
Submitted:
16 August 2023
Posted:
17 August 2023
You are already at the latest version
Abstract
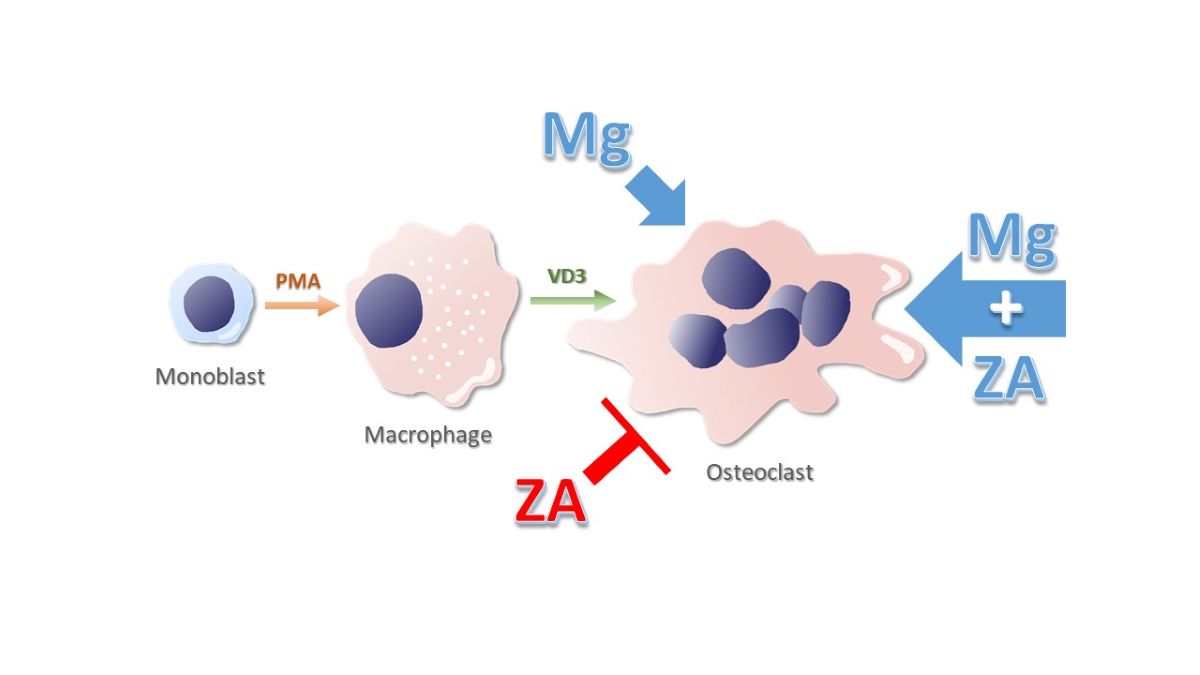
Bisphosphonates (BPs) are successfully used to cure a number of bone diseases characterized by metabolic osteopenia, such as Osteoporosis, or neoplastic osteolysis, such as Multiple Myeloma and Bone Metastases. These drugs exert their therapeutic effect inducing a systemic osteoclast depletion that, in turn, is responsible for a reduced bone resorption. Unfortunately, in addition to their beneficial activity, BPs can also determine a frightening side effect known as Osteonecrosis of the Jaw (ONJ). It is generally believed that the inability of osteoclasts to get rid of inflamed / necrotic bone tissue represents the main physio-pathological aspect of ONJ. In principle, a therapeutic strategy able to elicit a local re-activation of osteoclast production could counteract ONJ and promote the healing of its lesions. We have previously demonstrated that Magnesium (Mg) is a powerful inducer of osteoclast differentiation. Using a model of Vitamin D3 – dependent osteoclastogenesis, based on the U937 cell line, here we show that, surprisingly, this property of Mg is greatly enhanced by the presence of Zoledronic Acid (ZA), chosen for our study because it is the most effective among BPs. This finding allow us to hypothesize that Mg might play an important role in the topical therapy of ONJ.
Keywords:
bisphosphonates
; zoledronate
; ONJ
; osteoclasts
; magnesium
; differentiation
1. Introduction
Several diseases give rise to a metabolic reduction of bone density, called osteopenia, that is for example observed in osteoporosis, or a neoplastic destruction of bone tissue, named osteolysis, that is instead detected in Multiple Myeloma and Bone Metastases of Solid Tumours. All these conditions can be cured using a class of drugs, defined Bisphosphonates (BPs), that counteract the loss of bone tissue by inhibiting the function of osteoclasts, i.e. the cells normally mediating bone resorption [1]. It has to be pointed out that, in addition to their anti-resorptive effect, BPs are also endowed with an intrinsic anti - tumour activity allowing them to directly limit the proliferation of malignant cells [2,3,4]. The practical consequences of a treatment with BPs are thus represented, in general, by a reduction of bone pains and a lower risk to develop bone fractures that, in the specific case of bone tumours, can also be accompanied by a slowdown of disease progression. The denomination of BPs reflects the presence of two phosphate groups in their molecule rendering them structurally similar to Pyrophosphate (PP). Thanks to this feature, once administered to the patient, BPs are preferentially delivered to the bone tissue, where they bind to the hydroxyapatite crystals of its extra-cellular matrix [5]. In this location BPs accumulate at very high concentrations, persisting there for long periods of time, even for the entire life of patient. When bone is resorbed, BPs are released and internalized in great amounts by osteoclasts, causing their suppression that can take place with different mechanisms, depending on the category they belong to. In fact, based on the presence of nitrogen in their molecule, BPs are divided in two groups: Nitrogen – containing (N- BPs) and Non – Nitrogen – containing (Non-N-BPs). N-BPs, like Zoledronate and Pamidronate, act inhibiting the farnesylation of Ras, required for tyrosine-kinase receptor signaling [6,7]. This effect is reached through a competition with a set of PP – bound lipids mediating such reaction. Non-N-BPs, like Clodronate and Etidronate, instead, act replacing PP in the ATP molecule, thus impairing its hydrolysis and the contextual release of energy [8]. From a biological point of view, it is therefore evident that both mechanisms are based on the capacity of BPs to mimic PP structure, finally leading to a common effect that is always represented by the apoptosis of osteoclasts [9,10]. On the other hand, among the more clinical aspects diversifying the two BP categories, it is worth mentioning that N-BPs are more powerful and prevalently used in neoplastic bone disorders whereas Non-N-BPs are less active and substantially used in metabolic bone diseases [7]. Unfortunately, despite their beneficial activity, BPs can also determine a frightening side effect named Osteonecrosis of the Jaw (ONJ) [11,12,13,14]. This condition appears, as a rule, with a gingival ulceration that is less frequently associated to suppuration and pain. Although ONJ can arise spontaneously [15,16], it is more often triggered by a surgical intervention, typically a tooth extraction [17,18]. According to the current view, the primary event causing ONJ is the invasion of alveolar bone by bacteria deriving from the oral cavity [19]. This circumstance is then responsible for a situation of infection, inflammation and necrosis of the interested site. It is conceivable that the osteoclast depletion, induced by BP treatment [20,21,22], favors both the entry of bacteria into the initially healthy bone and then the inability to eliminate the damaged bone. As expected, the probability to undergo ONJ correlates with the efficacy of the responsible BP and, on this basis, it is higher with N-BPs as compared to Non-N-NPs. With Zoledronate, the most powerful but also the most hazardous among BPs, the cumulative risk to develop an ONJ has been estimated 1% for each year of treatment. To date there is no an available medical therapy to cure this condition that can be exclusively approached with a surgical treatment consisting of the removal of necrotic bone [10,23,24,25,26]. The local re-activation of osteoclast function, inhibited by the systemic therapy with BPs might, in theory, cure and, perhaps, also prevent ONJ [27,28]. Using an experimental model of Vitamin D3 dependent osteoclastogenesis, based on the U937 cell line, we have previously demonstrated that Magnesium is a powerful inducer of osteoclast differentiation [29,30,31,32]. Starting from this assumption, and using the same system, we tried therefore to clarify whether it is able to maintain its activity in presence of Zoledronate, selected for our project in light of its peculiar features, already explained above. The results obtained were really surprising since they not only demonstrated the inability of Zoledronate to contrast the osteoclast differentiation capacity of Magnesium, but they also put in evidence that the latter was strongly potentiated by the presence of the former.
2. Materials and Methods
2.1. Cell cultures
The U937 cell line was obtained from the American Type Culture Collection (ATCC; Rockville, MD, USA) and cultured at 37°C, 5% CO2 in RPMI1640 medium (Euroclone, Devon, UK), supplemented with 10% heat inactivated Foetal Bovine Serum (FBS) (Sigma-Aldrich, St. Louis, MO, USA) and 1 mM L-Glutamine (Euroclone). Zoledronic Acid (ZA) and Magnesium Chloride (MgCl2) were both purchased from Sigma-Aldrich, dissolved in sterile water and added to cell cultures at the desired concentrations. The osteoclast differentiation of U937 cells was induced by a 2 - day stimulation with 48 nM Phorbol 12-Myristate 13-Acetate (PMA) (Sigma-Aldrich), followed by a PBS rinse, aimed to remove non adhering undifferentiated cells, and then by a 3 - day stimulation with 10-8 M 1α, 25 di-hydroxy Vitamin D3 (VD3) (Sigma – Aldrich). Cell counts were performed with a TC20 automated cell counter (Bio-rad, Munich, Germany) always considering alive cells in the size range between 8 and 18 μm. Morphological analysis was carried out using a Evos M500 Imaging System (Thermo Fisher Scientific, Waltham, MA, USA) digital microscope on cyto-centrifuged and by May-Grunwald- Giemsa stained specimens.
2.2. Flow cytometry analysis
Analysis of cell cycle and apoptosis was performed on proliferating U937 cells upon a 30 min incubation at 4°C with Nicoletti’s solution, containing 20 μg / ml Propidium Iodide (PI) 0,1 % Triton X–100, 0,1 % Tri-sodium citrate, all provided by Sigma and dissolved in distilled water. Immune-phenotypic analysis of differentiated U937 cells was conducted by a 30 min incubation at 4°C with phycoerythrin-conjugated (PE) mouse anti-human CD11b Monoclonal Antibody (MoAb) (Miltenyi Biotec, Auburn, CA, USA), in PBS containing 5% FCS and 1% FcR blocking reagent (Miltenyi). At the end of labeling, cells were washed twice and finally re-suspended with PBS. All samples undergoing flow cytometry were finally analyzed using an Attune NxT flow cytometer (Thermo Fisher Scientific, Waltham, MA, USA).
2.3. RNA extraction and QRT-PCR reaction
Total RNA was purified using the Qiagen RNeasy plus mini kit (Qiagen, Valencia, CA, USA) and, once extracted, its concentration was assessed using a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific). Messenger RNA expression was analyzed with a Quantitative Real Time – Polymerase Chain Reaction (QRT-PCR) procedure based on the Taqman method. For this purpose, 200 ng of each RNA sample were subjected to Reverse Transcription (RT) using the HiScript III RT SuperMix for qPCR (Vazyme Biotech, Nanjing, China), that was handled according the manufacturer’s instructions. Amplification was then performed by adding to the cDNA sample the Taqman Gene Expression Master Mix (Thermo Fisher Scientific) and the proper Taqman Gene Expression Assays (Thermo Fisher Scientific), listed below together with their gene symbol and catalogue numbers: D3 cyclin (CCND3; Hs01017690_g1), Cyclin Dependent Kinase Inhibitor 1 A (CDKN1A, also known as p21; Hs00355782_m1), Nuclear Factor of Activated T cells 1 (NFATC1; Hs00542678_m1), Dendritic Cell Specific Transmembrane protein 1 (DCST1; Hs00984780_m1), Acid Phosphatase (ACP5, also known as Tartrate Resistant Acis Phosphatase or TRAP; Hs00356261_m1), Cathepsin (CTSK; Hs00166156_m1), Matrix Metallo Proteinase 9 (MMP9; Hs00234579_m1), Musculo – Aponeurotic Fibrosarcoma oncogene homologue B (MAFB; Hs00534343_s1), Cluster Designation 14 (CD14; Hs02621496_s1), Cluster Designation 163 (CD163; Hs00174705_m1), Glyceraldehyde - 3 - Dehydrogenase (GAPDH; Hs02786624_m1). QRT-PCR reaction was run in triplicate in a Light Cycler 480 detection system (Roche Diagnostics, Mannheim, Germany) thermal cycler. Normalization of signals was obtained using the Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) mRNA as endogenous control. Statistical analysis of QRT-PCR results was conducted using the 2-ΔΔCt method which calculates the relative changes in gene expression of the considered target mRNA normalized to the endogenous control and related to a calibrator sample. The values obtained were represented in terms of relative quantity of mRNA level variations.
2.4. Statistical Analysis
All experiments were repeated at least three times and results were presented as mean ± S.E.M. values. Pairwise comparisons were carried out using the Student’s t-test whereas multiple comparisons were performed with the Anova procedure. Results of statistical analysis were considered significant when exhibiting p-values ≤ 0.05 and highly statistically significant when exhibiting p-values ≤ 0.01 and have been reported in Results or in Table 1.
3. Results
3.1. Proliferative and apoptotic effects determined on cycling U937 cells by scalar concentrations of Zoledronate
To assess the biological effects determined by Zoledronate (Zoledronic Acid, ZA) on proliferation and apoptosis, U937 cells were treated for up to four days with scalar concentrations of this drug ranging from 0 to 100 μM and then subjected to cell count, flow cytometry analysis upon PI staining and QRT-PCR analysis of D3 cyclin and p21 mRNA expression [33]. Statistical analysis was carried out by a Student’s t test, performed on pairwise comparisons between control untreated cells and cells treated with each tested ZA concentration.
A daily count revealed that U937 cells, treated with ZA, undergo a time and dose – dependent decrease of cell number (Figure 1, upper panel). In fact, at the end of experiment, cell concentration, indicated as million / ml, averaged 2.7 ± 0.1 in control untreated cells (0 μM) whereas the same parameter was 2.5 ± 0.2, 2.2 ± 0.2 and 0.7 ± 0.1 in samples respectively treated with 1, 10 and 100 μM ZA. Therefore, the reduction of cell number was remarkable with the highest drug concentration (100 μM) whereas it was quite limited with the lower ones (1 and 10 μM). Consistently, p values appeared highly significant for the former, already starting from the second day of treatment (0.005 at day 2, 0.0009 at day 3, 0.00003 at day 4), whereas they were not significant for the latter.
In agreement with the data presented so far, ZA treatment also elicited a dose dependent apoptotic effect (Figure 1, middle panel), indicated by the fact that, always at day 4, the mean percentage of apoptotic cells resulted 7.2 ± 1.8 in control untreated cells (0 μM) and 16.1 ± 1.8, 27.8 ± 6.8 and 64.2 ± 6.5 in cells that had been respectively treated with 1, 10 and 100 μM ZA. The corresponding p values were at least statistically significant (0.01, 0.03 and 0.007 respectively). Exposure to the same amounts of the investigated drug gave rise to a similar and not significant distribution in the various cell cycle phases with the exception of the highest, 100 μM ZA concentration, promoting a reduction of cells in the G0/G1 phase (41.9 ± 8.0 % versus 67.1 ± 5.7 % of control) and, in our opinion, a relative increase in the other cell cycle phases (for S phase, 40.9 ± 4.8 % versus 19.5 ± 5.6 % respectively). This finding suggests that, as already observed by other authors, apoptosis is preferentially activated in the G1 phase of cell cycle [2].
QRT-PCR analysis disclosed a 4.5 ± 0.9 fold increase of D3 cyclin transcript and a 4.8 ± 0.7 fold induction of p21 mRNA levels, that were exclusively observed with a 100 μM concentration of ZA, confirming the capacity of this drug to determine a growth arrest in the G1 phase of cell cycle (Figure 1, lower panel). The mRNA expression of the two analyzed genes remained, on the contrary, unaffected with the lower ZA concentrations.
Taken together, these data indicate that ZA concentrations between 1 and 10 μM, coinciding with the therapeutic range reached in vivo in the bone tissue of patient under treatment, exert the expected apoptotic effect on target cells and, at the same time, are well tolerated avoiding the strong growth arrest observed with the higher tested concentration. Consequently, the 10 μM concentration of ZA was chosen for the subsequent experiments.
3.2. Proliferative and apoptotic effects determined on cycling U937 cells by a supra- physiological concentration of MgCl2
We have previously demonstrated that 10 mM is the optimal concentration of MgCl2 favoring the osteoclast differentiation of U937 cells [30,34], but its effect on the proliferation and apoptosis of these cells has not been assessed before. For this reason, they were exposed to the mentioned concentration of MgCl2, under current cell culture conditions, and then analyzed as already explained for ZA treatment.
A cell count, performed at day 2 of the experiment and again indicated as millions / ml, put in evidence an about 15 % statistically significant reduction of cell number, from 1.2 ± 0.1 of control untreated to 1.0 ± 0.0 of MgCl2 treated cells (p = 0.03) whereas, at day 4, the same treatment gave rise to an apparently inverted effect characterized by a 5 % increase of cell number, from 2.0 ± 0.1 to 2.1 ± 0.2 respectively, although statistically not significant (Figure 2, upper panel). At the same time point, flow cytometry examination revealed similar and statistically not significant mean percentage values of apoptotic cells (8.1 ± 2.8 in MgCl2 treated versus 6.8 ± 2.1 of control untreated cells) and proliferating cells in the various cell cycle phases (respectively: 71.6 ± 3.2 versus 72.7 ± 4.9, for G0/G1, 9.6 ± 1.8 versus 8.0 ± 1.6, for S, 18.9 ± 4.6 versus 19.4 ± 4.6, for G2/M) (Figure 2, middle panel).
QRT-PCR analysis disclosed a modest but statistically significant up-regulation of the mRNA expression of both the D3 cyclin and the p21 genes, respectively undergoing a 2.3 ± 0.1 (p = 0.05) and a 2.1 ± 0.3 (p = 0.03) - fold increase in response to MgCl2 treatment (Figure 2, lower panel).
These data globally indicate that MgCl2 is able to determine a weak and transitory inhibition on the proliferation of U937 cells. This effect can be probably interpreted as a delayed cell cycle entry and / or a prolonged doubling time. However, a 10 mM concentration of the considered compound appeared overall well tolerated by cells and suitable for further experimentation.
3.3. Capacity of MgCl2 and ZA to modulate the osteoclast differentiation of U937 cells induced by stimulation with Phorbol Esters and Vitamin D3
To assess the effect determined by MgCl2 and ZA on osteoclast differentiation, we adopted an experimental model of VD3 dependent osteoclastogenesis, already used in the past in our laboratory, and consisting of U937 cells undergoing an initial 2 day stimulation with 48 nM PMA followed by a 3 day stimulation with 10-8 M VD3 [29,30,32]. Treatment with 10 mM MgCl2 and 10 μM ZA was carried out, alone or in combination, for the entire stimulation period using untreated cells as a control. At the end of cell culture, all samples were subjected to molecular, immunophenotype and morphological analysis.
Molecular analysis was performed by QRT-PCR to evaluate the mRNA expression levels of a number of osteoclast and monocyte – macrophage differentiation markers. The osteoclast markers included the following: NFATC1, the master regulator of osteoclast differentiation [35]; DCST1, a surface antigen mediating the osteoclast cell fusion [36]; ACP5, a phosphatase responsible for the degradation of the mineral component of bone extra-cellular matrix; CTSK and MMP9, two proteases responsible for the degradation of the protein component of the same matrix [37]. The monocyte – macrophage markers included instead: MAFB, the master regulator of monocyte – macrophage differentiation [38]; CD14, a surface antigen more related to monocytes and M1 activation; CD163, a surface antigen more related to macrophages and M2 activation [37,39,40]. The results of this analysis are presented in Figure 3 and Figure 4, as bar histograms, and in Table 1, as numerical values, always reporting the RQ values obtained by the various QRT-PCR reactions in terms of mean ± S.E.M. As expected, treatment with MgCl2 promoted an up-regulated mRNA expression of all the studied osteoclast differentiation markers. Among them, the most remarkable was represented by ACP5, exhibiting a 7.5 ± 1.4 - fold increase of its transcript levels, whereas for the other genes of the same group the degree of mRNA induction was: 3.0 ± 0.6 - fold for MMP9, 2.0 ± 0.3 - fold for CTSK, 1.6 ± 0.1 - fold for DCST1 and 1.3 ± 0.1 - fold for NFATC1. Among the monocyte – macrophage markers, a similar result was observed for the mRNA expression of the CD163 antigen, undergoing a 5.0 ± 1.7 - fold increase, whereas the transcript levels of the MAFB transcription factor and the CD14 antigen appeared only minimally affected by MgCl2 treatment. With the only exception of ACP5, presenting a 2.0 ± 0.3 - fold increase of its mRNA expression, treatment with ZA was substantially unable to determine an appreciable effect on both the considered gene categories. Surprising data arose, on the contrary, by the combined treatment with ZA and MgCl2, disclosing a synergic effect on virtually all the analyzed osteoclast markers. The most striking result was, again, observed for ACP5, showing a 20.9 ± 4.0 - fold up-regulation of its mRNA expression, implying an about tripled effect in comparison with MgCl2 alone. The other genes of the same group, although to a lesser extent, confirmed this trend. In fact, the entity of mRNA induction was: 3.2 ± 0.6 - fold for CTSK, 2.1 ± 0.5 - fold for DCST1 and 1.9 ± 0.1 - fold for NFATC1, thus sensibly higher values as compared to those, already listed before, elicited by MgCl2 alone. In this regard, the MMP9 gene represented an exception, exhibiting identical values under the two considered treatment conditions (3.0 ± 0.7 versus 3.0 ± 0.6, respectively). The same effect was not observed on monocyte – macrophage markers, although it is worth underlining that the transcript levels of CD163 remained as high as those observed with MgCl2 alone (4.5 ± 1.7 versus 5.0 ± 1.7). Statistical analysis, conducted with the Anova procedure, resulted highly significant for all osteoclast markers but one (DCST1) and for 1 out of the 3 monocyte – macrophage markers (CD163) (see Table 1, right column).
The table shows the numerical values of QRT-PCR described by histograms in Figure 3 and Figure 4. All experiments were repeated at least three times. Comparisons were performed between treated (Mg, MgCl2; ZA, Zoledronate; ZA + Mg, combined treatment) and untreated (Ctr) osteoclasts derived from U937 cells upon stimulation with PMA and VD3. The Table reports the RQ values obtained by the various QRT-PCR reactions in terms of mean ± SEM. Comparisons were performed with the Anova procedure, shown in the last column. Results of statistical analysis were considered significant when exhibiting p-values ≤ 0.05 and highly statistically significant when exhibiting p-values ≤ 0.01.
Immune phenotype analysis was carried out by flow cytometry assessment of the CD11b antigen for which we estimated the positivity percentage and the mean fluorescence intensity (MFI) (Figure 5) [41]. In comparison with undifferentiated U937 cells, not stimulated with PMA and VD3, all samples receiving such stimulation exhibited a CD11b positivity higher than 95 % (not shown) indicating that, at least for this parameter, the analyzed treatment conditions apparently did not affect the entity of osteoclast differentiation. In spite of this, treatment with MgCl2 resulted in a more than twice increase of CD11b MFI (151.8 ± 24.3 versus 66.8 ± 12.3 of control untreated cells) whereas ZA determined an almost 10 % reduction of the same value when used alone (62.3 ± 10.7 versus, again, 66.8 ± 12.3 of control untreated cells) and an about 15 % decrease when added to MgCl2 (129.5 ± 19.5 versus 151.8 ± 24.3 of MgCl2). Statistical analysis, conducted using the Anova procedure, attested that the differences observed among the CD11b MFI values, so far listed, were highly significant (p = 0.007).
Morphological analysis, performed on cyto-centrifuged samples upon staining with May –Grumwald - Giemsa, put in evidence that, as expected, 91 % of control untreated cells presented a morphologically differentiated phenotype, although osteoclasts belonging to this sample contained only 1 or 2 nuclei (Figure 6). Treatment with MgCl2 resulted in an apparently reduced percentage of cells with a terminally differentiated morphology (54 %) but, in this case, osteoclasts contained 3 or 4 nuclei, testifying anyway a more differentiated phenotype than control. Treatment with ZA was responsible for a dramatic reduction of fully mature cells (20 %) that were also characterized by the complete absence of multi-nucleated osteoclasts. The simultaneous treatment with ZA and MgCl2 gave rise to a differentiation effect (88 %) that was numerically similar to that of the control but with osteoclasts containing 5 to 10 nuclei.
The conclusions emerging from this part of the work are therefore that ZA alone exert an inhibitory effect on osteoclast differentiation that we can define: absent, at the molecular level, weak, on immune phenotype, and strong, on morphology. The addition of MgCl2, to ZA treated cells, determines not only a reversion of ZA effect but even the appearance of a synergism between the two compounds which became especially evident on the molecular and morphological phenotype.
4. Discussion
BPs are anti-resorptive drugs that, due their capacity to inhibit osteoclast activity, are used in several skeletal disorders all sharing the loss of bone tissue as major feature. Unfortunately, in addition to their therapeutic effect, these compounds can also provoke a serious side effect known as ONJ. The depletion of osteoclasts, responsible for a reduced ability to fight the bacterial invasion and to eliminate the infected bone, probably plays a crucial role in the development of this condition. Based on this premise we postulate that a local re-activation of osteoclast function might cure or even prevent ONJ. The starting point of our work has been represented by the observation that MgCl2 is a powerful inducer of osteoclast differentiation. To assess whether this activity is also maintained in the presence of BPs, we have used an in vitro experimental model in which U937 cells, differentiated to osteoclasts through a sequential stimulation with PMA and VD3, have been concomitantly subjected to treatment with MgCl2 and / or ZA [20,28]. The choice of this system is, in our opinion, a crucial aspect of the project and bases its biological rationale on the fact that, besides the physiological role played in the regulation of osteoclast differentiation, VD3 is also usually administered to patients under treatment with BPs. As previously observed, MgCl2 determined a remarkable induction of osteoclast differentiation that was especially evident on molecular and immune phenotype. Morphological analysis revealed the presence of two cell subpopulations one of which was represented by osteoclasts more differentiated than control whereas the other consisted of undifferentiated cells still presenting a precursor aspect. This finding was consistent with the delay of proliferation that, in parallel, was observed on the same sample by flow cytometry analysis of cell cycle. On the other hand, ZA strongly inhibited osteoclast differentiation at the morphological level whereas this effect appeared only weakly exerted on immune phenotype and totally absent at the molecular level. These data indicate a post-translational block of osteoclast differentiation that was in perfect agreement with the universally accepted mechanism of action of ZA. In fact, according to the current view, ZA and other N-BPs act inhibiting the Farnesyl Pyro-Phosphate Synthase (FPPS) enzyme which is required to add a lipid anchor to a number of G proteins promoting, in addition to other biological functions, the cytoskeleton arrangements and morphological changes that are necessary to accomplish terminal differentiation. Co- treatment with ZA and Mg resulted in a quite unexpected situation since, not only ZA was unable to inhibit the powerful differentiation effect of MgCl2 but, on the contrary, this combination gave rise to a powerful synergistic effect on osteoclast differentiation, that was especially evident at the molecular and morphological level. This conclusion was, in fact, respectively testified by the over 20 – fold increase of ACP5 mRNA expression and the massive formation of giant multi-nucleated osteoclasts containing 5 to 10 nuclei, implying that the presence of ZA amplified of about three times the osteoclast differentiation effect of MgCl2. If confirmed by proper studies in vivo, this finding would mean that, in various forms (solution, gel, foam, membrane, powder), MgCl2 or other salts, or more generically other compounds, containing Mg might be used in a topical therapy to cure or prevent ONJ. It is worth underlining that, although, based on our results, the best efficacy is expected in patients under treatment with ZA, or probably other BPs, a therapeutic effect is anyway expected also in their absence considering the capacity of MgCl2, clearly demonstrated in our experimental system, to induce osteoclast differentiation even when used alone. The molecular mechanism by which MgCl2 favors osteoclast differentiation, and other biological functions, is complex and poorly understood. What is known is that the Mg2+ cation binds to ATP4- forming MgATP2- that in turn activate about three hundred of enzymes comprising Protein Kinases and ATPases regulating a huge number of cell processes. A similar operation is mediated by Mg, through a binding activity with GTP, on GTPases. It is therefore possible that osteoclast differentiation is one of the cell processes regulated by this modality. An additional issue that remains to be elucidated is represented by the molecular mechanism by which ZA potentiates, rather than making the opposite, the osteoclast differentiation effect of MgCl2. A possible explanation, in this case, could rely in the fact that the interaction of both PP and N-BPs with the active catalytic site of FPPS enzyme is mediated by the Mg2+ cation [42,43]. It is therefore possible that, perhaps acting through a conformational change mechanism, an increase of Mg2+ concentration might determine opposite effects on the recruitment of PP and N-BP, favoring the former and inhibiting the latter. However, this hypothesis can only partly explain our data because, as far as known, the FPPS enzyme is devoid of transcriptional effects that are instead clearly observed in U937 derived osteoclasts subjected to co-treatment with ZA and MgCl2. An aspect deserving some attention is, in our opinion, that a couple of analyzed genes, more in particular MMP9 and CD163, although clearly induced by MgCl2, did not undergo a further up-regulation upon ZA addition but anyway maintained a high mRNA expression level. Interestingly, both these antigens are M2 macrophage polarization markers. Several authors have stated that osteoclast phenotype is very close to that of M1 polarized macrophages. An interpretation of our data might be that MgCl2 favors the differentiation of variant osteoclasts, characterized by an M2 phenotype, rendering them different from the typical osteoclasts, exhibiting instead a classical M1 phenotype. The presence of VD3, which is endowed with a known capacity to promote M2 macrophage polarization, could perhaps contribute to the appearance of this phenotype. In light of the recognized involvement of M2 macrophage polarization in the process of post inflammation / infection tissue repair [39,40], we think that the biological feature under consideration might represent an important advantage in the recovery from ONJ. Further investigation is, of course, necessary to confirm this hypothesis and to clarify a number of other issues concerning our observation such as, for example, how MgCl2 interact with other BPs or even non – BP anti – osteoclast drugs.
5. Conclusions
The conclusions of our work is that, in addition to their capacity to induce osteoclast apoptosis, BPs are also characterized by a strong inhibition activity on osteoclast differentiation. This effect is not only abrogated by MgCl2 but even transformed, through a sort of paradox effect, in an opposite and striking potentiation of osteoclast differentiation. In our opinion these results open the possibility to use MgCl2 as a topic agent to cure or prevent ONJ although this circumstance needs, of course, to be properly verified in vivo.
6. Patents
The results presented in this manuscript have been previously used to file the Italian Patent entitled “Therapy of Osteonecrosis of the Jaw”, Inventors Alexis Grande, Patent Status Pending, Priority Number 102023000002301, Priority Date 10/02/2023, License International, Commercial Rights Exclusive, Availability Available; viewable at the link https://www.knowledge-share.eu/en/patent/therapy-of-osteonecrosis-of-the-jaw/
Author Contributions
Conceptualization, A.G., S.R. and T.Z.-M.; Methodology, A.G., S.R., R.P., F.L., F.G., L.C. and M.M.; Software, S.R., F.L. and M.M.; Validation, A.G., S.R., R.P., F.L., F.G., L.C. and M.M.; Formal Analysis, A.G., S.R., F.L. and M.M.; Investigation, A.G., S.R., R.P., F.L., R.P., F.G. and L.C.; Resources, A.G.; Data Curation, A.G., S.R. and M.M.; Writing – Original Draft Preparation, A.G., T.Z.-M., V.Z. and. A.A.; Writing – Review & Editing, A.G., T.Z-M., V.Z. and A.A.; Visualization, A.G., T.Z-M., V.Z. and A.A.; Supervision, A.G., A.A.; Project Administration, A.G.; Funding Acquisition, A.G. All authors have read and agreed to the published version of the manuscript.++
Funding
We would like to thank the Department of Biomedical, Metabolic and Neural Sciences, University of Modena and Reggio Emilia for providing the financial support (FAR project).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cremers, S.; Drake M., T.; Ebetino, F.; Bilezikian J., P. ; R. Graham G. Russell. Pharmacology of Bisphosphonates. Br J Clin Pharmacol 2019, 85, 1052–62. [Google Scholar] [CrossRef]
- An, Y.; Zhao, J. Functionalized Selenium Nanotherapeutics Synergizes With Zoledronic Acid to Suppress Prostate Cancer Cell Growth Through Induction of Mitochondria-Mediated Apoptosis and Cell Cycle S Phase Arrest. Front. Oncol. 6857. [Google Scholar] [CrossRef]
- Wang, B.; Zhan, Y.; Yan, L.; Hao, D. How Zoledronic Acid Improves Osteoporosis by Acting on Osteoclasts. Front. Pharmacol. 9619. [Google Scholar] [CrossRef]
- Wang, L.; Fang, D.; Xu, J.; Luo, R. Various Pathways of Zoledronic Acid against Osteoclasts and Bone Cancer Metastasis: A Brief Review. BMC Cancer, 1059. [Google Scholar] [CrossRef]
- Gong, L.; Altman, R. B.; Klein, T. E. Bisphosphonates Pathway. Pharmacogenet 2011, 21, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, D. B. Mechanism of Action, Pharmacokinetic and Pharmacodynamic Profile, and Clinical Applications of Nitrogen-Containing Bisphosphonates. J. Dent. Res. 2007, 86, 1022–33. [Google Scholar] [CrossRef] [PubMed]
- Troeltzsch, M.; Woodlock, T.; Kriegelstein, S.; Steiner, T.; Messlinger, K.; Troeltzsch, M. Physiology and Pharmacology of Nonbisphosphonate Drugs Implicated in Osteonecrosis of the Jaw. J Can Dent Assoc.
- Moriyama, Y.; Nomura, M. Clodronate: A Vesicular ATP Release Blocker. Trends in Pharmacological Sciences 2018, 39, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Kenkre, J. S.; Bassett, J. The Bone Remodelling Cycle. Ann. Clin. Biochem. 2018, 55, 308–27. [Google Scholar] [CrossRef] [PubMed]
- Khan, A. A.; Morrison, A.; Hanley, D. A.; Felsenberg, D.; McCauley, L. K.; O’Ryan, F.; Reid, I. R. Diagnosis and Management of Osteonecrosis of the Jaw: A Systematic Review and International Consensus. J. Bone Miner. Res. 2015; 30. [Google Scholar] [CrossRef]
- Hoefert, S.; Schmitz, I.; Weichert, F.; Gaspar, M.; Eufinger, H. Macrophages and Bisphosphonate-Related Osteonecrosis of the Jaw (BRONJ): Evidence of Local Immunosuppression of Macrophages in Contrast to Other Infectious Jaw Diseases. Clin Oral Investig 2015, 19, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Marx, R. E. Pamidronate (Aredia) and Zoledronate (Zometa) Induced Avascular Necrosis of the Jaws: A Growing Epidemic. J. Oral Maxillofac. Surg. 2003, 61, 1115–17. [Google Scholar] [CrossRef] [PubMed]
- Nicolatou-Galitis, O.; Kouri, M.; Papadopoulou, E.; Vardas, E.; Galiti, D. ; Epstein,J. B.; Elad, S. et al. Osteonecrosis of the Jaw Related to Non-Antiresorptive Medications: A Systematic Review. Support Care Cancer. [CrossRef]
- Ruggiero, S. L.; Dodson, T. B.; Aghaloo, T.; Carlson, E. R.; Ward, B. B.; Kademani, D. American Association of Oral and Maxillofacial Surgeons’ Position Paper on Medication-Related Osteonecrosis of the Jaws—2022 Update. J. Oral Maxillofac. Surg 2022, 80, 920–43. [Google Scholar] [CrossRef]
- Aghaloo, T.; Hazboun, R.; Tetradis, S. Pathophysiology of Osteonecrosis of the Jaws. Oral Maxillofac Surg Clin North Am 2015, 27, 489–96. [Google Scholar] [CrossRef]
- Chang, J.; Hakam, A. E.; McCauley L., K. Current Understanding of the Pathophysiology of Osteonecrosis of the Jaw. Curr. Osteoporos. Rep. 2018, 16, 584–95. [Google Scholar] [CrossRef]
- Otto, S.; Tröltzsch, M.; Jambrovic, V.; Panya, S.; Probst, F.; Ristow, O.; Ehrenfeld, M.; Pautke, C. Tooth Extraction in Patients Receiving Oral or Intravenous Bisphosphonate Administration: A Trigger for BRONJ Development? J Craniomaxillofac Surg 2015, 43, 847–54. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Yamori, M.; Ishizaki, T.; Asai, K.; Goto, K.; Takahashi, K.; Nakayama, T.; Bessho, K. Increased Incidence of Osteonecrosis of the Jaw after Tooth Extraction in Patients Treated with Bisphosphonates: A Cohort Study. Int J Oral Maxillofac Surg 2012, 41, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, M.; Kuroshima, S.; Sawase, T. Clinical Considerations for Medication-Related Osteonecrosis of the Jaw: A Comprehensive Literature Review. Int. J. Implant Dent. [CrossRef]
- Asagiri, M.; Takayanagi, H. The Molecular Understanding of Osteoclast Differentiation. Bone 2007, 40, 251–64. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-L.; Huang, L.-Y.; Cheng, Y.-T.; Li, F.; Zhou, Q.; Wu, C.; Shi, Q.-H.; Guan, Z.-Z.; Liao, J.; Hong, W. Zoledronic Acid Inhibits Osteoclast Differentiation and Function through the Regulation of NF-ΚB and JNK Signalling Pathways. Int. J.Mol. Med. 2019, 44, 582–92. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Yu, W.; Zhao, H.; Su, J.; Sheng, Q. Skeletal Site-Specific Effects of Zoledronate on in Vivo Bone Remodeling and in Vitro BMSCs Osteogenic Activity. Sci. Rep. 3612. [Google Scholar] [CrossRef]
- Fliefel, R.; Tröltzsch, M.; Kühnisch, J.; Ehrenfeld, M.; Otto, S. Treatment Strategies and Outcomes of Bisphosphonate-Related Osteonecrosis of the Jaw (BRONJ) with Characterization of Patients: A Systematic Review. Int J Oral Maxillofac Surg 2015, 44, 568–85. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Morrison, A.; Cheung, A.; Hashem, W.; Compston, J. Osteonecrosis of the Jaw (ONJ): Diagnosis and Management in 2015. Osteoporos Int 2016, 27, 853–59. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, S.; Soutome, S.; Yanamoto, S.; Fujita, S.; Hasegawa, T.; Komori, T.; Kojima, Y. Evaluation of the Treatment Strategies for Medication-Related Osteonecrosis of the Jaws (MRONJ) and the Factors Affecting Treatment Outcome: A Multicenter Retrospective Study with Propensity Score Matching Analysis. J. Bone Miner. Res. 2017; 32. [Google Scholar] [CrossRef]
- Rupel, K.; Ottaviani, G. ; Gobbo, M; Contardo, L.; Tirelli, G.; Vescovi, P.; Di Lenarda, R.; Matteo Biasotto. A Systematic Review of Therapeutical Approaches in Bisphosphonates-Related Osteonecrosis of the Jaw (BRONJ). Oral Oncol. 1049. [Google Scholar] [CrossRef]
- Anesi, A.; Generali, L.; Sandoni, L.; Pozzi, S. ; Grande. From Osteoclast Differentiation to Osteonecrosis of the Jaw: Molecular and Clinical Insights. Int. J. Mol. Sci. 4925. [Google Scholar] [CrossRef]
- Boyle, W. J.; Scott Simonet, W.; Lacey, D. L. Osteoclast Differentiation and Activation. Nature 2003, 423, 337–42. [Google Scholar] [CrossRef] [PubMed]
- Parenti, S.; Sandoni, L.; Montanari, M.; Zanocco-Marani, T.; Anesi, A.; Iotti, S.; Manfredini, R.; Frassineti, C.; Davalli, P.; Grande, A. Magnesium Favors the Capacity of Vitamin D3 to Induce the Monocyte Differentiation of U937 Cells. Magnes. Res. 2021, 34, 114–29. [Google Scholar] [CrossRef]
- Mammoli, F.; Castiglioni, S.; Parenti, S.; Cappadone, C.; Farruggia, G.; Iotti, S.; Davalli, P.; Maier, J.; Grande, A.; Frassineti, C. Magnesium Is a Key Regulator of the Balance between Osteoclast and Osteoblast Differentiation in the Presence of Vitamin D3. Int. J. Mol. Sci. [CrossRef]
- Amoui, M.; Suhr, S.-M.; Baylink, D. J.; Lau, K.-H. W. An Osteoclastic Protein-Tyrosine Phosphatase May Play a Role in Differentiation and Activity of Human Monocytic U-937 Cell-Derived, Osteoclast-like Cells. Am. J. Physiol., Cell Physiol. [CrossRef]
- Takahashi, N.; Udagawa, N.; Suda, T. Vitamin D Endocrine System and Osteoclasts. BoneKEy Rep. [CrossRef]
- Li, P.; Zhao, Z.; Wang, L.; Jin,X. ; Shen, Y.; Nan, C.; Liu, H. Minimally Effective Concentration of Zoledronic Acid to Suppress Osteoclasts in Vitro. Exp. Ther. Med. 2018, 15, 5330–36. [Google Scholar] [CrossRef]
- Wu, L.; Feyerabend, F.; Schilling, A. F.; Willumeit-Römer, R.; Luthringer, B. J. C. Effects of Extracellular Magnesium Extract on the Proliferation and Differentiation of Human Osteoblasts and Osteoclasts in Coculture. Acta Biomater 2015, 27, 294–304. [Google Scholar] [CrossRef]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; et al. Autoamplification of NFATc1 Expression Determines Its Essential Role in Bone Homeostasis. Exp. Biol. Med. (Maywood) 2005, 202, 1261–69. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T. Regulators of Osteoclast Differentiation and Cell-Cell Fusion. Keio J Med 2011, 60, 101–5. [Google Scholar] [CrossRef] [PubMed]
- Pelekanou, V.; Villarroel-Espindola, F.; Schalper, K. A.; Pusztai, L.; Rimm, D. L. CD68, CD163, and Matrix Metalloproteinase 9 (MMP-9) Co-Localization in Breast Tumor Microenvironment Predicts Survival Differently in ER-Positive and -Negative Cancers. Breast Cancer, 20. [CrossRef]
- Kim, K.; Kim J., A.; Lee, J. J.; Jin, H. J.; Kook, H.; Kim, K. K.; Lee, S. Y.; Kim, N. MafB Negatively Regulates RANKL-Mediated Osteoclast Differentiation. Blood 2007, 109, 3253–59. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Kou, X.; Luo, Q.; Yang, R.; Liu, D.; Wang, X.; Song, Y.; et al. Enhanced M1/M2 Macrophage Ratio Promotes Orthodontic Root Resorption. J. Dent. Res. 2015, 94, 129–39. [Google Scholar] [CrossRef] [PubMed]
- 40. Fukui, Shoichi, Naoki Iwamoto, Ayuko Takatani, Takashi Igawa, Toshimasa Shimizu, Masataka Umeda, Ayako Nishino, et al.. M1 and M2 Monocytes in Rheumatoid Arthritis: A Contribution of Imbalance of M1/M2 Monocytes to Osteoclastogenesis. Frontiers in Immunology, 1958. [CrossRef]
- Yang, G.; Chen, X.; Yan, Z.; Zhu, Q.; Yang, C. CD11b Promotes the Differentiation of Osteoclasts Induced by RANKL through the Spleen Tyrosine Kinase Signalling Pathway. J. Cell. Mol. Med. 2017, 21, 3445–52. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Li, X.; Xia, Y.; Yu, Z.; Cai, N.; Malwal, S. R.; Han, X.; Oldfield, E.; Zhang, Y. Farnesyl Pyrophosphate Synthase as a Target for Drug Development: Discovery of Natural-Product-Derived Inhibitors and Their Activity in Pancreatic Cancer Cells. J. Med. Chem. 2019, 62, 10867–96. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G. A. Bone Mass Homeostasis and Bisphosphonate Action. Bone 1997, 20, 1–4. [Google Scholar] [CrossRef]
Figure 1.
Proliferative and apoptotic effects exerted by scalar concentrations of Zoledronate on U937 cells. U937 cells underwent treatment with scalar concentrations of Zoledronate (ZA) ranging from 0 to 100 μM for up to 4 days. Upper panel shows the results of a daily cell count represented as growth curve. The histogram of middle panel depicts the percentage of apoptotic cells, and cells distributed in the various cell cycle phases, as assessed by flow cytometry analysis performed at day 4 of the experiment. Lower panel presents a histogram indicating the mRNA expression levels of Cyclin D3 and p21 genes, estimated by QRT-PCR analysis at day 4 of experiment. Statistical analysis was always carried out by a Student’s t test, performed on pairwise comparisons between control untreated cells and cells treated with each tested ZA concentration.
Figure 1.
Proliferative and apoptotic effects exerted by scalar concentrations of Zoledronate on U937 cells. U937 cells underwent treatment with scalar concentrations of Zoledronate (ZA) ranging from 0 to 100 μM for up to 4 days. Upper panel shows the results of a daily cell count represented as growth curve. The histogram of middle panel depicts the percentage of apoptotic cells, and cells distributed in the various cell cycle phases, as assessed by flow cytometry analysis performed at day 4 of the experiment. Lower panel presents a histogram indicating the mRNA expression levels of Cyclin D3 and p21 genes, estimated by QRT-PCR analysis at day 4 of experiment. Statistical analysis was always carried out by a Student’s t test, performed on pairwise comparisons between control untreated cells and cells treated with each tested ZA concentration.
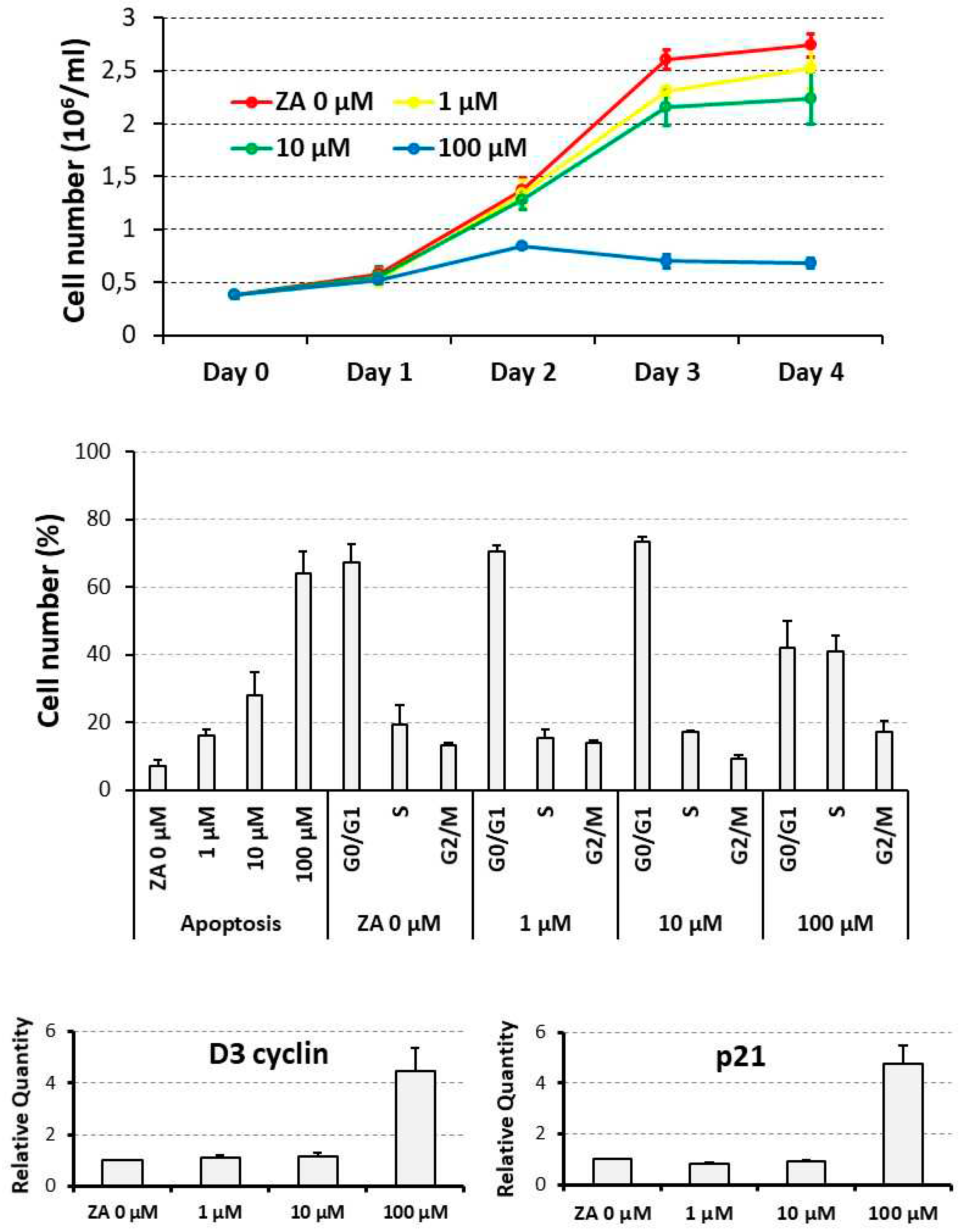
Figure 2.
Proliferative and apoptotic effects exerted by a supra-physiological concentration of MgCl2 on U937 cells. U937 cells were exposed to a 10 mM concentration of MgCl2 and then subjected to a cell count at 2 day intervals (upper panel), a flow cytometry analysis, at day 4 of the experiment, to assess the percentage of apoptotic cells, and cells distribute in the various cell cycle phases (middle panel), and a QRT-PCR analysis, always at day 4 of the experiment, to evaluate the transcript levels of D3 cyclin and p21 genes (lower panel). All the results have been presented as histograms. Ctr, control, untreated cells; Mg, cells treated with MgCl2.
Figure 2.
Proliferative and apoptotic effects exerted by a supra-physiological concentration of MgCl2 on U937 cells. U937 cells were exposed to a 10 mM concentration of MgCl2 and then subjected to a cell count at 2 day intervals (upper panel), a flow cytometry analysis, at day 4 of the experiment, to assess the percentage of apoptotic cells, and cells distribute in the various cell cycle phases (middle panel), and a QRT-PCR analysis, always at day 4 of the experiment, to evaluate the transcript levels of D3 cyclin and p21 genes (lower panel). All the results have been presented as histograms. Ctr, control, untreated cells; Mg, cells treated with MgCl2.
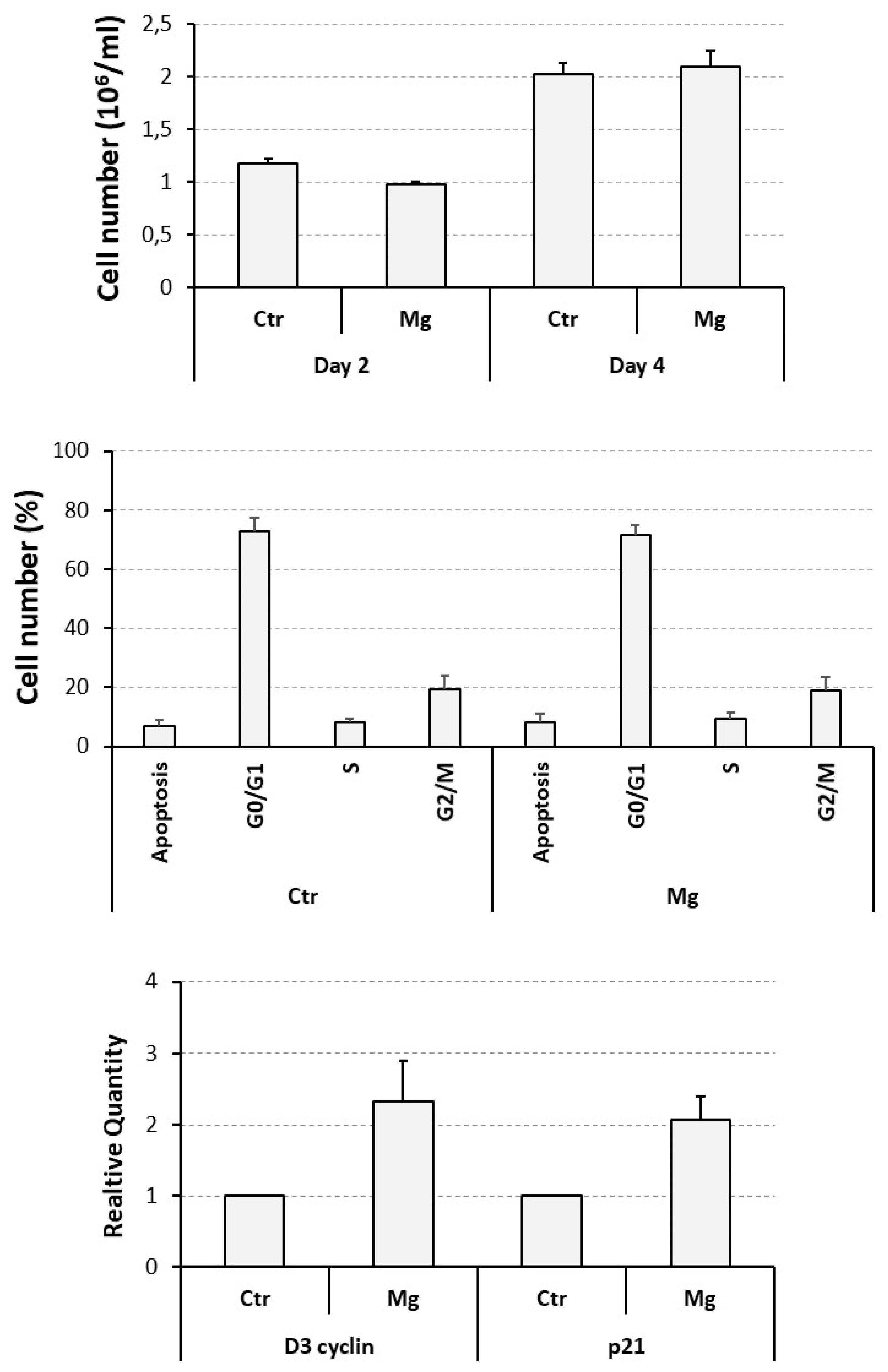
Figure 3.
Effect of ZA and MgCl2 on the mRNA expression of osteoclast differentiation markers in osteoclasts derived from U937 cells upon stimulation with PMA and VD3. Analysis was performed by QRT-PCR. In each histogram expression levels are expressed on the y axis as “Relative Quantity” (RQ), represented as mean ± S.E.M. The x-axis shows the different treatments: None (Ctr), Zoledronate (ZA), MgCl2 (Mg) or both ZA + Mg. Analyzed genes are listed on the top of each histogram.
Figure 3.
Effect of ZA and MgCl2 on the mRNA expression of osteoclast differentiation markers in osteoclasts derived from U937 cells upon stimulation with PMA and VD3. Analysis was performed by QRT-PCR. In each histogram expression levels are expressed on the y axis as “Relative Quantity” (RQ), represented as mean ± S.E.M. The x-axis shows the different treatments: None (Ctr), Zoledronate (ZA), MgCl2 (Mg) or both ZA + Mg. Analyzed genes are listed on the top of each histogram.
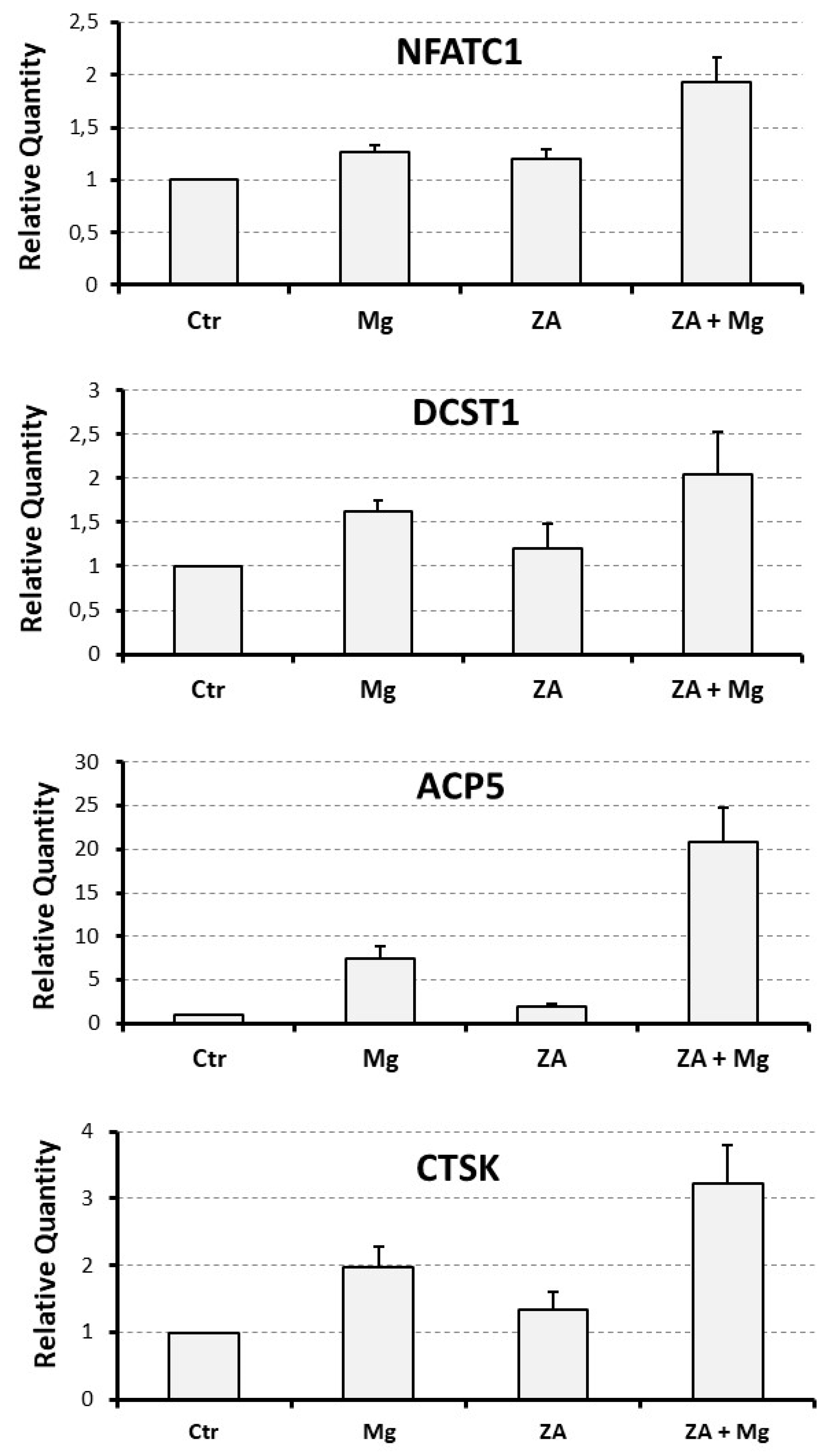
Figure 4.
Effect of Za and MgCl2 on the mRNA expression of monocyte / macrophage differentiation markers in osteoclasts derived from U937 cells upon stimulation with PMA and VD3. Analysis was performed by QRT-PCR. In each histogram expression levels are expressed on the y axis as “Relative Quantity” (RQ), represented as mean ± S.E.M. The x-axis shows the different treatments: None (Ctr), Zoledronate (ZA), MgCl2 (Mg) or both (ZA + Mg). Analyzed genes are listed on the top of each histogram.
Figure 4.
Effect of Za and MgCl2 on the mRNA expression of monocyte / macrophage differentiation markers in osteoclasts derived from U937 cells upon stimulation with PMA and VD3. Analysis was performed by QRT-PCR. In each histogram expression levels are expressed on the y axis as “Relative Quantity” (RQ), represented as mean ± S.E.M. The x-axis shows the different treatments: None (Ctr), Zoledronate (ZA), MgCl2 (Mg) or both (ZA + Mg). Analyzed genes are listed on the top of each histogram.
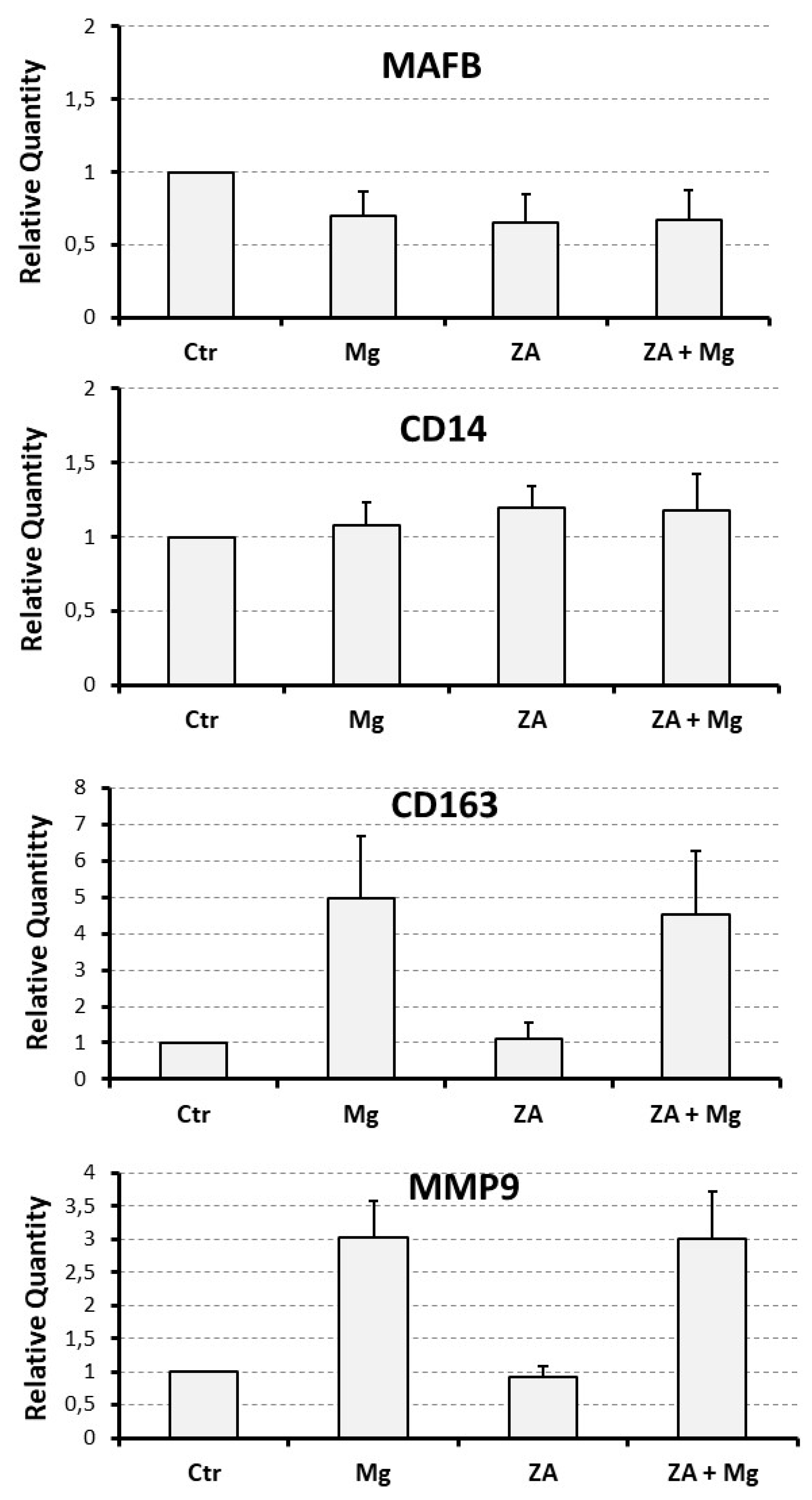
Figure 5.
Analysis of the myeloid differentiation marker CD11b performed upon treatment with ZA, MgCl2, or both in osteoclasts derived from U937 cells obtained by stimulation with PMA and VD3. Expression of the CD11b surface antigen was assessed by flow cytometry. The upper panel is a histogram showing on the y axis the Mean Fluorescent Intensity (MFI) measured following staining with and anti - CD11b PE antibody. The x axis shows the different treatments received by U937 derived osteoclasts: None (Ctr), MgCl2 (Mg), Zoledronate (ZA) or both ZA + Mg. The lower panel is a histogram showing the flow cytometry results of a representative experiment. Y axis shows the percentage of cells, while x axis shows the intensity of fluorescence. The treatments are highlighted by different colors. Ctr- refers to undifferentiated U937 cells, that were not stimulated with PMA and VD3, while Ctr+ are differentiated U937 cells that were stimulated with PMA and VD3 but didn’t receive ZA or Mg treatment.
Figure 5.
Analysis of the myeloid differentiation marker CD11b performed upon treatment with ZA, MgCl2, or both in osteoclasts derived from U937 cells obtained by stimulation with PMA and VD3. Expression of the CD11b surface antigen was assessed by flow cytometry. The upper panel is a histogram showing on the y axis the Mean Fluorescent Intensity (MFI) measured following staining with and anti - CD11b PE antibody. The x axis shows the different treatments received by U937 derived osteoclasts: None (Ctr), MgCl2 (Mg), Zoledronate (ZA) or both ZA + Mg. The lower panel is a histogram showing the flow cytometry results of a representative experiment. Y axis shows the percentage of cells, while x axis shows the intensity of fluorescence. The treatments are highlighted by different colors. Ctr- refers to undifferentiated U937 cells, that were not stimulated with PMA and VD3, while Ctr+ are differentiated U937 cells that were stimulated with PMA and VD3 but didn’t receive ZA or Mg treatment.
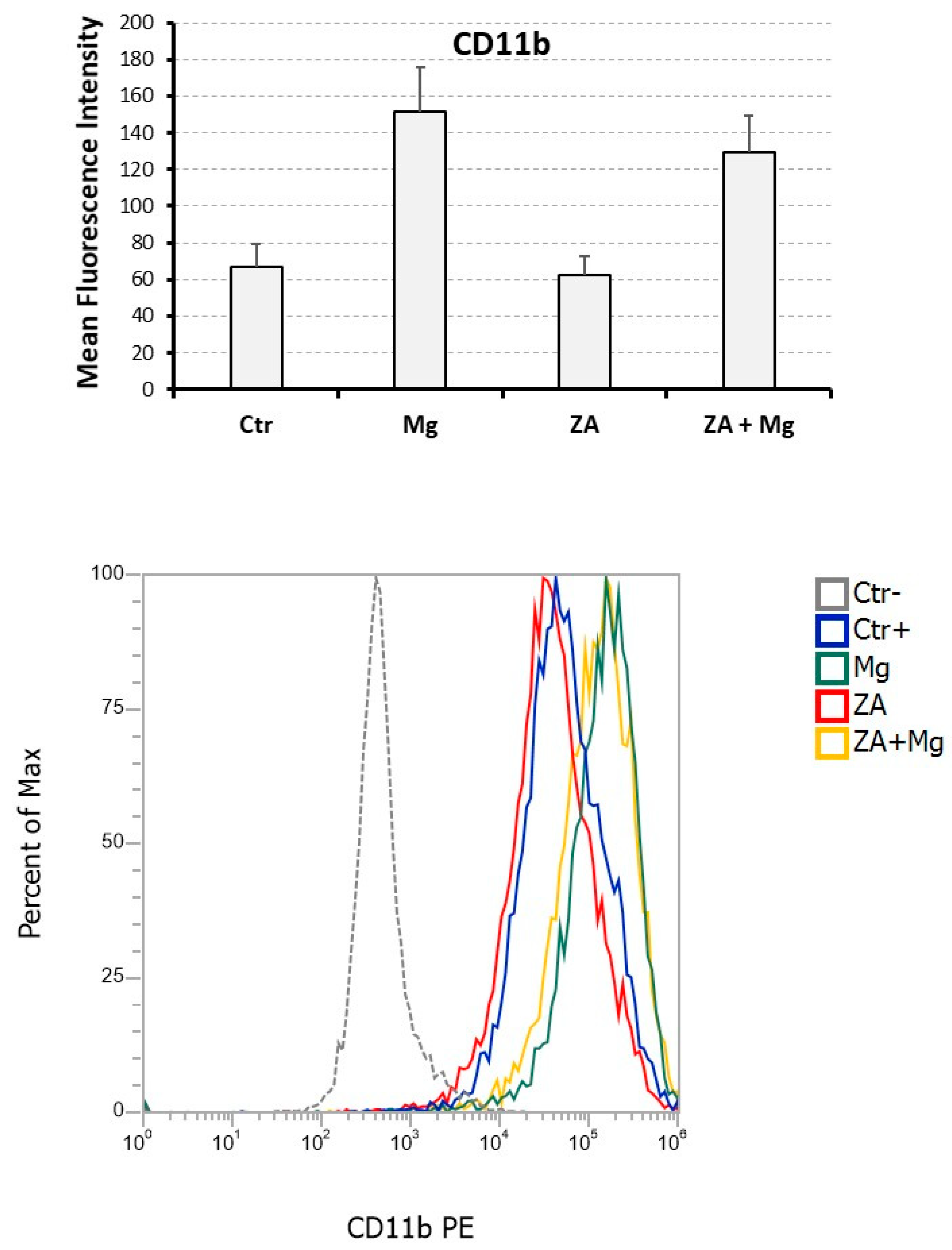
Figure 6.
Morphological analysis performed upon treatment with ZA, MgCl2, or both in osteoclasts derived from U937 cells obtained by stimulation with PMA and VD3. U937 cells, under the same experimental conditions of previous figures (Figures 3 to 5), were subjected to cyto-centrifugation, May-Grunwald-Giemsa staining and microscope examination. Ctr represents osteoclasts derived from U937 cells stimulated with PMA and VD3, that did not receive further treatments. Treated samples were instead exposed to Zoledronate (ZA), MgCl2 (Mg) or both (ZA + Mg).
Figure 6.
Morphological analysis performed upon treatment with ZA, MgCl2, or both in osteoclasts derived from U937 cells obtained by stimulation with PMA and VD3. U937 cells, under the same experimental conditions of previous figures (Figures 3 to 5), were subjected to cyto-centrifugation, May-Grunwald-Giemsa staining and microscope examination. Ctr represents osteoclasts derived from U937 cells stimulated with PMA and VD3, that did not receive further treatments. Treated samples were instead exposed to Zoledronate (ZA), MgCl2 (Mg) or both (ZA + Mg).
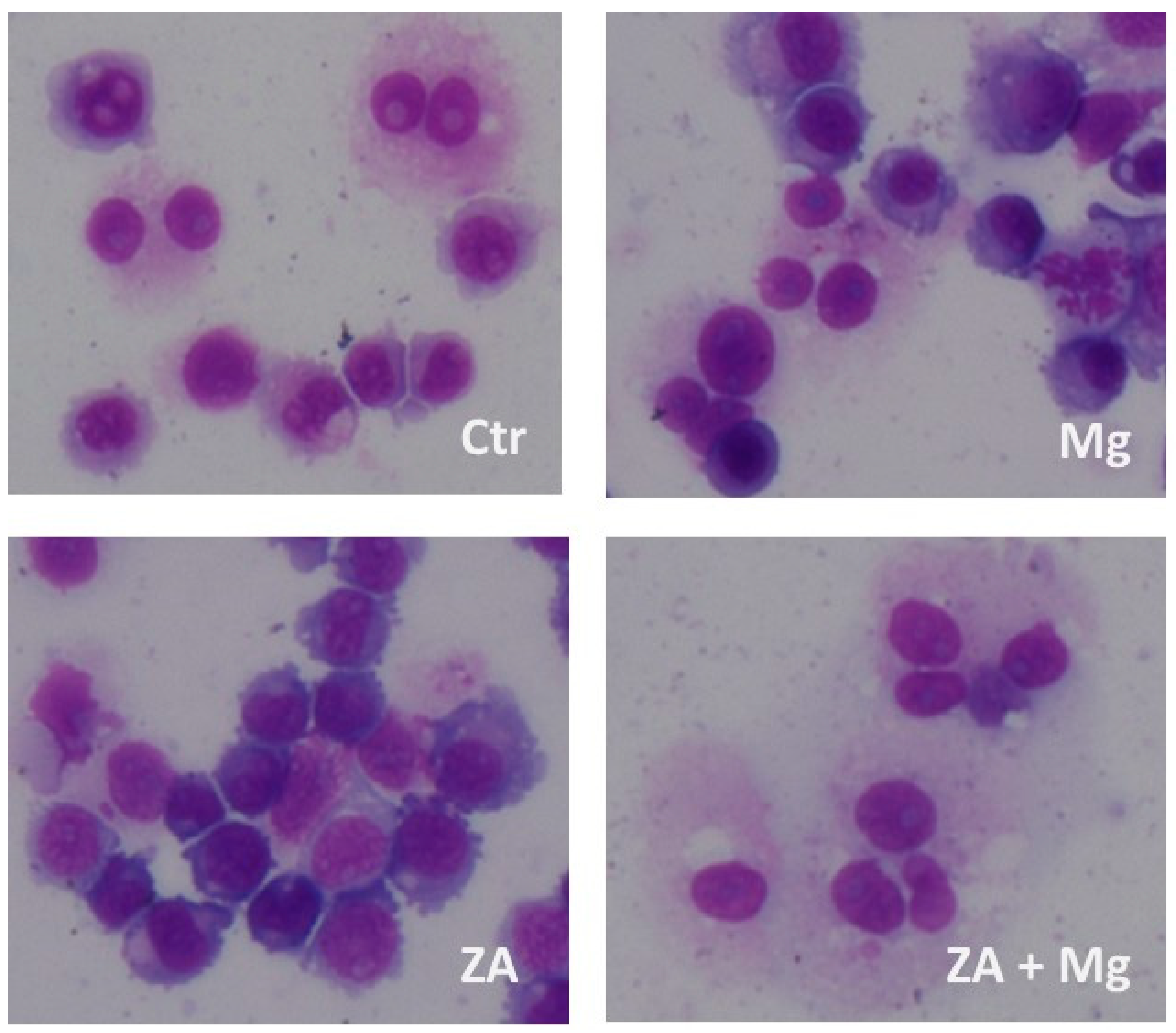

Table 1.
Numerical values and statistical data of the molecular analysis performed by QRT-PCR on different osteoclast and monocyte – macrophage differentiation markers.
Table 1.
Numerical values and statistical data of the molecular analysis performed by QRT-PCR on different osteoclast and monocyte – macrophage differentiation markers.
| Analyzed Marker |
Ctr | Mg | ZA | ZA + Mg | Anova, p value |
|---|---|---|---|---|---|
| NFATC1 | 1 | 1.3 ± 0.1 | 1.2 ± 0.1 | 1.9 ± 0.2 | 0.005 |
| DCST1 | 1 | 1.6 ± 0,1 | 1.2 ± 0.3 | 2.1 ± 0.5 | 0.3 |
| ACP5 | 1 | 7.5 ± 1.4 | 2.0 ± 0.3 | 20.9 ± 4.0 | 0.002 |
| CTSK | 1 | 2.0 ± 0.3 | 1.3 ± 0.3 | 3.2 ± 0.6 | 0.01 |
| MMP9 | 1 | 3.0 ± 0.6 | 0.9 ± 0.2 | 3.0 ± 0.7 | 0.01 |
| MAFB | 1 | 0.7 ± 0.2 | 0.7 ± 0.2 | 0.7 ± 0.2 | 0.4 |
| CD14 | 1 | 1.1 ± 0.2 | 1.2 ± 0.2 | 1.1 ± 0.3 | 0.8 |
| CD163 | 1 | 5.0 ± 1.7 | 1.1 ± 0.4 | 4.5 ± 1.7 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



