
Submitted:
16 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
Over the past several decades, the extensive use of pyrethroids has led to the development of resistance in many insect populations, including the economically damaging pest tarnished plant bug (TPB), Lygus lineolaris on cotton. To manage TPB resistance, several commercially formulated pyrethroid-containing binary mixtures, in combination with neonicotinoids or avermectin are recommended for TPB control and resistance management in the Mid-south USA. This study aimed to evaluate the toxicity and resistance risks of four formulated pyrethroid-containing binary mixtures (Endigo, Leverage, Athena, and Hero) on one susceptible and two resistant TPB populations which were field-collected in July (Field-R1) and October (Field-R2), respectively. Based on LC50 values, both resistant TPB populations displayed variable tolerance to the four binary mixtures, with Hero showing the highest resistance and Athena the lowest. Notably, the Field-R2 exhibited 1.5-3-fold higher resistance compared to the Field-R1 for all four binary insecticides. Moreover, both resistant TPB populations demonstrated significantly higher resistance ratios toward Hero and Leverage compared to their corresponding individual pyrethroid, while Endigo and Athena showed similar or lower resistance. This study also utilized the calculated additive index (AI) and co-toxicity coefficient (CTC) analysis, which revealed that the two individual components in Leverage exhibited antagonist effects against the two resistant TPB populations. In contrast, the two individual components in Endigo, Hero and Athena displayed synergistic interactions. Considering that Hero is a mixture of two pyrethroids which can enhance the development of TPB resistance, our findings suggest that Endigo and Athena are likely superior products for slowing down resistance development in TPB populations. This study provides valuable insight for selecting the most effective mixtures to achieve better TPB control through synergistic toxicity analysis, while simultaneously reducing economic and environmental risks associated with resistance development in the insect pest.
Keywords:
Lygus lineolaris
; pyrethroids binary mixtures
; bioassay
; resistance management
; synergism
; toxicity
1. Introduction
The tarnished plant bug (TPB) Lygus lineolaris (Palisot de Beauvois) (Hemiptera: Miridae) is a significant major pest in cotton-growing regions across the Mid-southern USA. With the eradication of the boll weevil and the adoption of Bt cotton to control lepidopteran pests, TPB infestations have increased yearly, resulting in severe damage to cotton fruiting buds [1]. Both TPB nymphs and adults use specialized piercing-sucking stylets and digestive enzymes to feed on cotton fruiting buds (squares) and small fruit (bolls), causing fruit abscission and damage to seeds and lint [2,3,4]. The prevalence of TPB adults is the highest during the pre-flowering stage of cotton, while nymphs are more commonly found during the flowering period. As a result, TPBs primarily inflict the most significant damage from the first square stage to the early flowering stages of cotton growth [2].
The use of insecticide has been the primary method for preventing and controlling TPB in the Mid-south cotton growing area, with a variety of classes employed such as organophosphates, carbamates, neonicotinoids, pyrethroids, insect growth regulators and sulfoximine [5,6,7,8,9]. Pyrethroids, which account for 30% of the global pesticide market, are synthetic insecticides based on natural pyrethrins found in Chrysanthemum flowers [10]. Over the past few decades, pyrethroids were extensively used to control agricultural crop pests and human disease vectors [11]. However, the overuse of pyrethroids led to the development of resistance in many insect pest populations, including the TPBs [12]. Pyrethroid resistance in TPBs collected from cotton was first reported in Mississippi in 1993 [13,14], likely the result of selective pressure from early season insecticide applications targeting lepidopteran pests when TPBs were present. By 1999, resistance to pyrethroids in TPB was widespread in the Mid-south region [15,16]. Pyrethroid resistance significantly reduced the effectiveness of chemical control and increased the cost and quantity of insecticides required to control this pest. Due to resistance development, pyrethroids are no longer recommended for TPB control in cotton in Mississippi [17]. To combat resistance, insecticide mixtures and rotation are proposed as important tools for resistance management. Currently, several commercially formulated pyrethroid binary mixtures in combination with neonicotinoids or avermectin are listed for TPB control and resistance management in Mississippi Delta region which include: Brigadier (bifenthrin +imidacloprid), Leverage (imidacloprid + β-cyfluthrin), Endigo (thiamethoxam + λ -cyhalothrin), Athena (bifenthrin + avermectin) and the two pyrethroid mixture Hero (bifenthrin +ζ-cypermethrin) (https://extension.msstate.edu/sites/default/files/ publications/publications/P2471_web.pdf ). Mixing insecticides with different modes of action is more effective in resistance management programs compared to rotational strategies [18]. Previously, mixtures consisted of a pyrethroid with carbamate [19]; or a pyrethroid with organophosphorus [20,21,22,23]. More recently, the development of neonicotinoid insecticides with reduced toxicity to human compared to previously used organophosphates and carbamates can now provide a broad spectrum to control numerous crop-damaging insects. Pyrethroids target insect voltage-gated sodium channels [11] and neonicotinoids act as agonists for the nicotinic acetylcholine receptors (nAChR), and both classes of insecticide impact the central nervous system of insects (Table 1) [24]. Mixtures of neonicotinoids and pyrethroids are useful for resistance management as highly effective tools against some of the world's most destructive crop pests [25]. Avermectin, on the other hand, allosterically activates glutamate-gated chloride channels (GluCls) in insect nerve and muscle cells, causing cell hyperpolarization, eventually resulting in insect paralysis and death [26]. Pyrethroids can also be mixed with avermectin and applied for insect pest control. Insecticide mixtures afford two key advantages: targeting a broad spectrum of pest species and managing pesticide resistance.
Insecticide resistance research in the past predominantly focused on individual insecticides. Testing on commercialized formulated mixtures is not well studied, especially in resistant TPB populations. Here, we conducted a comprehensive investigation using two resistant field TPB populations collected in July and October, respectively, from wild-host plants in Coahoma County, a cotton growing area in the northern Mississippi Delta region, USA. We conducted dose-response bioassays of four formulated pyrethroid-containing binary mixtures using a modified Potter spray tower. Additionally, we analyzed the potential interaction between the two individual components in the four binary mixtures on susceptible and two resistant TPB populations. Our findings provide valuable information for selecting the most effective mixtures to achieve better TPB control. By understanding the interactions between the individual components in the binary mixtures, we can optimize their use and develop targeted resistance management strategies.
2. Materials and Methods
2.1. Insect Populations
The laboratory susceptible TPB population (Lab-S) was collected from the hills and wooded area of Crossett, AR, where insecticide was infrequently used. This population is historically recognized as a susceptible location and was used in previous studies [8,15,27]. Lab-S TPB was reared on an artificial diet and without any exposure to insecticides, following the method outlined in Portilla et al. [28,29]. The resistant TPB population was collected from Coahoma County in July (Field-R1) and October 2022 (Field-R2),respectively, located in the northern Delta cotton growing region of Mississippi. For the bioassays, the TPB populations were maintained under laboratory conditions (27 ± 2 °C, 65 ± 10% RH and a 12:12 h (L:D) photoperiod) prior to and during the experiments.
2.2. Insecticides
The insecticides used in this study were all formulated and include: Endigo 2,06ZC (Syngenta), Warrior II (Syngenta), Centric 40 WG (Syngenta), Leverage 360 EC (Bayer Crop Science), Baythroid XL (Bayer), Advise ®Four (Winfield), Hero 1.24 (FMC), Mustang® Maxx (FMC), Tundra® EC (Winfield), Athena (FMC) and Epi-Mek (Agri-Mek 0.15EC, Syngenta). Insecticides were purchased from local agricultural suppliers near Leland, MS with the manufacturers and mode of action given in Table 1.
2.3. Laboratory spray tower bioassays
The TPB adults from the lab-S (7-9 d old) or field-R mixed age populations were placed into plastic cups (500 mL round wide-mouth polypropylene cup DxH: 9.3x10 cm, 20 TPBs/cup) with fabric mesh covered holes (5.0 cm in diameter) cut in both the lid and bottom. A modified spray tower was constructed with Plexiglass to fit into a fume hood, incorporated an original spray nozzle of a Potter Spray Tower (Burkard Scientific Ltd, Uxbridge, U.K.) and supplied nearly the same air pressure (69 kPa or 10 psi) and insecticide delivery (spray distance= 22 cm) as those in the Potter Spray Tower. Pesticide solutions were dissolved in deionized H2O and diluted serially to obtain the five desired concentrations. Two 7-8 cm long whole green beans were placed at the bottom of each cup, and 0.5 mL pesticide solution was sprayed into the cup to cover the inner wall, green beans, and TPB adults. Prior to spraying insecticide solutions, a control was conducted by spraying 0.5 mL of deionized water. After treatment, the caged TPB adults were placed in an environmental incubator at 27 ± 2°C, 65 ± 10% RH, and a 12:12 (L:D) photoperiod. Mortality was determined 48 hours after the spray treatment. The TPB adults were considered dead if they were unable to walk or fly. Each treatment was replicated three or four times (for each concentration of insecticide) with 60-80 adults per concentration. Five concentrations of each insecticide were used to determine the LC50 values and 95% confidence intervals.
2.4. Evaluation of potential interaction of two individual component in formulated binary mixtures
To analyze the potential interaction of two individual components in four binary insecticides, the synergistic, additive or antagonistic effects in four formulated binary mixtures (Endigo, Leverage, Hero and Athena) was assessed using Marking’s additive index (AI) methods [30] and Sun and Johnson [31] methods, which based on the LC50 values from individual pesticide and their mixture to characterize the interactions of pesticides. The AI was adopted to assess binary toxicity as follows:
where S represents the sum of the toxicity of pesticides A and B; Am indicates the LC50 of pesticide A in mixture; Ai is the LC50 of individual pesticide A; Bm indicates the LC50 of pesticide B in mixture; Bi is the LC50 of individual pesticide B.
The AI value was determined from the sum of S based on the appropriate formulas as follows:
AI = for S < 1.0 and AI = 1- S for S ≥ 1.0
Binary toxicities were classified as antagonistic effect (AI ≤ -0.2), additive (-0.2 < AI ≤ 0.25) or synergistic effect (AI > 0.25) accordingly. The greater the AI value, the greater the pesticide synergy [32].
Sun and Johnson methods[31]:
(1). Toxicity Index (T.I.) (using A as standard, A and B are individual components in formulated mixture) T.I. of A=100; T.I. of B=ALC50/BLC50 x100.
(2). Actual Toxicity Index (ATI) of mixture (using A as standard) ATI=LC50 of A/ LC50 of (A+B) x 100.
(3). Theoretical Toxicity Index (TTI) of mixture: TTI= T.I. of A x % of A in mixture + T.I. of B x % of B in mixture. From the actual and theoretical toxicity of the mixture, the mixture toxicity can be calculated by the following CTC equation.
(4) CTC (co-toxicity coefficient): CTC=ATI/TTI x 100; When the co-toxicity coefficient of the formulated mixture is 100, the effect of this mixture indicates the probability of similar action. If the mixture gives a coefficient significantly greater than 100, it indicates a synergistic action.
2.5. Data analysis
For the bioassay data, LC50 values and 95% confidence intervals were calculated by Probit analysis using SPSS software (version 19.0, SPSS Inc., Chicago, Illinois, USA, 2003), and data were presented as the means ± S.D. If there was no overlap in the 95% confidence intervals, LC50 values of different populations were considered significantly different. Resistance ratios (RR) were calculated as the ratio of LC50 value of the resistant populations (Field-R1/Field-R2) to that of the lab-S TPB populations. The comparison of resistance ratio (RR) was plotted using JMP software with one-way analysis of variance followed by Tukey’s HSD was used to determine statistically significant differences (P < 0.05).
3. Results
3.1. Toxicity of the four formulated mixtures against the laboratory susceptible strain
Four pyrethroid-containing formulated binary insecticides (Endigo, Leverage, Athena, and Hero) were applied to Lab-S and two field-collected resistant TPB populations (Field-R1 & Field-R2). Following exposure to spray treatment for 48 hours, the LC50 values for Endigo, Leverage, Hero, and Athena in Lab-S TPB were 22.54, 20.53, 20.39, and 73.60 µg/mL, respectively (Table 2).
3.2. Toxicity of formulated mixtures against the two field-collected resistant TPB populations
In contrast, the Field-R1 TPB population collected from Coahoma County, MS in July 2022 displayed significantly higher resistance to the four binary insecticides. The LC50 values for this population were significantly elevated to 166.64, 273.44, 329.96, and 479.60 µg/mL for Endigo, Leverage, Hero, and Athena, respectively (Table 2). The corresponding resistance ratios (RR) for each binary mixture were 7.39-fold for Endigo, 13.3-fold for Leverage, 16.2-fold for Hero and 6.56-fold for Athena (Table 2, Figure 1A). Among the four binary insecticides, Hero exhibited the highest RR, while Athena showed the lowest RR (Table 2, Figure 1A).
Additionally, another Field-R2 TPB population collected from Coahoma County, MS, in October 2022 also demonstrated increased resistance to the four binary insecticides. The LC50 values for this population were significantly elevated to 315.18, 513.94, 970.67, and 708.30 µg/mL for Endigo, Leverage, Hero, and Athena, respectively (Table 2). The corresponding resistance ratios (RR) for each binary mixture were 13.98-fold for Endigo, 25.0-fold for Leverage, and 47.60-fold for Hero and 9.62-fold for Athena (Table 2, Figure 1B). Similar to the Field-R1 TPB population, Hero exhibited the highest RR, while Athena displayed the lowest RR in Field-R2 TPB as well (Table 2, Figure 1B). Furthermore, when comparing the Field-R2 TPB population collected in October with the Field-R1 collected in July, Endigo and Leverage showed approximately 2-fold higher resistance, Hero’s resistance increased up to 3-fold, whereas Athena only displayed 1.4-fold increase in resistance (Table 2, Figure 1).
3.3. Toxicity comparison of the individual pyrethroid with the binary mixture
The bioassay of the corresponding individual components and binary mixture insecticides were examined simultaneously. For the Field-R1 TPB resistant population, both Leverage and Hero exhibited significant higher resistance levels compared to the corresponding individual pyrethroids present in their formulations (Figure 1A) [33]. In contrast, Endigo and Athena displayed similar resistance levels to the corresponding individual pyrethroid (Figure 1A) [33].
Similarly, for the Field-R2 TPB population, Endigo showed a similar resistance level to the individual λ-cyhalothrin (Figure 1B). However, Leverage and Hero displayed significantly higher resistance levels compared to their respective individual pyrethroids (unpublished data). On the other hand, Athena exhibited a significant lower resistance level to the individual pyrethroid bifenthrin in its formulation (Figure 1B).
3.4. Analysis of the potential interaction of two individual component in the four formulated binary mixtures
To analyze the potential interaction of the two individual components in the four formulated binary mixtures, we employed the additive index (AI) and co-toxicity coefficient (CTC) analysis. According to calculated AI value (AI > 0) and CTC value (CTC > 100), synergistic effects were determined on the Lab-S strain for all four formulated pyrethroid-containing binary insecticides (Table 3).
Calculated AI (AI > 0) and CTC values (CTC > 100) for Endigo and Athena also indicated synergistic interaction between the two components in both Field-R1 and Field-R2 TPB populations (Table 3). Although Hero showed the highest resistance ratio among all four tested pesticides, two pyrethroids in it exhibited a synergistic effect with an AI value of 0.93 for Field-R1 and 0.55 for Field-R2, or CTC value of 213.6 for Field-R1 and 154.9 for Field-R2, respectively (Table 3). However, calculated AI value (-0.38 for Field-R1 and -0.28 for Field-R2), or CTC value (72.6 for Field-R1 and 88.0 for Field-R2) for Leverage indicated antagonistic interaction between imidacloprid and β-cyfluthrin in Leverage for both Field-R1 and Field-R2 TPB populations (Table 3).
4. Discussion
The primary objective of this study was to evaluate the toxicity and resistance levels in two field resistant TPB populations against four formulated pyrethroid-containing binary insecticides through spray treatments. Our results elucidated significant resistance development in both Field-R1 and Field-R2 TPB populations to four binary insecticides. Among the tested four binary formulations, TPBs consistently exhibited the highest resistance ratio towards Hero, indicating greater tolerance to this mixture in both Field-R1 and Field-R2 TPB populations. On the other hand, TPBs displayed the lowest resistance ratio to Athena and highlights its continued effectiveness against resistant TPB populations. Interestingly, the Field-R2 TPB population collected in October displayed about 1.4-3-fold increase in resistance compared to the Field-R1 TPB population collected in July. These findings align with our previous investigations on individual insecticides examined in both resistant TPB populations , which suggested that the resistance levels in TPB populations escalated and resulted in a rapid evolution of insecticide resistance due to selection pressure accumulated within a single growing season [33].
Among the four pyrethroid-containing binary insecticides tested, both Field-R1 and Field-R2 TPB populations exhibited higher resistance levels towards Leverage and Hero compared to their corresponding individual pyrethroid. On the other hand, both field resistant TPBs exhibited similar or lower resistance to Endigo and Athena. These findings suggested that Endigo and Athena are likely superior products for slowing down resistance development compared to their corresponding individual pyrethroid in resistant TPB populations.
In the current study, the utilization of the calculated additive index (AI) and co-toxicity coefficient (CTC) assessed the interaction between the individual components in the four binary mixtures. Results indicated that the two individual components in Endigo, Leverage, Hero and Athena exhibited synergistic effects in the Lab-S TPB population, and increased pyrethroid toxicity when mixed with neonicotinoids or avermectin. Previous studies also reported synergistic interaction between pyrethroids and neonicotinoid insecticides in binary mixtures for other insect pest species, such as mosquitos [34,35,36]; Cimex lectularius [37]; Cimex hemipterus [25], Bombyx mori [38] and Drosophila [39]. Corbett [19] proposed a general theory to explain the synergistic interactions among different insecticide mixtures. According to this theory, one insecticide in the mixture interferes with the metabolic detoxification of the other insecticide, thereby enhancing the toxicity of the latter one. Applications of either Endigo or Leverage on TPBs bind both pyrethroids and neonicotinoids to the monooxygenase P450 [40,41], which hydrolyzes cytochrome P450 to catalyze both insecticides. Consequently, this prevents subsequent binding of neonicotinoid or pyrethroid insecticides catalyzed by monooxygenase enzymes [42]. In the case of Athena, the mixture of the pyrethroid bifenthrin and abamectin may allosterically activate the glutamate-gated chloride channel (GluCl) combined with bifenthrin, leading to synergistic action [26].
In addition, the calculated additive index (AI) and co-toxicity coefficient (CTC) also indicated two individual components in Endigo, Hero, and Athena (but not Leverage) exhibited synergistic effects against both Field-R1 and Field-R2 TPB populations. Furthermore, both resistant TPB populations showed similar or lower resistance levels for Athena and Endigo than their corresponding individual pyrethroid. Pre-mixed binary insecticides which contain two different active ingredients (i.e., different mode of action and target sites) may be more likely to induce synergistic toxicity, facilitate uptake and slow the development of resistance [39,42]. Endigo and Athena contain two components with differing modes of action and target either the nicotinic acetylcholine receptor (nAChR) or Glutamate-gated chloride channel (GluCl), respectively (Table 1). The probability of cross-resistance between pyrethroids and neonicotinoids or avermectin is low, and the development of multiple resistance based on different resistance alleles is also low, thus individuals with multiple resistance mechanisms will be rare. This may explain the observed similar or lower resistance levels in resistant TPBs exposed to Endigo and Athena compared to their corresponding individual pyrethroid in their formulation. However, resistant TPBs exposed to Hero, a mixture of bifenthrin and ζ-cypermethrin with the same mode of action (target on sodium channels, Table 1), exhibited much higher resistance ratio than the corresponding individual components. Developed resistance to multiple insecticides under long-term selection pressure in the field may explain the field-collected TPBs with higher resistance to Hero. An increase in insecticide applications rates in cotton producing areas of the Delta since 1999 (0.3 applications/year) may also contribute to the observed resistance [43]. Even though Hero induced synergism in both resistant TPB populations, the probability of cross-resistance between bifenthrin and ζ-cypermethrin is high [17], and led to the highest observed resistance ratio (47.60-fold) among all tested insecticides. Therefore, Hero is not a suitable choice for long-term TPB multiple resistance management, while Endigo and Athena are more effective and recommended for such management. Also, TPBs displayed a higher resistance level towards Leverage than the corresponding individual pyrethroid β-cyfluthrin and demonstrated antagonistic action in the two resistant TPB populations. The more recent intensive use of the individual component in Leverage, imidacloprid, to control sucking insect pests may explain the increased resistance to the binary mixture [44]. Although Brigadier (bifenthrin + imidacloprid) was not tested on TPBs in this study, previous research showed that resistant TPBs exposed to tank mixtures of bifenthrin and imidacloprid exhibited antagonistic effects consistent with the antagonism observed with Leverage currently [44]. Moreover, multiple resistance mechanisms, including decreased cuticular penetration and target-site insensitivity may contribute to the antagonistic effect as well [42].
Although manufacturers may optimize component ratios in pyrethroid-containing binary insecticides(e.g. Endigo, Leverage and Athene), tank mixing of insecticides is common practice in integrated pest management for row crops where multiple insects and plant pathogens are targeted in the field [45]. For instance, tank mixing bifenthrin plus acephate is synergistic on TPBs and is often used to control TPBs and other cotton pests [44]. Additionally, antagonistic interactions between tank mixing pyrethroids and organophosphates have been reported in other insects, such as Bemisia tabaci [46]. Similarly, a binary mixture of deltamethrin and chlorpyrifos showed antagonistic effects against bollworm at 96 hour [22]. While tank mixing lacks an optimized ratio, the interaction of bifenthrin and fipronil was additive on Musca domestica when in the ratio 1:1, whereas used in the ratio of LC50: LC50, the mixtures produced a synergistic effect [42]. Therefore, the antagonistic or synergistic interactions among tank mixed binary insecticides depends on the type of insecticides used, the ratios of components and genetic background of the test organism [42].
The use of binary mixtures is a common practice for controlling resistant insect species, and two of the mixtures in the current study are recommended options for managing resistant TPB populations. Our study highlights the importance of understanding the interactions between different insecticides to develop effective pest management strategies. Considering the control cost, two or more compounds might be mixed to obtain the best control effects [35]. However, long-term use of insecticide mixtures can pose risk to beneficial organisms [47]. Some mixtures including neonicotinoids are toxic to the bee Apis mellifera [48,49], while others can disrupt the survival and development of non-target aquatic organisms [50,51,52]. Despite these risks, our study found that two formulated pyrethroid-containing mixtures Endigo and Athena exhibited synergistic interactions between pyrethroid and non-pyrethroid mixtures, which could effectively manage resistant TPB populations. Our work contributes to the development of more efficient and sustainable approaches for combating TPB resistance and enhancing overall pest management strategies in cotton cultivation.
Author Contributions
Conceived and designed the study: Y.D. and Y.Z. Performed the experiments: Y.D. and S.S. Methodology: M.P. Wrote the manuscript: Y.D. and J.C. All authors took part in discussing, reading and approving the manuscript.
Funding
The work is supported by USDA-ARS Research Project# 6066-22000-090-00DInsect. Control and Resistance Management in Corn, Cotton, Sorghum, Soybean, and Sweet Potato, and Alternative Approaches to Tarnished Plant Bug Control in the Southern United States.
Acknowledgments
The authors would like to thank Amiyah Hampton, ARS-USDA, Southern Insect Management Research Unit (SIMRU), for her help with bioassays.
Conflicts of Interest
None.
References
- Snodgrass, G.L. Insecticide restance in field populations of the tarnished plant bug (Heteroptera:Miridae) in cotton in the Mississippi Delta. J. Econ. Entomol. 1996, 89, 783-790.
- Layton, M.B. Biology and damage of the tarnished plant bug, Lygus lineolaris, in cotton; 2000; Volume 23, pp. 7-20.
- Zhu, Y.-C.; Yao, J.; Luttrell, R. Identification of genes potentially responsible for extra-oral digestion and overcoming plant defense from salivary glands of the tarnished plant bug (Hemiptera: Miridae) using cDNA sequencing. J. Insect Science 2016, 16. [CrossRef]
- George, J.; Glover, J.P.; Gore, J.; Crow, W.D.; Reddy, G.V.P. Biology, ecology, and pest management of the tarnished plant bug, Lygus lineolaris (Palisot de Beauvois) in southern row crops. Insects 2021, 12. [CrossRef]
- Hollingsworth, R.; Steinkraus, D.; Tugwell, N. Responses of Arkansas populations of tarnished plant bugs (Heteroptera: Miridae) to insecticides, and tolerance differences between nymphs and adults. J. Econ. Entomol. 1997, 90, 21-26. [CrossRef]
- Pankey, J.; Leonard, B.; Graves, J.; Burris, E. Toxicity of acephate, cypermethrin, and oxamyl to tarnished plant bugs in vial bioassays and cage studies on cotton. In Proceedings of the Beltwide Cotton Conferences (USA), 1996.
- Snodgrass, G.L.; Gore, J. Status of insecticide resistance for the tarnished plant bug. In Proceedings of the Proc. Beltwide Cotton Conf., National Cotton Council, Memphis, Tennessee, 2007; pp. 56-61.
- Snodgrass, G.L.; Gore, J.; Abel, C.A.; Jackson, R. Acephate resistance in populations of the tarnished plant bug (Heteroptera: Miridae) from the Mississippi River Delta. J. Econ. Entomol. 2009, 102, 699-707. [CrossRef]
- Snodgrass, G.L.; Scott, W.P. Effect of ULV malathion use in boll weevil (Coleoptera: Curculionidae) eradication on resistance in the tarnished plant bug (Heteroptera: Miridae). J. Econ. Entomol. 2003, 96, 902-908. [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [CrossRef]
- Dong, K.; Du, Y.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.; Silver, K.; Zhorov, B.S. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 2014, 50, 1-17. [CrossRef]
- Parys, K.A.; Luttrell, R.G.; Snodgrass, G.L.; Portilla, M.R. Patterns of tarnished plant bug (Hemiptera: Miridae) resistance to pyrethroid insecticides in the lower mississippi delta for 2008-2015: Linkage to pyrethroid use and cotton insect management. J. Insect Sci. 2018, 18. [CrossRef]
- Snodgrass, G.L. Pyrethroid resistance in a field population ot the tarnished plant bug in the Mississippi Delta. In Proceedings of the Proc. Beltwide Cotton Conf., National Cotton Council, Memphis, Tennessee, 1994; pp. 1186-1187.
- Snodgrass, G.L.; Elzen, G.W. Insecticide resistance in a tarnished plant bug population in cotton in the Mississippi Delta. Southwest Entomol 1995, 20, 317-323. [CrossRef]
- Snodgrass, G.L.; Scott, W.P. Seasonal changes in pyrethroid resistance in tarnished plant bug (Heteroptera: Miridae) populations during a three-year period in the delta area of Arkansas, Louisiana, and Mississippi. J. Econ. Entomol. 2000, 93, 441-446. [CrossRef]
- Snodgrass, G.; Scott, W. discriminating-dose bioassay for detecting pyrethroid resistance in tarnished plant bug (Heteroptera: Miridae) populations. Southwest. Entomol. 1999.
- Layton, M.B. Cotton Insect Control Guide 2002. Mississippi State University Extension Service Publication 343.Mississippi State University, MS 2003. [CrossRef]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annual review of entomology 2000, 45, 371-391. [CrossRef]
- Corbel, V.; Raymond, M.; Chandre, F.; Darriet, F.; Hougard, J.M. Efficacy of insecticide mixtures against larvae of Culex quinquefasciatus (Say) (Diptera: Culicidae) resistant to pyrethroids and carbamates. Pest Manag. Sci. 2004, 60, 375-380. [CrossRef]
- Martin, T.; Ochou, O.G.; Vaissayre, M.; Fournier, D. Organophosphorus insecticides synergize pyrethroids in the resistant strain of cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) from West Africa. J. Econ. Entomol. 2003, 96, 468-474. [CrossRef]
- Ahmad, M. Observed potentiation between pyrethroid and organophosphorus insecticides for the management of Spodoptera litura (Lepidoptera: Noctuidae). Crop protection 2009, 28, 264-268. [CrossRef]
- Ahmad, M.; Saleem, M.A.; Sayyed, A.H. Efficacy of insecticide mixtures against pyrethroid- and organophosphate-resistant populations of Spodoptera litura (Lepidoptera: Noctuidae). Pest Manag. Sci. 2009, 65, 266-274. [CrossRef]
- Ascher, K.; Eliyahu, M.; Ishaaya, I.; Zur, M.; Ben-Moshe, E. Synergism of pyrethroid—organophosphorus insecticide mixtures in insects and their toxicity against spodoptera littoralis larvae. Phytoparasitica 1986, 14, 101-110.
- Jeschke, P.; Nauen, R. Neonicotinoids-from zero to hero in insecticide chemistry. Pest Manag. Sci. 2008, 64, 1084-1098. [CrossRef]
- Dang, K.; Doggett, S.L.; Lee, C.Y. Performance of pyrethroid-neonicotinoid mixture formulations against dield-xollected atrains of the tropical bed bug (Hemiptera: Cimicidae) on different substrates. J. Econ. Entomol. 2022. [CrossRef]
- Wolstenholme, A.J.; Rogers, A.T. Glutamate-gated chloride channels and the mode of action of the avermectin/milbemycin anthelmintics. Parasitol. 2005, 131 Suppl, S85-95. [CrossRef]
- Parys, K.A.; Snodgrass, G.L.; Luttrell, R.G.; Allen, K.C.; Little, N.S. Baseline susceptibility of Lygus lineolaris (Hemiptera: Miridae) to novaluron. J. Econ. Entomol. 2015, 109, 339-344. [CrossRef]
- Portilla, M.; Reddy, G.V.P. Development of a method for rearing Nezara viridula (Heteroptera: Pentatomidae) on a semi-solid artificial diet. J. Insect Sci. 2021, 21. [CrossRef]
- Portilla, M.; Snodgrass, G.; Streett, D. Effect of modification of the NI artificial diet on the biological fitness parameters of mass reared western tarnished plant bug, Lygus hesperus. J. Insect Sci. 2011, 11. [CrossRef]
- Marking, L. Toxicity of chemical mixtures. Fundamentals of aquatic toxicology: methods and applications. Hemisphere Publishing Corporation Washington DC 1985, 164-176.
- Sun, Y.-P.; Johnson, E.R. Analysis of joint action of insecticides against house flies. J. Econ. Entomol. 1960, 53, 887-892. [CrossRef]
- Wang, Y.; Dai, D.; Yu, Y.; Yang, G.; Shen, W.; Wang, Q.; Weng, H.; Zhao, X. Evaluation of joint effects of cyprodinil and kresoxim-methyl on zebrafish, Danio rerio. J. Hazard. Mater.2018, 352, 80-91. [CrossRef]
- Du, Y.; Zhu, Y.C.; Portilla, M.; Zhang, M.; Reddy, G.V.P. The mechanisms of metabolic resistance to pyrethroids and neonicotinoids fade away without selection pressure in the tarnished plant bug Lygus lineolaris. Pest Manag. Sci. 2023. [CrossRef]
- Fongnikin, A.; Houeto, N.; Agbevo, A.; Odjo, A.; Syme, T.; N'Guessan, R.; Ngufor, C. Efficacy of Fludora® Fusion (a mixture of deltamethrin and clothianidin) for indoor residual spraying against pyrethroid-resistant malaria vectors: laboratory and experimental hut evaluation. Parasit Vectors 2020, 13, 466. [CrossRef]
- Darriet, F.; Chandre, F. Efficacy of six neonicotinoid insecticides alone and in combination with deltamethrin and piperonyl butoxide against pyrethroid-resistant Aedes aegypti and Anopheles gambiae (Diptera: Culicidae). Pest Manag. Sci. 2013, 69, 905-910. [CrossRef]
- Ngufor, C.; Fongnikin, A.; Rowland, M.; N'Guessan, R. Indoor residual spraying with a mixture of clothianidin (a neonicotinoid insecticide) and deltamethrin provides improved control and long residual activity against pyrethroid resistant Anopheles gambiae sl in Southern Benin. PLoS One 2017, 12, e0189575. [CrossRef]
- Wang, C.; Singh, N.; Cooper, R. Field Study of the Comparative Efficacy of Three Pyrethroid/Neonicotinoid Mixture Products for the Control of the Common Bed Bug, Cimex lectularius. Insects 2015, 6, 197-205. [CrossRef]
- Yu, R.X.; Wang, Y.H.; Hu, X.Q.; Wu, S.G.; Cai, L.M.; Zhao, X.P. Individual and Joint Acute Toxicities of Selected Insecticides Against Bombyx mori (Lepidoptera: Bombycidae). J. Econ. Entomol. 2015, 109, 327-333. [CrossRef]
- Luong, H.N.B.; Damijonaitis, A.; Nauen, R.; Vontas, J.; Horstmann, S. Assessing the anti-resistance potential of public health vaporizer formulations and insecticide mixtures with pyrethroids using transgenic Drosophila lines. Parasit Vectors 2021, 14, 495. [CrossRef]
- Zoh, M.G.; Bonneville, J.-M.; Tutagata, J.; Laporte, F.; Fodjo, B.K.; Mouhamadou, C.S.; Sadia, C.G.; McBeath, J.; Schmitt, F.; Horstmann, S.; et al. Experimental evolution supports the potential of neonicotinoid-pyrethroid combination for managing insecticide resistance in malaria vectors. Sci. Rep. 2021, 11, 19501. [CrossRef]
- Scott, J.G. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757-777. [CrossRef]
- Khan, H.A.; Akram, W.; Shad, S.A.; Lee, J.J. Insecticide mixtures could enhance the toxicity of insecticides in a resistant dairy population of Musca domestica L [corrected]. PLoS One 2013, 8, e60929. [CrossRef]
- Fleming, D.E.; Krishnan, N.; Catchot, A.L.; Musser, F.R. Susceptibility to insecticides and activities of glutathione S-transferase and esterase in populations of Lygus lineolaris (Hemiptera: Miridae) in Mississippi. Pest Manag. Sci. 2016, 72, 1595-1603. [CrossRef]
- Jones, M.M.; Duckworth, J.L.; Robertson, J. Toxicity of Bifenthrin and Mixtures of Bifenthrin Plus Acephate, Imidacloprid, Thiamethoxam, or Dicrotophos to Adults of Tarnished Plant Bug (Hemiptera: Miridae). J. Econ. Entomol. 2018, 111, 829-835. [CrossRef]
- Studebaker, G.; Davis, J.; Hopkins, J.D.; Johnson, D.R.; Loftin, K., Lorenz,; G., S., N., ; Spradley, P.; Zawislak, J. 2016 Insecticide Recommendations for Arkansas. University of Arkansas Cooperative Extension Service 2016, MP-144.
- Ahmad, M. Potentiation/Antagonism of pyrethroids with organophosphate insecticides in Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 2007, 100, 886-893. [CrossRef]
- Attique, M.; Khaliq, A.; Sayyed, A. Could resistance to insecticides in Plutella xylostella (Lep., Plutellidae) be overcome by insecticide mixtures? J.Appl. Entomol. 2006, 130, 122-127. [CrossRef]
- Iwasa, T.; Motoyama, N.; Ambrose, J.T.; Roe, R.M. Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop prot. 2004, 23, 371-378.
- Belden, J.B. The acute toxicity of pesticide mixtures to honeybees. Integr. Environ. Assess Manag. 2022. [CrossRef]
- Houndji, M.A.B.; Imorou Toko, I.; Guedegba, L.; Yacouto, E.; Agbohessi, P.T.; Mandiki, S.N.M.; Scippo, M.L.; Kestemont, P. Joint toxicity of two phytosanitary molecules, lambda-cyhalothrin and acetamiprid, on African catfish (Clarias gariepinus) juveniles. J. Environ. Sci. Health B 2020, 55, 669-676. [CrossRef]
- Guedegba, N.L.; Imorou Toko, I.; Agbohessi, P.T.; Zoumenou, B.; Douny, C.; Mandiki, S.N.M.; Schiffers, B.; Scippo, M.L.; Kestemont, P. Comparative acute toxicity of two phytosanitary molecules, lambda-cyhalothrin and acetamiprid, on Nile Tilapia (Oreochromis Niloticus) juveniles. J. Environ. Sci. Health B 2019, 54, 580-589. [CrossRef]
- Kunce, W.; Josefsson, S.; Örberg, J.; Johansson, F. Combination effects of pyrethroids and neonicotinoids on development and survival of Chironomus riparius. Ecotoxicol. Environ. Saf. 2015, 122, 426-431. [CrossRef]
Figure 1.
The resistance ratios (RR) of formulated pyrethroid-containing binary insecticides (colored bars) and corresponding individual components (black bars) in two field resistant TPB populations. (a) Field-R1: collected in July and (b) Field-R2: collected in October after spray application for 48h. Resistance ratios were determined as the ratio of LC50 of the field resistant TPB population (Field-R1/Field R2) divided by LC50 of the Lab-S strain. LC50 values and 95% confidence intervals were calculated by Probit analyses using SPSS software and data were presented as the means ± S.D. Within each group of resistance ratio, means sharing different letter on the top of bars are significantly different, as determined by one-way analysis of variance with Tukey’s HSD test, and significant values were set at P < 0.05.
Figure 1.
The resistance ratios (RR) of formulated pyrethroid-containing binary insecticides (colored bars) and corresponding individual components (black bars) in two field resistant TPB populations. (a) Field-R1: collected in July and (b) Field-R2: collected in October after spray application for 48h. Resistance ratios were determined as the ratio of LC50 of the field resistant TPB population (Field-R1/Field R2) divided by LC50 of the Lab-S strain. LC50 values and 95% confidence intervals were calculated by Probit analyses using SPSS software and data were presented as the means ± S.D. Within each group of resistance ratio, means sharing different letter on the top of bars are significantly different, as determined by one-way analysis of variance with Tukey’s HSD test, and significant values were set at P < 0.05.
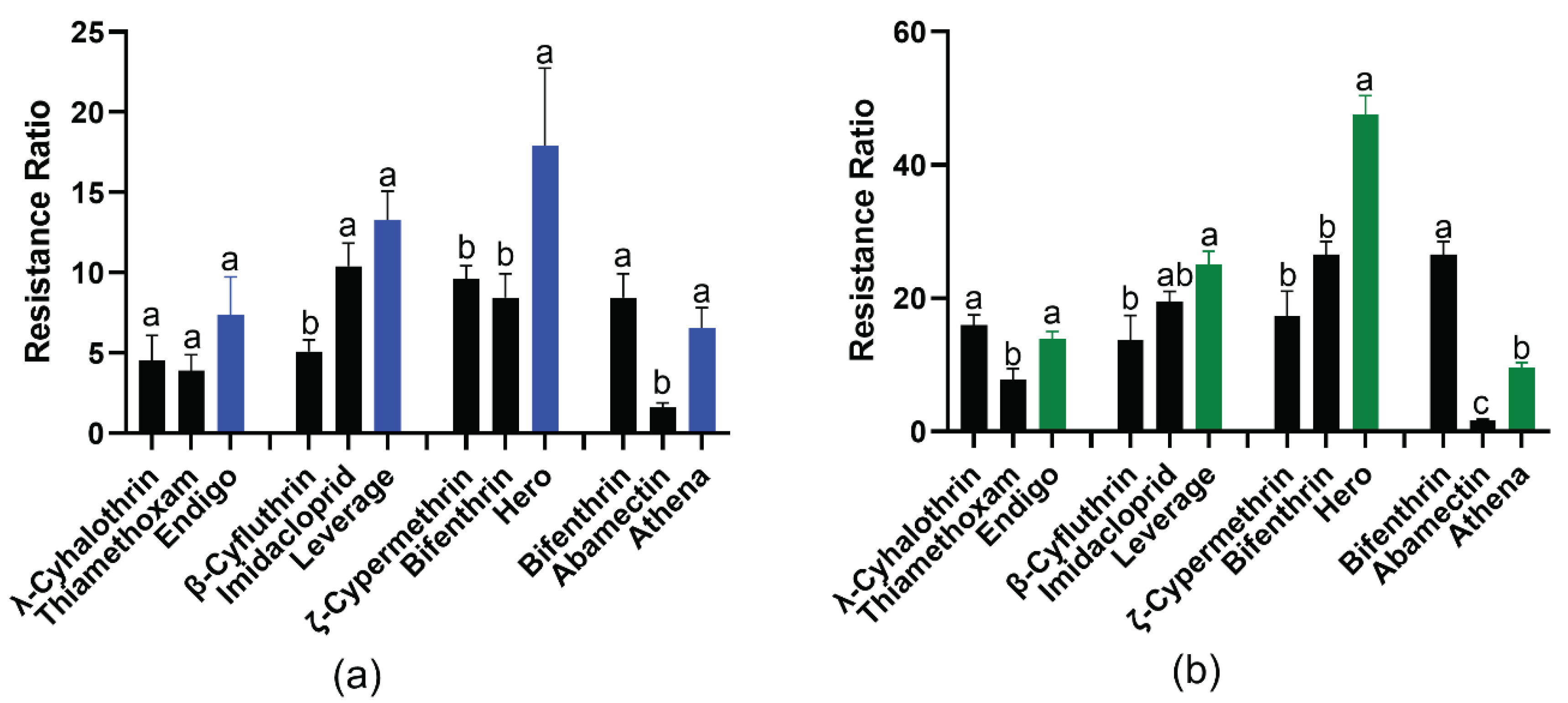

Table 1.
The commercial name, common name (with percentage active ingredient), manufacturer, and mode of action (MOA) for insecticides used in the current study.
Table 1.
The commercial name, common name (with percentage active ingredient), manufacturer, and mode of action (MOA) for insecticides used in the current study.
| Commercial name | Common name (Percentage of active ingredient) | Manufacturer | Mode of action | |
|---|---|---|---|---|
| 1 | Endigo 2.06ZC | Thiamethoxam (12.60%) + λ -Cyhalothrin (9.48%) | Syngenta | 3A+4A |
| 2 | Warrior II | λ-cyhalothrin (22.8%) | Syngenta | 3A |
| 3 | Centric 40WG | Thiamethoxam (40%) | Syngenta | 4A |
| 4 | Leverage 360EC | Imidacloprid (21.0%) + β -Cyfluthrin (10.5%) | Bayer Crop Science | 4A+3A |
| 5 | Baythroid XL | β-cyfluthrin (12.7%) | Bayer | 3A |
| 6 | Advise® Four | Imidacloprid (40.4%) | Winfield | 4A |
| 7 | Hero 1.24 | Bifenthrin (11.25%) + ζ -Cypermethrin (3.75%) | FMC | 3A+3A |
| 8 | Tundra®EC | Bifenthrin (25.1%) | Winfield | 3A |
| 9 | Mustang Maxx | ζ-cypermethrin (9.15%) | FMC | 3A |
| 10 | Athena | Bifenthrin (8.84%) + Avermectin B1(1.33%) | FMC | 3A+6 |
| 11 | Epi-Mek | Abamectin (15%) | Syngenta | 6 |
Table 2.
The toxicity of formulated pyrethroid-containing binary insecticides against the susceptible and two resistant populations of Lygus lineolaris after spray application for 48h.
Table 2.
The toxicity of formulated pyrethroid-containing binary insecticides against the susceptible and two resistant populations of Lygus lineolaris after spray application for 48h.
| Compounds | Strain a | Slope | LC50 (μg/mL) b | 95% Confidence Limits (μg/mL) | χ2 | P | RRc |
|---|---|---|---|---|---|---|---|
| Endigo |
Lab-S | 2.877 ± 0.296 | 22.54 | 19.19 - 26.49 | 1.53 | 0.68 | -- |
| Field-R1 | 2.364 ± 0.562 | 166.64 | 113.75 - 242.05 | 4.74 | 0.32 | 7.39 | |
| Field-R2 | 1.947 ± 0.334 | 315.18 | 254.95 – 397.47 | 1.07 | 0.90 | 13.98 | |
| Leverage | Lab-S | 2.613 ± 0.278 | 20.53 | 17.35 - 24.16 | 3.27 | 0.35 | -- |
| Field-R1 | 5.049 ± 0.911 | 273.44 | 237.29 - 318.85 | 2.39 | 0.30 | 13.3 | |
| Field-R2 | 1.628 ± 0.346 | 513.94 | 384.53 – 883.0 | 1.75 | 0.78 | 25.0 | |
| Hero | Lab-S | 2.929 ± 0.285 | 20.39 | 17.59 - 23.66 | 2.20 | 0.53 | -- |
| Field-R1 | 2.308 ± 0.427 | 329.96 | 266.61 – 439.07 | 2.11 | 0.35 | 16.2 | |
| Field-R2 | 3.377 ± 0.411 | 970.67 | 836.94 – 1177.82 | 5.72 | 0.33 | 47.60 | |
| Athena | Lab-S | 2.876 ± 0.351 | 73.60 | 62.84 - 86.62 | 2.74 | 0.26 | -- |
| Field-R1 | 3.363 ± 0.648 | 479.60 | 389.88 – 598.0 | 1.07 | 0.59 | 6.56 | |
| Field-R2 | 3.283 ± 0.403 | 708.30 | 619.44 – 828.43 | 6.70 | 0.15 | 9.62 |
a Lab-S: laboratory-reared susceptible strain; Field- R: field-collected resistant TPB populations, collected in July (Field-R1) and October (Field-R2). b LC50 values and 95% confidence intervals were calculated by Probit analyses using SPSS software. c Resistant ratio (RR) calculated by dividing LC50 of the lab-S by LC50 of field- collected resistant TPB populations.
Table 3.
The calculated additive index (AI) and co-toxicity coefficient (CTC) value of binary insecticides in the Lab-S and two field resistant Lygus lineolaris populations (Field-R1 and Field-R2) after spray application for 48h.
Table 3.
The calculated additive index (AI) and co-toxicity coefficient (CTC) value of binary insecticides in the Lab-S and two field resistant Lygus lineolaris populations (Field-R1 and Field-R2) after spray application for 48h.
| Compounds | Strain | AI | CTC |
|---|---|---|---|
| Endigo |
Lab-S | 1.28 | 227.7 |
| Field-R1 | 0.26 | 126.4 | |
| Field-R2 | 0.79 | 157.4 | |
| Leverage | Lab-S | 0.32 | 131.6 |
| Field-R1 | -0.38 | 72.6 | |
| Field-R2 | -0.28 | 88.0 | |
| Hero | Lab-S | 2.71 | 369.3 |
| Field-R1 | 0.93 | 213.6 | |
| Field-R2 | 0.55 | 154.9 | |
| Athena | Lab-S | 1.22 | 222.5 |
| Field-R1 | 1.41 | 233.8 | |
| Field-R2 | 2.16 | 316.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



