
Submitted:
18 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
The aim of this study was to investigate the biochemical properties and gel-forming capacity of duck myofibrillar proteins under the effects of 2,2'-azobis (2-amidinopropane) dihydrochloride (AAPH)-mediated oxidation. Duck myofibrillar proteins were extracted and treated with different concentrations of AAPH solutions (0, 1, 3, 5, 10 mmol/L) and then analysed for carbonyl content, dynamic rheology, protein profiles and gel-forming properties (colour, water holding capacity, hardness, protein profile, microstructure). The results showed that with increasing AAPH concentration, the carbonyl content of protein showed an increasing trend (p < 0.05); SDS-PAGE pattern results indicated that moderate oxidation (3 mmol/L AAPH) induced myosin aggregation via covalent bonds including disulfide, enhanced protein-protein, and thus improved protein-water interactions and gel strength of DMPs heat-induced gels. However, high oxidation (5 and 10 mmol/L AAPH) led to partial degradation of myosin heavy chain (MHC), as evidenced by lower storage modulus and irregular microstructure, which significantly reduced gelation ability. These results suggest that the internal relationship between alkanoperoxy radical-induced oxidation should be taken into account in the processing of duck meat, as mild protein oxidation is conducive to improving gel quality.
Keywords:
AAPH
; Protein oxidation
; Myofibrillar protein
; Gel properties
1. Introduction
For a long time, lipids oxidation and the role of micro-organisms have been considered to be an important factor in the deterioration of meat product quality. In recent years, the influence of protein oxidation on the quality of meat products has been gradually recognized [1,2]. Muscle protein could promote the tissue structure of meat products, in particular its gelling property plays an important role in the quality and sensory performance of meat products [3]. However, myofibrillar proteins is a very sensitive substance to reactive oxygen free radicals. Oxidation of meat is inevitable during processing and storage. In addition, oxidative damage will alter the physical, chemical and functional properties of proteins, thus reducing the sensory and nutritional quality of products [4,5]. Under the action of multi-layer metal ions, the oxidation of myofibrillar proteins would lead to the formation of protein carbonyls, which greatly reduces the properties of gel [6]. At the same time, Liu et al. found that mild oxidation can promote the cross-linking of disulfide bonds between protein molecules, thus making the protein network structure more stable [7]. Our previous papers mainly investigated the oxidation modification on the gel-forming capacity of duck myofibrillar proteins (DMPs), revealed that excessive oxidation led to explicit cross-linking of DMPs, which negatively affected the gel-forming capacity of DMPs by hydroxy radicals and malondialdehyde, but moderate oxidation of malondialdehyde was beneficial to improve the gel properties [8,9]. 2,2'-Azobis (2-Amidinopropane) Dihydrochloride (AAPH) generates alkanoperoxy radicals on degradation, which preferentially oxidize methionine and tryptophan residues, respectively, thereby causing protein oxidation [10,11]. In general, the actual meat product system is composed of protein and lipids. When lipids are rapidly oxidized, proteins are subsequently susceptible to oxidation as promoted by lipid-derived radicals and hydroperoxides [12]. However, to our knowledge, limited literature has discussed the oxidation effect of alkyl peroxide radical system on the gel-forming capacity of duck myofibrillar proteins. Some recent studies have shown the effect of alkanoperoxy radicals on the oxidation of porcine myofibrillar protein [13] and Casein [14]. Partial main chain breakage of proteins was also found. In this study, AAPH was used as a typical free radical intermediate for lipids oxidation [15], and the effects on different degrees of oxidation status (carbonyl, cross-linking) and also the properties of DMPs thermal gel, including rheological characteristics and textures, were investigated and their internal correlation was also discussed. The aim is to provide a theoretical basis to lay a foundation for further control of the oxidation level of duck meat and the rational use of oxidants.
2. Results and Discussion
3.1. Total Carbonyl Content
As shown in Figure 1, With increasing AAPH concentration, the protein carbonyl content tended to increase gradually, reaching a maximum of 12.0 nmol/mg protein at 10 mmol/L AAPH. It has been reported amino acids with NH or NH2 including arginine, lysine, threonine and proline, on their side chains are easily attacked by ROS. In the alkanoperoxy radical system, the peptide chains of proteins through the oxidation reaction of amino acid residues was easily attacked, promoting the formation of carbonyl derivatives [16]. The direct oxidation of glutamyl and aspartyl can break peptide bonds [17], subsequently leading to peptide cleavage, as a result of α-amidation pathway, β-scission reaction and glutamyl side-chain oxidation reaction, the cleavage of backbones is considered a major mechanism of protein carbonylation [18]. Therefore, oxidative damage to proteins can be reflected by carbonyl derivatives [19,20].
3.2. SDS-PAGE Pattern
Oxidation cross-links proteins and simultaneously produces a variety of polymers, and the cross-linking is mostly associated with the myosin component of DMPs [21]. As shown in Figure 2 that the concentration of AAPH has a certain effect on the structural composition of proteins. When AAPH concentration is 0, 1 and 3 mmol/L, it can promote the cross-linking of proteins and generate protein polymers. When the AAPH concentration is 5-10 mmol/L, it can partially degrade protein components. In the case of non-reducing conditions (- DTT), because there are a large number of high molecular weight myosin cross-links in DMPs, proteins are clustered in large quantities in up-layer of stacking gel; it can be speculated that the protein is attacked by alkanoperoxy radicals, causing the sulfhydryl group to form a disulfide bond and other covalent bond, leading to cross-linking of the protein, increasing its molecular weight and aggregation. Furthermore, the band intensity of myosin heavy chain (MHC) firstly increased and then decreased with increasing AAPH concentration, reaching a maximum at 3 mmol/L AAPH concentration and then decreasing with increasing AAPH concentration. It is notable to mention that myofibrillar proteins have a large number of sulfhydryl groups, among which myosin alone contains 42 sulfhydryl groups [22], easily attacked by reactive oxygen species to form disulphide bonds, leading to changes in protein structure and further affecting protein function. This stage is a reversible oxidation reaction, whereas irreversible oxidation reactions could also produce sulfinic acid and sulfonic acid [23]. Under the reducing condition (+ DDT), the high molecular weight protein almost completely disappeared in up-layer of stacking gel, and the intensity of MHC and actin significantly increased, indicating that disulfide bonds play an important role in protein cross-linking, and MHC and actin proteins are the main proteins that undergo cross-linking and aggregation during oxidation [24]; Around 17 kD, samples without DTT have clear bands of myosin light chain 3 and fuzzy bands of myosin light chain 2, whereas in the reduced condition, the intensity of the myosin light chain 3 band of the samples decreases significantly and myosin light chain 2 could be clearly seen. At the same time, higher concentration of AAPH could induce aggregation of myosin and denaturation of myosin, troponin and tropomyosin, and also lead to protein degradation [25]. In particular, both actin and myosin light chain interacted with other peptides. This resulted in an increase in molecular weight, as indicated by the migration site. As reported, this means that alkanoperoxy radicals would induce formation of secondary free radicals which self-react to produce inter- and intra- molecular covalent bonds. Formation of ditryptophan bonds has also been implicated in oxidative cross-linking of proteins. It is known that carbonyl groups undergo secondary reactions with basic residues (N-terminal amino groups) to generate schiff bases inducing protein cross-linking [26].
3.3. Dynamic Rheological Properties
From Figure 3, it can be seen that the change in rheological properties of DMP oxidation is also related to the concentration of AAPH. At the end of the heating phase, the storage modulus of the protein increases with increasing AAPH concentration, especially at 3 mmol/L. Meanwhile, the trend of the loss modulus is similar to that of the storage modulus. At the same time, at the heating stage, the value of the storage modulus is always greater than that of the loss modulus at the same temperature. The research of Feng and Xiong et al. [27] showed that at a high temperature of 65 ℃, the denaturation of myosin causes the expansion of the structure, and the activated groups are also exposed. This change promotes cross-linking of the protein, which improves the viscoelasticity of the colloid and increases the storage modulus (G '). It was noteworthy that according to the analysis of SDS-PAGE in section 3.2, after treatment with higher concentration of AAPH, MHC is degraded to a certain extent [28]. Among them, the change range of protein G' after 3 mmol/L AAPH oxidation treatment is steeper and higher than that of other concentration of AAPH oxidation treatment. Simultaneously, the decrease in myosin thermal stability is caused by the partial degradation of MHC caused by high concentration oxidation, it causes the denaturation of myosin at the later stage of heating, which remarkably reduces the G' of the system. Apparently, moderate oxidation is conducive to improving the strength of the protein gel. Much protein aggregation is caused by protein oxidation. Protein aggregation or change in protein structure will have a major effect on its functional properties and the most obvious is the change in gel strength.
3.4. Gel strength
Figure 4 shows the effect of oxidation on the gel strength of myofibrillar protein gel. It can be seen from the figure that the strength of the myofibrillar protein gel first subtle increases and then decreases. This shows that protein oxidation can improve the gel strength of myofibrillar protein to a certain extent. The higher the gel strength, the denser, stronger and more stable the structure of the gel. Xiong [29] have shown that myofibrillar proteins can easily form protein gel under thermal action after low concentration oxidation treatment, and the strength of the gel is increased due to the interaction between proteins. Utrera et al [30] showed that protein carbonylation caused by oxidation also changed the gel properties of the protein. According to the results of SDS-PAGE, high concentration oxidation treatment will significantly reduce the polymerisation ability of protein gel and increase the brittleness of gel.
3.5. Whiteness and water holding capacity of gel
Protein denaturation affects the whiteness of the gel [31]. As shown in Figure 5(a), the whiteness of the gel decreased significantly with increasing AAPH concentration (P < 0.05). The water holding capacity (WHC) of the gel can reflect the roughness of the internal structure of the gel, which means that the gel could hold more water or prevent water seepage under different physical and chemical conditions. As can be seen from Figure 5(b), as the AAPH concentration increases, the water holding capacity of the protein gel tends to first increase and then decrease. When the AAPH concentration is 4 mmol/L, the maximum water holding capacity is reached. When the AAPH concentration is 10 mmol/L, the water holding capacity of the gel decreases, which may be due to the degradation of protein components under high concentration oxidation, making it more difficult to form gel. Lund et al. [32] have indicated the consequences of protein oxidation in muscle food have often been associated with changes in solubility and protein functionality such as gelation and emulsifying properties, or WHC.
3.6. Water status in gel
It can be seen from Figure 6 that the concentration of AAPH has an effect on the NMR relaxation time of the DMPs heat-induced gel. Han Minyi et al [33] previously reported that there were generally 4 types of peaks in the NMR relaxation curve of myofibrillar protein gel after fitting, which corresponded to the states of 4 kinds of water, including bound bond water (T2b), moderately immobilised water (T21), immobilised water (T22), and free water (T23). The second peak and the third peak were collectively classified as immobilised water. In our study, it could be seen from the figure that the T2 relaxation intensity has a peak of moderately immobilised water between 10-95 Ms (T21), a peak of immobilised water between 100-1000 Ms (T22) and a peak of free water after 1000 Ms (T23). The peak with the largest peak area in the figure corresponds to immobilized water, indicating that myofibrillar proteins cross-link with each other when heated to form a network structure that binds a large amount of water molecules. When the concentration of AAPH is relatively low (0 - 3 mmol/L), the area of free water gradually decreases. whereas the result is the opposite when the concentration of AAPH is higher than 5 mmol/L. Li Yin et al combined the traditional drying method with the low field NMR technology and proved that the free water content is inversely proportional to the water holding capacity of the gel [34]. This means that our NMR results are just right and show the same trend as the previous WHC results.
3.7. Gel microstructure
The microstructure of the protein gel is an important method for studying the structure and properties of the gel. The SEM image of the protein gel treated with 4 % glutaraldehyde is shown in Figure 7. The results showed that the internal structure of the gel changed significantly after AAPH oxidised the protein. Proper oxidation can promote protein cross-linking, thus encouraging irregular aggregates to participate in the gel [35], thus improving the network structure. As shown in Figure 7, the structure of the DMPs gel deteriorates with increasing AAPH concentration and the degree of deterioration also increases. When the concentration of AAPH is 10 mmol/L, the micropores continue to expand and break, forming fibre fragments, and the gel structure begins to collapse with irregular cracks.
3. Materials and Methods
3.1. Sample Preparation and Reagents
Duck breasts, obtained from local Market in Lishui District, Nanjing, Jiangsu Province, were taken to lab at 4 °C, then divided into 30 g per sealed bag and stored at - 80 °C before use. The reagents including 2,2-Azobis (2-methylpropyl imido) dihydrochloride, AAPH) were obtained from shangha imacklin biochemical co. Ltd. Shanghai, China, which are analytically pure.
3.2. Extraction of Duck Myofibrillar Proteins (DMPs)
Duck myofibrillar proteins were extracted according to zhu et al.[36] Briefly, the -80 °C frozen duck breast was taken out, poured into centrifuge tubes, and then added 5 times the volume of extraction solution (100 mmol/L NaCl, 2 mmol/L MgCl2, 1 mmol/L EGTA, 10 mmol/L K2HPO4, pH 7. 0) and then centrifuged at 4 °C, 4000 × g for 10 min, and The supernatant was then discarded. The above steps were repeated for three times, also filtered through two layers of sterilized gauze in the meantime; Subsequently, Precipitation was homogenised with 5 times the volume of extract (12.5 mmol/L NaCl, 2.5 mmol/L MgCl2, 1. 25 mmol/L EGTA, 12.5 mmol/L K2HPO4, 1% Triton X-100, pH 7.0), then centrifuged at 4 °C 4000 × g for 10 min to remove membrane protein; finally, 4 times the volume of 0. 1 mol/L NaCl was added, then centrifuged at 4 °C, 4000×g, for 10 min, Final precipitate was collected to obtain duck myofibrillar proteins.
3.3. Oxidation of Duck Myofibrillar Proteins
The protein content of DMPs was firstly measured by the biuret method and then diluted to 30 mg/mL. The oxidation solution for duck myofibrillar proteins was prepared according to Table 1 and then incubated for 24 h at 4 °C with shaking and avoidance of light. After oxidation, DMPs was immediately centrifuged at 4 °C 8000 × g for 10 min. The precipitate was washed twice with an appropriate volume of distilled water and centrifuged again under the same conditions. The supernatant was then discarded and the precipitate was considered as a sample of oxidised myofibrillar protein.
3.4. Carbonyl Content
The carbonyl content of protein was determined according to the method of Soglia et al.[37], with slight modifications, and the carbonyl content of protein was determined by the 2,4-dinitrophenylhydrazine colour development method. In brief The carbonyl content is expressed in nmol/mg protein in terms of the molar extinction coefficient 22,000 L/(mol·cm), then calculated according to Equation (1)
3.5. Sodium Dodecyl Sulfonate Polyacrylamide Gel Electrophoresis Analysis (SDS-PAGE)
SDS-PAGE was performed according to the method of Jia et al [38], with slight modifications, the concentration of protein solution was adjusted to 2 mg/ml, mixed with an equal amount of 2×standard SDS sample loading buffer with or without dithiothreitol (DTT), and heated for 3 min in a dry heater (100 °C). SDS-PAGE electrophoresis run at 220V for 45 min using NuPAGETM 5-12% Bis-Tris gel (Invitrogen). The gel was then stained with Coomassie Brilliant Blue G-250 before analyse.
3.6. Dynamic Rheological Test
Protein solution was measured using a rheometer (MCR-301, Anton Paar, Graz, Austria) in oscillatory mode as described by Zhuang et al [39]. Brief parameters were set as follow: 50 mm flat plate with a gap of 1 mm, frequency of 0.1 Hz, strain of 2 %, temperature rise from 25 °C to 85 °C at rate of 2 °C/min and a temperature fall rate of 5 °C/min. The storage modulus (G') and loss modulus (G’’) was then recorded.
3.7. Gel Preparation
According to the method of Xia et al.[40], protein solution (40 mg/ml) were heated at 80 ℃ for 35 minutes to induce gelation, then cooled with ice water for 30 minutes, and stored 4 ℃ to equilibrate overnight.
3.8. Gel Strength Measurement
The strength of the gel was measured using the texture analyser (TAXT plus Plaser, Stable Micro Systems, Godalming, UK). Parameters: use P/0.5r probe, 0.5 mm/s rate, 0.5 mm/s test rate, 0.5 mm/s post test rate, 5 mm probe depth distance and trigger force of 4 g, and repeat each treatment sample for 3 times.
3.9. Gel Whiteness and Water Holding Capacity
The whiteness of gel was measured with with Minolta CR-400 (illuminant D65) equipment (Minolta Camera, Osaka, Japan), calibrated with a standard white board. C light source was used as the light source for the measurement, and L* , a* and b* of the gel were recorded. Whiteness of the gel is calculated according to salvador et al. [41] as follow equation (2):
The water holding capacity (WHC) of the gel was determined by centrifugation methods according to Zhu et al. [42].
3.10. Water Status in Gel
The T2 relaxation time of the water in samples was measured using an NMR analyzer (MesoMR23-060H-1, Niumag electric Co., Shanghai, China) according to Xia et al. [43]. Firstly, a standard oil sample was calibrated, then about 2 g of sample was placed in a centrifuge tube and placed in the instrument, and the CPMG sequence was selected according to the spin-spin relaxation time. The proton resonance frequency was set at 22.6 MHz and the measurement was performed at 32 °C. Parameters were set as follow: the number of repeated samples is 4, the waiting time is 2000 ms, the number of echoes is 9000. The resulting curve is an exponential decay sample curve.
3.11. Gel Microstructure
Gel sample were firstly cutted into cubes (3 × 3 × 3 mm3), fixed with 4 % glutaraldehyde, then observed with a Hitachi S-3000N scanning electron microscope (Tokyo, Japan) at an accelerating voltage of 20 kV.
3.12. Statistical Analysis
SPSS 19.0 (version 20, SPSS Inc., Chicago, IL, USA) was used for one-way ANOVA, and Duncan's multiple comparison method was used for statistical analysis (P < 0.05 indicates significant difference)
4. Conclusions
As the concentration of AAPH increases, the carbonyl group content of the protein increases. At low concentrations of AAPH (1-3 mmol/L), the protein is attacked by peroxy radicals, which cross-link the protein, increase its molecular weight and form aggregates. Under the action of high concentrations of AAPH (5-10 mmol/L), protein components are degraded. In particular, at 10 mmol/L AAPH, the gel structure begins to collapse and large irregular cracks appear, making gel formation more difficult. It can be seen that the peroxyl and alkoxyl radical system has an obvious effect on the gel properties of duck myofibrils, and at a suitable concentration of AAPH (3 mmol/L) the texture properties and water retention capacity of the gel can be effectively improved.
Author Contributions
Conceptualization, project administration, writing-review and editing, methodology; funding acquisition,Xueshen Zhu; Sample collection, investigation, data curation, writing-original draft, Jin Zhang; investigation, Xinyu Zhang; Visualization, validation, Qun Dai; Supervision, Qingquan Fu; All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by National Natural Science Foundation of China (31301507), Natural science fund for colleges and universities in Jiangsu Province (21KJA550002).
Institutional Review Board Statement
Not applicable.
Acknowledgments
The authors wish to thank Elife Technology Co., Ltd. for providing the samples and facilities for sample preparation, as well as Zhuran Wang for his assist statistical analysis of data of the paper.
Conflicts of Interest
There are no conflicts to declare.
Sample Availability
Samples of the compounds are available from the authors.
References
- Johns, A.M.; Birkinshaw, L.H.; Ledward, D.A. Catalysts of lipid oxidation in meat-products. Meat Science 1989, 25, 209-220. https://doi.org/10.1016/0309-1740(89)90073-9. [CrossRef]
- Dominguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.G.; Lorenzo, J.M. A Comprehensive Review on Lipid Oxidation in Meat and Meat Products. Antioxidants 2019, 8. https://doi.org/10.3390/antiox8100429. [CrossRef]
- Sohaib, M.; Anjum, F.M.; Sahar, A.; Arshad, M.S.; Rahman, U.U.; Imran, A.; Hussain, S. Antioxidant proteins and peptides to enhance the oxidative stability of meat and meat products: A comprehensive review. Int. J. Food Prop. 2017, 20, 2581-2593. https://doi.org/10.1080/10942912.2016.1246456. [CrossRef]
- Bao, Y.; Ertbjerg, P.; Estevez, M.; Yuan, L.; Gao, R. Freezing of meat and aquatic food: Underlying mechanisms and implications on protein oxidation. Compr. Rev. Food. Sci. Food Saf. 2021, 20, 5548-5569. https://doi.org/10.1111/1541-4337.12841. [CrossRef]
- Zhang, L.T.; Li, Q.; Bao, Y.L.; Tan, Y.Q.; Lametsch, R.; Hong, H.; Luo, Y.K. Recent advances on characterization of protein oxidation in aquatic products: A comprehensive review. Critical Reviews in Food Science and Nutrition. https://doi.org/10.1080/10408398.2022.2117788. [CrossRef]
- Bao, Y.L.; Ertbjerg, P. Effects of protein oxidation on the texture and water-holding of meat: a review. Critical Reviews in Food Science and Nutrition 2019, 59, 3564-3578. https://doi.org/10.1080/10408398.2018.1498444. [CrossRef]
- Liu, Z.; Xiong, Y.L.; Chen, J. Protein Oxidation Enhances Hydration but Suppresses Water-Holding Capacity in Porcine Longissimus Muscle. Journal of Agricultural and Food Chemistry 2010, 58, 10697-10704. https://doi.org/10.1021/jf102043k. [CrossRef]
- Zhu, X.; Shi, X.; Liu, S.; Gu, Y.; Liu, J.; Fu, Q.; Wang, R. Physicochemical properties and gel-forming ability changes of duck myofibrillar protein induced by hydroxyl radical oxidizing systems. Frontiers in Nutrition 2022, 9. https://doi.org/10.3389/fnut.2022.1029116. [CrossRef]
- Zhu, X.; Ma, Z.; Zhang, X.; Huang, X.; Liu, J.; Zhuang, X. Effect of Malondialdehyde-Induced Oxidation Modification on Physicochemical Changes and Gel Characteristics of Duck Myofibrillar Proteins. Gels 2022, 8. https://doi.org/10.3390/gels8100633. [CrossRef]
- Dion, M.Z.; Wang, Y.J.; Bregante, D.; Chan, W.M.; Andersen, N.; Hilderbrand, A.; Leiske, D.; Salisbury, C.M. The Use of a 2,2 '-Azobis (2-Amidinopropane) Dihydrochloride Stress Model as an Indicator of Oxidation Susceptibility for Monoclonal Antibodies. Journal of Pharmaceutical Sciences 2018, 107, 550-558. https://doi.org/10.1016/j.xphs.2017.09.022. [CrossRef]
- Sadeghinejad, N.; Sarteshnizi, R.A.; Gavlighi, H.A.; Barzegar, M. Pistachio green hull extract as a natural antioxidant in beef patties: Effect on lipid and protein oxidation, color deterioration, and microbial stability during chilled storage. LWT-Food Sci. Technol. 2019, 102, 393-402. https://doi.org/10.1016/j.lwt.2018.12.060. [CrossRef]
- Bridi, R.; Giordano, A.; Peñailillo, M.F.; Montenegro, G. Antioxidant Effect of Extracts from Native Chilean Plants on the Lipoperoxidation and Protein Oxidation of Bovine Muscle. Molecules 2019, 24, 3264. [CrossRef]
- Zhou, F.; Zhao, M.; Zhao, H.; Sun, W.; Cui, C. Effects of oxidative modification on gel properties of isolated porcine myofibrillar protein by peroxyl radicals. Meat Science 2014, 96, 1432-1439. https://doi.org/10.1016/j.meatsci.2013.12.001. [CrossRef]
- Fuentes-Lemus, E.; Silva, E.; Barrias, P.; Aspee, A.; Escobar, E.; Lorentzen, L.G.; Carroll, L.; Leinisch, F.; Davies, M.J.; López-Alarcón, C. Aggregation of α- and β- caseins induced by peroxyl radicals involves secondary reactions of carbonyl compounds as well as di-tyrosine and di-tryptophan formation. Free Radical Biology and Medicine 2018, 124, 176-188. https://doi.org/10.1016/j.freeradbiomed.2018.06.005. [CrossRef]
- Werber, J.; Wang, Y.J.; Milligan, M.; Li, X.; Ji, J.A. Analysis of 2,2 '-Azobis (2-Amidinopropane) Dihydrochloride Degradation and Hydrolysis in Aqueous Solutions. Journal of Pharmaceutical Sciences 2011, 100, 3307-3315. https://doi.org/10.1002/jps.22578. [CrossRef]
- 1Wu, W.; Zhang, C.M.; Kong, X.Z.; Hua, Y.F. Oxidative modification of soy protein by peroxyl radicals. Food Chemistry 2009, 116, 295-301. https://doi.org/10.1016/j.foodchem.2009.02.049. [CrossRef]
- Essex, D.W.; Li, M.R.; Miller, A.; Feinman, R.D. Protein disulfide isomerase and sulfhydryl-dependent pathways in platelet activation. Biochemistry 2001, 40, 6070-6075. https://doi.org/10.1021/bi002454e. [CrossRef]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein Oxidation: Basic Principles and Implications for Meat Quality. Critical Reviews in Food Science and Nutrition 2013, 53, 1191-1201. https://doi.org/10.1080/10408398.2011.577540. [CrossRef]
- Stadtman, E.R.; Levine, R.L. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids 2003, 25, 207-218. https://doi.org/10.1007/s00726-003-0011-2. [CrossRef]
- Estevez, M. Protein carbonyls in meat systems: A review. Meat Science 2011, 89, 259-279. https://doi.org/10.1016/j.meatsci.2011.04.025. [CrossRef]
- Grune, T.; Jung, T.; Merker, K.; Davies, K.J.A. Decreased proteolysis caused by protein aggregates, inclusion bodies, plaques, lipofuscin, ceroid, and 'aggresomes' during oxidative stress, aging, and disease. International Journal of Biochemistry & Cell Biology 2004, 36, 2519-2530. https://doi.org/10.1016/j.biocel.2004.04.020. [CrossRef]
- Hofmann, K.; Hamm, R. Sulfhydryl and Disulfide Groups in Meats**Dedicated to Professor Dr. Alfons Schoberl, Hannover (Germany), a pioneer in the chemistry of organic sulfur compounds. In Advances in Food Research, Chichester, C.O., Ed.; Academic Press: 1978; Volume 24, pp. 1-111.
- Morzel, M.; Gatellier, P.; Sayd, T.; Renerre, M.; Laville, E. Chemical oxidation decreases proteolytic susceptibility of skeletal muscle myofibrillar proteins. Meat Science 2006, 73, 536-543. https://doi.org/10.1016/j.meatsci.2006.02.005. [CrossRef]
- Dorta, E.; Avila, F.; Fuentes-Lemus, E.; Fuentealba, D.; Lopez-Alarcon, C. Oxidation of myofibrillar proteins induced by peroxyl radicals: Role of oxidizable amino acids. Food Research International 2019, 126. https://doi.org/10.1016/j.foodres.2019.108580. [CrossRef]
- Relkin, P.; Fabre, M.; Guichard, E. Effect of Fat Nature and Aroma Compound Hydrophobicity on Flavor Release from Complex Food Emulsions. Journal of Agricultural and Food Chemistry 2004, 52, 6257-6263. https://doi.org/10.1021/jf049477a. [CrossRef]
- Fuentes-Lemus, E.; Silva, E.; Leinisch, F.; Dorta, E.; Lorentzen, L.G.; Davies, M.J.; López-Alarcón, C. α- and β-casein aggregation induced by riboflavin-sensitized photo-oxidation occurs via di-tyrosine cross-links and is oxygen concentration dependent. Food Chemistry 2018, 256, 119-128. https://doi.org/10.1016/j.foodchem.2018.02.090. [CrossRef]
- Feng, J.; Xiong, Y.L. Interaction and Functionality of Mixed Myofibrillar and Enzyme-hydrolyzed Soy Proteins. Journal of Food Science 2003, 68, 803-809. https://doi.org/10.1111/j.1365-2621.2003.tb08246.x. [CrossRef]
- Zhu, Z.; Yang, J.; Zhou, X.; Khan, I.A.; Bassey, A.P.; Huang, M. Comparison of two kinds of peroxyl radical pretreatment at chicken myofibrillar proteins glycation on the formation of N-epsilon-carboxymethyllysine and N-epsilon-carboxyethyllysine. Food Chemistry 2021, 353. https://doi.org/10.1016/j.foodchem.2021.129487. [CrossRef]
- Xiong, Y.L.; Blanchard, S.P.; Ooizumi, T.; Ma, Y.Y. Hydroxyl Radical and Ferryl-Generating Systems Promote Gel Network Formation of Myofibrillar Protein. Journal of Food Science 2010, 75, C215-C221. https://doi.org/10.1111/j.1750-3841.2009.01511.x. [CrossRef]
- Utrera, M.; Estevez, M. Oxidation of Myofibrillar Proteins and Impaired Functionality: Underlying Mechanisms of the Carbonylation Pathway. Journal of Agricultural and Food Chemistry 2012, 60, 8002-8011. https://doi.org/10.1021/jf302111j. [CrossRef]
- Hwang, J.-S.; Lai, K.-M.; Hsu, K.-C. Changes in textural and rheological properties of gels from tilapia muscle proteins induced by high pressure and setting. Food Chemistry 2007, 104, 746-753. https://doi.org/10.1016/j.foodchem.2006.11.075. [CrossRef]
- Lund, M.N.; Heinonen, M.; Baron, C.P.; Estevez, M. Protein oxidation in muscle foods: A review. Molecular Nutrition & Food Research 2011, 55, 83-95. https://doi.org/10.1002/mnfr.201000453. [CrossRef]
- Han, M.; Wang, P.; Xu, X.; Zhou, G. Low-field NMR study of heat-induced gelation of pork myofibrillar proteins and its relationship with microstructural characteristics. Food Research International 2014, 62, 1175-1182. https://doi.org/10.1016/j.foodres.2014.05.062. [CrossRef]
- Li, Y.; Li, X.; Wang, J.-z.; Zhang, C.-h.; Sun, H.-m.; Wang, C.-q.; Xie, X.-l. Effects of Oxidation on Water Distribution and Physicochemical Properties of Porcine Myofibrillar Protein Gel. Food Biophysics 2014, 9, 169-178. https://doi.org/10.1007/s11483-013-9329-9. [CrossRef]
- Wang, Z.; He, Z.; Gan, X.; Li, H. Effect of peroxyl radicals on the structure and gel properties of isolated rabbit meat myofibrillar proteins. International Journal of Food Science and Technology 2018, 53, 2687-2696. https://doi.org/10.1111/ijfs.13878. [CrossRef]
- Zhu, X.S.; Ruusunen, M.; Gusella, M.; Zhou, G.H.; Puolanne, E. High post-mortem temperature combined with rapid glycolysis induces phosphorylase denaturation and produces pale and exudative characteristics in broiler Pectoralis major muscles. Meat Science 2011, 89, 181-188. https://doi.org/10.1016/j.meatsci.2011.04.015. [CrossRef]
- Soglia, F.; Petracci, M.; Ertbjerg, P. Novel DNPH-based method for determination of protein carbonylation in muscle and meat. Food Chemistry 2016, 197, 670-675. https://doi.org/10.1016/j.foodchem.2015.11.038. [CrossRef]
- Jia, N.; Zhang, F.X.; Liu, Q.; Wang, L.T.; Lin, S.W.; Liu, D.Y. The beneficial effects of rutin on myofibrillar protein gel properties and related changes in protein conformation. Food Chemistry 2019, 301. https://doi.org/10.1016/j.foodchem.2019.125206. [CrossRef]
- Zhuang, X.; Wang, L.; Jiang, X.; Chen, Y.; Zhou, G. Insight into the mechanism of myofibrillar protein gel influenced by konjac glucomannan: Moisture stability and phase separation behavior. Food Chemistry 2021, 339, 127941. https://doi.org/10.1016/j.foodchem.2020.127941. [CrossRef]
- Xia, T.; Xu, Y.; Zhang, Y.; Xu, L.; Kong, Y.; Song, S.; Huang, M.; Bai, Y.; Luan, Y.; Han, M.; et al. Effect of oxidation on the process of thermal gelation of chicken breast myofibrillar protein. Food Chemistry 2022, 384, 132368. https://doi.org/10.1016/j.foodchem.2022.132368. [CrossRef]
- Salvador, P.; Toldra, M.; Saguer, E.; Carretero, C.; Pares, D. Microstructure-function relationships of heat-induced gels of porcine haemoglobin. Food Hydrocolloids 2009, 23, 1654-1659. https://doi.org/10.1016/j.foodhyd.2008.12.003. [CrossRef]
- Zhu, X.; Zhang, J.; Liu, S.; Gu, Y.; Yu, X.; Gao, F.; Wang, R. Relationship between Molecular Structure and Heat-Induced Gel Properties of Duck Myofibrillar Proteins Affected by the Addition of Pea Protein Isolate. Foods 2022, 11, 1040, https://doi.org/10.3390/foods11071040. [CrossRef]
- Xia, T.L.; Cao, Y.Y.; Chen, X.; Zhang, Y.L.; Xue, X.W.; Han, M.Y.; Li, L.; Zhou, G.H.; Xu, X.L. Effects of chicken myofibrillar protein concentration on protein oxidation and water holding capacity of its heat-induced gels. Journal of Food Measurement and Characterization 2018, 12, 2302-2312. https://doi.org/10.1007/s11694-018-9847-8. [CrossRef]
Figure 1.
Effect of AAPH-mediated oxidation (0, 1, 3, 5 and 10 mmol/L) on carbonyl content of duck myofibrillar proteins.
Figure 1.
Effect of AAPH-mediated oxidation (0, 1, 3, 5 and 10 mmol/L) on carbonyl content of duck myofibrillar proteins.
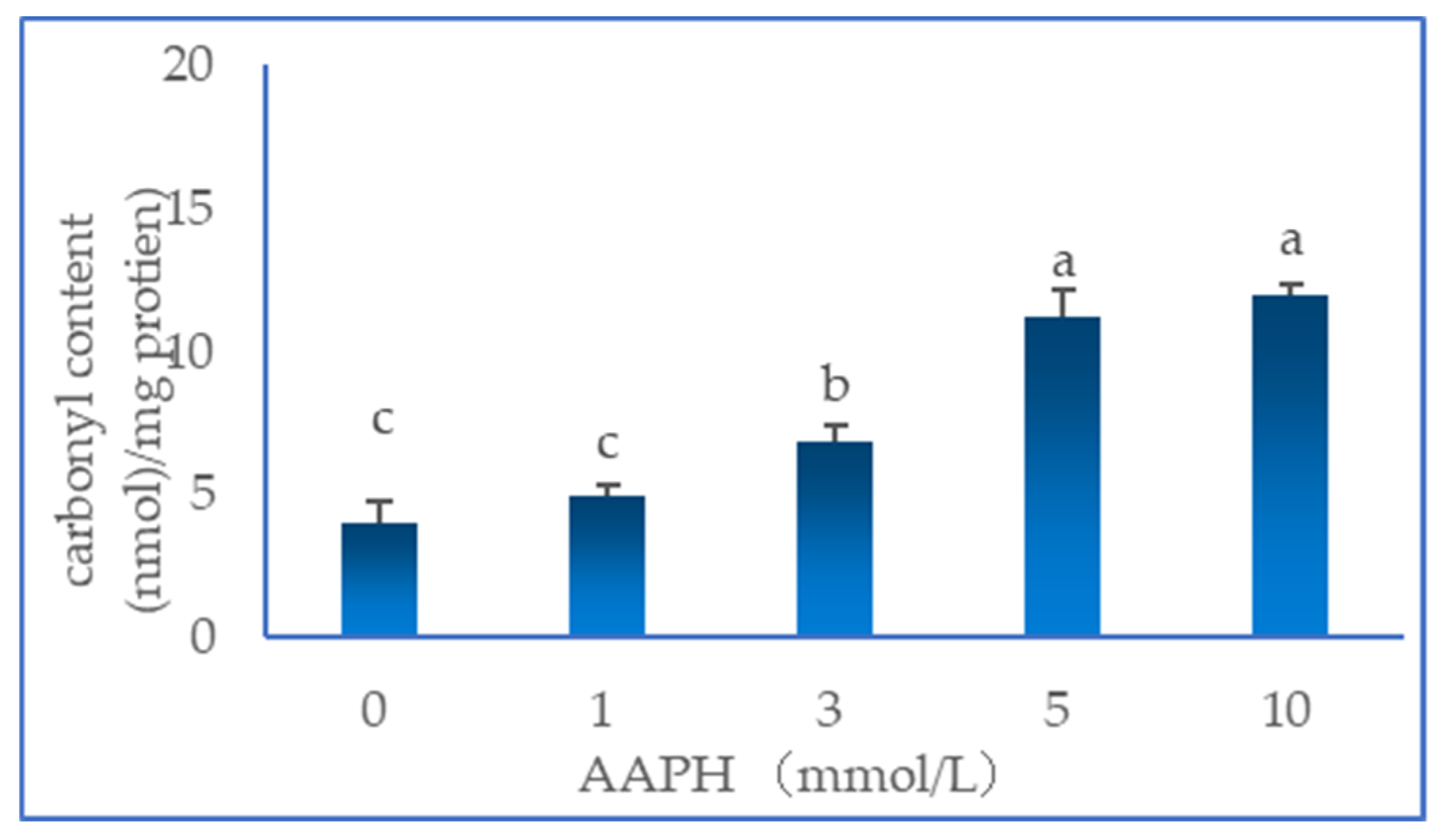
Figure 2.
Results of gel electrophoresis (SDS-PAGE) of duck myofibrillar proteins at different concentrations of AAPH (0, 1, 3, 5 and 10 mmol/L). Note: 0 is marker; 1-5 are without DTT and AAPH concentrations are 0, 1, 3, 5 and 10 mmol/L; 6-10 are with DTT and AAPH concentrations are 0, 1, 3, 5 and 10 mmol/L. α-Actinin:α-Actin, MHC: myosin heavy chain, actin: actin, MLC: myosin light chain.
Figure 2.
Results of gel electrophoresis (SDS-PAGE) of duck myofibrillar proteins at different concentrations of AAPH (0, 1, 3, 5 and 10 mmol/L). Note: 0 is marker; 1-5 are without DTT and AAPH concentrations are 0, 1, 3, 5 and 10 mmol/L; 6-10 are with DTT and AAPH concentrations are 0, 1, 3, 5 and 10 mmol/L. α-Actinin:α-Actin, MHC: myosin heavy chain, actin: actin, MLC: myosin light chain.
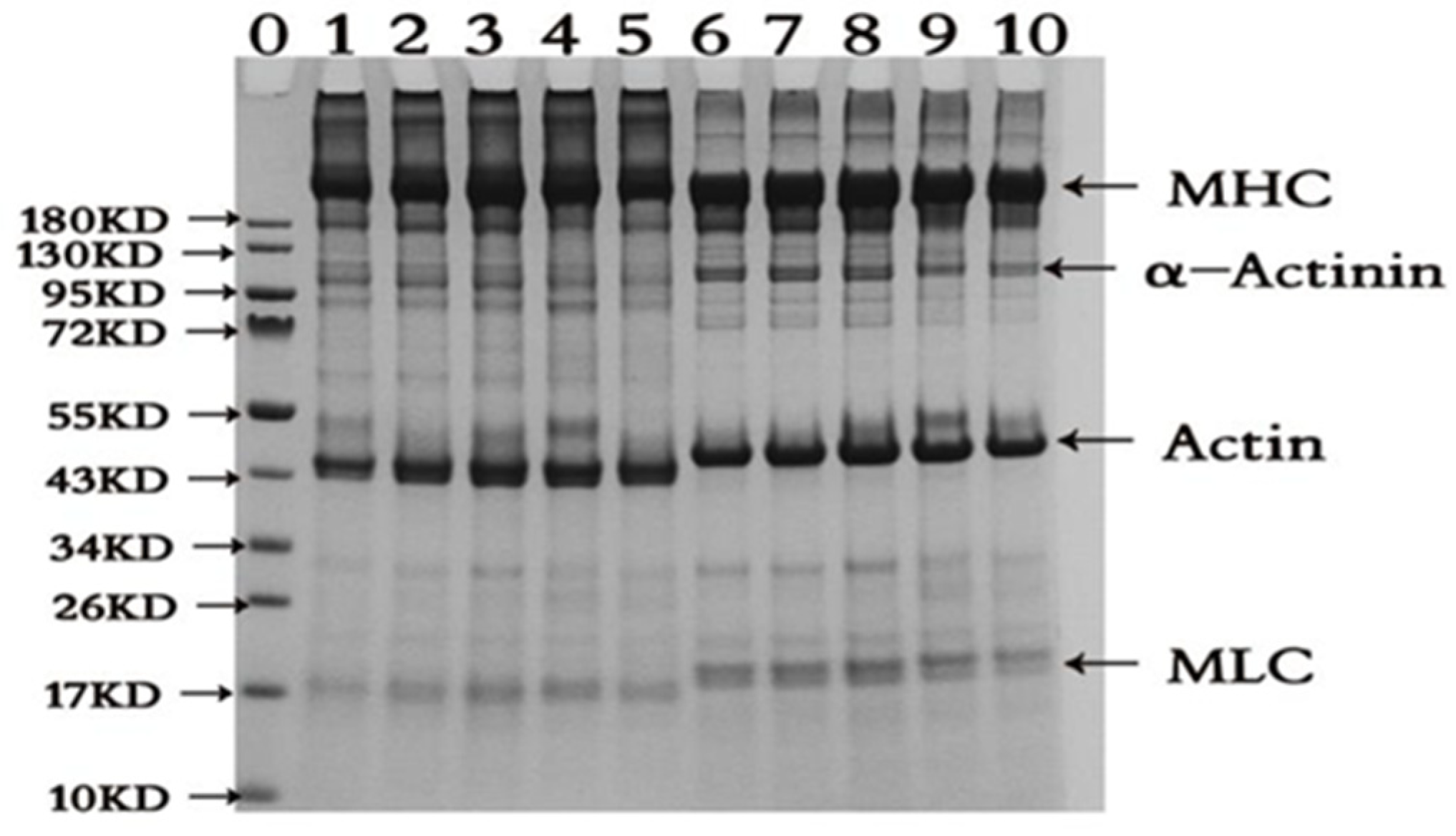
Figure 3.
Change in the storage modulus (G') of duck myofibrillar proteins at different concentrations of AAPH (0, 1, 3, 5 and 10 mmol/L).
Figure 3.
Change in the storage modulus (G') of duck myofibrillar proteins at different concentrations of AAPH (0, 1, 3, 5 and 10 mmol/L).
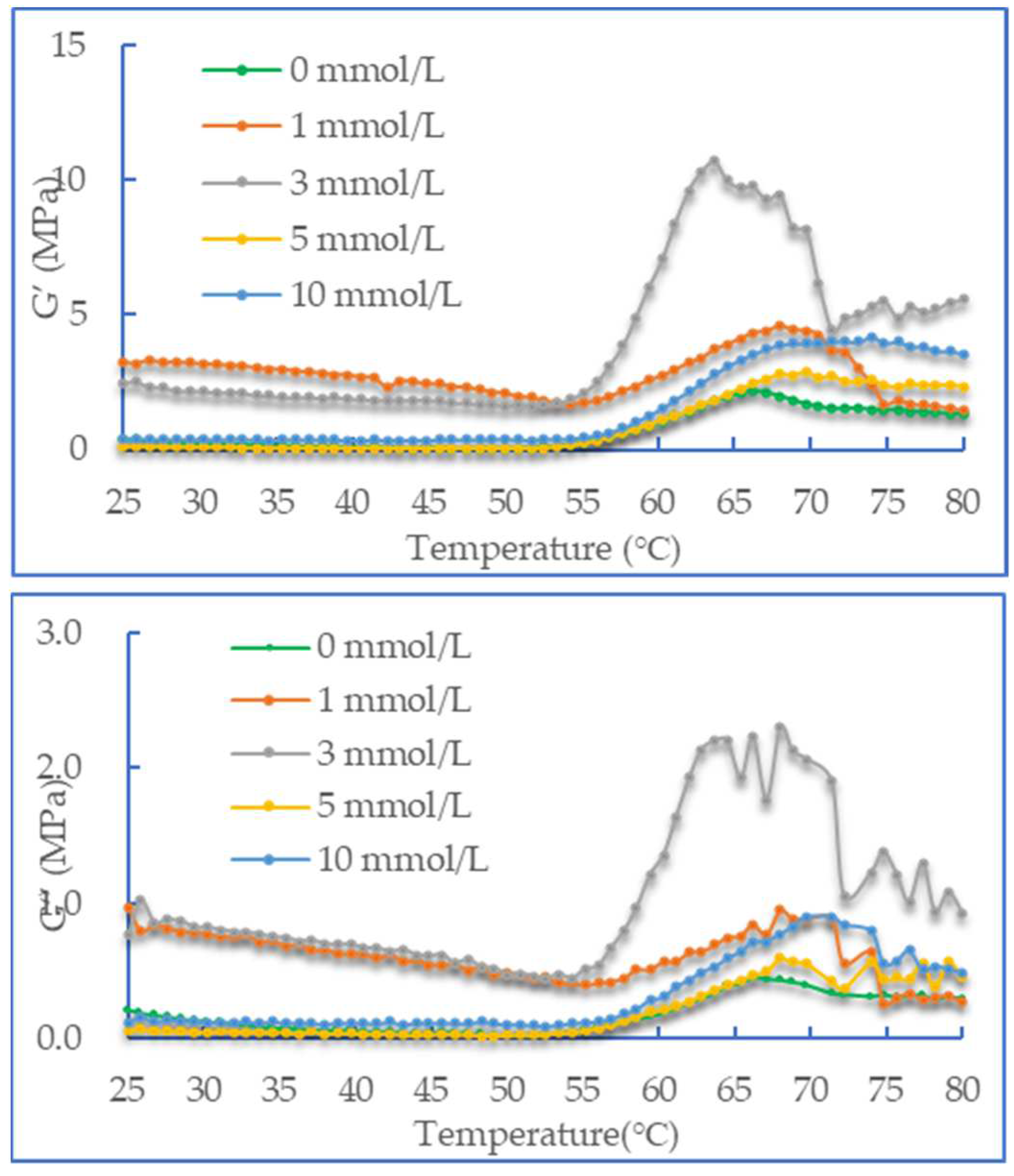
Figure 4.
Effect of AAPH-mediated oxidation (0, 1, 3, 5 and 10 mmol/L) on gel strength of duck myofibrillar proteins.
Figure 4.
Effect of AAPH-mediated oxidation (0, 1, 3, 5 and 10 mmol/L) on gel strength of duck myofibrillar proteins.
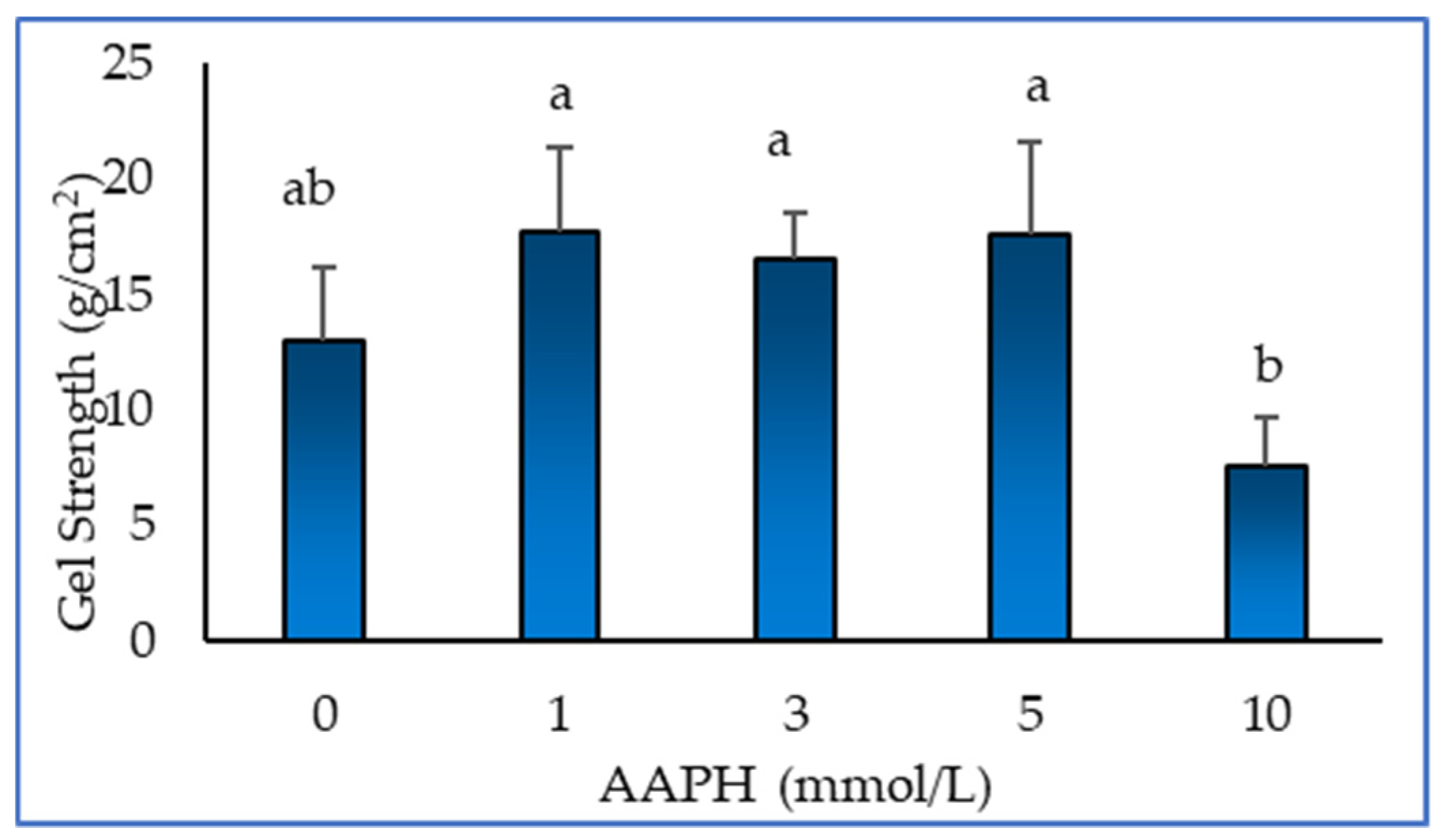
Figure 5.
Effect of AAPH-mediated oxidation (0, 1, 3, 5 and 10 mmol/L) on gel whiteness (a) and water holding capacity (b) of duck myofibrillar protein gels.
Figure 5.
Effect of AAPH-mediated oxidation (0, 1, 3, 5 and 10 mmol/L) on gel whiteness (a) and water holding capacity (b) of duck myofibrillar protein gels.
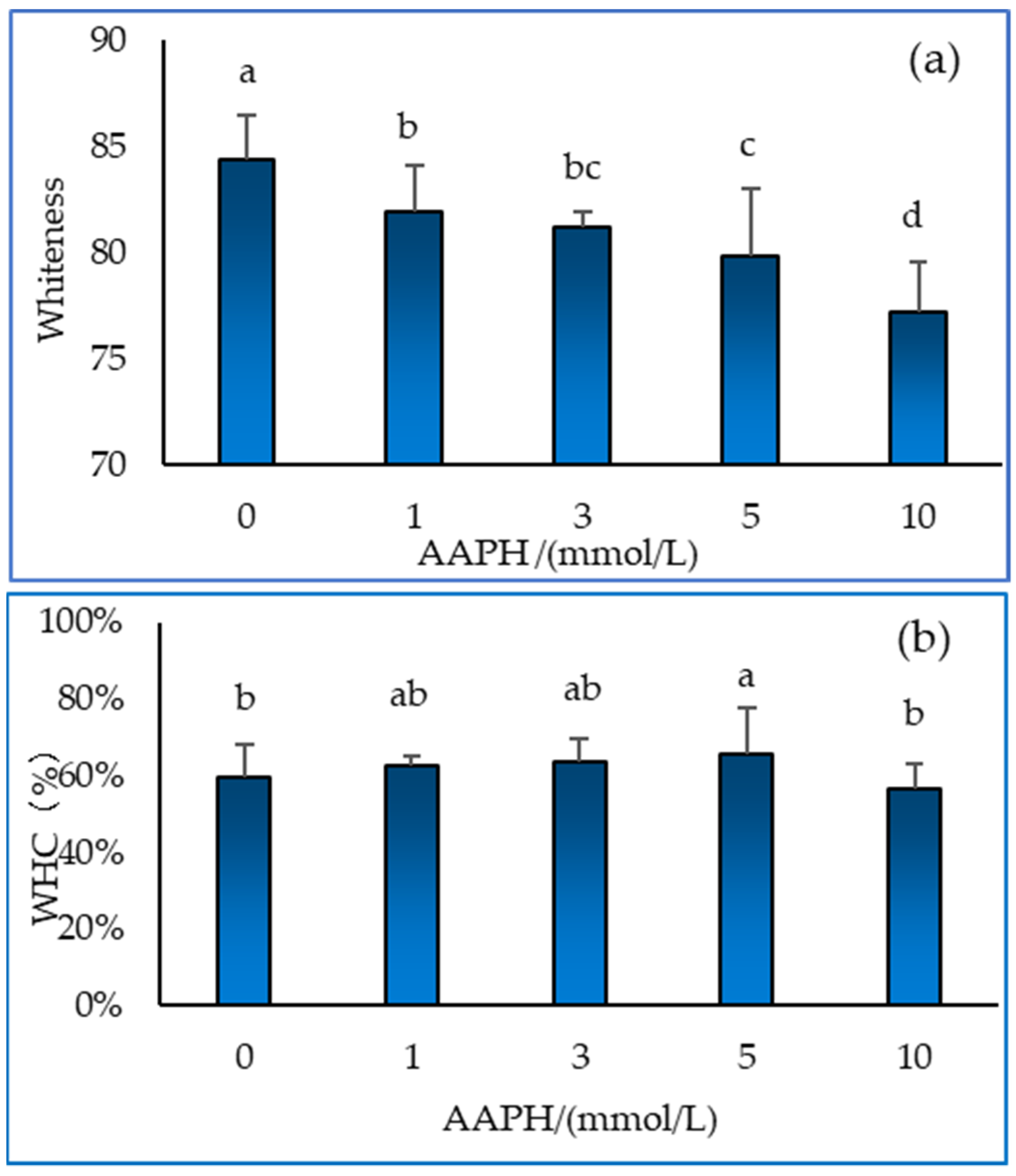
Figure 6.
NMR characterisation of duck myofibrillar protein gels under the influence of different concentrations of AAPH (0, 1, 3, 5 and 10 mmol/L).
Figure 6.
NMR characterisation of duck myofibrillar protein gels under the influence of different concentrations of AAPH (0, 1, 3, 5 and 10 mmol/L).
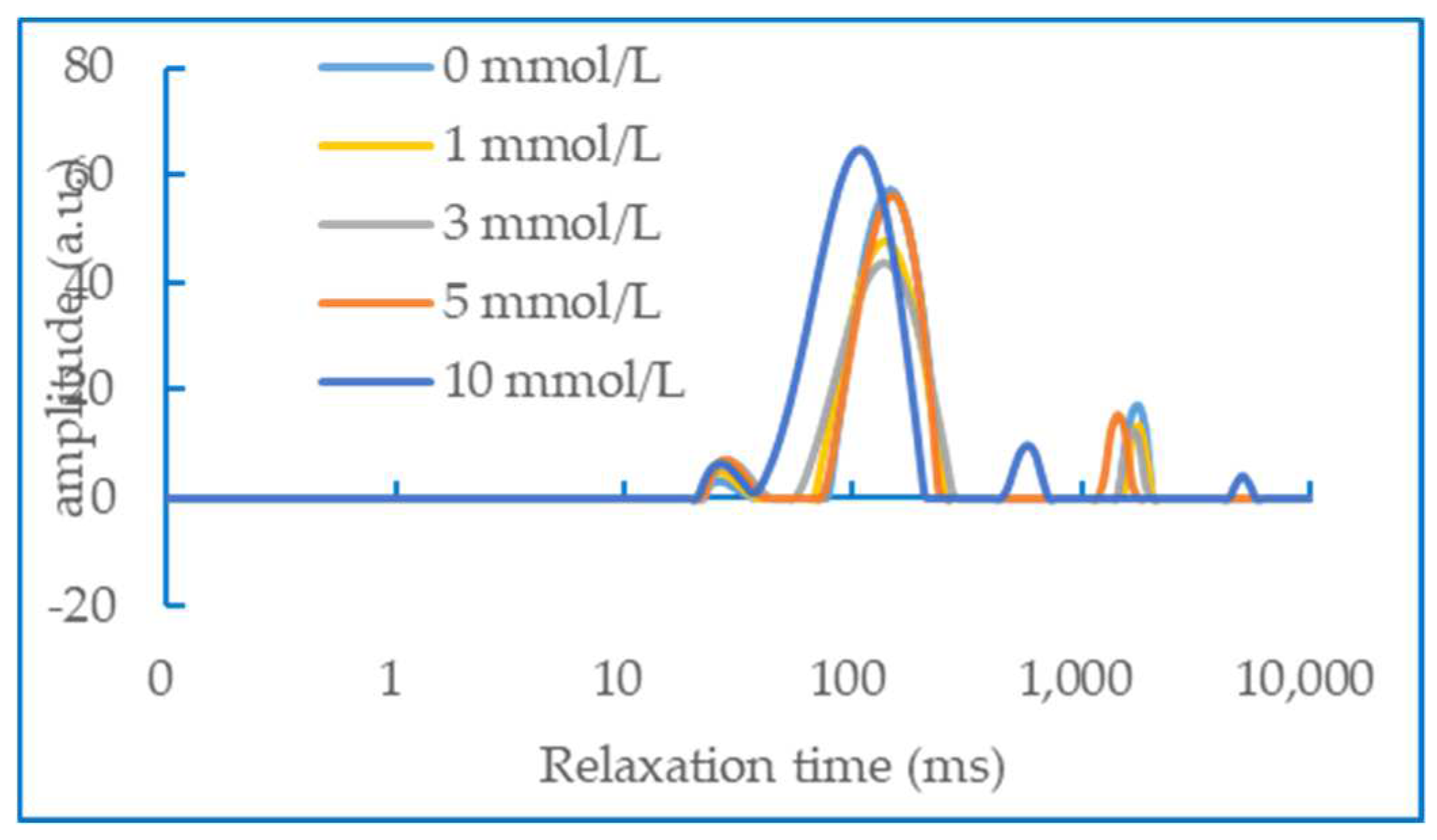
Figure 7.
Gel microstructure analysis of duck myofibrillar proteins treated with AAPH concentrations of 0 mmol/L (a), 1 mmol/L (b), 3 mmol/L (c), 5 mmol/L (d), and 10 mmol/L (e).
Figure 7.
Gel microstructure analysis of duck myofibrillar proteins treated with AAPH concentrations of 0 mmol/L (a), 1 mmol/L (b), 3 mmol/L (c), 5 mmol/L (d), and 10 mmol/L (e).


Table 1.
Table 1 Preparation of myofibrillar proteins oxidation solution.
Table 1.
Table 1 Preparation of myofibrillar proteins oxidation solution.
| AAPH concentraton (mmol/L) | 0 | 1 | 3 | 5 | 10 |
|---|---|---|---|---|---|
| DMPs (mL) | 6 | ||||
| AAPH Stock solution (mL) | 0 | 0.2 | 0.6 | 1 | 2 |
| PBS (mL) | 4 | 3.8 | 3.4 | 3 | 2 |
Note:AAPH Stock solution concentration is 50 mmol/L.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



