Submitted:
18 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
Cabbage (Brassica oleracea L. var. capitata) plays a very important role in the annual national vegetable supply and export trade. In recent years, the cadmium content of agricultural soils has increased. As a result, cabbage yields and quality have declined significantly. Studies have shown that selenium can counteract the harmful effects of cadmium on plants and can alleviate the stress caused by cadmium during the growth of plants. However, the mechanism of exogenous selenium application in the alleviation of cadmium stress in cabbage seedlings has not been thoroughly investigated. In this study, exogenous selenium (10μMol/L) was applied under cadmium (25μMol/L) stress and the physiological mechanisms such as biomass, photosynthetic pigment, leaf stomata parameters, selenium and cadmium content, chloroplast ultrastructure, active oxygen accumulation, leaf membrane esterification and antioxidant enzyme activity were determined. The protective mechanism of exogenous selenium on cabbage seedlings under cadmium stress was investigated. The re-sults showed that exogenous application of selenium could effectively alleviate the decrease in growth, photosynthetic pigment and gas exchange characteristics of cabbage seedlings under cadmium stress, im-prove cabbage root vitality, reduce root leaf cadmium content and alleviate cadmium stress-induced damage. Ultrastructural observation showed that cadmium stress caused the disruption of the internal structure of chloroplasts of cabbage leaves, while exogenous selenium treatment alleviated the chloroplast damage to some extent, improved the stability of the inner capsule membrane and alleviated the cadmium stress-induced damage to photosynthetic organs. Cadmium stress also caused oxidative damage and ex-cessive accumulation of ROS in the leaves of cabbage seedlings as evidenced by significant accumulation of O2-, H2O2, MDA and electrolyte leakage. On the other hand, after exogenous selenium treatment, cadmium stress- induced oxidative damage could be reduced by up-regulating the activities of antioxidant enzymes SOD, POD and APX. At the same time, cadmium stress significantly increased GSH levels, and exogenous selenium treatment further increased GSH levels, thereby increasing the tolerance of cabbage to cadmium stress. In conclusion, by protecting the photosynthetic system, eliminating excessive accumulation of reactive oxygen species in cadmium-stressed cabbage seedlings, alleviating oxidative stress and reducing cadmium levels in the plant, exogenous selenium can further improve cadmium tolerance in cabbage seedlings.
Keywords:
selenium
; cadmium stress
; cabbage (Brassica oleracea L. var. capitata)
; photosynthesis
; physiological characteristics
1. Introduction
With the rapid development of industry, heavy metal pollution caused by mining, waste incineration, heavy use of pesticides and fertilizers has become increasingly serious [1]. Cadmium is a type of harmful and non-essential toxic heavy metal for plants, and once absorbed, it inhibits plant growth and development, resulting in slower root development, curled or chlorotic and etiolated leaves, and in severe cases, plant death [2,3]. Cadmium inhibits plant growth by affecting physiological processes such as photosynthesis, oxidative damage and mineral nutrition, resulting in growth retardation, leaf yellowing and biomass reduction. Some studies have shown that the chloroplast granule and thylakoid lamellar structure of wheat were disrupted under cadmium stress, and the efficiency of light capture and photosynthetic electron transport were significantly reduced [4,5]. Many studies have reported that active oxygen species and membrane peroxidation of rapeseed and Arabidopsis thaliana increased under cadmium stress [6,7]. At the same time, cadmium stress can inhibit the absorption, transport and distribution of many mineral elements in plants [8,9]. Studies in barley, tomato and other plants have shown that cadmium stress significantly inhibits the uptake of K, Mg, Fe, Ca, Zn and Mn, resulting in stagnation of plant growth [10,11]. In addition, cadmium not only affects plant growth and development, but also has a negative impact on human health when it accumulates in plants and enters the food chain. Therefore, agricultural and environmental researchers have focused on finding a method that can effectively mitigate cadmium stress on crops.
In recent years, the exogenous application of plant growth regulators has become an important way of reducing cadmium damage to plants and even reducing cadmium uptake. It has been reported that external foliar application of fulvic acid can reduce cadmium toxicity in lettuce [12] and ascorbic acid can reduce cadmium toxicity and absorption in maize [13]. During plant growth, tolerance to abiotic stress increased after selenium application [14,15]. Studies have shown that selenium can counteract and reverse the harmful effects of cadmium on plants, improve plant photosynthesis, regulate the balance of plant mineral elements, reduce cadmium levels in plants and increase plant antioxidant capacity to alleviate cadmium stress in plants. FILEK et al. found that cadmium stress caused chloroplast membrane degradation in rapeseed, whereas exogenous selenium application resulted in ultrastructural reconstruction of the chloroplast, recombination of thylakoid and stromal lamellar structure, increased fatty acid unsaturation of chloroplast size and fluidity of the plasma membrane [16]. Zhang et al. found that surface application of selenium increased photosystem II and electron transfer rate of Brassica napus leaves under cadmium stress, while root application of selenium increased photosynthetic rate, stomatal conductance, transpiration rate and stomatal threshold under cadmium stress [17]. The study on the effect of selenium on the absorption of mineral nutrients under cadmium stress showed that with the increase of cadmium concentration, the absorption of trace elements in plants was unbalanced, while the low concentration of selenium increased the content of Ca, Cu in the stem and Fe, Cu and Zn in the leaves, which effectively alleviated the toxic effect of cadmium and restored the normal growth of plants [18].Selenium can enhance the capacity of plant antioxidant defense system, reduce membrane lipid peroxidation and increase resistance to oxidative stress by scavenging intracellular free radicals, increasing antioxidant oxidase activity and non-enzymatic antioxidant content [19]. Research has shown that selenium can reduce the level of cadmium in the cell by increasing the retention of cadmium in the cell wall, and selenium can increase the synthesis of pectin, cellulose, hemicellulose and lignin in the cell wall of plant roots to trap more cadmium in the cell wall to reduce cadmium stress [20].
Cabbage (Brassica oleracea L. var. capitata), is an annual or biennial herb of the Brassica genus in the cruciferous family, originating from the Mediterranean region, with high adaptability, easy storage and transport resistance, and high yield and occupies a very important position in the national annual vegetable supply and export trade. As a cruciferous plant, cabbage usually has a strong ability to absorb and accumulate heavy metals [21], and the increased cadmium content in the soil and farmland led to a significant reduction in yield and quality. Therefore, it is a matter of concern to reduce the accumulation of cadmium in cabbage. This study aimed to investigate the protective mechanism of exogenous selenium on cadmium stress in cabbage seedlings, that is, exogenous selenium may improve the growth, photosynthesis and gas exchange ability of cabbage to protect the chloroplast ultrastructure of leaves, prevent oxidative damage by enhancing antioxidant activity, and mediate the absorption and transport of cadmium to enhance the tolerance of plants to cadmium. This study provides new insights into the potential physiological mechanism of exogenous selenium application in alleviating cadmium stress in cabbage seedlings, and certain indications for promoting yield and resistance to cadmium stress in cabbage seedlings.
2. Materials and Methods
2.1. Plant Materials and Handling
The cabbage variety ‘Fuer’ was provided by the College of Horticulture, Northwest A&F University. Seeds were soaked in 5% sodium hypochlorite for 5 min, rinsed twice with distilled water and then germinated for 24 hours in the dark at 25°C.
Seedlings were transplanted into a cavity dish with a soil: vermiculite ratio of 3:1 and grown in an incubator at 25/20°C(day/night), with a 14h/10h(day/night) photoperiod, 150μmol·m-2·s-1 light intensity and 70-80% relative humidity. Seedlings were transplanted into plastic POTS (25 cm × 20 cm × 10 cm; six plants per pot). To ensure normal growth of the cabbage, the cadmium and selenium concentration tests were carried out after 2 days of slow germination in Hogland’s nutrient solution, and the nutrient solution was renewed every 5 days.
The experiment was a completely randomized block design with four treatments: T1: Hoagland solution; T2: Hoagland solution +10μMol/L selenium; T3: Hoagland’s solution+25μMol/L cadmium; T4: Hoagland solution +25μMol/L cadmium+10μMol/L selenium, where the concentration of cadmium (25μMol/L) was chosen according to the previous study. After 14 days of cadmium stress, samples were collected and immediately cryopreserved in -80 °C liquid nitrogen for further analysis.
2.2. Measurement of Growth Parameters
Fresh and dry weight of cabbage leaves and roots, stem diameter, plant height and root length were measured after 10 days of cadmium treatment. Stem diameter, plant height and root length of cabbage were measured with calipers. Fresh weight was determined after washing the seedlings in distilled water, and dry weight was determined by drying the plants to constant weight at 65 °C.
2.3. Determination of Root Activity
Root activity determination was performed according to the method described by Luo et al. [24]. Briefly, 0.5 g of root sample and 5 mL of phosphate buffer (60 mM, pH 7.0) were added to the tube and reacted at 37°C for 2.5 h, then 2 mL of 1M sulfuric acid was added to the tube to stop the chemical reaction. Grind the roots in 3-4 mL of ethyl acetate with a mortar and pestle, transfer the red grinding liquid to the test tube, make up to 10 mL with ethyl acetate and record the absorbance at 485 nm.
2.4. Determination of Chlorophyll Content
The leaf sample (0.5 g) was placed in 10 mL of 95% ethanol at room temperature, extracted in the dark for 24 h and the supernatant centrifuged at 5000× g for 10 min. Absorbance values were measured with a spectrophotometer at 649 nm and 665 nm to calculate the contents according to the method of Wintermans and De-Mots [25].
2.5. Determination and Analysis of Leaf Stomata Parameters
Three cabbage seedlings were randomly selected for each treatment, and the stomatal exchange parameters of the fourth fully expanded leaf were determined from 9:00 to 10:00 using the Plant Photosynthesis Analyzer 6800 (LI-6800, LI-COR Corporation of the United States). The parameters were set as follows: photosynthetic photon density (PPFD) 1000 μmol m-2 s-1, leaf temperature (25°C), CO2 concentration (400 μmol CO2 mol-1).
2.6. Determination and Analysis of Leaf Chlorophyll Fluorescence Parameters
We randomly selected three cabbage seedlings from each treatment and measured the relevant parameters of the fourth fully unfolded leaf using a portable modulated chlorophyll fluorescence analyzer (PAM2500, WALZ, Germany), including maximum photochemical efficiency (Fv/Fm), actual photochemical efficiency Y(II), non-photochemical quenching (NPQ) and the quantum yield for the regulation of energy dissipation in photochemical quenching (qP).
2.7. Determination of Selenium and Cadmium Contents
The root and leaf tissues were dried at 65°C until the sample reached constant weight, then the dried plant samples (0.5 g) were ground to powder, mixed with nitric acid/perchloric acid (4:1, v/v) and boiled at 220°C until clear and used for constant volume determination. The cadmium content was determined by flame atomic absorption spectrophotometer and the selenium content by liquid phase atomic fluorescence photometer.
2.8. Chloroplast Ultrastructure Analysis
The fourth fully expanded leaf (approximately 1 mm2) of cabbage from each treatment was fixed in 2.5% (v/v) glutaraldehyde (0.1M phosphate buffer, pH 7.2) overnight, soaked in 1% (v/v) osmic acid and washed with 0. 1M PBS (pH 7.4) after 1-2 h. Then the sample was dehydrated in 30%-50%-70%-80%-95%-100% alcohol sequentially, dehydrated in 100% ethanol for 15 min 3 times and immersed in Epon-812 epoxy resin at 60°C overnight. Finally, the samples were semi-sectioned, stained, rinsed with dye solution, fixed in copper mesh, scanned by transmission electron microscopy, and photographed for observation [26].
2.9. Determination and Analysis of Relative Water Content
The fourth fully unfolded leaf of the seedling under each treatment was selected to determine the electrical conductivity and the specific measurement method was taken from Bajji. [27].
2.10. Pro Content Determination and Analysis
0.5 g of cabbage leaves were weighed and pulverized with liquid nitrogen, 7.5 mL of 3% sulfosalicylic acid was added to regrind completely and then centrifuged at 4°C for 20 min (11000×g). 2 mL of the supernatant was removed and mixed by adding 2 mL of ninhydrin and 2 mL of 3.5% acetic acid, and the mixed solution was boiled at 100°C for 1 h. After cooling, 4 mL of toluene was added to the reaction mixture, and the absorbance value was measured at 520 nm [28].
2.11. MDA Content Determination and Analysis
Weigh 0.5 g of cabbage leaves, add 5 mL of 0.1 % trichloroacetic acid (TCA), crush completely in a pre-cooled mortar and centrifuge at 4000 × g for 15 min at 4 °C. The supernatant is the liquid to be measured. The supernatant was the liquid to be determined. 1 mL of the supernatant was absorbed and 4 mL of a 20 % TCA mixture containing 0.65 % (w/v) TBA was added. The mixture was reacted in boiling water for 15 min, immediately cooled and centrifuged again to determine light absorption at 450, 532 and 660 nm.
2.12. Analysis of O2− and H2O2 Contents
Tissue from 0.5 g of fresh leaves was taken and 5 mL of 50 mM phosphate buffer (pH 7.8) was added. Take 0.5 g of fresh leaf tissue, ground with 5mL of 50 mM phosphate buffer (pH 7.8), and the mixture centrifuged at 10,000×g for 20 min. The O2- content was measured in the supernatant and calculated from the standard curve based on sodium nitrite, expressed as μmol·g−1·min−1 FW. To determine H2O2 content, 0.5 g of fresh leaf tissue was ground in an ice bath with 5 mL 100% cold water acetone, centrifuged at 10,000×g 4°C for 20 min and the supernatant was collected immediately for H2O2 analysis. The concentration of H2O2 was measured according to the standard curve and expressed as μmol·g−1 FW.
2.13. H2O2 and O2- Histochemical Staining
The nitrogen blue tetrazole (NBT) method was used for O2- staining analysis of cabbage leaves: The leaves were soaked in 0.5 mg·mL-1 NBT solution (pH 7.8) in the dark for 1 h, then removed, rinsed 3 times with distilled water and then heated with 95% ethanol for 10 min for decolorizations. The blue spots on the leaves indicated the production of O2-. The 3, 3-diaminobenzidine (DAB) staining method was used to stain the tissue of cabbage leaves for H2O2: The leaves were soaked in 1 mg·mL-1 DAB solution (pH 7.8) at 25 °C for 5 h, then decolored with 95% (v/v) ethanol every 20 min at 80 °C until the green background was completely removed from the leaves. Red-brown spots on the leaves indicate H2O2 production.
2.14. Determination and Analysis of GSH
The level of GSH was determined using the Solarbio BC1175 kit for the content of reduced glutathione.
2.15. APX Determination and Analysis
The determination was carried out by means of the Solarbio BCO220 Ascorbate Peroxidase Kit.
2.16. Antioxidant Enzyme System Determination and Analysis
The 0.5 g leaf sample was weighed and ground to a homogenate with 3 mL of phosphate buffer consisting of 0.2 mM EDTA and 2% (w/v) PVP. The homogenate was centrifuged at 4°C for 20 min at 13,000×g, and the supernatant was the enzyme liquid to be measured, which was used for the determination of SOD and POD enzyme activities. APX assay: 3 mL of the reaction mixture contained 50 mM PBS (pH 7.0), 9 mM ASA, 12.5 mM H2O2, and 100μL of enzyme solution, and the enzyme activity was calculated based on the change in absorbance at 290 nm every 1 min.
2.17. Statistical Analysis
All data were analyzed using one-way ANOVA followed by Duncan multiple range tests. P value < 0.05 was considered significant. Values are presented as mean ± standard deviation (SD) of at least three independent experiments with three replicates for each treatment (P ≤ 0.05).
3. Results
3.1. Exogenous Selenium Can Promote the Growth of Cabbage Seedlings under Cadmium Stress
To study the effect of exogenous selenium on the growth and development of cabbage seedlings under cadmium stress, we measured plant height, stem diameter, root length, root vitality, total chlorophyll content, and dry and fresh weight of leaves and roots of cabbage seedlings. Compared with the control, cadmium stress caused significant leaf chlorosis and curling, and inhibited plant and root growth. Exogenous spraying of selenium improved the growth of cabbage seedlings and alleviated the symptoms of leaf yellowing (Figure 1). Under normal conditions, exogenous selenium increased the growth of cabbage seedlings. Compared with the control, FW and DW of leaves and roots, plant height, stem diameter and root length of cabbage seedlings were significantly decreased under cadmium stress. Compared with cadmium treatment alone, the application of exogenous selenium under cadmium stress could significantly increase the FW and DW of leaves and roots, plant height, stem diameter and root length of cabbage seedlings by 30.14%,34.36%, 21.21%,60%, 18.30%, 16.67% and 28.20%, respectively (Figure 2A–G). In addition, compared with the control group, root activity was significantly decreased under cadmium stress, while compared with cadmium stress alone, root activity of cabbage seedlings was significantly increased by 82.92% when exogenous selenium was applied under cadmium stress. The results showed that exogenous selenium could promote the growth of cabbage seedlings under cadmium stress by increasing root water uptake and nutrient use efficiency, and enhancing root activity (Figure 2I). Compared with the control group, the total chlorophyll content of cabbage seedlings decreased significantly under cadmium stress alone, but increased when treated with exogenous selenium alone. Compared with the treatment of cadmium stress alone, the total chlorophyll content of cabbage seedlings increased by 55.56% when exogenous selenium was applied under cadmium stress, indicating that exogenous selenium could alleviate the decrease of photosynthetic pigment content in cabbage seedlings under cadmium stress (Figure 2H).
3.2. Effects of Exogenous Selenium on Stomatal Parameters of Cabbage Seedlings under Cadmium Stress
To study the effect of exogenous selenium on the stomatal parameters of cabbage seedlings under cadmium stress, we evaluated the stomatal parameters of cabbage seedlings. Compared to the control, cadmium treatment significantly reduced the stomatal exchange capacity of cabbage leaves. When compared with cadmium treatment alone, exogenous selenium increased the content of Pn (65.4%), Gs (32.5%) and Tr (72.84%), respectively, in cabbage leaves under cadmium stress, while decreasing the content of Ci (14.59%). When compared with the control, no significant difference was found in Pn, Gs, Tr and Ci (Figure 3A, Figure 3D, Figure 3C, Figure 3B), but the effect was alleviated by the application of exogenous selenium. When treated with selenium alone, the contents of Pn, Gs and Tr in the leaves of cabbage seedlings were not significantly different from the control group, while Ci was significantly increased.
3.3. Effects of Exogenous Selenium on Chlorophyll Fluorescence Parameters of Cabbage Seedlings under Cadmium Stress
Cadmium stress alone inhibited chlorophyll synthesis, decreased stomatal exchange parameters and reduced PSII activity in leaves of cabbage seedlings. Compared with the control group, maximum photochemical efficiency (Fv/Fm), actual photosynthetic capacity Y(II) and photochemical quenching (qP) of leaves of cabbage seedlings were reduced by 78.5%, 68.51% and 59.55%, respectively. However, exogenous selenium applied under cadmium stress can significantly reduce the cadmium-induced decreases in Fv/Fm, Y(II) and qP. The above results indicated that the application of exogenous selenium to cabbage seedlings could significantly alleviate cadmium stress and increase the maximum photochemical efficiency of leaves under cadmium stress. On the contrary, the NPQ value of leaves of cabbage seedlings treated with cadmium alone was the highest, but this phenomenon was alleviated by spraying exogenous selenium. In addition, there was no significant difference between the leaves of cabbage seedlings and the control treatment when exogenous selenium was sprayed alone (Figure 4A–D).
3.4. Exogenous Selenium Can Enhance the Tolerance of Cabbage Seedlings under Cadmium Stress
To further elucidate the mechanism of cadmium tolerance induced by exogenous selenium in leaves of cabbage seedlings, we determined the effects of exogenous selenium on cadmium and selenium contents in leaves and roots of cabbage seedlings. The results showed that the cadmium content in leaves and roots was highest under cadmium treatment alone. Exogenous application of selenium significantly reduced the cadmium content in leaves and roots of cabbage seedlings, and compared with cadmium treatment alone, the cadmium content in leaves and root tissues of cabbage seedlings was significantly reduced to 73% and 76%, respectively (Figure 5A). Selenium content in leaves and roots of cabbage seedlings was increased by selenium application alone, indicating that cabbage seedlings had a certain ability of selenium accumulation, while compared with selenium and cadmium combined treatment, selenium content in roots and leaves was not significantly increased (Figure 5B), indicating that exogenous selenium is antagonistic to cadmium. The cadmium content in leaves was significantly lower than that in roots after exogenous selenium treatment, indicating that exogenous selenium could inhibit the transfer of cadmium from the underground to the aboveground part by regulating the cadmium content in different parts of cabbage seedlings, thus improving the tolerance of cabbage seedlings to cadmium stress.
3.5. Observation of Chloroplast Ultrastructure in Leaves of Cabbage Seedlings
The internal structure of chloroplasts in leaves of cabbage seedlings was further observed to determine whether exogenous application of selenium can reduce chloroplast damage caused by cadmium stress. The results showed that the control group had good morphology, complete cell structure, uniform thickness of cell wall (CW), tight adhesion between cell membrane (CM) and cell wall, and no obvious plasma wall separation. Chloroplasts (Ch) were abundant, mostly in short fusiform form, a small amount of starch granules (SG) and granular lamellae (GL) were found in the interior, and a small amount of osmophilic granules (OG) were found in the leaf green (Figure 6A–C). Seedlings treated with selenium alone were similar to the control group in that the cell structure was intact, the cell wall (CW) was intact, the membrane (CM) was closely attached to the cell wall and no obvious plasma wall separation was observed. The number of chloroplasts (Ch) was normal and most were short fused, with a small amount of starch granules (SG), a regular arrangement of grana lamellae (GL) and a fuzzy structure. The mitochondria (M) are elliptical and the vacuole (V) membrane is intact (Figure 6D–F). Cadmium alone caused partial disintegration of the cell structure, the cell wall (CW) remained intact, the cell membrane (CM) was damaged in a small area, the cell matrix dissolved and the plasmalemma wall separated. The number of chloroplasts (Ch) decreased significantly, some morphological structures were damaged or disappeared, a small number of chloroplasts were spherical, the grana lamella (GL) was reduced, the structure was unclear, and a small number of chloroplasts were expanded, and a small number of osmophilic particles (OG) without uniform size were found in the chloroplasts. In addition, the small number of osmophilic particles in the leaf green were different in size compared to the control group, and no obvious starch particles were found (Figure 6G–I), indicating that the photosynthetic organs in the leaves of cabbage seedlings were damaged under cadmium stress. After exogenous application of selenium, chloroplast morphology was obviously restored, cell structure was mostly present, cell wall (CW) was intact, cell membrane (CM) was damaged and disappeared, and plasmalemma wall was separated. The number of chloroplasts (Ch) was normal, free and most of them were in good condition. The arrangement of grana lamellae (GL) was regular and some of them were moderately dilated. There were more starch granules (SG) in the chloroplasts with larger volume. A small number of free osmophilic particles (OG) could be seen in the cells (Figure 6J–L), indicating that exogenous selenium application alleviated the damage caused by cadmium stress on chloroplasts to a certain extent, improved the stability of the inner capsule membrane, and alleviated the damage caused by cadmium stress on photosynthetic organs.
3.6. Effects of Exogenous Selenium on Reactive Oxygen Species Accumulation and Leaf Membrane Esterification of Cabbage Seedlings under Cadmium Stress
To further elucidate the degree of damage to the plasma membrane of cabbage seedlings under cadmium stress, the membrane integrity and the degree of membrane lipid peroxidation of leaves were observed by diaminobenzidine method and nitro tetrazolium chloride reagent staining, respectively. When cadmium was treated alone, the leaves of cabbage seedlings were stained by diaminobenzidine method and nitro tetrazolium chloride blue reagent, respectively. The leaves of cabbage seedlings were stained blue and yellow, respectively, and the degree of staining was worse than that of the control, indicating that cadmium treatment alone could damage the integrity of the cell membrane of cabbage seedlings and aggravate membrane lipid peroxidation of leaves. There was no significant difference between selenium treatment alone and the control. The degree of blue and yellow staining of leaves of cabbage seedlings was reduced after exogenous application of selenium under cadmium stress, indicating that exogenous application of selenium can alleviate the oxidative damage of leaves of cabbage seedlings under cadmium stress (Figure 7A,B).
The degree of oxidative damage in leaves under cadmium stress is related to the excessive accumulation of ROS. The levels of O2- and H2O2 in leaves of cabbage seedlings were measured and it was found that the levels of O2- and H2O2 in leaves of cadmium-stressed cabbage seedlings accumulated most when treated with cadmium alone. There was no significant difference between the Se treatment alone and the control. However, compared with cadmium stress alone, exogenous application of selenium treatment under cadmium stress reduced the content of O2- and H2O2 in leaves of cabbage seedlings to 13.63% and 40.83%, respectively (Figure 8A,B). In addition, cadmium alone had the greatest effect on the electrical conductivity, proline and malondialdehyde contents of cabbage leaves. Compared with cadmium treatment alone, exogenous application of selenium under cadmium stress significantly reduced the electrical conductivity and malondialdehyde content of cabbage seedlings to 50.63% and 33.87%, respectively, and further increased the proline content of cabbage seedlings by 80.3%. There was no significant difference between selenium addition alone and the control group (Figure 8C–E).
3.7. Effects of Exogenous Selenium on Reactive Oxygen Species Accumulation and Leaf Membrane Esterification of Cabbage Seedlings under Cadmium Stress
The activities of superoxide dismutase (SOD), peroxidase (POD), ascorbate peroxidase (APX) and the content of reduced glutathione (GSH) in leaves of cabbage seedlings were determined to elucidate the role of exogenous selenium in the antioxidant system under cadmium stress. The activities of SOD, POD and GSH and APX in leaves of cabbage seedlings treated with selenium alone were similar to the control and the activity of APX was increased. The activities of SOD, POD, APX and GSH were significantly higher than the control when treated with cadmium alone. However, when treated with the combination of cadmium and selenium, the activities of SOD, POD, APX and GSH were further increased by 23.36%, 52.28%, 36.54% and 8.33%, respectively (Figure 9A–D).Exogenous selenium could alleviate the cadmium-induced damage, indicating that selenium could alleviate the cadmium-induced oxidative stress by regulating the antioxidant activity of leaves.
4. Discussion
Heavy metal contamination of soil is a global problem and poses a serious threat to public health [27,28,29]. Cadmium is highly toxic to plants [30], and the accumulation of cadmium in edible parts of crops is one of the main threats to human health. Reducing the damage of abiotic stress to plants by applying exogenous substances has gradually become the focus of research [31]. As one of the essential elements for animals and humans, selenium has been proven to reduce the toxicity of cadmium, lead Pb and arsenic As to plants [32,33,34]. Therefore, the application of exogenous selenium to alleviate cadmium stress in cabbage seedlings is of great significance for agricultural production.
In this study, we analyzed the physiological and biochemical parameters of cabbage seedlings regulated by exogenous selenium application under cadmium stress. Under cadmium stress, cabbage plants showed obvious symptoms such as leaf rolling, yellowing, inhibition of root elongation and reduction of biomass (Figure 1), which was consistent with studies in Brassica napus, mustard, tomato and cucumber [35,36,37,38]. On the contrary, exogenous application of selenium could alleviate these phenomena, and exogenous application of selenium promoted the growth of cabbage seedlings under cadmium stress, which was reflected in the increase of plant height, stem diameter and biomass (Figure 2A-G). This was consistent with the research results of Filek M and Qi W. [39,40], which indicated that selenium could reduce the growth inhibition of cabbage seedlings under cadmium stress(Figure 2I). The root activity may reflect the growth status of plants, and the activities of roots treated with cadmium alone were significantly reduced, while they were apparently restored after exogenous application of Se, which was consistent with the results of Jiang J ‘s study [41]. These results indicated that selenium could enhance root vitality by improving the efficiency of root water and nutrient uptake to promote plant growth under cadmium stress.
Our research showed that the cadmium content in leaves and roots of cabbage seedlings was significantly increased under cadmium stress, and was higher in roots than in leaves (Figure 5A), indicating that cadmium is trapped in the roots and a small amount is subsequently transported to the leaves. The results are consistent with those of previous studies that cadmium accumulates more in roots than in leaves. However, the cadmium content in leaves and roots was reduced after exogenous selenium application, which is similar to the results of the study by Li L et al. in mustard. In conclusion, exogenous selenium application has a positive effect on reducing cadmium content in cabbage seedlings, and the difference in the positive effect of selenium on different plant parts may be related to the accumulation of cadmium in the upper part of the plant.
Cadmium stress can lead to a decrease in photosynthetic pigment content in plants, which has been reported in cabbage, broccoli, pepper and other plants [41,42,43]. Our results showed that cadmium treatment alone resulted in a significant decrease in total chlorophyll content, and at the same time, cadmium stress also significantly decreased gas exchange characteristics such as Pn, Gs and Tr and increased Ci (Figure 3A, Figure 3D, Figure 3C, Figure 3B). We speculated that cadmium stress led to stomatal closure, reduced mesophyll conductance and CO2 fixation, and ultimately photoinhibition. selenium can promote growth and development of plants under various abiotic stresses. In this study, exogenous selenium application effectively alleviated the cadmium induced decrease in chlorophyll content and photosynthetic characteristics, which was consistent with previous studies [44].
Chlorophyll fluorescence can reflect the photosynthetic efficiency and photosynthetic inhibition of plants under heavy metal stress. In this study [45], cadmium treatment significantly reduced Fv/Fm, Y(II) and qP parameters, but increased NPQ value (Figure 4A-D), indicating that PSII under cadmium stress had an imbalance between light energy absorption and photosynthetic electron transfer [46,47]. However, exogenous selenium can significantly increase chlorophyll content and photosynthetic capacity and reduce cadmium stress-induced photoinhibition by increasing Fv/Fm, Y(II) and qP [48,49].
Chloroplasts, as the sites of photosynthesis in plants, are involved in the regulation of various physiological responses and are also organelles sensitive to cadmium stress. In this study, the ultrastructure of leaf mesophyll cells of cabbage seedlings was significantly damaged under cadmium stress, as shown by chloroplast deformation and damage, poor lamellar arrangement, reduction of starch granules and appearance of osmophilic particles, which was consistent with previous studies [50,51]. The results showed that cadmium stress significantly decreased the photosynthetic rate, chlorophyll content and starch granule accumulation in cabbage chloroplasts, which was consistent with the results of H. Y. Sun et al. Therefore, we speculate that the structural changes in chloroplasts under cadmium stress may be the direct cause of damage to the photosynthetic system and chlorophyll degradation, which ultimately leads to the inhibition of photosynthesis.
However, exogenous application of selenium restored the damage to the chloroplast structure, increased the number of starch granules in cabbage leaves, and effectively improved the stability and integrity of the chloroplast structure (Figure 6), indicating that selenium could protect the growth of cabbage seedlings and alleviate the damage to the chloroplast structure under cadmium stress, thus promoting photosynthesis, which played a key role in alleviating cadmium stress in cabbage seedlings.
To investigate the effect of exogenous selenium application on the scavenging of ROS in seedling leaves, we examined the levels of O2- and H2O2 in leaves of cabbage seedlings. The results showed that cadmium caused a significant increase in the production of O2- and H2O2, resulting in excessive ROS production (Figure 7, Figure 8), which was consistent with the results of He S et al. [52]. However, exogenous application of selenium minimized the production of ROS induced by cadmium stress, possibly due to its function of protecting cells from cadmium-induced oxidative damage.
At the same time, the levels of MDA, proline and electrolyte leakage are used as important indicators of lipid peroxidation in plants to reflect the degree of membrane damage [53]. Our results showed that cadmium treatment significantly increased the generation of MDA, proline and electrolyte leakage in leaf tissues compared to the control (Figure 8D). According to the histochemical detection, the staining degree of leaves of cabbage seedlings treated with cadmium alone was higher, indicating that cadmium stress caused severe cellular REDOX imbalance and oxidative damage, increased lipid peroxidation and excessive ROS accumulation. It can be concluded that the oxidative damage caused by cadmium in plants is the reason for the destruction of cell membrane, inhibition of nutrient absorption and enhanced lipid peroxidation [54].
Plants induce resistance to biotic and abiotic stresses by producing antioxidant enzymes, and oxidative damage is closely related to antioxidant defense mechanisms. The results showed that the activities of SOD, POD, APX and GSH of cabbage seedlings were increased under cadmium stress, while the exogenous application of selenium further improved the activity of cabbage seedling leaves (Figure 9), indicating that selenium could significantly reduce the oxidative damage caused by cadmium stress by enhancing the antioxidant response. Our results were consistent with those of Qi W et al. who confirmed that Cadmium-stressed Brassica napus plants could enhance the activity of antioxidant enzymes, and the enzyme activity was further improved after selenium application, and antioxidant enzymes were closely related to cadmium tolerance [40]. These results suggest that exogenous selenium may induce REDOX homeostasis by activating both enzyme-supported and non-enzyme-supported antioxidant systems, thereby protecting cabbage seedlings from cadmium stress.
5. Conclusions
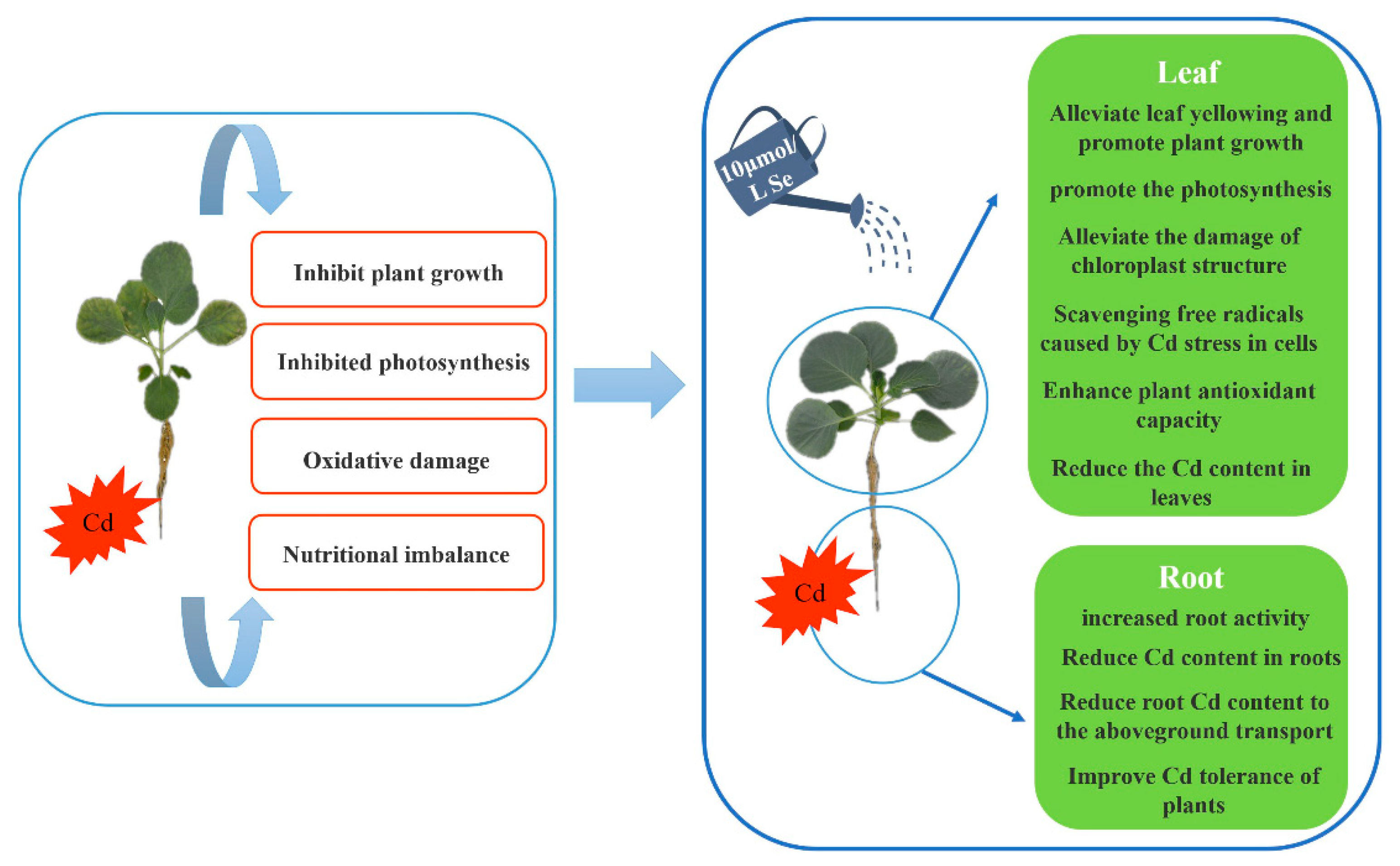
In this study, we found that exogenous selenium can effectively alleviate the stress of cadmium on the growth and physiological and biochemical characteristics of cabbage seedlings, inhibit the transport of cadmium from root to leaf, and restore the negative effect of cadmium stress. Exogenous selenium application is an important way to alleviate cadmium stress.
Cadmium stress severely inhibited the growth and photosynthesis of cabbage seedlings, increased the levels of O2-, H2O2, MDA, electrolyte leakage and proline, decreased the antioxidant capacity of SOD, POD, APX and GSH, and disturbed the nutrient balance. However, exogenous application of selenium increased the biomass and photosynthetic efficiency of cabbage seedlings and decreased the concentration of cadmium, which was conducive to maintaining the growth of cabbage seedlings and eliminating excess ROS by upregulating antioxidant enzymes to protect the ultrastructure of chloroplasts. Therefore, the results confirmed the ameliorative effect of exogenous selenium application on cadmium stress in cabbage seedlings, which laid the foundation for the study of exogenous selenium application to reduce cadmium stress in cabbage (Figure 10).
In conclusion, this study elucidates the role and regulatory mechanism of selenium under cadmium stress in cabbage, which is helpful for the application of selenium under cadmium stress in agricultural production to promote plant growth and increase yield.
Author Contributions
K.J. was the experimental designer and performer of the experimental study, and completed the data analysis and first draft of the paper; Z.Z., B.W., W.W. and W.W. participated in the experimental design and analysis of the experimental results; Z.X., W.H. and D.L. were the conceptualizers and leaders of the project, and led the experimental design, data analysis, paper writing and revision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2016YFD0101702), the Special Funds Project for the Construction of National Modern Agricultural Industrial Technology System (CARS-23-G22) and the Science and Technology Program of Xi’an Municipality (20NYTX0001).
Data Availability Statement
Data are contained within the article.
Acknowledgments
We thank Ruihong Chen and Lijuan Lu (Horticulture Science Research Center, Northwest A&F University, Yangling, China) for their assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khalid, U.; et al. Comparative effects of conventional and nano-enabled fertilizers on morphological and physiological attributes of Caesalpinia bonducella plants. Journal of the Saudi Society of Agricultural Sciences. 2022, 21, 61–72. [Google Scholar] [CrossRef]
- Rizwan, M.; et al. A critical review on the effects of zinc at toxic levels of cadmium in plants. Environmental Science and Pollution Research. 2019, 26, 6279–6289. [Google Scholar] [CrossRef] [PubMed]
- Baruah, N.; et al. Influence of Heavy Metals on Seed Germination and Seedling Growth of Wheat, Pea, and Tomato. Water, Air, & Soil Pollution. 2019, 230, 273. [Google Scholar]
- Alves, L.R.; et al. Selenium improves photosynthesis and induces ultrastructural changes but does not alleviate cadmium-stress damages in tomato plants. Protoplasma. 2020, 257, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.Y.; Wei, X.J.; Wang, H.Z. Cadmium tolerant and sensitive wheat lines: their differences in pollutant accumulation, cell damage, and autophagy. Biologia Plantarum. 2018, 62, 379–387. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; et al. Cadmium Stress Leads to Rapid Increase in RNA Oxidative Modifications in Soybean Seedlings. Front Plant Sci. 2017, 8, 2219. [Google Scholar] [CrossRef]
- Lv, Y.; et al. Metallothioneins BcMT1 and BcMT2 from Brassica campestris enhance tolerance to cadmium and copper and decrease production of reactive oxygen species in Arabidopsis thaliana. Plant and Soil. 2013, 367, 507–519. [Google Scholar] [CrossRef]
- Tran, T.A.; Popova, L.P. Functions and toxicity of cadmium in plants: recent advances and future prospects. Turkish Journal of Botany. 2013, 37, 1–13. [Google Scholar] [CrossRef]
- Kumar, S., S. Kumar and T. Mohapatra, Interaction Between Macro- and Micro-Nutrients in Plants. Front Plant Sci. 2021, 12, 665583. [Google Scholar] [CrossRef]
- GUO, T.; et al. Influence of Aluminum and Cadmium Stresses on Mineral Nutrition and Root Exudates in Two Barley Cultivars* *Project supported by the Chinese Ministry of Science and Technology (China-Australian Special Link Research Program) and the Grains Research and Development Corporation of Australia (No. UT-8). Pedosphere. 2007, 17, 505–512. [Google Scholar]
- Borges, K.L.R.; et al. Nutritional status and root morphology of tomato under Cd-induced stress: Comparing contrasting genotypes for metal-tolerance. Scientia Horticulturae. 2019, 246, 518–527. [Google Scholar] [CrossRef]
- Wang, Y.; et al. Exogenous foliar application of fulvic acid alleviate cadmium toxicity in lettuce (Lactuca sativa L.). Ecotoxicol Environ Saf. 2019, 167, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; et al. Exogenous application of ascorbic acid mitigates cadmium toxicity and uptake in Maize (Zea mays L.). Environ Sci Pollut Res Int. 2019, 26, 19261–19271. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Selenium Pretreatment Upregulates the Antioxidant Defense and Methylglyoxal Detoxification System and Confers Enhanced Tolerance to Drought Stress in Rapeseed Seedlings. Biological Trace Element Research. 2011, 143, 1758–1776. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Chapter 16 - Silicon and Selenium: Two Vital Trace Elements that Confer Abiotic Stress Tolerance to Plants, In Emerging Technologies and Management of Crop Stress Tolerance. P. Ahmad and S. Rasool, P. Ahmad and S. Rasool^Editors. 2014, Academic Press: San Diego. 377-422.
- Battin, E.E.; Brumaghim, J.L. Antioxidant activity of sulfur and selenium: a review of reactive oxygen species scavenging, glutathione peroxidase, and metal-binding antioxidant mechanisms. Cell Biochem Biophys. 2009, 55, 1–23. [Google Scholar] [CrossRef]
- Zhang, Z.W.; et al. Selenium Enhances Cadmium Accumulation Capability in Two Mustard Family Species-Brassica napus and B. juncea. Plants. 2020, 9, 7. [Google Scholar] [CrossRef]
- Qin, X.; et al. Influence of selenium on root morphology and photosynthetic characteristics of winter wheat under cadmium stress. Environmental and Experimental Botany. 2018, 150, 232–239. [Google Scholar] [CrossRef]
- Silva, V.M.; et al. Physiological, biochemical, and ultrastructural characterization of selenium toxicity in cowpea plants. Environmental and Experimental Botany. 2018, 150, 172–182. [Google Scholar] [CrossRef]
- Yu, H.; et al. Characteristics of cadmium immobilization in the cell wall of root in a cadmium-safe rice line (Oryza sativa L.). Chemosphere. 2020, 241, 125095. [Google Scholar] [CrossRef]
- Rizwan, M.; et al. Cadmium phytoremediation potential of Brassica crop species: A review. Science of The Total Environment. 2018, 631–632, 1175–1191. [Google Scholar] [CrossRef]
- Luo, Z.; et al. Heavy metal accumulation and signal transduction in herbaceous and woody plants: Paving the way for enhancing phytoremediation efficiency. Biotechnology Advances. 2016, 34, 1131–1148. [Google Scholar] [CrossRef] [PubMed]
- Wintermans, J.F.; de Mots, A. Spectrophotometric characteristics of chlorophylls a and b and their pheophytins in ethanol. Biochim Biophys Acta. 1965, 109, 448–53. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; et al. Growth, physiological, and biochemical responses of Camptotheca acuminata seedlings to different light environments. Front Plant Sci. 2015, 6, 321. [Google Scholar] [CrossRef] [PubMed]
- Bajji, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regulation. 2004, 36, 61–70. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant and Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Järup, L.; Akesson, A. Current status of cadmium as an environmental health problem. Toxicol Appl Pharmacol. 2009, 238, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Vesey, D.A.; Gobe, G.C. Current health risk assessment practice for dietary cadmium: Data from different countries. Food Chem Toxicol. 2017, 106, 430–445. [Google Scholar] [CrossRef]
- Zhao, F.J.; et al. Soil contamination in China: current status and mitigation strategies. Environ Sci Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef]
- Satarug, S.; et al. A global perspective on cadmium pollution and toxicity in non-occupationally exposed population. Toxicology Letters. 2003, 137, 65–83. [Google Scholar] [CrossRef]
- Zaki, H.E.M.; Radwan, K.S.A. The use of osmoregulators and antioxidants to mitigate the adverse impacts of salinity stress in diploid and tetraploid potato genotypes (Solanum spp.). Chemical and Biological Technologies in Agriculture. 2022, 9, 19. [Google Scholar] [CrossRef]
- CAMARA, A.Y.; et al. Effect of selenium on uptake and translocation of arsenic in rice seedlings (Oryza sativa L.). Ecotoxicology and Environmental Safety. 2018, 148, 869–875. [Google Scholar] [CrossRef]
- Huang, Q.; et al. Selenium application alters soil cadmium bioavailability and reduces its accumulation in rice grown in Cd-contaminated soil. Environ Sci Pollut Res Int. 2018, 25, 31175–31182. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; et al. Indications of Selenium Protection against Cadmium and Lead Toxicity in Oilseed Rape (Brassica napus L.). Frontiers in Plant Science. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Baszyński, T. , Interference of Cd2+ in functioning of the photosynthetic apparatus of higher plants. Acta Societatis Botanicorum Poloniae. 1986, 55, 291–304. [Google Scholar] [CrossRef]
- Zhang, F.; et al. Cadmium-induced accumulation of hydrogen peroxide in the leaf apoplast of Phaseolus aureus and Vicia sativa and the roles of different antioxidant enzymes. J Hazard Mater. 2009, 168, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Komarkova, M.; et al. Physiological and biochemical responses of Brassica napus L. cultivars exposed to Cd stress. Plant, Soil and Environment. 2022, 68, 431–440. [Google Scholar] [CrossRef]
- Ahmad, P.; et al. Exogenous Application of Selenium Mitigates Cadmium Toxicity in Brassica juncea L. (Czern & Cross) by Up-Regulating Antioxidative System and Secondary Metabolites. Journal of Plant Growth Regulation. 2016, 35, 936–950. [Google Scholar]
- Filek, M.; et al. The protective role of selenium in rape seedlings subjected to cadmium stress. Journal of Plant Physiology. 2008, 165, 833–844. [Google Scholar] [CrossRef]
- Qi, W.; et al. Selenium nanoparticles ameliorate Brassica napus L. cadmium toxicity by inhibiting the respiratory burst and scavenging reactive oxygen species. Journal of Hazardous Materials. 2021, 417, 125900. [Google Scholar] [CrossRef]
- Jiang, J.; et al. Exogenous tryptophan application improves cadmium tolerance and inhibits cadmium upward transport in broccoli (Brassica oleracea var. italica). Front Plant Sci. 2022, 13, 969675. [Google Scholar] [CrossRef]
- Yang, L.; et al. Physiological Mechanism of Exogenous 5-Aminolevulinic Acid Improved the Tolerance of Chinese Cabbage (Brassica pekinensis L.) to Cadmium Stress. Front Plant Sci. 2022, 13, 845396. [Google Scholar] [CrossRef] [PubMed]
- Alshegaihi, R.M.; et al. Effective citric acid and EDTA treatments in cadmium stress tolerance in pepper (Capsicum annuum L.) seedlings by regulating specific gene expression. South African Journal of Botany. 2023, 159, 367–380. [Google Scholar] [CrossRef]
- Zhao, Y.; et al. Selenium reduces cadmium accumulation in seed by increasing cadmium retention in root of oilseed rape (Brassica napus L.). Environmental and Experimental Botany. 2019, 158, 161–170. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. A chlorophyll fluorescence analysis of photosynthetic efficiency, quantum yield and photon energy dissipation in PSII antennae of Lactuca sativa L. leaves exposed to cinnamic acid. Plant Physiology and Biochemistry. 2011, 49, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; et al. Acetylcholine ameliorates the adverse effects of cadmium stress through mediating growth, photosynthetic activity and subcellular distribution of cadmium in tobacco (Nicotiana benthamiana). Ecotoxicol Environ Saf. 2020, 198, 110671. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V.; Johnson, M.P.; Duffy, C.D.P. The photoprotective molecular switch in the photosystem II antenna. Biochimica et Biophysica Acta (BBA) – Bioenergetics. 2012, 1817, 167–181. [Google Scholar] [CrossRef]
- Wu, H.; et al. Roles of salicylic acid in selenium-enhanced salt tolerance in tomato plants. Plant and Soil. 2023, 484, 569–588. [Google Scholar] [CrossRef]
- Hirotsu, N.; et al. The photosynthetic properties of rice leaves treated with low temperature and high irradiance. Plant Cell Physiol. 2005, 46, 1377–83. [Google Scholar] [CrossRef]
- Vaculík, M.; Pavlovič, A.; Lux, A. Silicon alleviates cadmium toxicity by enhanced photosynthetic rate and modified bundle sheath’s cell chloroplasts ultrastructure in maize. Ecotoxicol Environ Saf. 2015, 120, 66–73. [Google Scholar] [CrossRef]
- Sun, H.; et al. Selenium modulates cadmium-induced ultrastructural and metabolic changes in cucumber seedlings. RSC Adv. 2020, 10, 17892–17905. [Google Scholar] [CrossRef]
- He, S.; et al. Nano silicon dioxide reduces cadmium uptake, regulates nutritional homeostasis and antioxidative enzyme system in barley seedlings (Hordeum vulgare L.) under cadmium stress. Environ Sci Pollut Res Int. 2023, 30, 67552–67564. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; et al. Rhizobacterial Strain Bacillus megaterium BOFC15 Induces Cellular Polyamine Changes that Improve Plant Growth and Drought Resistance. Int J Mol Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; et al. Biochar alleviates Cd phytotoxicity by minimizing bioavailability and oxidative stress in pak choi (Brassica chinensis L.) cultivated in Cd-polluted soil. Journal of Environmental Management. 2019, 250, 109500. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of exogenous selenium on plant and leaf morphology of cabbage seedlings under cadmium stress.
Figure 1.
Effect of exogenous selenium on plant and leaf morphology of cabbage seedlings under cadmium stress.

Figure 2.
Effect of exogenous selenium on the growth of cabbage seedlings under cadmium stress. (A) leaf fresh weight, (B) leaf dry weight, (C) root fresh weight, (D) root dry weight, (E) plant height, (F) stem thick, (G) root length, (H) Chla+b content, and (I) root activity. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
Figure 2.
Effect of exogenous selenium on the growth of cabbage seedlings under cadmium stress. (A) leaf fresh weight, (B) leaf dry weight, (C) root fresh weight, (D) root dry weight, (E) plant height, (F) stem thick, (G) root length, (H) Chla+b content, and (I) root activity. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
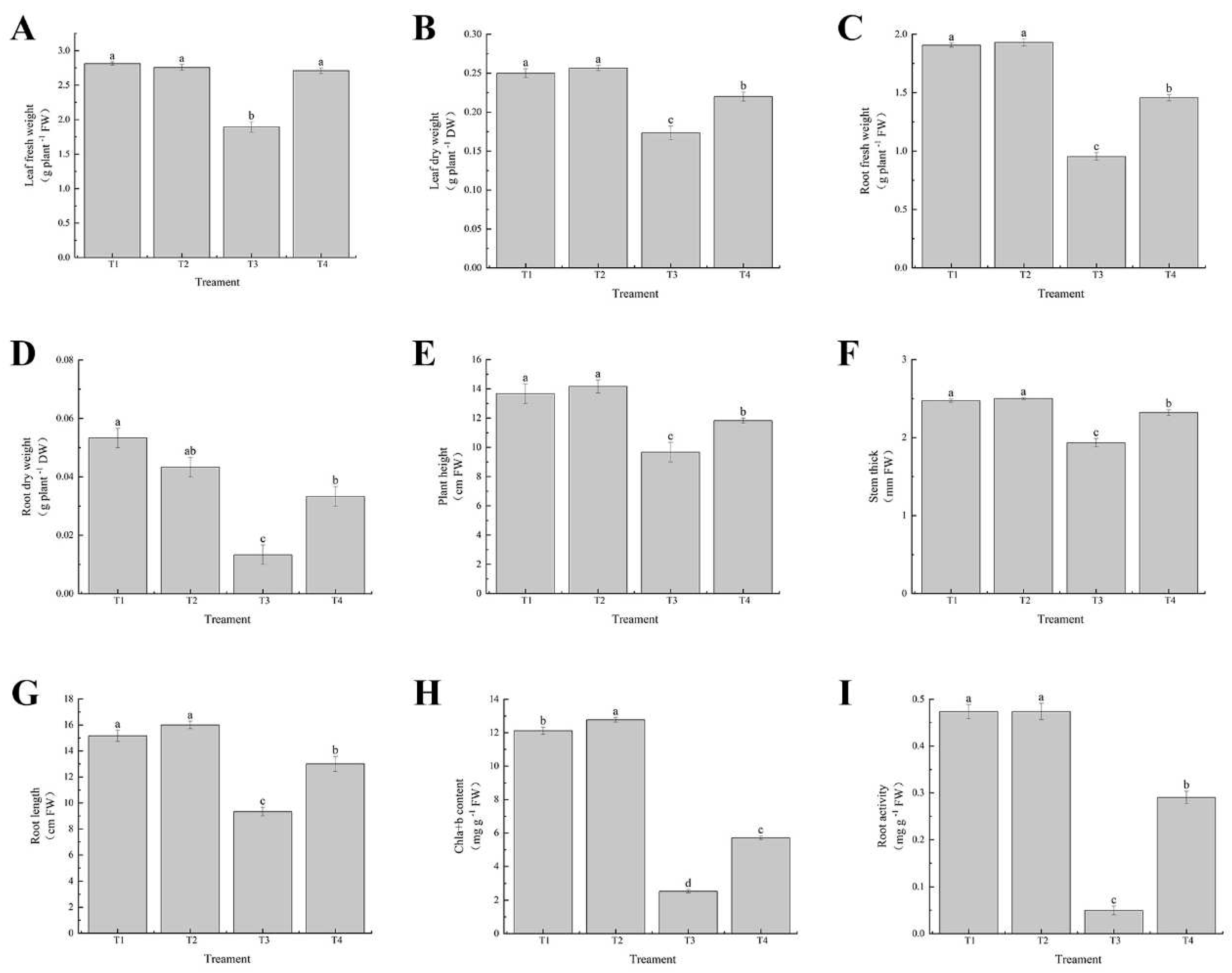
Figure 3.
Effects of exogenous selenium on stomatal parameters of cabbage seedlings under cadmium stress. (A) Pn, (B) Ci, (C) Tr, (D) Gs. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
Figure 3.
Effects of exogenous selenium on stomatal parameters of cabbage seedlings under cadmium stress. (A) Pn, (B) Ci, (C) Tr, (D) Gs. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
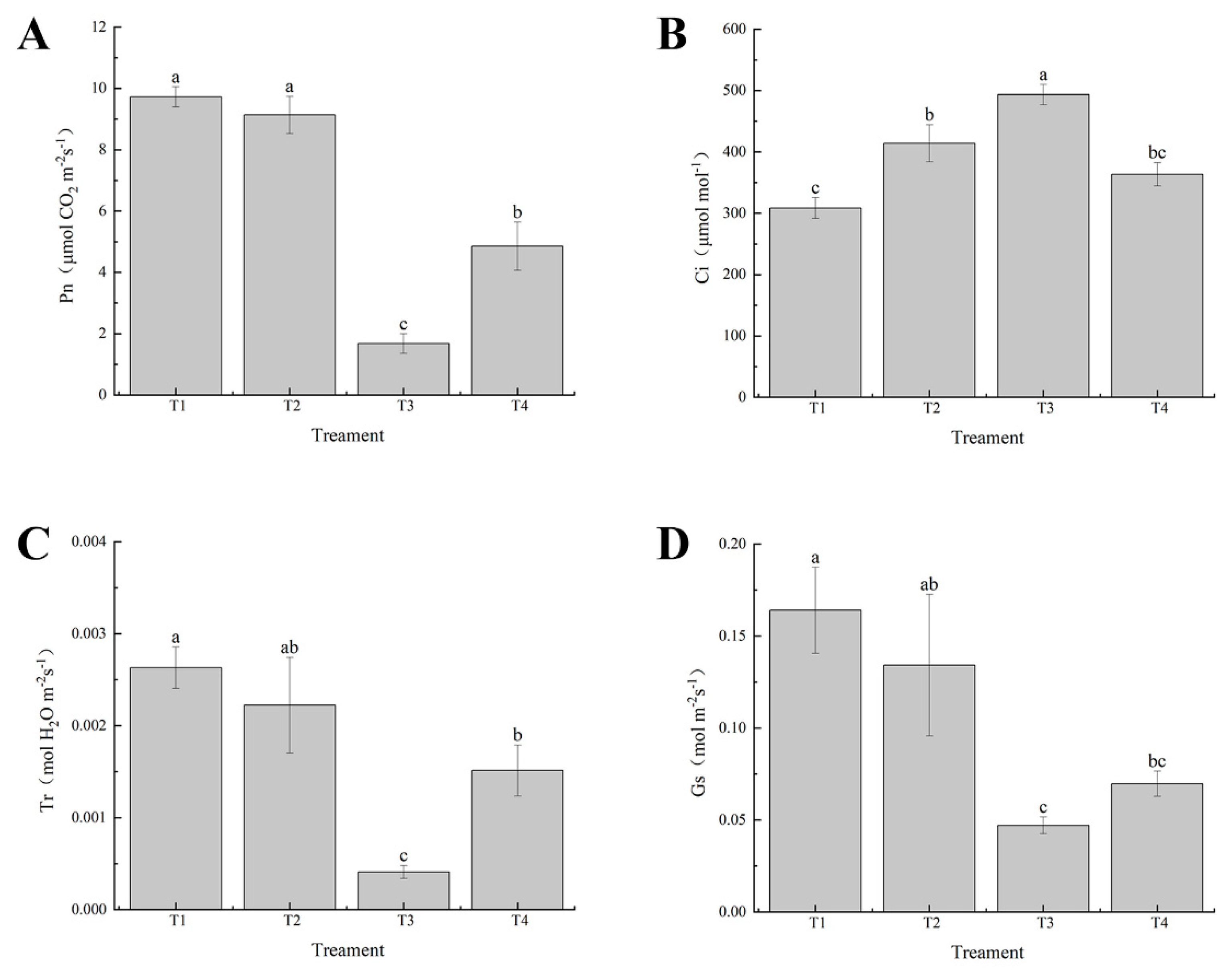
Figure 4.
Effects of exogenous selenium on chlorophyll fluorescence parameters of cabbage seedlings under cadmium stress. (A) Fv/Fm, (B) Y (Ⅱ), (C) Tr, (D) qP. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
Figure 4.
Effects of exogenous selenium on chlorophyll fluorescence parameters of cabbage seedlings under cadmium stress. (A) Fv/Fm, (B) Y (Ⅱ), (C) Tr, (D) qP. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
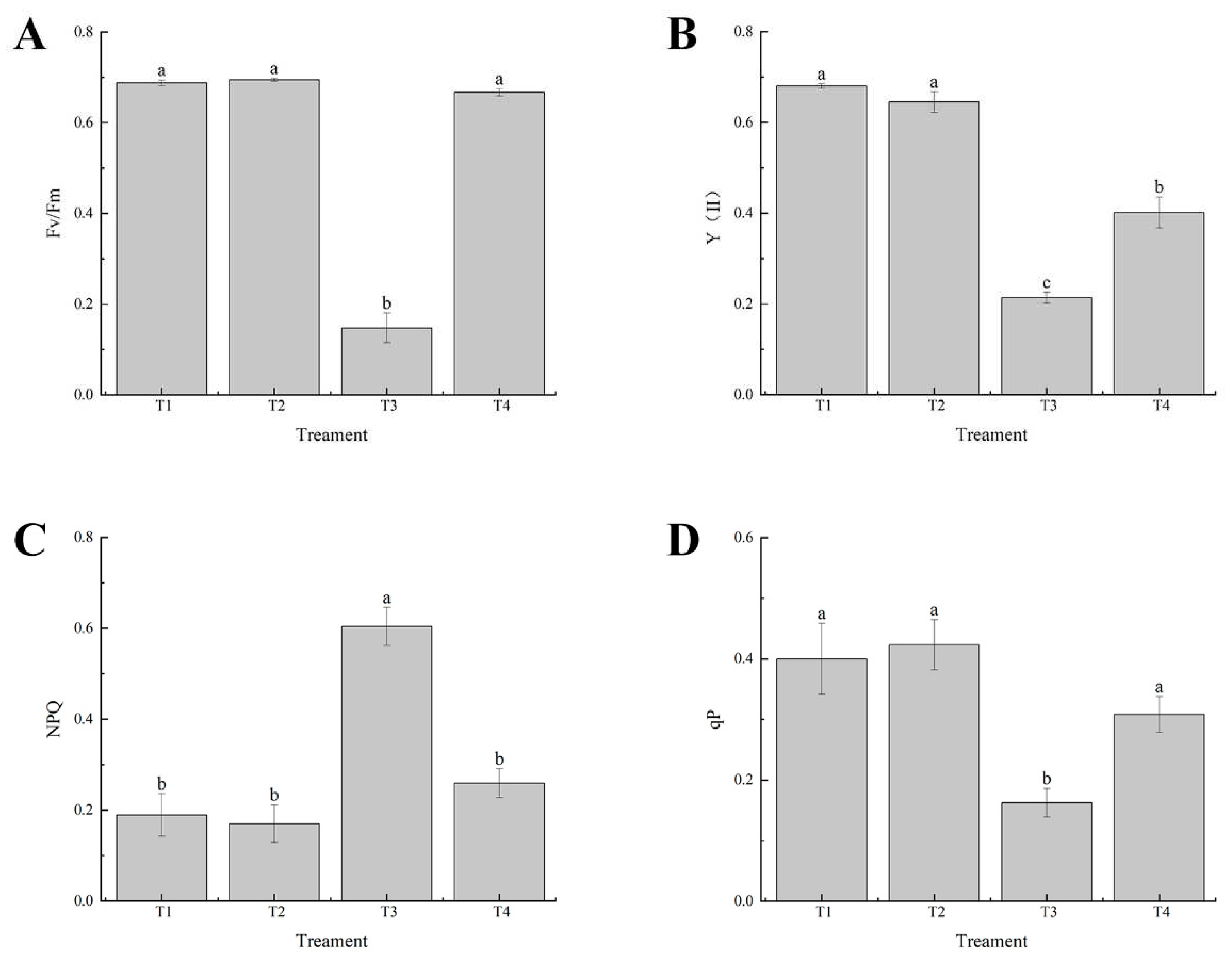
Figure 5.
Effects of exogenous selenium on tolerance of cabbage seedlings under cadmium stress. (A) Cd content (B) Se content. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
Figure 5.
Effects of exogenous selenium on tolerance of cabbage seedlings under cadmium stress. (A) Cd content (B) Se content. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
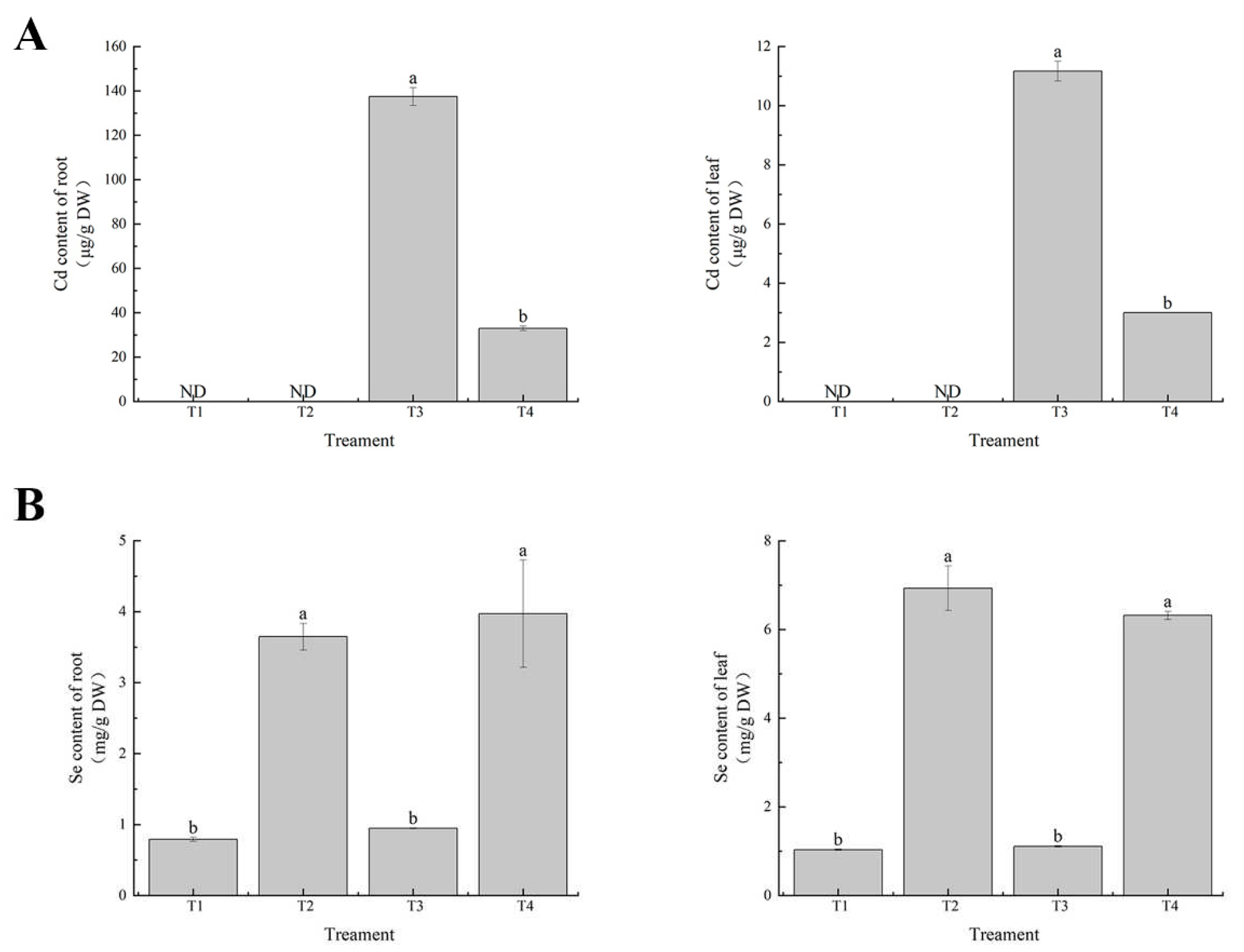
Figure 6.
Observation on chloroplast ultrastructure of cabbage seedling leaves. T1. (A–C), T2. (D–F), T3. (G–I), T4. (J–L). Abbreviation: CW, cell wall. CM, cell membrane. Ch, chloroplast. OG, osmiophilic globule. GL, grana lamellae. M, mitochondria. V, vacuole. SG, starch granules.
Figure 6.
Observation on chloroplast ultrastructure of cabbage seedling leaves. T1. (A–C), T2. (D–F), T3. (G–I), T4. (J–L). Abbreviation: CW, cell wall. CM, cell membrane. Ch, chloroplast. OG, osmiophilic globule. GL, grana lamellae. M, mitochondria. V, vacuole. SG, starch granules.
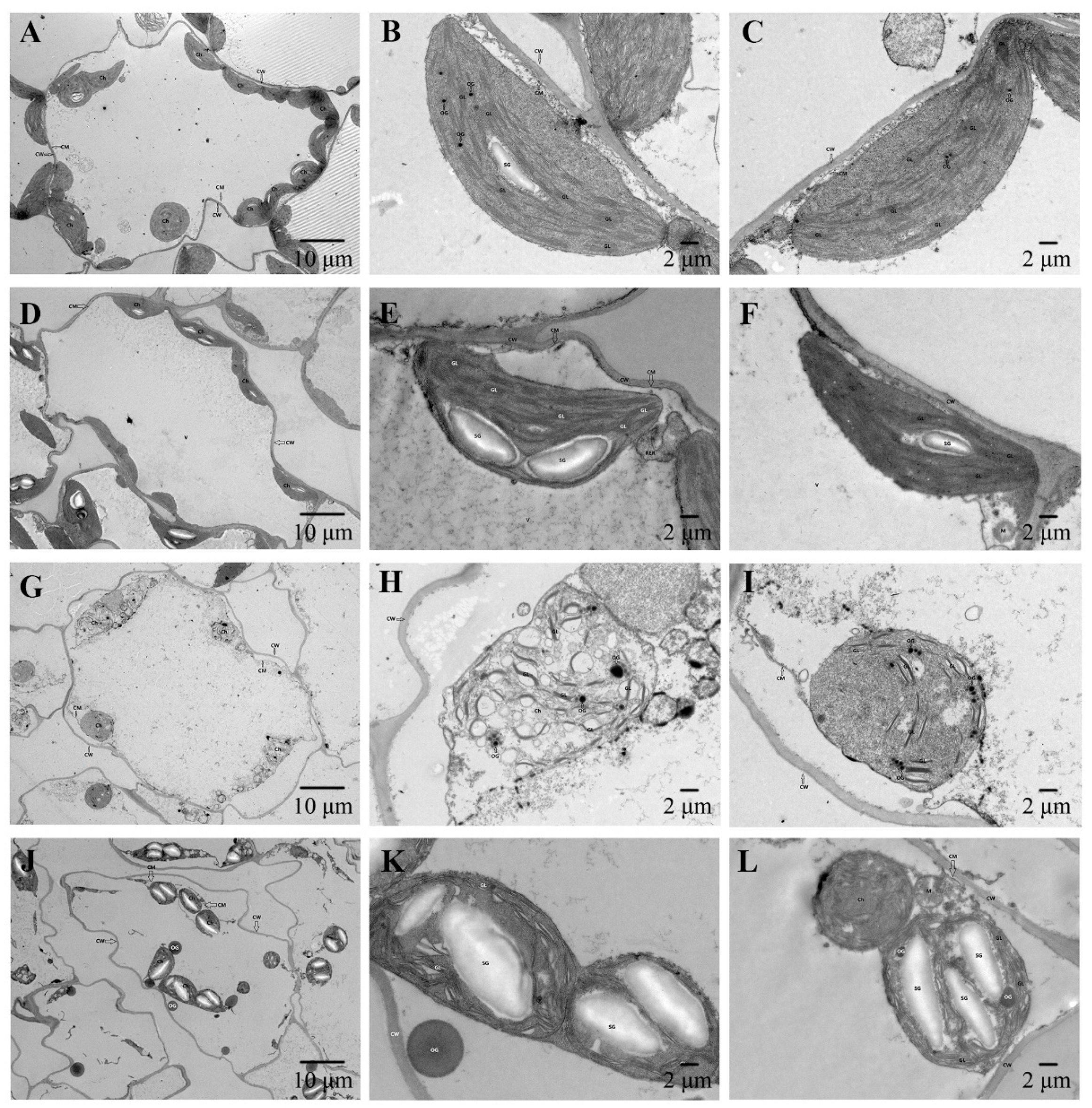
Figure 7.
Effects of exogenous selenium on DAB and NBT staining of leaves of cabbage seedlings under cadmium stress (A,B).
Figure 7.
Effects of exogenous selenium on DAB and NBT staining of leaves of cabbage seedlings under cadmium stress (A,B).

Figure 8.
Effects of exogenous selenium onO2- content, H2O2 content, Proline content, MDA content, Electrolyte leakage of cabbage seedlings under cadmium stress (A–E). Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
Figure 8.
Effects of exogenous selenium onO2- content, H2O2 content, Proline content, MDA content, Electrolyte leakage of cabbage seedlings under cadmium stress (A–E). Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
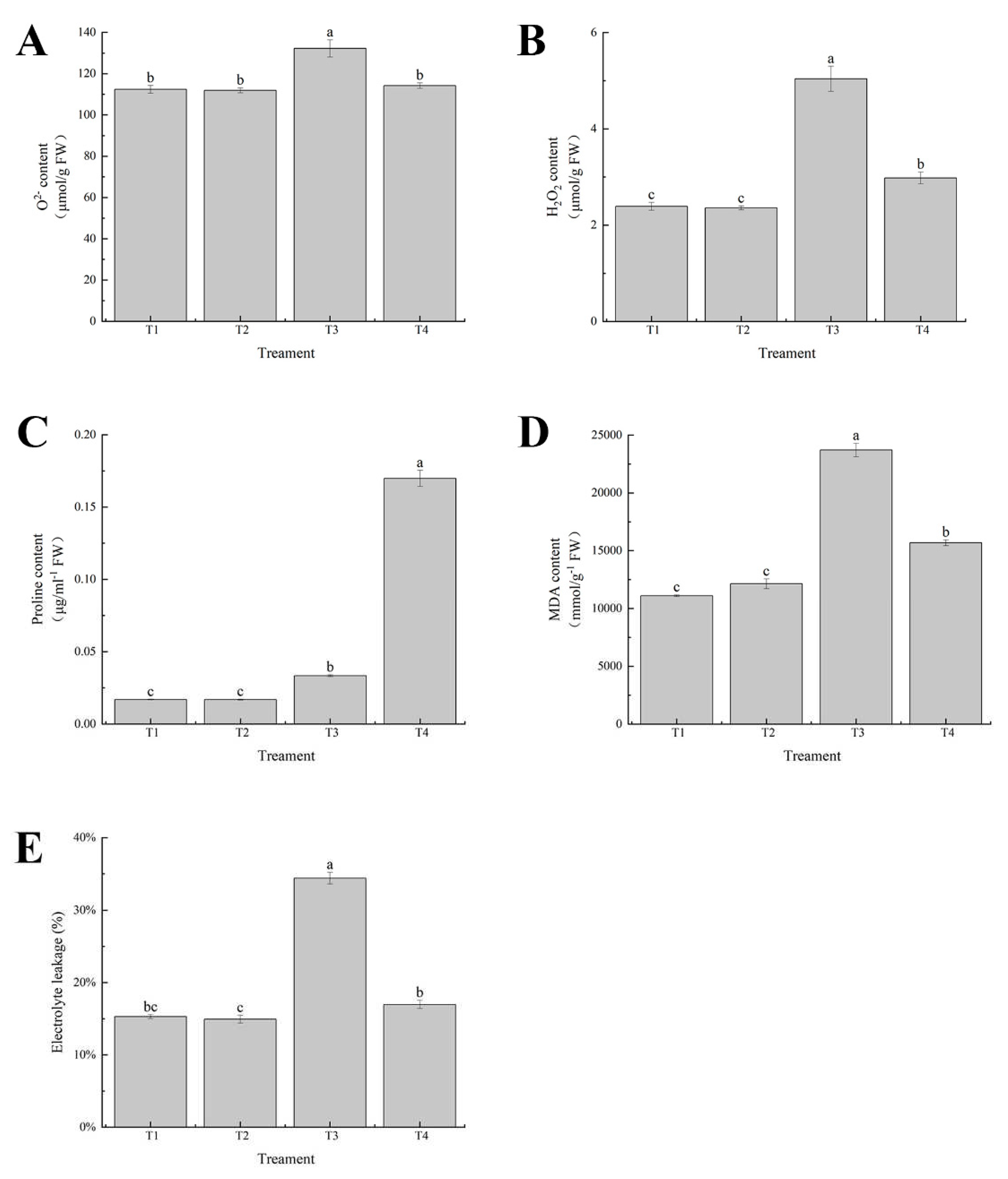
Figure 9.
Effects of exogenous selenium on antioxidant enzyme activities of cabbage seedlings under cadmium stress. (A) SOD activity, (B) POD activity, (C) APX activity, (D) GSH content. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
Figure 9.
Effects of exogenous selenium on antioxidant enzyme activities of cabbage seedlings under cadmium stress. (A) SOD activity, (B) POD activity, (C) APX activity, (D) GSH content. Values are means of three replicates ± SD. Different letters indicate significant difference at P ≤ 0.05.
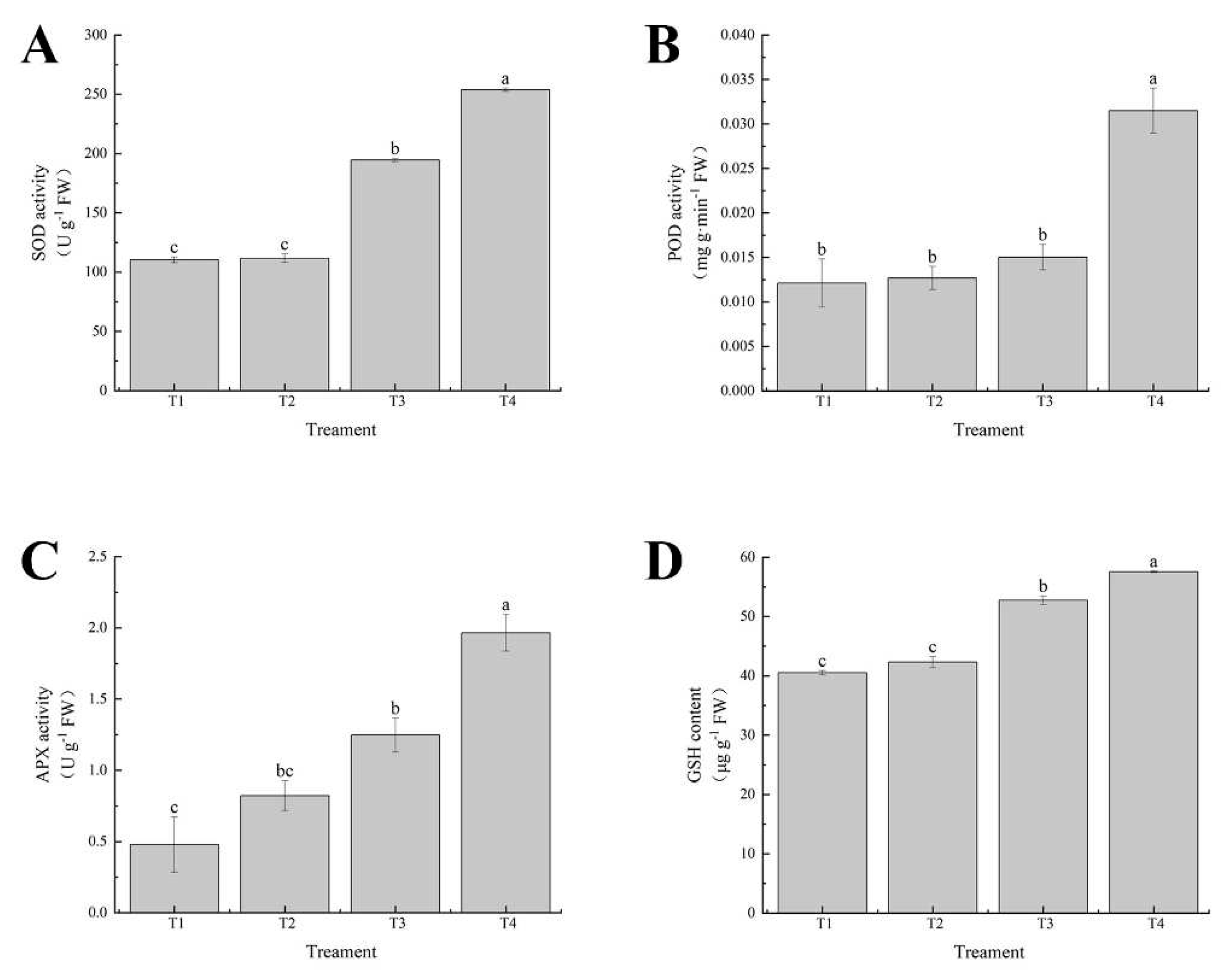
Figure 10.
Exogenous selenium has an alleviating effect on the physiological mechanism of cabbage seedlings.
Figure 10.
Exogenous selenium has an alleviating effect on the physiological mechanism of cabbage seedlings.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



