitrosylation sites.
Most of the identified S-nitrosylated proteins predicted by the three and four programs were localized in the cytoplasm and mitochondria (Figure 4). Interestingly, S-nitrosylation sites in the proteins involved in protein biosynthesis, stress response, and carbohydrate metabolism were predicted by three programs, while the modification sites in the proteins involved in cytoskeleton function, protein folding, and general stress response were predicted by four programs.
3.4. Protein-ligand docking and protein-protein interactions
Protein-ligand docking was performed to computer simulate possible S-nitrosylation of proteins using GSNO, a biological source of NO [32], as a ligand.
Prediction of the molecular interactions of proteins with GSNO using the SwissDock web service and Chimera 1.14 software demonstrated that GSNO interacts with cysteine in triosophosphate isomerase (Figure 5) and mitochondrial ADP/ATP transporter (data not shown) at previously predicted positions of hypothetical S-nitrosylation (Table 1). Interestingly, GSNO interacted with cysteine in spermidine synthase at a position 43 that was not predicted by S-nitrosylation site search programs (Table 1). Protein-ligand modelling suggested that for spermidine synthase this PTM is high energy consuming and therefore may be quite rare in a living system. Interestingly, while heat shock proteins and the 14-3-3 like protein GF14 lacked interactions with GSNO (data not shown), software predicted hypothetical S-nitrosylation sites (Table 1).
To assess possible involvement of the proteins identified by nanoLC-MS/MS and containing prospective S-nitrosylation sites (Section 3.2) in the autophagy, PPI networks were constructed using the STRING online service. For this, we added to the list of identified proteins several autophagic (ATG) proteins involved in four different stages of autophagosome formation: ATG8-conjugation system (ATG8, ATG4, ATG7, ATG3); ATG12-conjugation system (ATG12, ATG7, ATG10, ATG5, ATG16); ATG1 protein kinase complex (ATG1, ATG13, ATG17, ATG29); Vps34 PI3 kinase complex (Vps34, Vps15, ATG14, ATG6; ATG9); and ATG2-ATG18 complex (ATG2, ATG18, ATG9). Moreover, we added to this list two further proteins - target of rapamycin (TOR) kinase and SNF1-related kinase (SnRK1), which are the negative and positive regulators of autophagic processes. To link the proteins identified in this study with the ATG proteins, 13 predicted functional partners proposed by the STRING program were also introduced (Table 2). Network analysis demonstrated that selected proteins have more interactions with each other than it could be expected for a random set of proteins. This enrichment indicates that the proteins are potentially biologically related (p = 1.0e-16). From an expected number of 364 edges, the number of nodes is 56 (avg. local clustering coefficient is 0.775).
Importantly, we found possible PPIs between key ATG proteins and proteins identified by LC-MS/MS identified proteins with predicted S-nitrosylation sites (Figure 6, Table 2, Table S9). The following interactions were found: 2-Cys peroxiredoxin BAS1 (BAS-1) with ATG10 (combined score 0.6) and ATG6 (combined score 0.5); heat shock 70 kDa protein (BIP2) with VPS34 (combined score 0.7), ATG7, ATG6, ATG3, and ATG5 (combined score 0.6); 26S proteasome non-ATPase regulatory subunit 12 (EMB2107) with ATG7 (combined score 0.5); 14-3-3-like protein (GRF4) with TOR (combined score 0.6). It is noteworthy that ATG proteins could also interact with proteins identified by nanoLC-MS/MS and could affect their expression (Table S9).
4. Discussion
S-nitrosylation of proteins is well-known as a key mechanism of NO-mediated signaling in cells. Given that only fragmentary information is available about PTM-mediated regulation of autophagy in plants, it is important to develop an efficient approach to search for S-nitrosylated target proteins. Therefore, in this study we applied a combination of methods including immunoblotting, protein identification using a standard bottom-up proteomics approach, predictive algorithms, and molecular docking to search for target proteins, i.e. for the potential candidates for S-nitrosylation. While only a few ATG proteins are predicted to be capable of S-nitrosylation, this study identified several S-nitrosylated proteins potentially involved in autophagic flux in wheat roots. Using PPI networks, we found that some of these proteins identified by LC-MS/MS can directly interact with ATG proteins, while others interact with ATG proteins indirectly via key multifunctional regulatory proteins.
Immunoblotting with monoclonal anti-nitroso-L-cysteine antibodies revealed the accumulation of S-nitrosylated proteins in the cells of the roots treated with the NO donor KNO2 and the mitochondrial inhibitor antimycin A (Figure 1b). The accumulation of S-nitrosylated proteins in the roots following treatment with KNO2 is probably a result of the increase in intracellular NO levels that occurs during the reduction of nitrite to NO by nitrite:NO reductase at the plasma membrane [33] and the mitochondrial ETC [34]. Previously we showed that treatment of wheat roots with the inhibitor of mitochondrial complex III antimycin A induces autophagy, which was confirmed by the accumulation of autophagosomes in the cells [25] and an increase in ATG gene expression (Minibayeva et al. unpublished). Mitochondrial ETC is known as one of the greatest intracellular sources of NO [34]. Disruption of mitochondrial complexes causes an increase in mitochondrial NO levels, which can intensify the process of S-nitrosylation of proteins.
Functional annotation of the identified here proteins demonstrated that out of 298 proteins the most representative group of proteins are involved in protein biosynthesis and metabolism, transport, and carbohydrate metabolism (Figure S2). Among these proteins, 178, which were predicted to contain S-nitrosylation sites in their sequence, are involved in general universal stress response (Figure 4b). The search for PTMs in the total proteome of the organism leads to a large number of single predictions and a relatively tight correction for multiple comparisons (FDR correction) of the probabilities of each of them. In turn, this leads to the loss of relatively low-confidence peptide spectra, which typically include posttranslationally modified peptides. To avoid these losses of valuable information, such spectra are typically manually inspected. Indeed, as a result of the low contents of modified peptides in the total hydrolysates, the intensity of the corresponding MS signals is low. Moreover, pronounced ion suppression, which is characteristic for electrospray ionization [35], further reduces signal strength. The low signal intensity of MS1 spectra results in a low intensity of fragment spectra and insufficient representation of individual signals of b- and y-fragment series. In turn, this reduces the probability of identifying the corresponding peptides and Xcorr values obtained in the Sequest search. Such peptides with low probability values do not pass FDR correction and are rejected by the search engine as false positives.
A search for the thionitroso (SNO) group in the identified proteins revealed that this group is lost during sample preparation for mass spectrometric analysis. The SNO group is known to be highly labile and sensitive to experimental conditions. Indeed, in the absence of immediate chemical derivatization of the nitroso group (e.g., by the biotin switch method), the reverse reaction - denitrosylation - begins to prevail. It leads to the degradation of the modification and the impossibility of its detection [36]. It seems likely that the main reason for the secondary degradation of the modification is the use of the relatively strong reducing agent DTT during sample preparation for nanoLC-MS/MS analysis [37]. Nevertheless, the presence of S-nitrosylated proteins is confirmed by the interaction of proteins in extracts with monoclonal antibodies for nitroso-L-cysteine (Fig. 1b), and in silico identification of S-nitrosylation sites in proteins identified by LC-MS/MS. Three programs predict seventy-eight proteins with a S-nitrosylation site, while eighteen proteins are predicted by all four programs (Table 1). Interestingly, among the identified S-nitrosylated proteins, PPs localized in the cytoplasm and mitochondria predominate. Identified proteins with predicted SNO group include those involved in protein biosynthesis, responses to stressors, carbohydrate metabolism, cytoskeletal functioning, protein folding, and energy metabolism. For example, modeling of triosophosphate isomerase, a glycolytic enzyme involved in carbohydrate metabolism, confirms possible S-nitrosylation at Cys127 (Figure 4). Preliminary analysis using S-nitrosylation site prediction software shows that isoforms of the autophagic proteins ATG4 and ATG16 have hypothetical S-nitrosylation sites (Table S8). This, in turn, may indicate the possibility of S-nitrosylation of ATG proteins.
Using PPI networks, we found proteins such as 14-3-3 proteins (GRF4), heat shock protein 70 kDa (Hsp70, BIP2), 2-Cys peroxiredoxin (BAS-1), and ADP/ATP transporter (AAC1), can directly interact with ATG proteins, while triosephosphate isomerase (TPI), 60S ribosomal protein (RPL7AB), and 26S proteasome non-ATPase regulatory subunit (EMB2107). These proteins can be divided into three groups: (1) proteins that negatively regulate autophagy (14-3-3 protein, heat shock 70 kDa, ADP/ATP carrier); (2) proteins that can be selectively taken up by autophagosomes (26s proteasome sub., 60s ribosomal protein); and (3) proteins whose expression depends on the expression of ATG proteins (2-Cys peroxiredoxin, triosephosphate isomerase). Negative regulators of autophagy include proteins with diverse effects on autophagy flux. For example, Hsp70 is known to regulate autophagy activity by interacting with p62 protein [37]. However, recent studies showed that the stress response induced Hsp70 can also inhibit autophagy by activating RAC-alpha serine/threonine-protein kinase (Akt), which in turn phosphorylates and stimulates mTOR. An obvious negative regulator of autophagy is the family of 14-3-3 proteins, which are involved in various multiple molecular interactions and implicated in subcellular localization, scaffolding and stability of proteins. The 14-3-3 proteins block the formation of autophagosomes by interacting with phosphorylated ULK1 (ATG1), [38]. Furthermore, in a later stage of autophagosome formation, 14-3-3 proteins can interfere with the activity of hVps34 Beclin-1 (ATG6), thereby inhibiting the autophagy process [39]. Moreover, in plants the 14-3-3-like protein GRF10 and ATG10 are linked through a product of the RNA degradation, namely 2′,3′-cyclic adenosine monophosphate (2′,3′-cAMP) [40]. Accumulation of Br-2′,3′-cAMP/2′,3′-cAMP decreases the expression levels of 14-3-3-like protein but increases the levels of ATG10. Another protein that can stimulate or repress autophagy is the mitochondrial ADP/ATP transporter (AAC), the major transport protein of the inner mitochondrial membrane. This protein exchanges mitochondrial ATP for cytosolic ADP thereby controlling the production of cellular ATP. It was experimentally shown that in yeast AAC proteins are involved in the degradation of mitochondria by facilitating permeabilization of mitochondrial membranes and thus triggering PCD [41].
A second regulatory group includes proteins that are involved in selective autophagy, such as selective degradation of ribosomes via ribophagy. Ribosomal assemblies and the abundance of individual ribosomal proteins can be controlled by both the ubiquitin system and autophagy. Interestingly, in human cell lines, it was shown that an increase in NO levels can decrease translational activity and induce ribosome collision, leading to the ribotoxic stress response [42]. Therefore, a surge of NO may be one of the stimuli for the selective removal of ribosomes. Another NO-sensitive system is the 26S proteasome, a multi-subunit proteolytic machine. It has been shown that in endothelial cells eNOS-derived NO functions as a physiological suppressor of the 26S proteasome in vascular endothelial cells [43]. Notably, nuclear 26S proteasomes can be substrates of autophagy after nitrogen starvation or inactivation. In plants, excess or damaged proteasomes can be degraded in autophagy pathways mediated by signals from the nutrient-responsive kinase ATG1, the ubiquitin subunit, and autophagy receptors, including RPN10. RPN10 acts as a selective autophagy receptor that targets inactive 26S proteasomes through simultaneous interactions with ubiquitylated subunits/targets of proteasomes and lipidated ATG8 lining enveloped autophagic membranes [44].
Positive regulation of autophagy by 2-Cys peroxiredoxin (2-CP) is confirmed by the co-expression of transcripts encoding 2-Cys peroxiredoxin and ATG in tomato plants exposed to heat stress [45]. Silencing ATG5 or ATG7 increased transcript and protein levels of 2-CP but decreased heat stress tolerance, suggesting that peroxiredoxin plays an important role in stress adaptation of plants through activation of autophagy [45]. Another protein, triosephosphate isomerase is also involved in the autophagy process. For example, it was shown that airborne PM0.1 nanoparticles (diameter <100 nm) induce autophagic cell death, which was characterized by the expression of ATG3, 7 and 8 proteins and triosephosphate isomerase in SH-SY5Y cells [46].
Taken together, our data demonstrate that numerous proteins involved in autophagy in wheat roots are capable of S-nitrosylation. Mechanisms of their involvement in the autophagic machinery are diverse and can include both direct interactions with ATG proteins and indirect interactions with components of signaling transduction pathways, e.g. receptors and protein kinases, resulting in suppression or activation of autophagic flux.
5. Conclusions
In this study we show that S-nitrosylation is a key mechanism of NO-mediated regulation of autophagy in wheat roots. Detection of the -SNO group directly in a mixture of proteins by mass spectrometry is known to be a challenging experimental task. An additional difficulty is that the S-NO bond is sensitive to hydrolysis and unstable, which makes detection of SNO-modifications of proteins even more difficult. Nevertheless, the methods of immunoblotting and standard bottom-up proteomics in combination with in silico predictive algorithms allowed us to identify proteins with potential S-nitrosylation sites. Using PPI networks enabled us to find the proteins that can directly interact with ATG proteins, and those that can interact with them indirectly via key multifunctional regulatory proteins. The results obtained in this work can contribute to the development of effective protocols for the identification and analysis of proteins capable of S-nitrosylation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Visualization of S-nitrosylated proteins by Western blot. Protein bands that were excised from the gel and subjected to in-gel trypsin cleavage for protein identification are marked on the blot; Figure S2: Functional annotation of 298 proteins identified proteins; Table S1: Parameters of the nanoHPLC separation method employed in the nanoLC-QqTOF-MS-based proteomics experiments; Table S2: Instrument settings applied for ESI-QqTOF-MS DDA experiments employed in the nanoLC-QqTOF-MS-based proteomics experiments; Table S3: PEAKS Studio 10.6 parameters for database search settings; Table S4: Peptides, proteins, and protein groups, identified in Triticum aestivum; Table S5: Functional classes of proteins, identified in Triticum aestivum; Table S6: Localization of proteins, identified in Triticum aestivum; Table S7: Hypothetical S-nitrosylation sites of identified proteins predicted by three programs; Table S8: Potential S-nitrosylation sites of autophagic proteins; Table S9: PPI of identified proteins, ATG proteins, and predicted functional partners.
Author Contributions
Conceptualization, F.M., A.F.; methodology, A.M., N.G., J.S.; software, A.M., J.S., E.R.; investigation, A.M., N.G., J.S.; resources, A.F.; writing—original draft preparation, A.M., F.M., J.S., A.F.; writing—review and editing, F.M., A.F.; visualization, A.M., J.S.; supervision, F.M., A.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially performed within the framework of the state assignment of the FRC KazSC RAS (AM, NG, FM). The Kazan Federal University Strategic Academic Leadership Program (PRIORITY-2030) is thanked for partial financial support (FM). The Russian Science Foundation (grant № 23-44-00101) supported the proteomics methodology (JS).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Authors greatly acknowledge the support of the Research Park of St. Petersburg State University (St. Petersburg, Russia) for providing the equipment: the UHPLC-QqTOF-MS/MS measurements were performed in the Centre for Molecular and Cell Technologies. Authors greatly appreciate Galina Novikova (Timiryazev Institute of Plant Physiology, Russian Academy of Sciences) and Richard Beckett (University of KwaZulu-Natal, South Africa) for valuable discussions, Ilya Leksin (Kazan Institute of Biochemistry and Biophysics, FRC Kazan Scientific Center, Russian Academy of Sciences) is thanked for the help with Python programming. JS thanks the infrastructural support provided by the Timiryazev Institute of Plant Physiology, Russian Academy of Sciences (theme no. 1021052706080-4-1.6.11).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Siyiannis, V.F.; Protonotarios, V.E.; Zechmann, B.; Chorianopoulou, S.N.; Müller, M.; Hawkesford, M.J.; Bouranis, D.L. Comparative spatiotemporal analysis of root aerenchyma formation processes in maize due to sulphate, nitrate or phosphate deprivation. Protoplasma 2012, 249, 671–686. [Google Scholar] [CrossRef] [PubMed]
- Hanamata, S.; Kurusu, T.; Okada, M.; Suda, A.; Kawamura, K.; Tsukada, E.; Kuchitsu, K. In vivo imaging and quantitative monitoring of autophagic flux in tobacco BY-2 cells. Plant Signal. Behav. 2013, 8, e22510. [Google Scholar] [CrossRef] [PubMed]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: the clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Sláviková, S.; Shy, G.; Yao, Y.; Glozman, R.; Levanony, H.; Pietrokovski, S.; Elazar, Z.; Galili, G. The autophagy-associated atg8 gene family operates both under favourable growth conditions and under starvation stresses in arabidopsis plants. J. Exp. Bot. 2005, 56, 2839–2849. [Google Scholar] [CrossRef]
- Shi, J.; Feng, H.; Lee, J.; Ning Chen, W. Comparative proteomics profile of lipid-cumulating oleaginous yeast: an ITRAQ-coupled 2-D LC-MS/MS analysis. PLoS ONE 2013, 8, e85532. [Google Scholar] [CrossRef] [PubMed]
- Melia, T.J.; Lystad, A.H.; Simonsen, A. Autophagosome biogenesis: from membrane growth to closure. J. Cell Biol. 2020, 219, e202002085. [Google Scholar] [CrossRef]
- Liu, B.; Huang, X.; Li, Y.; Liao, W.; Li, M.; Liu, Y.; He, R.; Feng, D.; Zhu, R.; Kurihara, H. JS-K, a nitric oxide donor, induces autophagy as a complementary mechanism inhibiting ovarian cancer. BMC Cancer 2019, 19, 645. [Google Scholar] [CrossRef]
- Sarkar, S.; Korolchuk, V.I.; Renna, M.; Imarisio, S.; Fleming, A.; Williams, A.; Garcia-Arencibia, M.; Rose, C.; Luo, S.; Underwood, B.R.; et al. Complex inhibitory effects of nitric oxide on autophagy. Mol. Cell 2011, 43, 19–32. [Google Scholar] [CrossRef]
- Zhang, X.; Jin, L.; Tian, Z.; Wang, J.; Yang, Y.; Liu, J.; Chen, Y.; Hu, C.; Chen, T.; Zhao, Y.; et al. Nitric oxide inhibits autophagy and promotes apoptosis in hepatocellular carcinoma. Cancer Sci. 2019, 110, 1054–1063. [Google Scholar] [CrossRef]
- Dmitrieva, S.A.; Ponomareva, A.A.; Gurjanov, O.P.; Mazina, A.B.; Andrianov, V.V.; Iyudin, V.S.; Minibayeva, F.V. Spermine induces autophagy in plants: possible role of NO and reactive oxygen species. Dokl. Biochem. Biophys. 2018, 483, 341–343. [Google Scholar] [CrossRef]
- Kuo, E.Y.; Chang, H.-L.; Lin, S.-T.; Lee, T.-M. High light-induced nitric oxide production induces autophagy and cell death in Chlamydomonas reinhardtii. Front. Plant Sci. 2020, 11, 772. [Google Scholar] [CrossRef]
- Lee, J.; Giordano, S.; Zhang, J. Autophagy, mitochondria and oxidative stress: cross-talk and redox signalling. Biochem. J. 2012, 441, 523–540. [Google Scholar] [CrossRef] [PubMed]
- Bignon, E.; Allega, M.F.; Lucchetta, M.; Tiberti, M.; Papaleo, E. Computational structural biology of S-nitrosylation of cancer targets. Front. Oncol. 2018, 8, 272. [Google Scholar] [CrossRef]
- Astier, J.; Kulik, A.; Koen, E.; Besson-Bard, A.; Bourque, S.; Jeandroz, S.; Lamotte, O.; Wendehenne, D. Protein S-nitrosylation: what’s going on in plants? Free Radic. Biol. Med. 2012, 53, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- León, J. Protein tyrosine nitration in plant nitric oxide signaling. Front. Plant Sci. 2022, 13, 859374. [Google Scholar] [CrossRef]
- Corpas, F.J.; González-Gordo, S.; Palma, J.M. Protein nitration: a connecting bridge between nitric oxide (NO) and plant stress. Plant Stress 2021, 2, 100026. [Google Scholar] [CrossRef]
- Montagna, C.; Rizza, S.; Maiani, E.; Piredda, L.; Filomeni, G.; Cecconi, F. To eat, or NOt to eat: S -nitrosylation signaling in autophagy. FEBS J. 2016, 283, 3857–3869. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Wang, L.; Wang, P.; Xue, Y.; Li, X.; Qiao, X.; Zhang, X.; Xu, T.; Liu, G.; et al. Autophagy impairment mediated by S-nitrosation of ATG4B leads to neurotoxicity in response to hyperglycemia. Autophagy 2017, 13, 1145–1160. [Google Scholar] [CrossRef]
- Oh, C.; Dolatabadi, N.; Cieplak, P.; Diaz-Meco, M.T.; Moscat, J.; Nolan, J.P.; Nakamura, T.; Lipton, S.A. S-nitrosylation of p62 inhibits autophagic flux to promote α-synuclein secretion and spread in Parkinson’s disease and Lewy body dementia. J. Neurosci. 2022, 42, 3011–3024. [Google Scholar] [CrossRef]
- Wright, C.; Iyer, A.K.V.; Kulkarni, Y.; Azad, N. S -nitrosylation of Bcl-2 negatively affects autophagy in lung epithelial cells. J. Cell. Biochem. 2016, 117, 521–532. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, C.; Liu, Q. PTEN S-nitrosylation by NOS1 inhibits autophagy in NPC cells. Cell Death Dis. 2019, 10, 306. [Google Scholar] [CrossRef]
- Nagarajan, N.; Oka, S.; Nah, J.; Wu, C.; Zhai, P.; Mukai, R.; Xu, X.; Kashyap, S.; Huang, C.-Y.; Sung, E.-A.; et al. Thioredoxin 1 promotes autophagy through transnitrosylation of Atg7 during myocardial ischemia. J. Clin. Investig. 2023, 133, e162326. [Google Scholar] [CrossRef] [PubMed]
- Guha, P.; Harraz, M.M.; Snyder, S.H. Cocaine elicits autophagic cytotoxicity via a nitric oxide-GAPDH signaling cascade. Proc. Natl. Acad. Sci. U.S.A. 2016, 113, 1417–1422. [Google Scholar] [CrossRef]
- Zhan, N.; Wang, C.; Chen, L.; Yang, H.; Feng, J.; Gong, X.; Ren, B.; Wu, R.; Mu, J.; Li, Y.; et al. S-nitrosylation targets GSNO reductase for selective autophagy during hypoxia responses in plants. Mol. Cell 2018, 71, 142–154. [Google Scholar] [CrossRef]
- Minibayeva, F.; Dmitrieva, S.; Ponomareva, A.; Ryabovol, V. Oxidative stress-induced autophagy in plants: the role of mitochondria. Plant Physiol. Biochem. 2012, 59, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Mazina, A.B.; Dmitrieva, S.A.; Minibayeva, F.V. (2017). Donors of nitric oxide as autophagy indicators in wheat cells. In Proceedings of Molecular Aspects of Plant Redox Metabolism, Ufa, Russia, 351-354., June 2017. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970, 227, 680. [Google Scholar] [CrossRef]
- Bassal, M.; Abukhalaf, M.; Majovsky, P.; Thieme, D.; Herr, T.; Ayash, M.; Tabassum, N.; Al Shweiki, M.R.; Proksch, C.; Hmedat, A.; et al. Reshaping of the Arabidopsis thaliana proteome landscape and co-regulation of proteins in development and immunity. Mol. Plant 2020, 13, 1709–1732. [Google Scholar] [CrossRef] [PubMed]
- Spiller, S.; Frolov, A.; Hoffmann, R. Quantification of specific glycation sites in human serum albumin as prospective type 2 diabetes mellitus biomarkers. Protein Pept. Lett. 2018, 24, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Shumilina, J.; Kiryushkin, A.S.; Frolova, N.; Mashkina, V.; Ilina, E.L.; Puchkova, V.A.; Danko, K.; Silinskaya, S.; Serebryakov, E.B.; Soboleva, A.; et al. Integrative proteomics and metabolomics analysis reveals the role of small signaling peptide rapid alkalinization factor 34 (RALF34) in cucumber roots. Int. J. Mol. Sci. 2023, 24, 7654. [Google Scholar] [CrossRef]
- Chen, S.-J.; Liao, D.-L.; Chen, C.-H.; Wang, T.-Y.; Chen, K.-C. Construction and analysis of protein-protein interaction network of heroin use disorder. Sci. Rep. 2019, 9, 4980. [Google Scholar] [CrossRef]
- Begara-Morales, J.C.; Chaki, M.; Valderrama, R.; Sánchez-Calvo, B.; Mata-Pérez, C.; Padilla, M.N.; Corpas, F.J.; Barroso, J.B. Nitric oxide buffering and conditional nitric oxide release in stress response. J. Exp. Bot. 2018, 69, 3425–3438. [Google Scholar] [CrossRef] [PubMed]
- Galeeva, E.I.; Trifonova, T.V.; Ponomareva, A.A.; Viktorova, L.V.; Minibayeva, F.V. Nitrate reductase from Triticum aestivum leaves: regulation of activity and possible role in production of nitric oxide. Biochem. (Mosc.). 2012, 77, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Kumari, A.; Florez-Sarasa, I.; Fernie, A.R.; Igamberdiev, A.U. Interaction of nitric oxide with the components of the plant mitochondrial electron transport chain. J. Exp. Bot. 2018, 69, 3413–3424. [Google Scholar] [CrossRef] [PubMed]
- Antonova, K.; Vikhnina, M.; Soboleva, A.; Mehmood, T.; Heymich, M.-L.; Leonova, T.; Bankin, M.; Lukasheva, E.; Gensberger-Reigl, S.; Medvedev, S.; et al. Analysis of chemically labile glycation adducts in seed proteins: case study of methylglyoxal-derived hydroimidazolone 1 (MG-H1). Int. J. Mol. Sci. 2019, 20, 3659. [Google Scholar] [CrossRef] [PubMed]
- Benhar, M.; Forrester, M.T.; Stamler, J.S. Protein denitrosylation: enzymatic mechanisms and cellular functions. Nat. Rev. Mol. Cell Biol. 2009, 10, 721–732. [Google Scholar] [CrossRef]
- Leiper, J.; Murray-Rust, J.; McDonald, N.; Vallance, P. S -nitrosylation of dimethylarginine dimethylaminohydrolase regulates enzyme activity: further interactions between nitric oxide synthase and dimethylarginine dimethylaminohydrolase. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 13527–13532. [Google Scholar] [CrossRef]
- Sisti, G.; Kanninen, T.T.; Ramer, I.; Witkin, S.S. Interaction between the inducible 70-KDa heat shock protein and autophagy: effects on fertility and pregnancy. Cell Stress and Chaperones 2015, 20, 753–758. [Google Scholar] [CrossRef]
- Hurley, J.H.; Young, L.N. Mechanisms of autophagy initiation. Annu. Rev. Biochem. 2017, 86, 225–244. [Google Scholar] [CrossRef]
- Jia, H.; Liang, Z.; Zhang, X.; Wang, J.; Xu, W.; Qian, H. 14-3-3 proteins: an important regulator of autophagy in diseases. Am. J. Transl. Res. 2017, 9, 4738–4746. [Google Scholar]
- Chodasiewicz, M.; Kerber, O.; Gorka, M.; Moreno, J.C.; Maruri-Lopez, I.; Minen, R.I.; Sampathkumar, A.; Nelson, A.D.L.; Skirycz, A. 2′,3′-CAMP treatment mimics the stress molecular response in Arabidopsis thaliana. Plant Physiol. 2022, 188, 1966–1978. [Google Scholar] [CrossRef]
- Pereira, C.; Chaves, S.; Alves, S.; Salin, B.; Camougrand, N.; Manon, S.; Sousa, M.J.; Côrte-Real, M. Mitochondrial degradation in acetic acid-induced yeast apoptosis: the role of pep4 and the ADP/ATP carrier: mitochondria degradation in apoptosis. Mol. Microbiol. 2010, 76, 1398–1410. [Google Scholar] [CrossRef] [PubMed]
- Ryder, L.; Arendrup, F.S.; Martínez, J.F.; Snieckute, G.; Pecorari, C.; Shah, R.A.; Lund, A.H.; Blasius, M.; Bekker-Jensen, S. Nitric oxide-induced ribosome collision activates ribosomal surveillance mechanisms. Cell Death Dis. 2023, 14, 467. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, S.; Zhang, H.; Xu, J. Identification of nitric oxide as an endogenous inhibitor of 26S proteasomes in vascular endothelial cells. PLoS ONE 2014, 9, e98486. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.S.; Li, F.; Gemperline, D.C.; Book, A.J.; Vierstra, R.D. Autophagic degradation of the 26S proteasome is mediated by the dual ATG8/ubiquitin receptor RPN10 in Arabidopsis. Mol. Cell 2015, 58, 1053–1066. [Google Scholar] [CrossRef]
- Cheng, F.; Yin, L.-L.; Zhou, J.; Xia, X.-J.; Shi, K.; Yu, J.-Q.; Zhou, Y.-H.; Foyer, C.H. Interactions between 2-Cys peroxiredoxins and ascorbate in autophagosome formation during the heat stress response in Solanum lycopersicum. J. Exp. Bot. 2016, 67, 1919–1933. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.-M.; Lee, M.-Y. Airborne nanoparticles (PM 0.1) induce autophagic cell death of human neuronal cells: autophagic cell death by PM 0.1. J. Appl. Toxicol. 2016, 36, 1332–1342. [Google Scholar] [CrossRef]
Figure 1.
Electropherogram of total water-soluble proteins from wheat roots stained with colloidal Coomassie Brilliant blue G-250 (a). Visualization of S-nitrosylated proteins by Western blot after incubation with mouse monoclonal primary anti-Nitroso-L-cysteine antibodies and secondary anti-mouse IgG antibodies conjugated with horseradish peroxidase (b).
Figure 1.
Electropherogram of total water-soluble proteins from wheat roots stained with colloidal Coomassie Brilliant blue G-250 (a). Visualization of S-nitrosylated proteins by Western blot after incubation with mouse monoclonal primary anti-Nitroso-L-cysteine antibodies and secondary anti-mouse IgG antibodies conjugated with horseradish peroxidase (b).

Figure 2.
Venn diagram: intersection of proteins from the samples of different treatments. The numbers of the proteins identified by the tandem mass spectra of corresponding peptides in the digests obtained from the protein isolates of the roots treated with KNO2, Antimycin A and GSNO (a); prediction of cellular localization of identified proteins.
Figure 2.
Venn diagram: intersection of proteins from the samples of different treatments. The numbers of the proteins identified by the tandem mass spectra of corresponding peptides in the digests obtained from the protein isolates of the roots treated with KNO2, Antimycin A and GSNO (a); prediction of cellular localization of identified proteins.
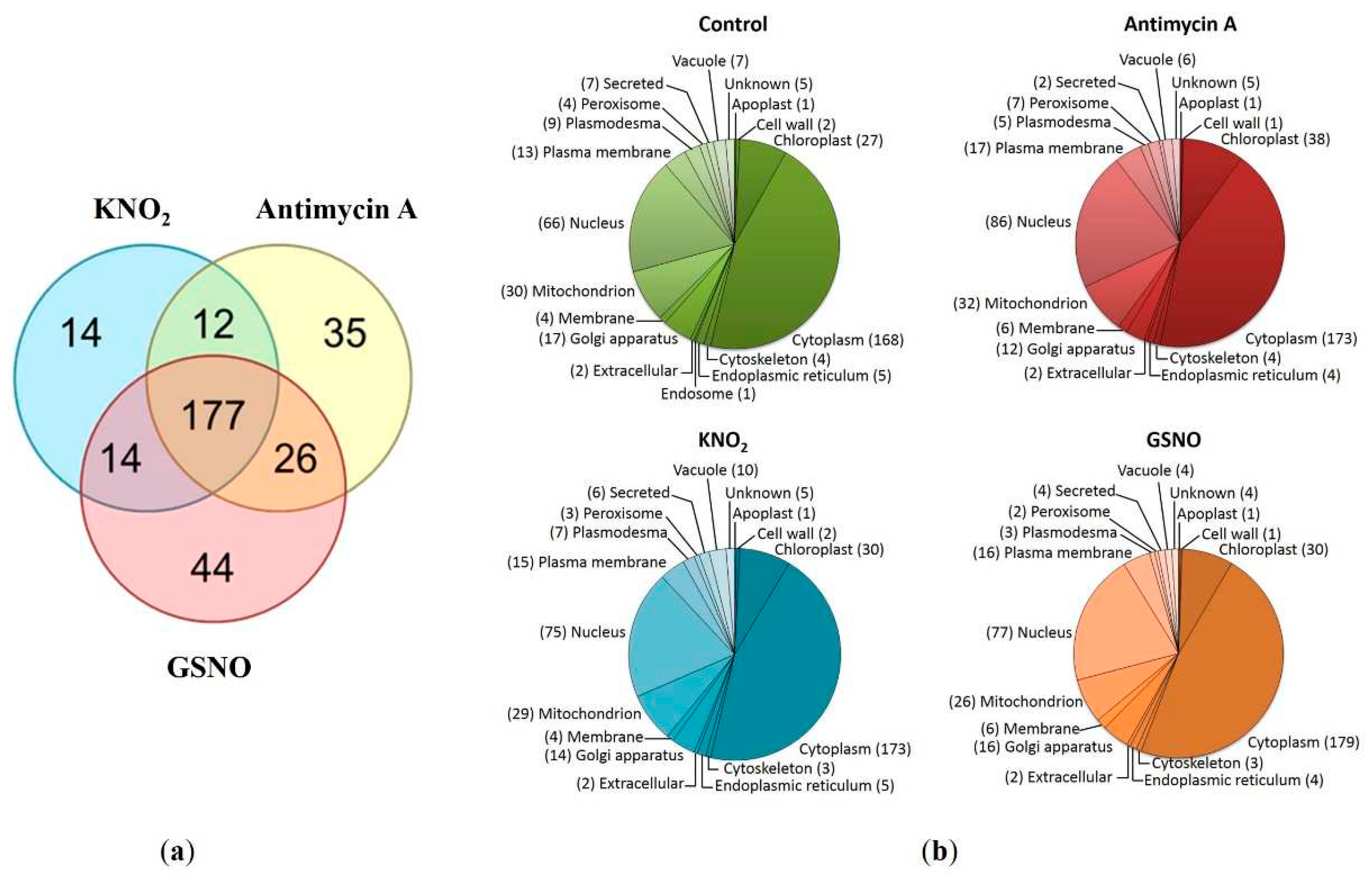
Figure 3.
Functional annotation of the total identified proteins. The proteins representing the major (a) and minor (b) functional categories (i.e. the functional groups with the most and the least represented number of proteins, respectively).
Figure 3.
Functional annotation of the total identified proteins. The proteins representing the major (a) and minor (b) functional categories (i.e. the functional groups with the most and the least represented number of proteins, respectively).
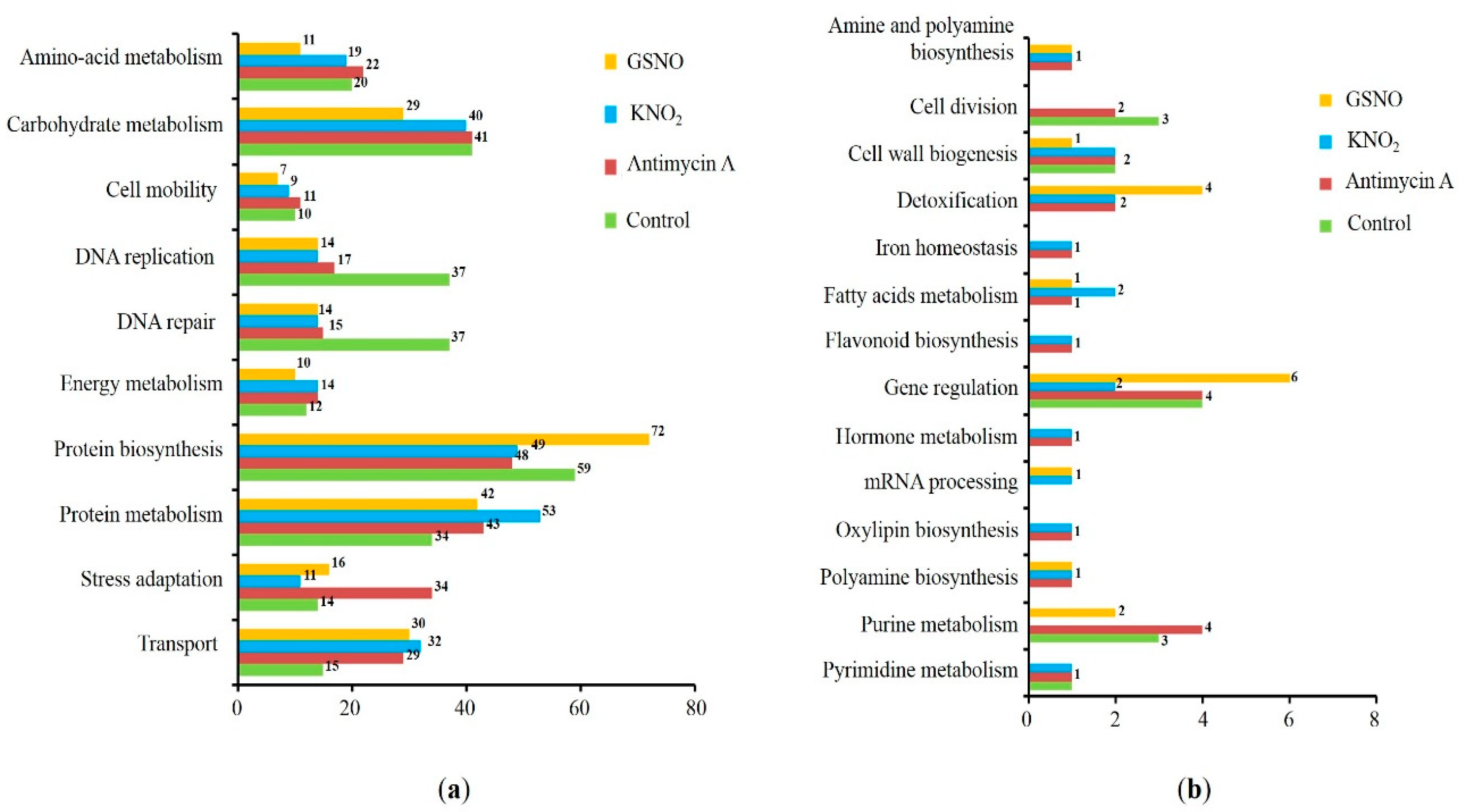
Figure 4.
Predicted intracellular localization (a) and functional annotation (b) of S-nitrosylated proteins in wheat roots treated with antimycin A, KNO2 and GSNO predicted by four bioinformatic programs (see Table 1).
Figure 4.
Predicted intracellular localization (a) and functional annotation (b) of S-nitrosylated proteins in wheat roots treated with antimycin A, KNO2 and GSNO predicted by four bioinformatic programs (see Table 1).
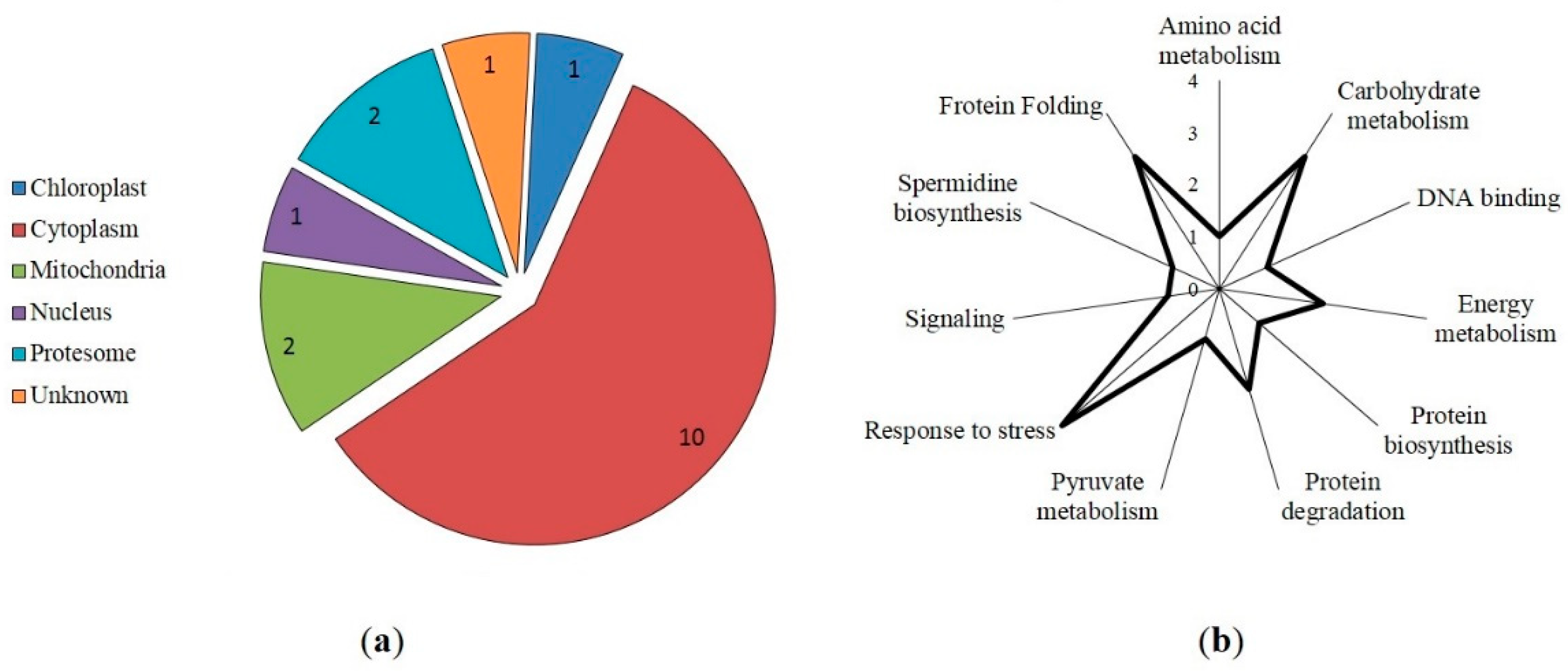
Figure 5.
Protein-ligand molecular docking. Interaction (highlighted in green) of cysteine (highlighted in red) of triosophosphate isomerase with GSNO.
Figure 5.
Protein-ligand molecular docking. Interaction (highlighted in green) of cysteine (highlighted in red) of triosophosphate isomerase with GSNO.
Figure 6.
STRING protein-protein interaction network based on the homology to proteins from Arabidopsis thaliana. Proteins marked in red were identified and predicted to have a hypothetical S-nitrosylation site; blue proteins were autophagy-related proteins; navy blue proteins were multifunctional regulatory proteins involved in autophagy. Colored lines between proteins indicate different types of interaction evidence: blue - from curated databases, pink - experimentally determined, yellow – textmining and black - co-expression.
Figure 6.
STRING protein-protein interaction network based on the homology to proteins from Arabidopsis thaliana. Proteins marked in red were identified and predicted to have a hypothetical S-nitrosylation site; blue proteins were autophagy-related proteins; navy blue proteins were multifunctional regulatory proteins involved in autophagy. Colored lines between proteins indicate different types of interaction evidence: blue - from curated databases, pink - experimentally determined, yellow – textmining and black - co-expression.
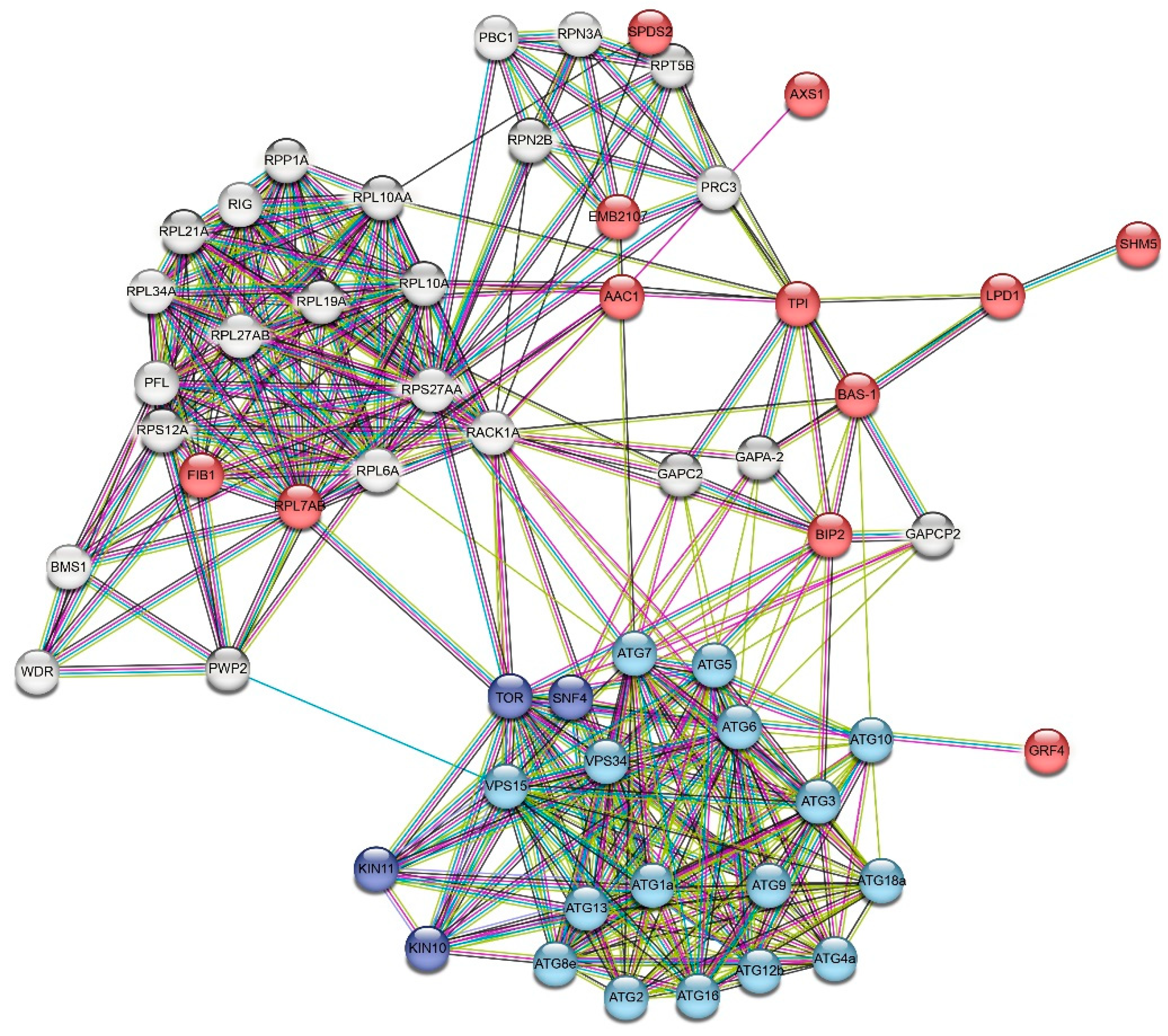

Table 1.
Hypothetical S-nitrosylation sites of identified proteins predicted by iSNOPseAAC, iSNO-AAPair, GPS-SNO 1.0 and pLMSNOSite.
Table 1.
Hypothetical S-nitrosylation sites of identified proteins predicted by iSNOPseAAC, iSNO-AAPair, GPS-SNO 1.0 and pLMSNOSite.
| Uniprot ID | Protein | Hypothetical S-nitrosylation sites |
|---|---|---|
| P46077 | 14-3-3-like protein GF14 phi | 106 |
| Q9FIB6 | 26S proteasome non-ATPase regulatory subunit 12 homolog A | 397 |
| Q9LNU4 | 26S proteasome non-ATPase regulatory subunit 3 homolog A | 141 |
| P80602 | 2-Cys peroxiredoxin BAS1 chloroplastic (Fragment) | 64, 185 |
| Q9LZH9 | 60S ribosomal protein L7a-2 | 193 |
| P31167 | ADP ATP carrier protein 1 mitochondrial | 130 |
| Q41629 | ADP ATP carrier protein 1 mitochondrial | 81, 206 |
| A8MS68 | Dihydrolipoyl dehydrogenase 1 chloroplastic | 400 |
| Q9S7C0 | Heat shock 70 kDa protein 14 | 268 |
| F4HQD4 | Heat shock 70 kDa protein 15 | 268 |
| Q39043 | Heat shock 70 kDa protein BIP2 | 298 |
| Q9FEF8 | rRNA 2'-O-methyltransferase fibrillarin 1 | 252 |
| Q9SVM4 | Serine hydroxymethyltransferase 5 | 324 |
| O48661 | Spermidine synthase 2 | 43 |
| P48491 | Triosephosphate isomerase cytosolic | 13, 127 |
| Q9ZUY6 | UDP-D-apiose/UDP-D-xylose synthase 1 | 187 |
| Q9SGE0 | UDP-D-apiose/UDP-D-xylose synthase 2 | 187 |
Table 2.
List of the proteins included in the STRING protein-protein interaction network: proteins with hypothetical S-nitrosylation sites, ATG proteins and 13 predicted functional partners proposed by the STRING program.
Table 2.
List of the proteins included in the STRING protein-protein interaction network: proteins with hypothetical S-nitrosylation sites, ATG proteins and 13 predicted functional partners proposed by the STRING program.
| Name | Description (UNIPROT) | UNIPROT ID |
|---|---|---|
| GRF4 | 14-3-3-like protein GF14 phi | P46077 |
| EMB2107 | 26S proteasome non-ATPase regulatory subunit 12 homolog A | Q9FIB6 |
| BAS-1 | 2-Cys peroxiredoxin BAS1 chloroplastic (Fragment) | Q9C5R8 |
| RPL7AB | 60S ribosomal protein L7a-2 | Q9LZH9 |
| AAC1 | ADP ATP carrier protein 1 mitochondrial | P31167 |
| LPD1 | Dihydrolipoyl dehydrogenase 1 chloroplastic | A8MS68 |
| BIP2 | Heat shock 70 kDa protein BIP2 | Q39043 |
| FIB1 | rRNA 2'-O-methyltransferase fibrillarin 1 | Q9FEF8 |
| SHM5 | Serine hydroxymethyltransferase 5 | Q9SVM4 |
| SDS2 | Spermidine synthase 2 | O48661 |
| TPI | Triosephosphate isomerase cytosolic | P48491 |
| AXS1 | UDP-D-apiose/UDP-D-xylose synthase 1 | Q9ZUY6 |
| ATG1a | Serine/threonine-protein kinase ATG1a | Q94C95 |
| ATG2 | Autophagy-related protein 2 | F8S296 |
| ATG3 | Autophagy-related protein 3 | Q0WWQ1 |
| ATG4a | Cysteine protease ATG4a | Q8S929 |
| ATG5 | Autophagy protein 5 | Q9FFI2 |
| ATG6 | Beclin-1-like protein | Q9M367 |
| ATG7 | Ubiquitin-like modifier-activating enzyme atg7 | Q94CD5 |
| ATG8e | Autophagy-related protein 8e | Q8S926 |
| ATG9 | Autophagy-related protein 9 | Q8RUS5 |
| ATG10 | Ubiquitin-like-conjugating enzyme ATG10 | Q8VZ52 |
| ATG12b | Ubiquitin-like protein ATG12B | Q9LVK3 |
| ATG13 | Autophagy-related protein 13a | Q9SCK0 |
| ATG16 | Autophagy-related protein 16 | Q6NNP0 |
| ATG18a | Autophagy-related protein 18a | Q93VB2 |
| TOR | Serine/threonine-protein kinase TOR | Q9FR53 |
| VPS15 | Serine/threonine-protein kinase VPS15 | Q9M0E5 |
| VPS34 | Phosphatidylinositol 3-kinase VPS34 | P42339 |
| KIN10 | SNF1-related protein kinase catalytic subunit alpha KIN10 | Q38997 |
| KIN11 | SNF1-related protein kinase catalytic subunit alpha KIN11 | P92958 |
| SNF4 | Sucrose nonfermenting 4-like protein | Q944A6 |
| GAPC2 | Glyceraldehyde-3-phosphate dehydrogenase GAPC2, cytosolic | Q9FX54 |
| GAPA-2 | Glyceraldehyde-3-phosphate dehydrogenase GAPA2, chloroplastic | Q9LPW0 |
| GAPCP2 | Glyceraldehyde-3-phosphate dehydrogenase GAPC2, cytosolic | Q9FX54 |
| PRC3 | Proteasome subunit alpha type-2-A | O23708 |
| RPT5B | 26S proteasome regulatory subunit 6A homolog B | O04019 |
| RPN3A | 26S proteasome non-ATPase regulatory subunit 3 homolog A | Q9LNU4 |
| PBC1 | Proteasome subunit beta type-3-A | Q9XI05 |
| RPN2B | 26S proteasome non-ATPase regulatory subunit 1 homolog B | Q9MAT0 |
| RPL10AA | Large ribosomal subunit protein uL1z | Q8VZB9 |
| RPL10A | Large ribosomal subunit protein uL16z | Q93VT9 |
| RPS27AA | Ubiquitin-ribosomal protein eS31z fusion protein | P59271 |
| RACK1A | Small ribosomal subunit protein RACK1z | O24456 |
| RPL6A | Large ribosomal subunit protein eL6z | Q9FZ76 |
| RPL19A | Large ribosomal subunit protein eL19x | Q9SRX2 |
| RPP1A | Large ribosomal subunit protein P1w | Q8LCW9 |
| RIG | Small ribosomal subunit protein uS19u | Q08112 |
| RPL21A | Large ribosomal subunit protein eL21z/eL21y | Q43291 |
| RPL27AB | Large ribosomal subunit protein uL15y | Q9LR33 |
| RPL34A | Large ribosomal subunit protein eL34z | Q42351 |
| PFL | Small ribosomal subunit protein uS13z/uS13y/uS13x | P34788 |
| RPS12A | Small ribosomal subunit protein eS12z | Q9S9P1 |
| BMS1 | P-loop containing nucleoside triphosphate hydrolases superfamily protein | F4IDR3 |
| WDR | Uncharacterized protein At1g15425 | Q8L403 |
| PWP2 | Periodic tryptophan protein 2 | Q8VYZ5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



