Submitted:
22 August 2023
Posted:
23 August 2023
You are already at the latest version
Abstract
Grazing exclusion (GE) and rest grazing (RG) are important management to restore grassland ecosystems. In order to evaluate the effects and mechanisms of different grazing management on species diversity, vegetation community indices and soil variables were determined in the plots of Qilian Mountains in Gansu Province. The results showed that diversity effects and their regulating mechanisms had space-scale dependence under different grazing management. Species richness and species diversity indices of RG in grassland were significantly higher than that of GE at the regional scale. Additionally, three grazing management in mountain meadows had only a significant effect on species richness, but different management in mountain meadows and temperate steppes had a significantly different effect on species diversity indices. Meanwhile, soil variables only influenced species diversity at the regional scale. Most of community and soil variables at each scale had positive effects on species diversity, except that the direction of biodiversity effects was negative for species coverage, mean plant height, soil porosity (SK) and bulk density (BD) under two contrasting grazing management. In conclusion, choosing RG at the regional scale and select grazing management according to different grassland types at the local scale to restore degraded grassland vegetation.
Keywords:
Grazing exclusion
; rest grazing
; vegetation community
; scale effect
; species diversity
; soil characteristics
1. Introduction
Alpine grasslands cover above 60% of Qinghai-Tibetan Plateau and 25% of Chinese total territorial lands [1,2,3], which is an important location of livestock production. Owing to the vulnerability of the ecosystem and anthropological activity, alpine grasslands on the Qinghai-Tibetan Plateau has been severely degraded and caused dynamic change of ecosystem function by long-term grazing at multiple scales in recent decades, including plant diversity, vegetation biomass, soil properties, soil retention, water conservation [3,4]. Nowadays, a series of grazing management strategies were launched to protect and make sustainable use of grasslands in China [5,6,7], which included “Grazing for Green Program”, “Grassland Ecological Protection Program”, and “Return Grazing Land to Grassland Program” [8,9,10,11]. Numerous studies have considered different grazing management practices that mainly included GE, rotational grazing and RG (compared to no-grazing) to restore degraded grasslands [12,13], but to date there have been few comparisons between grazing regimes, especially on the biodiversity and natural habitats [14,15]. Thus, a better understanding of the effect of grazing management practices on biodiversity and behind mechanisms is needed for alpine grasslands restoration.
The effects of grazing management above-mentioned on plant diversity have not been conclusive, which are controversial [16,17,18]. Some studies indicated that GE increased plant diversity, whereas others reported a decrease or no change [19,20,21,22,23]. For example, Song et al. [24] reported that long-term GE reduced species diversity at four treatments in the alpine meadow ecosystem near the Haibei national field research station in China. Plant diversity was higher in GE site than that in the grazing sites in rangelands in Saral [25]. McDonald et al. [26] found that RG had different effects on plant diversity when compared to grazed areas or ungrazed areas. But to our knowledge, most of these studies consisted of site-specific controlled-grazing management experiments based on grazed-ungrazed contrasts (i.e. grazing versus enclosure, or rest grazing) [17,27], indicating that the effects of grazing regimes on plant diversity was spatial-scale dependence, which is important to understand the effect of grazing regimes on plant diversity at a larger scale (dependence) from a management strategy perspective.
The mechanisms that the effect of grazing management types on plant diversity have reported various and even contradictory results [18,27,28]. Previous studies have shown that the effects of grazing regimes on species diversity depended largely on local variables (site-specific) and regional variables (vegetation types and the duration of grazing exclusion) [17,29,30]. Previous studies reported that GE had varying effects on the plant diversity in different grasslands types [16,31]. For example, Schultz et al. [32] found that the effects of GE on plant species richness and phytomass accumulation varied across a regional productivity gradient. Species richness is scale-dependent [33,34,35], especially species-area relationship [36]. Li et al.[37] reported that moderate grazing was the possible reason for RG as a sustainable grazing management strategy. Most of these studies were restricted to specific vegetation types, or at single scale, but very few studies attempted to derive general conclusions especially at different spatial scales from grazing management strategy perspective [27,38,39].
The Qilian Mountains are important ecological security barrier in north-west China, where most grasslands have been degraded because of climate change, over-grazing and inadequate integrated management in the past [40,41]. In the present study, an investigation was conducted to evaluated the response of degraded grassland ecosystems to grazing management to address the following questions: (1) How have grazing management affected the vegetation and soil characteristics? (2) Are the effects of different grazing management (GE and RG) on grassland species diversity scale-dependent? (3) What are the regulatory mechanisms of grassland species diversity of different grazing management (GE and RG) at different spatial scales?
2. Materials and Methods
2.1. Study sites
The study was conducted in Qilian Mountains, in Tianzhu and Yongchang counties of Gansu Province (102°01′-103°40′ E and 36°30′-37°35′N), which is located in the arid and semi-arid desert region of Northwest China, at the intersection of Qinghai-Tibet Plateau, Inner Mongolia Plateau and Loess Plateau with elevation ranging from 2040 m to 4874 m above sea level (asl) [2]. The climate in the northern and southern is a temperate continental semi-arid and continental highland monsoon, respectively, which are separated by the Wushao Mountains with a total yearly solar radiation of 130 kcal/cm2, a maximum sunshine duration of 4,434 h and a frost-free season of approximately 130 days. The annual average temperature and precipitation range from 0.2℃ to 4℃ and from 260 mm to 500 mm respectively. The annual evaporation ranges from 1200 mm to 1700 mm.
At the study site, Tianzhu County and Yongchang County have cumulatively established mesh fencing with a total of 360,000 hm2 and 200,000 hm2 since 2018, respectively.
2.2. Experimental design
The experiment was established in 2019 to study the effects of grazing management on plant diversity. In this study, in a randomized block design, five grassland management types were included to make 4 experimental sites which was divided into 32 plots (33.3m×33.3 m) with 96 subplots (Table 1). Experimental sites included in Songshan Township with temperate steppe, in Zhuaxi Xiulong Township with alpine meadow and mountain meadow, in Haxi Township with alpine meadow and mountain meadow, and in Xinchengzi Township with mountain meadow. Five grazing management types were selected, including three GE types and two RG types. GE types included 10 -month, 4- year and a long-term (more than 20 years) grazing exclusion. The long-term grazing exclusion site was selected within the Haxi Nature Reserve. RG types included winter grazing and summer grazing (the duration of rest grazing was not specified).
2.3. Sampling and measurements
2.3.1. Vegetation sampling and measurements
Plant communities were surveyed within each plot using three 1 m × 1 m quadrats to measure plant species abundance and biomass (including aboveground biomass and belowground biomass). In each quadrat, plant species were identified, and the abundance of each species (clumps) was counted [42]. The height of each plant species in the subplot was investigated using a metal rod and a tape measure. We estimated plant community cover and the relative cover of each species. Aboveground biomass (AGB) was determined by clipping all living plant to ground level, oven dried at 65℃ for 48h to a constant weight, and then weighed. Belowground biomass was calculated by soil cores method as described below.
The species richness index (S), evenness index [Pielou index (E)] and diversity indices [Shannon-Wiener index (H') and Simpson index (D)] were calculated to reflect species diversity using the method described in Ma et al. [43].
where N is the total number of species, S is the number of species in the sample plot, and Pi is the ratio of the number of individuals of species i to the total number of individuals.
Richness index (S) = N
Shannon-Wiener index (H') = -∑ (Pi) (lnPi)
Pielou index (E) = H/lnS
2.3.2. Soil sampling and measurements
We collected soil samples (in 0-10cm, 10-20 cm and 20-30 cm) using a soil drill and soil cores (5cm diameter×5cm depth) to analyze soil physical-chemical properties. Soil cores were used to measure soil BD, SP, field capacity (FC) and belowground biomass. Other physical-chemical variables were calculated by soil in soil sampler. All the samples were stored in ice coolers and transported to a laboratory for analysis within 1 week.
In each plot, three cores in 0-10cm, 10-20 cm and 20-30 cm were set up for a total of 9 soil cores in each plot, respectively. Soil cores were kept 10 cm belowground to obtain the most roots since most species have shallow root system in the Qilian Mountains grassland. A soil column was immersed in water for 24 h and then carefully washed to obtain the roots of co-occurring species. Water content and BD were calculated by using the oven-dried weight of the soil sample at 105℃. SP was calculated by BD and specific gravity. Soil texture was analysed by laser diffraction (MALVERN Instruments, Mastersizer 2000) [44]. At the time of pot filling, moisture content of soil at field capacity was gravimetrically determined by watering the pots filled with soil to flooding and the pots were left for overnight. The air dry-weight of the soil samples and the wet weight after drainage over-night were recorded [45].
The soil organic carbon (SOC) content was determined via potassium dichromate-ferric sulfate titration. Soil total nitrogen content (TN) was determined using the semi-micro Kjeldahl method, and soil total phosphorus (TP) content was measured using the sulfuric acid-perchloric acid digestion-molybdenum antimony colorimetric method. Soil total potassium (TK) was measured using by hydrofluoric acid-perchloric acid digestion, flame photometric method. The above determination methods are the Soil and Agricultural Chemistry Analysis [46].
2.4. Statistical analysis
Various environmental variables especially soil variables in different layers were also measured at each individual plot and then averaged them when we used them.
In order to analyze the effect of different grazing management types on plant diversity at the regional scale in grassland, generalized linear mixed models (GLMM) were constructed using the lmer function in the lme4 package in R 4.2.1. The response variables, fixed variables and random variables in GLMM were species diversity indices, grazing management types and grassland types, respectively.
At the local scale, different grazing management types and grassland types were included as fixed variables in GLMM. However, as their interactions were not significant, indicating that the effect of different grazing management types on plant communities in grasslands were not dependent on grassland types. Therefore, one-way ANOVA followed by a post-hoc Tukey’s test and paired t test were used to analyze the effects of different grazing management types in the same grassland type on species diversity and plant community characteristics. There was only one grazing exclusion plot for alpine meadow, so it was not included in the comparison analysis of mean values for the relevant indicators.
All statistical analyses were performed using the SPSS 17.0 statistical software (SPSS Inc., Chicago, IL,USA), and graphs were created using the Sigma Plot 11.0 program (Systat Software Inc., San Jose, CA, USA).
3. Results
3.1. Responses of Different Grazing Management on Vegetation and Soil Characteristics
For vegetation community characteristics, two contrasting grazing management had no significant effect on community height, community coverage, and community biomass at different scales (P>0.05), except that community coverage in mountain meadows under the 10-month GE was merely significantly smaller than those under other two grazing management (P<0.05) (Table 2).
For soil characteristics, the two contrasting grazing management significantly affected SOC content and sand content (0.02-2mm) at the regional scale (P<0.05), but had different effects on soil variables for different grasslands types. The two contrasting grazing management significantly affected TN content and BD in temperate steppes (P<0.05) (Table 2). The SOC content of RG in mountain meadows was significantly higher than those of other two management (P<0.01). The TN content in mountain meadows under the 10-month GE was significantly smaller than those under other grazing management (P<0.01). The TK content of GE in mountain meadows was significantly smaller than those of other grazing management (P<0.05). FC and clay content (<0.002mm) of RG in mountain meadows were merely significantly different than that under the 10-month GE (P<0.05). It should be noted that the two contrasting grazing management had no significant effect (P>0.05) on other soil variables (Table 2).
3.2. Responses of Different Grazing management on Species Diversity
3.2.1. Species Richness
There were significant differences for species richness between GE and RG at the regional scale (df=28.82, t=2.02, P=0.05, R2=0.84), whereas different grazing management in mountain meadows had only significant effect on species richness (df=2, F=34.03, P=0). Specifically, species richness of GE in grasslands was significantly smaller than that of RG. Species richness of RG in grasslands was approximately 30.30% higher than that of GE. In addition, species richness in mountain meadows was significantly higher (P<0.05) under long-term GE than those under winter grazing (RG) and 10-month GE, followed by winter grazing, long-term GE, 10-month GE in a decreasing order (Table 2).
3.2.2. Diversity Indices
There were significant differences for species diversity between GE and RG at different scales (P<0.05). The Shannon-Weiner index of GE was significantly smaller (df=25.74, t=2.22, P=0.036, R2=0.71) than that of RG (Figure 1). The Simpson index of GE was significantly lower (df=27.65, t=2.36, P=0.026, R2=0.69) than that of RG (Figure 2). Additionally, grazing management in mountain meadows had significant effects on diversity indices (df=2, F=6.35, P=0.012). The Shannon-Weiner index and Simpson index in mountain meadows under the 10-month GE were both significantly smaller than those under other two grazing management (P<0.05). For temperate steppe, the Shannon-Weiner index and Simpson index of GE was both significantly lower than those of RG (Shannon Weiner index F=17.25, P=0.003; Simpson index F=8.32, P=0.02) (Table 2).
3.2.3. Evenness
Grazing management did not significantly affect the evenness in grasslands at different scales (P>0.05) (Table 2).
3.3. Regulatory Mechanisms of Different Grazing Management on Species Diversity
Our results showed that species diversity indices were mainly regulated by community coverage, above- and below-ground biomass, water-retention and nutrient variables in soil at different scales.
3.3.1. Regulatory Mechanisms of Community and Soil Characteristics on Species Richness
Specifically, a highly significant positive linear correlation of RG was found between community coverage and species richness (P<0.01) at the regional scale, even between community coverage and belowground biomass under GE (P<0.01). It should be noted that neither belowground biomass of RG nor aboveground biomass of GE was significantly associated with species richness (P=0.069 and 0.087, respectively) (Table 3). In addition, species richness increased with SOC、TN、FC and C/K of RG (P=0.001,0.003,0.027 and 0.002, respectively), and increased with TN、C/K and N/P of GE (P =0, 0.001, 0, respectively), but decreased with TK and BD of GE (P=0.026 and 0.047, respectively).
At the local scale, different community characteristics influence species richness in different grassland types under different grazing management. There was a highly significant positive relationship between community coverage and species richness in mountain meadows under GE (P<0.01), but a significant negative correlation between species coverage and species richness in temperate steppe under RG (P<0.05). Additionally, species richness was slight positively correlated with aboveground biomass of RG in temperate steppe (P=0.089) and in alpine meadows (P=0.078), but negatively correlated with community height and species coverage (0.05<P<0.1) (Table 4). It should be noted there were no relationships between species richness and other community characteristics in grasslands under grazing management (P>0.1).
3.3.2. Regulatory Mechanisms on Species Diversity by Community and Soil Characteristics
At the regional scale, the Shannon-Weiner index of RG was positively correlated with community coverage (P<0.01), but the Simpson index of RG was a slight correlated with community coverage under RG (P=0.069). Specifically, the Shannon-Weiner index and Simpson index of GE were positively correlated with both community coverage (P<0.01) and biomass (aboveground and belowground, P<0.05). At the same time, species diversity indices increased with SOC content、TP content、SP、 FC、C/N and C/K of RG (P=0.002, 0.005, 0, 0.001, 0.018 and 0.023 for Shannon-Wiener index, respectively; P=0.011, 0.011, 0, 0.004, 0.011, no data for CK for Simpson index, respectively). Species diversity increased with SOC content、TN content、C/P, C/K and N/P of GE (P=0.004, 0, 0.001, 0 and 0.013 for Shannon-Wiener index, respectively; P=0.004, 0.002, 0.001, 0.001, 0.044 for Simpson index, respectively), but decreased with BD of GE (P=0.006 and 0.006 for Shannon-Wiener index and Simpson index, respectively).
At the local scale, different community characteristics influenced species diversity indices in different grassland types under different grazing management. There was a significant positive and linear correlation of RG in alpine meadows between the aboveground biomass and both the Shannon-Weiner index and Simpson index (P<0.05). Specifically, the Shannon-Wiener index and Simpson index in mountain meadows were strongly associated with species coverage of RG and community coverage of GE (P<0.05). In addition, the Shannon-Wiener index was also significant correlated with community height of GE in mountain meadows (P<0.05). But species diversity was slight positively correlated with community coverage of RG in mountain meadows (P=0.092), community height in mountain meadows of GE (P=0.056), community height and species coverage of GE in temperate steppes (0.05<P<0.1). There were no relationships between species diversity and community characteristics in grasslands under different grazing management (P>0.1) (Table 4).
3.3.3. Regulatory Mechanisms on Evenness by Community Characteristics
At the regional scale, there was no significant relationship between community characteristics and the Pielou index under two contrasting grazing management (P>0.1). At the same time, the Pielou index increased with SP and C/N of RG in grasslands (P=0.005 and 0.019, respectively), and with SOC content of GE in grasslands (P=0.039), but decreased with N/P (P=0.04). At the local scale, different community characteristics affected the Pielou index in different grassland types under different grazing management. The Pielou index of RG was significant linear positive associated with aboveground biomass in alpine meadows (P<0.05), and species coverage in mountain meadows (P<0.05). In addition, the Pielou index was slightly correlated with community coverage of RG (P=0.086) in mountain meadows, aboveground biomass and species coverage of RG in temperate steppes (P=0.1), plant density per clump and species coverage of GE (0.05 < P < 0.1) in temperate steppes (Table 4). However, there was no relationship between community characteristics and the Pielou index in grasslands under different grazing management (P>0.1).
4. Discussion
4.1. Significant Effects of Different Grazing management on Species Diversity
Existing research results have shown that the effects of different grazing management on species diversity were controversial (i.e. increasing, decreasing, or no significant changes). In the present study, there were significant effects of two contrasting grazing management on species richness and diversity indices in grasslands at different scales but no effect on species evenness. Species richness and species diversity indices in grassland were higher under RG than under GE at the regional scale, which were consistent with previous studies in the Qilian Mountains [47] and Hulunbeir grasslands [48]. At the same time, our results showed that species richness and species diversity indices of RG only in mountain meadows and temperate steppes under rest grazing were greater than those under short-term grazing exclusion, which were consistent with previous studies reported [49,50,51,52]. The reason for this result may be that moderate grazing was beneficial to increase species diversity in degraded grasslands [53,54]. Additionally, RG was more suitable for nutritional growth, seed maturation of edible forage, the renewal and utilization of grassland resources and sustainable development of grassland, which supported the “intermediate disturbance” hypothesis [9,52]. Then RG in grasslands made community characteristics maintain stable, particularly utilize the growth attributes of grasslands without harming them. In particular, RG is usually seasonal, which reduces intense species competition. On the contrary, past studies reported that GE had negative impacts on species diversity and species evenness [55,56]. Generally speaking, GE allows pasture to rest, regenerate, and regrow, which resulted in competition among species for limited resources such as light and nutrients. As reported in previous studies, appropriate grazing disturbance can increase species diversity in grasslands, while GE may lead to a decrease in species diversity [57,58]. However, GE may make plant community structures uniform and lead to a decrease in community anti-interference ability. Also, our findings about RG are contrary to some past findings [58] and Bai et al. [50]. This may be due to the differences of selecting season for rest grazing [13], grazing intensity influencing grassland vegetation [51], and other environmental conditions [59].
It should be noted that there were no significant differences in species richness and species diversity indices between long-term grazing exclusion and RG in mountain meadows at the local scale. This is consistent with the findings of [53] and [60] in terms of species richness. As mentioned earlier, that could be because the grazing period of RG in mountain meadows was approximately 9 months under only winter grazing management that caused high grazing intensity. High grazing intensity would offset the positive benefits from rest grazing, resulting in species diversity indices being consistent with long-term grazing exclusion results. Meanwhile, long-term grazing exclusion and RG both increased the amounts of taller plants in grasslands in our study that mainly included Ranunculus tanguticus, Cirsium souliei, Poa annua, and Polygonum viviparum under long-term grazing exclusion, and Elymus nutans, Oxytropis ochrocephala, Polygonum viviparum, Poa annua, and Medicago archiducis-nicolai under rest grazing. These resulted in decreasing space for short plant to grow. Therefore, the differences in species diversity between two contrasting grazing management were not significant after a certain period of GE or RG.
4.2. Regulatory Mechanisms of Scale-effect on Species Diversity by Different Grazing management
In an era of rapid species extinction, exploring the regulatory mechanisms of species diversity under different grazing management is an urgent issue. Past studies found that the decline of species diversity in grasslands is associated with factors such as nutrient overloading and grazing [61,62]. Our study showed that species diversity under different grazing management at multiple scales had different regulatory mechanisms in grasslands of the Qilian Mountains. Positive and negative interactions between species diversity and their regulating factors that represent facilitation and competition respectively occurred simultaneously in our study. Particularly, negative diversity effects suggested that resource competition is more prevalent than niche complementarity at small scale. However, at large scale competition pressure may be reduced and increased facilitation thus occurring positive diversity effects.
In the current study, soil variables only affected species diversity at the regional scale under two contrasting grazing management. That could be attributed to the fact that heterogeneous distribution of soil condition at broad scale may cause community patches and thus influence species diversity, but the nonsignificant effects of soil variables at local scale may be homogeneous soil condition in our sampling sites. In terms of soil physics, negative diversity effect about BD under GE may be that GE increased species diversity, resulting in increasing soil water storage, consequently low BD [63,64]. For example, earlier studies have reported that grazing increased BD, resulting in reducing SP [63,64,65], but RG kept BD in alpine meadows [8]. It should be noted that positive diversity effect about FC and SP under RG occurred as above-mentioned, which was in accordance with other findings reported [63,66,67]. Likewise, from soil nutrients perspective, the same regulating factors of species diversity under two contrasting managements were SOC content and TN content, while the regulating factors under RG and GE were TP content and TK content, respectively. Positive diversity effect about soil nutrients under two contrasting grazing management was similar with other studies stated [68], which could be due to the fact that high levels of soil nutrients support plant growth and further promote species diversity under two grazing management, and heterogeneous distribution of soil nutrients as above-mentioned. SOC content and TN content play vital roles in plant growth, therefore increased species diversity [17,63]. Dong et al. [18] stated that RG notably increased TP content owing to excreting into by herbivores in the form of dung and urine [63,66]. Besides, our result shown negative richness effect, which may be due to the fact that herbivores didn’t excrete into dung and urine under GE [66]. In line with our results, past findings found higher TK content in grazed areas as compared to in protected areas, indicating that livestock trampling and from droppings can accumulated potassium [69,70].
Regrading to soil stoichiometric ratios, the same regulating factor of species diversity under two contrasting managements was C/K, while the regulating factors were C/N of RG, N/P and C/P of GE, respectively. Firstly, some studies reported that C/K had positive correlation with SOC content [13], then both affecting species diversity. On the other hand, C/N under RG were positively correlated with species diversity. The probable explanation was due to the accumulation of plant litter that alleviated species diversity and promoted the decomposition rate, consequently lower C/N [13,52]. It is widely acknowledged that soil C/N reflects the decomposition rate of soil organic matter and a lower C/N ratio with a faster decomposition rate. Compared to RG, GE had a lower mineralization and species diversity because of the lower decomposition rate of large litter, thereby resulting lower C/P and N/P [66].
With respect to the plant community, vegetation community characteristics influenced species diversity at each scale with different indicators. Only at the small scale, species diversity indices were negatively associated with mean plant height for GE and species coverage for RG in mountain meadows except species coverage also negatively regulated Pielou index for RG in mountain meadows, but species coverage was also negatively correlated with species richness under RG in temperate steppes in our study. Conversely, aboveground biomass was positively correlated with species diversity and evenness (Pielou index) in alpine meadows under RG. One possible reason for our results is that small scale can lead to light competition among species, thereby affecting plant diversity variation, whereas species coverage, aboveground biomass, plant height are important indicators of light competition changes [61,71,72]. Interesting to note at the local scale species diversity had different controlling factors according to different grassland types, which was in line with other findings as reported [17,73,74]. It is widely known that alpine meadow, mountain meadow and temperate steppe was three grassland types along altitude gradient, thus leading to the discrepancy of soil water content. Alpine meadow had higher productivity and soil moisture compared to other grassland types [17]. Alternatively, mountain meadow and temperate steppe were much sparser than alpine meadows in grass species due to limited nutrients and soil moisture [73,74]. On the one hand, moderate grazing promoted litter removal and increased species diversity, resulting in increasing grazing tolerance of most species, leading to decreasing species occupation area (coverage) owing to herbivores eating and trampling under rest grazing not only in mountain meadows but in temperate steppes. In addition, regarding positive relationships between aboveground biomass and diversity indices and evenness in alpine meadows, stems and leaves of plants had priority over root for growth under limited light, thereby increasing community coverage and aboveground biomass under RG according to optimal allocation theory [61,62,75]. Short-term grazing under RG decreased the evenness due to reducing palatable plants such as gramineous species, sedge species and leguminous species, leading to low aboveground biomass because of palatable plants with high amounts of shoots and leaves in alpine meadows. On the other hand, GE are prior to taller plants that increased resource competition among species, resulting in high mean plant height, thus leading to decreasing species diversity and compared to RG [52,76].
At the regional scale, species diversity indices and richness were positively associated with community coverage and belowground biomass (slightly correlation) under RG. In line with other studies, RG increased community coverage and resulting in positive correlation with species diversity in previous studies reported at broad scale [49,77,78]. Herbivores consumption of taller plants facilitated the growth and assembly of smaller plants during short-term grazing periods, which increased available light for smaller plants and increased species coverage as discussed above, thus increasing community coverage [34,59]. Subsequently, moderate grazing under RG promoted root biomass allocation [18]. For GE, species diversity indices were positively associated with community coverage and above-and below-ground biomass, which was consistent with other studies [79,80]. That may be because GE alleviated the pressure of long-term grazing on grassland ecosystems, leading to increasing species diversity and productivity [59,81]. Besides, alpine meadow plants are relatively low in height after grazing; therefore GE promoted the growth of taller plants and increases community coverage and aboveground biomass as discussed above in RG. Meanwhile, GE promoted the growth of the reproduction and regeneration of suppressed species (such as gramineous species) due to the selective grazing behavior of livestock under grazing, and then increase the species richness. The eating and trampling on soil by livestock were eliminated in grassland under GE, resulting in increased belowground biomass accumulation [82]. Past studies stated that plants reduced the proportion of AGB and allocated large amounts of biomass to BGB so as to germinate and resist grazing pressures as an adaptive strategy in alpine grassland [83,84]. So the aboveground biomass may be mainly came from increasing palatable and taller plants in GE. In fact, relative proportion of gramineous families in GE in our study was higher than that of in RG. Thus, there was no relationship between species richness and aboveground biomass.
5. Conclusions
This study highlights the effect of rest grazing and grazing exclusion on plant composition, diversity and soil characteristics in grasslands of the Qilian mountains. Our findings suggest that community structure characteristics, species diversity indices, soil nutrition variables under rest grazing in grasslands except Pielou index, biomass, soil bulk density and mean plant eight were relatively higher than those under grazing exclusion. Our analysis indicates also that grazing-induced changes in species diversity indices excluding evenness and their regulating factors are scale-dependent. Although different community and soil variables regulated species diversity at each scale, our results indicated that temperature and soil moisture behind those variables regulated species diversity to a certain extent under different grazing management. The space-scale dependence of environmental controls on species diversity has important implications in restoring and managing degraded grasslands. We recommended that the improvement of the grazing management system in arid steppe is mandatory to prevent further land degradation due to biodiversity loss and large-scale biotic homogenization. The implementation of “short-term grazing exclusion” in many sites at the same time and the application of season-grazing in the surrounding areas could enhance the ecological value of protected steppes.
Author Contributions
L.L., Y. H and W. L contributed to the conception of the study. L.L. performed the data analyses and wrote the manuscript. L.L. performed the experiment. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Biodiversity Survey, Monitoring and Assessment Project of Ministry of Ecology and Environment, China (Grants No. 2019HB2096001006).
Data Availability Statement
Not applicable.
Acknowledgments
We thank Zhao Changming and his team members for their assistance with field experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dong, S.K.; Sherman, R. Enhancing the resilience of coupled human and natural systems of alpine rangelands on the Qinghai-Tibetan Plateau. Rangel. J. 2015, 31, 1–3. [Google Scholar] [CrossRef]
- Qian, D.W.; Du, Y.G.; Li, Q.; Guo, X.W. , Cao, G.M. Alpine grassland management based on ecosystem service relationships on the southern slopes of the Qilian Mountains, China. J. Environ. Manage. 2021, 288, 112447. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Liu, X.Y.; Zhao, C.Y.; Wang, H. , Zang, F. The trade-offs and synergies of the ecological-production-living functions of grassland in the Qilian mountains by ecological priority. J. Environ. Manage. 2023, 327, 116883. [Google Scholar] [CrossRef] [PubMed]
- Qian, D.W.; Li, Q.; Fan, B.; Lan, Y.T.; Cao, G.M. Characterization of the spatial distribution of plateau pika burrows along an alpine grassland degradation gradient on the Qinghai–Tibet Plateau. Ecol. Evol. 2021, 11, 14905–14915. [Google Scholar] [CrossRef] [PubMed]
- Steffens, M.; K Lbl, A.; Totsche, K.U.; K Gel-Knabner, I. Grazing effects on soil chemical and physical properties in a semiarid steppe of Inner Mongolia (P.R. China). Geoderma 2008, 143, 63–72. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Y.; Zhao, X.; Awada, T.; Shang, W.; Han, J. Effects of grazing exclusion on soil properties and on ecosystem carbon and nitrogen storage in a sandy rangeland of Inner Mongolia, northern China. J. Environ. Manage. 2012, 50, 622–632. [Google Scholar] [CrossRef]
- Miao, F.H.; Guo, Z.G.; Xue, R.; Wang, X.Z.; Shen, Y.Y. Effects of grazing and precipitation on herbage biomass, herbage nutritive value, and yak performance in an alpine meadow on the Qinghai – Tibetan Plateau. PLoS. One. 2015, 10, e127275. [Google Scholar] [CrossRef]
- Li, W.; Cao, W.X.; Wang, J.L.; Li, X.L.; Xu, C.L.; Shi, S.L. Effects of grazing regime on vegetation structure, productivity, soil quality, carbon and nitrogen storage of alpine meadow on the Qinghai-Tibetan Plateau. Ecol. Eng. 2017, 98, 123–133. [Google Scholar] [CrossRef]
- Ma, C.H.; Hao, X.H.; He, F.C.; Baoyin, T.G. , Yang, J.J., Dong, S.K. Effects of seasonal grazing on plant and soil microbial diversity of typical temperate grassland. Front. Plant. Sci. 2022, 13, 1040377. [Google Scholar] [CrossRef]
- Zhan, T.Y.; Zhao, W.W.; Feng, S.Y.; Hua, T. Plant community traits respond to grazing exclusion duration in alpine meadow and alpine steppe on the Tibetan plateau. Front. Plant. Sci. 2022, 13, 863246. [Google Scholar] [CrossRef]
- Cai, D.W.; Ge, Q.S.; Wang, X.M.; Liu, B.L.; Goudie, A.S.; Hu, S. Contribution of ecological programs to vegetation restoration in arid and semiarid China. Environ. Res. Lett. 2020, 15, 114046. [Google Scholar] [CrossRef]
- Wang, S.Z.; Fan, J.W.; Li, Y.Z.; Huang, L. Effects of grazing exclusion on biomass growth and species diversity among various grassland types of the Tibetan Plateau. Sustainability-Basel. 2019, 11, 1705. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Z.W.; Huang, X.T.; Li, L.H. How does grazing exclusion influence plant productivity and community structure in alpine grasslands of the Qinghai-Tibetan Plateau? Glob.Ecol.Conserv. 2020, 23, e01066. [Google Scholar] [CrossRef]
- Fedrigo, J.K.; Ataide, P.F.; Filho, J.A.; Oliveira, L.V.; Jaurena, M.; Laca, E.A.; Overbeck, G.E.; Nabinger, C. Temporary grazing exclusion promotes rapid recovery of species richness and productivity in a long-term overgrazed Campos grassland. Restor. Ecol. 2018, 26, 677–685. [Google Scholar] [CrossRef]
- McDonald, S.E.; Lawrence, R.; Kendall, L.; Rader, R. Ecological, biophysical and production effects of incorporating rest into grazing regimes: A global meta-analysis. J.Appl. Ecol. 2019, 56, 2723–2731. [Google Scholar] [CrossRef]
- Bi, X.; Li, B.; Fu, Q.; Fan, Y.; Ma, L.X.; Yang, Z.H.; Nan, B.; Dai, X.H.; Zhang, X.S. Effects of grazing exclusion on the grassland ecosystems of mountain meadows and temperate typical steppe in a mountain-basin system in Central Asia's arid regions, China. Sci.Total.Environ. 2018, 630, 254–263. [Google Scholar] [CrossRef]
- Gao, J.L.; Carmel, Y. A global meta-analysis of grazing effects on plant richness. Agriculture. Agr.Ecosyst. Environ. 2020, 302, 107072. [Google Scholar] [CrossRef]
- Dong, S.K.; Shang, Z.H.; Gao, J.X.; Boone, R.B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agr. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]
- Mekuria, W.; Betemariam, E. Exclosure land management for restoration of the soils in degrade communal grazing lands in northern Ethiopia. Land. Degrad.Dev. 2011, 24, 528–538. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, W. Vegetation and soil property response of short-time fencing in temperate desert of the Hexi Corridor, northwestern China. Catena 2015, 133, 4351. [Google Scholar] [CrossRef]
- Meissner, R.A.; Facelli, J.M. Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrublands of South Australia. J. Arid. Environ. 1999, 42, 117–128. [Google Scholar] [CrossRef]
- Altesor, A.; Oesterheld, M.; Leoni, E.; Lezama, F.; Rodríguez, C. Effect of grazing on community structure and productivity of a Uruguayan grassland. Plant. Ecol. 2005, 179, 83–91. [Google Scholar] [CrossRef]
- Wang, D.; Wu, G.L.; Zhu, Y.J.; Shi, Z.H. Grazing exclusion effects on above- and below-ground C and N pools of typical grassland on the Loess Plateau (China). Catena 2014, 123, 113120. [Google Scholar] [CrossRef]
- Song, S.S.; Zhu, J.L.; Zheng, T.L.; Tang, Z.Y.; Zhang, F.; Ji, C.J.; Shen, Z.H.; Zhu, J.X. Long-term grazing exclusion reduces species diversity but increases community heterogeneity in an alpine grassland. Front. Ecol. Evol. 2020; 8, 66. [Google Scholar] [CrossRef]
- Karami, P.; Bandak, I.; Gorgin Karaji, M. Comparing the effects of continuous grazing and long term exclosure on floristic composition and plant diversity in rangeland ecosystems of Saral, Iran. Int. J. Environ. Sci. Te. 2019, 16, 7769–7776. [Google Scholar] [CrossRef]
- McDonald, S.E.; Lawrence, R.; Rader, R. A Systematic Review of Ecological and Production Outcomes under Rest-Grazing Systems. XXIV International Grassland Congress /XI International Rangeland Congress, 2021.
- Herrero-Jáuregui, C.; Oesterheld, M. Effects of grazing intensity on plant richness and diversity: A meta-analysis. Oikos 2018, 127, 757–766. [Google Scholar] [CrossRef]
- Filazzola, A.; Brown, C.; Dettlaff, M.A.; Batbaatar, A.; Grenke, J.; Bao, T.; Heida, I.P.; Cahill Jr, J.F. The effects of livestock grazing on biodiversity are multi-trophic: a meta-analysis. Ecol. Lett. 2020, 23, 1298–1309. [Google Scholar] [CrossRef]
- Olff, H.; Ritchie, M.E. Effects of herbivores on grassland plant diversity. Trends. Eco. Evo. 1998, 13, 261–265. [Google Scholar] [CrossRef]
- Bakker, E.S.; Ritchie, M.E.; Olff, H.; Milchunas, D.G.; Knops, J.M.H. Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size. Eco. Lett. 2006, 9, 9,780–788. [Google Scholar] [CrossRef]
- Deng, L.; Shangguan, Z.P.; Wu, G.L.; Chang, X.F. Effects of grazing exclusion on carbon sequestration in China’s grassland. Earth-Sci. Rev 2017, 173, 84–95. [Google Scholar] [CrossRef]
- Schultz, N.L.; Morgan, J.W.; Lunt, I.D. Effects of grazing exclusion on plant species richness and phytomass accumulation vary across a regional productivity gradient. J. Veg. Sci. 2011, 22, 130–142. [Google Scholar] [CrossRef]
- FernándezLugo, S. , De, N.L., Mellado, M., Arévalo, J.R. Grazing effects on species richness depends on scale: a 5-year study in Tenerife pastures (Canary Islands). Plant. Eco. 2011, 212, 423–432. [Google Scholar] [CrossRef]
- Li, W.H.; Zhan, S.X. , Lan, Z.C., Ben Wu, X.B., Bai, Y.F. Scale-dependent patterns and mechanisms of grazing-induced biodiversity loss: evidence from a field manipulation experiment in semiarid steppe. Landscape. Eco. 2015, 30, 1751–1765. [Google Scholar] [CrossRef]
- Mardari, C. Plant diversity-environment relationships in xeric grasslands of north-eastern romania. Appl.Ecol.Env.Res. 2016, 14, 111–127. [Google Scholar] [CrossRef]
- Sandel, B.; Corbin, J.D. Scale-dependent responses of species richness to experimental manipulation of productivity and disturbance in Californian coastal grasslands. J. Veg. Sci. 2012, 23, 906–918. [Google Scholar] [CrossRef]
- Li, W. The evaluation of the research on the grazing optimization hypothesis. Grassland.China. 1999, 4, 62–67. [Google Scholar]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; McIntyre, S.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meir, I.; Landsberg, J.; Zhang, W.; Clark, H.; Campbell, B.D. Plant trait responses to grazing-a global synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Li, Z.X.; Feng, Q.; Li, Z.J.; Wang, X.F.; Gui, J.; Zhang, B.J.; Li, Y.C.; Deng, X.H.; Xue, J.; Gao, W.D.; Yang, A.L.; Nan, F.S.; Liang, P.F. Reversing conflict between humans and the environment-The experience in the Qilian Mountains. Renew. Sust. Energ. Rev. 2021, 148, 111333. [Google Scholar]
- Xu, W.H.; Xiao, Y.; Zhang, J.J.; Yang, W.; Zhang, L.; Hull, V.; Wang, Z.; Zheng, H.; Liu, J.G.; Polasky, S.; Jiang, L.; Xiao, Y.; Shi, X.W.; Rao, E.M.; Lu, F.; Wang, X.K.; Daily, G.C.; Ouyang, Z.Y. 2017. Strengthening protected areas for biodiversity and ecosystem services in China. PNAS 2017, 114, 1601–1606. [Google Scholar] [CrossRef]
- Chen, L.L.; Xu, H.B.; Wu, S.Y.; Baoyin, T. Plant and soil properties mediate the response of soil microbial communities to moderate grazing in a semiarid grassland of northern China. J. Environ. Manage. 2021, 284, 112005. [Google Scholar] [CrossRef]
- Ma, K.P.; Huang, J.H.; Yu, S.L.; Chen, L.Z. Plant community diversity in Dongling Mountain, Beijing, China. II. Species richness, evenness and species diversities. Acta. Ecol. Sin. 1995, 15, 268–277. [Google Scholar]
- Gomboš, M.; Pavelkova, D.; Kandra, B.; Tall, A. Quantification of temporal and spatial changes in soil suspension density, specific surface area and parameters of the distribution of the texture of the dispersed soil phase. IOP Conference Series: Materials Science and Engineering. IOP Publishing 2022, 1252, 012034. [Google Scholar]
- Mango, L.; Nhete, M.; Mutanda, G.W. Varying effects of field capacity watering regimes in distinct soil types on the growth characteristics of wild sorghum. Octa. J. Biosciences. 2022, 10, 10,62–75. [Google Scholar]
- Lu, R.K. Soil and Agricultural Chemistry Analysis; Agricultural Science and Technology Press: Beijing, China, 2000; pp. 18–96. [Google Scholar]
- Du, K. Response of Vegetation and Soil Properties, Carbon and Nitrogen Pools to Different Grazing Patterns in Alpine Meadow in Qilian Mountains. Ma.Sc dissertation, Gansu Agricultural University, Lan Zhou, 2020. [Google Scholar]
- Jiang, L.X.; Tian, Y.; Liu, X.Y.; Zhang, K.B. Effects of different grazing methods on the composition and structure of plant functional groups in grassland community. J.Beijing.Forest.Univ. 2022, 44, 77–86. [Google Scholar]
- Zhang, W.N.; Ganjurjav; Li, Y. W.; Gao, Q.Z.; Wan, Y.F.; Li, Y.E.; DanJiu, L.B.; XiRao, Z.M.; BaiMa, Y.Z. Effects of Banning Grazing and Delaying Grazing on Species Diversity and Biomass of Alpine Meadow in Northern Tibet. J.Agr.Sci.Tech-Iran. 2013, 15, 143–149. [Google Scholar]
- Bai, W.L.; Hu, F.C.; Li, C.T. Study on enclosure period and utilization mode of Tianzhu County grassland. J.Animal.Sci.Vet.Med. 2015, 34, 40–43. [Google Scholar]
- Xiang, M.X.; Guo, Y.J.; Gu, S.Q.Z.; Zhang, X.Q.; Pan, Y.; Wu, J.X.; LHA, D. Effects of grazing intensity on plant community and species diversity of temperate steppe in Lhasa river valley. Acta.Agrestia.Sin. 2019, 27, 668–674. [Google Scholar]
- Tian, L.H.; Bai, Y.F.; Wang, W.W.; Qu, G.P. , Deng, Z.H., Li, R.C., Zhao, J.X. Warm-and cold-season grazing affect plant diversity and soil carbon and nitrogen sequestration differently in Tibetan alpine swamp meadows. Plant Soil 2021, 458, 458,151–164. [Google Scholar] [CrossRef]
- Wang, C.T.; Long, R.J.; Wang, Q.L.; Jing, Z.C.; Shi, J.J. Changes in plant diversity, biomass and soil C, in alpine meadows at different degradation stages in the head-water region of three Rivers, China. Land. Degrad.Dev. 2007, 18, 578–588. [Google Scholar]
- Liu, M.; Liu, G.H.; Wu, X.; Wang, H.; Chen, L. Vegetation traits and soil properties in response to utilization patterns of grassland in Hulun Buir City, Inner Mongolia, China. Chinese. Geogr.Sci. 2014, 24, 471–478. [Google Scholar] [CrossRef]
- Hao, B.T. Effects of Different Rest-Grazing on Typical Grassland Plant Communities and Soil Stoichiometry. Ma.Sc dissertation, Inner Mongolia University, Huhhot, 2019. [Google Scholar]
- Yan, C.C.; Wang, Y.; Yang, Z.F.; Liu, P.J.; Wang, G.Q.; Hao, H.T.; Xi, L.Q. Effects of long-term enclosure on plant diversity and soil physicochemical properties of meadow grassland on the south slope of Tianshan Mountains. Heilongjiang. Animal. Sci. Vet. Med. 2021, 13, 13,91–96. [Google Scholar]
- Xu, G.P.; Zhang, D.G.; Xu, C.L.; Pu, X.P.; Liu, X.J.; Liu, X.N.; Chen, J.G. Effect of grazing disturbance on species diversity of alpine grassland plant community in Eastern Qilian Mountains. J. Gansu. Agric. Univ. 2005, 6, 789–796. [Google Scholar]
- Song, X.Y.; Wei, Z.J.; Zheng, S.H.; Li, L.H.; Chang, S.J.; Yang, Y.; Liu, A.J. Impacts of Different Disturbances on Ecosystem Characteristic of Typical Steppe in Hulun Buir. Ecol. Environ. Sci. 2018, 27, 27,1405–1410. [Google Scholar]
- Cheng, Y.T. Research on Grassland Vegetation and Soil Recovery after Fencing in China: A Meta-analysis. Ma.Sc dissertation, East China Normal University, Shanghai, 2020. [Google Scholar]
- Zhang, Q.; Yang, J.; Yao, B.H.; Cai, Z.Y.; Wang, X.Y. ,Su, J.H. Effects of grazing modes on soil physical, chemical properties and species diversity in alpine meadow in the eastern margin of Qilian Mountains. Grassland.Turf. 2021, 41, 105–112. [Google Scholar]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 324,636–638. [Google Scholar] [CrossRef] [PubMed]
- Eskelinen, A.; Harpole, W.S.; Jessen, M.T.; Virtanen, R.; Hautier, Y. Light competition drives herbivore and nutrient effects on plant diversity. Nature 2022, 611, 301–305. [Google Scholar] [CrossRef]
- Ren, G.H.; Wang, C.X.; Dong, K.H.; Zhu, H.S.; Wang, Y.C.; Zhao, X. Effects of grazing exclusion on soil–vegetation relationships in a semiarid grassland on the Loess Plateau, China. Land. Degrad. Dev. 2018, 29, 4071–4079. [Google Scholar] [CrossRef]
- Chai, Q.L.; Ma, Z.Y.; Chang, X.F.; Wu, G.L.; Zheng, J.Y.; Li, Z.W.; Wang, G.J. Optimizing management to conserve plant diversity and soil carbon stock of semi-arid grasslands on the Loess Plateau. Catena 2019, 172, 781–788. [Google Scholar] [CrossRef]
- Sun, Y.; Schleuss, P.M.; Pausch, J.; Xu, X.L.; Kuzyakov, Y. Nitrogen pools and cycles in Tibetan Kobresia pastures depending on grazing. Biol. Fert. Soils. 2018, 54, 569–581. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, D.W.; Li, Q.; Wang, Q.C. Vertical distributions of soil nutrients and their stoichiometric ratios as affected by long term grazing and enclosing in a semi-arid grassland of Inner Mongolia. Agriculture 2020, 10, 382. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J.; Bautista, S.; Bellot, J.; Vallejo, R. Small scale environmental heterogeneity and spatiotemporal dynamics of seedling establishment in a semiarid degraded ecosystem. Ecosystems 2003, 6, 630–643. [Google Scholar] [CrossRef]
- Sigcha, F.; Pallavicini, Y.; Camino, M.J.; Martínez-Ruiz, C. Effects of short-term grazing exclusion on vegetation and soil in early succession of a Subhumid Mediterranean reclaimed coal mine. Plant Soil 2018, 426, 426,197–209. [Google Scholar] [CrossRef]
- Kohandel, A.; Arzani, H.; Hosseini, M. Effect of grazing intensity on N. P. K of soil. Iran-watershed manage. Sci. Eng. 2006, 3, 59–65. [Google Scholar]
- Zarekia, S.; Jafari, M.; Arzani, H.; Javadi, S.A.; Jafari, A.A. Grazing effects on physical and chemical properties of soil. World. Appl. Sci.J. 2012, 20, 205–212. [Google Scholar]
- Sala, O.E.; Stuart Chapin, F.; Armesto, J.J.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.L.; Liao, L.R.; Wang, J.; Zhang, L.; Ye, Z.C.; Liu, G.B.; Zhang, C. The diversity-Godron stability relationship of alpine grassland and its environmental drivers. Acta. Pratac. Sin. 2023, 2, 1–12. [Google Scholar]
- Zhu, J.T.; Jiang, L.; Zhang, Y.J.; Jiang, Y.B.; Tao, J.; Tian, L.; Zhang, T.; Xi, Y. Below-ground competition drives the self-thinning process of Stipa purpurea populations in northern Tibet. J. Veg. Sci. 2015, 26, 166–174. [Google Scholar] [CrossRef]
- Wu, J.S.; Li, M.; Fiedler, S.; Ma, W.L.; Wang, X.T.; Zhang, X.Z.; Tietjenm, B. Impacts of grazing exclusion on productivity partitioning along regional plant diversity and climatic gradients in Tibetan alpine grasslands. J. Environ. Manage. 2019, 231, 635–645. [Google Scholar] [CrossRef]
- Wang, W.Y.; Ma, Y.G.; Xu, J.; Wang, H.C.; Zhu, J.F.; Zhou, H.K. The uptake diversity of soil nitrogen nutrients by main plant species in Kobresia humilis alpine meadow on the Qinghai-Tibet Plateau. Sci.China.Earth.Sci. 2012, 55, 1688–1695. [Google Scholar] [CrossRef]
- Wu, G.L.; Wang, D.; Liu, Y.; Ding, L.M.; Liu, Z.H. Warm-season grazing benefits species diversity conservation and topsoil nutrient sequestration in alpine meadow. Land.Degrad.Dev. 2017, 28, 28,1311–1319. [Google Scholar]
- Li, Y. Study on clonal modules of Kobresia humilis under different degenerative gradation in Kobresia Meadow. Pratac. Animal. Husbandry. 2007, 27, 6–9. [Google Scholar]
- Wang, K.B.; Deng, L.; Ren, Z.P.; Li, J.P.; Shangguan, Z.P. Grazing exclusion significantly improves grassland ecosystem C and N pools in a desert steppe of Northwest China. Catena 2016, 137, 441–448. [Google Scholar] [CrossRef]
- Wu, G.L.; Du, G.Z.; Liu, Z.H.; Thirgood, S. Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau. Plant Soil 2009, 319, 115–126. [Google Scholar] [CrossRef]
- Zhao, J.X.; Li, X.; Li, R.C.; Tian, L.H.; Zhang, T. Effect of grazing exclusion on ecosystem respiration among three different alpine grasslands on the central Tibetan Plateau. Ecol. Eng. 2016, 94, 599–607. [Google Scholar] [CrossRef]
- Yin, X.Y.; Ye, M.; Guo, J.X.; Zhang, K.L.; Zhao, F.F. Relationship Between species diversity and productivity of different grassland types in Buerjin Forest area of Altai Mountains. J.Soil.Water.Conserv. 2022, 36, 110–115. [Google Scholar]
- Gao, Y.H.; Zeng, X.Y.; Schumann, M.; Chen, H. Effectiveness of exclosures on restoration of degraded alpine meadow in the eastern Tibetan Plateau. Arid.Land.Res.Manag. 2011, 25, 164–175. [Google Scholar] [CrossRef]
- Ma, W.L.; Shi, P.L.; Li, W.H.; He, Y.T.; Zhang, X.Z.; Shen, Z.X.; Chai, S.Y. Changes in individual plant traits and biomass allocation in alpine meadow with elevation variation on the Qing-hai-Tibetan Plateau. Sci.China. Life.Sci. 2010, 53, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, X.D.; Cheng, G.W.; Wu, J.B.; Hong, J.T.; Niu, S.L. Effects of grazing regimes on plant traits and soil nutrients in an alpine steppe, northern Tibetan Plateau. PLoS. One 2014, 9, 108821. [Google Scholar] [CrossRef]
Figure 1.
Comparison of Shannon-Wiener index based on generalized mixed model in grassland in Qilian Mountains.
Figure 1.
Comparison of Shannon-Wiener index based on generalized mixed model in grassland in Qilian Mountains.
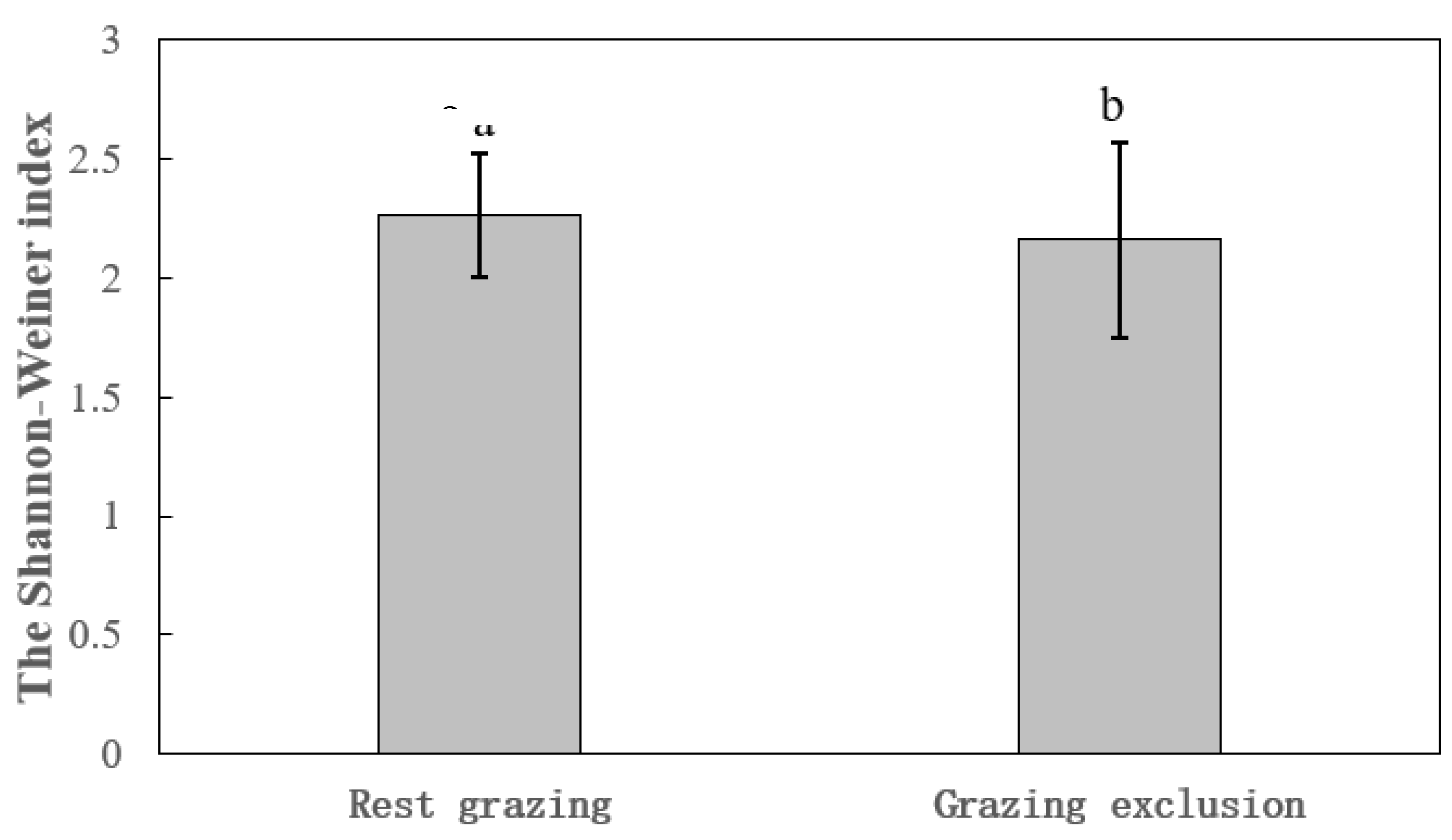
Figure 2.
Comparison of Simpson index based on generalized mixed model in grassland in Qilian Mountains.
Figure 2.
Comparison of Simpson index based on generalized mixed model in grassland in Qilian Mountains.
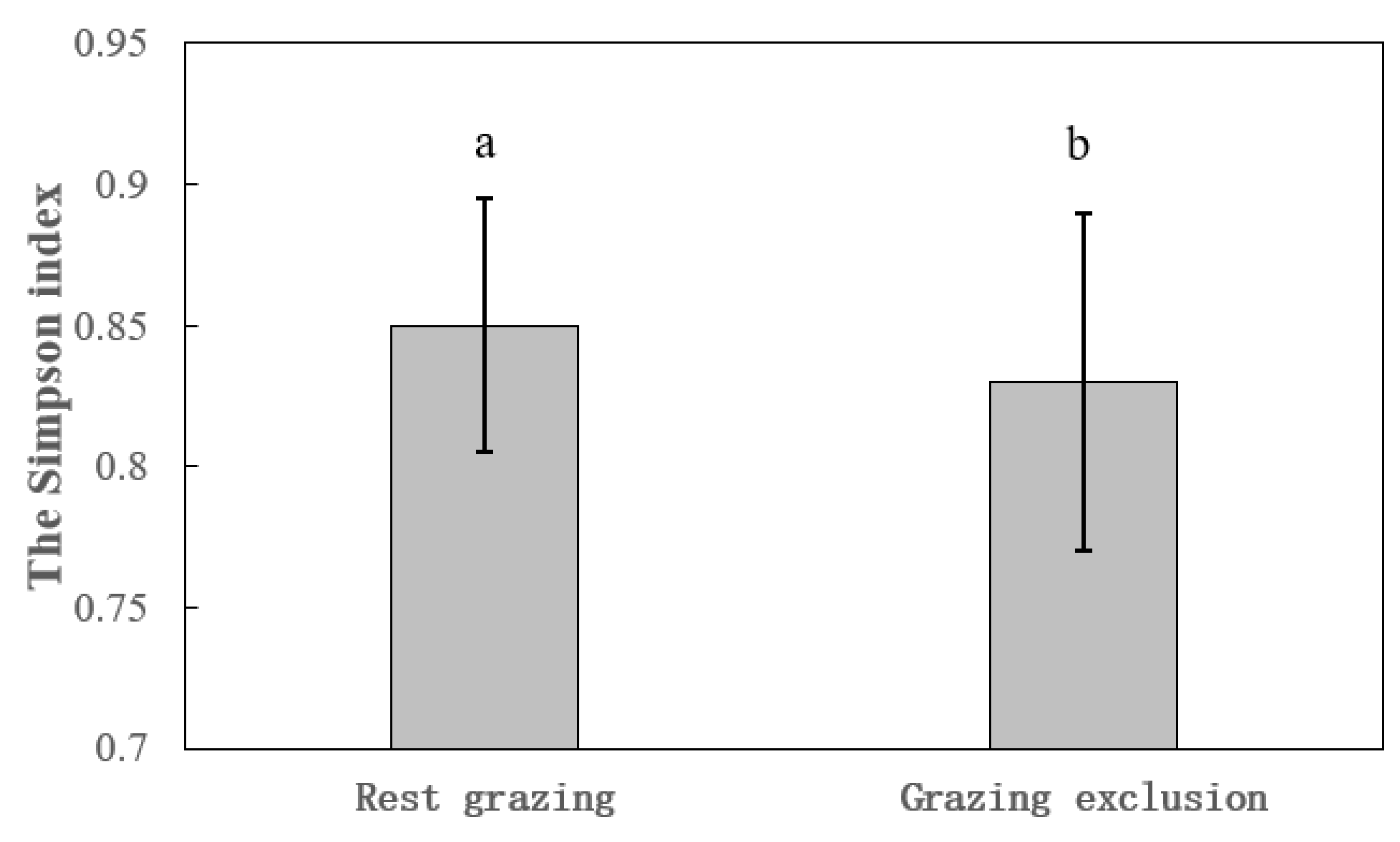

Table 1.
Plot information.
| Grassland Types | Number of plots | Returning grazing lands to grasslands | Years |
|---|---|---|---|
| Mountain meadow | 5 | Grazing exclusion | >20 years |
| Mountain meadow | 6 | Grazing in the non-growing seasons | unknown |
| Mountain meadow | 5 | Grazing exclusion | 10 months |
| Alpine meadow | 1 | Grazing exclusion | >20 years |
| Alpine meadow | 5 | Grazing in the growing seasons | unknown |
| Temperate grassland | 5 | Grazing exclusion | 4 years |
| Temperate grassland | 5 | Grazing in the non-growing seasons | unknown |
Table 2.
Vegetation and soil characteristics in different grasslands.
| Index | Mountain meadow | Alpine meadow | Temperate steppe | Region | |||||
|---|---|---|---|---|---|---|---|---|---|
| Grazing exclusion | Rest-grazing | Grazing exclusion 10 months | Grazing exclusion | Rest-grazing | Grazing exclusion | Rest-grazing | Grazing exclusion | Rest-grazing | |
| Coverage (%) | 99.20a | 96.83 a | 66.20c | 81.00 | 89.80 | 61.80a | 64.80a | 76.06a | 84.63a |
| Height(cm) | 9.81ab | 15.58a | 19.89a | 4.57 | 2.40 | 12.90a | 7.14a | 13.60a | 8.83a |
| Species richness | 68b | 69a | 35c | 16 | 60 | 24a | 36a | 98b | 128a |
| Pielou index | 0.78a | 0.76a | 0.79a | 0.78 | 0.73 | 0.70a | 0.74a | 0.76a | 0.74a |
| Shannon-Wiener index | 2.63a | 2.41a | 2.09cb | 2.22 | 2.27 | 1.73b | 2.07a | 2.16b | 2.26a |
| Simpson | 0.89a | 0.87a | 0.84c | 0.86 | 0.84 | 0.76b | 0.82a | 0.83b | 0.85a |
| Aboveground biomass(g/m2) | 377.08a | 355.42a | 350.77a | 185.36 | 128.26 | 123.11a | 287.19a | 277.51a | 263.11a |
| belowground biomass(g/m2) | 1898.60a | 1407.60a | / | / | 1253.40 | 405.27a | 613.00a | 1151.93a | 1091.33a |
| Soil organic carbon (mg/g) | 39.32c | 69.48a | 25.27c | 48.52 | 43.09 | 17.68a | 24.55a | 28.74b | 45.71a |
| Soil nitrogen (mg/g) | 5.58a | 6.82a | 2.79c | 4.97 | 5.70 | 2.30b | 2.88a | 3.64a | 5.29a |
| Soil total phosphorus (mg/g) | 0.61a | 0.72a | 0.73a | 0.62 | 0.89 | 0.66a | 0.55a | 0.67a | 0.72a |
| Soil total potassium (mg/g) | 3.90b | 5.75a | 5.75ac | 4.11 | 4.19 | 5.42a | 5.38a | 4.97a | 5.11a |
| Soil bulk density (g/cm3) | 0.68a | 0.72a | 0.89a | 0.52 | 0.74 | 1.07a | 0.94b | 0.86a | 0.80a |
| Soil porosity (%) | 46.57a | 54.54a | 40.17a | 27.69 | 43.31 | 45.03a | 39.73a | 42.91a | 45.86a |
| Field capacity (%) | 58.79ab | 74.95a | 40.63bc | 35.39 | 45.62 | 45.52a | 42.03a | 47.50a | 54.20a |
| <0.002mm clay content (%) | 7.59ab | 8.63b | 9.93a | 7.67 | 10.09 | 9.02a | 9.65a | 8.70a | 9.46a |
| 0.002-0.02mm silt content (%) | 64.77a | 79.78a | 81.42a | 74.53 | 77.02 | 76.18a | 76.67a | 73.60a | 77.82a |
| 0.02-2mm sand content (%) | 10.98a | 11.59a | 8.64a | 17.80 | 12.89 | 14.80a | 13.68a | 11.82b | 12.72a |
Note: Belowground biomass is not allowed to collect in alpine meadows under GE and in Mountain meadow under the 10-month grazing exclusion. The same letter indicates P>0.05, adjacent letters without the same letter indicate P<0.05, and letters that are neither the same nor adjacent indicate P<0.01.
Table 3.
Relationship between community characteristics and species diversity in regional grassland.
Table 3.
Relationship between community characteristics and species diversity in regional grassland.
| Management | Community characteristics | Species richness | Shannon-Wiener index | Simpson index | ||||||
| Rest-grazing | Regression equation | R2 | P | Regression equation | R2 | P | Regression equation | R2 | P | |
| Coverage (%) | y=0.24x+1.35 | 0.65 | 0 | y=0.011x+1.32 | 0.4 | 0.009 | y=0.0014x+0.73 | 0.22 | 0.069 | |
| belowground biomass(g/m2) | y=0.0027x+17.98 | 0.23 | 0.069 | / | / | / | / | / | / | |
| Grazing exclusion | Coverage (%) | y=0.34x-8.27 | 0.62 | 0 | y=0.015x+1.01 | 0.54 | 0.001 | y=0.002x+0.69 | 0.42 | 0.007 |
| Aboveground biomass(g/m2) | y=0.026x+10.07 | 0.2 | 0.087 | y=0.002x+1.71 | 0.33 | 0.021 | y=0.0002x+0.77 | 0.35 | 0.017 | |
| belowground biomass(g/m2) | y=0.007x+12.59 | 0.64 | 0.005 | y=0.0003x+1.79 | 0.5 | 0.02 | y=0.000046x+0.77 | 0.47 | 0.028 | |
Table 4.
Relationship between soil characteristics and species diversity in different grassland.
| Index | Management | Species richness | Shannon-Wiener index | Simpson index | Pielou index | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Regression equation | R2 | P | Regression equation | R2 | P | Regression equation | R2 | P | Regression equation | R2 | P | ||
| SOC | RG | y=0.12x+15.24 | 0.46 | 0.001 | y=0.008x+1.87 | 0.54 | 0.002 | y=0.001x+0.79 | 0.4 | 0.011 | |||
| GE | y=0.018x+1.64 | 0.46 | 0.004 | y=0.003x+0.76 | 0.46 | 0.004 | y=0.002x+0.70 | 0.27 | 0.039 | ||||
| TN | RG | y=1.63x+12.46 | 0.54 | 0.003 | |||||||||
| GE | y=3.88x+3.19 | 0.63 | 0 | y=0.18x+1.49 | 0.62 | 0 | y=0.024x+0.74 | 0.52 | 0.002 | ||||
| TP | RG | y=0.57x+1.83 | 0.47 | 0.005 | y=0.097x+0.77 | 0.4 | 0.011 | ||||||
| GE | |||||||||||||
| TK | RG | ||||||||||||
| GE | y=-3.56x+35.01 | 0.31 | 0.026 | ||||||||||
| BD | RG | ||||||||||||
| GE | y=-18.192x+32.945 | 0.25 | 0.047 | y=-1.12x+3.12 | 0.42 | 0.006 | y=-0.16x+0.97 | 0.43 | 0.006 | ||||
| SP | RG | y=0.015x+1.53 | 0.71 | 0 | y=0.003x+0.72 | 0.65 | 0 | y=0.003x+0.59 | 0.47 | 0.005 | |||
| GE | |||||||||||||
| FC | RG | y=0.11x+14.93 | 0.32 | 0.027 | y=0.009x+1.77 | 0.56 | 0.001 | y=0.001x+0.76 | 0.48 | 0.004 | |||
| GE | |||||||||||||
| C/N | RG | y=0.065x+1.69 | 0.38 | 0.018 | y=0.012x+0.74 | 0.43 | 0.011 | y=0.017x+0.59 | 0.38 | 0.019 | |||
| GE | |||||||||||||
| C/P | RG | ||||||||||||
| GE | y=0.30x+3.88 | 0.48 | 0.003 | y=0.016x+1.45 | 0.59 | 0.001 | y=0.002x+0.732 | 0.54 | 0.001 | ||||
| C/K | RG | y=0.83x+13.38 | 0.55 | 0.002 | y=0.039x+1.89 | 0.34 | 0.023 | ||||||
| GE | y=1.98x+5.10 | 0.58 | 0.001 | y=0.099x+1.55 | 0.63 | 0 | y=0.014x+0.75 | 0.56 | 0.001 | ||||
| N/P | RG | y=-0.005x+0.79 | 0.31 | 0.04 | |||||||||
| GE | y=1.74x+6.69 | 0.66 | 0 | y=0.062x+1.78 | 0.37 | 0.013 | y=0.008x+0.79 | 0.26 | 0.044 | ||||
Table 5.
Relationship between community characteristics and species diversity in different grassland.
Table 5.
Relationship between community characteristics and species diversity in different grassland.
| Grassland | Management | Community characteristics | Species richness | Shannon-Wiener index | Simpson index | Pielou index | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Regression equation | R2 | P | Regression equation | R2 | P | Regression equation | R2 | P | Regression equation | R2 | P | |||
| Alpine meadow | RG | Aboveground biomass(g/m2) | y=0.035x+17.94 | 0.7 | 0.078 | y=0.013x+0.56 | R2=0.89 | 0.016 | y=0.002x+0.52 | 0.85 | 0.025 | y=0.004x+0.23 | 0.89 | 0.016 |
| Mountain meadow | RG | Coverage (%) | y=0.036x-1.05 | 0.55 | 0.092 | y=0.0067x+0.10 | 0.56 | 0.086 | ||||||
| Species coverage | y=-0.078x+3.64 | 0.81 | 0.015 | y=-0.01x+1.02 | 0.87 | 0.007 | y=-0.014x+0.98 | 0.79 | 0.018 | |||||
| GE | Coverage (%) | y=0.39x-12.13 | 0.71 | 0.002 | y=0.013x+1.33 | 0.55 | 0.014 | y=0.0012x+0.76 | 0.4 | 0.049 | ||||
| Height(cm) | y=-0.026x+2.75 | 0.4 | 0.048 | y=-0.003x+0.91 | 0.38 | 0.056 | ||||||||
| Temperate steppe | RG | Aboveground biomass(g/m2) | y=0.01x+13.86 | 0.67 | 0.089 | y=-0.0003x+0.82 | 0.64 | 0.1 | ||||||
| Species coverage | y=-0.69x+24.18 | 0.86 | 0.023 | y=0.018x+0.54 | 0.63 | 0.1 | ||||||||
| GE | Height(cm) | y=-0.40x+17.34 | 0.64 | 0.1 | y=-0.016x+1.94 | 0.65 | 0.099 | |||||||
| Number of plants/clumps* | y=0.004x+0.62 | 0.65 | 0.098 | |||||||||||
| Species coverage | y=-0.82x+21.21 | 0.71 | 0.072 | y=-0.032x+2.09 | 0.67 | 0.09 | y=0.008x+0.62 | 0.66 | 0.096 | |||||
Note: * means plant density per clumps.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



