
Submitted:
17 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
Methodologies for the synthesis and purification of metabolites, which have been developed following their discovery, analysis and structural identification, have been involved in numerous life science milestones. The renewed focus on the small molecule domain of biological cells has also created an increasing awareness of the rising gap between the metabolites identified and the metabolites which have been prepared as pure compounds. Due to the large number and molecular diversity of metabolites the design and engineering of resource-efficient and straightforward synthetic methodologies for their production have attracted much interest. The variety of metabolic pathways in biological cells provide a wonderful blueprint for designing simplified and resource-efficient synthetic routes to desired metabolites. Therefore, biocatalytic systems have become key enabling tools for the synthesis of an increasing number of metabolites, which can then be utilized as standards, enzyme substrates, inhibitors or products, or for the discovery of novel biological functions.
Keywords:
metabolite synthesis
; biocatalytic systems
; product recovery
; metabolite purification
1. Introduction
Traditional knowledge of the bioresources for microbial, plant and animal metabolites, as well as their processing and application, has been contributing tremendously to the quality of life for thousands of years already. Small-molecular weight natural products have accompanied mankind and supported the quality of life in highly relevant areas, such as nutrition, diagnostics and therapy of diseases, colours, cosmetics and well-being, with an ever increasing knowledge base. Numerous milestones have been achieved in the isolation and purification of metabolites from natural bioresources and the elucidation of their molecular structures, demonstrating their large structural diversity. Renewed interest in the small molecule domain of biology [1] and the structures and functions of natural products and metabolites has brought back the spotlight onto metabolism [2]. Manifold interactions of metabolites with other biomolecules, such as proteins, DNA, RNA or other metabolites, within the same as well as with other biological cells are of much fundamental interest for biological sensing, controlling and regulatory processes at the genetic, epigenetic, transcriptomic and proteomic levels. As precise experimental investigations of metabolites and its interactions with other biomolecules, such as being a substrate, inhibitor or activator of an enzyme, are only possible by having available pure metabolites, synthetic access is essential.
The concept of the metabolome [3] has contributed to revitalize the interest in metabolites and metabolic pathways. Analyses of large numbers of metabolites have been facilitated by great advances in powerful analytical methodologies with high information content [4,5,6], on which future developments towards identifying by mass spectrometry (MS) [7] and nuclear magnetic resonance (NMR) [8] known and unknown metabolites can be based. A growing number of large databases, which are focussing on a) metabolite analysis by MS [7] or NMR [9], b) metabolites, metabolic pathways, natural products and small molecules of biological interest [10,11,12], and c) species-specific metabolites and metabolic pathways (see table 1) [13,14,15,16,17,18,19,20,21,22], provide fast access to information on the increasing number of identified metabolites and metabolic pathways.
The development of analytical methodologies for identifying biologically active metabolites, interactions of metabolites with proteins and other biomolecules [23,24,25] offers great opportunities for delineating molecular mechanisms of numerous biological processes. These can range from the activities of proteins and their modulation by activators, inhibitors, allosteric regulation, or post-translational modification, to the sensing of metabolites by riboswitches or post-transcriptional modification of RNA, and to controlling gene expression.
The use of isotopes for labelling small molecules continues to be essential for analytical methodologies, from measuring metabolite concentrations in biological matrices, determining metabolite fluxes in biological organisms to the discovery of metabolic pathways [26,27,28]. Biocatalytic systems have thereby been important for the synthesis of isotope-labelled metabolites, whether by whole cell systems or by isolated enzymes [29,30]. Synthesis has also been key for proving the correct molecular structure [31], and for the production of larger amounts of the pure metabolite [32,33]. While synthetic organic chemistry provides a huge repertoire of well-established reactions, tools and strategies for the synthesis of metabolites, safety, health, environment and sustainability aspects have become of increasing importance in industrial manufacturing processes for metabolites [34]. Metabolic pathways used by biological organisms to prepare valuable metabolites from raw materials available in their environment have also been successfully utilized and developed into industrial bioprocesses for manufacturing of metabolites, metabolite-like compounds, non-natural chemical entities and metabolites thereof [35,36,37]. The metabolic pathways which start from highly functionalized biobased raw materials instead of hydrocarbons provide also inspirations for biocatalytic defunctionalization reactions in transitioning towards raw materials from bioresources [38]. The desired use of biobased raw materials needs however to consider also other goals, such as biodiversity, sustainability and supply chain issues, if biologically endangered and rare biological species are required or the amount and quality of the biobased raw material is variable and subject to various environmental factors [39]. Therefore, the molecular and engineering fundamentals of how nature achieves the biosynthesis of metabolites by using biocatalytic reactions in microbes, plants, animals and humans have attracted much interest as a blueprint for optimized biocatalytic systems of metabolite production [40].
The purpose of this work is to give an overview of design and engineering approaches for biocatalytic systems in metabolite production and their application in manufacturing processes. The significance of biocatalytic systems for metabolite production is connected with general strategic advantages of using biocatalysis in synthesis, such as high selectivity, shortened synthetic routes, safety, health and sustainability benefits [41]. Biocatalytic system design and engineering towards the synthesis of metabolites starts with route selection and includes the preparation of suitable biocatalysts and raw materials, reaction engineering, process integration, intensification and scaling of the selected metabolite manufacturing processes [34]. The main aim of the work focusses on designing and engineering biocatalytic systems in order to produce metabolites. The great progress in delineating natural and engineering synthetic metabolic pathways, advanced methodologies and tools for finding and applying biocatalysts and last but not least, product recovery and purification have brought biocatalytic systems into a privileged position for producing not only natural metabolites, but also metabolite-like compounds, metabolites derived from the transformation of new chemical entities by biological organisms and isotope-labelled metabolites.
Knowing a recipe how to synthesize a metabolite or metabolite-like compound, or even more convenient, having already available at hand the pure product, has been and continues to be highly important and relevant [42,43] for a number of reasons, as shown in Figure 1A for fundamental and applied sciences, as well as for a great variety of applications in industry and medicine, as shown in Figure 1B.
In view of millions of protein sequences genomic enzymology tools are highly valuable for generating clues and hypotheses guiding experiments towards the correct assignment of enzyme and metabolic functions, as the functions of a significant fraction of protein sequences are either unknown, or uncertain, or even misassigned [44]. The final proof of the assignments is not possible without experimental verification of the predicted enzyme functions, which requires the corresponding metabolites as enzyme substrate for demonstrating its conversion to the predicted product and for performing the enzyme activity assays. and discovery of novel biological functions. In case of chiral enzyme substrates or products, or for investigations of enzymatic reaction mechanisms, enantiopure metabolites can resolve fundamental questions and provide more detailed insights. Applications in industry are growing and include food supplements, pharmaceuticals. flavours, fragrances, cosmetics, dyes, agrochemicals. Therefore safety, health, environment and sustainability aspects are not only important for manufacturing metabolites at industrial large scale, but also for applications and the whole product life cycle. Improving energy and resource efficiency in manufacturing metabolites, such as reducing energy use and avoiding protection-deprotection schemes in lengthy chemical synthesis from fossil-based raw materials or minimizing biological waste in low-yield extractive procedures from biobased raw materials, is essential in building more resilient and sustainable manufacturing routes to metabolites. The blueprint of nature with the molecular transformations catalyzed by enzymes and the metabolic pathways as well as the great advances of biocatalysis provide rich sources of knowledge for designing and engineering biocatalytic systems for the synthesis of metabolites.
Biocatalytic systems using whole cells have been developed into numerous fermentation processes as well as biotransformation processes at industrial large scale for the production of metabolites [45,46,47,48]. The vast knowledge base and the rich diversity of biocatalytic whole cell systems have lead to biocatalytic metabolite production by suitable growing or resting whole cells (see Figure 2). Metabolic engineering and synthetic biology enable improvements in titer, rate and space-time yield of metabolite synthesis from the starting materials a) by increasing the performance of the biosynthetic pathways to the metabolite, and b) by deleting any biocatalytic degradation reaction of the final metabolite and of any metabolic intermediates. Biocatalytic whole cell systems are also connected with a high degree of complexity, which can be reduced by using cell-free biocatalytic systems (see Figure 2) in different forms of purification, from crude cell-free extracts to isolated and purified enzymes. Whatever biocatalytic system is considered, bioprocess design and engineering need to address and optimize various parameters such as biocatalytic pathway selection, form and status of the biocatalysts, reaction engineering, downstream processing and purification of the metabolites [34,49]. Cell-free biocatalytic systems have the advantage of reducing the bioprocess complexity by the absence of interfering and degrading enzymes or the removal of mass-transfer limitations for substrates and products [49].
While the step economy of a biosynthetic pathway is already considered in the design phase, possible improvements such as complexity reduction, intermediate purification steps, the number of separate reactors needed, are also taken into account by the degree of process integration of a biocatalytic metabolite synthesis. Depending on how many reaction steps are needed for transforming the starting materials to the metabolites, one-step to multistep enzymatic reactions are developed, if possible in one pot. If all enzymatic reactions of a biosynthetic pathway can be performed well, enzymatic total synthesis can be achieved [50].
2. Synthesis of Naturally Occurring Metabolites
The milestone discoveries that small molecules of life could not only be isolated from nature, but could also be synthesized in the laboratory from inorganic chemical precursors, started the new era of synthetic organic chemistry [51]. The synthesis of urea by Friedrich Wöhler in 1828 [52] or acetic acid by Hermann Kolbe in 1845 [53] were clear demonstrations that organic compounds, which are found and formed naturally in living organisms, could be prepared starting from inorganic materials. It sparked a tremendous interest in synthetic organic chemistry and the art and science of total synthesis of numerous more complex natural products [54,55,56,57,58]. Total synthesis has not only been key to the final proof of structures, or for correcting incorrect structures [59], but has also been a key driver for novel synthetic methods in organic chemistry, which are at its heart, up to the present time [60]. The tools and methodologies for broadly applicable synthetic reactions, either reactions which already well established, or newly discovered and emerging from modern organic chemistry, have enabled the total synthesis of an impressive number of naturally occurring metabolites, from very simple structures without any stereogenic center to the most complex metabolites, such as the marine natural product palytoxin, which contains 63 stereogenic centers as well as four trans- and three cis-carbon-carbon double bonds [61,62,63,64,65,66]. Total synthesis without using protecting groups is of much interest for reducing the complexity, cost and the number of steps, as shown by marine natural products synthesis [67]
The milestone discoveries of living microbial whole cells [68], their biosynthetic capabilities and the elucidation of the organic chemistry of the underlying biocatalytic reactions exerted by cell-free extracts [69] have started the era of synthetic biochemistry. Although biological formation of urea was known, the elucidation of urea biosynthesis in animals from ammonia and carbon dioxide [70,71] required major scientific breakthroughs. Numerous bioprocesses for producing naturally occurring metabolites have been developed to industrial large scale [72,73,74,75,76,77].
Natural metabolites continue to be highly important as sources for small molecule pharmaceuticals for treating diseases such as infectious diseases, cancer, cardiovascular diseases, diabetes, glaucoma or multiple sclerosis [78], and large-scale manufacturing is key for adequate supply. Therefore efficient and reliable bioprocesses to metabolites have been important to replace a) extraction processes from endangered biological species which may become extinct and which may give yields depending on various environmental factors, or b) non-sustainable production procedures combining extraction from biological species and synthetic modifications. A sustainable plant cell fermentation process for producing the natural diterpenoid paclitaxel (registered trade name Taxol(R)) at industrial large scale (see Figure 3) preserves Taxus plants and is able to provide the required amounts of this Taxus species metabolite, which the WHO lists as Essential Medicine and is used in cancer treatment [79,80]. Human health and the quality of life of large numbers of people around the world have greatly benefitted from a long history of dedicated work to discover and develop small molecules formed by biological organisms as gifts from nature for their use as anti-infectives in the 20th century [81,82]. It is however not the time for complacency today, as the appearance of new infectious agents, human negligence and economic boundary conditions require actions towards the discovery, development and reliable production of novel anti-infectives [83,84,85].
Beyond manufacturing of metabolites by bioprocesses at industrial large scale as anticancer drugs, anti-infectives and other pharmaceuticals for human health [73,77] biomanufacturing of a variety of other metabolites, such as vitamins, organic acids, flavours and fragrances have become increasingly attractive for various industrial sectors [72,74,75,76,86,87,88]. The efficient enzymatic cyclization of (E,E)-homofarnesol, which can be produced from the fermentation product (E)-β-farnesene, to the fragrance ingredient (-)-ambrox catalyzed by engineered squalene hopene cyclase at industrial scale (see Figure 4) represents a significant improvement in carbon efficiency and sustainability [87].
Another important area is the synthesis of metabolites for analytical or diagnostic applications, such as the use of metabolites in analytical or diagnostic devices, as standards, or for measuring enzyme activities. The synthesis of metabolites which act as ionophores has been of much interest for the analysis and monitoring of biomedically and environmentally relevant ions via ion-selective electrodes and sensors [89], for example the biomanufacturing of highly pure nonactin as neutral ionophore in monitoring ammonium ions [90,91].
Metabolites need also to be synthesized for analytical investigations involving the measurement of enzyme activities, such as the analysis of enzyme activities relevant to clinical chemistry, food analysis, enzymology, enzyme production, environment, verification or discovery of novel enzyme functions, development of enzyme inhibitors, or the analysis of activating, signalling or regulatory functions.
Energy metabolism and glycolytic pathways are central to biological organisms. As the metabolites of the monosaccharide catabolic pathways are essential. It is therefore desirable to synthesize in pure and stable form the metabolites of the most common pathways for the breakdown of D-glucose, the Emden-Meyerhof-Parnas, the Entner-Doudoroff and the Pentose Phosphate pathway, because these are central to all kingdoms of life. The enantiomerically pure metabolite D-glyceraldehyde-3-phosphate, which occurs in all the three most common D-glucose catabolic pathways, as well as the enantiomerically pure L-glyceraldehyde 3-phosphate, which is toxic to cells, have been synthesized from the corresponding aldehydes by biocatalytic ATP-dependent phosphorylation using enantiocomplementary kinases and phosphoenol pyruvate/pyruvatekinase for regenerating ATP. Glycerol kinase has been used in the enantioselective phosphorylation of L-glyceraldehyde [92,93], which could be prepared by glycerol dehydrogenase-catalyzed resolution of racemic glyceraldehyde [94,95]. Dihydroxyacetone kinase has been found as the corresponding enantiocomplementary enzyme to enantioselectively catalyze the phosphorylation of D-glyceraldehyde [96]. The enolase substrate D-glycerate-2-phosphate has been prepared by phosphorylating D-glycerate using recombinant glycerate-2-kinase, ATP as cofactor and phosphoenolpyruvate/pyruvatekinase for ATP regeneration [97]. In the pentose phosphate pathway D-xylulose-5-phosphate has been prepared by two different routes, either by transketolase-catalyzed condensation of D-glyceraldehyde-3-phosphate [98,99] with hydroxypyruvate, which serves as irreversible C2-donor, or by xylulokinase-catalyzed ATP-dependent phosphorylation of D-xylulose [100,101]. This latter approach has also been extended to the synthesis of L-xylulose-5-phosphate by using an enantiocomplementary xylulokinase [100,101]. In the D-tagatose catabolic pathways D-tagatose-6-phosphate 1-kinase-catalyzed phosphorylation of D-tagatose-6-phophate enabled the preparation of the central metabolite D-tagatose-1,6-diphosphate [102,103]. A characteristic metabolite for the Entner-Doudoroff pathway is 2-keto-3-deoxy-6-phosphogluconate, which can be synthesized in one step by eliminating water from 6-phosphogluconate using 6-phosphogluconate dehydratase [104]. In a similar way metabolites of other monosaccharide non-phosphorylative catabolic pathways can be synthesized in a straightforward way from the corresponding sugar acid, such as 2-keto-3-deoxy-D-galactonate from D-galactonate, 2-keto-3-deoxy-D-xylonate from D-xylonate using D-xylonate dehydratase [105]. D-gluconate dehydratase-catalyzed water elimination from D-gluconate allows the straightforward preparation of 2-keto-3-deoxy-D-gluconate [106]. In energy metabolism the high energy of the phosphorus-nitrogen-bond in phosphagens is a key energy source and selective biocatalytic synthesis enables straightforward access, such as the one step synthesis of Nω-phospho-L-arginine .
Biocatalytic methods are also very useful in synthesizing a number of key metabolites from other metabolic pathways. Shikimic acid-3-phosphate can be prepared by shikimate kinase-catalyzed ATP-dependent phosphorylation of shikimic acid and ATP regeneration using phosphoenolpyruvate and pyruvate kinase [109,110]. Pyridoxamine-5'-phosphate was synthesized from pyridoxal-5-phosphate by biocatalytic transamination using an ω-transaminase [111]. The biocatalytic L-arginine addition reaction to fumaric acid enabled an efficient one-step access to the urea cycle metabolite L-argininosuccinate [112,113].
Simple biocatalytic routes are also of special interest for disease-specific metabolites in order to support and simplify the diagnostics, for example of inborn errors of metabolism, cancers, cardiovascular and metabolic diseases.
3. Synthesis of Isotope-labelled Metabolites
The use of radioactive isotopes such as 3H (tritium), 14C or 32P has been instrumental for the discovery of major metabolic pathways, such as the path of carbon in photosynthesis [114]. Biocatalytic methods which have been developed for the synthesis of metabolites labelled with a radioactive isotope at a specific position, such as tritium- or 14C-labelled NAD+ or 14C-labelled nicotinamide riboside [115,116], can also be translated to methods for the synthesis of the corresponding metabolites labelled with a stable isotope at a specific position, such as 13C-labelled NAD+ or 13C-labelled nicotinamide riboside [116]. In contrast to radioactive labels, working with stable isotope labels such as the biogenic isotopes 2H (deuterium), 13C, 15N or 18O does not involve any health hazard and is not subject to regulations regarding radiation safety. The technology of isotope separation has enabled a continuous increase of the production of stable isotopes of light elements [117]. Compounds in which an atom like 1H, 12C, 14N, or 16O is replaced by a corresponding isotope with a higher atomic mass are of much interest to numerous applications, because the chemical structure and physical properties remain unchanged. The impressive advances of highly sensitive MS and NMR instrumentation, with its powerful methodologies and analyses with high information content, have shifted the interest to the use of the stable isotopes 2H, 13C, 15N or 18O [118,119,120]. As the isotope separation for the production of stable isotope labelled starting chemicals, with high chemical and isotopic purity, is demanding and requires highly specialized equipment and facilities, the precious stable isotope labelled starting materials should then be fully utilized for the synthesis of the desired metabolites. Therefore highly selective synthetic methods, which are able to efficiently incorporate to a high degree the stable isotope from the starting material into a defined position of the product and to completely convert the starting material to the target metabolite, are needed. The thermodynamics of biochemical reactions and the universe of biocatalysts provide a huge knowledge base from which suitable biocatalytic reactions for selective labelling with stable isotopes can be selected (see Figure 5).
Biocatalytic methods for site- and stereoselective deuteration are of much interest for short routes to deuterated metabolites, for example in the synthesis of selectively deuterated phosphatidyl-sn-glycerol, amino acids deuterated in the α- and/or β-position, deuterated NAD+/ NADH cofactors, or deuterated aldehydes [121,122,123,124,125,126]. The biocatalytic synthes of 3',4',5',5'-tetradeuterated metabolite 5-phospho-D-ribosyl α-1-pyrophosphate (PRPP) was prepared from 3',4',5',5'-tetradeuterated D-ribose by ribokinase-catalyzed phosphorylation and PRPP synthetase-catalyzed pyrophosphorylation [127]. The tetradeuterated PRPP was then converted by multistep enzymatic processes in high yields to the 3',4',5',5'-tetradeuterated nucleotides ATP, CTP, GTP and UTP [127].
The synthesis of 13C-labelled metabolites has been very useful for various metabolomics applications, such as for growing cells on media containing 13C-labelled carbon sources as nutrients [128], detailed investigations of cellular metabolism and the functional properties of complex metabolic networks by 13C-based metabolic flux analysis [26,129], or for overcoming matrix effects in accurate and reliable metabolite analyses and quantitative metabolomics [130]. Key for these applications and the discovery of novel biosynthetic pathways like the deoxyxylulose phosphate pathway [131] has been the synthesis of 13C-labelled biochemicals, such as 13C-labelled acetate or isotope isomers isotopomers) of D-glucose, where specific 12C-atoms are replaced by their 13C-isotopes. Enzymatic methods have facilitated access to 13C-labelled metabolites, such as monosaccharides [132] as well as 13C-labelled amino acids [133,134,135,136]. The use of very simple and inexpensive 13C-labelled precursors is thereby attractive, such as 13C-labelled pyruvate for the enzymatic synthesis of 13C-labelled aromatic amino acids [137]. Direct utilization of 13carbon dioxide is not only of much interest for investigating the metabolism of photosynthetic organisms, but also for the biocatalytic synthesis of 13C-labelled biochemicals, such as 13C-labelled L-malate [138].
As many natural products contain nitrogen, the introduction of the stable nitrogen isotope 15N is very useful for discovering natural products and characterizing its biosynthetic pathway and metabolic intermediates [139]. Biocatalytic synthesis of 15N-labelled metabolites has been achieved in a straightforward way by introducing the stable isotope from 15NH4 salts, by using biocatalytic systems with isolated enzymes or by making use of biosynthesis in whole cells growing in media containing 15NH4 salts. A very efficient NAD+-dependent amino acid dehydrogenase-catalyzed preparation of 15N-labelled L-serine, L-methionine and L-glutamic acid from the corresponding α-keto acids has been demonstrated using alanine dehydrogenase, leucine dehydrogenase and glutamate dehydrogenase, respectively, whereby NADH regeneration was done with the glucose/glucose dehydrogenase system [140]. The four 15N-labelled cobalamin standards hydroxocobalamin, adenosylcobalamin, methylcobalamin and cyanocobalamin have been prepared through biosynthesis by growing Propionibacterium freudenreichii whole cells in chemically defined medium containing (15NH4)2SO4 instead of (14NH4)2SO4 [141].
In addition to using a single type of stable isotope for labelling, biocatalytic synthesis has also been attractive for the introduction of more than one type of stable isotope. Pentose phosphate and purine pathway enzymes, together with biocatalytic regeneration cycles for the nucleoside triphosphate, folate, aspartate, glutamine, and NAD, have been utilized for labelling purine nucleotides with 13C and 15N [29]. This flexible and robust one-pot biocatalytic system enabled the preparation of uniformly 13C- and/or 15N-labelled GTP, or 13C-labelled ATP in the C2- and C8-position from labelled serine, ammonium, glucose and carbon dioxide [29].
4. Synthesis of Pharmaceutical Drug Metabolites
The development of new molecular entities for the effective treatment of human diseases with minimized side effects requires an understanding of its interactions with the biological cells of humans and their microbiome. The investigation of potential in vitro and in vivo pharmaceutical drug metabolism involving human and microbial enzymes, the biocatalytic reactions converting administered pharmaceutical drugs to derived metabolites, and the identity and biological activity of pharmaceutical drug metabolites, are of key importance for the treatment response, drug safety and side effects. The knowledge about pharmaceutical drug metabolism reactions, data on drug metabolizing enzymes, and structures of drug metabolites, has been growing very much over the years, as shown by its increasing numbers in the DrugBank database and its most recent version DrugBank 5.0 [142]. The complexity of pharmaceutical drug metabolism is increased further because pharmaceutical drugs not only interact with human metabolism but also with the human microbiome [143], as demonstrated by the biotransformation capabilities of the human gut microbiome towards numerous pharmaceutical drugs [144,145,146].
When applications of a pharmaceutical drug candidate show that it is enzymatically converted, from the site where it is administered to the desired drug action site, to a less active drug metabolite, this results in a poor treatment response and requires further drug development. Although pharmaceutical drug metabolism often leads to inactivation, there are also cases where therapeutic benefits may derive from biotransformation to a pharmacologically more active metabolite [147], for example when a prodrug with better cell permeability is enzymatically converted to the active pharmaceutical in diseased cells. Pharmacologically inactive, or less active by three orders of magnitude, small molecular weight compounds which are enzymatically converted in vivo to the active pharmaceutical have been developed by different paths, either being discovered by chance, from rescueing a drug discovery project, or by designing a prodrug [148]. Chemically reactive or toxic drug metabolites have received increased attention and the investigation of potential side effects, safety and toxicity issues of pharmaceutical drug metabolites which may be potentially be formed by biotransformations in the human body, has also evolved with the “Metabolites In Safety Testing" (MIST) guidance and the framework for identifying, quantifying, and assessing human drug metabolite safety [149,150,151]. As the investigations of the possible effects of such modified drugs require sufficient amounts of pure drug metabolites, straightforward methods for their synthetic access are highly desirable and can be highly significant for the timeline of projects. As selective chemical modification of complex drugs with stereocenters may be challenging [152], using biocatalysts which are involved in human drug metabolism provides an attractive selective approach [153].
When the orally active synthetic pharmaceutical drug dydrogesterone is used for treating progesterone deficiency and various gynecological conditions, human metabolism is responsible for the formation of the drug metabolite 20α-dihydrodydrogesterone. This drug metabolite has been prepared by an efficient stereo- and regioselective reduction of dydrogesterone (see Figure 6) catalyzed by recombinant human 20α-hydroxysteroid dehydrogenase AKR1C1 expressed in Schizosaccharomyces pombe [154]. With chemical reduction of the C20-keto group in dydrogesterone leading only to the 20β-dihydrodydrogeste-rone, biocatalytic reduction is key for obtaining 20α-dihydrodydrogesterone, which is also pharmacologically active [154]. The drug metabolite (S)-fingolimod-phosphate, which is a modulator of sphingosine 1-phosphate receptor 1, is formed in vivo by sphingosine kinase 2-catalyzed phosphorylation (see Figure 6) of the drug fingolimod, which has been approved as pharmaceutical drug for the therapy of multiple sclerosis in more than 80 countries [155]. As different forms of mycophenolic acid have various therapeutic applications as pharmaceutical drug, for example as immunosuppressant, its pharmacologically active metabolite mycophenolic acid acylglucuronide has attracted interest as it also inhibits inosine monophosphate dehydrogenase II like mycophenolic acid [156]. For the biocatalytic synthesis of the acylglucuronide of mycophenolic only the horse liver homogenate was found to catalyze the glucuronidation of mycophenolic acid, using UDP-glucuronic acid as donor, but the acylglucuronide was formed in a 1:1 mixture with the 7-O-glucuronide [156]. By optimization of the reaction temperature, the concentrations of the liver homogenate and the UDP-glucuronide the degree of conversion was increased to 54% and an acylglucuronide to 7-O-glucuronide ratio of 4.9:1 could be obtained, leading to the drug metabolite mycophenolic acid acylglucuronide (see Figure 6) in >95% purity and 34% isolated yield [156].
Biocatalytic transformations of synthetic pharmaceutical drug derivatives by human metabolism in vivo are important aspects in the design of prodrugs in order to overcome barriers in delivering and releasing the parent drug, and to improve cell permeability and water solubility [157].
Although many drug metabolites are pharmacologically inactive or much less active than the parent drug, such as for example the major mycophenolic acid metabolite in humans, the pharmacologically inactive 7-O-glucuronide of mycophenolic acid, synthetic access is important [158]. The 7-O-glucuronide of mycophenolic was obtained in 97% purity by biocatalytic glucuronidation of mycophenolic acid, using horse liver homogenate as biocatalyst and UDP-glucuronic acid as donor [156].
Chemically reactive drug metabolites which are formed by human metabolism can react with relevant molecules of biological cells and thereby lead to functional changes and adverse drug reactions [159]. In the drug metabolism of the nonsteroid anti-inflammatory drug diclofenac, which is used for treating rheumatoid disorders, the two metabolic pathways of the cytochrome P450-catalyzed oxidation to the corresponding chemically reactive quinone imine metabolites and the enzymatic glucuronidation are thought to be involved with several adverse drug reactions [160,161]. Cytochrome P450 enzymes have been shown to catalyze the drug conversion to chemically reactive metabolites which can cause toxic effects, such as the oxidation of acetaminophen to the toxic metabolite N-acetyl-p-benzoquinone imine catalyzed by human cytochrome P450 2E1, 1A2, and 3A4 [161,162,163].
5. Synthesis of Metabolite-like Compounds
The concepts of natural product-likeness [164,165] and metabolite-likeness [166,167] are attractive because the transport of natural products, nutrients and metabolites is omnipresent in the biological organisms. Natural-product-like and metabolite-like structures have been found to be present in a significant number of approved pharmaceutical drugs [168,169].
Variations of metabolite and natural products structures have attracted increasing interest due to the functional group differences which have been found to exist between natural products and synthetic molecules [170], the changing structural characteristics and properties of approved pharmaceutical drugs over time [171,172] and the interactions metabolites and natural products with target proteins, biosynthetic and transport proteins [173]. With the increasing knowledge about specific interactions between metabolites of the human microbiome and human disease-relevant biomolecules, such as ligand -receptor or enzyme inhibitor-enzyme interactions, the concept of utilizing microbial metabolites as a rich molecular space is very attractive for discovering metabolite-like compounds as novel pharmaceutical drugs for highly precise therapies and derisking of adverse drug reactions [174,175]. The development of metabolite-like compounds from bacterial tryptophan metabolism for discovering novel ligands, which bind directly to the pregnane X receptor and are not cytotoxic, is of much interest for further development and opens up a wide range of opportunities [176]. Excellent biocatalytic tools and methodologies have been developed for synthesizing non-natural chiral amino acids with high enantioselectivity, and the diversity of possible functional groups and chiral centers in non-natural amino acids is attractive for their use as fragments in small molecule pharmaceutical drugs (for some examples see Figure 7) for the therapy of diseases [177,178,179,180].
Combining biocatalytic with chemical reaction steps in chemoenzymatic syntheses offers new opportunities for overcoming challenges and diversifying metabolite structures. Advances have been achieved in the synthesis of diversified compounds of the plant metabolite cis-(+)-12-oxophytodienoic acid [181], a variety of non-natural nucleosides and nucleoside building blocks (see Figure 7) as well as in the synthesis of nucleoside analogue drugs by enzymatic cascades [182,183].
6. Discussion
The growing number of metabolite structures identified in biological organisms and the renewed interest in the elucidation of fundamental roles and useful properties of metabolites have also increased the necessity to develop analytical and synthetic methods and tools for a great diversity of metabolites. The identification of natural metabolic pathways has provided great starting points for the development of straightforward biocatalytic synthetic routes, inspired by nature, to naturally occurring metabolites, stable isotope-labelled metabolites, and drug metabolites. The molecular economy and reduced complexity of highly selective and protecting group-free biocatalytic reactions have enabled resource-efficient and robust production procedures to these metabolite classes. When the extraction yields of naturally occurring metabolites from biological resources are rather low and quite variable, or if challenging and lengthy chemical routes using protection-deprotection schemes require significant purification steps, biocatalytic production procedures using recombinant biocatalysts are a preferred choice. Biocatalytic systems offer particular benefits, not only for producing naturally occurring metabolites but also for isotope-labelled and drug metabolites. When manufacturing at larger scale is needed due to increasing demand, raw materials, resource efficiency, reliability issues have become increasingly important. Sustainability benefits can also be achieved when biological resources, which are rare and in danger becoming extinct, are preserved, and abundant bio-based resources are used for metabolite production using biocatalytic systems. Finally, the synthesis of non-natural metabolite-like compounds can also benefit from the power of biocatalysis.
8. Future Directions
Excellent opportunities appear for exploring uncharted territory regarding novel metabolites and their biological functions [184], natural product drugs [185,186], metabolite-like compounds. Discovering and characterizing novel enzyme functions and pathways from nature as well as engineering and evolving enzyme which catalyze reactions new to nature [187,188] are important for extending the frontiers of biocatalytic reactions in synthesis. Unlocking the power of enzymes can transform the synthesis of metabolites, natural products and non-natural small molecules derived thereof in various ways, from individual enzymatic reactions in chemoenzymatic synthesis to their full utilization in enzymatic total synthesis [189,190,191]. Many roads can lead to discovering novel enzymes from nature, such as guiding the correct assignment of enzyme functions to gene annotations and domains of unknown functions on genomes [192,193,194], unlocking natural product biosynthetic enzymes from metagenomes [195], or identifying missing enzymes in biosynthetic pathways to metabolites [196]. Mining microbial genomes for biosynthetic gene clusters [197] and deciphering precise genome-metabolome relationships of bacteria and fungi are very promising approaches for finding novel biosynthetic enzymes and pathways to novel metabolites [198,199,200,201]. Advanced metabolic engineering and synthetic biology tools and methodologies [202] and efficient gene expression to highly functional and fit-for-use enzymes are key for engineering novel biocatalytic pathways in viable and sustainable production processes of metabolites [203,204] and metabolite-like compounds of pharmaceutical interest [205,206,207]. Therefore, the future of synthesizing metabolites and metabolite-like compounds by using biocatalytic systems looks bright.
Funding
This research received no external funding.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The author declares no conflict of interest.
References
- Schreiber, S.L. Small molecules: the missing link in the central dogma. Nat. Chem. Biol. 2005, 1(2), 64–66. [Google Scholar] [CrossRef] [PubMed]
- McKnight, S.L. Back to the future: molecular biology meets metabolism. In: Cold Spring Harbor Symposia on Quantitative Biology 2011, 76, 403-411. Cold Spring Harbor Laboratory Press. [CrossRef]
- Kell, D.B.; Oliver, S. The metabolome 18 years on: a concept comes of age. Metabolomics 2016, 12, 148. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: the combination of targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2017, 114, 30.4.1–30.4.32. [Google Scholar] [CrossRef] [PubMed]
- Plumb, R.S.; Gethings, L.A.; Rainville, P.D.; Isaac, G.; Trengove, R.; King, A.M.; Wilson, I.D. Advances in high throughput LC/MS based metabolomics: A review. Trends Anal. Chem. 2023, 116954. [Google Scholar] [CrossRef]
- Emwas, A.H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; Wishart, D.S. NMR spectroscopy for metabolomics research. Metabolites 2019, 9(7), 123. [Google Scholar] [CrossRef]
- Guijas, C; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; Wolan, D.W.; Spilker, M.E.; Benton, H.P.; Siuzdak, G. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef]
- Wishart, D.S.; Cheng, L.L.; Copié, V.; Edison, A.S.; Eghbalnia, H.R.; Hoch, J.C.; Gouveia, G.J.; Pathmasiri, W.; Powers, R.; Schock, T.B.; Sumner, L.W. NMR and metabolomics—A roadmap for the future. Metabolites 2022, 12(8), 678. [Google Scholar] [CrossRef]
- Hoch, J.C.; Baskaran, K.; Burr, H.; Chin, J.; Eghbalnia, H.R.; Fujiwara, T.; Gryk, M.R.; Iwata, T.; Kojima, C.; Kurisu, G.; Maziuk, D.; Miyanoiri, Y.; Wedell, J.R.; Wilburn, C.; Yao, H.; Yokochi, M. Nucleic Acids Res. 2023, 51(D1), D368–D376. [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: back to metabolism in KEGG. Nucleic Acids Res. 2014, 42(D1), D199–D205. [Google Scholar] [CrossRef]
- Hastings, J.; Owen, G.; Dekker, A.; Ennis, M.; Kale, N.; Muthukrishnan, V.; Turner, S.; Swainston, N.; Mendes, P.; Steinbeck, C. ChEBI in 2016: Improved services and an expanding collection of metabolites. Nucleic Acids Res. 2016, 44(D1), D1214–D1219. [Google Scholar] [CrossRef]
- van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; Little, J.L.C.; Delgadillo, D.A.; Dorrestein, P.C.; Duncan, K.R.; Egan, J.M.; Galey, M.M.; Haeckl, F.P.J.; Hua, A.; Hughes, A.H.; Iskakova, D.; Khadilkar, A.; Lee, J.H.; Lee, S.; LeGrow, N.; Dennis, Y. Liu, D.Y.; Macho, J.M.; McCaughey, C.S.; Medema, M.H.; Neupane, R.P.; O’Donnell, T.J.; Paula, J.S.; Sanchez, L.M.; Shaikh, A.F.; Soldatou, S.; Terlouw, B.R.; Tran, T.A.; Valentine, M.; van der Hooft, J.J.J.; Vo, D.A.; Wang, M.; Wilson, D.; Zink, K.E.; Linington, R.G. The Natural Products Atlas: An Open Access Knowledge Base for Microbial Natural Products Discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [PubMed]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L; Berjanskii, M. HMDB 5.0: the human metabolome database for 2022. Nucleic Acids Res. 2022, 50(D1), D622–D631. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Oler, E.; Peters, H.; Guo, A.; Girod, S.; Han, S.; Saha, S.; Lui, V.W.; LeVatte, M.; Gautam, V.; Kaddurah-Daouk, R.; Karu, N. MiMeDB: the Human Microbial Metabolome Database. Nucleic Acids Res. 2023, 51(D1), D611–D620. [Google Scholar] [CrossRef]
- Sajed, T.; Marcu, A.; Ramirez, M.; Pon, A.; Guo, A.; Knox, C.; Wilson, M.; Grant, J.; Djoumbou, Y.; Wishart, D. ECMDB 2.0: A richer resource for understanding the biochemistry of E. coli. Nucleic Acids Res. 2016, 44(D1), D495–D501. [Google Scholar] [CrossRef]
- Huang, W.; Luke, K. Brewer, L.K.; Jace W. Jones, J.W.; Angela T. Nguyen, A.T.; Ana Marcu, A.; David S. Wishart, D.S.; Amanda G. Oglesby-Sherrouse, A.G.; Maureen A. Kane, M.A.; Angela Wilks, A. PAMDB: a comprehensive Pseudomonas aeruginosa metabolome database. Nucleic Acids Res. 2018, 46(D1), D575–D580. [Google Scholar] [CrossRef] [PubMed]
- Moumbock, A.F.A.; Gao, M.; Qaseem, A.; Li, J.; Kirchner, P.A.; Ndingkokhar, B.; Bekono, B.D.; Simoben, C.V.; Babiaka, S.M.; Malange, Y.I.; Sauter, F.; Zierep, P.; Ntie-Kang, F.; Günther, S. StreptomeDB 3.0: an updated compendium of streptomycetes natural products. Nucleic Acids Res. 2021, 49(D1), D600–D604. [Google Scholar] [CrossRef]
- Jones, M.R.; Pinto, E.; Torres, M.A.; Dörr, F.; Mazur-Marzec, H.; Szubert, K.; Tartaglione, L.; Dell’Aversano, C.; Miles, C.O.; Beach, D.G.; McCarron, P.; Sivonen, K.; Fewer, D.P.; Jokela, J.; Janssen, E.M.L. CyanoMetDB, a comprehensive public database of secondary metabolites from cyanobacteria. Water Res. 2021, 196, 117017. [Google Scholar] [CrossRef]
- Wang, D.G.; Wang, C.Y.; Hu, J.Q.; Wang, J.J.; Liu, W.C.; Zhang, W.J.; Du, X.R.; Wang, H.; Zhu, L.L.; Sui, H.Y.; Li, Y.Z.; Wu, C. Constructing a Myxobacterial Natural Product Database to Facilitate NMR-Based Metabolomics Bioprospecting of Myxobacteria. Anal. Chem. 2023, 95, 12, 5256–5266. [Google Scholar] [CrossRef]
- Ramirez-Gaona, M.; Marcu, A.; Pon, A.; Guo, A.C.; Sajed, T.; Wishart, N.A.; Karu, N.; Djoumbou Feunang, Y.; Arndt, D.; Wishart, D.S. YMDB 2.0: a significantly expanded version of the yeast metabolome database. Nucleic Acids Res. 2017, 45, D440–D445. [Google Scholar] [CrossRef]
- Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Piri-Moghadam, H.; Zheng, J.; Guo, A.; Li, C.; Guan, L.L.; Wishart, D.S. The Bovine Metabolome. Metabolites 2020, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Takeshi Ara, T.; Sakurai, N.; Takahashi, S.; Waki, N.; Suganuma, H.; Aizawa, K.; Matsumura, Y.; Kawada, T.; Shibata, D. TOMATOMET: A metabolome database consists of 7118 accurate mass values detected in mature fruits of 25 tomato cultivars. Plant Direct. 2021, 5, e00318. [Google Scholar] [CrossRef]
- Link, H.; Kochanowski, K; Sauer, U. Systematic identification of allosteric protein-metabolite interactions that control enzyme activity in vivo. Nat. Biotechnol. 2013, 31, 357–361. [Google Scholar] [CrossRef]
- Piazza, I.; Kochanowski, K.; Cappelletti, V.; Fuhrer, T.; Noor, E.; Sauer, U.; Picotti, P. A map of protein-metabolite interactions reveals principles of chemical communication. Cell 2018, 172(1), 358–372. [Google Scholar] [CrossRef] [PubMed]
- Rinschen, M.R.; Ivanisevic, J.; Giera, M.; Siuzdak, G. Identification of bioactive metabolites using activity metabolomics. Nat. Rev. Mol. Cell Biol. 2019, 20(6), 353–367. [Google Scholar] [CrossRef]
- Buescher, J.M.; Antoniewicz, M.R.; Boros, L.G.; Burgess, S.C.; Brunengraber, H.; Clish, C.B.; DeBerardinis, R.J.; Feron, O.; Frezza, C., Ghesquiere, B.; Gottlieb, E.; Hiller, K., Jones, R.G.; Kamphorst, J.J.; Kibbey, R.G.; Kimmelman, A.C.; Locasale, J.W.; Lunt, S.Y.; Maddocks, O.D.K.; Malloy, C.; Metallo, C.M., Meuillet, E.J.; Munger, J.; Nöh, K.; Rabinowitz, J.D.; Ralser, M.; Sauer, U.; Stephanopoulos, G.; St-Pierre, J.; Tennant, D.A.; Wittmann, C.; Vander Heiden, M.G.; Vazquez, A.; Vousden, K.; Young, J.D. Zamboni, N.; Fendt, S.M. A roadmap for interpreting 13C metabolite labeling patterns from cells. Curr. Opin. Biotechnol. 2015, 34, 189–201. [CrossRef] [PubMed]
- Di Yu, D.; Zhou, L.; Liu, X.; Xu, G. Stable isotope-resolved metabolomics based on mass spectrometry: Methods and their applications. Trends Anal. Chem. 2023, 160, 116985. [Google Scholar] [CrossRef]
- Han, S.; Van Treuren, W.; Fischer, C.R.; Merrill, B.D.; DeFelice, B.C.; Sanchez, J.M.; Higginbottom, S.K.; Guthrie, L.; Fall, L.A.; Dodd, D.; Fischbach, M.A.; Sonnenburg, J.I. A metabolomics pipeline for the mechanistic interrogation of the gut microbiome. Nature 2021, 595, 415–420. [Google Scholar] [CrossRef]
- Schultheisz, H.L.; Szymczyna, B.R.; Scott, L.G.; Williamson, J.R. Pathway Engineered Enzymatic de novo Purine Nucleotide Synthesis. ACS Chem Biol. 2008, 3(8), 499–511. [Google Scholar] [CrossRef]
- Rowbotham, J.S.; Ramirez, M.A.; Lenz, O.; Reeve, H.A.; Vincent, K.A. Bringing biocatalytic deuteration into the toolbox of asymmetric isotopic labelling techniques. Nat. Commun. 2020, 11(1), 1454. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Snyder, S.A. Chasing molecules that were never there: misassigned natural products and the role of chemical synthesis in modern structure elucidation. Angew. Chem. Int. Ed. 2005, 44(7), 1012–1044. [Google Scholar] [CrossRef] [PubMed]
- Sunazuka, T.; Hirose, T.; Omura, S. Efficient Total Synthesis of Novel Bioactive Microbial Metabolites. Acc. Chem. Res. 2008, 41(2), 302–314. [Google Scholar] [CrossRef]
- Nicolaou, K.C. Organic synthesis: the art and science of replicating the molecules of living nature and creating others like them in the laboratory. Proc. R. Soc. A 2014, 470, 20130690. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Route Selection and Reaction Engineering for Sustainable Metabolite Synthesis. React. Chem. Eng. 2023, Advance Article. [Google Scholar] [CrossRef]
- Demain, A.L. From natural products discovery to commercialization: a success story. J. Ind. Microbiol. Biotechnol. 2006, 33, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Hoff, B.; Plassmeier, J.; Blankschien, M.; Letzel, A.C.; Kourtz, L.; Schröder, H.; Koch, W.; Zelder, O. Unlocking Nature's Biosynthetic Power—Metabolic Engineering for the Fermentative Production of Chemicals. Angew. Chem. Int. Ed. 2021, 60(5), 2258–2278. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Eun, H.; Prabowo, C.P.S.; Cho, S.; Lee, S.Y. Metabolic and cellular engineering for the production of natural products. Curr. Opin. Biotechnol. 2022, 77, 102760. [Google Scholar] [CrossRef]
- Wohlgemuth, R. 2022. Selective biocatalytic defunctionalization of raw materials. ChemSusChem 2022, 15(9), e202200402. [Google Scholar] [CrossRef]
- Kaiser, R. Scent of the Vanishing Flora. ISBN 13: 978-3-906390-64-2, Wiley-VHCA AG, Zürich, Switzerland, 2010.
- Walsh, C.T.; Tang, Y. Natural product biosynthesis – Chemical logic and enzymatic machinery. Royal Society of Chemistry, London, UK, 2017.
- Alcántara, A.R.; Dominguez de Maria, P.; Littlechild, J.A.; Schürmann, M.; Sheldon, R.A.; Wohlgemuth, R. Biocatalysis as key to sustainable industrial chemistry. ChemSusChem 2022, 15(9), e202102709. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Tools and ingredients for the biocatalytic synthesis of metabolites. Biotechnol. J. 2009, 4(9), 1253–1265. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Horizons of systems biocatalysis and renaissance of metabolite synthesis. Biotechnol. J. 2018, 13(6), 1700620. [Google Scholar] [CrossRef] [PubMed]
- Oberg, N.; Zallot, R.; Gerlt, J.A. EFI-EST, EFI-GNT, and EFI-CGFP: Enzyme Function Initiative (EFI) Web Resource for Genomic Enzymology Tools. J. Mol. Biol. 2023, 435, 168018. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, C.; Liao, J.C. (Eds.) Industrial Biotechnology: Microorganism. First Edition, Wiley-VCH, Weinheim, Germany, 2017. ISBN: 978-3-527-34179-5.
- Wittmann, C.; Liao, J.C. (Eds.) Industrial Biotechnology: Products and Processes. First Edition, Wiley-VCH, Weinheim, Germany, 2017. ISBN: 978-3-527-34181-8.
- Lee, S.Y.; Nielsen, J.; Stephanopoulos, G. (Eds.) Metabolic Engineering - Concepts and Applications. First Edition, Wiley-VCH, Weinheim, Germany, 2021. ISBN: 978-3-527-34662-2.
- Flickinger, M.C. (Ed.) Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology. Vol. 1-7, John Wliey & Sons, Hoboken, New Jersey, USA, 2010. ISBN: 978-0-471-79930-6.
- Wohlgemuth, R.; Littlechild, J. Complexity reduction and opportunities in the design, integration and intensification of biocata-lytic processes for metabolite synthesis. Front. Bioeng. Biotechnol. 2022, 10, 958606. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Biocatalysis–Key enabling tools from biocatalytic one-step and multi-step reactions to biocatalytic total synthesis. New Biotechnol. 2021, 60, 113–123. [Google Scholar] [CrossRef]
- Wentrup, C. Origins of Organic Chemistry and Organic Synthesis. Eur. J. Org. Chem. 2022, e202101492. [Google Scholar] [CrossRef]
- Wöhler, F. Ueber künstliche Bildung des Harnstoffs. Ann. Phys. 1828, 88(2), 253–256. [Google Scholar] [CrossRef]
- Kolbe, H. Beiträge zur Kenntnis der gepaarten Verbindungen. Ann. Chem. Pharm. 1845, 54(2), 145–188. [Google Scholar] [CrossRef]
- Sheehan, J.C.; Henery-Logan, K.R. The total synthesis of penicillin V. J. Am. Chem. Soc. 1957, 79, 5, 1262–1263. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Vourloumis, D.; Winssinger, N.; Baran, P.S. The art and science of total synthesis at the dawn of the twenty-first century. Angew. Chem. Int. Ed. 2000, 39(1), 44–122. [Google Scholar] [CrossRef]
- Veitch, G.E.; Boyer, A.; Ley, S.V. The Azadirachtin Story. Angew. Chem. Int. Ed. 2008, 47, 9402–9429. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Rigol, S. Perspectives from nearly five decades of total synthesis of natural products and their analogues for biology and medicine. Nat. Prod. Rep. 2020, 37(11), 1404–1435. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Han, J.C.; Zhang, W.; Gu, C.C.; Zou, Y.P.; Li, C.C. Strategies and Lessons Learned from Total Synthesis of Taxol. Chem. Rev. 2023, 123, 8, 4934–4971. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Snyder, S.A. Chasing molecules that were never there: misassigned natural products and the role of chemical synthesis in modern structure elucidation. Angew. Chem. Int. Ed. 2005, 44, 1012–1044. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.S.; Pitts, C.R.; McClymont, K.S.; Stratton, T.P.; Bi, C.; Baran, P.S. Ideality in Context: Motivations for Total Synthesis. Acc. Chem. Res. 2021, 54, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Cardwell, H.M.E.; Cornforth, J.W.; Duff, S.R.; Holtermann, H.; Robinson, R. Total synthesis of androgenic hormones. Chem. Ind. 1951, 20, 389–90. [Google Scholar]
- Woodward, R.B.; Sondheimer, F.; Taub, D. The total synthesis of cholesterol. J. Am. Chem. Soc. 1951, 73(7), 3548–3548. [Google Scholar] [CrossRef]
- Eschenmoser, A.; Wintner, C.E. Natural Product Synthesis and Vitamin B12: Total synthesis of vitamin B12 provided a framework for exploration in several areas of organic chemistry. Science 1977, 196(4297), 1410–1420. [Google Scholar] [CrossRef]
- Woodward, R.B. The total synthesis of vitamin B12. Pure Appl. Chem. 1973, 33(1), 145–178. [Google Scholar] [CrossRef] [PubMed]
- Kishi, Y. Palytoxin: an inexhaustible source of inspiration - personal perspective. Tetrahedron 2002, 58, 6239–6258. [Google Scholar] [CrossRef]
- Sears, J.E.; Boger, D.L. Total Synthesis of Vinblastine, Related Natural Products, and Key Analogues and Development of Inspired Methodology Suitable for the Systematic Study of Their Structure−Function Properties. Acc. Chem. Res. 2015, 48, 653–662. [Google Scholar] [CrossRef]
- Baran, P.S.; Maimone, T.J.; Richter, J.M. Total synthesis of marine natural products without using protecting groups. Nature 2007, 446, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Pasteur, L. Mémoire sur la fermentation appelée lactique. Comptes Rendus Chimie 1857, 45, 913–916. [Google Scholar]
- Buchner, E. Alkoholische Gährung ohne Hefezellen. Berichte der Deutschen Chemischen Gesellschaft 1897, 30, 117–124. [Google Scholar] [CrossRef]
- Krebs, H.A.; Henseleit, K. Untersuchungen über die Harnstoffbildung im Tierkörper. Klin. Wochenschr. 1932, 11, 757–759. [Google Scholar] [CrossRef]
- Krebs, H.A., Henseleit, K. Untersuchungen über die Harnstoffbildung im Tierkörper. II. Klin. Wochenschr. 1932, 11, 1137–1139. [CrossRef]
- Amato, A.; Becci, A.; Beolchini, F. Citric acid bioproduction: the technological innovation change. Crit. Rev. Biotechnol. 2020, 40(2), 199–212. [Google Scholar] [CrossRef] [PubMed]
- Elander, R.P. Industrial production of β-lactam antibiotics. Appl. Microbiol. Biotechnol. 2003, 61, 385–392. [Google Scholar] [CrossRef]
- Sanchez, S.; Rodríguez-Sanoja, R.; Ramos, A.; Demain. A.L. Our microbes not only produce antibiotics, they also overproduce amino acids. J. Antibiotics 2018, 71, 26–36. [Google Scholar] [CrossRef]
- Vandamme, E.J.; Revuelta, J.L. Industrial Biotechnology of Vitamins, Biopigments, and Antioxidants. Wiley-VCH, Weinheim, Germany, 2016. [CrossRef]
- Meyer, H.P.; Robins, K.T. Large scale bioprocess for the production of optically pure L-carnitine. Monatsh. Chemie 2005, 136, 1269–1277. [Google Scholar] [CrossRef]
- Ramírez-Rendon, D.; Passari, A.K.; Ruiz-Villafán, B.; Rodríguez-Sanoja, R.; Sánchez, S.; Demain, A.L. Impact of novel microbial secondary metabolites on the pharma industry. Appl. Microbiol. Biotechnol. 2022, 106, 1855–1878. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83(3), 770–803. [Google Scholar] [CrossRef] [PubMed]
- Bringi, V.; Kadkade, P.G.; Prince, C.L.; Roach, B.L. Enhanced production of taxol and taxanes by cell cultures of Taxus species. US 2013/0017582 A1, 2013.
- Sharma, A.; Bhatia, S.K.; Banyal, A.; Chanana, I.; Kumar, A.; Chand, D.; Kulshrestha, S.; Kumar, P. An Overview on Taxol Production Technology and Its Applications as Anticancer Agent. Biotechnol. Bioprocess Engin. 2022, 27, 706–728. [Google Scholar] [CrossRef]
- Omura, S. A Splendid Gift from the Earth: The Origins and Impact of the Avermectins (Nobel Lecture). Angew. Chem. Int. Ed. 2016, 55, 10190–10209. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y. Artemisinin—A Gift from Traditional Chinese Medicine to the World (Nobel Lecture). Angew. Chem. Int. Ed. 2016, 55, 10210–10226. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, M.A.; Walsh, C.T. Antibiotics for Emerging Pathogens. Science 2009, 325(5944), 1089–1093. [Google Scholar] [CrossRef]
- Lewis, K. The Science of Antibiotic Discovery. Cell 2020, 181, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B., Moreira, R.; Li, Y.; Luzhetskyy, A.; Medema, M.H.; Pernodet, J.L.; Stadler, M.; Tormo, J.R.; Genilloud, O.; Truman, A.W.; Weissman, K.J.; Takano, E.; Sabatini, S.; Stegmann, E.; Brötz-Oesterhelt, H.; Wohlleben, W.; Seemann, M.; Empting, M.; Hirsch, A.K.H.; Loretz, B.; Lehr, C.M.; Titz, A.; Herrmann, J.; Jaeger, T.; Alt, S.; Hesterkamp, T.; Winterhalter, M.; Schiefer, A.; Pfarr, K.; Hoerauf, A.; Graz, H.; Graz, M.; Lindvall, M.; Ramurthy, S.; Karlén, A.; van Dongen, M.; Petkovic, H.; Keller, A.; Peyrane, F.; Stefano Donadio, S.; Fraisse, L.; Piddock, L.J.V.; Gilbert, I.H.; Moser, H.E.; Müller, R.. Towards the sustainable discovery and development of new antibiotics. Nat Rev Chem 2021, 5, 726–749. [CrossRef]
- Meyer, H.P.; Eichhorn, E.; Hanlon, S.; Lütz, S.; Schürmann, M.; Wohlgemuth, R.; Coppolecchia, R. The use of enzymes in organic synthesis and the life sciences: perspectives from the Swiss Industrial Biocatalysis Consortium (SIBC). Cat. Sci. Technol. 2013, 3(1), 29–40. [Google Scholar] [CrossRef]
- Eichhorn, E.; Locher, E.; Guillemer, S.; Wahler, D.; Fourage, L.; Schilling, B. 2018. Biocatalytic process for (−)-Ambrox production using squalene hopene cyclase. Adv. Synth. Cat. 2018, 360(12), 2339–2351. [Google Scholar] [CrossRef]
- Calvillo, A.; Pellicer, T.; Carnicer, M.; Planas, A. Bioprocess Strategies for Vitamin B12 Production by Microbial Fermentation and Its Market Applications. Bioengineering 2022, 9, 365. [Google Scholar] [CrossRef]
- Bühlmann, P.; Pretsch, E.; Bakker, E. Carrier-Based Ion-Selective Electrodes and Bulk Optodes. 2. Ionophores for Potentiometric and Optical Sensors. Chem. Rev. 1998, 98, 1593−1687. [Google Scholar] [CrossRef]
- Jani, P.; Emmert, J.; Wohlgemuth, R. Process analysis of macrotetrolide biosynthesis during fermentation by means of direct infusion LC-MS. Biotechnol. J. 2008, 3(2), 202–208. [Google Scholar] [CrossRef] [PubMed]
- Cuartero, M.; Colozza, N.; Fernández-Pérez, B.M.; Crespo, G.A. Why ammonium detection is particularly challenging but insightful with ionophore-based potentiometric sensors – an overview of the progress in the last 20 years. Analyst 2020, 145, 3188–3210. [Google Scholar] [CrossRef] [PubMed]
- Gauss, D.; Schoenenberger, B.; Wohlgemuth, R. Chemical and enzymatic methodologies for the synthesis of enantiomerically pure glyceraldehyde 3-phosphates. Carbohydrate Res. 2014, 389, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Molla, G.S.; Kinfu, B.M.; Chow, J.; Streit, W.; Wohlgemuth, R.; Liese, A. Bioreaction engineering leading to efficient synthesis of L-glyceraldehyd-3-phosphate. Biotechnol. J. 2017, 12(3), 1600625. [Google Scholar] [CrossRef]
- Richter, N., Neumann, M., Liese, A., Wohlgemuth, R., Eggert, T. and Hummel, W., 2009. Characterisation of a Recombinant NADP-Dependent Glycerol Dehydrogenase from Gluconobacter oxydans and its Application in the Production of L-Glyceraldehyde. ChemBioChem 2009, 10(11), 1888–1896. [CrossRef]
- Richter, N.; Neumann, M.; Liese, A.; Wohlgemuth, R.; Weckbecker, A.; Eggert, T.; Hummel, W. Characterization of a whole-cell catalyst co-expressing glycerol dehydrogenase and glucose dehydrogenase and its application in the synthesis of L-glyceraldehyde. Biotechnol. Bioeng. 2010, 106(4), 541–552. [Google Scholar] [CrossRef] [PubMed]
- Gauss, D.; Sánchez-Moreno, I.; Oroz-Guinea, I.; García-Junceda, E.; Wohlgemuth, R. Phosphorylation catalyzed by dihydroxyacetone kinase. Eur. J. Org. Chem. 2018, 23, 2892–2895. [Google Scholar] [CrossRef]
- Hardt, N.; Kinfu, B.M.; Chow, J.; Schoenenberger, B.; Streit, W.R.; Obkircher, M.; Wohlgemuth, R. Biocatalytic Asymmetric Phosphorylation Catalyzed by Recombinant Glycerate-2-Kinase. ChemBioChem 2017, 18(15), 1518–1522. [Google Scholar] [CrossRef] [PubMed]
- Shaeri, J.; Wohlgemuth, R.; Woodley, J.M. Semiquantitative process screening for the biocatalytic synthesis of D-xylulose-5-phosphate. Org. Proc. Res. Dev. 2006, 10(3), 605–610. [Google Scholar] [CrossRef]
- Shaeri, J.; Wright, I.; Rathbone, E.B.; Wohlgemuth, R.; Woodley, J.M. Characterization of enzymatic D-xylulose 5-phosphate synthesis. Biotechnol. Bioeng. 2008, 101(4), 761–767. [Google Scholar] [CrossRef] [PubMed]
- Hardt, N.; Kind, S.; Schoenenberger, B.; Eggert, T.; Obkircher, M.; Wohlgemuth, R. Facile synthesis of D-xylulose-5-phosphate and L-xylulose-5-phosphate by xylulokinase-catalyzed phosphorylation. Biocat. Biotrans. 2020, 38(1), 35–45. [Google Scholar] [CrossRef]
- Hardt, N.; Kind, S.; Schoenenberger, B.; Eggert, T.; Obkircher, M.; Wohlgemuth, R. Kinase-Catalysed Phosphorylations of Xylulose Substrates and Synthesis of Xylulose-5-Phosphate Enantiomers. In: Applied Biocatalysis, Whittall, J.; Sutton, P.W. (Eds.) John Wiley & Sons, Hoboken, USA, 2021, 397-401.
- Schoenenberger, B.; Kind, S.; Meier, R.; Eggert, T.; Obkircher, M.; Wohlgemuth, R. Efficient biocatalytic synthesis of D-tagatose 1, 6-diphosphate by LacC-catalysed phosphorylation of D-tagatose 6-phosphate. Biocat. Biotrans. 2020, 38(1), 53–63. [Google Scholar] [CrossRef]
- Schoenenberger, B.; Kind, S.; Meier, R.; Eggert, T.; Obkircher, M.; Wohlgemuth, R. Kinase-Catalysed Phosphorylations of Ketohexose Phosphates and LacC-Catalysed Synthesis of D-Tagatose-1, 6-Diphosphate Lithium Salt. In: Applied Biocatalysis, Whittall, J.; Sutton, P.W. (Eds.) John Wiley & Sons, Hoboken, USA, 2021, 393-397.
- Krevet, S.; Shen, L.; Bohnen, T.; Schoenenberger, B.; Meier, R.; Obkircher, M.; Bangert, K.; Koehling, R.; Allenspach, E.; Wohlgemuth, R.; Siebers, B. Enzymatic synthesis of 2-keto-3-deoxy-6-phosphogluconate by the 6-phosphogluconate-dehydratase from Caulobacter crescentus. Front. Bioeng. Biotechnol. 2020, 8, 185. [Google Scholar] [CrossRef]
- Shen, L.; Kohlhaas, M.; Enoki, J.; Meier, R.; Schönenberger, B.; Wohlgemuth, R.; Kourist, R.; Niemeyer, F.; van Niekerk, D.; Bräsen, C.; Niemeyer, J. Snoep, J.; Siebers, B. A combined experimental and modelling approach for the Weimberg pathway optimisation. Nat. Commun. 2020, 11(1), 1098. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Köhling, R.; Schönenberger, B.; Kouril, T.; Esser, D.; Bräsen, C.; Siebers, B.; Wohlgemuth, R. One-step synthesis of 2-keto-3-deoxy-D-gluconate by biocatalytic dehydration of D-gluconate. J. Biotechnol. 2014, 191, 69–77. [Google Scholar] [CrossRef]
- Schoenenberger, B.; Wszolek, A.; Milesi, T.; Brundiek, H.; Obkircher, M.; Wohlgemuth, R. Synthesis of Nω-Phospho-L-arginine by Biocatalytic Phosphorylation of L-Arginine. ChemCatChem 2017, 9(1), 121–126. [Google Scholar] [CrossRef]
- Schoenenberger, B.; Wszolek, A.; Milesi, T.; Brundiek, H.; Obkircher, M.; Wohlgemuth, R. Phosphoramidates by Kinase-Catalysed Phosphorylation and Arginine Kinase-Catalysed Synthesis of N𝛚-Phospho-L-Arginine. In: Applied Biocatalysis, Whittall, J.; Sutton, P.W. (Eds.) John Wiley & Sons, Hoboken, USA, 2021, 401-407.
- Schoenenberger, B.; Wszolek, A.; Meier, R.; Brundiek, H.; Obkircher, M.; Wohlgemuth, R. Recombinant AroL-Catalyzed Phosphorylation for the Efficient Synthesis of Shikimic Acid 3-Phosphate. Biotechnol. J. 2018, 13(8), 1700529. [Google Scholar] [CrossRef]
- Schoenenberger, B.; Wszolek, A.; Meier, R.; Brundiek, H.; Obkircher, M.; Wohlgemuth, R. Shikimate Kinase-Catalysed Phosphorylations and Synthesis of Shikimic Acid 3-Phosphate by AroL-Catalysed Phosphorylation of Shikimic Acid. Applied Biocatalysis, Whittall, J.; Sutton, P.W. (Eds.) John Wiley & Sons, Hoboken, USA, 2021, 386-393.
- Schell, U.; Wohlgemuth, R.; Ward, J.M. Synthesis of pyridoxamine 5′-phosphate using an MBA: pyruvate transaminase as biocatalyst. J. Mol. Catal. B: Enzymatic 2009, 59(4), 279–285. [Google Scholar] [CrossRef]
- Schoenenberger, B.; Wszolek, A.; Meier, R.; Brundiek, H.; Obkircher, M.; Wohlgemuth, R. Biocatalytic asymmetric Michael addition reaction of l-arginine to fumarate for the green synthesis of N-(([(4S)-4-amino-4-carboxy-butyl] amino) iminomethyl)-L-aspartic acid lithium salt (L-argininosuccinic acid lithium salt). RSC Adv. 2017, 7(77), 48952–48957. [Google Scholar] [CrossRef]
- Schoenenberger, B.; Wszolek, A.; Meier, R.; Brundiek, H.; Obkircher, M.; Wohlgemuth, R. Biocatalytic Asymmetric Aza-Michael Addition Reactions and Synthesis of L-Argininosuccinate by Argininosuccinate Lyase ARG4-Catalysed Aza-Michael Addition of L-Arginine to Fumarate. In: Applied Biocatalysis, Whittall, J.; Sutton, P.W. (Eds.) John Wiley & Sons, Hoboken, USA, 2021, 204-210.
- Calvin, M. The Path of Carbon in Photosynthesis: The carbon cycle is a tool for exploring chemical biodynamics and the mechanism of quantum conversion. Science 1962, 135(3507), 879–889. [Google Scholar] [CrossRef]
- Rising, K.A.; Schramm, V.L. Enzymatic Synthesis of NAD+ with the Specific Incorporation of Atomic Labels. J. Am. Chem. Soc. 1994, 116, 6531–6536. [Google Scholar] [CrossRef]
- Tran, A.; Yokose, R.; Cen, Y. Chemo-enzymatic synthesis of isotopically labelled nicotinamide riboside. Org. Biomol. Chem. 2018, 16, 3662–3671. [Google Scholar] [CrossRef]
- Khoroshilov, A.V. Production of stable isotopes of light elements: past, present and future. J. Phys.: Conf. Ser. 2018, 1099, 012002. [Google Scholar] [CrossRef]
- Fan, T.W.M.; Lane, A.N. Applications of NMR spectroscopy to systems biochemistry. Progr. NMR Spectroscopy 2016, 92–93, 18–53. [Google Scholar] [CrossRef] [PubMed]
- Yu, D., Zhou, L., Liu, X. and Xu, G. Stable isotope-resolved metabolomics based on mass spectrometry: Methods and their applications. Trends Anal. Chem. 2023, 160, 116985. [CrossRef]
- Giraudeau, P. Quantitative NMR spectroscopy of complex mixtures. Chem. Commun., 2023, 59, 6627–6642. [Google Scholar] [CrossRef]
- Wohlgemuth, R.; Waespe-Sarcevic, N.; Seelig, J. Bilayers of Phosphatidylglycerol. A Deuterium and Phosphorus NuclearMagnetic Resonance Study of the Head-Group Region. Biochemistry 1980, 19, 3315–3321. [Google Scholar] [CrossRef]
- Chun, S.W.; Narayan, A.R.H. Biocatalytic, Stereoselective Deuteration of α-Amino Acids and Methyl Esters. ACS Catal. 2020, 10(13), 7413–7418. [Google Scholar] [CrossRef]
- Doyon, T.J.; Buller, A.R. Site-Selective Deuteration of Amino Acids through Dual-Protein Catalysis. J. Am. Chem. Soc. 2022, 144, 16, 7327–7336. [Google Scholar] [CrossRef]
- Rowbotham, J.S.; Ramirez, M.A.; Lenz, O.; Reeve, H.A.; Vincent, K.A. Bringing biocatalytic deuteration into the toolbox of asymmetric isotopic labelling techniques. Nat. Commun. 2020, 11, 1454. [Google Scholar] [CrossRef]
- Rowbotham, J.S.; Hardy, A.P.; Reeve, H.A.; Vincent, K.A. Synthesis of [4S-2H] NADH, [4R-2H] NADH, [4-2H2] NADH and [4-2H] NAD+ cofactors through heterogeneous biocatalysis in heavy water J. Label. Compd. Radiopharm. 2021, 64, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lou, Y.; Wang, L.; Wang, Z.; Xu, W.; Ma, W.; Chen, Z.; Chen, X.; Wu, Q. Rational Design of Biocatalytic Deuteration Platform of Aldehydes. ACS Catal. 2021, 11, 21, 13348–13354. [Google Scholar] [CrossRef]
- Tolbert, T.J.; Williamson, J.R. Preparation of Specifically Deuterated RNA for NMR Studies Using a Combination of Chemical and Enzymatic Synthesis. J. Am. Chem. Soc. 1996, 118, 7929–7940. [Google Scholar] [CrossRef]
- Bennett, B.D.; Yuan, J.; Kimball, E.H.; Rabinowitz, J.D. Absolute quantitation of intracellular metabolite concentrations by an isotope ratio-based approach. Nat. Protoc. 2008, 3(8), 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Sauer, U. Metabolic networks in motion: 13C-based flux analysis. Molecular Systems Biol. 2006, 2, 62. [Google Scholar] [CrossRef] [PubMed]
- Arrivault, S.; Guenther, M.; Fry, S.C.; Fuenfgeld, M.F.F.F.; Veyel, D.; Mettler-Altmann, T.; Stitt, M.; Lunn, J.E. Synthesis and Use of Stable-Isotope-Labelled Internal Standards for Quantification of Phosphorylated Metabolites by LC–MS/MS. Anal. Chem. 2015, 87, 13, 6896–6904. [Google Scholar] [CrossRef]
- Eisenreich, W.; Schwarz, M.; Cartayrade, A.; Arigoni, D.; Zenk, M.H.; Bacher, A. The deoxyxylulose phosphate pathway of terpenoid biosynthesis in plants and microorganisms. Chem. Biol. 1998, 5(9), R221–R233. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, S.; Serianni, A.S. Labeling monosaccharides with stable isotopes. Methods Enzymol. 2015, 565, 423–458. [Google Scholar] [CrossRef] [PubMed]
- Goux, W.J.; Rench, L.; Weber, D.S. Stereoselective synthesis of stable isotope labeled L-α-amino acids: The enzymatic preparation of 13C-labeled L-glutamic acids. J. Label. Compd. Radiopharm. 1993, 33(3), 181–193. [Google Scholar] [CrossRef]
- Maeda, H.; Takata, K.; Toyoda, A.; Niitsu, T.; Iwakura, M.; Shibata, K. Production of L-[3-13C] serine from [13C] formaldehyde and glycine using an enzyme system combined with tetrahydrofolate regeneration. J. Ferment. Bioeng. 1997, 83(1), 113–115. [Google Scholar] [CrossRef]
- Jemielity, J.; Kańska, M.; Kański, R. Enzymatic Synthesis of [1-13C]-and [1-14C]-L-Phenyl-Alanine. Isot. Environ. Health Stud. 1998, 34(4), 335–339. [Google Scholar] [CrossRef]
- Akita, H.; Suzuki, H.; Doi, K.; Ohshima, T. Efficient synthesis of D-branched-chain amino acids and their labeled compounds with stable isotopes using D-amino acid dehydrogenase. Appl. Microbiol. Biotechnol. 2014, 98, 1135–1143. [Google Scholar] [CrossRef]
- Van Raad, D.; Huber, T.; Otting, G. Improved spectral resolution of [13C,1H]-HSQC spectra of aromatic amino acid residues in proteins produced by cell-free synthesis from inexpensive 13C-labelled precursors. J. Biomol. NMR 2023. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, Y.; Wang, Q.; Wang, X.; Li, Q.; Liu, W.; Zhao, Z.K. Non-natural Cofactor and Formate-Driven Reductive Carboxylation of Pyruvate. Angew. Chem. Int. Ed. 2020, 59(8), 3143–3146. [Google Scholar] [CrossRef]
- Morgan, K.D. The use of nitrogen-15 in microbial natural product discovery and biosynthetic characterization. Front. Microbiol. 2023, 14, 1174591. [Google Scholar] [CrossRef]
- Chiriaca, M.; Lupan, I.; Popa, F.; Palibroda, N.; Popescu, O. Enzymatic synthesis of some 15N-labelled L-amino acids. Isot. Environ. Health Stud. 2010, 46(2), 249–254. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Asam, S.; Chen, J.; Ehrmann, M.; Rychlik, M. Production of Four 15N-Labelled Cobalamins via Biosynthesis Using Propionibacterium freudenreichii. Front. Microbiol. 2021, 12, 713321. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; Assempour, N.; Iynkkaran, I.; Liu, Y.; Maciejewski, A.; Gale, N.; Wilson, A.; Chin, L.; Cummings, R.; Le, D.; Pon, A.; Knox, C.; Wilson, M. DrugBank 5.0: a major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46(D1), D1074–D1082. [Google Scholar] [CrossRef]
- Clayton, T.A.; Baker, D.; Lindon, J.C.; Everett, J.R.; Nicholson, J.K. Pharmacometabonomic identification of a significant host-microbiome metabolic interaction affecting human drug metabolism. Proc. Natl. Acad. Sci. 2009, 106(34), 14728–14733. [Google Scholar] [CrossRef]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A. L. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Javdan, B.; Lopez, J.G.; Chankhamjon, P.; Lee, Y.C.J.; Hull, R.; Wu, Q.; Wang, X.; Chatterjee, S.; Donia, M.S. Personalized mapping of drug metabolism by the human gut microbiome. Cell 2020, 181, 1661–1679.e22. [Google Scholar] [CrossRef] [PubMed]
- Heinken, A.; Hertel, J.; Acharya, G.; Ravcheev, D.A.; Nyga, M.; Okpala, O.E.; Hogan, M.; Magnúsdóttir, S.; Martinelli, F.; Nap, B.; Preciat, G.; Edirisinghe, J.N.; Henry, C.S.; Fleming, R.M.T.; Thiele, I. Genome-scale metabolic reconstruction of 7,302 human microorganisms for personalized medicine. Nat. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Fura, A.; Shu, Y.Z.; Zhu, M.; Hanson, R.L.; Roongta, V.; Humphreys, W.G. Discovering Drugs through Biological Transformation: Role of Pharmacologically Active Metabolites in Drug Discovery. J. Med. Chem. 2004, 47(18), 4339–4351. [Google Scholar] [CrossRef]
- Rautio, J.; Meanwell, N.A.; Di, L.; Hageman, M.J. The expanding role of prodrugs in contemporary drug design and development. Nat. Rev. Drug Discov. 2018, 17, 559–587. [Google Scholar] [CrossRef]
- Schadt, S.; Bister, B.; Chowdhury, S.K.; Funk, C.; Hop, C.E.C.A.; Humphreys, W.G.; Igarashi, F.; James, A.D.; Kagan, M.; Khojasteh, S.C.; Nedderman, A.N.R.; Prakash, C.; Runge, F.; Scheible, H.; Spracklin, D.K.; Swart, P.; Tse, S.; Yuan, J.; Obach, R.S. A Decade in the MIST: Learnings from Investigations of Drug Metabolites in Drug Development under the “Metabolites in Safety Testing” Regulatory Guidance. Drug Metab. Dispos. 2018, 46(6), 865–878. [Google Scholar] [CrossRef] [PubMed]
- US Food and Drug Administration (FDA), Safety Testing of Drug Metabolites, 2020. https://www.fda.gov/media/72279/download.
- Luffer-Atlas, D.; Obach, R.S.; Smith, D.A. A MIST conception: what has been learned from twenty years of human metabolite safety assessment? Med. Chem. Res. 2023. [Google Scholar] [CrossRef]
- Chhatrapati Bisen, A.; Nashik Sanap, S.; Agrawal, S.; Biswas, A.; Sankar Bhatta, R. Chemical metabolite synthesis and profiling: Mimicking in vivo biotransformation reactions. Bioorganic Chemistry 2023, 139, 106722. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.; Geier, M.; Hanlon, S.P.; Nidetzky, B.; Glieder, A. Human enzymes for organic synthesis. Angew. Chem. Int. Ed. 2018, 57(41), 13406–13423. [Google Scholar] [CrossRef] [PubMed]
- Naumann, J.M.; Zöllner, A.; Drăgan, C.A.; Messinger, J.; Adam, J.; Bureik, M. Biotechnological Production of 20-alpha-Dihydrodydrogesterone at Pilot Scale. Appl. Biochem. Biotechnol. 2011, 165, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Billich, A.; Baumruker, T.; Heining, P.; Schmouder, R.; Francis, G.; Aradhye, S.; Burtin, P. Fingolimod (FTY720): discovery and development of an oral drug to treat multiple sclerosis. Nat. Rev. Drug Discov. 2010, 9(11), 883–897. [Google Scholar] [CrossRef] [PubMed]
- Kittelmann, M.; Rheinegger, U.; Espigat, A.; Oberer, L.; Aichholz, R.; Francotte, E.; Ghisalba, O. Preparative Enzymatic Synthesis of the Acylglucuronide of Mycophenolic Acid. Adv. Synth. Catal. 2003, 345, 825–829. [Google Scholar] [CrossRef]
- Rautio, J.; Kumpulainen, H.; Heimbach, T.; Oliyai, R.; Oh, D.; Järvinen, T.; Savolainen, J. Prodrugs: design and clinical applications. Nat. Rev. Drug Discov. 2008, 7, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Wolff, N.A.; Burckhardt, B.C.; Burckhardt, G.; Oellerich, M.; Armstrong, V.W. Mycophenolic acid (MPA) and its glucuronide metabolites interact with transport systems responsible for excretion of organic anions in the basolateral membrane of the human kidney. Nephrol. Dial. Transplant. 2007, 22, 2497–2503. [Google Scholar] [CrossRef]
- Park, B.K.; Boobis, A.; Clarke, S.; Goldring, C.E.P.; Jones, D.; Kenna, J.G.; Lambert, C.; Laverty, H.G.; Naisbitt, D.J.; Nelson, S.; Nicoll-Griffith, D.A.; Obach, R.S.; Routledge, P.; Smith, D.A.; Tweedie, D.J.; Vermeulen, N.; Williams, D.P.; Wilson, I.D.; Baillie, T.A. Managing the challenge of chemically reactive metabolites in drug development. Nat. Rev. Drug Discov. 2011, 10, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Tateishi, Y.; Ohe, T.; Ogawa, M.; Takahashi, K.; Nakamura, S.; Mashino, T. Development of Novel Diclofenac Analogs Designed to Avoid Metabolic Activation and Hepatocyte Toxicity. ACS Omega 2020, 5, 32608–32616. [Google Scholar] [CrossRef]
- Guengerich, F.P. A history of the roles of cytochrome P450 enzymes in the toxicity of drugs. Toxicol. Res. 2021, 37, 1–23. [Google Scholar] [CrossRef]
- Dahlin, D.C.; Miwa, G.T.; Lu, A.Y.; Nelson, S.D. N-acetyl-p-benzoquinone imine: a cytochrome P-450-mediated oxidation pro-duct of acetaminophen. Proc. Natl. Acad. Sci. 1984, 81(5), 1327–1331. [Google Scholar] [CrossRef]
- Bender, R.P.; Lindsey, Jr., R.H.; Burden, D.A.; Osheroff, N. N-Acetyl-p-benzoquinone Imine, the Toxic Metabolite of Acetaminophen, Is a Topoisomerase II Poison. Biochemistry 2004, 43, 3731–3739. [CrossRef]
- Ertl, P.; Roggo, S.; Schuffenhauer, A. Natural Product-likeness Score and Its Application for Prioritization of Compound Libraries. J. Chem. Inf. Model, 2008; 48 (1), 168–174. [Google Scholar] [CrossRef]
- Tay, D.W.P.; Yeo, N.Z.X.; Adaikkappan, K.; Lim, Y.H.; Ang, S.J. 67 million natural product-like compound database generated via molecular language processing. Sci Data 2023, 10(1), 296. [Google Scholar] [CrossRef]
- Dobson, P.D.; Patel, Y.; Kell, D.B. Metabolite-likeness’ as a criterion in the design and selection of pharmaceutical drug libraries. Drug Discovery Today 2009, 14(1–2), 31–40. [Google Scholar] [CrossRef] [PubMed]
- O'Hagan, S.; Kell, D.B. Understanding the foundations of the structural similarities between marketed drugs and endogenous human metabolites. Front. Pharmacol. 2015, 6, 105. [Google Scholar] [CrossRef] [PubMed]
- O′Hagan, S.; Swainston, N.; Handl, J.; Kell, D.B. A ‘rule of 0.5’ for the metabolite-likeness of approved pharmaceutical drugs. Metabolomics 2015, 11, 323–339. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83(3), 770–803. [Google Scholar] [CrossRef]
- Ertl, P. Substituents of life: The most common substituent patterns present in natural products. Bioorg. Med. Chem. 54 (2022) 116562. [CrossRef] [PubMed]
- Shultz, M.D. Two Decades under the Influence of the Rule of Five and the Changing Properties of Approved Oral Drugs. J. Med. Chem. 2019, 62, 1701–1714. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.G.; Wobst, J.J. A Decade of FDA-Approved Drugs (2010−2019): Trends and Future Directions. J. Med. Chem. 2021, 64, 2312–2338. [Google Scholar] [CrossRef]
- Young, R.J.; Flitsch, S.L.; Grigalunas, M.; Leeson, P.D.; Quinn, R.J.; Turner, N.J.; Waldmann, H. The Time and Place for Nature in Drug Discovery. JACS Au 2022, 2, 2400–2416. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Rajpal, D.K.; Brown, J.R. Human microbial metabolites as a source of new drugs. Drug Discov. Today 2016, 21(4), 692–698. [Google Scholar] [CrossRef]
- Li, H.; Ranhotra, H.S.; Mani, S.; Dvořák, Z.; Sokol, H.; Müller, R. Human microbial metabolite mimicry as a strategy to expand the chemical space of potential drugs. Drug Discov. Today 2020, 25(9), 1575–1579. [Google Scholar] [CrossRef]
- Dvorák, Z.; Kopp, F.; Costello, C.M.; Kemp, J.S.; Li, H.; Vrzalová, A.; Stepánková, M.; Bartonková, I.; Jiskrová, E.; Poulíková, K.; Vyhlídalová, B.; Nordstroem, L.U.; Karunaratne, C.V.; Ranhotra, H.S.; Mun, K.S.; Naren, A.P.; Murray, I.A.; Perdew, G.H.; Brtko, J.; Toporova, L.; Schön, A.; Wallace, B.D.; Walton, W.G.; Redinbo, M.R.; Sun, K.; Beck, A.; Kortagere, S.; Neary, M.C.; Chandran, A.; Vishveshwara, S.; Cavalluzzi, M.M.; Lentini, G.; Cui, J.Y.; Gu, H.; March, J.C.; Chatterjee, S.; Matson, A.; Wright, D.; Flannigan, K.L.; Hirota, S.A.; Sartor, R.B.; Mani, S. Targeting the pregnane X receptor using microbial metabolite mimicry. EMBO Mol. Med. 2020, 12, e11621. [Google Scholar] [CrossRef]
- Xue, Y.P.; Cao, C.H.; Zheng, Y.G. Enzymatic asymmetric synthesis of chiral amino acids. Chem. Soc. Rev. 2018, 47, 1516–1561. [Google Scholar] [CrossRef] [PubMed]
- Herger, M., van Roye, P., Romney, D.K., Brinkmann-Chen, S., Buller, A.R. and Arnold, F.H. Synthesis of β-branched tryptophan analogues using an engineered subunit of tryptophan synthase. J. Am. Chem. Soc. 2016, 138(27), 8388–8391. [CrossRef]
- Ellis, J.M.; Campbell, M.E.; Kumar, P.; Geunes, E.P.; Bingman, C.A.; Buller, A.R. Biocatalytic synthesis of non-standard amino acids by a decarboxylative aldol reaction. Nat. Catal. 2022, 5, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Alfonzo, E.; Das, A.; Arnold, F.H. New Additions to the Arsenal of Biocatalysts for Noncanonical Amino Acid Synthesis. Curr. Opin. Green Sustain. Chem. 2022, 38, 100701. [Google Scholar] [CrossRef] [PubMed]
- Löwe, J.; Dietz, K.J.; Gröger, H. From a Biosynthetic Pathway toward a Biocatalytic Process and Chemocatalytic Modifications: Three-Step Enzymatic Cascade to the Plant Metabolite cis-(+)-12-OPDA and Metathesis-Derived Products. Adv. Sci. 2020, 7(13), 1902973. [Google Scholar] [CrossRef]
- Westarp, S.; Kaspar, F.; Neubauer, P.; Kurreck, A. Industrial potential of the enzymatic synthesis of nucleoside analogs: existing challenges and perspectives. Curr. Opin. Biotechnol. 2022, 78, 102829. [Google Scholar] [CrossRef]
- Cosgrove, S.C.; Miller, G.J. Advances in biocatalytic and chemoenzymatic synthesis of nucleoside analogues, Expert Opin. Drug Discov. 2022, 17(4), 355–364. [Google Scholar] [CrossRef]
- Trung, M.N.; Kieninger, S.; Fandi, Z.; Qiu, D.; Liu, G.; Mehendale, N.K.; Saiardi, A.; Jessen, H.; Keller, B.; Fiedler, D. Stable Isotopomers of myo-Inositol Uncover a Complex MINPP1-Dependent Inositol Phosphate Network. ACS Cent. Sci. 2022, 8(12), 1683–1694. [Google Scholar] [CrossRef]
- Shen, B. A New Golden Age of Natural Products Drug Discovery. Cell 2015, 163, 1297–1300. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G. Atanasov, A.G., Zotchev, S.B., Dirsch, V.M. and Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20(3), 200–216. [Google Scholar] [CrossRef]
- Reetz, M.T.; Sun, Z.; Qu, G. Enzyme Engineering: Selective Catalysts for Applications in Biotechnology, Organic Chemistry, and Life Science. Wile-VCH, Weinhem, Germany, 2023. ISBN 978-3-527-35033-9.
- Chen, K.; Arnold, F.H. Engineering new catalytic activities in enzymes. Nat. Catal. 2020, 3(3), 203–213. [Google Scholar] [CrossRef]
- Zetzsche, L.E.; Chakrabarty, S.; Narayan, A.R. The transformative power of biocatalysis in convergent synthesis. J. Am. Chem. Soc. 2022, 144(12), 5214–5225. [Google Scholar] [CrossRef]
- Stout, C.N.; Wasfy, N.M.; Chen, F.; Renata, H. Charting the Evolution of Chemoenzymatic Strategies in the Syntheses of Complex Natural Products. J. Am. Chem. Soc. 2023. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Deng, H.; Renata, H. Chemoenzymatic approaches for exploring structure–activity relationship studies of bioactive natural products. Nat. Synth. 2023, 2, 708–718. [Google Scholar] [CrossRef]
- Hermann, J.C., Marti-Arbona, R., Fedorov, A.A., Fedorov, E., Almo, S.C., Shoichet, B.K. and Raushel, F.M. Structure-based activity prediction for an enzyme of unknown function. Nature 2007, 448(7155), 775–779.
- Zallot, R.; Oberg, N.; Gerlt, J.A. The EFI web resource for genomic enzymology tools: leveraging protein, genome, and metagenome databases to discover novel enzymes and metabolic pathways. Biochemistry 2019, 58(41), 4169–4182. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Wetmore, K.M.; Waters, R.J.; Callaghan, M.; Ray, J.; Liu, H.; Kuehl, J.V.; Melnyk, R.A.; Lamson, J.S.; Suh, Y.; Carlson, H.K.; Esquivel, Z.; Sadeeshkumar, H.; Chakraborty, R.; Zane, G.M.; Rubin, B.E.; Wall, J.D.; Visel, A.; Bristow, J.; Blow, M.J.; Arkin, A.P.; Deutschbauer, A.M. Mutant phenotypes for thousands of bacterial genes of unknown function. Nature 2018, 557, 503–509. [Google Scholar] [CrossRef]
- Robinson, S.L.; Piel, J.; Sunagawa, S. A roadmap for metagenomic enzyme discovery. Nat. Prod. Rep., 2021, 38, 1994–2023. [Google Scholar] [CrossRef]
- Caputi, L.; Franke, J.; Farrow, S.C.; Chung, K.; Payne, R.M.; Nguyen, T.D.; Dang, T.T.T.; Soares Teto Carqueijeiro, I.; Koudounas, K.; Dugé de Bernonville, T.; Ameyaw, B.; Jones, D.M.; Vieira, I.J.C.; Courdavault, V.; O’Connor, S.E. Missing enzymes in the biosynthesis of the anticancer drug vinblastine in Madagascar periwinkle. Science 2018, 360(6394), 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 7.0: new and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef] [PubMed]
- Caesar, L.K.; Montaser, R.; Keller, N.P.; Kelleher, N.L. Metabolomics and genomics in natural products research: complementary tools for targeting new chemical entities. Nat. Prod. Rep. 2021, 38(11), 2041–2065. [Google Scholar] [CrossRef]
- Scherlach, K.; Hertweck, C. Mining and unearthing hidden biosynthetic potential. Nat. Commun. 2021, 12(1), 3864. [Google Scholar] [CrossRef] [PubMed]
- Klapper, M.; Hübner, A.; Ibrahim, A.; Wasmuth, I.; Borry, M.; Haensch, V.G.; Zhang, S.; Al-Jammal, W.K.; Suma, H.; Fellows Yates, J.A.; Frangenberg, J.; Velsko, I.M.; Chowdhury, S.; Herbst, R.; Bratovanov, E.V.; Dahse, H.M.; Horch, T.; Hertweck, C.; González Morales, M.R.; Straus, L.G.; Vilotijevic, I.; Warinner, C.; Stallforth, P. Natural products from reconstructed bacterial genomes of the Middle and Upper Paleolithic. Science 2023, 380(6645), 619–624. [Google Scholar] [CrossRef]
- Caesar, L.K.; Butun, F.A.; Robey, M.T.; Ayon, N.J.; Gupta, R.; Dainko, D.; Bok, J.W.; Nickles, G.; Stankey, R.J.; Johnson, D.; Mead, D.; Cank, K.B.; Earp, C.E.; Raja, H.A.; Oberlies, N.H.; Keller, N.P.; Kelleher, N.L. Correlative metabologenomics of 110 fungi reveals metabolite–gene cluster pairs. Nat. Chem. Biol. 2023, 19, 846–854. [Google Scholar] [CrossRef]
- Smanski, M.J.; Zhou, H.; Claesen, J.; Shen, B.; Fischbach, M.; Voigt, C.A. Synthetic biology to access and expand nature’s chemical diversity. Nat. Rev. Microbiol. 2016, 14(3), 135–149. [Google Scholar] [CrossRef]
- Erb, T.J.; Patrik R Jones, P.R.; Bar-Even, A. Synthetic metabolism: metabolic engineering meets enzyme design. Curr. Opin. Chem. Biol. 2017, 37, 56–62. [Google Scholar] [CrossRef]
- Yi, J.; Li, Z. Artificial multi-enzyme cascades for natural product synthesis. Curr. Opin. Biotechnol. 2022, 78, 102831. [Google Scholar] [CrossRef]
- Huffman, M.A.; Fryszkowska, A.; Alvizo, O.; Borra-Garske, M.; Campos, K.R.; Canada, K.A.; Devine, P.N.; Duan, D.; Forstater, J.H.; Grosser, S.T.; Halsey, H.M.; Hughes, G.J.; Jo, J.; Joyce, L.A.; Kolev, J.N.; Liang, J.; Maloney, K.M.; Mann, B.F.; Marshall, N.M.; McLaughlin, M.; Moore, J.C.; Murphy, G.S.; Nawrat, C.C.; Nazor, J.; Novick, S.; Patel, N.R.; Rodriguez-Granillo, A.; Robaire, S.A.; Sherer, E.C.; Truppo, M.D.; Whittaker, A.M.; Verma, D.; Xiao, L.; Xu, Y.; Yang, H. Design of an in vitro biocatalytic cascade for the manufacture of islatravir. Science 2019, 366(6470), 1255–1259. [Google Scholar] [CrossRef]
- McIntosh, J.A.; Benkovics, T.; Silverman, S.M.; Huffman, M.A.; Kong, J.; Maligres, P.E.; Itoh, T.; Yang, H.; Verma, D.; Pan, W.; Ho, H.I.; Vroom, J.; Knight, A.M.; Hurtak, J.A.; Klapars, A.; Fryszkowska, A.; Morris, W.J.; Strotman, N.A.; Murphy, G.S.; Maloney, K.M.; Fier, P.S. Engineered ribosyl-1-kinase enables concise synthesis of molnupiravir, an antiviral for COVID-19. ACS Cent. Sci. 2021, 7(12), 1980–1985. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.A.; Liu, Z.; Andresen, B.M.; Marzijarani, N.S.; Moore, J.C.; Marshall, N.M.; Borra-Garske, M.; Obligacion, J.V.; Fier, P.S.; Peng, F.; Forstater, J.H.; Winston, M.S.; An, C.; Chang, W.; Lim, J.; Huffman, M.A.; Miller, S.P.; Tsay, F.R.; Altman, M.D.; Lesburg, C.A.; Steinhuebel, D.; Trotter, B.W.; Cumming, J.N.; Northrup, A.; Bu, X.; Mann, B.F.; Biba, M.; Hiraga, K.; Murphy, G.S.; Kolev, J.N.; Makarewicz, A.; Pan, W.; Farasat, I.; Bade, R.S.; Stone, K.; Duan, D.; Alvizo, O.; Adpressa, D.; Guetschow, E.; Hoyt, E.; Regalado, E.L.; Castro, S.; Rivera, N.; Smith, J.P.; Wang, F.; Crespo, A.; Verma, D.; Axnanda, S.; Dance, Z.E.X.; Devine, P.N.; Tschaen, D.; Canada, K.A.; Bulger, P.G.; Sherry, B.D.; Truppo, M.D.; Ruck, R.T.; Campeau, L.C.; Bennett, D.J.; Humphrey, G.R.; Campos, K.R.; Maddess, M.L. A kinase-cGAS cascade to synthesize a therapeutic STING activator. Nature 2022, 603(7901), 439–444. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Selected reasons for the synthesis of metabolites and metabolite-like compounds A) for basic and applied sciences, and B) for industry and medicine.
Figure 1.
Selected reasons for the synthesis of metabolites and metabolite-like compounds A) for basic and applied sciences, and B) for industry and medicine.
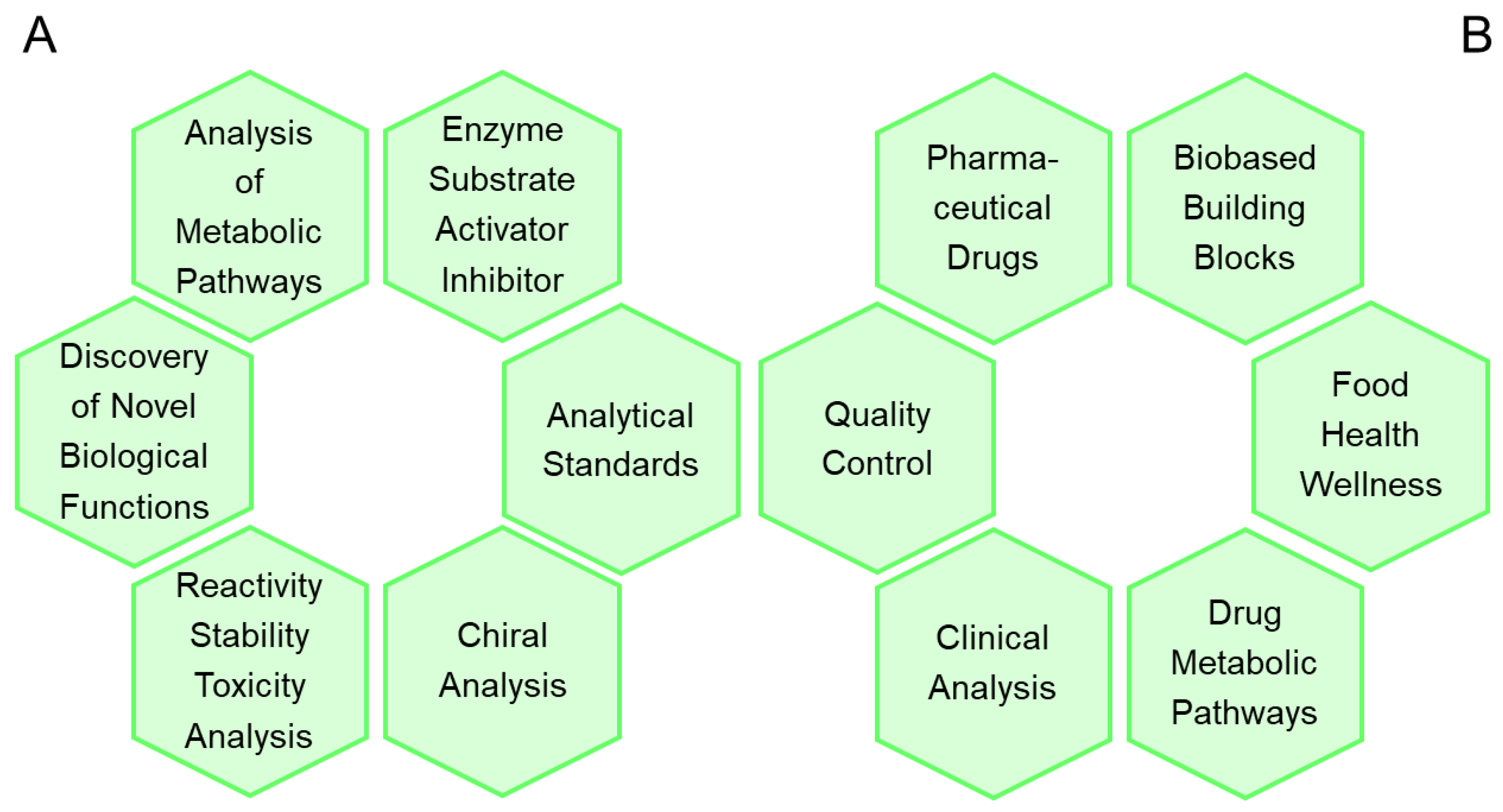
Figure 2.
Basic types of biocatalytic systems for the synthesis of metabolites and metabolite-like compounds.
Figure 2.
Basic types of biocatalytic systems for the synthesis of metabolites and metabolite-like compounds.

Figure 3.
Plant cell fermentation for the industrial production of paclitaxel.
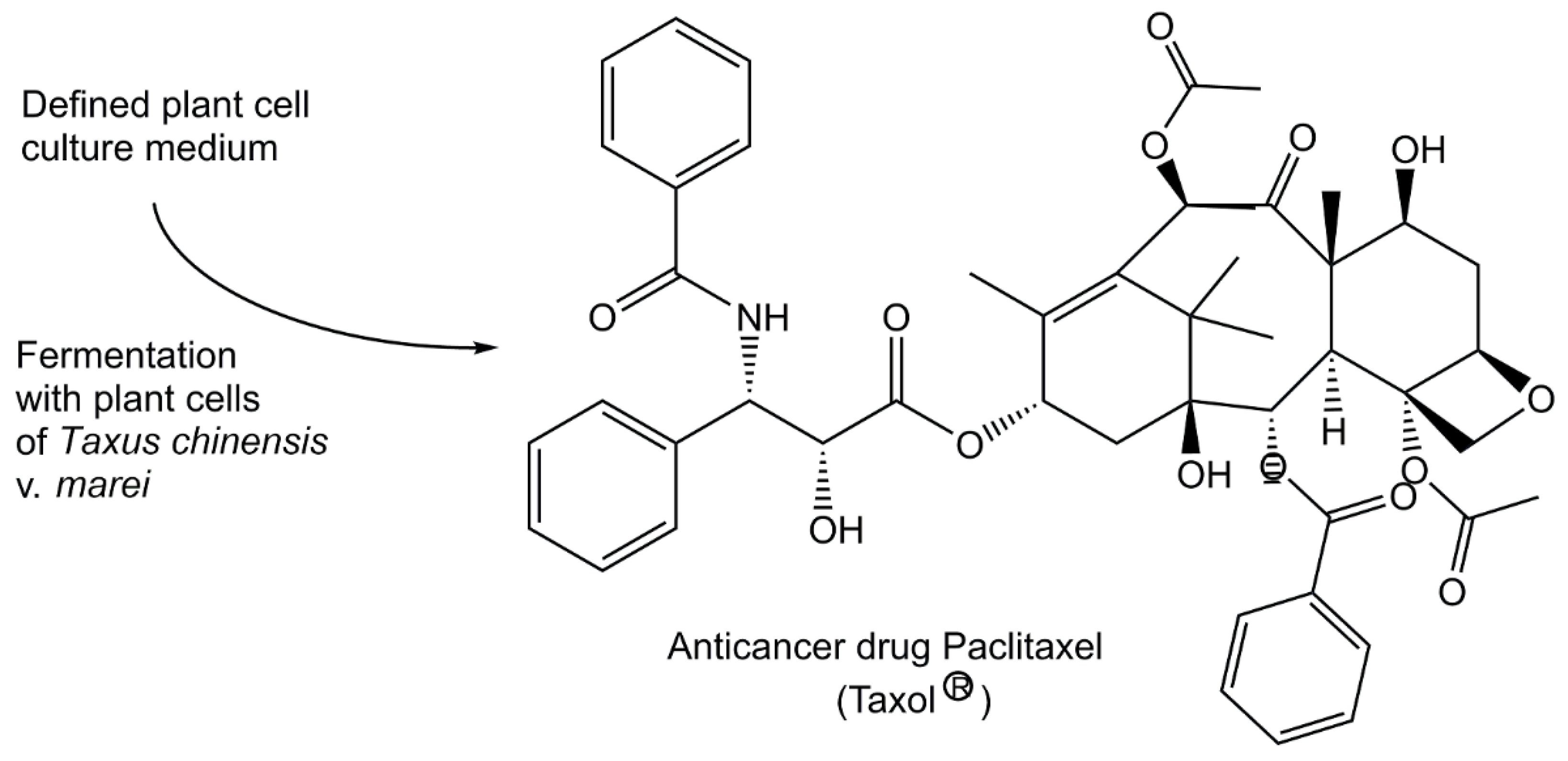
Figure 4.
Enzymatic process for the industrial production of the fragrance ingredient (-)-Ambrox.
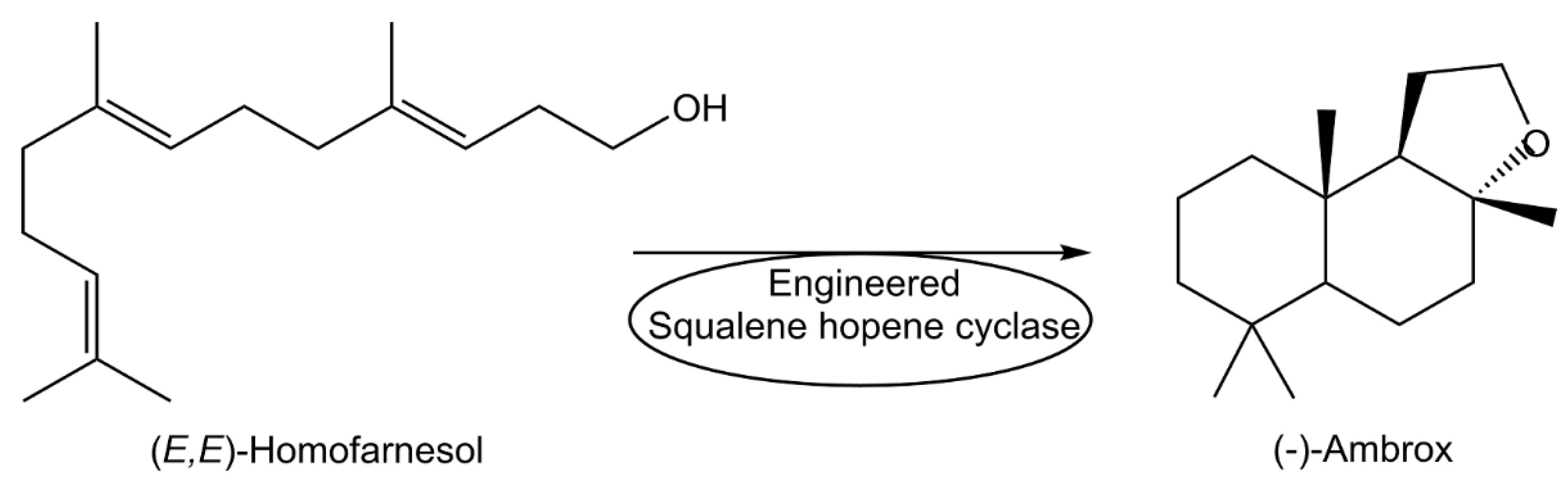
Figure 5.
Selected biocatalytic systems for the synthesis of metabolites labelled with 2H-, 13C- or 15N-stable isotopes.
Figure 5.
Selected biocatalytic systems for the synthesis of metabolites labelled with 2H-, 13C- or 15N-stable isotopes.
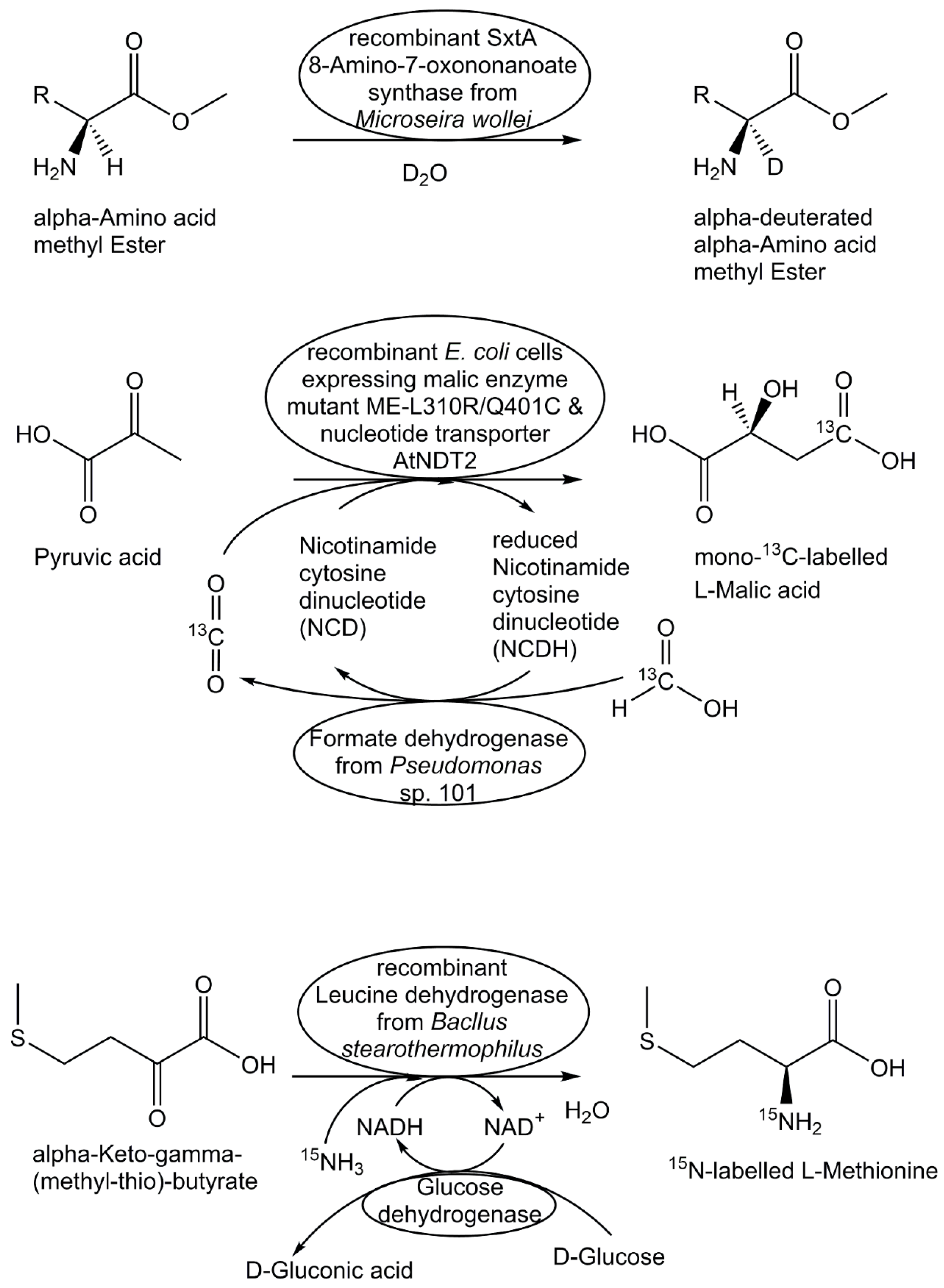
Figure 6.
Selected biocatalytic systems for pharmaceutical drug metabolite synthesis.
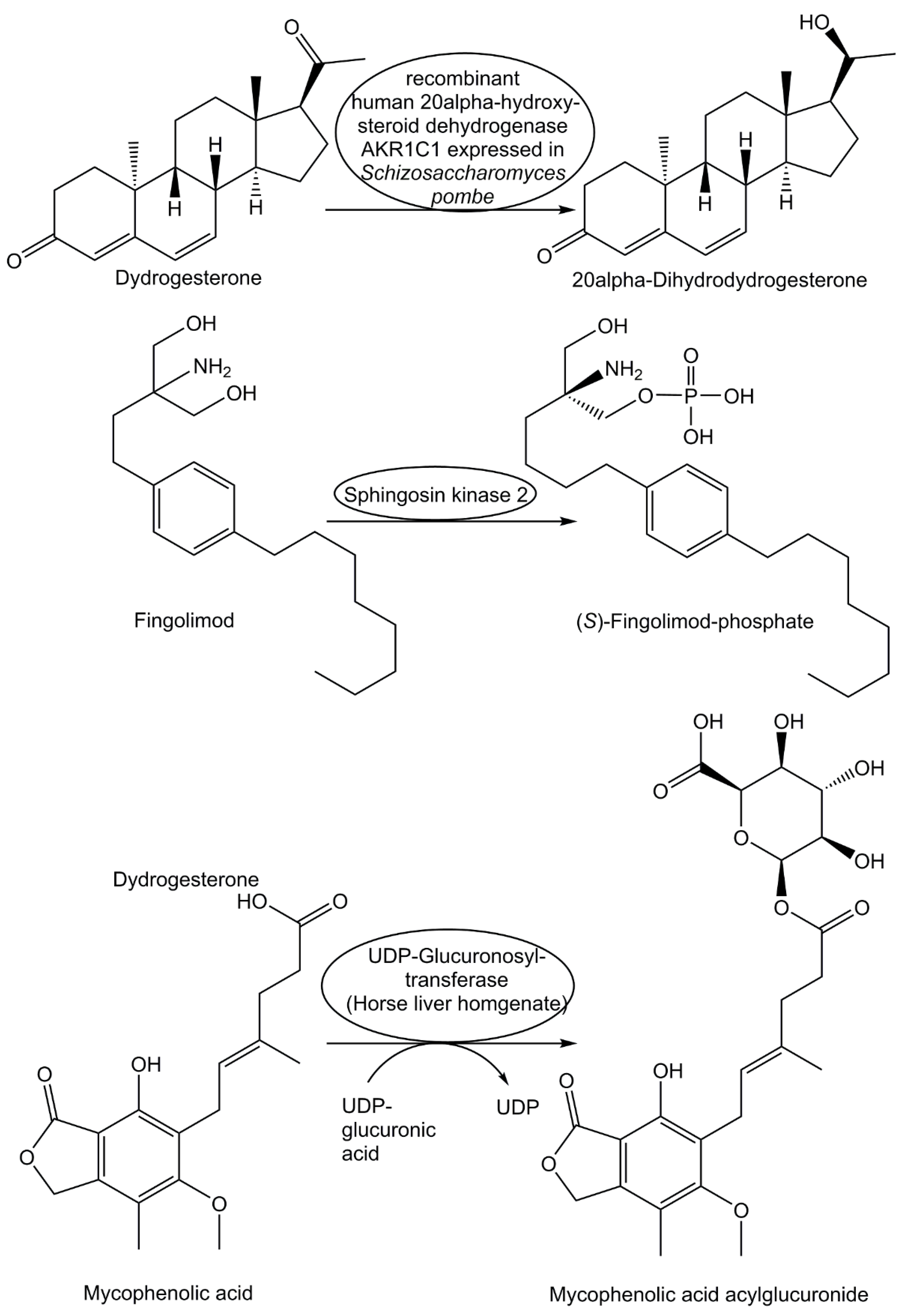
Figure 7.
Biocatalytic synthesis of selected metabolite-like compounds.
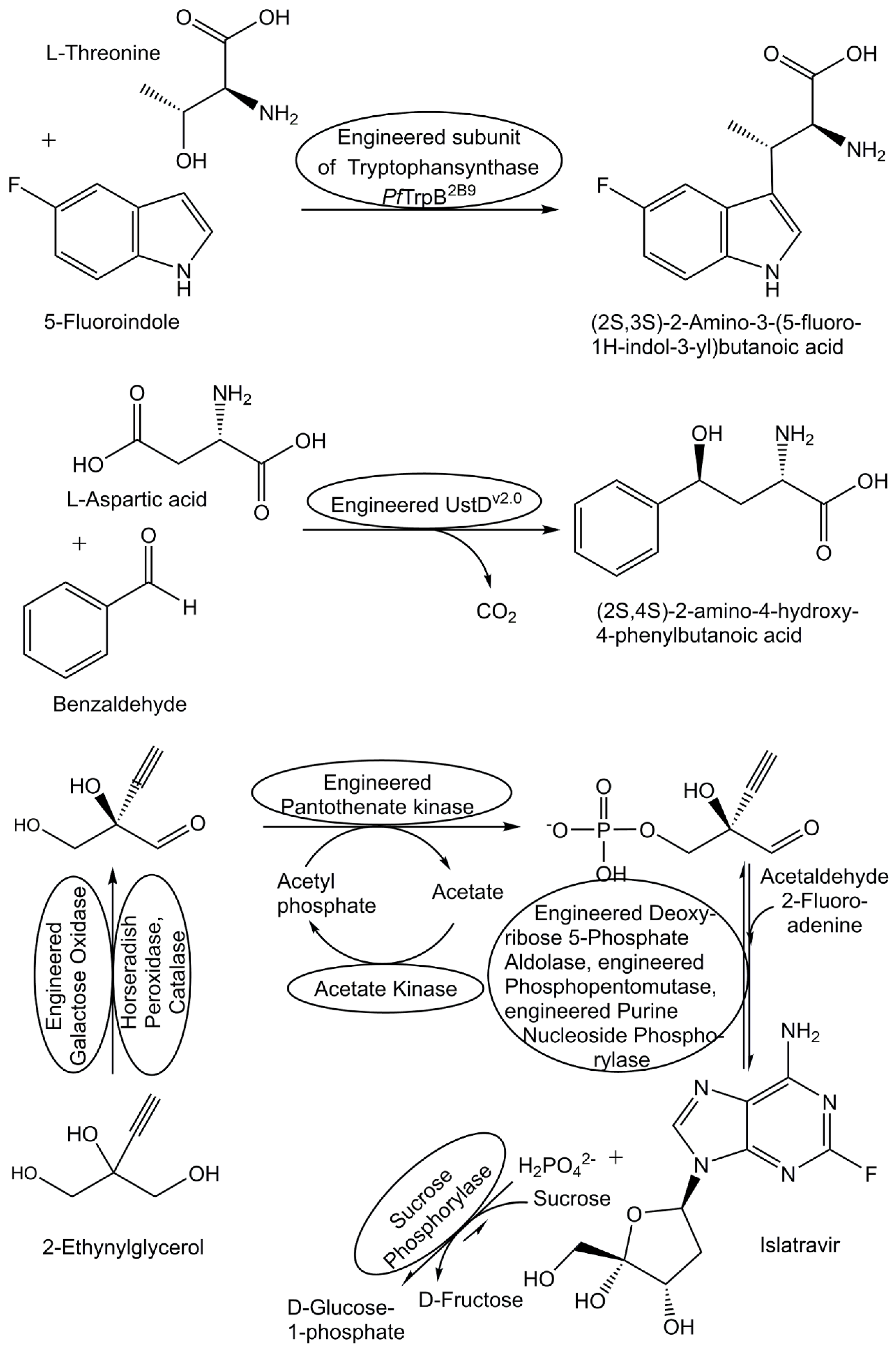
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



