Submitted:
22 August 2023
Posted:
23 August 2023
You are already at the latest version
Abstract
Abstract
Plant uptake and assimilation of essential nitrogen compounds from the soil are mainly in the form of nitrate and ammonium ions. Plant nitrate transporters (NRTs) play a crucial role in nitrate uptake from the soil into the roots. NRT structural analysis reveals the mechanisms by which these transporters function. Similarly, plant ammonium transporters (AMTs) mediate the uptake of ammonium ions, and their structural analysis has provided valuable information on their mechanisms. After nitrogen uptake by plant roots, the nitrogen metabolism pathway involves the incorporation of nitrogen into organic compounds. Glutamine synthetase (GS) and glutamate synthase (GoGAT) are the master players in this pathway. They work together to convert ammonium ions into glutamine and glutamate, respectively. Different isoforms of GS and GoGAT exist, enabling plants to fine-tune nitrogen metabolism based on environmental cues. Under severe abiotic stress conditions, nitric oxide (NO) has been found to enhance plant survival under drought. Furthermore, the interaction between salinity stress and nitrogen availability in plants has been studied, with NO identified as a potential mediator of responses to salt stress. On the other hand, excessive use of nitrate fertilizers can lead to health and environmental issues. Therefore, alternative strategies, such as establishing nitrogen fixation in plants through diazotrophic microbiota, have been explored to reduce reliance on synthetic fertilizers. Ultimately, genomics can identify new genes related to nitrogen fixation, which could be harnessed to improve plant productivity. By deciphering the genetic basis of nitrogen-fixing traits, researchers aim to develop crops that efficiently utilize nitrogen from the environment.
Keywords:
Nitrogen uptake
; nitrogen metabolism
; plant stress response
; plant-microbe interaction
Introduction
Soil erosion, compaction, acidification, and contamination are significant factors causing soil degradation affecting 50% of agricultural land and food production [1,2,3]. Consequently, 75 billion tons of fertile soil are lost worldwide due to degradation annually [4]. Moreover, massive amounts of synthetic agro inputs to fertilize crops used by farmers harm environmental health [5,6]. In this sense, applying nitrogen fertilizers has increased sharply by 7.4 times compared with crop productivity, which only increased by 2.4 times, indicating that crops have reduced their ability to use N efficiently [7], increasing food insecurity [8,9]. Therefore, innovative and sustainable strategies in the agro-biotechnological sector have received more attention from botanical scientists worldwide over the last century to solve food security challenges and develop environment-friendly products [10]. Throughout time, microbial diversity and biomass have been assessed as partners in the plant-microbe association in maintaining soil multifunctionality [11].
Nitrogen uptake
Nitrogen (N2) is an essential macronutrient for plants, and it can be acquired from the soil in various forms. The most common inorganic forms are nitrate (NO3-) and ammonium (NH4+), which are transported across the root plasma membrane by different families of transporters. Four families of transporters mediate nitrate uptake: NRT1, NRT2, CLC-1, and SLAC1/SLAH. These transporters have distinct characteristics in affinity, capacity, regulation, and localization [12]. Ammonium uptake is facilitated by AMTs, high-affinity transporters expressed mainly in the root hairs and epidermis [13]. Ammonium is the dominant form of N in flooded or acidic soils, and AMT-mediated acquisition is crucial for the N demand of plants growing in such conditions [14]. Besides inorganic N, plants can also absorb organic N in the form of amino acids (AAs), which are abundant in soils that receive organic amendments such as manure or compost [15]. Several AAs transporters have been identified in plant roots, including AAP1, AAP5, ProT2, LHT1, and LHT6 [16,17]. These transporters have different substrate specificities and expression patterns, contributing to the uptake of a range of amino acids from the soil solution [18].
Plant nitrate transporters (NRTs)
Plants acquire nitrate (NO3-) from the soil through various transporters that belong to different families [19]. The NRT1 and NRT2 families are the primary transporters involved in NO3- uptake, and they have different roles in plant growth and seed development. The NRT1 family is a large and diverse group of transporters in the NPF family. In Arabidopsis, there are 53 NRT1 genes, while 93 in rice [20]. These genes can be classified into 8–10 subfamilies; NRT1.1 and NRT1.2 are responsible for NO3- root uptake. The NRT1 family can transport NO3- and other substrates such as hormones, nitrite, amino acids, peptides, chloride, glucosinolates, and jasmonate-isoleucine [21]. The NRT2 family is a smaller, more specific group of transporters expressed mainly under low NO3- conditions. In Arabidopsis, four NRT2 transporters (NRT2.1, NRT2.2, NRT2.4, and NRT2.5) function in nitrate influx and account for 95% of nitrate uptake under low NO3- concentrations [22]. However, NRT2.1 and NRT2.2 are significant members of the NRT2 family for nitrate uptake. The NRT2 family has different spatial and temporal expression patterns, and they transport NO3- from different sources to different tissues. For example, NRT2.4 and NRT2.5 absorb NO3- from the soil root hairs, while NRT2.1 and NRT2.2 transport it from the apoplast to the cortex and endodermis cells. Moreover, NRT2.5 is induced by long-term starvation and enhances NO3- uptake from both shoots and roots of mature plants [22,23]. In addition to the NRT2 family, another transporter family called NRT3 is also involved in NO3- transport in plants. The NRT3 family consists of two members: NRT3.1 and NRT3.2. These transporters form a complex with NRT2 transporters and regulate their activity and stability [24]. The NRT1.1 transceptor is a crucial component of NO3- signaling in plants, as it can sense the external NO3- concentration and modulate the root growth accordingly [25]. The activity and specificity of NRT1.1 are regulated by two CIPKs, CIPK8 and CIPK23, which interact with NRT1.1 and mediate low- and high-affinity responses, respectively [26]. At high NO3- concentrations, NRT1.1 activates the expression of ANR1, a transcription factor that promotes lateral root growth and proliferation [27]. At low NO3- concentrations, NRT1.1 inhibits lateral root development by controlling auxin levels and meristem activation [28]. To transmit the NO3- signals from NRT1.1 to the nucleus, calcium acts as a secondary messenger that modulates the expression of NO3- responsive genes. Three calcium-dependent protein kinases (CPK10, CPK30, and CPK32) and their partner CBL are involved in this process [29]. After being sensed and transported by NRT1.1, NO3- is assimilated into amino acids in different plant tissues. Several other NRT transporters are involved in the loading and unloading NO3- from the xylem and phloem, which affects the distribution and availability of NO3– within the plant. For example, NRT1.5, NRT1.8, and NRT1.9 are expressed in the xylem and phloem, and they function in the influx/efflux, removal, and loading of NO3- from/to the root vascular tissues, respectively [29,30]. Several studies have shown that overexpressing NRTs in different plant species can improve nitrogen use efficiency (NUE), which is the ratio of plant biomass or yield to nitrogen input. However, the effects of NRT overexpression may depend on the tissue specificity, nitrate concentration, and interaction with other genes. For instance, only NRT1 transporters have been successfully overexpressed in both leaves and roots to enhance NUE, while the role of NRT2 transporters in this process is still unclear. Recently, a chimeric NRT1 transporter, AtNC4N, was overexpressed in the phloem of old leaves in Arabidopsis, rice, and tobacco, and it increased nitrate uptake and NUE under low nitrate conditions [31]. Another NRT1 transporter, OsNRT1.1A (OsNPF6.3), was found to improve NUE, flowering time, yield, and maturity in rice [32]. Several other NRTs, such as OsNRT1.1B (OsNPF6.3B), OsNRT2.1, OsNRT2.3a/b, OsPTR9, OsAMT1.1, and qNGR9, have also been reported to enhance NUE under high or low nitrate levels in rice [32,33,34,35].
Moreover, some genes that regulate or interact with NRTs have been identified as potential targets for improving NUE in plants. For example, the transcription factor OsNAC42 activates the expression of several NRTs and increases nitrate assimilation and NUE in rice. The nitrate reductase gene OsNR2 converts nitrate to ammonium, affecting the expression of NRTs and other nitrogen-related genes in rice. The nitrate transporter OsNPF4.5 is regulated by a microRNA (miR827) and modulates nitrate distribution and remobilization in rice. These studies demonstrate the complexity and diversity of nitrate transport and signaling in plants and provide valuable insights for improving NUE and crop production [36].
NRT Structural analysis
The nitrate transporter system includes three distinct genes: NRT1, NRT2, and NRT3; they have different structural conformations (Figure 1A-C). As a result, Pfam database collects these genes under distinct families: PF00854 for NRT1, PF07690 for NRT2, and PF16974 for NRT3 [37]. PF00854 refers to the proton-dependent oligopeptide transporter (POT) family (also known as the peptide transporter (PTR) family) and is a class of energy-dependent transporters discovered in organisms ranging from bacteria to humans [38]. However, some family members are nitrate permeases, while others are in histidine transport [39]. The NRT1 transporter adopts a standard major facilitator superfamily (MFS) fold, which is described by 12 TMHs with a pseudo 2-fold axis connecting the N-terminal (TMH1–6) and C-terminal (TMH7–12) domains [40].
NRT2 members belong to the PF07690 (MSF) family, representing the largest family of secondary transporters with members ranging from Archaea to Homo sapiens. MFS proteins target a diverse range of substrates in both directions across the membrane, including ions, carbohydrates, lipids, amino acids and peptides, nucleosides, and other small molecules, in many cases catalyzing active transport by converting the energy stored in a proton electrochemical gradient into a concentration gradient of the substrate [41]. NRT3 candidates lie under the PF16974 family, which contains a C-terminal transmembrane domain required for high-affinity nitrate absorption [42,43]. NRT3 is implicated in the inhibition of lateral root initiation in the presence of high sucrose-to-nitrogen ratios in the medium. As a result, the NRT2 and NRT3 component systems are believed to be implicated in the signaling pathway that integrates nutritional inputs for the regulation of lateral root architecture [44]. In Arabidopsis, the functional unit of the high-affinity nitrate influx complex is most likely a tetramer composed of two NRT2 and NRT3 subunits each [45].
Plant ammonium transporters (AMTs)
In most natural and agricultural environments, ammonium and nitrate are the two principal inorganic nitrogen sources absorbed by higher plant roots [46,47]. Because ammonium absorption requires less energy than nitrate assimilation, ammonium is the preferred form of nitrogen uptake when plants are nitrogen deficient [48,49]. Although ammonium uptake is more effective, too much ammonium in the plant can be hazardous [50,51,52]; thus, the ammonium uptake system in plants receives special attention [53,54,55]. Recent research indicates that AMTs also play a role in a variety of other physiological processes, such as transporting ammonium from symbiotic fungi to plants [56,57], delivering ammonium from roots to shoots [58], transmitting ammonium in leaves and reproductive organs [59,60,61], and encouraging resistance to plant diseases through ammonium transport [62,63]. Aside from being a transporter, many AMTs are essential for root growth in the presence of ammonium [54,64,65,66]. To avoid the adverse effects of insufficient or excessive ammonium nitrogen intake on plant growth and development, AMTs activities are fine-tuned not only at the transcriptional level through the involvement of at least four transcription factors [67,68,69,70] but also at the protein level through phosphorylation [71,72], pH [73], and endocytosis [74].
AMTs Structural analysis
The ammonium transporter domain (PF00909) comprises two structural copies of five helices each, plus one extra C-terminal helix (Figure 1D). It has been defined as an 11-times-spanning membrane channel. An AMT is composed of 11–12 transmembrane spots, with feature sequences "D (F Y W S) A G (G S C) X2 (L I V) (E H) X2 (G A S) (G A) X2 (G A S) (L F)" at transmembrane spot 5 and "D D X (L I V M F C) (E D G A) (L I V AC) X3 H (G A L I V) X2 (G S) X (L I V A W) G” at transmembrane region 10 [54]. AMTs can be divided into two subtypes: AMT1 and AMT2 or methylammonium permeases (MEP) [54,58,75].
Nitrogen metabolism pathway
Nitrogen is among the most widely distributed elements in the lithosphere, atmosphere, hydrosphere, and biosphere [76]. The lithosphere contains 94% of all nitrogen on Earth (e.g., NO3−, NO2−, NH4+), with the remaining 6% in the atmosphere (e.g., NO, N2O, N2) and a trace (0.006%) in the hydrosphere and biosphere (e.g., NO3−, NO2−, NH4+). Nitrogen is the fourth most prevalent element in the biosphere, after oxygen, carbon, and hydrogen, and is an essential component of total biomass [77,78]. The relative quantity of nitrogen in the biosphere reflects the relevance of nitrogen to living organisms. Nitrogen may produce a wide range of stable chemical and inorganic compounds with oxidation states ranging from 3 to +5. Nitrogen has two forms, organic and inorganic, such as nitrate and nitric oxide, which are paramount as a keystone of the global nitrogen cycle and used as essential signaling molecules. In addition, organic nitrogen, such as amino acids, hormones, and nucleic acids, is vital to life on Earth [79,80,81,82]. The global nitrogen cycle is a web of chemical reactions that recycle nitrogen between the atmosphere, biosphere, hydrosphere, and lithosphere. The global nitrogen cycle is dominated by inorganic nitrogen molecules (i.e., nitrate (NO3), nitrite (NO2), nitrous oxide (N2O), nitric oxide (NO), dinitrogen (N2), and ammonia (NH3) [79,83,84,85] (Figure 2).
Nitrate is the most oxidized form of nitrogen, while NH3 is the most reduced form. Many microorganisms, mainly microbes, drive the global nitrogen cycle by metabolizing nitrogen through a variety of redox processes for energy transduction, detoxification, or assimilation via various sub-pathways such as Nitrogen fixation (KEGG;M00175), Assimilatory nitrate reduction (KEGG;M00531), Dissimilatory nitrate reduction (KEGG;M00530), Denitrification (KEGG;M00529), Nitrification (KEGG;M00528), Complete nitrification, complete ammonia oxidation (comammox) (KEGG;M00804) and Anammox (KEGG;M00973) (Figure 2).
Moreover, each sub-pathway from these previous sub-pathways consists of a list of genes and their encoded enzymes that are involved in the various biogenic processes in the nitrogen cycle. The nitrogen fixation path consists of Nitrogenase molybdenum-iron protein alpha chain (NIFD), Vanadium-dependent nitrogenase alpha chain (VNFD), and Nitrogenase delta subunit (ANFG) genes [86,87,88], see (Figure 2) and Table 1.

Table 1.
List of genes and enzymes involved in the nitrogen cycle.
| No. | KEGG ID | Gene Symbol | Gene Name | EC number | Reference | |
| 1 | K04561 | NORB | Nitric oxide reductase subunit B | 1.7.2.5 | Heiss et al., 1989; Suzuki et al., 2006 | |
| 2 | K15877 | CYP55 | Fungal nitric oxide reductase | 1.7.1.14 | Kizawa et al., 1991; Zhang and Shoun, 2008 | |
| 3 | K00376 | NOSZ | Nitrous-oxide reductase | 1.7.2.4 | Coyleet al., 1985; Arai et al., 2003; Zumft and Kroneck, 2007 | |
| 4 | K02586 | NIFD | Nitrogenase molybdenum-iron protein alpha chain | 1.18.6.1 | Joerger et al., 1989; Fani et al., 2000 | |
| 5 | K22896 | VNFD | Vanadium-dependent nitrogenase alpha chain | 1.18.6.2 | Joerger et al., 1990 | |
| 6 | R05186 | NIFF | Nitrogenase | 1.19.6.1 | Deistung et al., 1985 | |
| 7 | K01455 | FORMAMIDASE | Formamidase | 3.5.1.49 | ||
| 8 | K20935 | HDH | Hydrazine dehydrogenase | 1.7.2.8 | Kartal et al., 2011 | |
| 9 | K20932 | K20932 | Hydrazine synthase subunit | 1.7.2.7 | 1.7.2.7 | |
| 10 | K01672 | CA | Carbonic anhydrase | 4.2.1.1 | Breton, 2001; Pastorekova et al., 2004 | |
| 11 | K01725 | CYNS | Cyanate lyase | 4.2.1.104 | Harano et al., 1997 | |
| 12 | K00368 | NIRK | Nitrite reductase (NO-forming) | 1.7.2.1 | Cantera and Stein, 2007 | |
| 13 | K02575 | NRT2, NARK, NRTP, NASA | MFS transporter, NNP family, nitrate/nitrite transporter | ------ | Noji et al., 1989 | |
| 14 | K15576 | NRTA, NRTB, NRTC, NASD | Nitrate/nitrite transport system substrate-binding protein | 7.3.2.4 | Maeda and Omata, 2009 | |
| 15 | K00370 | NARG, NARZ, NXRA | Nitrate reductase/nitrite oxidoreductase, alpha subunit | 1.7.5.11.7.99.- | Blasco et al., 1990; Sohaskey and Wayne, 2003 | |
| 16 | K10534 | NR | Nitrate reductase (NAD(P)H) | 1.7.1.1 1.7.1.2 1.7.1.3 | Okamoto and Marzluf, 1993 | |
| 17 | K00367 | NARB | Ferredoxin-nitrate reductase | 1.7.7.2 | Rubio et al., 1996 | |
| 18 | K02567 | NAPA | Nitrate reductase (cytochrome) | 1.9.6.1 | Stolz and Basu, 2002; Jepson et al., 2007 | |
| 19 | K17877 | NIT-6 | Nitrite reductase (NAD(P)H) | 1.7.1.4 | Exley et al., 1993 | |
| 20 | K00366 | NIRA | Ferredoxin-nitrite reductase | 1.7.7.1 | Takahashi et al., 2001 | |
| 21 | K00362 | NIRB | Nitrite reductase (NADH) large subunit | 1.7.1.15 | Harborne et al., 1992 | |
| 22 | K03385 | NRFA | Nitrite reductase (cytochrome c-552) | 1.7.2.2 | Einsle et al., 1999 | |
| 23 | K00459 | NCD2, NPD | Nitronate monooxygenase | 1.13.12.16 | Gadda and Francis, 2010 | |
| 24 | K19823 | NAO | Nitroalkane oxidase | 1.7.3.1 | Daubner et al., 2002 | |
Cont. Table 1.
List of genes and enzymes involved in the nitrogen cycle.
| No. | KEGG ID | Gene Symbol | Gene Name | EC number | Reference |
| 25 | R00143 | Hydroxylamine reductase (NADH) | 1.7.1.10 | ||
| 26 | K05601 | HCP | Hydroxylamine reductase | 1.7.99.1 | Wolfe et al., 2002 |
| 27 | K10535 | HAO | Hydroxylamine dehydrogenase | 1.7.2.6 | Hommes et al., 2001 |
| 28 | R10230 | Hydroxylamine oxidase (cytochrome) | 1.7.3.6 | Wehrfritz et al., 1996 | |
| 29 | K10944 | PMOA-AMOA | Methane/ammonia monooxygenase subunit A | 1.14.18.3 1.14.99.39 | Norton et al., 2002 |
| 30 | K00926 | ARCC | Carbamate kinase | 2.7.2.2 | Durbecq et al., 1997 |
| 31 | K01948 | CPS1 | Carbamoyl-phosphate synthase (ammonia) | 6.3.4.16 | Haraguchi et al., 1991 |
| 32 | K00260 | GUDB, ROCG | Glutamate dehydrogenase | 1.4.1.2 | Belitsky and Sonenshein, 1998 |
| 33 | K00261 | GLUD1_2, GDHA | Glutamate dehydrogenase (NAD(P)+) | 1.4.1.3 | Frigerio et al., 2008 |
| 34 | K00262 | E1.4.1.4, GDHA | Glutamate dehydrogenase (NADP+) | 1.4.1.4 | Riba et al., 1988 |
| 35 | K01915 | GLNA, GLUL | Glutamine synthetase | 6.3.1.2 | Newsholme et al., 2003 |
| 36 | K01501 | E3.5.5.1 | Nitrilase | 3.5.5.1 | Zhu et al., 2008 |
| 37 | K00265 | GLTB | Glutamate synthase (NADPH) large chain | 1.4.1.13 | Sonawane and Röhm, 2004 |
| 38 | K00264 | GLT1 | Glutamate synthase (NADH) | 1.4.1.14 | Lancien et al., 2002 |
| 39 | K00284 | GLU, GLTS | Glutamate synthase (ferredoxin) | 1.4.7.1 | Coschigano et al., 1998 |
| 40 | K00372 | NASC, NASA | Assimilatory nitrate reductase catalytic subunit | 1.7.99.- | Ogawa et al., 1995; Lin et al., 1993 |
| 41 | K26139 | NASD, NASB | Nitrite reductase [NAD(P)H] large subunit | 1.7.1.4 | Ogawa et al., 1995 |
| 42 | K15864 | NIRS | Nitrite reductase (NO-forming) / hydroxylamine reductase | 1.7.2.1 1.7.99.1 | Rees et al., 1997; Hole et al., 1996 |
| 43 | K00531 | ANFG | Nitrogenase delta subunit | 1.18.6.1 | Joerger et al., 1989 |
| 44 | K04561 | NORB | Nitric oxide reductase subunit B | 1.7.2.5 | Heiss et al., 1989; Suzuki et al., 2006 |
In this sub-pathway these previous genes were used to convert N2 to NH3 by oxidoreductases reaction, and this NH3 molecules are widely used in different vital pathways for living organisms such as, Arginine biosynthesis (KEGG;map00220), Purine metabolism (KEGG;map00230), Alanine, aspartate and glutamate metabolism (KEGG;map00250), Glycine, serine and threonine metabolism (KEGG;map00260), Cyanoamino acid metabolism (KEGG;map00460), Glyoxylate and dicarboxylate metabolism (KEGG;map00630), Lipoic acid metabolism (KEGG;map00785), Nitrogen metabolism (KEGG;map00910), Metabolic pathways (KEGG;map01100), Biosynthesis of secondary metabolites (KEGG;map01110), Microbial metabolism in diverse environments (KEGG;map01120), Carbon metabolism (KEGG;map01200), Biosynthesis of amino acids (KEGG;map01230), Proximal tubule bicarbonate reclamation (KEGG;map04964), Collecting duct acid secretion (KEGG;map04966), Protein digestion and absorption (KEGG;map04974), Vibrio cholerae infection (KEGG;map05110) and Epithelial cell signaling in Helicobacter pylori infection (KEGG;map05120) [86,87,88].
In addition, the Anammox path consists of Hydrazine dehydrogenase (HDH), Hydrazine synthase subunit (K20932), and Nitrite reductase (NO-forming) (NIRK) genes, and these previous genes can convert NO2 to N2 or NH3 using nitrite reductase enzyme [89,90] (Figure 2) and Table 1. In that context, the Denitrification path consists of four genes Nitrous-oxide reductase (NOSZ), Nitrite reductase (NO-forming) (NIRK), Nitrate reductase (cytochrome) (NAPA), and nitric oxide reductase subunit B (NORB) [91,92,93]. In this sub-pathway, the Denitrification genes convert NO3 to N2 through NO2, NO, and NO2. Furthermore, Assimilatory and dissimilatory nitrate reduction paths have different genes, such as Nitrate reductase/nitrite oxidoreductase, alpha subunit (NARG, NARZ, and NXRA), Nitrate reductase (cytochrome) (NAPA), Nitrite reductase (NADH) large subunit (NIRB), Nitrite reductase (cytochrome C-552) (NRFA), Ferredoxin-nitrate reductase (NARB), Nitrate reductase (NAD-(P)-H) (NR), Assimilatory nitrate reductase catalytic subunit (NASC and NASA), Nitrite reductase (NAD-(P)-H) (NIT-6), Ferredoxin-nitrite reductase (NIRA) and Nitrite reductase [NAD(P)H] large subunit (NASD and NASB) that can convert NO2 to NH3 [94,95,96,97].
Function and regulation of glutamine synthetase and glutamate synthase in nitrogen metabolism
In various biological systems, the interconversion of glutamate and glutamine plays a central role in nitrogen assimilation, transport, and recycling. Glutamine synthetase (GS) and glutamate synthase or glutamine 2-oxoglutarate aminotransferase (GOGAT) are vital enzymes orchestrating these interconversions, ensuring optimal nitrogen utilization, and maintaining nitrogen homeostasis.
The process of nitrogen uptake by plants from the soil for growth has been well studied. Plants acquire nitrogen in either ammonium or nitrate. Nitrate is converted into ammonium, then combined with glutamate to be assimilated into glutamine [98]. The principal site of ammonium assimilation is the roots, where the enzyme GS facilitates this conversion [99,100]. Following this, one of the amino groups from glutamine is transferred to 2-oxoglutarate (2-OG) to synthesize glutamate. This reaction is catalyzed by GOGAT (Figure 3). The coordinated action of GS and GOGAT is responsible for these interconnected processes. The GS/GOGAT cycle is the plants' central pathway for ammonium assimilation [101,102,103]. Experimental studies involving reverse genetic analysis have demonstrated that the absence of genes encoding these ammonium-responsive GS/GOGAT isoenzymes reduces ammonium assimilation, particularly in the roots, resulting in impaired growth [104,105,106,107]. These findings suggest that the ammonium-responsive forms of GS/GOGAT have a central role in the primary assimilation of ammonium within the roots [108]. As the expression of these isoenzyme-encoding genes increases with ammonium availability, it is plausible to infer that plants possess a regulatory transcriptional network that modulates their gene expression in response to ammonium availability [105].
Plant GS (EC6.3.1.2), a highly conserved enzyme, catalyzes glutamine synthesis from glutamate and ammonium ions, conserving the nitrogen atom in an organic molecule. This conversion is fueled by the energy released through the hydrolysis of ATP to ADP, making GS a crucial component of nitrogen metabolism across different life forms [109], in addition, the GOGAT is the critical enzyme in synthesizing glutamate, [101]. This reaction involves transferring the amide amino group of glutamine to 2-oxoglutarate through a reductant-driven process, yielding two glutamate molecules [110]. Within the realm of plants, this enzyme exists in two distinct variants: one employs reduced ferredoxin (Fd) as the electron contributor (EC 1.4.7.1), while the other employs NADH as the electron contributor (EC 1.4.1.14). The Fd-dependent form of the enzyme is notably abundant within the chloroplasts of photosynthetic tissues. In these regions, it utilizes light energy directly to serve as a reductant source.
Conversely, the NADH-dependent variant, also present within plastids, is primarily concentrated in non-photosynthesizing cells. The reductant is sourced from the pentose phosphate pathway [111]. Notably, the expression of the Fd- and NADH-dependent forms of glutamate synthase seems to exhibit dissimilar patterns across distinct plant tissues. It has been suggested that in most plants, two genes encode each form of glutamate synthase [110].
Isoforms of GS and GoGAT
According to subcellular distribution patterns, investigations into plant GS enzymes have identified two main categories. The initial group, designated as GS1, is primarily situated within the confines of the cytoplasm. Conversely, the second group, GS2, is predominantly found within plastid structures [112,113]. Various multigenic families are responsible for encoding multiple isoforms of GS1, whereas the plastidial GS2 arises from a limited number of nuclear genes [114]. Typically, GS2 is linked to the process of (re)assimilation of NH4+ in leaves, whereas GS1 is associated with recycling nitrogen within the plant. However, the relative activity levels exhibited by GS1 and GS2 are subject to variation based on species, specific plant organs, nitrogen sources, developmental stages, and prevailing environmental conditions, including abiotic stress factors. This variability underscores the intricate involvement of these isozymes, reflecting a multifaceted role [113,115]. In wheat research, a comprehensive exploration has unveiled four distinct variations of GS genes: GS1, GS2, GSr, and GSe [112]. Employing advanced high-performance liquid chromatography (HPLC) methods, [116] successfully separated GS and discovered two subtypes in leaves: cytoplasmic subtype GS1 and chloroplast subtype GS2. Both subtypes' content and activity fluctuate in tandem with developmental processes. The subcellular localizations of GSr and GSe align with the cytoplasmic compartment [117]. In addition, the advancement of genome sequencing has unveiled various isoenzymes of GS/GOGAT present in plants. Among these, specific isoenzymes, such as GS1;2 and NADH-GOGAT1 from rice [118], as well as GLN1;2 and NADH-GOGAT (GLT1) from Arabidopsis [119,120], are expressed in plant roots in response to ammonium supply. These isoenzymes' expression at transcript and protein levels highlights their significance in ammonium assimilation.
Biological function of GS in response to abiotic stress
The pivotal function of GS within the nitrogen metabolism pathway of plants has been extensively investigated. Many research endeavors have underscored GS's capacity to regulate nitrogen levels within plants, exerting an indirect influence over diverse photosynthesis and physiological metabolic processes. Consequently, these regulatory actions have far-reaching implications on the trajectory of growth, development, crop yield, and overall quality of the produce. Nevertheless, the intricate diversity exhibited by GS isoforms implies a multifaceted role for GS in navigating through various dimensions of abiotic stress tolerance [113,121]; for instance, in rice plants exposed to salinity stress, GS1 increased in root at seedling stage and old leaves with no change in young leaves, while GS2 was down-regulated in old leaves and young leaves [122]. Under drought stress, GS1 and GS2 exhibited differential expression among different plant organs and species, with a prominent increase of GS1 in roots and leaves, while the total GS activity showed a decrease in the potato root [123,124]. A comparative proteomic analysis under drought stress conditions in wheat was studied [125]. They found that drought-tolerant wheat varieties exhibited a significant upregulation of GS2 expression, indicating its potential role in enhancing drought tolerance. Similar results have been obtained by [126], who explored GS activity in wheat leaves and observed that older leaves had lower GS activity than flag leaves in drought-tolerant and sensitive varieties under normal conditions. During drought stress, GS activity in sensitive varieties' flag leaves decreased compared to tolerant varieties. This suggested that GS activity could indicate drought stress sensitivity and tolerance, with elevated cytoplasmic-to-plastid GS ratio and premature senescence as markers. Exposure to heavy metals such as Cadmium, Molybdenum, copper, and Aluminum led to the down-regulation of GS and decreased its total activity in different plant species, including rice, soybean, tomato, and tea [127,128,129,130]
Nitrogen-induced resilience in plants against drought stress
Drought stress represents one of the most significant abiotic stresses that significantly damages crops worldwide, particularly in arid and semi-arid regions. Crops are constantly exposed to drought and temperature pressures during the growing season in arid and semiarid regions [131]. Plants adapt to drought via avoidance (stomata adjustment to reduce transpiration rate) and tolerance (antioxidants and osmolytes production) of drought [132] (Figure 4). The concentrations of NO3- and NH4+ have remarkable impacts on plants under drought conditions; the application of NH4+ alleviates the effect of drought on plant development, while NO3- has the opposite impact [13]. Nitrogen fertilizers might enhance the physiological responses to drought stress by elevating the concentrations of N and chlorophyll and promoting PSII photochemical activity [133]. It has been shown that adding N increases water use efficiency (WUE) by increasing plant dry mass and decreasing water loss [132].
Additionally, nitrogen may lessen the drought's inhibitory effects on photosynthesis and prevent C starvation [134]. On the other hand, low N availability can make plants more sensitive to dehydration and cause protein degradation, which lowers the amount of N-containing osmolytes like proline [132,134]. Nitrogen supply enhances osmotic adjustment and the activity of antioxidant enzymes via proline accumulation induction to mitigate the physiological harm from drought stress. However, the nitrogen effects depend on the ionic form of nitrogen supplied [135]. NH4+ is essential in promoting plant drought tolerance by increasing root numbers (surface area) and subsequently improving and facilitating water uptake. Also, decreased aerenchyma development may contribute to NH4+ promoted drought tolerance [13]. The influence of NO3- on drought responses is linked to NO3- transport/assimilation in plants. Numerous NO3- transport/assimilation-related genes, such as NRT2.5, GS, and GOGAT, are repressed in response to drought stress. Plants respond better to drought when genes involved in NO3- absorption or assimilation pathways are disrupted [13]. Drought stress and N deficiency have similar plant physiological responses. They reduce photosynthesis and use similar adaptive mechanisms such as osmolyte accumulation, longer roots, induction of antioxidative enzymes, reduction of photosynthesis and stomatal conductance, and induction of reactive oxygen species (ROS) [136]. Therefore, a high nitrogen concentration enhances root growth and photosynthetic traits, alleviates drought stress via stomatal conductance, and increases the photosynthetic rate [137].
Additionally, at high nitrogen supply, more nitrogen forms soluble nitrogenous molecules like free amino acids such as proline, which function as osmotica to balance the water status under abiotic stress [138]. Plant nutrient management is crucial to promote abiotic stress tolerance in cotton plants. The use of nitrogen fertilizer either in the soil or foliar spray can decrease the toxicity of ROS caused by abiotic stresses through the increase the peroxidase (POD), catalase (CAT), and superoxide dismutase (SOD) levels in plant cells [131,139]. These enzymes improve the photosynthetic rate, promoting stress tolerance by scavenging ROS and reducing photooxidation to preserve chloroplast membrane integrity [139,140,141]. Applying nitrogen fertilizers (N60 kg.ha-1) can increase the tolerance of drought stress in wheat plants combined with the highest yield [131].
Nitric oxide enhances plant survival under drought stress
Nitric oxide (NO) is a highly reactive, redox-active nitrogen species (RNS) formed in living cells under normal and stressful biotic and abiotic conditions. NO may function as a detoxifier and lessen adverse effects when a plant's ROS content reaches dangerous levels [142]. It was reported that the synthesis of NO increased in plant cells under drought stress conditions, and it has been suggested that this may help enhance plants' acclimation responses to cope with water limitation [143]. Drought stress enhanced NO synthesis in beat, pea, tobacco, cucumber, grapevine, and rice plants. Also, the activity of NO synthase (NOS)-like and NO release rate have increased under dehydration conditions [144]. NO has a vital role in respiratory function, called pathways of electron transport in mitochondria; it adjusts ROS and therefore activates defense strategies via promoted antioxidant formation in plants under various abiotic stresses [142]. NO plays a crucial role in mediating hormonal activities, interacting with ROS, and influencing gene expression as well as protein activity during sensing and signaling processes that enable plants to respond to and survive in environmental stress circumstances. Exogenously supply of NO increased drought tolerance in various plants such as maize, rice, and cucumber due to the increase of antioxidant defense, which acts to scavenge ROS, improve cellular membrane stability, and preserve photosynthesis as well as water status [144,145].
Drought induces the synthesis of abscisic acid (ABA), which regulates several critical plant processes, like seed germination and stomatal conductance, with the help of signaling molecules like NO and ROS [146]. NO plays an essential role in hormonal signaling pathways, where NO acts as a critical signaling molecule promoting ABA-induced stomatal closure in case of hyperosmotic stress [147]. NO is able to reduce the toxic levels of hydrogen peroxide (H2O2), superoxide anions (O2−), and lipid-free radicals (R*). Moreover, NO can alleviate the negative impact of ROS through lipid peroxidation, enhancing photosynthesis as well as the expression of antioxidant enzymes via mitogen-activated protein kinase (MAPK) and other signaling pathways [142]. It was found that exogenous application of NO caused a significant increase in seedling growth of cucumber by decreasing seedlings injury, enhancing the antioxidant activities and chlorophyll and free proline contents under stress conditions. Moreover, an exogenous supply of NO in wheat markedly preserved high relative water content via decreasing transpiration, increasing K content, and mitigated cell membrane damage [148] (Figure 4).
Salinity stress and nitrogen in plant
Salinity stress is one of the harmful abiotic stress elements that impact the metabolism and productivity of crop plants worldwide. Salinity increases represent global hazards and threats to food security, resulting in the remarkable conversion of agricultural arable soil into unproductive land [149]. The high level of salt ions in agricultural soil may be harmful to plants causing salt stress, where sodium (Na+) and chlorine (Cl−) concentrations increase in saline soil and affect the natural physiological process in plants [150]. The most efficient way to improve plant development under salt stress is to apply nitrogen (N) due it is considered one of the most influencing plant growth macronutrients in saline environments. Nitrogen is a necessary, powerful nutrient that mediates various molecular, physiological, and cellular responses essential to plant survival as well as different signal transduction pathways linked to plant defense systems and salt-stress tolerance [151,152]. Moreover, salt stress affects ammonification and nitrification in the soil due to chloride (Cl-) competes with nitrate (NO3-), ammonium (NH4+) competes with sodium (Na+) and creates ion toxicities and inequalities, which can limit the processes of N uptake, transport, and assimilation [153,154]. Salt stress has inhibited various enzyme activities that contribute to N assimilation in plants such as maize, tomato, rice, cowpea, and mung bean, reducing N uptake and utilization [155].
Interestingly, it was reported that the interaction between salt stress and N had an impact on plant growth, and the effect of N on salt-exposed plants was dependent on the N rate [150]. In the case of maize, excessive N treatment can successfully mitigate the deleterious effects of salt stress [156]. Similar findings in tomatoes showed that supra-optimum N was better than optimum N in reducing salt stress [138]. Under salinity stress, the application of N caused a significant increase in N and K uptake and decreased Na accumulation in wheat seedlings. Furthermore, N application can protect wheat seedlings in saline conditions via the upregulation of osmolytes, antioxidant system, and secondary metabolite accumulation [149]. In contrast, excessive N application exacerbated the negative effect of salt stress on wheat and rice, while moderate N supply could ameliorate the deleterious effect of salt stress. Therefore, applying an appropriate N rate is the key to mitigating the adverse effects caused by salt stress [150].
On the contrary, it was reported that the application of NO3- has resulted in an increase of Na+ uptake and loading of the sodium into the xylem, increasing root inhibition caused by the salinity. The co-transportation of Na+ and NO3- was suggested based on NO3- dependent Na+ uptake data at various Na+ concentrations [13]. Similarly, NO3-dependent transport mechanisms in salt-stress environments encourage Na+ ion uptake and loading into the xylem, which may represent a main pathway for Na+ accumulation in Arabidopsis shoots [151].
Nitric oxide and salt stress
It has been demonstrated that nitric oxide can act as a signal to induce the resistance of salt stress by raising the K: Na ratio in plant cells, which is based on an increase in the H1-ATPase activity of plasma membrane in calluses from maize seedlings, reed (Phragmites communis) plants, and poplar trees [144]. Exogenous NO can improve salt tolerance under salinity conditions by stimulating the K+/Na+ ratio, the Na+/H+ antiport in the tonoplast, and proton-pump activity [157]. Under salt stress, NO improve the absorption and translocation of several macro- and micronutrients, including K, Fe, Mg, and Zn, which improve respiration and chlorophyll biosynthesis [158]. The uptake of mineral nutritional elements such as nitrogen (N), zinc (Zn), iron (Fe), potassium (K), and calcium (Ca) was improved as a result of NO application and thereby diminished the harmful effect of high salinity on strawberry plants [159]. The exogenous NO supply can protect chickpea plants from salt stress-induced oxidative harm by promoting the biosynthesis of photosynthetic pigment, and antioxidant enzymes, osmolytes accumulation, and thereby improving plant development under saline stress [160]. Furthermore, under salt stress conditions, exogenous NO treatment enhanced wheat seed germination by converting starch to sugars, where the application of NO led to elevate in the effectiveness of α- and β-amylase enzymes [158,161]. Under salt stress, NO treatment boosted plant development and demonstrated a protective ability against salinity-triggered oxidative stress by increasing the effectiveness of peroxidase (POD), superoxide dismutase (SOD), and catalase (CAT) enzymes in Triticum aestivum [159].
Nitrate fertilizers have hazardous health issues
Nutrient deficiency and/or availability are crucial to plant production [162,163]. One of the severe issues the world faces is the adverse effects of fertilizers on the environment. Farmers have been using fertilizers since ancient times, but long-term use has affected soil fertility adversely. Due to over-fertilization in many agricultural areas, environmental pollution was established [164]. The seriousness of N pollution is a critical factor affecting food sustainability. For example, Nitrate pollution in groundwater is a severe problem worldwide, affecting animal and human health. In addition, Nitrate leaching decreases N availability for crops and can cause water pollution. In addition, high rates of N2 fertilizer application to crops led to low N fertilizer use efficiency and reduced N availability by Nitrate leaching, which may cause groundwater nitrate pollution [165]. In general, the improper application of fertilizers damages the farmlands, leading to soil and water pollution. It also causes heavy metals contaminations and soil erosion. Nitrous oxide and methane lead to greenhouse gas (GHG) emissions and reduce food quality.
While ammonium sulfate cause soil pollution, nitrogen oxides and ammonia cause air pollution. Nitrate is the final breakdown product of the nitrogen applications accumulating in groundwater, contaminating drinking water [166,167]. Since Nitrate levels above 10 mg L−1 N cause methemoglobinemia which is fatal. The U.S. Environmental Protection Agency set human drinking water to maintain nitrate levels below a Maximum Contaminant Level (MCL) at a critical value of 10 mg L−1 nitrate-nitrogen N [168]. In Argentina, the national food law set the limit for human consumption at 11.3 mg L−1 NO3−-N. Consumption of nitrate-rich water has been associated with methemoglobinemia (Blue Child Syndrome) and different forms of cancer [169,170,171]. Furthermore, diabetes, adverse reproductive outcomes (especially neural tube defects), and thyroid conditions are related to nitrate-contaminated drinking water.
A notable study by Maghanga et al. (2013) was conducted to monitor changes in surface water nitrate levels in ten rivers within a Kenyan tea plantation for three years. Water samples were obtained before and after fertilizer application in 2004, 2005, and 2006. For three years, there was no established trend between surface water nitrate levels and the time of fertilizer applications. However, the highest nitrate-nitrogen levels were in river Temochewa in 2005 during the first fertilizer applications (4.9 mg L−1 to 8.2 mg L−1). Furthermore, fertilizer application increased nitrate levels, and the study indicated that initial nitrate-nitrogen levels in most rivers were high, causing surface water contamination [172].
Establishing nitrogen fixation in plants by diazotrophic microbiota
Rapid human population growth and declining agricultural soils intensify resource competition, affecting human food hunger and sustainable agriculture systems worldwide [173]. Hence, developing alternative strategies to increase the current food demand is an inevitable need [174]. Accordingly, crop productivity is the pillar of nutritional food security and heavily depends on applying nitrogen fertilizers [175]. Nitrogen element is a critical requirement for crop productivity; While applying N fertilizers consumes vast amounts of energy, the excess harms environmental health. In addition, the expensive cost, environmental hazards, soil fertility reduction, and other negative consequences [176,177]. Therefore, increasing plant N use efficiency (NUE) is essential for developing sustainable agriculture [178].
Ultimately nitrogen fixation has multiple benefits through improving the yields of landraces that often receive little attention from breeders and empowering farmers using a sustainable and less expensive strategy. Thus, this technology can improve farmers' nutrition by enhancing access to indigenous microbial biodiversity, income, and resilience. Additionally, it reduces the need for fossil fuels requiring synthetic nitrogen fertilizers [179,180]. Overall, the symbiotic relationship between leguminous plants and rhizobia is a powerful adaptation strategy that enables these plants to reduce their dependence on synthetic nitrogen fertilizers and thrive in nitrogen-deficient soils. The ability to fix atmospheric dinitrogen (N2) and transfer it to legumes has a distinct advantage over non-leguminous plants in nitrogen-stressed environments, contributing to their ecological significance and agricultural importance in sustainable farming practices [181,182]. Taking together, improving the microbial activities (Rhizobium and Bacillus spp.) that contribute positively to boosting plant growth to sustain and increase food production under diverse climate situations is necessary [183]. In that sense, several beneficial mutualistic microbes have been discovered; however, their dependable utilization as biofertilizers in soil conditions is still challenged. Thus, improving the microbial inoculants, exploring their diversity under variable conditions via testing in vitro, and improving their performance in the external habitat could lead to understanding the knowledge gap between the microbial performance in vitro and the external habitat. Besides, biofertilizers have the advantage of being eco-friendly and cheap [184]. Applying mineral fertilizers at site-specific doses, increasing organic waste in agriculture, and bio-fertilizers with nitrogen-fixing bacteria and legumes in crop rotation seem more environmentally sustainable. In addition, N supply is necessary to provide N required for nucleotide and protein biosynthesis with other functions, i.e., carbon fixation capacity [185].
Bacterial endophytes associated with crops, i.e., corn and wheat, can produce various sugar compositions, such as arabinose [186]. They stimulate the production of a thick gelatinous layer (mucilage) around the root systems of these crops. This layer allows crops to better update fertilizer by forming a continuous boundary layer between roots and soil particles, protecting the root system from drought, and supporting other microbes (Figure 5). Most interestingly, mucilage structures associated with complex microbiota contribute to nutrient acquisition play a vital role in plant growth and defence [180]. Since inorganic fertilizers are produced from fossil fuels and are estimated to use 1% to 2% of the global energy supply, the structure of a complex polysaccharide (e.g., fucose, arabinose, and xylose) comprising the mucilage might provide the energy source to fuel nitrogenase activity [187].
Remarkably, a novel concept related to the protective plant strategy under stress [188] regarding nitrogen fixation in an indigenous maize landrace grown in nitrogen-stressed fields near Oaxaca, Mexico, was recently reported. High levels of a complex nitrogen-fixing microbiome were associated with the abundant production of a sugar-rich mucilage-supported nitrogen fixation [180]. They hypothesized a general mechanism of N2 fixation for cereals using mucilage structure to support associations with microbial diazotrophs. Setaria viridis and Setaria italica had a significant amount of fixed nitrogen associated with Azospirillum brasilense, contributing to growth promotion [189,190]. Several factors, such as plant root communities, plant genotypes, plant growth, and root microbe associations, considerably influence the microbial abundance and the quantity of mucilage exudation that modify the rhizodeposition [191,192]. In this context, mucilage exudation significantly increased under drying environmental conditions. In addition, mucilage secreted from plants grown in water-stressed soil had a higher contact angle than mucilage from plants grown in wet soil [180,191]. This research will open a new avenue in cereal crop-microbe interactions by transferring atmospheric nitrogen with significant economic and environmental benefits to farmers.
Moreover, reducing the chemical N fertilizer application by modifying N2-fixing bacteria in the microbial structure of cereal crop rhizospheres will improve the N-fixing efficiency. Improving the sustainability of biologically fixed N2 to cereal crops is a future need to enhance crop productivity, representing a significant breakthrough in N-fixation research. To achieve this goal, engineering key regulators of N2-fixing by activating the symbiotic signaling pathway between diazotrophic bacteria and cereal crops is a promising strategy for sustainable development [181].
Fortunately, establishing new Omics techniques, such as metabolomics, metagenomics, metatranscriptomics, and metaproteomics, will help in-depth research and identify the rhizospheric microbes using next-generation sequencing approaches [193]. Accordingly, manipulating the promising microbes that replace synthetic fertilizers and improve food security independently is a solution. Rhizobium-plant symbiosis is now becoming challenging with diverse genetic modifiers of the symbiosis relationship, not only in the genomic variability of partners but also in the soil microbiota. In that sense, there is a future direction to improve the specificity of the partnership during symbiotic colonization using different methods (e.g., axenic cultures) by eliminating other microbes. Therefore, identifying and developing bioinoculant strains and genotypes by adapting and evolving elite strains in both lab-scale analyses and trials to open-field applications to increase crop yield becomes crucial. Considering the human–microbe interaction to find non-pathogenic microbes (especially in the rhizobia genomes) that can colonize their host plant in field conditions.
Can microbes replace synthetic fertilizers?
Variable climatic conditions, abiotic stressors, and imbalance in nitrogen cycling are the threatening factors that limit crop production and sustainable agriculture. Fertile soil is a primary component in the backbone of sustainable agriculture; however, land degradation and rapid desertification cause an estimated global loss of 24 billion tons of fertile arable land [194]. Noticeably, eutrophication and decline of soil fertility result from massive application of chemical fertilizers, causing significant environmental concern [195]. Therefore, the production and application of microbial fertilizers are increasing globally due to the negative impacts of the overuse of chemical-based fertilizers [196,197]. Moreover, with the limited N resources and increasing agricultural demands for nitrogen supplies, there is a need to discover diverse mutualistic interactions between plant roots and rhizosphere microbiome (Figure 5).
Soils are habitats for diverse microorganisms, either harmful (pathogenic) or beneficial (symbiotic) to plants, comprising organic substances, water, and nutrients. Food production requires essential nutrients, metabolites, and water that the soil provides to plants. Thus, soil fertility is a vital natural resource from which most global food is produced, directly related to the quantity and quality of produced food [173,198,199]. Besides the nutrient resources in the soil, plants attract microbes via root exudates, root border cells, and mucilages that serve as food for the rhizosphere microbiome and their assembly [200]. Interestingly, soil contains an extensive reservoir of microorganisms (1 × 109 microbial cells g−1 dry soil) and microbial diversity (1 × 105 microbial species g−1 dry soil) [201,202]. Rhizospheric microbes are crucial in diverse biological processes, including soil structure, climate regulation, disease control, and organic matter decomposition. Rhizosphere microbiome plays a functional role in plant growth via various aspects, including nutrient uptake, cycling water availability, and the formation of stable aggregates to reduce the risks of soil erosion [203,204,205]. In addition, these microbes can improve plant health and serve as biocontrol agents against plant pathogens as well as biofertilizers (eco-friendly alternatives to hazardous synthetic fertilizers) (i.e., Rhizobium, Azotobacter, Bacillus subtilis) [184,206]. Accordingly, the functional role of these microbial communities has had enormous efforts from botanical scientists over two decades to eliminate hunger and improve soil fertility by reducing synthetic fertilizers [207,208,209,210]. Protein (amino acid) deficiencies, primarily in Africa and South Asia, negatively impact people globally [211]. In addition, metabolites secreted by plant roots, such as flavonoids, benzaldehyde, biochanin A, salicylic acid, terpenoids, and tryptophan.
Furthermore, some bacterial metabolites in this signaling help build a beneficial association between microbes and their host. For example, 2,4-diacetyl phloroglucinol, hydrogen cyanide (HCN), indole-acetic-acid (IAA), lipopeptides, phenazine-1-carboxamide salicylic acid, and siderophores play a vital role in the plant-microbe signaling and promoting the bacterial colonization [212,213]. For example, N-fixer microbes convert nitrogen from the atmosphere into reactive nitrogen that plants can use in the natural rhizosphere as an alternative to chemical fertilization practices. Nitrogen is required to synthesize chlorophyll and other amino acids during crop production. Currently, nitrogen starvation is expected during fuel price increases, affecting synthetic fertilizer manufacture [214,215].
Consequently, synthetic nutrients will be unaffordable for million smallholder farmers worldwide [216]. Fortunately, legume plants represent a solution, including fava beans, soybeans, lentils, and cowpea. They recruit symbiotic rhizobia bacteria from the soil into specialized root organs called nodules (N2-fixation factory) through biological nitrogen fixation (BNF) (Figure 5). Nitrogen fixation is an eco-friendly and manageable strategy by harnessing nitrogen-fixing endophytic and free-living rhizobacteria to increase nitrogen levels in agricultural land. Numerous studies revealed that some beneficial microbes, i.e., Rhizobium, Pseudomonas, Azospirillum, and Bacillus, positively impacted crops by enhancing above and belowground biomass and could play vital roles in achieving sustainable agriculture outcomes [217]. Biological nitrogen fixation converts dinitrogen (N2) to the fixed form of ammonia (NH3), essential for crop productivity, as fixed nitrogen is a critical component of many biological molecules [218].
The biologically fixed amounts of atmospheric nitrogen into ammonia inside root nodules by legumes account for about 65% of nitrogen utilization in global agriculture via mutualistic symbiotic relationships with nitrogen-fixing rhizobia. Rhizobia can remain viable in mature nodules [219]. Accordingly, this process allows legume plants to grow well in N-deficient soils, eliminating N fertilizer application [220]. The inoculation of rhizobia strains (native or commercial) was studied on the growth and nodulation of three cowpea (Vigna unguiculata) genotypes in semiarid regions of Kenya. Nitrogen (N2) is one of the limiting nutrients in semiarid regions, adversely affecting plant growth. Field trials were performed in a randomized complete block design with three replications, and the symbiotic efficiency (SE) of Rhizobium isolates was evaluated. In the field, rhizobia inoculation significantly (P < 0.05) increased nodulation and shoot DW compared to the uninoculated controls. Interestingly, Rhizobia inoculation significantly increased yields, whereas inoculation with native isolates recorded a 22.7% increase compared to uninoculated control in the first season and a 28.6% increase in yield in the second season. This study concluded that the efficient native rhizobia in smallholder farms with a promising potential to improve cowpea yield under a changing climate. In particular, R. tropici clone H53, Mesorhizobium sp. WSM3874 and R. pusense strain Nak353 showed more superiority in all the tested parameters. However, rhizobia inoculants improved nodulation and crop yield; cowpea genotypes did not prefer any inoculants. A practical and sustainable biofertilizer approach will empower cowpea production in subsistence farmers and establish effective symbiosis among legumes [221]. A recent study revealed that plant exudates and chemotaxis play a role in the development of bacterial biofilms in the soil, and creating microaerophilic conditions improves the nitrogen-fixing efficacy of diazotrophic bacteria in the soil, supplying plants with ammonia. In addition, plants support a diverse bacterial signal on the root surface or inside the tissues [222]. Flavonoids released by plant roots into the rhizosphere are crucial in regulating interactions between plants and microbes and controlling bacterial colonization [223]. The biofilm formation in the nitrogen-fixing bacterium Gluconacetobacter diazotrophicus [224] exhibited enhanced flavonoid compounds. This study also hypothesized that manipulating flavonoid synthetic pathways leads to the induction of biological nitrogen fixation in cereals through biofilm formation in soil diazotrophs [225,226]. Furthermore, flavonoid compounds are critical to the symbiotic relationship between legumes and rhizobia via inducing rhizobial Nod genes that signal nodule formation [227]. Biofilm formation (multicellular communities of microorganisms) is critical to the nitrogenase activity, which supported a free-living rhizospheric bacterium Pseudomonas stutzeri A1501, and Azospirillum brasilense to fix nitrogen under nitrogen-limiting conditions. Moreover, a positive correlation was observed between the nitrogen-fixing efficacy and the biofilm production of Gluconacetobacter diazotrophicus. Most interestingly, N-limiting signals very likely induce the production of exopolysaccharides which may function as a barrier to block excess oxygen in the air and provide a suitable microaerobic condition for bacteria inside the EPS to fix nitrogen [228,229]. A large amount of exopolysaccharide was reported in response to N limitation by cellulolytic bacteria (e.g., Bacillus, Pedobacter, Chryseobacterium, and Flavobacterium [230]. Therefore, exopolysaccharide production could be a general response of bacteria to N-limiting growth conditions. They hypothesized biofilm formation is a competitive carbon and nitrogen acquisition strategy and provides valuable insights linking nitrogen inputs to carbon sequestration. Applying the symbiotic association between rhizobia and leguminous plants is a significant research area to reduce the dependency on chemical fertilizers. Bradyrhizobium, Azospirillum, and Rhizobium had high plant growth-promoting capacity, N-fixing efficiency, and resistance to heavy metals (e.g., Cd contamination), indicating abundant functional microbial resources in extreme soil environments [231,232] (Figure 5).
Inoculation of Azospirillum, Burkholderia, and other nitrogen-fixing bacteria had little success in the field [233]. Thus, the evolution of a compatible host for the indigenous nitrogen-fixing bacteria is critical for the survival rate, colonizing the host, and improving nitrogenase activity.
In response to environmental pressure (e.g., nitrogen stress), leguminous plants may allocate more energy and resources for nodule formation by secreting phenolics (such as flavonoids) to induce the expression of rhizobial nodulation genes. These phenolics act in a pathway with the bacterial activator protein NodD to regulate gene expression that codes for Nod-factor synthesizing proteins as plant stimulants [234]. Legume crops may exhibit changes in the efficiency of nitrogen fixation within the nodules in response to nitrogen stress to maximize the nitrogen-fixing capacity of rhizobia through various molecular and physiological processes. In addition, plants elaborately adjust their growth and nitrogen uptake. Rhizobia and the host legume plant exchange various molecular signals to maximize symbiont performances; some are essential for symbiosis (such as flavonoids and Nod factors), while others influence the association [235]. Thus, we advocate more future research efforts to explore the elite rhizosphere microbiota under fluctuating climatic conditions and limited economic resources (Figure 5).
Arctic N cycling microbiota
By the end of this century, arctic land areas are expected to warm by 5.6–12.4 °C [236], resulting in significant permafrost thawing [237,238] and significant alterations in ecological functioning [239]. Massive nitrogen stocks (>67 billion tons) [240] deposited in permafrost thousands of years ago may now be available for breakdown, resulting in the release of nitrous oxide (N2O) into the atmosphere. N2O is an influential greenhouse gas, over 300 times more powerful than CO2 in terms of warming the climate. While the dynamics of carbon in the Arctic are well understood, the fact that Arctic soils store massive amounts of nitrogen has gotten little attention thus far [241]. Microorganisms can convert nitrogen compounds as reactive and harmful as nitric oxide or as inert and harmless as dinitrogen gas. In brief, a dinitrogen gas molecule is first fixed to ammonia, which then transforms into organic nitrogen (biomass). Ammonification, the decomposition of organic nitrogen, produces a molecule of ammonia, which is then oxidized to nitrate via nitrification (NH4+ to NO2- to NO3-) and eventually transformed back into a molecule of dinitrogen gas via denitrification (NO3- to NO2- to NO to N2O to N2) or anaerobic ammonium oxidation (anammox; NO2- + NH4+ to N2) [242]. In addition to methane and carbon dioxide fluxes, warming arctic soils release nitrogen, which can increase nitrous oxide (N2O) emissions, another potent greenhouse gas. Microbial mechanisms such as denitrification have a crucial role in controlling N2O release. The significance and amount of N2O released from Arctic soils are unknown, as are the bacteria responsible for greenhouse gas emissions.
Genomic approaches have a role in identifying new genes to improve nitrogen fixation
The identification of new genes involved in nitrogen fixation, a complete investigation of the gene organization of nitrogen-fixing species, and the discovery of novel diazotrophic species have all been made possible by genomics. This section of the review discusses the essential traits of nitrogen-fixing genomes as well as techniques for employing genome scanning to find and examine the genomes of new diazotrophs [243]. These diazotrophs are bacteria that can convert atmospheric nitrogen gas into ammonia, a more beneficial form of nitrogen. For example, Klebsiella pneumoniae and Azotobacter vinelandii may develop without external sources of fixed nitrogen [244,245]. The primary genes involved in nitrogen fixation were addressed. In order to explore nitrogen-fixing at the genetic, biochemical, and biophysical levels, we also evaluate methods for genome modification.
Additionally, one of the four types of nitrogenase enzymes is responsible for reducing nitrogen to ammonia. (EC 1.18.6.1; EC 1.19.6.1), which are generated by specific bacteria. All diazotrophs examined thus far employ at least one of these enzymes. The first three forms of nitrogenase are linked architecturally, phylogenetically, and mechanistically; however, they differ in that either vanadium or molybdenum comprise the heterometal component or possess a homometal cofactor that solely comprises iron. The fourth form of nitrogenase, a superoxide-dependent, molybdopterin-containing nitrogenase, was discovered in a single species, Streptomyces thermoautotrophicus. The genomic sequence of S. thermoautotrophicus and the amino acid sequence of this fourth kind of nitrogenase has not yet been established [246,247]. The fact that the inventory and distribution of species encoding for the fourth kind of nitrogenase are unknown highlights how few diazotrophs' genomes have been sequenced. A. vinelandii is one of the most extensively researched species among diazotrophs with sequenced genomes. In an aerobic culture, the free-living bacteria A. vinelandii can fix nitrogen. It has served as a model for nitrogen fixation for decades due to several factors, including (i) its genetic adaptability, (ii) its capacity to fix nitrogen during aerobic growth, (iii) its nutritional flexibility—which is demonstrated by its capacity to fix nitrogen via three different pathways and its capacity to adapt its metabolism to a variety of nutrients and media additives—(iv) and (v) the genome's comprehensive sequencing and manual editing [248,249]. A. vinelandii has a single circular chromosome with 5,365,318 base pairs in its genome, which is expected to code for 5051 proteins [249]. The diazotroph Pseudomonas stutzeri is the closest companion of A. vinelandii with a sequenced genome [250]. Even though P. stutzeri may grow aerobically and can only fix nitrogen in microaerobic conditions [251,252]. Alternatively, in ambient oxygen concentrations (20% O2), A. vinelandii can catalyze nitrogen fixation, an oxygen-sensitive process [253,254]. Its respiratory protection system makes it feasible for it to undertake two incompatible biological processes, oxidative phosphorylation, and nitrogen fixation, concurrently. A. vinelandii may modify its respiration rate to maintain a low amount of cytoplasmic oxygen while nitrogen-fixing is taking place [255]. Five terminal oxidases were found in the genome, along with additional NADH oxidoreductases and other respiratory complexes that provide electrons to terminal oxidases, raising oxygen consumption [249]. Some of these genes, such as cydAB I, have been linked to respiratory protection and are crucial for aerobic nitrogen fixation [246,247].
The Discovery of novel nitrogen-fixing organisms and the quantity and kind of nitrogenases coding in these species may also be targeted by genome sequencing utilizing NifD sequences as queries [256,257,258]. The genomes of diazotrophs also encode for other proteins comparable to NifD, in addition to the Nif/Vnf/AnfD sequences [258]. The discovery of new genes involved in nitrogen fixation in A. vinelandii and other diazotrophs might improve the efficacy of nitrogen fixation.
Conclusion
Nitrogen is an essential macronutrient required for the growth and development of all living organisms. The most common inorganic forms are nitrate (NO3-) and ammonium (NH4+), transported across the root plasma membrane. However, applying nitrogen fertilizers is necessary for plant growth and productivity, but massive amounts of synthetic agro inputs to fertilize crops used by farmers harm environmental health. In this sense, improving the use of N efficiency by plants is a promising strategy to reduce the negative impact of using synthetic fertilizers. Bacterial endophytes play a crucial role in the nitrogen cycle by converting atmospheric nitrogen gas into ammonia, a more beneficial form of nitrogen via biological nitrogen fixation (BNF). Nitrogen fixation is an eco-friendly and manageable strategy to increase nitrogen levels in agricultural land. Thus, improving nitrogen bacterial production by genome editing could replace synthetic fertilizers to reduce dependency on conventional chemical fertilizers. In addition, discovering new plant microbiomes and understanding their structure, abundance, and diversity will open a new avenue to better understanding the agricultural scenario to sustain global food security under limited nitrogen conditions.
Author Contributions
All authors listed have made substantial, direct, and intellectual contributions to the work and approved it for publication.
Funding
Ze-Chun Yuan’s lab appreciates the supports from Agriculture and Agri-Food Canada A-base project funds (J-002488 and J-003105), Mitacs fund, the Natural Sciences and Engineering Research Council of Canada (NSERC) Strategic Partnership Grant as well as the support from Ontario Greenhouse and Vegetable Growers Association.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lal, R. Restoring Soil Quality to Mitigate Soil Degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environment International 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Bogunovic, I.; Muñoz-Rojas, M.; Brevik, E.C. Soil ecosystem services, sustainability, valuation and management. Current Opinion in Environmental Science & Health 2018, 5, 7–13. [Google Scholar] [CrossRef]
- Pimentel, D.; Burgess, M. Soil Erosion Threatens Food Production. Agriculture 2013, 3, 443–463. [Google Scholar] [CrossRef]
- Díaz-Rodríguez, A.M.; Salcedo Gastelum, L.A.; Félix Pablos, C.M.; Parra-Cota, F.I.; Santoyo, G.; Puente, M.L.; Bhattacharya, D.; Mukherjee, J.; de los Santos-Villalobos, S. The Current and Future Role of Microbial Culture Collections in Food Security Worldwide. Frontiers in Sustainable Food Systems 2021, 4. [Google Scholar] [CrossRef]
- Arora, N.K.; Khare, E.; Maheshwari, D.K. Plant Growth Promoting Rhizobacteria: Constraints in Bioformulation, Commercialization, and Future Strategies. In Plant Growth and Health Promoting Bacteria; Maheshwari, D.K., Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2011; pp. 97–116. [Google Scholar] [CrossRef]
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving Nitrogen Use Efficiency in Crops for Sustainable Agriculture. Sustainability 2011, 3, 1452–1485. [Google Scholar] [CrossRef]
- Akinnifesi, F.K.; Ajayi, O.C.; Sileshi, G.; Chirwa, P.W.; Chianu, J. Fertiliser trees for sustainable food security in the maize-based production systems of East and Southern Africa. A review. Agronomy for Sustainable Development 2010, 30, 615–629. [Google Scholar] [CrossRef]
- Spiertz, H. Avenues to meet food security. The role of agronomy on solving complexity in food production and resource use. European Journal of Agronomy 2012, 43, 1–8. [Google Scholar] [CrossRef]
- Azadi, H.; Ho, P. Genetically modified and organic crops in developing countries: A review of options for food security. Biotechnology Advances 2010, 28, 160–168. [Google Scholar] [CrossRef]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Science of the Total Environment 2018, 634, 497–500. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture. Annals of botany 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.Y.; Tian, W.H.; Jin, C.W. Nitrogen in plants: from nutrition to the modulation of abiotic stress adaptation. Stress Biology 2022, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, N.; Sethi, M.; Kumar, A.; Dang, D.; Singh, J.; Chhuneja, P. Biochemical and genetic approaches improving nitrogen use efficiency in cereal crops: A review. Frontiers in plant science 2021, 12, 657629. [Google Scholar] [CrossRef]
- Delin, S.; Stenberg, B.; Nyberg, A.; Brohede, L. Potential methods for estimating nitrogen fertilizer value of organic residues. Soil Use and Management 2012, 28, 283–291. [Google Scholar] [CrossRef]
- Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino acid transporters in plants: Identification and function. Plants 2020, 9, 972. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Dong, C.; Yang, T.; Bao, S.; Fang, W.; Lucas, W.J.; Zhang, Z. The tea plant CsLHT1 and CsLHT6 transporters take up amino acids, as a nitrogen source, from the soil of organic tea plantations. Horticulture Research 2021, 8. [Google Scholar] [CrossRef]
- Feng, H.; Fan, X.; Miller, A.J.; Xu, G. Plant nitrogen uptake and assimilation: regulation of cellular pH homeostasis. Journal of Experimental Botany 2020, 71, 4380–4392. [Google Scholar] [CrossRef]
- Forde, B.G. Nitrate transporters in plants: structure, function and regulation. Biochimica et Biophysica Acta (BBA)-Biomembranes 2000, 1465, 219–235. [Google Scholar] [CrossRef]
- Iqbal, A.; Qiang, D.; Zhun, W.; Xiangru, W.; Huiping, G.; Hengheng, Z.; Nianchang, P.; Xiling, Z.; Meizhen, S. Growth and nitrogen metabolism are associated with nitrogen-use efficiency in cotton genotypes. Plant Physiology and Biochemistry 2020, 149, 61–74. [Google Scholar] [CrossRef]
- Léran, S.; Varala, K.; Boyer, J.-C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B. A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends in plant science 2014, 19, 5–9. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Hsu, P.-K.; Tsay, Y.-F. Uptake, allocation and signaling of nitrate. Trends in plant science 2012, 17, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Cheng, Y.-H.; Chen, K.-E.; Tsay, Y.-F. Nitrate transport, signaling, and use efficiency. Annual review of plant biology 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Kiba, T.; Feria-Bourrellier, A.-B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Bréhaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H. The Arabidopsis nitrate transporter NRT2. 4 plays a double role in roots and shoots of nitrogen-starved plants. The Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Chattha, M.S.; Ali, Q.; Haroon, M.; Afzal, M.J.; Javed, T.; Hussain, S.; Mahmood, T.; Solanki, M.K.; Umar, A.; Abbas, W. Enhancement of nitrogen use efficiency through agronomic and molecular based approaches in cotton. Frontiers in Plant Science 2022, 13, 994306. [Google Scholar] [CrossRef] [PubMed]
- Maghiaoui, A.; Gojon, A.; Bach, L. NRT1. 1-centered nitrate signaling in plants. Journal of Experimental Botany 2020, 71, 6226–6237. [Google Scholar] [CrossRef] [PubMed]
- Izmailov, S.; Nikitin, A. Nitrate signaling in plants: mechanisms of implementation. Russian journal of plant physiology 2020, 67, 31–44. [Google Scholar] [CrossRef]
- O'Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate transport, sensing, and responses in plants. Molecular plant 2016, 9, 837–856. [Google Scholar] [CrossRef]
- Fang, X.Z.; Fang, S.Q.; Ye, Z.Q.; Liu, D.; Zhao, K.L.; Jin, C.W. NRT1. 1 dual-affinity nitrate transport/signalling and its roles in plant abiotic stress resistance. Frontiers in plant science 2021, 12, 715694. [Google Scholar] [CrossRef]
- Chen, C.-Z.; Lv, X.-F.; Li, J.-Y.; Yi, H.-Y.; Gong, J.-M. Arabidopsis NRT1. 5 is another essential component in the regulation of nitrate reallocation and stress tolerance. Plant Physiology 2012, 159, 1582–1590. [Google Scholar] [CrossRef]
- Nazish, T.; Arshad, M.; Jan, S.U.; Javaid, A.; Khan, M.H.; Naeem, M.A.; Baber, M.; Ali, M. Transporters and transcription factors gene families involved in improving nitrogen use efficiency (NUE) and assimilation in rice (Oryza sativa L.). Transgenic Research 2021, 1–20. [Google Scholar] [CrossRef]
- Guo, B.; Li, Y.; Wang, S.; Li, D.; Lv, C.; Xu, R. Characterization of the Nitrate Transporter gene family and functional identification of HvNRT2. 1 in barley (Hordeum vulgare L.). PLoS One 2020, 15, e0232056. [Google Scholar]
- Aluko, O.O.; Kant, S.; Adedire, O.M.; Li, C.; Yuan, G.; Liu, H.; Wang, Q. Unlocking the potentials of nitrate transporters at improving plant nitrogen use efficiency. Frontiers in Plant Science 2023, 14, 1074839. [Google Scholar] [CrossRef]
- Ji, L.; Song, L.; Zou, L.; Li, S.; Zhang, R.; Yang, J.; Wang, C.; Zhang, Y.; Wang, X.; Yun, L. Cassava Nitrate Transporter NPF5. 4 promotes both yield potential and salt tolerance in rice. 2022. [Google Scholar]
- Zhao, L.; Chen, P.; Liu, P.; Song, Y.; Zhang, D. Genetic effects and expression patterns of the nitrate transporter (NRT) gene family in Populus tomentosa. Frontiers in Plant Science 2021, 12, 661635. [Google Scholar] [CrossRef] [PubMed]
- Qi(杞金芳), J.; Yu(郁露), L.; Ding(丁静丽), J.; Ji(姬晨晨), C.; Wang(汪社亮), S.; Wang(王创), C.; Ding(丁广大), G.; Shi(石磊), L.; Xu(徐芳森), F.; Cai(蔡红梅), H. Transcription factor OsSNAC1 positively regulates nitrate transporter gene expression in rice. Plant Physiology 2023, 192, 2923–2942. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, K.; Xie, J.; Liu, J.; Tan, P.; Peng, F. Genome-Wide Identification and Expression Analysis of AMT and NRT Gene Family in Pecan (Carya illinoinensis) Seedlings Revealed a Preference for NH4+-N. International Journal of Molecular Sciences 2022, 23, 13314. [Google Scholar] [CrossRef]
- Schuldiner, S.; Shirvan, A.; Linial, M. Vesicular neurotransmitter transporters: from bacteria to humans. Physiological reviews 1995, 75, 369–392. [Google Scholar] [CrossRef]
- Tsay, Y.-F.; Chiu, C.-C.; Tsai, C.-B.; Ho, C.-H.; Hsu, P.-K. Nitrate transporters and peptide transporters. FEBS letters 2007, 581, 2290–2300. [Google Scholar] [CrossRef]
- Sun, J.; Bankston, J.R.; Payandeh, J.; Hinds, T.R.; Zagotta, W.N.; Zheng, N. Crystal structure of the plant dual-affinity nitrate transporter NRT1. 1. Nature 2014, 507, 73–77. [Google Scholar] [CrossRef]
- Madej, M.G.; Dang, S.; Yan, N.; Kaback, H.R. Evolutionary mix-and-match with MFS transporters. Proceedings of the National Academy of Sciences 2013, 110, 5870–5874. [Google Scholar] [CrossRef] [PubMed]
- Orsel, M.; Chopin, F.; Leleu, O.; Smith, S.J.; Krapp, A.; Daniel-Vedele, F.o.; Miller, A.J. Characterization of a Two-Component High-Affinity Nitrate Uptake System in Arabidopsis. Physiology and Protein-Protein Interaction. Plant Physiology 2006, 142, 1304–1317. [Google Scholar] [CrossRef]
- Okamoto, M.; Kumar, A.; Li, W.; Wang, Y.; Siddiqi, M.Y.; Crawford, N.M.; Glass, A.D.M. High-Affinity Nitrate Transport in Roots of Arabidopsis Depends on Expression of the NAR2-Like Gene AtNRT3.1. Plant Physiology 2006, 140, 1036–1046. [Google Scholar] [CrossRef]
- Benková, E.; Bielach, A. Lateral root organogenesis — from cell to organ. Current Opinion in Plant Biology 2010, 13, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Yong, Z.; Kotur, Z.; Glass, A.D. Characterization of an intact two-component high-affinity nitrate transporter from Arabidopsis roots. The Plant Journal 2010, 63, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Bowen, G.D.; Smith, S.E. THE EFFECTS OF MYCORRHIZAS ON NITROGEN UPTAKE BY PLANTS. Ecological Bulletins 1981, 237–247. [Google Scholar]
- Öhlund, J.; Näsholm, T. Growth of conifer seedlings on organic and inorganic nitrogen sources. Tree Physiology 2001, 21, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wirén, N. Three Functional Transporters for Constitutive, Diurnally Regulated, and Starvation-Induced Uptake of Ammonium into Arabidopsis Roots. The Plant Cell 1999, 11, 937–947. [Google Scholar] [CrossRef]
- Bloom, A.J.; Meyerhoff, P.A.; Taylor, A.R.; Rost, T.L. Root Development and Absorption of Ammonium and Nitrate from the Rhizosphere. Journal of Plant Growth Regulation 2002, 21, 416–431. [Google Scholar] [CrossRef]
- Bittsánszky, A.; Pilinszky, K.; Gyulai, G.; Komives, T. Overcoming ammonium toxicity. Plant Science 2015, 231, 184–190. [Google Scholar] [CrossRef]
- Renard, J.J.; Calidonna, S.E.; Henley, M.V. Fate of ammonia in the atmosphere—a review for applicability to hazardous releases. Journal of Hazardous Materials 2004, 108, 29–60. [Google Scholar] [CrossRef]
- Hachiya, T.; Watanabe, C.K.; Fujimoto, M.; Ishikawa, T.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Nitrate Addition Alleviates Ammonium Toxicity Without Lessening Ammonium Accumulation, Organic Acid Depletion and Inorganic Cation Depletion in Arabidopsis thaliana Shoots. Plant and Cell Physiology 2012, 53, 577–591. [Google Scholar] [CrossRef]
- von Wirén, N.; Gazzarrini, S.; Gojon, A.; Frommer, W.B. The molecular physiology of ammonium uptake and retrieval. Current Opinion in Plant Biology 2000, 3, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Loqué, D.; von Wirén, N. Regulatory levels for the transport of ammonium in plant roots. Journal of Experimental Botany 2004, 55, 1293–1305. [Google Scholar] [CrossRef]
- Glass, A.D.M.; Britto, D.T.; Kaiser, B.N.; Kinghorn, J.R.; Kronzucker, H.J.; Kumar, A.; Okamoto, M.; Rawat, S.; Siddiqi, M.Y.; Unkles, S.E.; et al. The regulation of nitrate and ammonium transport systems in plants. Journal of Experimental Botany 2002, 53, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.-L.; Zhou, J.-Y.; Yang, S.-Y.; Qi, W.; Yang, K.-J.; Su, Y.-H. Function and Regulation of Ammonium Transporters in Plants. International Journal of Molecular Sciences 2020, 21, 3557. [Google Scholar] [CrossRef]
- Behie, S.W.; Bidochka, M.J. Ubiquity of Insect-Derived Nitrogen Transfer to Plants by Endophytic Insect-Pathogenic Fungi: an Additional Branch of the Soil Nitrogen Cycle. Applied and Environmental Microbiology 2014, 80, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Giehl, R.F.H.; Laginha, A.M.; Duan, F.; Rentsch, D.; Yuan, L.; von Wirén, N. A Critical Role of AMT2;1 in Root-To-Shoot Translocation of Ammonium in Arabidopsis. Molecular Plant 2017, 10, 1449–1460. [Google Scholar] [CrossRef]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytologist 2007, 174, 137–150. [Google Scholar] [CrossRef]
- Esmaeilzadeh-Salestani, K.; Samandari_Bahraseman, M.R.; Tohidfar, M.; Khaleghdoust, B.; Keres, I.; Mõttus, A.; Loit, E. Expression of AMT1;1 and AMT2;1 is stimulated by mineral nitrogen and reproductive growth stage in barley under field conditions. Journal of Plant Nutrition 2023, 46, 1246–1258. [Google Scholar] [CrossRef]
- Wu, X.; Yang, H.; Qu, C.; Xu, Z.; Li, W.; Hao, B.; Yang, C.; Sun, G.; Liu, G. Sequence and expression analysis of the AMT gene family in poplar. Frontiers in Plant Science 2015, 6. [Google Scholar] [CrossRef]
- Wu, X.X.; Yuan, D.P.; Chen, H.; Kumar, V.; Kang, S.M.; Jia, B.; Xuan, Y.H. Ammonium transporter 1 increases rice resistance to sheath blight by promoting nitrogen assimilation and ethylene signalling. Plant Biotechnology Journal 2022, 20, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Pastor, V.; Gamir, J.; Camañes, G.; Cerezo, M.; Sánchez-Bel, P.; Flors, V. Disruption of the ammonium transporter AMT1.1 alters basal defenses generating resistance against Pseudomonas syringae and Plectosphaerella cucumerina. Frontiers in Plant Science 2014, 5. [Google Scholar] [CrossRef]
- Lima, J.E.; Kojima, S.; Takahashi, H.; von Wirén, N. Ammonium Triggers Lateral Root Branching in Arabidopsis in an AMMONIUM TRANSPORTER1;3-Dependent Manner. The Plant Cell 2010, 22, 3621–3633. [Google Scholar] [CrossRef] [PubMed]
- Rogato, A.; D’Apuzzo, E.; Chiurazzi, M. The multiple plant response to high ammonium conditions. Plant Signaling & Behavior 2010, 5, 1594–1596. [Google Scholar] [CrossRef]
- Rogato, A.; D’Apuzzo, E.; Barbulova, A.; Omrane, S.; Parlati, A.; Carfagna, S.; Costa, A.; Schiavo, F.L.; Esposito, S.; Chiurazzi, M. Characterization of a Developmental Root Response Caused by External Ammonium Supply in Lotus japonicus. Plant Physiology 2010, 154, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Liu, Q.; Liu, X.; Ma, Z.; Zhang, J.; Li, X.; Shen, L.; Yuan, J.; Zhang, Q. Phosphatidic acid regulates ammonium uptake by interacting with AMMONIUM TRANSPORTER 1;1 in Arabidopsis. Plant Physiology 2023, 10.1093/plphys/kiad421. [Google Scholar] [CrossRef]
- Giovannetti, M.; Volpe, V.; Salvioli, A.; Bonfante, P. Chapter 7 - Fungal and Plant Tools for the Uptake of Nutrients in Arbuscular Mycorrhizas: A Molecular View. In Mycorrhizal Mediation of Soil; Johnson, N.C., Gehring, C., Jansa, J., Eds.; Elsevier, 2017; pp. 107–128. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, W.; Wei, J.; Yoon, H.; An, G. Transcription Factor OsDOF18 Controls Ammonium Uptake by Inducing Ammonium Transporters in Rice Roots. Mol Cells 2017, 40, 178–185. [Google Scholar] [CrossRef]
- Yuan, L.; Gu, R.; Xuan, Y.; Smith-Valle, E.; Loqué, D.; Frommer, W.B.; von Wirén, N. Allosteric Regulation of Transport Activity by Heterotrimerization of Arabidopsis Ammonium Transporter Complexes in Vivo. The Plant Cell 2013, 25, 974–984. [Google Scholar] [CrossRef]
- Lanquar, V.; Loqué, D.; Hörmann, F.; Yuan, L.; Bohner, A.; Engelsberger, W.R.; Lalonde, S.; Schulze, W.X.; von Wirén, N.; Frommer, W.B. Feedback Inhibition of Ammonium Uptake by a Phospho-Dependent Allosteric Mechanism in Arabidopsis. The Plant Cell 2009, 21, 3610–3622. [Google Scholar] [CrossRef]
- Loqué, D.; Lalonde, S.; Looger, L.L.; von Wirén, N.; Frommer, W.B. A cytosolic trans-activation domain essential for ammonium uptake. Nature 2007, 446, 195–198. [Google Scholar] [CrossRef]
- Søgaard, R.; Alsterfjord, M.; MacAulay, N.; Zeuthen, T. Ammonium ion transport by the AMT/Rh homolog TaAMT1;1 is stimulated by acidic pH. Pflügers Archiv - European Journal of Physiology 2009, 458, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhao, Y.; Luo, W.; Li, R.; He, Q.; Fang, X.; Michele, R.D.; Ast, C.; von Wirén, N.; Lin, J. Single-particle analysis reveals shutoff control of the <i>Arabidopsis</i> ammonium transporter AMT1;3 by clustering and internalization. Proceedings of the National Academy of Sciences 2013, 110, 13204–13209. [Google Scholar] [CrossRef]
- Koltun, A.; Maniero, R.A.; Vitti, M.; de Setta, N.; Giehl, R.F.H.; Lima, J.E.; Figueira, A. Functional characterization of the sugarcane (Saccharum spp.) ammonium transporter AMT2;1 suggests a role in ammonium root-to-shoot translocation. Frontiers in Plant Science 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Nitrogen. Principles of plant nutrition 2001, 397–434. [Google Scholar]
- Sparacino-Watkins, C.; Stolz, J.F.; Basu, P. Nitrate and periplasmic nitrate reductases. Chemical Society Reviews 2014, 43, 676–706. [Google Scholar] [CrossRef] [PubMed]
- Rosswall, T. Some perspectives of the major biogeochemical cycles. Likens, GE., editor. Vol. 2. New York, USA: John Wiley & Sons Ithaca: 1981.
- Ward, M.H.; DeKok, T.M.; Levallois, P.; Brender, J.; Gulis, G.; Nolan, B.T.; VanDerslice, J. Workgroup report: drinking-water nitrate and health—recent findings and research needs. Environmental health perspectives 2005, 113, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Powlson, D.S.; Addiscott, T.M.; Benjamin, N.; Cassman, K.G.; de Kok, T.M.; van Grinsven, H.; L'hirondel, J.L.; Avery, A.A.; Van Kessel, C. When does nitrate become a risk for humans? Journal of environmental quality 2008, 37, 291–295. [Google Scholar] [CrossRef]
- Bonner, F.T.; Hughes, M.N. The aqueous solution chemistry of nitrogen in low positive oxidation states. Comments on Inorganic Chemistry 1988, 7, 215–234. [Google Scholar] [CrossRef]
- Jaffe, D.A. 12 The Nitrogen Cycle. In International Geophysics, Butcher, S.S., Charlson, R.J., Orians, G.H., Wolfe, G.V., Eds. Academic Press: 1992; Vol. 50, pp. 263–284.
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of Earth’s nitrogen cycle. science 2010, 330, 192–196. [Google Scholar] [CrossRef]
- Ishii, S.; Ikeda, S.; Minamisawa, K.; Senoo, K. Nitrogen cycling in rice paddy environments: past achievements and future challenges. Microbes and environments 2011, 26, 282–292. [Google Scholar] [CrossRef]
- Elrys, A.S.; Zhu, Q.; Jiang, C.; Liu, J.; Sobhy, H.H.; Shen, Q.; Uwiragiye, Y.; Wu, Y.; El-Tarabily, K.A.; Meng, L. Global soil nitrogen cycle pattern and nitrogen enrichment effects: Tropical versus subtropical forests. Global Change Biology 2023, 29, 1905–1921. [Google Scholar] [CrossRef] [PubMed]
- Joerger, R.; Jacobson, M.; Premakumar, R.; Wolfinger, E.; Bishop, P. Nucleotide sequence and mutational analysis of the structural genes (anfHDGK) for the second alternative nitrogenase from Azotobacter vinelandii. Journal of bacteriology 1989, 171, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Joerger, R.; Loveless, T.; Pau, R.; Mitchenall, L.; Simon, B.; Bishop, P. Nucleotide sequences and mutational analysis of the structural genes for nitrogenase 2 of Azotobacter vinelandii. Journal of bacteriology 1990, 172, 3400–3408. [Google Scholar] [CrossRef]
- Fani, R.; Gallo, R.; Liò, P. Molecular evolution of nitrogen fixation: the evolutionary history of the nifD, nifK, nifE, and nifN genes. Journal of molecular evolution 2000, 51, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cantera, J.J.L.; Stein, L.Y. Molecular diversity of nitrite reductase genes (nirK) in nitrifying bacteria. Environmental microbiology 2007, 9, 765–776. [Google Scholar] [CrossRef]
- Kartal, B.; Maalcke, W.J.; De Almeida, N.M.; Cirpus, I.; Gloerich, J.; Geerts, W.; Op den Camp, H.J.; Harhangi, H.R.; Janssen-Megens, E.M.; Francoijs, K.-J. Molecular mechanism of anaerobic ammonium oxidation. Nature 2011, 479, 127–130. [Google Scholar] [CrossRef]
- Jepson, B.J.; Mohan, S.; Clarke, T.A.; Gates, A.J.; Cole, J.A.; Butler, C.S.; Butt, J.N.; Hemmings, A.M.; Richardson, D.J. Spectropotentiometric and structural analysis of the periplasmic nitrate reductase from Escherichia coli. Journal of Biological Chemistry 2007, 282, 6425–6437. [Google Scholar] [CrossRef]
- Heiss, B.; Frunzke, K.; Zumft, W. Formation of the NN bond from nitric oxide by a membrane-bound cytochrome bc complex of nitrate-respiring (denitrifying) Pseudomonas stutzeri. Journal of bacteriology 1989, 171, 3288–3297. [Google Scholar] [CrossRef]
- Suzuki, M.; Arai, H.; Ishii, M.; Igarashi, Y. Gene structure and expression profile of cytochrome bc nitric oxide reductase from Hydrogenobacter thermophilus TK-6. Bioscience, biotechnology, and biochemistry 2006, 70, 1666–1671. [Google Scholar] [CrossRef]
- Ogawa, K.-i.; Akagawa, E.; Yamane, K.; Sun, Z.-W.; LaCelle, M.; Zuber, P.; Nakano, M.M. The nasB operon and nasA gene are required for nitrate/nitrite assimilation in Bacillus subtilis. Journal of bacteriology 1995, 177, 1409–1413. [Google Scholar] [CrossRef]
- Exley, G.E.; Colandene, J.D.; Garrett, R.H. Molecular cloning, characterization, and nucleotide sequence of nit-6, the structural gene for nitrite reductase in Neurospora crassa. Journal of bacteriology 1993, 175, 2379–2392. [Google Scholar] [CrossRef] [PubMed]
- Sohaskey, C.D.; Wayne, L.G. Role of narK2X and narGHJI inHypoxic Upregulation of Nitrate Reduction by Mycobacteriumtuberculosis. Journal of bacteriology 2003, 185, 7247–7256. [Google Scholar] [CrossRef] [PubMed]
- Stolz, J.F.; Basu, P. Evolution of nitrate reductase: molecular and structural variations on a common function. Chembiochem 2002, 3, 198–206. [Google Scholar] [CrossRef]
- Engels, C.; Marschner, H. Plant uptake and utilization of nitrogen. Nitrogen fertilization in the environment 1995, 41–81. [Google Scholar]
- Vega-Mas, I.; Cukier, C.; Coleto, I.; González-Murua, C.; Limami, A.M.; González-Moro, M.B.; Marino, D. Isotopic labelling reveals the efficient adaptation of wheat root TCA cycle flux modes to match carbon demand under ammonium nutrition. Scientific Reports 2019, 9, 8925. [Google Scholar] [CrossRef]
- Fujita, T.; Beier, M.P.; Tabuchi-Kobayashi, M.; Hayatsu, Y.; Nakamura, H.; Umetsu-Ohashi, T.; Sasaki, K.; Ishiyama, K.; Murozuka, E.; Kojima, M. Cytosolic glutamine synthetase GS1; 3 is involved in rice grain ripening and germination. Frontiers in Plant Science 2022, 13, 835835. [Google Scholar] [CrossRef]
- Lea, P.J.; Miflin, B.J. Nitrogen assimilation and its relevance to crop improvement. Annu. Plant Rev. Volume 2011, 42, 1–40. [Google Scholar]
- Miflin, B.J.; Lea, P.J. The pathway of nitrogen assimilation in plants. Phytochemistry 1976, 15, 873–885. [Google Scholar] [CrossRef]
- Oaks, A.; Stulen, I.; Jones, K.; Winspear, M.J.; Misra, S.; Boesel, I.L. Enzymes of nitrogen assimilation in maize roots. Planta 1980, 148, 477–484. [Google Scholar] [CrossRef]
- Tamura, W.; Hidaka, Y.; Tabuchi, M.; Kojima, S.; Hayakawa, T.; Sato, T.; Obara, M.; Kojima, M.; Sakakibara, H.; Yamaya, T. Reverse genetics approach to characterize a function of NADH-glutamate synthase1 in rice plants. Amino Acids 2010, 39, 1003–1012. [Google Scholar] [CrossRef]
- Kojima, S.; Minagawa, H.; Yoshida, C.; Inoue, E.; Takahashi, H.; Ishiyama, K. Coregulation of glutamine synthetase1; 2 (GLN1; 2) and NADH-dependent glutamate synthase (GLT1) gene expression in Arabidopsis roots in response to ammonium supply. Frontiers in Plant Science 2023, 14, 1127006. [Google Scholar] [CrossRef]
- Yasuda, T.; Konishi, N.; Kojima, S. Ammonium uptake capacity and response of cytosolic glutamine synthetase 1; 2 to ammonium supply are key factors for the adaptation of ammonium nutrition in Arabidopsis thaliana. Soil Science and Plant Nutrition 2017, 63, 553–560. [Google Scholar] [CrossRef]
- Konishi, N.; Saito, M.; Imagawa, F.; Kanno, K.; Yamaya, T.; Kojima, S. Cytosolic glutamine synthetase isozymes play redundant roles in ammonium assimilation under low-ammonium conditions in roots of Arabidopsis thaliana. Plant and Cell Physiology 2018, 59, 601–613. [Google Scholar] [CrossRef]
- Yamaya, T.; Kusano, M. Evidence supporting distinct functions of three cytosolic glutamine synthetases and two NADH-glutamate synthases in rice. Journal of experimental botany 2014, 65, 5519–5525. [Google Scholar] [CrossRef]
- Kusano, M.; Fukushima, A.; Redestig, H.; Saito, K. Metabolomic approaches toward understanding nitrogen metabolism in plants. Journal of Experimental Botany 2011, 62, 1439–1453. [Google Scholar] [CrossRef]
- Forde, B.G.; Lea, P.J. Glutamate in plants: metabolism, regulation, and signalling. Journal of experimental botany 2007, 58, 2339–2358. [Google Scholar] [CrossRef] [PubMed]
- Bowsher, C.G.; Lacey, A.E.; Hanke, G.T.; Clarkson, D.T.; Saker, L.R.; Stulen, I.; Emes, M.J. The effect of Glc6P uptake and its subsequent oxidation within pea root plastids on nitrite reduction and glutamate synthesis. Journal of experimental botany 2007, 58, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Habash, D.Z.; Bernard, S.; Schondelmaier, J.; Weyen, J.; Quarrie, S.A. The genetics of nitrogen use in hexaploid wheat: N utilisation, development and yield. Theoretical and Applied Genetics 2007, 114, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Yang, F.; He, X.; Du, X.; Mu, P.; Ma, W. Advances in the functional study of glutamine synthetase in plant abiotic stress tolerance response. The Crop Journal 2022, 10, 917–923. [Google Scholar] [CrossRef]
- Prinsi, B.; Espen, L. Mineral nitrogen sources differently affect root glutamine synthetase isoforms and amino acid balance among organs in maize. BMC plant biology 2015, 15, 1–13. [Google Scholar] [CrossRef]
- Zhang, Z.; Xiong, S.; Wei, Y.; Meng, X.; Wang, X.; Ma, X. The role of glutamine synthetase isozymes in enhancing nitrogen use efficiency of N-efficient winter wheat. Scientific Reports 2017, 7, 1000. [Google Scholar] [CrossRef] [PubMed]
- Habash, D.; Massiah, A.; Rong, H.; Wallsgrove, R.; Leigh, R. The role of cytosolic glutamine synthetase in wheat. Annals of Applied Biology 2001, 138, 83–89. [Google Scholar] [CrossRef]
- Gadaleta, A.; Nigro, D.; Marcotuli, I.; Giancaspro, A.; Giove, S.L.; Blanco, A. Isolation and characterisation of cytosolic glutamine synthetase (<i>GSe</i>) genes and association with grain protein content in durum wheat. Crop and Pasture Science 2014, 65, 38–45. [Google Scholar]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.)*. Journal of Experimental Botany 2007, 58, 2319–2327. [Google Scholar] [CrossRef]
- Ishiyama, K.; Inoue, E.; Watanabe-Takahashi, A.; Obara, M.; Yamaya, T.; Takahashi, H. Kinetic Properties and Ammonium-dependent Regulation of Cytosolic Isoenzymes of Glutamine Synthetase in <em>Arabidopsis</em>*. Journal of Biological Chemistry 2004, 279, 16598–16605. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.; Minagawa, H.; Yoshida, C.; Inoue, E.; Takahashi, H.; Ishiyama, K. Coregulation of glutamine synthetase1;2 (GLN1;2) and NADH-dependent glutamate synthase (GLT1) gene expression in Arabidopsis roots in response to ammonium supply. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Li, Q.; Liu, G.; Selvaraj, G.; Zheng, Z.; Zou, J.; Wei, Y. Roles of Cytosolic Glutamine Synthetases in Arabidopsis Development and Stress Responses. Plant and Cell Physiology 2019, 60, 657–671. [Google Scholar] [CrossRef]
- Yan, S.; Tang, Z.; Su, W.; Sun, W. Proteomic analysis of salt stress-responsive proteins in rice root. PROTEOMICS 2005, 5, 235–244. [Google Scholar] [CrossRef]
- Yousfi, S.; Márquez, A.J.; Betti, M.; Araus, J.L.; Serret, M.D. Gene expression and physiological responses to salinity and water stress of contrasting durum wheat genotypes. Journal of Integrative Plant Biology 2016, 58, 48–66. [Google Scholar] [CrossRef]
- Teixeira, J.; Pereira, S. High salinity and drought act on an organ-dependent manner on potato glutamine synthetase expression and accumulation. Environmental and Experimental Botany 2007, 60, 121–126. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, Y.; He, Q.; Li, H.; Zhang, X.; Zhang, F. Comparative proteomics illustrates the complexity of drought resistance mechanisms in two wheat (Triticum aestivum L.) cultivars under dehydration and rehydration. BMC Plant Biology 2016, 16, 188. [Google Scholar] [CrossRef]
- Nagy, Z.; Németh, E.; Guóth, A.; Bona, L.; Wodala, B.; Pécsváradi, A. Metabolic indicators of drought stress tolerance in wheat: Glutamine synthetase isoenzymes and Rubisco. Plant Physiology and Biochemistry 2013, 67, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Chien, H.-F.; Lin, C.C.; Wang, J.-W.; Chen, C.T.; Kao, C.H. Changes in ammonium ion content and glutamine synthetase activity in rice leaves caused by excess cadmium are a consequence of oxidative damage. Plant Growth Regulation 2002, 36, 41–47. [Google Scholar] [CrossRef]
- Balestrasse, K.B.; Benavides, M.P.; Gallego, S.M.; Tomaro, M.L. Effect of cadmium stress on nitrogen metabolism in nodules and roots of soybean plants. Functional Plant Biology 2003, 30, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Chaffei, C.; Pageau, K.; Suzuki, A.; Gouia, H.; Ghorbel, M.H.; Masclaux-Daubresse, C. Cadmium Toxicity Induced Changes in Nitrogen Management in Lycopersicon esculentum Leading to a Metabolic Safeguard Through an Amino Acid Storage Strategy. Plant and Cell Physiology 2004, 45, 1681–1693. [Google Scholar] [CrossRef] [PubMed]
- Rana, N.K.; Mohanpuria, P.; Yadav, S.K. Expression of tea cytosolic glutamine synthetase is tissue specific and induced by cadmium and salt stress. Biologia Plantarum 2008, 52, 361–364. [Google Scholar] [CrossRef]
- Sedri, M.H.; Amini, A.; Golchin, A. Evaluation of Nitrogen Effects on Yield and Drought Tolerance of Rainfed Wheat using Drought Stress Indices. Journal of Crop Science and Biotechnology 2019, 22, 235–242. [Google Scholar] [CrossRef]
- Song, J.; Wang, Y.; Pan, Y.; Pang, J.; Zhang, X.; Fan, J.; Zhang, Y. The influence of nitrogen availability on anatomical and physiological responses of Populus alba × P. glandulosa to drought stress. BMC Plant Biology 2019, 19, 63. [Google Scholar] [CrossRef]
- Xu, N.; Guo, W.; Liu, J.; Du, N.; Wang, R. Increased nitrogen deposition alleviated the adverse effects of drought stress on Quercus variabilis and Quercus mongolica seedlings. Acta Physiologiae Plantarum 2015, 37, 107. [Google Scholar] [CrossRef]
- Gessler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought-induced tree mortality and recovery. New Phytologist 2017, 214, 513–520. [Google Scholar] [CrossRef]
- Li, S.; Zhou, L.; Addo-Danso, S.D.; Ding, G.; Sun, M.; Wu, S.; Lin, S. Nitrogen supply enhances the physiological resistance of Chinese fir plantlets under polyethylene glycol (PEG)-induced drought stress. Scientific Reports 2020, 10, 7509. [Google Scholar] [CrossRef]
- Safavi-Rizi, V.; Uellendahl, K.; Öhrlein, B.; Safavi-Rizi, H.; Stöhr, C. Cross-stress tolerance: Mild nitrogen (N) deficiency effects on drought stress response of tomato (Solanum lycopersicum L.). Plant-Environment Interactions 2021, 2, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Cao, X.; Hu, J.; Zhu, L.; Zhang, J.; Huang, J.; Jin, Q. Nitrogen Metabolism in Adaptation of Photosynthesis to Water Stress in Rice Grown under Different Nitrogen Levels. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiology and Biochemistry 2016, 109, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Tan, D.K.Y.; Afridi, M.Z.; Luo, H.; Tung, S.A.; Ajab, M.; Fahad, S. Nitrogen fertility and abiotic stresses management in cotton crop: a review. Environmental Science and Pollution Research 2017, 24, 14551–14566. [Google Scholar] [CrossRef]
- Diao, M.; Ma, L.; Wang, J.; Cui, J.; Fu, A.; Liu, H.-y. Selenium Promotes the Growth and Photosynthesis of Tomato Seedlings Under Salt Stress by Enhancing Chloroplast Antioxidant Defense System. Journal of Plant Growth Regulation 2014, 33, 671–682. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, B.; Zhang, H.-J.; Weeda, S.; Yang, C.; Yang, Z.-C.; Ren, S.; Guo, Y.-D. Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). Journal of Pineal Research 2013, 54, 15–23. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.-G.; Yun, B.-W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environmental and Experimental Botany 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Pissolato, M.D.; Silveira, N.M.; Prataviera, P.J.C.; Machado, E.C.; Seabra, A.B.; Pelegrino, M.T.; Sodek, L.; Ribeiro, R.V. Enhanced Nitric Oxide Synthesis Through Nitrate Supply Improves Drought Tolerance of Sugarcane Plants. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef]
- Procházková, D.; Sumaira, J.; Wilhelmová, N.a.; Pavlíková, D.; Száková, J. Chapter 11 - Reactive Nitrogen Species and the Role of NO in Abiotic Stress. In Emerging Technologies and Management of Crop Stress Tolerance; Ahmad, P., Rasool, S., Eds.; Academic Press: San Diego, 2014; pp. 249–266. [Google Scholar] [CrossRef]
- Simontacchi, M.; Galatro, A.; Ramos-Artuso, F.; Santa-María, G.E. Plant Survival in a Changing Environment: The Role of Nitric Oxide in Plant Responses to Abiotic Stress. Frontiers in Plant Science 2015, 6. [Google Scholar] [CrossRef]
- Lau, S.-E.; Hamdan, M.F.; Pua, T.-L.; Saidi, N.B.; Tan, B.C. Plant Nitric Oxide Signaling under Drought Stress. Plants 2021, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Jiang, M.; Zhang, J.; Ding, H.; Xu, S.; Hu, X.; Tan, M. Nitric oxide induced by hydrogen peroxide mediates abscisic acid-induced activation of the mitogen-activated protein kinase cascade involved in antioxidant defense in maize leaves. New Phytologist 2007, 175, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Wu, X.; Zhong, Y. Exogenously Applied Nitric Oxide Enhances the Drought Tolerance in Hulless Barley. Plant Production Science 2015, 18, 52–56. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M.; El-Sheikh, M.A.; Alatar, A.A.; Zhang, L. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biology 2019, 19, 479. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, J.; Zhou, G.; Li, J.; Qian, C.; Lin, G.; Li, Y.; Zuo, Q. Moderate nitrogen application improved salt tolerance by enhancing photosynthesis, antioxidants, and osmotic adjustment in rapeseed (Brassica napus L.). Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef]
- Nazir, F.; Mahajan, M.; Khatoon, S.; Albaqami, M.; Ashfaque, F.; Chhillar, H.; Chopra, P.; Khan, M.I.R. Sustaining nitrogen dynamics: A critical aspect for improving salt tolerance in plants. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef]
- Han, M.-L.; Lv, Q.-Y.; Zhang, J.; Wang, T.; Zhang, C.-X.; Tan, R.-J.; Wang, Y.-L.; Zhong, L.-Y.; Gao, Y.-Q.; Chao, Z.-F.; et al. Decreasing nitrogen assimilation under drought stress by suppressing DST-mediated activation of Nitrate Reductase 1.2 in rice. Molecular Plant 2022, 15, 167–178. [Google Scholar] [CrossRef]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.M.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiology and Biochemistry 2019, 139, 171–178. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, Y.; Liu, X.; Korpelainen, H.; Li, C. Ammonium and nitrate affect sexually different responses to salt stress in Populus cathayana. Physiologia Plantarum 2022, 174, e13626. [Google Scholar] [CrossRef]
- Qu, C.; Liu, C.; Ze, Y.; Gong, X.; Hong, M.; Wang, L.; Hong, F. Inhibition of Nitrogen and Photosynthetic Carbon Assimilation of Maize Seedlings by Exposure to a Combination of Salt Stress and Potassium-Deficient Stress. Biological Trace Element Research 2011, 144, 1159–1174. [Google Scholar] [CrossRef]
- Akram, M. Effect of Nitrogen Nutrition on Solute Accumulation and Ion Contents of Maize under Sodium Chloride Stress. Communications in Soil Science and Plant Analysis 2014, 45, 86–100. [Google Scholar] [CrossRef]
- Zhao, F.G.; Qin, P. Protective effect of exogenous polyamines on root tonoplast function against salt stress in barley seedlings. Plant Growth Regulation 2004, 42, 97–103. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnology Reports 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Kapoor, D.; Singh, S.; Gautam, V.; Dhanjal, D.S.; Jan, S.; Ramamurthy, P.C.; Prasad, R.; Singh, J. Nitric Oxide: A Ubiquitous Signal Molecule for Enhancing Plant Tolerance to Salinity Stress and Their Molecular Mechanisms. Journal of Plant Growth Regulation 2021, 40, 2329–2341. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd_Allah, E.F.; Gucel, S.; Tran, L.-S.P. Nitric Oxide Mitigates Salt Stress by Regulating Levels of Osmolytes and Antioxidant Enzymes in Chickpea. Frontiers in Plant Science 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Liu, W.; Jing, Q.; Cao, W. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity. Environmental and Experimental Botany 2009, 67, 222–227. [Google Scholar] [CrossRef]
- Lopez, G.; Ahmadi, S.H.; Amelung, W.; Athmann, M.; Ewert, F.; Gaiser, T.; Gocke, M.I.; Kautz, T.; Postma, J.; Rachmilevitch, S.; et al. Nutrient deficiency effects on root architecture and root-to-shoot ratio in arable crops. Frontiers in Plant Science 2023, 13. [Google Scholar] [CrossRef]
- Falcioni, R.; Moriwaki, T.; Rodrigues, M.; de Oliveira, K.M.; Furlanetto, R.H.; dos Reis, A.S.; dos Santos, G.L.A.A.; Mendonça, W.A.; Crusiol, L.G.T.; Gonçalves, J.V.F.; et al. Nutrient deficiency lowers photochemical and carboxylation efficiency in tobacco. Theoretical and Experimental Plant Physiology 2023, 35, 81–97. [Google Scholar] [CrossRef]
- Sattelmacher, B. Addiscott, TM, AP whitmore and DS powlson: Farming, fertilizers and the nitrate problem. CAB international, 1991; 176 seiten. Paperback,£ 12.95. ISBN 0 85198 658 7. Wiley Online Library: 1993.
- Aparicio, V.; Costa, J.L.; Zamora, M. Nitrate leaching assessment in a long-term experiment under supplementary irrigation in humid Argentina. Agricultural water management 2008, 95, 1361–1372. [Google Scholar] [CrossRef]
- Mary, H. Ward. Too Much of a Good Thing? Nitrate from Nitrogen Fertilizers and Cancer. Reviews on Environmental Health 2009, 24, 357–363. [Google Scholar] [CrossRef]
- Ju, X.T.; Kou, C.L.; Zhang, F.S.; Christie, P. Nitrogen balance and groundwater nitrate contamination: Comparison among three intensive cropping systems on the North China Plain. Environmental Pollution 2006, 143, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Maghanga, J.K.; Kituyi, J.L.; Kisinyo, P.O.; Ng’etich, W.K. Impact of Nitrogen Fertilizer Applications on Surface Water Nitrate Levels within a Kenyan Tea Plantation. Journal of Chemistry 2013, 2013, 196516. [Google Scholar] [CrossRef]
- Bryan, N.S.; van Grinsven, H. Chapter Three - The Role of Nitrate in Human Health. In Advances in Agronomy, Sparks, D.L., Ed. Academic Press: 2013; Vol. 119, pp. 153–182.
- FRASER, P.; CHILVERS, C.; BERAL, V.; HILL, M.J. Nitrata and Human Cancer: A Review of the Evidence. International Journal of Epidemiology 1980, 9, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Knobeloch, L.; Salna, B.; Hogan, A.; Postle, J.; Anderson, H. Blue babies and nitrate-contaminated well water. Environmental Health Perspectives 2000, 108, 675–678. [Google Scholar] [CrossRef]
- Maghanga, J.; Kituyi, J.; Kisinyo, P.; Ng’Etich, W. Impact of nitrogen fertilizer applications on surface water nitrate levels within a Kenyan tea plantation. Journal of chemistry 2013, 2013. [Google Scholar] [CrossRef]
- Díaz-Rodríguez, A.M.; Salcedo Gastelum, L.A.; Félix Pablos, C.M.; Parra-Cota, F.I.; Santoyo, G.; Puente, M.L.; Bhattacharya, D.; Mukherjee, J.; de los Santos-Villalobos, S. The current and future role of microbial culture collections in food security worldwide. Frontiers in Sustainable Food Systems 2021, 4, 614739. [Google Scholar] [CrossRef]
- Fischer, R.A.; Connor, D.J. Issues for cropping and agricultural science in the next 20 years. Field Crops Research 2018, 222, 121–142. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, X.; Wu, K.; Fu, X. Nitrogen signaling and use efficiency in plants: what's new? Current Opinion in Plant Biology 2015, 27, 192–198. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annual Review of Plant Biology 2012, 63, 153–182. [Google Scholar] [CrossRef]
- Motavalli, P.P.; Goyne, K.W.; Udawatta, R.P. Environmental Impacts of Enhanced-Efficiency Nitrogen Fertilizers. Crop Management 2008, 7, 1–15. [Google Scholar] [CrossRef]
- Woods, J.; Williams, A.; Hughes, J.K.; Black, M.; Murphy, R. Energy and the food system. Philosophical Transactions of the Royal Society B: Biological Sciences 2010, 365, 2991–3006. [Google Scholar] [CrossRef] [PubMed]
- Ntatsi, G.; Karkanis, A.; Yfantopoulos, D.; Pappa, V.; Konosonoka, I.H.; Travlos, I.; Bilalis, D.; Bebeli, P.; Savvas, D. Evaluation of the field performance, nitrogen fixation efficiency and competitive ability of pea landraces grown under organic and conventional farming systems. Archives of Agronomy and Soil Science 2019, 65, 294–307. [Google Scholar] [CrossRef]
- Van Deynze, A.; Zamora, P.; Delaux, P.-M.; Heitmann, C.; Jayaraman, D.; Rajasekar, S.; Graham, D.; Maeda, J.; Gibson, D.; Schwartz, K.D.; et al. Nitrogen fixation in a landrace of maize is supported by a mucilage-associated diazotrophic microbiota. PLOS Biology 2018, 16, e2006352. [Google Scholar] [CrossRef]
- Santi, C.; Bogusz, D.; Franche, C. Biological nitrogen fixation in non-legume plants. Annals of Botany 2013, 111, 743–767. [Google Scholar] [CrossRef]
- Mus, F.; Crook, M.B.; Garcia, K.; Costas, A.G.; Geddes, B.A.; Kouri, E.D.; Paramasivan, P.; Ryu, M.-H.; Oldroyd, G.E.D.; Poole, P.S.; et al. Symbiotic Nitrogen Fixation and the Challenges to Its Extension to Nonlegumes. Applied and Environmental Microbiology 2016, 82, 3698–3710. [Google Scholar] [CrossRef] [PubMed]
- Barnawal, D.; Maji, D.; Bharti, N.; Chanotiya, C.S.; Kalra, A. ACC Deaminase-Containing Bacillus subtilis Reduces Stress Ethylene-Induced Damage and Improves Mycorrhizal Colonization and Rhizobial Nodulation in Trigonella foenum-graecum Under Drought Stress. Journal of Plant Growth Regulation 2013, 32, 809–822. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.S.; Saxena, A.K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatalysis and Agricultural Biotechnology 2020, 23, 101487. [Google Scholar] [CrossRef]
- Wen, A.; Havens, K.L.; Bloch, S.E.; Shah, N.; Higgins, D.A.; Davis-Richardson, A.G.; Sharon, J.; Rezaei, F.; Mohiti-Asli, M.; Johnson, A.; et al. Enabling Biological Nitrogen Fixation for Cereal Crops in Fertilized Fields. ACS Synthetic Biology 2021, 10, 3264–3277. [Google Scholar] [CrossRef]
- Compant, S.; Cambon, M.C.; Vacher, C.; Mitter, B.; Samad, A.; Sessitsch, A. The plant endosphere world – bacterial life within plants. Environmental Microbiology 2021, 23, 1812–1829. [Google Scholar] [CrossRef]
- Bacic, A.; Moody, S.F.; Clarke, A.E. Structural Analysis of Secreted Root Slime from Maize (Zea mays L.). Plant Physiology 1986, 80, 771–777. [Google Scholar] [CrossRef]
- Galloway, A.F.; Akhtar, J.; Marcus, S.E.; Fletcher, N.; Field, K.; Knox, P. Cereal root exudates contain highly structurally complex polysaccharides with soil-binding properties. The Plant Journal 2020, 103, 1666–1678. [Google Scholar] [CrossRef] [PubMed]
- Okon, Y.; Heytler, P.G.; Hardy, R.W.F. N<sub>2</sub> Fixation by <i>Azospirillum brasilense</i> and Its Incorporation into Host <i>Setaria italica</i>. Applied and Environmental Microbiology 1983, 46, 694–697. [Google Scholar] [CrossRef]
- Pankievicz, V.C.S.; do Amaral, F.P.; Santos, K.F.D.N.; Agtuca, B.; Xu, Y.; Schueller, M.J.; Arisi, A.C.M.; Steffens, M.B.R.; de Souza, E.M.; Pedrosa, F.O.; et al. Robust biological nitrogen fixation in a model grass–bacterial association. The Plant Journal 2015, 81, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.A.; Holz, M.; Woche, S.K.; Bachmann, J.; Carminati, A. Effect of soil drying on mucilage exudation and its water repellency: a new method to collect mucilage. Journal of Plant Nutrition and Soil Science 2015, 178, 821–824. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root exudation of phytochemicals in Arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PloS one 2013, 8, e55731. [Google Scholar] [CrossRef]
- Rane, N.R.; Tapase, S.; Kanojia, A.; Watharkar, A.; Salama, E.-S.; Jang, M.; Kumar Yadav, K.; Amin, M.A.; Cabral-Pinto, M.M.S.; Jadhav, J.P.; et al. Molecular insights into plant–microbe interactions for sustainable remediation of contaminated environment. Bioresource Technology 2022, 344, 126246. [Google Scholar] [CrossRef]
- Dubois, O. The state of the world's land and water resources for food and agriculture: managing systems at risk; Earthscan: 2011.
- Francis, B.; Aravindakumar, C.T.; Brewer, P.B.; Simon, S. Plant nutrient stress adaptation: A prospect for fertilizer limited agriculture. Environmental and Experimental Botany 2023, 213, 105431. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O. Biofertilizers and sustainable agriculture: exploring arbuscular mycorrhizal fungi. Applied Microbiology and Biotechnology 2017, 101, 4871–4881. [Google Scholar] [CrossRef]
- Meena, V.S.; Meena, S.K.; Verma, J.P.; Kumar, A.; Aeron, A.; Mishra, P.K.; Bisht, J.K.; Pattanayak, A.; Naveed, M.; Dotaniya, M.L. Plant beneficial rhizospheric microorganism (PBRM) strategies to improve nutrients use efficiency: A review. Ecological Engineering 2017, 107, 8–32. [Google Scholar] [CrossRef]
- Hartemink, A.E. The definition of soil since the early 1800s. Advances in Agronomy 2016, 137, 73–126. [Google Scholar]
- Campanhola, C.; Pandey, S. Sustainable food and agriculture: An integrated approach; Academic Press: 2018.
- Igiehon, N.O.; Babalola, O.O. Biofertilizers and sustainable agriculture: exploring arbuscular mycorrhizal fungi. Applied microbiology and biotechnology 2017, 101, 4871–4881. [Google Scholar] [CrossRef] [PubMed]
- Bodelier, P.L. Toward understanding, managing, and protecting microbial ecosystems. Frontiers in microbiology 2011, 2, 80. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, A.; Bhattarai, B.; Pandey, S. Variation of soil microbial population in different soil horizons. J Microbiol Exp 2015, 2, 00044. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Zhang, P.; Trivedi, P.; Riera, N.; Wang, Y.; Liu, X.; Fan, G.; Tang, J.; Coletta-Filho, H.D. The structure and function of the global citrus rhizosphere microbiome. Nature communications 2018, 9, 4894. [Google Scholar] [CrossRef]
- Hu, J.; Wei, Z.; Friman, V.-P.; Gu, S.-h.; Wang, X.-f.; Eisenhauer, N.; Yang, T.-j.; Ma, J.; Shen, Q.-r.; Xu, Y.-c. Probiotic diversity enhances rhizosphere microbiome function and plant disease suppression. MBio 2016, 7, 10–1128. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS microbiology reviews 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Mitter, E.K.; Tosi, M.; Obregón, D.; Dunfield, K.E.; Germida, J.J. Rethinking crop nutrition in times of modern microbiology: innovative biofertilizer technologies. Frontiers in Sustainable Food Systems 2021, 5, 606815. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher-Jenull, J.; Ceccherini, M.T.; Pietramellara, G.; Renella, G.; Schloter, M. Beyond microbial diversity for predicting soil functions: A mini review. Pedosphere 2020, 30, 5–17. [Google Scholar] [CrossRef]
- Johansson, J.F.; Paul, L.R.; Finlay, R.D. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS microbiology ecology 2004, 48, 1–13. [Google Scholar] [CrossRef]
- Saccá, M.L.; Barra Caracciolo, A.; Di Lenola, M.; Grenni, P. Ecosystem services provided by soil microorganisms. Proceedings of Soil biological communities and ecosystem resilience; pp. 9–24.
- Nannipieri, P.; Ascher, J.; Ceccherini, M.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. European journal of soil science 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Cross, H.R. Land-based production of animal protein: impacts, efficiency, and sustainability. Annals of the New York Academy of Sciences 2014, 1328, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Orozco-Mosqueda, M.d.C.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: a review. Biocontrol Science and Technology 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Bais, H.P.; Park, S.-W.; Weir, T.L.; Callaway, R.M.; Vivanco, J.M. How plants communicate using the underground information superhighway. Trends in plant science 2004, 9, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Soil health and human nutrition. In The Soil-Human Health-Nexus, CRC Press: 2020; pp. 315–325.
- Brunelle, T.; Dumas, P.; Souty, F.; Dorin, B.; Nadaud, F. Evaluating the impact of rising fertilizer prices on crop yields. Agricultural economics 2015, 46, 653–666. [Google Scholar] [CrossRef]
- Samberg, L.H.; Gerber, J.S.; Ramankutty, N.; Herrero, M.; West, P.C. Subnational distribution of average farm size and smallholder contributions to global food production. Environmental Research Letters 2016, 11, 124010. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O. Rhizosphere microbiome modulators: contributions of nitrogen fixing bacteria towards sustainable agriculture. International journal of environmental research and public health 2018, 15, 574. [Google Scholar] [CrossRef]
- Masson-Boivin, C.; Giraud, E.; Perret, X.; Batut, J. Establishing nitrogen-fixing symbiosis with legumes: how many rhizobium recipes? Trends in microbiology 2009, 17, 458–466. [Google Scholar] [CrossRef]
- Mendoza-Suárez, M.; Andersen, S.U.; Poole, P.S.; Sánchez-Cañizares, C. Competition, nodule occupancy, and persistence of inoculant strains: key factors in the rhizobium-legume symbioses. Frontiers in plant science 2021, 12, 690567. [Google Scholar] [CrossRef]
- Holland, P.L. Introduction: Reactivity of Nitrogen from the Ground to the Atmosphere. Chemical reviews 2020, 120, 4919–4920. [Google Scholar] [CrossRef]
- Nyaga, J.W.; Njeru, E.M. Potential of native rhizobia to improve cowpea growth and production in semiarid regions of Kenya. Frontiers in Agronomy 2020, 2, 606293. [Google Scholar] [CrossRef]
- Bogino, P.C.; Oliva, M.D.l.M.; Sorroche, F.G.; Giordano, W. The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations. International Journal of Molecular Sciences 2013, 14, 15838–15859. [Google Scholar] [CrossRef]
- Cesco, S.; Mimmo, T.; Tonon, G.; Tomasi, N.; Pinton, R.; Terzano, R.; Neumann, G.; Weisskopf, L.; Renella, G.; Landi, L.; et al. Plant-borne flavonoids released into the rhizosphere: impact on soil bio-activities related to plant nutrition. A review. Biology and Fertility of Soils 2012, 48, 123–149. [Google Scholar] [CrossRef]
- Yamada, Y.; Hoshino, K.-i.; Ishikawa, T. The phylogeny of acetic acid bacteria based on the partial sequences of 16S ribosomal RNA: the elevation of the subgenus Gluconoacetobacter to the generic level. Bioscience, biotechnology, and biochemistry 1997, 61, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Tajima, H.; Cline, L.C.; Fong, R.Y.; Ottaviani, J.I.; Shapiro, H.Y.; Blumwald, E. Genetic modification of flavone biosynthesis in rice enhances biofilm formation of soil diazotrophic bacteria and biological nitrogen fixation. Plant Biotechnology Journal 2022, 20, 2135–2148. [Google Scholar] [CrossRef] [PubMed]
- Cesco, S.; Mimmo, T.; Tonon, G.; Tomasi, N.; Pinton, R.; Terzano, R.; Neumann, G.; Weisskopf, L.; Renella, G.; Landi, L. Plant-borne flavonoids released into the rhizosphere: impact on soil bio-activities related to plant nutrition. A review. Biology and Fertility of Soils 2012, 48, 123–149. [Google Scholar] [CrossRef]
- Zhang, J.; Subramanian, S.; Stacey, G.; Yu, O. Flavones and flavonols play distinct critical roles during nodulation of Medicago truncatula by Sinorhizobium meliloti. The Plant Journal 2009, 57, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xu, A.; Elmerich, C.; Ma, L.Z. Biofilm formation enables free-living nitrogen-fixing rhizobacteria to fix nitrogen under aerobic conditions. The ISME journal 2017, 11, 1602–1613. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: an emergent form of bacterial life. Nature Reviews Microbiology 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Young, J.M.; Leschine, S.B.; Reguera, G. Reversible control of biofilm formation by Cellulomonas spp. in response to nitrogen availability. Environmental microbiology 2012, 14, 594–604. [Google Scholar] [CrossRef]
- Pankievicz, V.; Irving, T.B.; Maia, L.G.; Ané, J.-M. Are we there yet? The long walk towards the development of efficient symbiotic associations between nitrogen-fixing bacteria and non-leguminous crops. BMC biology 2019, 17, 1–17. [Google Scholar] [CrossRef]
- Ike, A.; Sriprang, R.; Ono, H.; Murooka, Y.; Yamashita, M. Bioremediation of cadmium contaminated soil using symbiosis between leguminous plant and recombinant rhizobia with the MTL4 and the PCS genes. Chemosphere 2007, 66, 1670–1676. [Google Scholar] [CrossRef] [PubMed]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernández, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martínez-Romero, E. Nitrogen fixation in cereals. Frontiers in Microbiology 2018, 9, 1794. [Google Scholar] [CrossRef] [PubMed]
- Broughton, W.J.; Zhang, F.; Perret, X.; Staehelin, C. Signals exchanged between legumes and Rhizobium: agricultural uses and perspectives. Plant and Soil 2003, 252, 129–137. [Google Scholar] [CrossRef]
- Checcucci, A.; Marchetti, M. The rhizosphere talk show: The rhizobia on stage. Frontiers in Agronomy 2020, 2, 591494. [Google Scholar] [CrossRef]
- Hesselbjerg Christensen, J. Climate phenomena and their relevance for future regional climate change Climate Change 2013: The Physical Science Basis ed TF Stocker et al. Cambridge: Cambridge University Press: 2013.
- Christiansen, H.H.; Etzelmüller, B.; Isaksen, K.; Juliussen, H.; Farbrot, H.; Humlum, O.; Johansson, M.; Ingeman-Nielsen, T.; Kristensen, L.; Hjort, J. The thermal state of permafrost in the nordic area during the international polar year 2007–2009. Permafrost and Periglacial Processes 2010, 21, 156–181. [Google Scholar] [CrossRef]
- Romanovsky, V.E.; Drozdov, D.; Oberman, N.G.; Malkova, G.; Kholodov, A.L.; Marchenko, S.; Moskalenko, N.G.; Sergeev, D.; Ukraintseva, N.; Abramov, A. Thermal state of permafrost in Russia. Permafrost and Periglacial Processes 2010, 21, 136–155. [Google Scholar] [CrossRef]
- Grosse, G.; Goetz, S.; McGuire, A.D.; Romanovsky, V.E.; Schuur, E.A. Changing permafrost in a warming world and feedbacks to the Earth system. Environmental Research Letters 2016, 11, 040201. [Google Scholar] [CrossRef]
- Harden, J.W.; Koven, C.D.; Ping, C.L.; Hugelius, G.; David McGuire, A.; Camill, P.; Jorgenson, T.; Kuhry, P.; Michaelson, G.J.; O'Donnell, J.A. Field information links permafrost carbon to physical vulnerabilities of thawing. Geophysical Research Letters 2012, 39. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: how well do we understand the processes and their controls? Philosophical Transactions of the Royal Society B: Biological Sciences 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Kuypers, M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nature Reviews Microbiology 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Weidner, S.; Pühler, A.; Küster, H. Genomics insights into symbiotic nitrogen fixation. Current opinion in biotechnology 2003, 14, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.; Kennedy, C.; Kavanagh, E.; Goldberg, R.B.; Hanau, R. Nitrogen fixation gene (nifL) involved in oxygen regulation of nitrogenase synthesis in K. peumoniae. Nature 1981, 290, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nature Reviews Microbiology 2004, 2, 621–631. [Google Scholar] [CrossRef]
- Burgess, B.K.; Lowe, D.J. Mechanism of molybdenum nitrogenase. Chemical reviews 1996, 96, 2983–3012. [Google Scholar] [CrossRef]
- Bjornsson, R.; Delgado-Jaime, M.U.; Lima, F.A.; Sippel, D.; Schlesier, J.; Weyhermüller, T.; Einsle, O.; Neese, F.; DeBeer, S. Molybdenum L-Edge XAS Spectra of MoFe Nitrogenase. Zeitschrift für anorganische und allgemeine Chemie 2015, 641, 65–71. [Google Scholar] [CrossRef]
- Setubal, J.C.; Dos Santos, P.; Goldman, B.S.; Ertesvåg, H.; Espin, G.; Rubio, L.M.; Valla, S.; Almeida, N.F.; Balasubramanian, D.; Cromes, L. Genome sequence of Azotobacter vinelandii, an obligate aerobe specialized to support diverse anaerobic metabolic processes. Journal of bacteriology 2009, 191, 4534–4545. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Yang, J.; Dou, Y.; Chen, M.; Ping, S.; Peng, J.; Lu, W.; Zhang, W.; Yao, Z.; Li, H. Nitrogen fixation island and rhizosphere competence traits in the genome of root-associated Pseudomonas stutzeri A1501. Proceedings of the National Academy of Sciences 2008, 105, 7564–7569. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Bennasar, A.; Bosch, R.; García-Valdés, E.; Palleroni, N.J. Biology of Pseudomonas stutzeri. Microbiology and molecular biology reviews 2006, 70, 510–547. [Google Scholar] [CrossRef]
- Dingler, C.; Kuhla, J.; Wassink, H.; Oelze, J. Levels and activities of nitrogenase proteins in Azotobacter vinelandii grown at different dissolved oxygen concentrations. Journal of bacteriology 1988, 170, 2148–2152. [Google Scholar] [CrossRef]
- Desnoues, N.; Lin, M.; Guo, X.; Ma, L.; Carreño-Lopez, R.; Elmerich, C. Nitrogen fixation genetics and regulation in a Pseudomonas stutzeri strain associated with rice. Microbiology 2003, 149, 2251–2262. [Google Scholar] [CrossRef]
- Oelze, J. Respiratory protection of nitrogenase in Azotobacter species: is a widely held hypothesis unequivocally supported by experimental evidence? FEMS microbiology reviews 2000, 24, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Curatti, L.; Brown, C.S.; Ludden, P.W.; Rubio, L.M. Genes required for rapid expression of nitrogenase activity in Azotobacter vinelandii. Proceedings of the National Academy of Sciences 2005, 102, 6291–6296. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.; Poole, R.; Yates, M.; Kennedy, C. Cloning and mutagenesis of genes encoding the cytochrome bd terminal oxidase complex in Azotobacter vinelandii: mutants deficient in the cytochrome d complex are unable to fix nitrogen in air. Journal of bacteriology 1990, 172, 6010–6019. [Google Scholar] [CrossRef] [PubMed]
- Dedysh, S.N.; Ricke, P.; Liesack, W. NifH and NifD phylogenies: an evolutionary basis for understanding nitrogen fixation capabilities of methanotrophic bacteria. Microbiology 2004, 150, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.; Siefert, J.L.; Staples, C.R.; Blankenship, R.E. The natural history of nitrogen fixation. Molecular biology and evolution 2004, 21, 541–554. [Google Scholar] [CrossRef]
- Henson, B.J.; Watson, L.E.; Barnum, S.R. The evolutionary history of nitrogen fixation, as assessed by nif D. Journal of molecular evolution 2004, 58, 390–399. [Google Scholar]
Figure 1.
Secondary structure of nitrogen plant transporters families. A) NRT1 family is represented by 4OH3; B) alphafold model of Q9LMZ9 represents NRT2 family; C) NRT3 family is represented by the alphafold model of Q9FGS5; D) alphafold model of P54144 represents AMT1 family.
Figure 1.
Secondary structure of nitrogen plant transporters families. A) NRT1 family is represented by 4OH3; B) alphafold model of Q9LMZ9 represents NRT2 family; C) NRT3 family is represented by the alphafold model of Q9FGS5; D) alphafold model of P54144 represents AMT1 family.
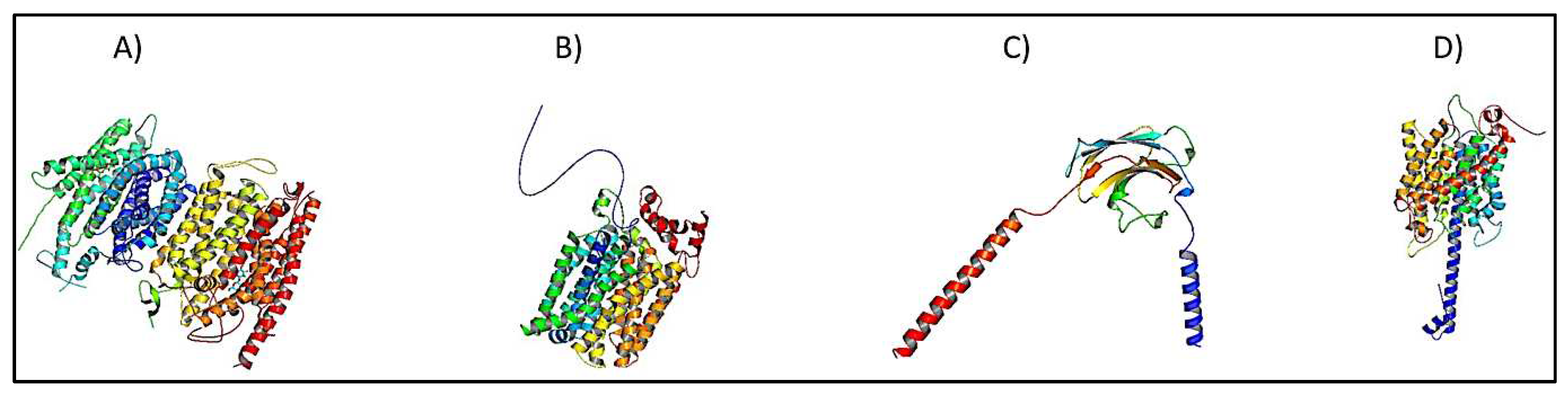
Figure 2.
Overview of nitrogen metabolism pathways. An illustration shows all the genes, enzymes, and substrates as defined in the KEGG Pathway database. It comprises six sub-pathways: Nitrogen fixation, Assimilatory nitrate reduction, Dissimilatory nitrate reduction, Denitrification, Nitrification, Complete nitrification, comammox, and Anammox.
Figure 2.
Overview of nitrogen metabolism pathways. An illustration shows all the genes, enzymes, and substrates as defined in the KEGG Pathway database. It comprises six sub-pathways: Nitrogen fixation, Assimilatory nitrate reduction, Dissimilatory nitrate reduction, Denitrification, Nitrification, Complete nitrification, comammox, and Anammox.
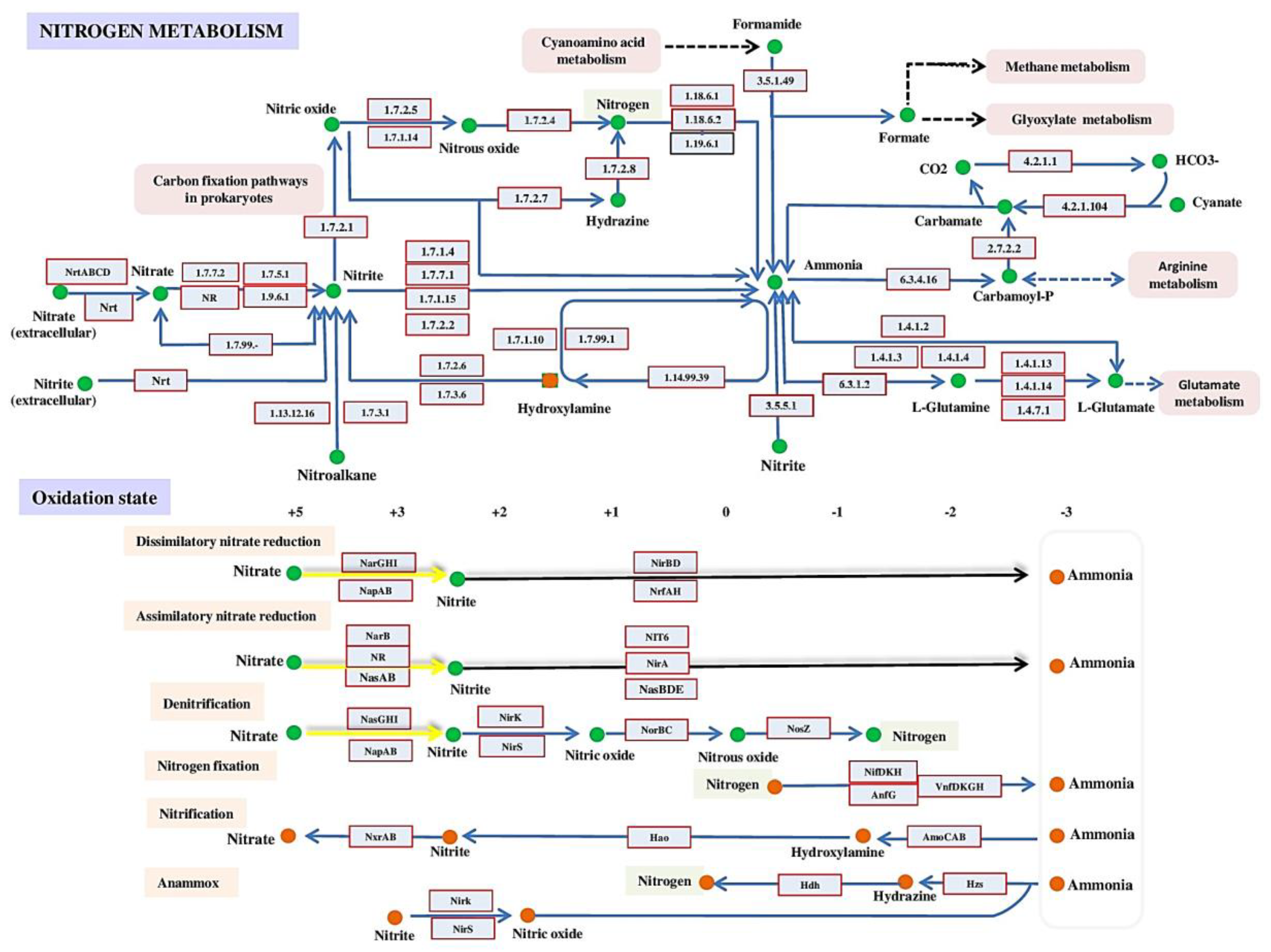
Figure 3.
The functional role of Glutamine synthetase (GS) isoforms and associated in response to abiotic stress. GS catalyzes the synthesis of glutamine from glutamate and ammonium ions and acts as a critical enzyme in the nitrogen metabolic pathway in organisms.
Figure 3.
The functional role of Glutamine synthetase (GS) isoforms and associated in response to abiotic stress. GS catalyzes the synthesis of glutamine from glutamate and ammonium ions and acts as a critical enzyme in the nitrogen metabolic pathway in organisms.
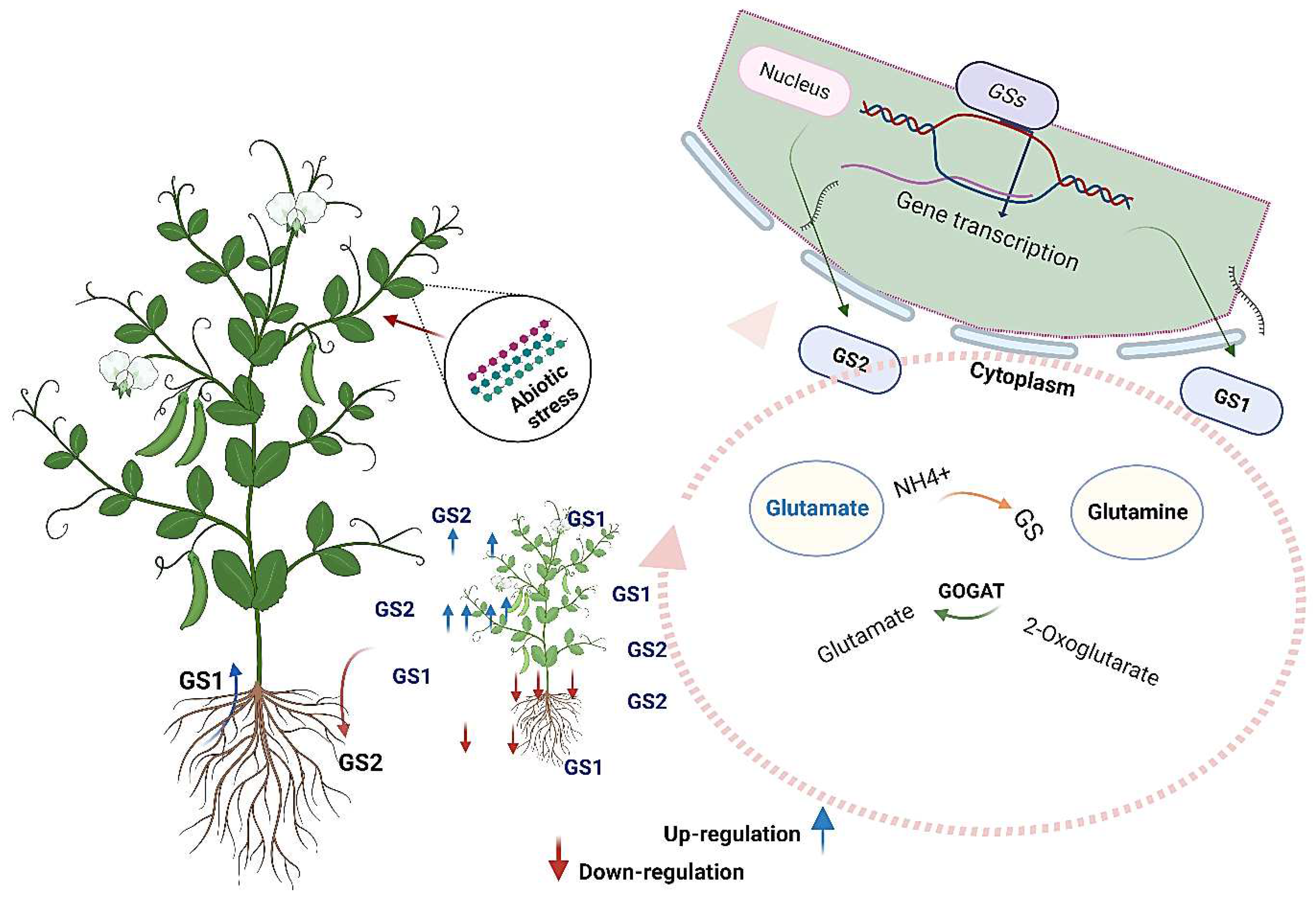
Figure 4.
Effect of drought stress on plant growth and activation of defense strategies by nitric oxide. When a plant is exposed to drought, it shows physiological and morphological changes to improve tolerance or avoid stress.
Figure 4.
Effect of drought stress on plant growth and activation of defense strategies by nitric oxide. When a plant is exposed to drought, it shows physiological and morphological changes to improve tolerance or avoid stress.
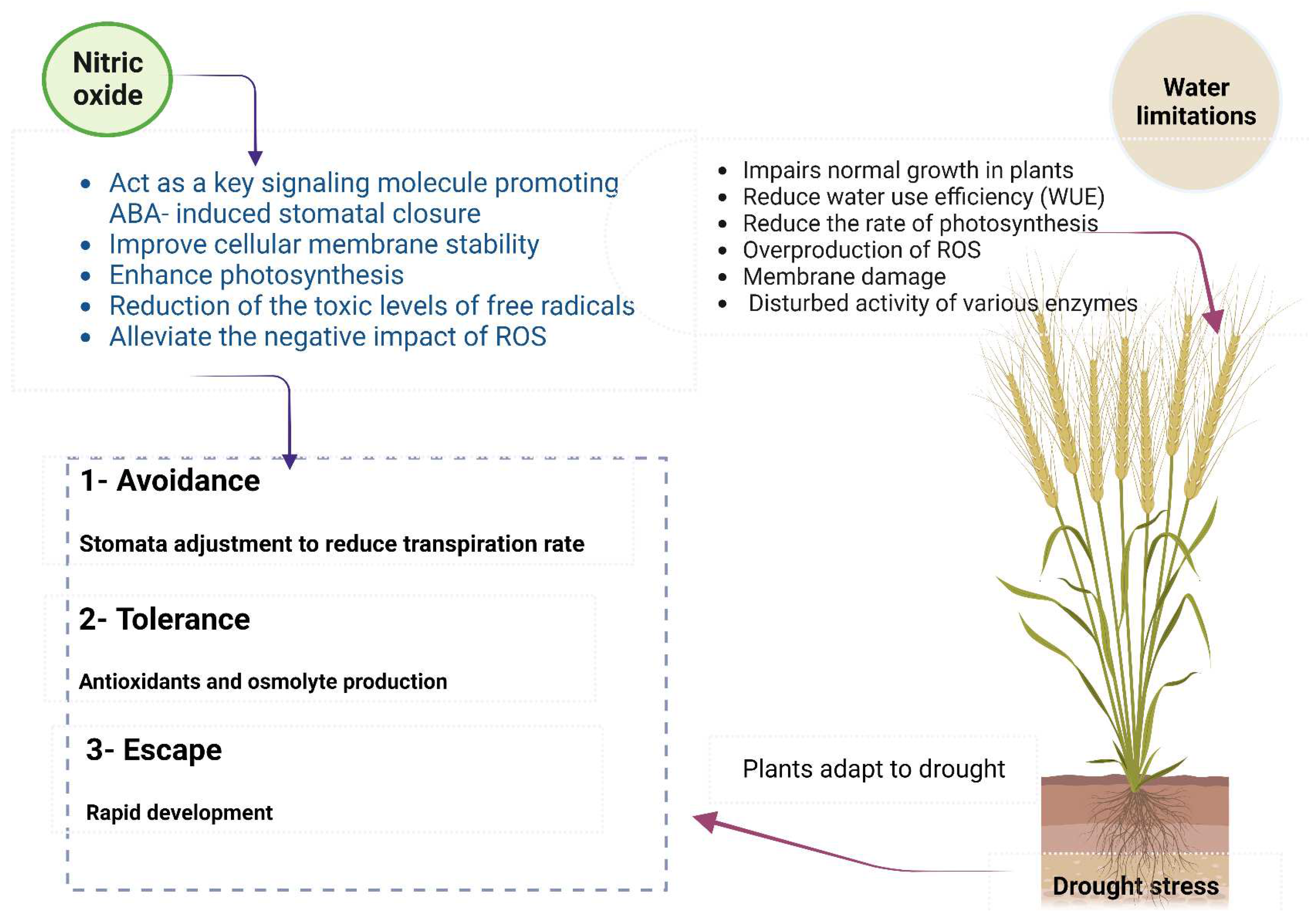
Figure 5.
A simplified model represents the microbial beneficial effects and the signal pathways involved in modulating the effects on nitrogen availability. Nitrogen source limitation also increases ammonium and nitrate uptake activity as a promising strategy to achieve the sustainability of agriculture. The availability of nitrogen is one of the critical elements in the process of biological nitrogen fixation that converts atmospheric nitrogen (N2) to ammonia (NH3) via a nodule structure.
Figure 5.
A simplified model represents the microbial beneficial effects and the signal pathways involved in modulating the effects on nitrogen availability. Nitrogen source limitation also increases ammonium and nitrate uptake activity as a promising strategy to achieve the sustainability of agriculture. The availability of nitrogen is one of the critical elements in the process of biological nitrogen fixation that converts atmospheric nitrogen (N2) to ammonia (NH3) via a nodule structure.
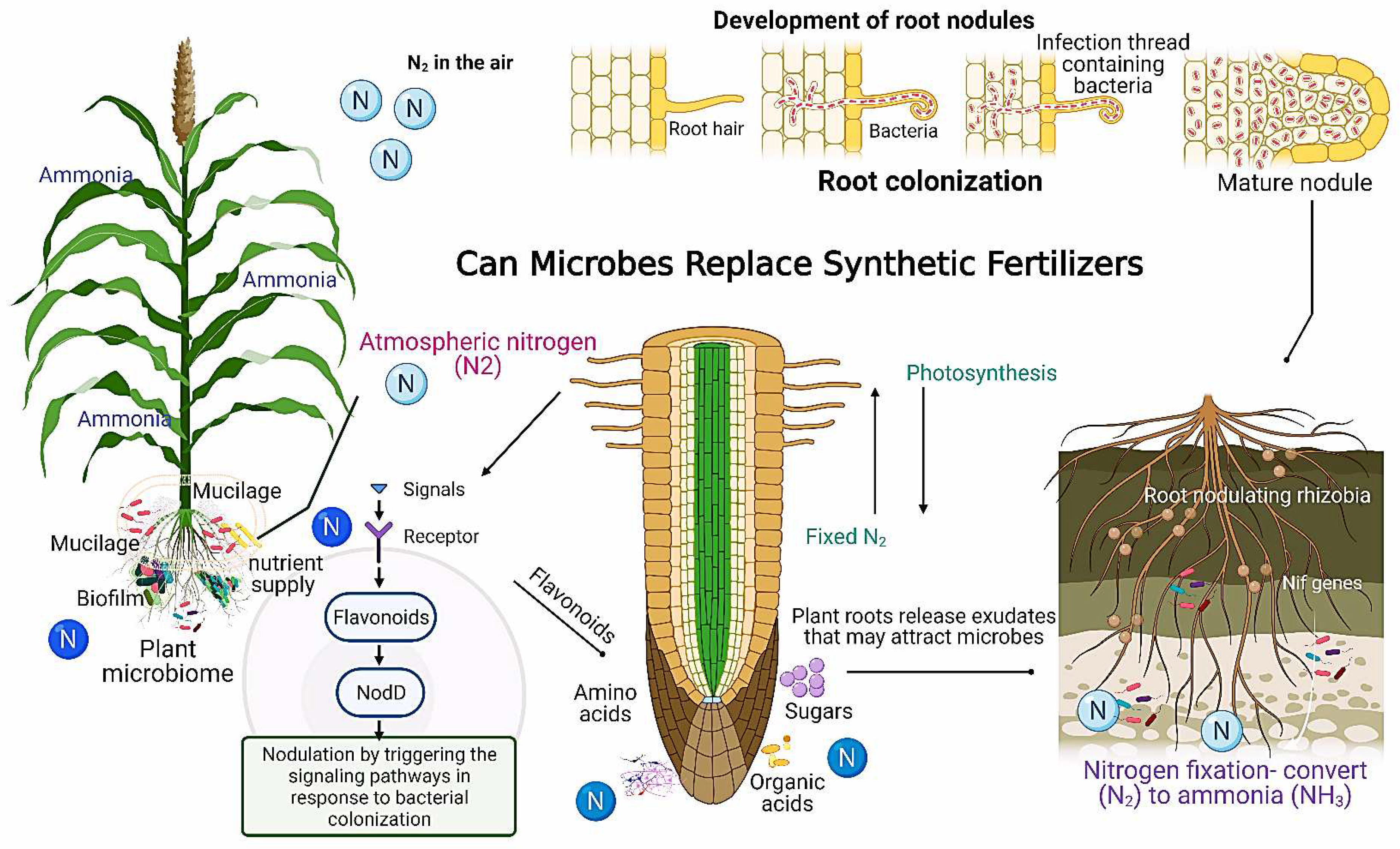
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



