
Submitted:
22 August 2023
Posted:
24 August 2023
You are already at the latest version
Abstract
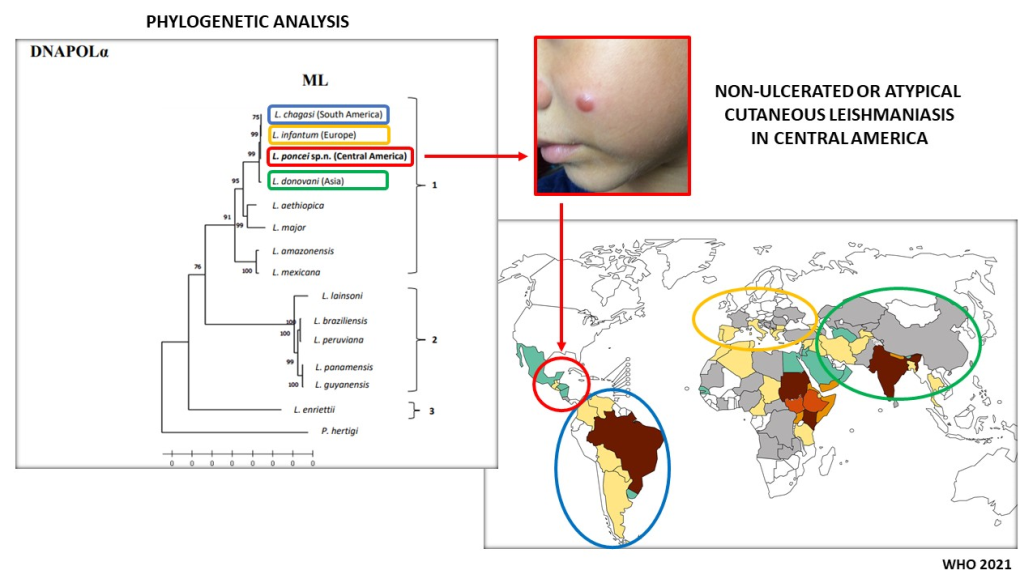
Non-ulcerated cutaneous leishmaniasis (NUCL) is an atypical and peculiar clinical form of the disease that was first described by Ponce et al. (1988) in Honduras (Central America), where iso-lated or disseminated popular and/or nodular skin lesions are commonly seen in adolescents and young adults. Leishmania (L.) chagasi was then incriminated as the causal agent of the disease, as well as of American visceral leishmaniasis (AVL), although NUCL has been shown to be more prevalent than AVL in the region. Considering, however, the dubious taxonomic character of the causal agent of NUCL, as there is no evidence of this form of disease associated with L. (L.) cha-gasi in South America, this study aimed to carry out a retrospective taxonomic analysis on a se-ries of studies on the diagnostic approach to the Honduran parasite in loco (El Tigre island, Amapala municipality, west coast of Honduras), where taxonomic characters of a biological (in vitro and in vivo), epidemiological (vector and canine reservoir), clinical-immunological (im-munopathological features of in situ skin lesion and systemic infection) and molecular (ge-nomic/phylogenetic analyses) nature were considered. The findings, particularly those of mo-lecular nature, such as: the Honduran parasite showed to be more ancestral (molecular clock analysis of the DNA polymerase alpha subunit gene) than all viscerotropic ones [L. (L.) chagasi, L. (L.) donovani and L. (L.) infantum], as well as presenting an absolutely unprecedented structural variation on chromosome 17 and the highest frequency of genomic SNPs (more than twice the number seen in the Brazilian parasite), in addition to representing a different phylogenetic lin-eage (mainly in the NJ - RNA Pol II tree) from those viscerotropic ones, have pointed to the characterization of a new leishmanine parasite, Leishmania (Leishmania) poncei n. sp. (Kinetoplas-tea: Trypanosomatidae), in honor of Professor Ponce, who was the first researcher to describe NUCL in Honduras.
Keywords:
Non-ulcerated cutaneous leishmaniasis
; visceral leishmaniasis
; biological
; epidemiological
; clinical-immunological
; and molecular taxonomic analysis
; Leishmania (Leishmania) poncei
; Honduras
; Central America
1. Introduction
Visceral leishmaniasis, also known as “kala-azar”, is an infectious, non-contagious disease caused by leishmanine protozoan parasites of the order Kinetoplastea, family Trypanosomatidae, and genus Leishmania Ross 1903. These have all been grouped together into the Leishmania (Leishmania) donovani complex, which comprises of three valid species: Leishmania (L.) donovani Laveran & Mesnil 1903 (Africa/India), Leishmania (L.) infantum Nicolle 1908 (Europe/North Africa), and Leishmania (L.) chagasi Lainson & Shaw 1987 (=Leishmania chagasi Cunha & Chagas 1937) (Central and South America) [1,2,3,4,5,6].
In Latin America, the disease is known as American visceral leishmaniasis (AVL) or even as “neotropical calazar”, and its main vector is the phlebotomine species Lutzomyia longipalpis (Psychodidae: Phlebotominae) [1,7]. AVL mainly affects children ages one to ten years-old, however, it can also affect young adults, with males being the most involved. The clinical features of the disease may appear gradually or suddenly, and it usually begins with a daily fever that lasts for up to two months, generalized body weakness, indisposition, loss of appetite, weight loss, mucocutaneous pallor, and abdominal distension that occurs due to the progressive enlargement of the liver and spleen (hepatosplenomegaly). This hepatosplenomegaly occurs as a result of hyperplasia and hypertrophy of the mononuclear phagocytic system (SFM) in the parenchyma of these viscera. In the absence of proper diagnosis and treatment, this condition almost always leads to death. Pancytopenia (anemia, leukopenia, and thrombocytopenia) and suppression of T-cell immune response (mainly CD4/Th1 cells) are significant immunopathological disorders that are responsible for intercurrences such as bronchopulmonary and intestinal infections. When these intercurrences occur with hemorrhages that result from uncontrolled coagulation, they account for the deaths that occur in the advanced stages of the disease [8,9,10].
Although AVL can be considered to be the main clinical-immunological manifestation that results from interactions between the parasite and human immune system from a medical and/or public health point of view [11], it is worth noting that in some Central American countries such as Honduras, El Salvador, Nicaragua and Costa Rica, the infection may also present atypically in teenagers and young adults. This atypical picture may only compromise the skin (with no evidence of visceral involvement), where non-ulcerated lesions with a papular and/or nodular appearance may be seen disseminated on the face, pinnae, thorax, back, and upper and lower limbs evolving over months or even years. This was initially described as a new variant of cutaneous leishmaniasis from the infection due to “Leishmania donovani chagasi” by the pioneering work of Professor Ponce’s research group in the Amapala municipality on the west coast of Honduras (El Tigre Island) [12,13].
Almost simultaneously, this new clinical variant of cutaneous leishmaniasis, currently recognized as non-ulcerated cutaneous leishmaniasis (NUCL) or atypical dermal leishmaniasis (ADL), was also described in Costa Rica [14]; however, it was associated with another leishmanine parasite, “Leishmania infantum”, which led these authors to consider that the parasite had been introduced from Europe to the New World. It is interesting to note, however, that a few years later, a genotypical analysis using the kDNA RFLP and RAPD techniques to identify the causal agent of both clinical forms (NUCL and AVL) in Honduras confirmed that L. (L.) chagasi was the causal agent of both forms of the disease [15].
It is therefore necessary to point out that the clinical picture referred to as NUCL or ADL comprises of a clinical entity that is exclusively characterized by cutaneous manifestations (with no previous history of AVL) that are mainly represented by closed lesions of a typically papular and/or nodular appearance or infiltrated plaque (without evidence of a necrotizing-ulcerative process), measuring about 5–10 mm in diameter on average. They are usually surrounded by a hypochromic halo and are commonly disseminated over the face, trunk and limbs (upper and lower), evolving over months or even years and associated, until then, to the infection by L. (L.) chagasi (Figure 1) [12,13,15]. This clinical picture clearly differs from that of AVL, although in some occasions AVL patients may simultaneously present with skin lesions (sometimes even with an ulcerative character) that seem to represent a dermal manifestation of the systemic infection by L. (L.) chagasi [16,17].
Another condition worth mentioning here refers to what was considered the first case of “active cutaneous leishmaniasis” caused by Leishmania donovani chagasi in the New World [18]. This case was seen in a 35-year-old female patient residing in the municipality of Rio de Janeiro, Rio de Janeiro State, Brazil and presented as a typical ulcerated skin lesion (3 cm in diameter) on the left lower limb with almost three months of evolution. This clearly differs from the clinical picture referred to as NUCL or the variant of cutaneous leishmaniasis described by Ponce et al. [12,13]. More recently, another similar rare case caused by Leishmania (L.) infantum chagasi was diagnosed as “American cutaneous leishmaniasis (ACL)”. This case showed three ulcerated skin lesions (also morphologically different from those with a papular and/or nodular aspect or infiltrated plaque typical of NUCL) on the face of an elderly patient (over eighty-years-old) that had been evolving for about seven months. The patient was also from the municipality of Rio de Janeiro, Rio de Janeiro State, Brazil, where there was no previous record of ACL by Leishmania (V.) braziliensis [19].
Based on the comments above, a diagnostic approach in the original area (El Tigre Island, Amapala municipality, west coast of Honduras) that Ponce et al. [12,13] carried out their pioneering study on the variant of cutaneous leishmaniasis (today, NUCL) caused by L. (L.) chagasi [15] was recently began (2017). This approach sought to better understand the biological and molecular (genomic) characteristics of the Honduran leishmanine parasite, as well as the epidemiological aspects of human (and also canine) infection and its physiopathogenic (or immunopathogenic) mechanisms responsible for the largest clinical-immunological spectrum of that infection in Latin America [20]. The evidence gathered so far (which is the reason for the publication of this manuscript), based on taxonomic characters of a biological [21,22], epidemiological [23,24], clinical-immunological [20,25,26,27,28,29,30,31] and molecular (genomic/phylogenetic) [4,5 Figueroa et al. unpublished data; Vasconcelos dos Santos et al. unpublished data] nature related to this leishmanine parasite, point to the characterization of a new leishmanine species, Leishmania (Leishmania) poncei n. sp. [Kinetoplastea: Trypanosomatidae] (Figure 2A,B), in honor of Professor Carlos Ponce, who was the first researcher to describe NUCL in Honduras, Central America.
2. Integrative taxonomic analysis of the biological, epidemiological, clinical-immunological and molecular characters of the leishmanine parasite causing both NUCL and AVL in Honduras, Central America
2.1. Study area
All the activities that led to the realization of the aforementioned works werecarried out on El Tigre Island in the Amapala municipality, (N13◦ 15.618, W87◦ 37.463), Valle Department, on the west coast of Honduras in April 2017. The municipality comprises of two islands: Zacate Grande and El Tigre and is located in the Gulf of Fonseca in southwestern Honduras with a total of 28 villages and an estimated population of 13,302 individuals. The natural vegetation cover is dominated by a dry tropical forest. Temperature ranges between 25 ◦C and 35 ◦C, and average annual rainfall is 2096 mm. The dominant topography is mountainous and rugged with an average altitude of 44 m. The ecological landscape markedly changes between the dry season (November-May) and the rainy season (June-October) [20].
2.2. Parasite
Four strains of the Honduran parasite were used in the in vitro and in vivo experiments that gave support to this diagnostic approach. These parasites have been described as follows: MHOM/HND/2017/AMA-65, MHOM/HND/2017/AMA-73, MHOM/HND/2018/AMA-161, and MHOM/HND/2018/AMA-614/Isla del Tigre/Amapala municipality/Honduras. The parasites were isolated from four human cases of non-ulcerated cutaneous leishmaniasis (NUCL) [4,5,21,22] and hitherto characterized as Leishmania (L.) infantum chagasi using the PCR-RFLP through Hae III digestion of a Hsp70 fragment [20].
2.3. Genomic/phylogenetic analyses
Genomic analyses were performed as described elsewhere [4,5]. Moreover, the target strain sequences (MHOM/HD/2017/M32502) derived from de novo whole-genome assembly (GenBank accession number JAGRQD000000000) were analyzed together with representative sequences of different Leishmaniine species, covering subgenera/complexes from both the New and Old World. Two different partial gene sequences were analyzed: the large subunit of RNA Polymerase II (RNA Pol II) and the DNA Pol 𝛼. Alignment and tree building was performed using the MEGA software. Sequences were aligned using Clustal W and the Kimura 2-parameter model gave the best-fitting model of sequence evolution and was used for tree construction using the maximum likelihood (ML). Models with the lowest Bayesian information criteria were applied and ML trees were generated for each data set, bootstrapping with 1000 replicates. Porcisia hertigi was included as a closely related out group.
2.4. Biological taxonomic characters of the Honduran parasite
Although L. (L.) chagasi is well known as the only etiological agent of AVL in Latin America [1,2,3,4,5,6,32], it is also responsible for the atypical NUCL that is observed in some Central American countries, such as Honduras, El Salvador, Nicaragua, and Costa Rica. Thus, even if the genetics of the immune response of infected individuals residing in these countries was considered to be the main factor that acts in sense of directing the infection to the skin, it is highly unlikely that the immune response would be able to solely orchestrate this atypical behavior of the infection. It therefore seems important to remember the crucial role of the species-specific Leishmania antigens in determining the outcome of the human T-cell immune response, preferentially towards the CD4+/Th1 or CD4+/Th2 poles, as has been shown in the Leishmania species of the subgenera L. (Viannia) and L. (Leishmania), respectively that act as etiological agents of American cutaneous leishmaniasis in the Brazilian Amazon [33,34,35,36]. After considering this guideline, we tried to evaluate some biological markers of L. (L.) chagasi strains that were isolated from NUCL of Honduras and compare them with those isolated from AVL and canine visceral leishmaniasis (CVL) of Brazil.
In vitro study
This experiment examined the biochemical and functional roles of lipophosphoglycans (LPGs), the major cell surface glycoconjugate of the Leishmania parasite, which have the ability to protect the parasites from complement-mediated lysis and facilitate its attachment and entry into macrophages and also inhibit phagolysosomal fusion and protein kinase C [37,38]. In this sense, the following dermotropic strains of L. (L.) chagasi: MHOM/HND/2017/AMA-65, HOM/HND/2017/AMA-73, MHOM/HND/2018/AMA-161, and HOM/HND/2018/AMA-614 isolated from NUCL were used during in vitro interaction with murine macrophages (BALB/c and C57BL/6) and compared with viscerotropic strains from AVL and CVL of Brazil (MHOM/BR/1970/BH46 and MCAN/BR/89/BA262). LPGs were extracted, purified, and their repeat units analyzed. This made it possible to demonstrate an unusual fact in these dermotropic strains of the Honduran parasite, i.e., they had no side chains consisting of Gal (β1,4) Man (α1)-PO4 that is common to all LPGs. The interaction of LPG from these dermotropic strains of the Honduran parasite with BALB/c and C57BL/6 peritoneal macrophages with a view to producing nitric oxide (NO) and cytokines (TNF-α and IL-6) revealed that LPGs from these dermotropic strains of the Honduran parasite were able to elicit higher levels of NO and cytokines compared to the viscerotropic strains (AVL and CVL) of Brazil. This led to the conclusion that LPG of the dermotropic strains of L. (L.) chagasi of Honduras is devoid of side chains and exhibits high proinflammatory activity [21].
In vivo study
To better understand the biological behavior of the Honduran parasite, hamsters were infected subcutaneously and intraperitoneally with L. (L.) chagasi promastigotes that were isolated from NUCL (MHOM/HND/2017/AMA-65 and MHOM/HND/2017/AMA-73) and AVL (MHOM/HND/2020/LV-3) patients in the stationary growth phase. After 15, 30, 60, 90, and 120-days post-infection, the parasite-load, histopathological changes, and cytokines production at the subcutaneous and visceral (liver and spleen) sites of infection were analyzed. Animals inoculated subcutaneously did not develop macroscopic skin lesions, and only few histopathological changes in the dermal layer at the inoculation point were observed, regardless of the cutaneous or viscerotropic strains used. Conversely, histopathological findings in the viscera revealed portal mononuclear inflammatory infiltrates and hepatic nodules, in addition to intense macrophage infiltrates in the spleen, which progressively increased during the course of the infection. The parasite load in the viscera also increased progressively, regardless of the route of inoculation. Apart from IL-10, which was evident in the infected animals, there was no other observed increase or decrease in the production of proinflammatory or anti-inflammatory cytokines when compared to the healthy control group. In this sense, L. (L.) chagasi strains isolated from patients with NUCL or AVL from Honduras were able to cause visceral lesions in hamsters, regardless of the route of inoculation [22].
2.5. Epidemiological taxonomic characters of infection of the Honduran parasite
Regarding the epidemiological character of the parasite, two studies are worthy of note, as they observed two situations that reflect the distinct epidemiological character of the parasite under analysis in relation to that (L. (L.) chagasi) responsible for AVL in endemic areas of South America, especially in Brazil.
Vector study
The first study carried out a survey on the sand fly fauna in the area (Honduran Pacific coast); however, it is necessary to remember that previous evidence had already revealed that Lutzomyia (Lutzomyia) longipalpis and Pintomyia (Pifanomyia) evansi were the most prevalent species found in the area [39,40]. These evidences also recognized these phlebotomine species as important vectors of AVL in other geographic regions of Latin America, but not in Honduras [7,41,42,43]. Conversely, it is also important to mention that both AVL and NUCL are endemic on the Pacific versant of the Central American Pacific, where NUCL is the most frequently seen clinical form of the disease [20]. Collections of sand flies were carried out on five consecutive nights (May, 2018), making it possible to capture 222 sand fly specimens of six different species. Lutzomyia longipalpis (81%) and Pintomyia evansi (16%) were the most prevalent species of sand flies seen. L. (L.) chagasi DNA was detected in nine of the 96 specimens analyzed. Seven of these specimens were identified as Lutzomyia longipalpis, while the remaining two were Pintomyia evansi with infection rates of 9.4% and 2.7%, respectively. Finally, this represented the first record of L. (L.) chagasi DNA in Pintomyia evansi (Psychodidae: Phlebotominae) in an NUCL endemic region (El Tigre Island, Amapala municipality, Pacific coast of Honduras) of Central America. This strongly suggest that this sand fly species could be acting as a secondary vector of L. (L.) chagasi in the transmission cycles of both NUCL and AVL in Central America [23].
Reservoir study
Although Ponce’s pioneering work and other collaborators suggested that dogs could be a potential reservoir of L. (L.) infantum chagasi in the study area based on the 17% seroprevalence of infection in this animal [13], recent data obtained from this diagnostic approach has questioned this premise. This survey was carried out to analyze the behavior of natural canine L. (L.) chagasi infection in NUCL endemic areas and compare it with that of the infection in AVL endemic areas of South America, using Brazil, where the domestic dog is highly susceptible to this infection, as a reference. This susceptibility is attributed to evolving with high parasite loads on the skin, which gives it the status of AVL’s domestic reservoir, in addition to presenting with a high death rate from canine visceral leishmaniasis (CVL) [44,45,46,47,48]. For the purpose of the study, 107 dogs residing with patients with NUCL were clinically examined, and their biological materials were collected with a view of a parasitological and immunological diagnosis. The vast majority of dogs were apparently asymptomatic, although some presented with weight loss (64%), alopecia (7%), onychogryphosis (5%) and skin lesions (1%). The overall seroprevalence of canine infection based on the DDP® quick test and/or in-house ELISA was 41%. A molecular diagnosis (PCR) of infection was confirmed in 94% of the dogs; however, the average parasite load in the buffy coat was only 6,09 parasites/μL (0.221 and 50.2). Furthermore, the histopathological analysis of the skin of seropositive dogs did not reveal any significant dermal change that was suggestive of inflammatory processes compatible with leishmaniasis or other parasites. These findings, unlike those of CVL in South America, mainly in Brazil, seem to suggest that the domestic dog is not a good source of infection for the phlebotomine sand fly vector in the NUCL endemic area in the west coast of Honduras [24].
2.6. Clinical-immunological taxonomic characters of infection by the Honduran parasite
The clinical-immunological taxonomic characters of L. (L.) chagasi infection in the NUCL endemic areas were also the target of this diagnostic approach, with the objective of trying to elucidate the mechanisms of the parasite’s interaction with the human immune response, and consequently, the possible clinical-immunological outcomes resulting from this interaction. It also sought to compare them with those already known that result from the interaction of the parasite (L. (L.) chagasi) with the human immune response in AVL endemic areas in South America, mainly in Brazil.
Immunopathological features of in situ skin lesion caused by NUCL
The first work studied the nature of the inflammatory response in the skin lesions caused by NUCL and sought to better understand the pathophysiologic (or immunopathogenic) mechanisms that are involved in the pathogenesis of the typical closed skin lesion with a papular and/or nodular aspect or infiltrated plaque that is commonly seen in NUCL. For this purpose, biopsies taken from skin lesions of 20 patients with NUCL received conventional histological technical treatment, including hematoxylin and eosin (H&E) staining. The main histopathological findings revealed a mononuclear inflammatory infiltrate in the dermal layer that ranged from mild to intense and were composed mainly of lymphocytes, macrophages (with mild parasitism), and few plasmocytes. Furthermore, there were epithelioid granulomatous reactions in 60% of the studied cases, which suggested that the inflammatory response in NULC skin lesions (although of the closed type and, therefore, not ulcerated) is mainly composed of a substrate of a cellular immune nature, given the high rate (60%) of the epithelioid granulomatous reactions in the skin lesions [27].
However, if the typical NUCL skin lesion is closed with a papular and/or nodular appearance or infiltrated plaque, how can one explain that the epithelioid granulomatous character of the inflammatory reaction, probably supported by a well-developed cellular immune substrate, is unable to externalize a necrotizing reaction with a consequent ulcerative process of the skin lesion? In an attempt to answer this question, the immune response of the skin lesion was examined in situ to explore the possible regulatory mechanisms that are involved in the pathogenesis of NUCL. This was done by investigating the anti-CD4, anti-FoxP3, anti-IL-10, and anti-TGF-β antibodies as target markers for this immunohistochemical analysis. These immunological markers (CD4+, FoxP3+, IL-10+, and TGF-β+) were observed in the dermal layer with inflammatory infiltrates and higher densities (P< 0.05) in all 20 of the NUCL skin lesion biopsies studied when compared to the normal skin controls. Moreover, a strong positive correlation between CD4+ and FoxP3+ cells and a moderate positive correlation between FoxP3+ and TGF-β+ was observed; however, this was not seen with IL-10+ cells. These findings suggest that T-regulatory FoxP3+ cells and the regulatory cytokines, mainly TGF-β+, play an important role in the pathogenesis of NUCL by modulating a moderate cellular immune response in skin lesions and avoiding tissue damage (a necrotizing reaction), thus, leading to low tissue parasitic persistence [25].
Following the analysis of the immune response in NUCL skin lesions made in situ, the presumed role of Th17 lymphocytes in the pathogenesis of the skin lesions was then investigated, taking into account previous findings that showed that CD8+ T-cells were the main lymphocyte population present in dermal infiltrates of NUCL, followed by CD4+ T-cells, which have been correlated with IFN-γ+ cells. The 20 biopsies of the NUCL skin lesions were again submitted for immunohistochemical analysis, and the anti-ROR-γt, anti-IL-17, anti-IL-6, anti-TGF-β, and anti-IL-23 antibodies were used to characterize the role of Th17 lymphocytes in the pathogenesis of NUCL skin lesion. All these immunologic markers: ROR-γt+, IL-17+, IL-6+, TGF-β+, and IL-23+ were observed in the dermal inflammatory infiltrates of NUCL skin lesions at higher (P< 0.05) densities when compared to normal skin controls. A positive correlation between CD4+ T-lymphocytes, ROR-γt+, and IL-17+ cells suggested that some CD4+ T-cells may be acting as Th17 lymphocytes. Furthermore, a positive correlation between ROR-γt+ cells and TGF-β+, IL-6+, IL-17+, and IL-23+ cells could also indicate the role of these cytokines in the differentiation and maintenance of Th17 lymphocytes in skin lesions, favoring a protective cellular immune action against infection [26].
It is high time to investigate the role of TCD4/CD8, B-lymphocytes, macrophages, NK cells, nitric oxide synthase (iNOS), and IFN-γ in the pathogenesis of NUCL through immunohistochemical analysis. The biopsies of the skin lesions that were taken from patients with NUCL were submitted for specific labeling with anti-CD4+ and anti-CD8+ T-lymphocytes, anti-B lymphocytes, anti-NK cells, anti-macrophages, anti-iNOS, and anti-IFN-γ antibodies. The immunohistochemical analysis of the samples revealed a higher expression of these immunological markers in the dermal layer of NUCL skin lesions compared to those of healthy individuals (P< 0.05). In addition, morphometric analysis performed on sections of the skin lesions showed a predominance of CD8+ T-lymphocytes in the mononuclear inflammatory infiltrate, followed by macrophages, mostly iNOS+, which indicates an overt cellular immune response that is most likely mediated by IFN-γ. These findings reinforce the crucial role of IFN-γ in mediating the cellular immune response in NUCL skin lesion and also highlight the pivotal role of CD8+ T-lymphocytes in the host’s defense mechanism against the parasite [28].
After considering the evident protective role of the cellular immune response that is capable of controlling the intramacrophagic multiplication of the parasite in NUCL, the phenotypic subpopulations of macrophages in those skin lesions were evaluated in situ. This was because it is already well known that macrophages play an important role in the innate and/or acquired immune response against the Leishmania parasite and that depending on its subset and/or activation status there may be an elimination or, otherwise, favorable environment for the intracellular multiplication of the Leishmania parasite. In this sense, the fate of the parasite can be influenced by the phenotype of the infected macrophage, i.e., linked to the classically activated resistant M1 macrophage, or alternatively, to the susceptible activated M2 macrophage. M1 and M2 macrophage subsets were examined by double-staining immunohistochemical analysis of the biopsies of skin lesions of patients with ACL caused by Leishmania (L.) amazonensis (anergic diffuse cutaneous leishmaniasis (ADCL): 5 cases, and localized cutaneous leishmaniasis (LCL): 4 cases, Leishmania (V.) panamensis (LCL): 10 cases, Leishmania (V.) braziliensis (LCL): 5 cases, as well as by L. (L.) chagasi (NCUL): 10 cases. The results obtained demonstrated a higher (P< 0.05) density of M1 macrophages (112 ± 12 cells/mm2) than those of M2 macrophages (43 ± 12 cells/mm2) in the NUCL caused by L. (L.) chagasi. Conversely, higher densities (P< 0.05) of M2 macrophage (616 ± 114 and 219 ± 29 cells/mm2) compared to those of M1 macrophage (195 ± 25 and 97 ± 24 cells/mm2) were observed in skin lesions of ADCL and LCL caused by L. (L.) amazonensis, respectively, and also in LCL caused by L. (V.) panamensis (M2= 164 ± 14 > M1= 71 ± 14). However, in the LCL caused by L. braziliensis, there were no differences in cell densities between M1 and M2 macrophages (M1= 50 ± 13, M2= 53 ± 10). These findings clearly suggest that polarization of M1 and M2 macrophages is strongly influenced by the dichotomy between Th1 and Th2-type cellular immune responses, i.e., in those clinical forms of the disease where the cellular immune response is predominantly of the Th2-type, as seen in the ADCL and LCL caused by L. (L.) amazonensis. In the LCL caused by L. (V.) panamensis, the expression of the macrophage cell density of the M2 phenotype was more prevalent, while in the NUCL, where the cellular immune response is predominantly of the Th1-type, the expression of cellular density of the M1 macrophage phenotype was more prevalent. However, although there is more expression of Th1-type cellular immune response than that of the Th2 type in the LCL caused by L. (V.) braziliensis, there has been no expression of a higher cellular density between M1 and M2 macrophages [29].
In order to further expand this knowledge, it was therefore analyzed the role of antigen-presenting cells in the pathogenesis of the NUCL through the double-staining immunohistochemical analysis using cellular and intracellular markers. In this sense, twenty-three skin biopsies from NUCL patients were examined, together with specific monoclonal antibodies against Langerhans cells, dermal dendritic cells, CD4 and CD8 T-lymphocytes, IL-12, IFN-γ, TNF-α and IL-10 cytokines, in addition to iNOS. The most significant results include the high expression of both dermal and Langerhans dendritic cells, together with that of pro-inflammatory IL-12 cytokine, greater (P< 0.05) than those of IFN-γ, TNF-α, iNOS, and IL-10. In addition, the activation of cellular immune response was marked by higher density of CD8 Tc1-lymphocytes followed by CD4 Th1-lymphocytes producing mainly IFN-γ. These findings suggest that antigen-presenting cells (both dermal and Langerhans dendritic cells) play a crucial role in the activation of immune response through the high production of pro-inflammatory cytokines, mainly IL-12, directing cellular immune response preferentially to Th1 and Tc1 types in the NUCL caused by L. (L.) chagasi in Honduras [30].
Clinical-immunological spectrum of the Honduran parasite-infection
Following the findings of the immune response in the skin lesions of patients in situ, we sought to understand how the humoral and cellular immune responses could be contributing to the pathogenesis of NUCL in a systemic way. As a result, 80 serum samples were selected in order to evaluate the cytokine and immunoglobulin profiles in patients with NUCL (n= 47), AVL (n= 5), and negative control group (n= 28) based on previous clinical and laboratory diagnosis of the infection. Cytokine profiles were evaluated using a Cytokine Bead Array (CBA) Human Th1/Th2/Th17 kit. IgG and IgM classes and IgG1 and IgG2 subclasses of immunoglobulins were evaluated by ELISA using species-specific L. (L.) chagasi antigens. The serum concentration of TNF-α, IFN-γ, IL-2, and IL-4 cytokines in the NUCL, AVL, and control group were below the detection threshold of the CBA kit. IL-6, IL-10, and IL-17A cytokine concentrations were lower (P< 0.05) in patients with NUCL compared to those with AVL. With regards to the immunoglobulin profile, it was noticed that patients with NUCL presented with 4.0 times more IgG than the control group, while patients with AVL presented with 6.6 times more IgG. There was a 1.6 times higher expression in NUCL and a 2.6 times higher expression in AVL patients than in the control group for IgM. Concerning the immunoglobulin subclass, only patients with AVL showed a positive expression of IgG1. There was no positive expression of IgG2 among the studied clinical immunological groups. Therefore, these findings suggest that there is a weak cellular and humoral systemic immune response in NUCL [31].
The final diagnostic approach of the Honduran leishmanine parasite did not exactly explore the main clinical-immunological manifestations of the infection, i.e., the variant of cutaneous leishmaniasis by Ponce et al. [12,13], however, the entire clinical-immunological spectrum of this infection represents, in fact, the broadest clinical-immunological spectrum hitherto known for a single Leishmania species. For this purpose, 576 individuals residing in the study area were examined from a clinical, parasitological, immunological [humoral (ELISA-IgG/IgM) and cellular (DTH skin test)] point of view, with the aim of making a clinical-laboratorial diagnosis of symptomatic and asymptomatic infections. All symptomatic infections (NUCL) were confirmed by parasitological diagnosis (104), and the prevalence rate of NUCL was 18%. Among the asymptomatic individuals examined (472), 320 showed reactivity for one or both immunological tests (ELISA-IgG/IgM, DHT, or both), generating a prevalence rate of 55.50% for asymptomatic infections. However, when these two categories of infection were associated: symptomatic (104) and asymptomatic (320), a higher overall prevalence rate 73.6% (424 cases/576 individuals) of infection was achieved. In addition, among the 320 asymptomatic infected individuals, there were identified three clinical-immunological profiles: (1) Indeterminate asymptomatic infection (IAI): IgG+/IgM+/DTH-, IgG+/IgM-/DTH-, and IgG-/IgM+/DTH-; (2) Resistant asymptomatic infection (RAI): IgG+/IgM+/DTH+, IgG+/IgM-/DTH+, and IgG-/IgM+/DTH+; and (3) Final asymptomatic infection (FAI): IgG-/IgM-/DTH+. On the other hand, among the 104 individuals with symptomatic infection (NUCL) [with positive parasitological diagnosis] it was possible to identify four clinical-immunological profiles: (1) Early symptomatic infection (ESI): IgM-/IgG-/DTH-; (2) Indeterminate symptomatic infection (ISI): IgG+/IgM+/DTH-, IgG+/IgM-/DTH-, and IgG-/IgM+/DTH-; (3) Resistant symptomatic infection (RSI): IgG+/IgM+/DTH+, IgG+/IgM-/DTH+, and IgG-/IgM+/DTH+; and (4) Final symptomatic infection (FSI): IgG-/IgM-/DTH+. As can be seen, this comprises the broadest clinical-immunological spectrum induced by a single Leishmania species [L. (L.) chagasi] identified so far, not counting the classic AVL that has not been registered in the study area for over ten years [20].
2.7. Molecular (genomic) taxonomic characters of the Honduran parasite
To finalize this integrative taxonomic analysis on the Honduran leishmanine agent of NUCL, or eventually of AVL, it is important to consider the main actor in the scenario under analysis, i.e., the parasite, which until recently has been recognized as L. (L.) chagasi/L. (L.) infantum chagasi [4,5]. Considering previous evidence that indicated close proximity of the New World leishmanine viscerotropic agent (L. (L.) chagasi) with that from the Old World (L. (L.) infantum) from a molecular point of view [49,50,51,52], the main target among these parasites was analyzed using the comparative genomic analysis, which focused mainly on their ancestry, genomic structure (number of chromosomes, number of reads/contigs, GC content, identity of the genome for each chromosome, and number of SNPs) and phylogeny in order to clarify their specific character. Therefore, after taking this guideline into account, an analysis was made by making molecular clock comparisons of the DNA polymerase alpha subunit gene of the whole genome sequence of L. (L.) chagasi isolated from the NUCL of Honduras (Central America) and compared with that of the same parasite isolated from the wild canid, Cerdocyon thous, of Brazil (South America), L. (L.) donovani (India), and L. (L.) infantum (Europe).
This showed that the Honduran parasite is significantly more ancient (382,800 ya) than that of Brazil (143,300 ya), L. (L.) donovani (33,776 ya), and L. (L.) infantum (13,000 ya) [5]. Furthermore, structural genomic comparisons among these leishmanine parasites were also carried out and mainly analyzed the variations in genomes and the number of genomic SNPs for each chromosome. This demonstrated that the Honduran parasite presents an absolutely unprecedented structural variation on chromosome 17, as well as the highest frequency of genomic SNPs (more than twice the number seen in the Brazilian parasite) [5].
Regarding the phylogeny of these parasites, it is important to note that the phylogenetic analysis using PCR examining three mitochondrial genetic targets of the maxi circle: 12S, 9S (highly conserved ribosomal), and ND7 in a sample of four isolates from the same clinical form (NUCL) and geographic area (El Tigre island, Amapala municipality, west coast of Honduras) of the disease revealed that the Honduran parasite (Central America) represents a different lineage from that of the Brazilian parasite (South America) [Figueroa-Fernandez et al. unpublished data]. Moreover, phylogenetic reconstructions using partial DNA and RNA polymerase gene sequences showed that the target strain (Honduran parasite) not only clustered in the subgenus L. (Leishmania) complex donovani, but also slightly diverged from that of the L. infantum (European)/L. chagasi (South American) types with well-supported bootstrap values. This divergence was most pronounced in the NJ-RNA Pol II tree, where the target strain diverged not only from that of the L. infantum (European)/L. chagasi (South American), but also from L. donovani (India) taxa (Figure 3A,B) [Vasconcelos dos Santos et al. unpublished data].
When taken together, these findings represent strong evidence that L. (L.) chagasi/L. (L.) infantum chagasi from Brazil (South America) is, undoubtedly, native to the New World and therefore, has a valid taxonomic status. Conversely, it was also demonstrated that the Honduran parasite; the most ancestral viscerotropic leishmanine parasite, showed biological, epidemiological, clinical-immunological, and genomic/phylogenetic taxonomic characters that are compatible with a new Leishmania species that causes both clinical forms of leishmaniases (NUCL and AVL) in Honduras (Central America).
Leishmania (Leishmania) poncei n. sp.
Taxonomic summary
Class Kinetoplastea Honigberg, 1963 emend., Vickerman, 1976
Order Trypanosomatida Kent, 1880 stat. nov. Hollande, 1952
Family Trypanosomatidae Doflein, 1951
Genus Leishmania Ross 1903
Subgenus L. (Leishmania) Ross 1903
Type host: Homo sapiens
Type locality and ecological features: El Tigre Island, Amapala municipality, Honduras (N13◦ 15.618, W87◦ 37.463).
Type material: Hapantotypes, cryopreserved promastigotes stored in liquid nitrogen at the Faculty of Medicine, University of São Paulo, Brazil and in the Ralph Lainson Leishmaniasis Lab., Instituto Evandro Chagas (IEC)/ Ministry of Health, Brazil.
Strain designation: MHOM/HND/2017/AM-65 (USP); M32502 (IEC).
Homologous strains: MHOM/HND/2017/AM-73, MHOM/HND/2018/AMA-161, and MHOM/HND/2018/AMA-614, all isolated from human cases of non-ulcerated cutaneous leishmaniasis (NUCL) [4,5,21,22].
Vector: Probably Lutzomyia (Lutzomyia) longipalpis and Pintomyia (Pifanomyia) evansi.
Reservoir(s): Unknown.
Representative DNA sequences: The de novo whole-genome assemblies and raw data have been deposited in the GenBank and SRA databases. The GenBank accession number is JAGRQD000000000 and the SRA accession number is SRR8608748, under BioProject number PRJNA722301 [4].
Zoobank registration: Available only after the acceptance.
Etymology: The species is named in honor of Professor Carlos Ponce (in memoriam), who was the first researcher to describe NUCL in Honduras, Central America.
Behavior in vitro: The parasite shows rich growth in the Schneider medium supplemented with 10% fetal bovine serum, 10_g/mL of 1% L-glutamine, and 100 IU/mL ampicillin.
Pathology: Non-ulcerated cutaneous leishmaniasis (NUCL) and American visceral leishmaniasis (AVL).
3. Discussion
This represents a unique integrative taxonomic analysis of the biological, epidemiological, clinical-immunological and molecular (genomic/phylogenetic) characters of the Honduran leishmanine parasite responsible for both clinical forms of leishmaniases (NUCL and AVL) in Honduras, Central America. This is quite different from other studies that have recently described new Leishmania species of the subgenus L. (Mundinia) Shaw, Camargo and Teixeira 2016, such as, Leishmania (Leishmania) martiniquensis [=Leishmania (Mundinia) martiniquensis emend: Shaw, Camargo and Teixeira 2016], Leishmania (Mundinia) orientalis, Leishmania (Mundinia) chancei and Leishmania (Mundinia) procaviensis [53,54,55,56], i.e., all representatives of a new taxonomic status within a subgenus that was recently described and, therefore, generating new taxonomic expectations. In contrast, the last valid species of the subgenus L. (Leishmania) that was described was Leishmania (L.) waltoni Shaw, Pratrong, Floeter-Winter, et al. 2015, a new causal agent of anergic diffuse cutaneous leishmaniasis (ADCL) in the Dominican Republic (Central America) after almost three decades following the first parasite isolates [57]. It does not seem like a very difficult task to understand the meaning of the present work, since more than three decades after the original description of NUCL in Honduras, Central America [12], the question that is still heard is whether this atypical clinical form of cutaneous leishmaniasis has the same causal agent as the visceral form of the disease (AVL). This implies that NUCL is just a clinical manifestation arising from man’s resistant immune response to this infection or that its etiological agent really has a specific taxonomic characteristic that not only differentiates it from the AVL agent (L. (L.) chagasi) currently recognized in Latin America, but also that it could be clinically translated by the atypical dermatological manifestation of the infection known as NUCL. As mentioned here, since the work of Noyes et al. [15] that addressed this issue by comparing some parasite strains isolated from the cutaneous (NUCL) and visceral (AVL) forms using molecular techniques (kDNA RFLP and RAPD), there has already been an almost conclusive answer suggesting that the etiological agent was the same in both forms of the disease. Therefore, the most reasonable answer is that NUCL seems to be the main clinical manifestation arising from man’s resistant immune response to the infection, as NUCL represents the most prevalent clinical form of the infection in Honduras [20].
It is important to make it clear that NUCL is a clinical condition that is frequently seen in the studied area (El Tigre Island, Amapala municipality, west coast of Honduras) and other Central American countries such as El Salvador, Nicaragua, and Costa Rica, rather than a sporadic clinical condition such as the ulcerative cutaneous leishmaniasis (~ACL) that is caused by L. (L.) chagasi in Brazil, which has only been reported on a few occasions [18,19,58]. Although there is previous evidence that suggests that the causal agent of NUCL and AVL is the same (i.e., both clinical forms had L. (L.) chagasi as the causal agent) [15], it is difficult to explain that the same causal agent is capable of inducing such diverse clinical conditions, i.e., the frequent NUCL seen in Central America and the sporadic ulcerative cutaneous leishmaniasis (~ACL) in South America. The different clinical characteristics of these two clinical conditions; on one hand, NUCL in Central America with clearly closed skin lesions of papular and/or nodular appearances or infiltrated plaques, and the ulcerative cutaneous leishmaniasis in South America on the other hand, seems to strongly signal that the etiologic agents involved (i.e., the one mainly responsible for NUCL and, less often, for AVL in Central America, and the other mainly responsible for AVL and, rarely, for ulcerative cutaneous leishmaniasis in South America) are, in fact, distinct parasitological entities with specific taxonomic characters, i.e., Leishmania (Leishmania) poncei, a new Leishmania sp. in honor of Professor Ponce, who was the first researcher to describe the atypical NUCL in Honduras, and Leishmania (L.) chagasi Lainson & Shaw 1987 (=Leishmania chagasi Cunha & Chagas 1937) mainly responsible for AVL and, less often, for ulcerative cutaneous leishmaniasis in Brazil (South America). This is shown by the integrative, taxonomic analysis of the biological, epidemiological, clinical-immunological, and molecular (genomic/phylogenetic) characteristics of the Honduran leishmanine parasite and confirms the specific taxonomic status of the causal agent of both clinical forms of leishmaniases (NUCL and AVL) in Honduras.
Let us consider the series of studies that took place in this integrative taxonomic analysis, starting with the analysis of the biological taxonomic character of the Honduran parasite [L. (L.) poncei]. These studies revealed two findings that deserve to be highlighted; firstly, on the biochemical and functional role of the main glycoconjugate molecule on the surface of Leishmania parasite, the lipophosphoglycan (LPG). This showed, in an unprecedented way, that the LPG molecule of the parasite, unlike other Leishmania species, does not present side chains consisting of Gal (β1,4)Man(α1)-PO4 that is common to all LPGs. Furthermore, with regards to in vitro L. (L.) poncei LPG interactions with macrophages from BALB/C and C57BL6 mice in view of producing nitric oxide (NO), TNF-α, and IL-6 cytokines, it was noticed that L. (L.) poncei LPG was able to elicit higher levels of NO and cytokines compared to L. (L.) chagasi strains that were isolated from cases of human (AVL) and canine visceral leishmaniasis (CVL) from Brazil. This was evidence that L. (L.) poncei LPG has a higher ability of producing proinflammatory activity [21].
Secondly, on the behavior of experimental L. (L.) poncei infection in hamsters inoculated with strains isolated from NUCL and AVL cases of Honduras through the subcutaneous and intraperitoneal routes, it was revealed that regardless of the route of inoculation, both L. (L.) poncei strains caused progressive viscerotropism and a similar parasite load in the viscera of both groups of inoculated animals (there was no observed development of skin lesions in animals inoculated subcutaneously) during the period of the experimental infection. This demonstrates that regardless of the route of inoculation, L. (L.) poncei strains isolated from NUCL and AVL from Honduras were capable of promoting visceral lesions in hamsters [22]. Without a doubt, these findings represent strong evidence on the individuality of the biological taxonomic character of L. (L.) poncei native to Central America, which is quite different from that of the other viscerotropic species, such as L. (L.) chagasi, which originally occurs in South America. This reveals that although the viscerotropism of the Honduran parasite can be expressed in an immunologically vulnerable host, such as a hamster or young man, i.e., under five years of age, dermal tropism seems to show an advanced biological stage of evolution and/or adaptation of the parasite in the human host substantiated by the higher prevalence of symptomatic infection in the skin (NUCL) than in the viscera (AVL) [20].
Regarding the epidemiological taxonomic character of L. (L.) poncei, it is worth highlighting two other findings that reflect this issue. In this case, we are talking about the main sand fly species involved in the transmission of the parasite, as well as the behavior of canine L. (L.) poncei infection in the studied area. Regarding the sand fly species, Lutzomyia longipalpis (81%) and Pintomyia evansi (16%) have been previously identified as the most prevalent species [39,40], with L. (L.) poncei infection rates of 9.4% and 2.7% detected by PCR respectively [23]. It is still too early to say what these findings represent even in terms of the vector capacity of these sand fly species for transmitting L. (L.) poncei. Considering previous records of the natural infection of these sand fly species by the closest Leishmania species, i.e., L. (L.) chagasi [41,42,43], it seems likely that Lutzomyia longipalpis and Pintomyia evansi play an important role in the transmission of L. (L.) poncei infection, which also represents the first record of this finding in Honduras, Central America.
Another epidemiological parameter that was used as a comparison between these two closely related parasitological entities, i.e., L. (L.) chagasi versus L. (L.) poncei, was the behavior of canine infection in the endemic area. As already well documented, canine L. (L.) chagasi infection (CVL) in AVL endemic areas in South America, using Brazil as a reference area, has systematically revealed that the domestic dog is highly susceptible to this infection, evolving with a high parasitic load on the skin, regardless of its clinical status (symptomatic or not), which gives it the status of domestic CVL reservoir, in addition to presenting a high mortality rate from the disease [44,45,46,47,48,59]. However, unlike what has been previously recorded in CVL by L. (L.) chagasi in Latin America, mainly in AVL endemic areas in Brazil (South America), canine L. (L.) poncei infection in the endemic area of NUCL in the present study has, from a clinical point of view, proven to be almost absolutely unnoticed such that there were no dogs with typical signs or reported cases of dog deaths from suspected CVL cases before the diagnostic approach of the canine infection in that area was carried out. It was possible to detect some clinical signs of canine infection such as weight loss (64%), alopecia (7%), onychogryphosis (5%), and skin lesions (1%) after requesting specialized veterinary professional support. Thus, although we have confirmed by immunological (seroprevalence of canine infection with DDP® quick test and/or in-house ELISA was 41%) and molecular (positive PCR in 94% of the dogs but with a very low parasite load in the buffy coat) parameters, the presence of canine L. (L.) poncei infection (but without histological evidence of the parasite and of an inflammatory process in the skin lesions) denotes that the pathogenesis of canine L. (L.) poncei infection is undoubtedly much less severe than that caused by L. (L.) chagasi in South America, mainly in Brazil. There is also a lower parasite load at the site of infection, mainly in the skin, which shows that there is a lower risk of dogs serving as a source of infection for the sand fly species that are potential vectors (Lutzomyia longipalpis and Pintomyia evansi) of the parasite in the NUCL endemic area of Honduras, Central America [24].
Therefore, although we have emphasized the epidemiological taxonomic character of canine L. (L.) poncei infection, it can also be seen that there is a crossing of this aspect with the pathogenesis of canine infection, which clinically reveals marked differences between canine infections by these two taxonomically distinct parasites, i.e., L. (L.) poncei and L. (L.) chagasi. Due to the clinical characteristics of canine and of human infections, it is possible to observe that L. (L.) poncei is capable of causing a clinical condition (NUCL) that can be recognized as closed skin lesions of the papular and/or nodular types or infiltrated plaques. Conversely, L. (L.) chagasi induces a clearly ulcerated skin lesion that has been compared to the picture of American cutaneous leishmaniasis (ACL). In line of these differences in the clinical character of infection between L. (L.) poncei and L. (L.) chagasi, it is worth remembering that the taxonomic classification of the genus Leishmania is phylogenetically well defined and can also be described in terms of host specificity (vertebrate and/or invertebrate) and clinical parameters [60].
In addition to the clinical characteristic of L. (L.) poncei infection, we could not fail to comment on its clinical-immunological taxonomic character, as both clinical and immunological characters go together. Due to the dynamics of the infection, this diagnostic approach considered the in situ immune response in NUCL skin lesion and the systemic humoral and cellular immune responses of the infection. It is necessary to emphasize the histopathological findings that border the inflammatory response in skin lesions that is represented by a mononuclear inflammatory infiltrate in the dermal layer ranging from mild to intense and composed mainly of lymphocytes, macrophages (with mild parasitism), and a few plasma cells, but with epithelioid granulomatous reaction in 60% of the cases studied prior to considering the in situ immune response of the main clinical manifestation of the infection. This suggests that the inflammatory response of the NUCL skin lesion (although of the closed papular and/or nodular type and, therefore, not the ulcerated type) seems to contain a significant substrate of a cellular immune nature, given the high rate (60%) of the epithelioid granulomatous reaction present in the lesion [27]. Following this histopathological approach, several factors inherent to the immunopathological mechanisms that were expected to take part in the pathogenesis of NUCL were analyzed. This made it possible to understand that the development of NUCL is sustained by a consistent cellular immune response against the parasite. This was represented by high expressions of CD8/CD4 T-cells and Th17 cells in such a way that the parasite load within infected macrophages (mainly the M1 phenotype) was visibly low. This shows a strong association with the Th1-type protective cellular immune response that is aided by significant production of the proinflammatory cytokines IL-2, TNF-α, INF-γ, and iNOS. In contrast, this is regulated by expressions of FoxP3, IL-10, and TGF-β without compromising the success of NUCL therapy with intralesional or systemic meglumine antimoniate [25,26,28,29,30]. Finally, although this condition does not configure a specific immune response against a particular Leishmania species or against L. (L.) poncei, it seems feasible to say that it would be in line with the suspicion of NUCL, especially when it is associated with the clinical characteristics of the disease.
Conversely, when analyzing the systemic, humoral, and cellular immune responses against L. (L.) poncei infection based on a clinical, parasitological, and immunological diagnostic support, a lower (P< 0.05) expression of regulatory cytokines: IL-6, IL-10, and IL-17 was evident, with a lower (P< 0.05) expression of IgG and IgM in NUCL compared to AVL. This therefore suggests that NUCL has lower expressions of systemic, humoral and cellular immune responses compared to AVL [31]. Furthermore, these same diagnostic parameters (clinical, parasitological, and immunological: ELISA-IgG/IgM and DHT) made it possible to identify the broadest clinical-immunological spectrum known so far for a single Leishmania species. This composed of three clinical-immunological profiles among the asymptomatic infected individuals [(1) Indeterminate asymptomatic infection (IAI): IgG+/IgM+/DTH-, IgG+/IgM-/DTH-, and IgG /IgM+/DTH-; (2) Resistant asymptomatic infection (RAI): IgG+/IgM+/DTH+, IgG+/IgM-/DTH+, and IgG-/IgM+/DTH+; and (3) Final asymptomatic infection (FAI): IgG-/IgM-/DTH+] and another four among the symptomatic infected ones (NUCL) [(1) Early symptomatic infection (ESI): IgM-/IgG-/DTH-; (2) Indeterminate symptomatic infection (ISI): IgG+/IgM+/DTH-, IgG+/IgM-/DTH-, and IgG-/IgM+/DTH-; (3) Resistant symptomatic infection (RSI): IgG+/IgM+/DTH+, IgG+/IgM-/DTH+, and IgG-/IgM+/DTH+; and (4) Final symptomatic infection (FSI): IgG-/IgM-/DTH+], which allows tracking and monitoring of the evolution of the infection in the endemic area [21], as has been done in the Brazilian Amazon [11,61].
Ultimately, a genomic analysis was used to consolidate the convictions regarding the significance of differences in the clinical nature of infections between L. (L.) poncei and L. (L.) chagasi, as this was, in fact, the major reason for carrying out this integrative taxonomic analysis of the Honduran leishmanine parasite (Central America). Part of this analysis, especially that related to the genomics and phylogenetics of the parasite, has also confirmed the thoughts regarding the autochthonous nature of L. (L.) chagasi in American territory [1,2,3,4,5,6] based on the simple ecological observation of the existence of primitive enzootic cycles of L. (L.) chagasi in the Brazilian Amazon (South America) in wild mammals, particularly small rodents (Proechimys sp., Proechimys cuvieri and Dasyprocta sp.) or canid (Cerdocyun thous). These animals carry the parasite inside the tropical forests in the states of Amapá (in the Wajãpi Indigenous territory, a rugged landscape covered by dense rainforest which is one of the world’s largest protected rainforests) and Pará (in an iron mining project in the Serra dos Carajás, municipality of Marabá, where the entry of domestic animals, especially dogs, is expressly prohibited). This represents irrefutable evidence of the autochthony of L. (L.) chagasi on the American soil [62,63].
Three findings are worth mentioning here: first, it was clearly demonstrated by molecular clock comparative analysis of the DNA polymerase alpha subunit gene of the genome sequence of L. (L.) poncei that was isolated from the NUCL that the Honduran parasite is significantly older (382,800 years) than L. (L.) chagasi of Brazil (143,300 years), L. (L.) donovani (33,776 years), and L. (L.) infantum (13,000 years) [4]. Second, structural genomic comparisons among these leishmanine parasites were also carried out and revealed that the Honduran parasite presents an unprecedented structural variation on chromosome 17 as well as the highest frequency of genomic SNPs (more than twice the number seen in the Brazilian parasite) [5]. Third, the phylogenetic analyses: 1) based on the PCR of three mitochondrial genetic targets of the maxi circle 12S and 9S (highly conserved ribosomal) and ND7 as has also been used to reconstruct the evolutionary relationships within the Trypanosomatidae family [64], in a sample of four isolates from the NUCL (one of which was used in the molecular clock and genomic structure analysis) revealed that the Honduran parasite represents a different lineage from that of the Brazilian parasite (Figueroa-Fernandez et al. unpublished data) and; 2) based on DNA and RNA polymerase gene sequences that showed that L. (L.) poncei is clustered in the subgenus L. (Leishmania) complex donovani and is also mainly diverged in the NJ-RNA Pol II tree from the L. infantum (European)/L. chagasi (South American) and L. donovani (India) taxa (Vasconcelos dos Santos et al. unpublished data).
4. Conclusions
In conclusion, following the aforementioned findings of this integrative taxonomic analysis, it seems quite consistent to propose that the new leishmanine parasite species, i.e., Leishmania (Leishmania) poncei, represents the causal agent of the two clinical forms of leishmaniases (NUCL and AVL) in Honduras, Central America from a clinical-epidemiological and/or public health point of view.
Author Contributions
Conceptualization: FTS, MDL; Data curation: FTS, TVS, GVAF, CMSP, ECS, WSO, CZV, VLM, CMCG, PKR, LVL, MBC, CEPC, MDL; Funding acquisition: FTS, MDL; Investigation: FTS, TVS, GVAF, CMSP, ECS, WSO, CZV, VLM, CMCG, PKR, LVL, MBC, CEPC, MDL; Methodology: FTS, TVS, GVAF, CMSP, ECS, WSO, CZV, VLM, CMCG, PKR, LVL, MBC, CEPC, MDL; Project administration: FTS, MDL; Resources: FTS, MDL; Software: ECS, TVS; Supervision: FTS, MDL; Validation: ECS, TVS; Visualization: FTS, MDL; Writing – original draft preparation: FTS, MDL; Writing – Review & Editing: FTS, TVS, GVAF, CMSP, ECS, WSO, CZV, VLM, CMCG, PKR, LVL, MBC, CEPC, MDL. All authors read and approved the final version of the manuscript.
Funding
This work was supported by Evandro Chagas Institute (Surveillance Secretary of Health and Environment, Ministry of Health, Brazil); Tropical Medicine Nucleus (Federal University of Pará, Brazil) and São Paulo Research Foundation #2016/50315-0 (São Paulo State, Brazil).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors thank to Ministry of Health of Honduras for allowing and assisting in the work in the endemic area, excellent technical assistance of Jessica Cardenas in endemic area of south Honduras, Raimundo Nonato Pires and Rodrigo Ribeiro Furtado for their laboratory assistance in the cultivation of the Honduran leishmanine parasite [Leishmania (L.) poncei] submitted to genomic sequencing at the Evandro Chagas Institute (Surveillance Secretary of Health and Environment, Ministry of Health), Pará State, Brazil.
Conflicts of Interest
The authors have no any conflict of interest concerning this work.
References
- Lainson: R.; Shaw, J.J. New World Leishmaniasis. In Topley & Wilson’s microbilogy & microbial infections: parasitology.; Hodder Arnold ASM Press, Ed.; London, 2005; pp. 313–349.
- Lainson, R. The Neotropical Leishmania Species: A Brief Historical Review of Their Discovery, Ecology and Taxonomy. Rev Panamazonica Saude 2010, 1, 13–32. [CrossRef]
- Silveira, F.T.; Corbett, C.E.P. Leishmania Chagasi Cunha & Chagas, 1937: Indigenous or Introduced? A Brief Review. Rev Panamazonica Saude 2010, 1, 143–147. [CrossRef]
- Silveira, F.T.; Junior, E.C.S.; Silvestre, R.V.; Costa-Martins, A.G.; Pinheiro, K. da C.; Ochoa, W.S.; Santos, T.V. dos; Ramos, P.K.; Casseb, S.; Silva, S.P. da; et al. Whole-Genome Sequencing of Leishmania Infantum Chagasi Isolates from Honduras and Brazil. Microbiol Resour Announc 2021, 10, 1–3. [CrossRef]
- Silveira, F.T.; Sousa Junior, E.C.; Silvestre, R.V.D.; Vasconcelos dos Santos, T.; Sosa-Ochoa, W.; Valeriano, C.Z.; Ramos, P.K.S.; Casseb, S.M.M.; Lima, L.V. do R.; Campos, M.B.; et al. Comparative Genomic Analyses of New and Old World Viscerotropic Leishmanine Parasites: Further Insights into the Origins of Visceral Leishmaniasis Agents. Microorganisms 2023, 11, 25. [CrossRef]
- Marcili, A.; Sperança, M.A.; da Costa, A.P.; Madeira, M. de F.; Soares, H.S.; Sanches, C. de O.C.C.; Acosta, I. da C.L.; Girotto, A.; Minervino, A.H.H.; Horta, M.C.; et al. Phylogenetic Relationships of Leishmania Species Based on Trypanosomatid Barcode (SSU RDNA) and GGAPDH Genes: Taxonomic Revision of Leishmania (L.) Infantum Chagasi in South America. Infection, Genetics and Evolution 2014, 25, 44–51. [CrossRef]
- Lainson, R.; Rangel, E.F. Lutzomyia Longipalpis and the Eco-Epidemiology of American Visceral Leishmaniasis, with Particular Reference to Brazil: A Review. Mem Inst Oswaldo Cruz 2005, 100, 811–827. [CrossRef]
- Silveira, F.T.; Lima, L.V. do R.; Santos, T.V. dos; Ramos, P.K.S.; Campos, M.B. Reviewing the Trajectory of American Visceral Leishmaniasis in Brazilian Amazon/Revendo a Trajetória Da Leishmaniose Visceral Americana Na Amazônia, Brasil: De Evandro Chagas Aos Dias Atuais. Rev Pan-Amaz Saude 2016, 15–22.
- Silveira, F.T.; Lainson, R.; de Souza, A.A.A.; Crescente, J.A.B.; Corbett, C.E.P. Leishmaniose Visceral Americana. In Medicina Tropical e Infectologia na Amazônia; Leão, R., Samauma, Eds.; Belem, Brazil, 2013; Vol. 2, pp. 1245–1274.
- Pearson, R.D.; Sousa, A. de Q. Clinical Spectrum of Leishmaniasis. Clinical Infectious Diseases 1996, 22, 1–13. [CrossRef]
- Furtado, R.R.; Alves, A.C.; Lima, L.V.R.; Vasconcelos Dos Santos, T.; Campos, M.B.; Ramos, P.K.S.; Gomes, C.M.C.; Laurenti, M.D.; da Matta, V.L.; Corbett, C.E.; et al. Visceral Leishmaniasis Urbanization in the Brazilian Amazon Is Supported by Significantly Higher Infection Transmission Rates Than in Rural Area. Microorganisms 2022, 10. [CrossRef]
- Ponce, C.; Ponce, E.; Morrison, A.; Cruz, A.; Neva, F. Leishmaniasis Cutânea Em Honduras Causada Por Leishmania Donovani Chagasi. XVI Congreso Centroamericano de Dermatología Y II Congreso Costarricense de Dermatología. Programa y Libro de Resúmenes, 1988, 73.
- Ponce, C.; Ponce, E.; Morrison, A.; Cruz, A.; Kreutzer, R.; McMahon-Pratt, D.; Neva, F. Leishmania Donovani Chagasi: New Clinical Variant of Cutaneous Leishmaniasis in Honduras. Lancet 1991, 337, 67–70. [CrossRef]
- Zeledón, R.; Hidalgo, H.; Víquez, A.; Urbina, A. Atypical Cutaneous Leishmaniasis in a Semiarid Region of North-West Costa Rica. Trans R Soc Trop Med Hyg 1989, 83, 786. [CrossRef]
- Noyes, H.; Chance, M.; Ponce, C.; Ponce, E.; Maingon, R. Leishmania Chagasi: Genotypically Similar Parasites from Honduras Cause Both Visceral and Cutaneous Leishmaniasis in Humans. Exp Parasitol 1997, 85, 264–273. [CrossRef]
- Vasconcelos, I. de A.; Sousa, A. de Q.; Vasconcelos, A.W.; Diógenes, M.J.; Momen, H.; Grimaldi Júnior, G.; Menezes, D.B.; Sleigh, A.C. Cutaneous Parasitism by Leishmania (Leishmania) Chagasi during South American Visceral Leishmaniasis. Bulletin de la Société de pathologie exotique 1933, 86, 101–105.
- Moura, C.R.L. de P.; Costa, C.H.N.; Moura, R. de D.; Braga, A.R.F.; Silva, V.C.; Costa, D.L. Cutaneous Parasitism in Patients with American Visceral Leishmaniasis in an Endemic Area. Rev Soc Bras Med Trop 2020, 53, e20190446. [CrossRef]
- Oliveira Neto, M.P.; Grimaldi, G.; Momen, H.; Pacheco, R.S.; Marzochi, M.C.; McMahon Pratt, D. Active Cutaneous Leishmaniasis in Brazil, Induced by Leishmania Donovani Chagasi. Mem Inst Oswaldo Cruz 1986, 81, 303–309.
- Lyra, M.R.; Pimentel, M.I.F.; Madeira, M. de F.; Antonio, L. de F.; Lyra, J.P. de M.; Fagundes, A.; Schubach, A. de O. First Report of Cutaneous Leishmaniasis Caused by Leishmania (Leishmania) Infantum Chagasi in an Urban Area of Rio de Janeiro, Brazil. Rev Inst Med Trop Sao Paulo 2015, 57, 451–454. [CrossRef]
- Sosa-Ochoa, W.; Zúniga, C.; Chaves, L.F.; Araujo Flores, G.V.; Sandoval Pacheco, C.M.; Ribeiro da Matta, V.L.; Pereira Corbett, C.E.; Tobias Silveira, F.; Dalastra Laurenti, M. Clinical and Immunological Features of Human Leishmania (L.) Infantum-Infection, Novel Insights Honduras, Central America. Pathogens 2020, 9, 554. [CrossRef]
- Cardoso, C.A.; Araujo, G. V.; Sandoval, C.M.; Nogueira, P.M.; Zúniga, C.; Sosa-Ochoa, W.H.; Laurenti, M.D.; Soares, R.P. Lipophosphoglycans from Dermotropic Leishmania Infantum Are More Pro-Inflammatory than Those from Viscerotropic Strains. Mem Inst Oswaldo Cruz 2020, 115, 1–6. [CrossRef]
- Araujo Flores, G. V.; Sandoval Pacheco, C.M.; Ferreira, A.F.; Tomokane, T.Y.; Nunes, J.B.; Colombo, F.A.; Sosa-Ochoa, W.H.; Zúniga, C.; Silveira, F.T.; Corbett, C.E.P.; et al. Leishmania (L.) Infantum Chagasi Isolated from Skin Lesions of Patients Affected by Non-Ulcerated Cutaneous Leishmaniasis Lead to Visceral Lesion in Hamsters. Parasitol Int 2023, 93, 102723. [CrossRef]
- Sosa-Ochoa, W.; Varela Amador, J.; Lozano-Sardaneta, Y.; Rodriguez Segura, G.; Zúniga Valeriano, C.; Araujo, G.V.; Sandoval Pacheco, C.M.; Laurenti, M.D.; Galvis-Ovallos, F. Detection of Leishmania Infantum DNA in Pintomyia Evansi and Lutzomyia Longipalpis in Honduras. Parasit Vectors 2020, 13, 593. [CrossRef]
- Segura, G.B.R.; Ochoa, W.H.S.; Matta, V.L.R.D.; Martínez, M.; Tercero, C.R.; Gonzalez, R.R.; Pacheco, C.M.S.; Flores, G.V.A.; Silveira, F.T.; Henriquez, M.M.R.; et al. Can Domestic Dogs Be Considered a Good Reservoir of Leishmania (L.) Infantum Chagasi in an Endemic Area of Nonulcerated Cutaneous Leishmaniasis in Southern Honduras? Rev Inst Med Trop Sao Paulo 2023, 65. [CrossRef]
- Araujo Flores, G.V.; Sandoval Pacheco, C.M.; Tomokane, T.Y.; Sosa Ochoa, W.; Zúniga Valeriano, C.; Castro Gomes, C.M.; Corbett, C.E.P.; Laurenti, M.D. Evaluation of Regulatory Immune Response in Skin Lesions of Patients Affected by Nonulcerated or Atypical Cutaneous Leishmaniasis in Honduras, Central America. Mediators Inflamm 2018, 2018, 1–7. [CrossRef]
- Araujo Flores, G.V.; Sandoval Pacheco, C.M.; Sosa Ochoa, W.; Castro Gomes, C.M.; Zúniga, C.; Corbett, C.P.; Laurenti, M.D. Th17 Lymphocytes in Atypical Cutaneous Leishmaniasis Caused by Leishmania (L.) Infantum Chagasi in Central America. Parasite Immunol 2020, 42, 1–6. [CrossRef]
- Sandoval Pacheco, C.M.; Araujo Flores, G.V.; Favero Ferreira, A.; Sosa Ochoa, W.; Ribeiro da Matta, V.L.; Zúniga Valeriano, C.; Pereira Corbett, C.E.; Dalastra Laurenti, M. Histopathological Features of Skin Lesions in Patients Affected by Non-Ulcerated or Atypical Cutaneous Leishmaniasis in Honduras, Central America. Int J Exp Pathol 2018, 99, 249–257. [CrossRef]
- Sandoval, C.; Araujo, G.; Sosa, W.; Avalos, S.; Silveira, F.; Corbett, C.; Zúniga, C.; Laurenti, M. In Situ Cellular Immune Response in Non-Ulcerated Skin Lesions Due to Leishmania (L.) Infantum Chagasi Infection. Journal of Venomous Animals and Toxins including Tropical Diseases 2021, 27, 1–8. [CrossRef]
- Sandoval Pacheco, C.M.; Araujo Flores, G. V.; Gonzalez, K.; De Castro Gomes, C.M.; Passero, L.F.D.; Tomokane, T.Y.; Sosa-Ochoa, W.; Zúniga, C.; Calzada, J.; Saldaña, A.; et al. Macrophage Polarization in the Skin Lesion Caused by Neotropical Species of Leishmania Sp. J Immunol Res 2021, 2021, 1–8. [CrossRef]
- Sandoval Pacheco, C.M.; Araujo Flores, G. V.; Ferreira, A.F.; da Matta, V.L.R.; de Castro Gomes, C.M.; Sosa-Ochoa, W.H.; Zúniga, C.; Silveira, F.T.; Corbett, C.E.P.; Laurenti, M.D. Role of Antigen-presenting Cells in Non-ulcerated Skin Lesions Caused by Leishmania (Leishmania) Infantum Chagasi. Parasite Immunol 2023, e12971. [CrossRef]
- Laurenti, M.D.; Sosa-Ochoa, W.; Araujo Flores, G.V.; Sandoval Pacheco, C.M.; Tomokane, T.Y.; Oliveira, L.M. da S.; Zúniga, C.; Silveira, F.T.; Corbett, C.E.P. Evaluation of Systemic Immunity in Atypical Cutaneous Leishmaniasis Caused by Leishmania (L.) Infantum Chagasi. Parasite Immunol 2021, 00, 1–6. [CrossRef]
- Lainson, R.; Shaw, J.J. Evolution, Classification and Geographical Distribution. In The leishmaniases in biology and medicine. Volume I. Biology and epidemiology; Wallace Peters, R.K.-K., Ed.; Academic Press: London, 1987; p. 120 ISBN 0-12-552101-4.
- Silveira, F.T.; Lainson, R.; Corbett, C.E. Clinical and Immunopathological Spectrum of American Cutaneous Leishmaniasis with Special Reference to the Disease in Amazonian Brazil: A Review. Mem Inst Oswaldo Cruz 2004, 99, 239–251. [CrossRef]
- Silveira, F.T.; Lainson, R.; De Castro Gomes, C.M.; Laurenti, M.D.; Corbett, C.E.P. Immunopathogenic Competences of Leishmania (V.) Braziliensis and L. (L.) Amazonensis in American Cutaneous Leishmaniasis. Parasite Immunol 2009, 31, 423–431. [CrossRef]
- T. Silveira, F.; B. Campos, M.; F. Müller, S.; K. Ramos, P.; V. Lima, L.; V. dos Santos, T.; Maria Gomes, C.; D. Laurenti, M.; Lucia da Matta, V.; Eduardo Corbett, C. From Biology to Disease: Importance of Species-Specific Leishmania Antigens from the Subgenera Viannia (L. Braziliensis) and Leishmania (L. Amazonensis) in the Pathogenesis of American Cutaneous Leishmaniasis . In Leishmania Parasites; IntechOpen: Croatia, 2023; pp. 1–29 ISBN 978-1-83768-311-6.
- Silveira, F.T. What Makes Mucosal and Anergic Diffuse Cutaneous Leishmaniases so Clinically and Immunopathogically Different? A Review in Brazil. Trans R Soc Trop Med Hyg 2019, 113, 505–516. [CrossRef]
- Giorgione, J.R.; Turco, S.J.; Epand, R.M. Transbilayer Inhibition of Protein Kinase C by the Lipophosphoglycan from Leishmania Donovani. Proc Natl Acad Sci U S A 1996, 93. [CrossRef]
- Späth, G.; Garraway, L.; Turco, S.; Beverley, S. The Role(s) of Lipophosphoglycan (LPG) in the Establishment of Leishmania Major Infections in Mammalian Hosts. Proc Natl Acad Sci U S A 2003, 100. [CrossRef]
- Sosa-Ochoa, W.; Morales Cortedano, X.; Argüello, S.; Zuniga, C.; Henríquez, J.; Mejía, R.; Mejía, A.; Araujo, G.; Sandoval, C.; Quan, D. Ecoepidemiología de La Leishmaniasis Cutánea No Ulcerada En Honduras. Revista Ciencia y Tecnología 2015, 115–128. [CrossRef]
- Mejía, Á.; Matamoros, G.; Fontecha, G.; Sosa-Ochoa, W. Bionomic Aspects of Lutzomyia Evansi and Lutzomyia Longipalpis, Proven Vectors of Leishmania Infantum in an Endemic Area of Non-Ulcerative Cutaneous Leishmaniasis in Honduras. Parasit Vectors 2018, 11. [CrossRef]
- Chaves, L.F.; Añez, N. Nestedness Patterns of Sand Fly (Diptera: Psychodidae) Species in a Neotropical Semi-Arid Environment. Acta Trop 2016, 153. [CrossRef]
- Travi, B.L.; Vélez, I.D.; Brutus, L.; Segura, I.; Jaramillo, C.; Montoya, J. Lutzomyia Evansi, an Alternate Vector of Leishmania Chagasi in a Colombian Focus of Visceral Leishmaniasis. Trans R Soc Trop Med Hyg 1990, 84, 676–677. [CrossRef]
- Travi, B.L.; Montoya, J.; Gallego, J.; Jaramillo, C.; Llano, R.; Velez, I.D. Bionomics of Lutzomyia Evansi (Diptera: Psychodidae) Vector of Visceral Leishmaniasis in Northern Colombia. J Med Entomol 1996, 33, 278–285. [CrossRef]
- Moreira, M.A.B.; Luvizotto, M.C.R.; Garcia, J.F.; Corbett, C.E.P.; Laurenti, M.D. Comparison of Parasitological, Immunological and Molecular Methods for the Diagnosis of Leishmaniasis in Dogs with Different Clinical Signs. Vet Parasitol 2007, 145, 245–252. [CrossRef]
- Silveira, F.T.; Carneiro, L.A.; Ramos, P.K.S.; Chagas, E.J.; Lima, L.V.R.; Campos, M.B.; Laurenti, M.D.; Gomes, C.M.C.; Corbett, C.E.P. A Cross-Sectional Study on Canine Leishmania (L.) Infantum Chagasi Infection in Amazonian Brazil Ratifies a Higher Prevalence of Specific IgG-Antibody Response than Delayed-Type Hypersensitivity in Symptomatic and Asymptomatic Dogs. Parasitol Res 2012, 111, 1513–1522. [CrossRef]
- Laurenti, M.D.; Rossi, C.N.; Matta, V.L.R. da; Tomokane, T.Y.; Corbett, C.E.P.; Secundino, N.F.C.; Pimenta, P.F.P.; Marcondes, M. Asymptomatic Dogs Are Highly Competent to Transmit Leishmania (Leishmania) Infantum Chagasi to the Natural Vector. Vet Parasitol 2013, 196, 296–300. [CrossRef]
- Aschar, M.; de Oliveira, E.T.B.; Laurenti, M.D.; Marcondes, M.; Tolezano, J.E.; Hiramoto, R.M.; Corbett, C.E.P.; da Matta, V.L.R. Value of the Oral Swab for the Molecular Diagnosis of Dogs in Different Stages of Infection with Leishmania Infantum. Vet Parasitol 2016, 225, 108–113. [CrossRef]
- Carneiro, L.A.; Lima, L.V.; Campos, M.B.; dos Santos, T.V.; Ramos, P.K.; Laurenti, M.D.; Silveira, F.T. Prevalence and Incidence of Canine Visceral Leishmaniasis and Its Clinical-Immunological Features in an Endemic Area of the Brazilian Amazon. Vet. Med. Sci. 2023, 1–12.
- Maurício, I.; Stothard, J.; Miles, M. The Strange Case of Leishmania Chagasi. Parasitol Today 2000, 16. [CrossRef]
- Lukes, J.; Mauricio, I.; Schönian, G.; Dujardin, J.; Soteriadou, K.; Dedet, J.; Kuhls, K.; Tintaya, K.; Jirků, M.; Chocholová, E.; et al. Evolutionary and Geographical History of the Leishmania Donovani Complex with a Revision of Current Taxonomy. Proc Natl Acad Sci U S A 2007, 104. [CrossRef]
- Leblois, R.; Kuhls, K.; François, O.; Schönian, G.; Wirth, T. Guns, Germs and Dogs: On the Origin of Leishmania Chagasi. Infect Genet Evol 2011, 11, 1091–1095. [CrossRef]
- Kuhls, K.; Alam, M.Z.; Cupolillo, E.; Ferreira, G.E.M.; Mauricio, I.L.; Oddone, R.; Feliciangeli, M.D.; Wirth, T.; Miles, M.A.; Schönian, G. Comparative Microsatellite Typing of New World Leishmania Infantum Reveals Low Heterogeneity among Populations and Its Recent Old World Origin. PLoS Negl Trop Dis 2011, 5, e1155. [CrossRef]
- Desbois, N.; Pratlong, F.; Quist, D.; Dedet, J.-P. Leishmania (Leishmania) Martiniquensis n. Sp. (Kinetoplastida: Trypanosomatidae), Description of the Parasite Responsible for Cutaneous Leishmaniasis in Martinique Island (French West Indies). Parasite 2014, 21. [CrossRef]
- Espinosa, O.A.; Serrano, M.G.; Camargo, E.P.; Teixeira, M.M.G.; Shaw, J.J. An Appraisal of the Taxonomy and Nomenclature of Trypanosomatids Presently Classified as Leishmania and Endotrypanum. Parasitology 2016, 145, 430–442. [CrossRef]
- Jariyapan, N.; Daroontum, T.; Jaiwong, K.; Chanmol, W.; Intakhan, N.; Sor-suwan, S.; Siriyasatien, P.; Somboon, P.; Bates, M.D.; Bates, P.A. Leishmania (Mundinia) Orientalis n. Sp. (Trypanosomatidae), a Parasite from Thailand Responsible for Localised Cutaneous Leishmaniasis. Parasit Vectors 2018, 11, 351. [CrossRef]
- Kwakye-Nuako, G.; Mosore, M.-T.; Boakye, D.; Bates, P.A. Description, Biology, and Medical Significance of Leishmania (Mundinia) Chancei n. Sp. (Kinetoplastea: Trypanosomatidae) from Ghana and Leishmania (Mundinia) Procaviensis n. Sp. (Kinetoplastea: Trypanosomatidae) from Namibia. Journal of Parasitology 2023, 109, 43–50. [CrossRef]
- Shaw, J.; Pratlong, F.; Floeter-Winter, L.; Ishikawa, E.; El Baidouri, F.; Ravel, C.; Dedet, J.-P. Characterization of Leishmania (Leishmania) Waltoni n.Sp. (Kinetoplastida: Trypanosomatidae), the Parasite Responsible for Diffuse Cutaneous Leishmaniasis in the Dominican Republic. Am J Trop Med Hyg 2015, 93, 552–558. [CrossRef]
- Castro, L.S.; França, A. de O.; Ferreira, E. de C.; Hans Filho, G.; Higa Júnior, M.G.; Gontijo, C.M.F.; Pereira, A.A.S.; Dorval, M.E.M.C.; PEREIRA, A.A.S.; DORVAL, M.E.M.C. Leishmania Infantum as a Causative Agent of Cutaneous Leishmaniasis in the State of Mato Grosso Do Sul, Brazil. Rev Inst Med Trop Sao Paulo 2016, 58, 23. [CrossRef]
- Lima, L.V.R.; Carneiro, L.A.; Campos, M.B.; Chagas, E.J.; Laurenti, M.D.; Corbett, C.E.P.; Lainson, R.; Silveira, F.T. Canine Visceral Leishmaniasis Due to Leishmania (L.) Infantum Chagasi in Amazonian Brazil: Comparison of the Parasite Density from the Skin, Lymph Node and Visceral Tissues between Symptomatic and Asymptomatic, Seropositive Dogs. Rev Inst Med Trop Sao Paulo 2010, 52, 259–266. [CrossRef]
- Butenko, A.; Kostygov, A.Y.; Sádlová, J.; Kleschenko, Y.; Bečvář, T.; Podešvová, L.; Macedo, D.H.; Žihala, D.; Lukeš, J.; Bates, P.A.; et al. Comparative Genomics of Leishmania (Mundinia). BMC Genomics 2019, 20, 726. [CrossRef]
- da Matta, V.L.R.; Gonçalves, A.N.; Gomes, C.M.C.; Chouman, I.H.; Ferreira, F.M.; Campos, M.B.; Lima, L. V; Vasconcelos Dos Santos, T.; Ramos, P.K.; Furtado, R.R.; et al. Gene Signatures of Symptomatic and Asymptomatic Clinical-Immunological Profiles of Human Infection by Leishmania (L.) Chagasi in Amazonian Brazil. Microorganisms 2023, 11. [CrossRef]
- Vasconcelos dos Santos, T.; Ramos, P.K.S.; da Silva, F.M.M.; Alves, A.C.; Lima, L.V.R.; Campos, M.B.; Furtado, R.R.; Silveira, F.T. Presence of Lutzomyia Longipalpis (Diptera: Psychodidae) and Natural Leishmania (L.) Infantum Chagasi-Infection in the Wild Rodent, Proechimys Sp. (Rodentia: Echimydae), in the “Serra Dos Carajás”, Southern of Pará State, Brazil. In Proceedings of the 6th World Congress of Leishmaniasis, Poster Section, Toledo, Span 2016.
- Moreno, E.S.; Sabioni, L.A.; Seixas, M.M.M. de; Souza Filho, J.A. de; Marcelino, A.P.; Shimabukuro, P.H.F. Evidence of a Sylvatic Enzootic Cycle of Leishmania Infantum in the State of Amapá, Brazil. Rev Soc Bras Med Trop 2020, 53, e20190169. [CrossRef]
- Kaufer, A.; Stark, D.; Ellis, J. Evolutionary Insight into the Trypanosomatidae Using Alignment-Free Phylogenomics of the Kinetoplast. Pathogens 2019, 8. [CrossRef]
Figure 1.
A typical case of non-ulcerated cutaneous leishmaniasis (NUCL) in a 14-year-old female adolescent presenting a papular cutaneous lesion on the left hemiface, surrounded by a hypochromic halo with a three-month evolution. Photograph courtesy of Marcia Dalastra Laurenti.
Figure 1.
A typical case of non-ulcerated cutaneous leishmaniasis (NUCL) in a 14-year-old female adolescent presenting a papular cutaneous lesion on the left hemiface, surrounded by a hypochromic halo with a three-month evolution. Photograph courtesy of Marcia Dalastra Laurenti.
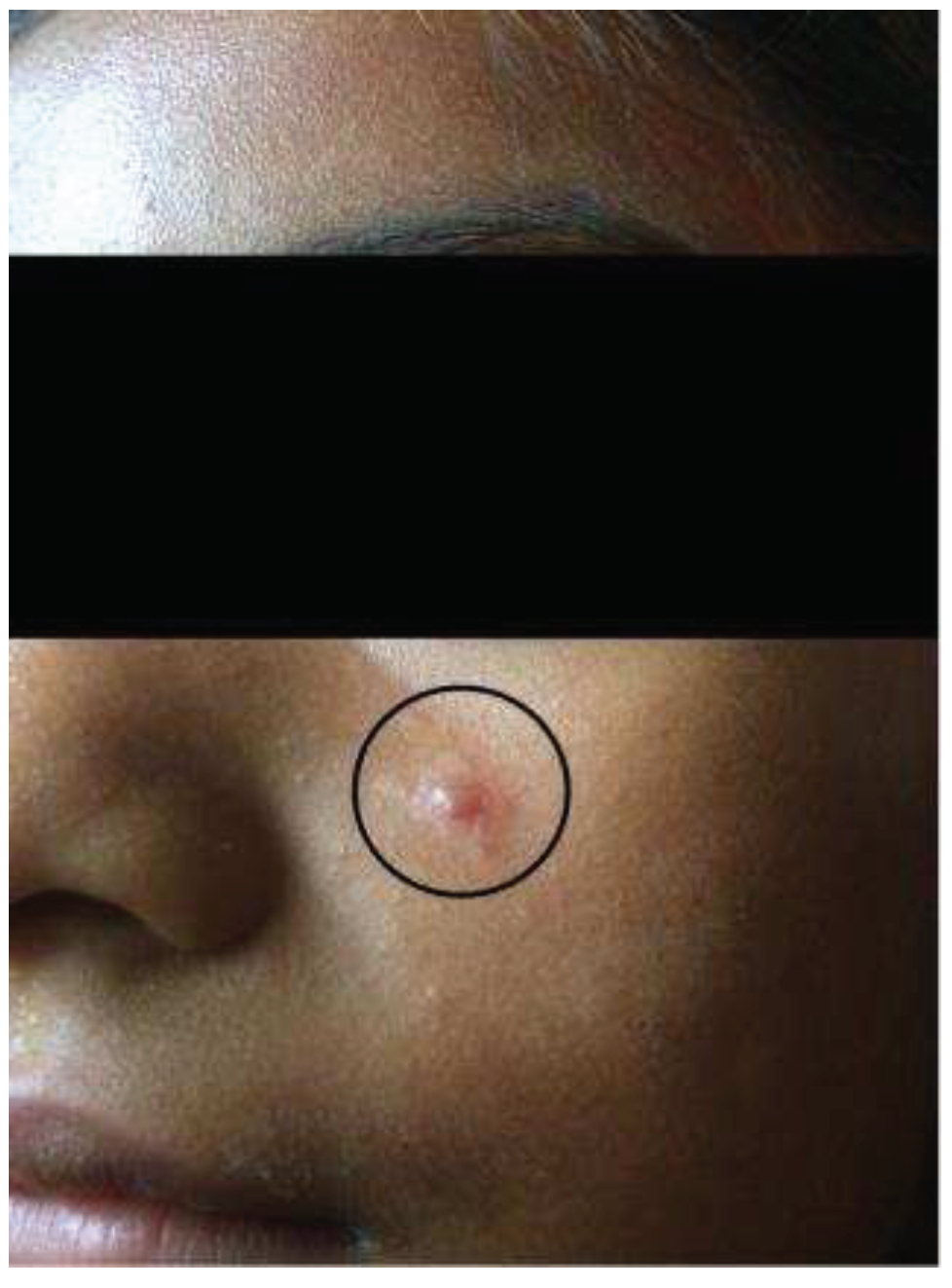
Figure 2.
A) Photomicrograph of Giemsa-stained amastigote forms of Leishmania (L.) poncei in spleen smear from a “hamster” experimentally infected with the parasite. B) Photomicrograph of Giemsa-stained promastigote forms of Leishmania (L.) poncei from Schneider culture medium. Scale-bar: 5 µm. Photographs courtesy of Gabriela V. Araujo Flores.
Figure 2.
A) Photomicrograph of Giemsa-stained amastigote forms of Leishmania (L.) poncei in spleen smear from a “hamster” experimentally infected with the parasite. B) Photomicrograph of Giemsa-stained promastigote forms of Leishmania (L.) poncei from Schneider culture medium. Scale-bar: 5 µm. Photographs courtesy of Gabriela V. Araujo Flores.
Figure 3.
A. Phylogenetic analysis of the alpha subunit of DNA Polymerase (DNA Pol alpha) sequences from Leishmania (Leishmania) poncei n. sp. analyzed together with representative sequences of L. (Leishmania) {1}, L. (Viannia) {2}, and L. (Mundinia) {3}, inferred by Maximum Likelihood (ML) and Neighbor-Joining (NJ) methods. Trees were generated for each data set, bootstrapping with 1,000 replicates. Porcisia hertigi was included as a closely related outgroup. B. Phylogenetic analysis of the large subunit of RNA Polymerase II (RNA Pol II) sequences from Leishmania (Leishmania) poncei n. sp. analyzed together with representative sequences of L. (Leishmania) {1}, L. (Viannia) {2}, L. (Mundinia) {3}, and L. (Sauroleishmania) {4}, inferred by Maximum Likelihood (ML) and Neighbor-Joining (NJ) methods. Trees were generated for each data set, bootstrapping with 1,000 replicates. Porcisia hertigi was included as a closely related outgroup.
Figure 3.
A. Phylogenetic analysis of the alpha subunit of DNA Polymerase (DNA Pol alpha) sequences from Leishmania (Leishmania) poncei n. sp. analyzed together with representative sequences of L. (Leishmania) {1}, L. (Viannia) {2}, and L. (Mundinia) {3}, inferred by Maximum Likelihood (ML) and Neighbor-Joining (NJ) methods. Trees were generated for each data set, bootstrapping with 1,000 replicates. Porcisia hertigi was included as a closely related outgroup. B. Phylogenetic analysis of the large subunit of RNA Polymerase II (RNA Pol II) sequences from Leishmania (Leishmania) poncei n. sp. analyzed together with representative sequences of L. (Leishmania) {1}, L. (Viannia) {2}, L. (Mundinia) {3}, and L. (Sauroleishmania) {4}, inferred by Maximum Likelihood (ML) and Neighbor-Joining (NJ) methods. Trees were generated for each data set, bootstrapping with 1,000 replicates. Porcisia hertigi was included as a closely related outgroup.
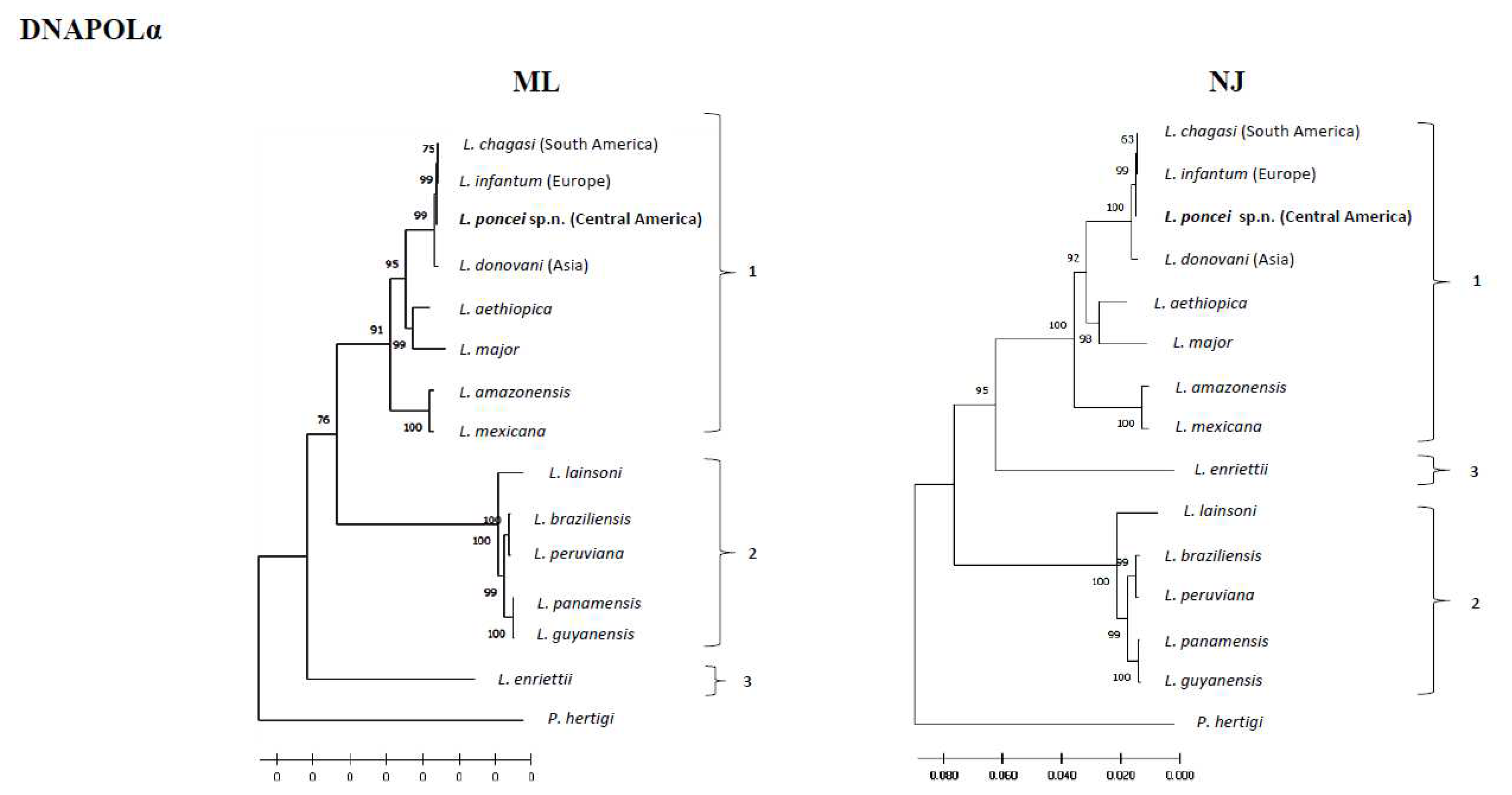
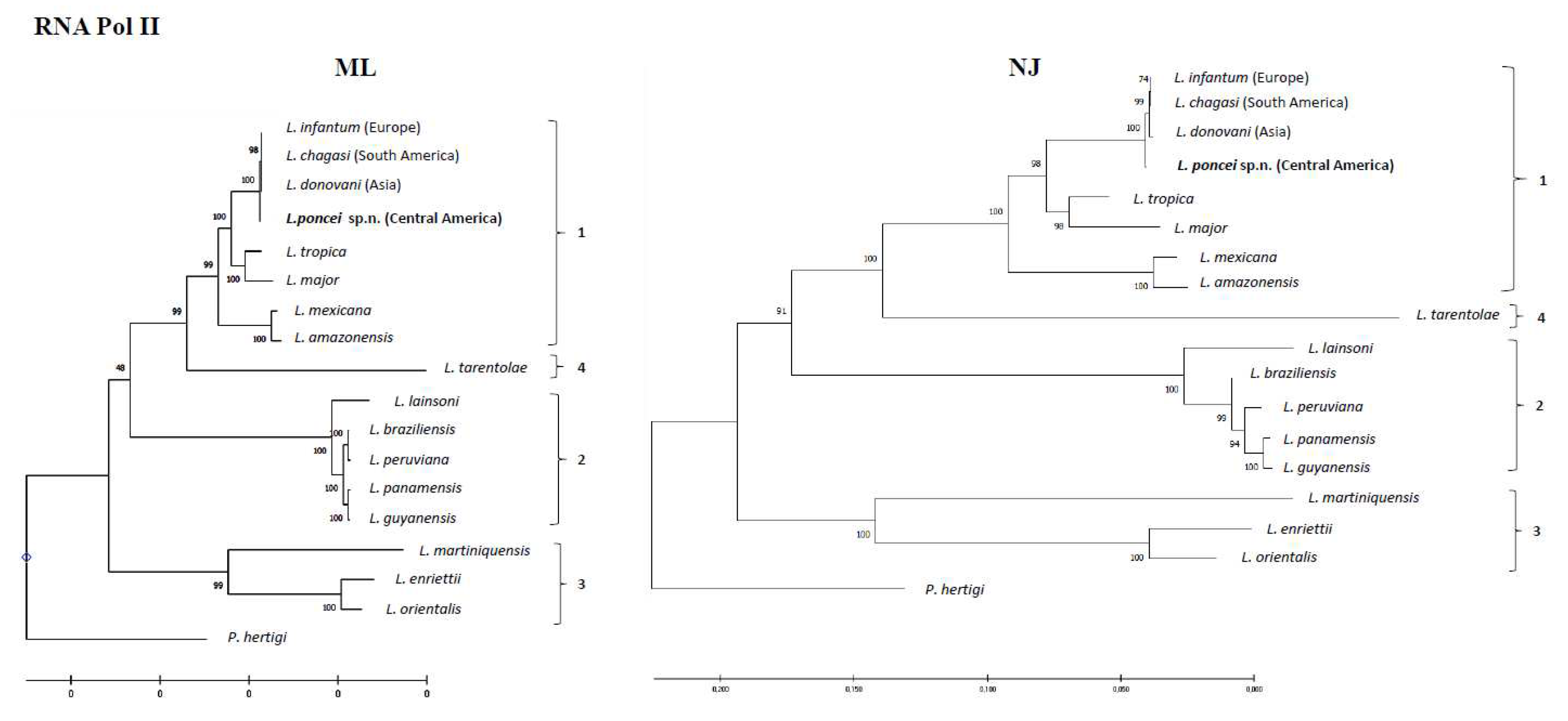
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



