
Submitted:
24 August 2023
Posted:
24 August 2023
You are already at the latest version
Abstract
Rapeseed (Brassica napus L.) is one of the most important oil crops in China. Improving oil production of rapeseed is an important way to ensure the safety of edible oil in China. Oil production is an important index to reflect the quality of rapeseed and is determined by oil content and yield. Applying nitrogen is an important way to ensure a high yield. However, increasing nitrogen application would decrease oil content. Thus, it is critical to screen elite germplasm resources with stable or improved oil content under high nitrogen and to investigate the molecular mechanisms of nitrogen regulation of oil accumulation. In this paper, we review the effects of nitrogen levels on rapeseed yields and oil content, aiming to reveal the mechanisms of nitrogen regulation in oil accumulation and thereby provide a theoretical basis for breeding varieties with high oil content.
Keywords:
rapeseed
; nitrogen
; yield
; oil content
; molecular mechanisms
1. Introduction
Rapeseed is one of the major oil crops in China. Oil production depends not only on the seed yield but also on the oil content [1]. Nitrogen, as a living element of plants, is particularly vital for their growth and development. Applying nitrogen is an important way to ensure a strong and stable yield. However, most varieties of rapeseed are extremely sensitive to nitrogen and have a reduced seed oil content after nitrogen application. The oil content decreased by 1.6% for every increase of 100 kg·hm-2 nitrogen application [2,3], and then equals a decrease of 4% in seed yield [3,4]. It is important to cultivate varieties of rapeseed that do not decrease in oil content after nitrogen application to ensure oil production. Moreover, the molecular mechanism of nitrogen-regulated oil accumulation in developing rapeseed is still poorly understood. Nitrogen levels affect the deposition of proteins and starch in maize and the accumulation of protein and oil in rapeseed [5,6,7]. Due to different target products, the mechanism of nitrogen on rapeseed is different from that of cereal crops such as maize and rice. It is important to study nitrogen’s effect on rapeseed grain yield and oil content.
2. Effects of Nitrogen on Plants
Sufficient nitrogen will insure large, vibrantly green leaves, additional branches, abundant flowers, and an increase in yield [8]. Excessive nitrogen can hinder the growth and development of plant roots. In Arabidopsis thaliana, lateral root elongation will be suppressed with increasing nitrogen concentration, while primary lateral root growth is insensitive to nitrogen [9]. Too much nitrogen can cause the stems and leaves to swell, reduce resistance to lodging and stress, and delay maturation time [7,10,11]. When nitrogen is insufficient, the organic matter in the plant will allocate to the root system priority and stimulate the growth of lateral roots [12]. Thus, the plant grows moderately, diminishes in size, gradually loses fresh leaves, becomes smaller, and has a lower chlorophyll content and redder leaves [13] and a reduction in the number of branches, flowers and grains [14]. The oil content of maize increases with the increase in nitrogen application rate [8]. In wheat, the effect of nitrogen on yield and protein yield is a quadratic curve [15]. In rapeseed, nitrogen can lead to a significant increase in yield, but can also cause a decrease in oil content. Protein, glucosinolate, and erucic acid content all tend to increase with higher nitrogen levels [1,7,10,16]. These results suggest different mechanisms by which nitrogen affects different plants.
3. Effects of Nitrogen on Rapeseed Yield
Nitrogen affects the growth and yields formation of rapeseed. It is reasonable to control the nitrogen application rate within 90~225kg·hm-2 [17,18,19]. Appropriate nitrogen can boost photosynthetic capacity, metabolic levels, morphogenesis, and increase yield [13]. The shortage of nitrogen can limit the photosynthesis ability, deplete the nutrient supply required for pod development, and reduce the count of productive pods on the main stem and the number of seeds per pod, therefore decreasing the weight per 1,000 grains and yielding [7,20]. Excessive nitrogen can induce over-growth during the vegetative stage and reduce nitrogen use efficiencies and yield during the maturity stage. This is because pre-flowering rapeseed consists primarily of vegetative morphogenesis, such as stems and leaves; After flowering, nutrients are transported to the grain [7]. An overabundance of nitrogenous fertilizers can cause stems and branches to produce an excessive number of ineffective shoots and branches, and lead to fertilizer waste and ecological destruction [14]. The siliques number per plant and seeds number per pod does not increase significantly with the nitrogen application rate beyond 180 kg·hm-2 [18].
3.1. Effects of Nitrogen on Photosynthetic Assimilation of Rapeseed
Nitrogen affects the efficiency of photosynthetic energy transport in leaves and then regulates the photosynthetic product. After nitrogen is assimilated, it is mainly transported to the mesophyll cells in the top of the plant through the xylem in the form of glutamine, aspartic acid, glutamic acid, and asparagine for carbon assimilation. Leaves are the primary organ of photosynthesis [14]. Chloroplasts are the sole site of photosynthetic carbon assimilation and the primary site of nitrogen assimilation in cells [21]. In chloroplasts, chlorophyll a and chlorophyll b have the function of collecting and transmitting sunlight energy [22]. The energy previously converted into chemical energy by sunlight is temporarily stored in ATP and NADPH for CO2 assimilation in dark reactions [23]. The way of CO2 assimilation is through the Calvin cycle pathway (C3) [24]. Triose phosphate (TP) is the net product of carbon assimilation that is assimilated through the pentose phosphate reduction pathway (RPPP) [14]. Further, TP is a major precursor to other biosynthetic reactions, including starch and sucrose biosynthesis, nitrogen and sulfur metabolism, fatty acid biosynthesis, cell wall biosynthesis, secondary metabolism, and other metabolic pathways [25]. The CO2 assimilation capacity is mainly affected by the nitrogen content of the leaves and is generally positively correlated with nitrogen content [26]. This is because rapeseed leaves capture sunlight energy processes, electron transport processes, and enzymatic reactions in carbon metabolism require large amounts of nitrogen to synthesize ATP, NADPH, and various proteases [26].
Under nitrogen deficiency conditions, the chlorophyll content of leaves decreased, the original chlorophyll in the cytoplasm of mesophyll cells disappeared, fresh chlorophyll could not be formed, and the photosynthetic rate decreased [13,27]. Increasing nitrogen can promote chlorophyll synthesis and increase Rubisco content, while the high activity of Rubisco can promote photosynthetic rate [7,15,28,29,30]. In addition, nitrogen can also regulate CO2 concentration in chloroplasts by increasing the surface area of mesophyll cell walls and chloroplast to regulate photosynthetic rates [31].
3.2. Nitrogen Affects the Substance Distribution of Rapeseed
The amino acids produced in the process of nitrogen assimilation are transported between different organs through the phloem and xylem [33]. Phloem mainly transfers amino acids from photosynthetic tissue (source organs) to non-photosynthetic tissues (sink organs) [34]. The phloem sap of rapeseed contains up to 650 millimolar of free amino acids [35,36]. In the source leaves, sucrose and amino acids are excreted from mature and senescent leaves, loaded into the phloem through cytoplasmic pathways or extracellular phloem loading, and transported over long distances to developing sink organs [37]. The transport and distribution of carbon and nitrogen assimilation products in different parts and at different times are regulated by the physiological state, source strength, and expression of the transporters. In the vegetative growth phase, the assimilates in the rapeseed leaves are mainly transported to the immature leaves and developing roots [38,39]. During the reproductive growth phase, assimilates in the leaves are transported to the flowers, siliques, and grains via phloem loaders and transporters, including sugar and amino acid transporters [40].
Nitrogen deficiency will accelerate leaf senescence and result in a decrease in protein content in leaves. This may change the amino acid transport activity but increase nitrogen remobilization [36,41,42]. Leaf proteins are degraded when carbon and nitrogen are needed to support the growth of the carbon sink organ. Appropriate application of nitrogen can significantly promote the accumulation of carbon and nitrogen products in leaves, pods, and stems and promote their transfer from source to sink in the late growth stage [7,17,19,43]. The seed is the primary sink tissue for photosynthetic products. Carbohydrates and amino acids are metabolized and then stored in the seed as proteins and oils [44]. To enable the transport and accumulation of carbon and nitrogen products in the seed, the seed coat and seed sink tissue need to coordinate through a membrane that transports sucrose and amino acids [21]. This membrane plays a crucial role in the movement of nutrients between the two tissues, and its effectiveness is closely related to the quality and quantity of seed development. After the nutrients in the seed coat are transmitted through the cytoplasm of the seed coat parenchyma, the assimilates are exported to the extracellular cavity and subsequently absorbed by the developing seeds [21].
3.3. Molecular Mechanism of Nitrogen Affecting Rapeseed Yield
With the rapid development of molecular biology techniques and intensive research on crop nitrogen use efficiency, genes that regulate plant nitrogen uptake, transport and utilization efficiency have been discovered. External nitrate and ammonium are absorbed by roots and then are assimilated by the nitrate transporter family (NRTs) and the ammonium transporter family (AMTs) in the roots [45,46,47]. Typically, the uptake of NO3- by roots is limited. Most of NO3- is reduced to NH4+ by nitrate reductase (NR) and nitrite reductase (NIR) in shoots [48]. NH4+ is then further assimilated into organic forms through the glutamine synthetase/glutamate synthetase (GS/GOGAT) cycle and participates in carbon metabolism and biosynthesis of other compounds in plants [50]. Once NH4+ is accumulated, plants will produce toxic symptoms [44,46,50,51]. A high concentration of NH4+ as the only nitrogen source will cause stress, and the nutrition of NO3- or NO3- plus a slight amount of NH4+ is suitable for rapeseed growth [52]. In addition, nitrogen levels had a significant effect on nitrate reductase (NR) activity in rapeseed, maize, and wheat [7,8,53,54]. Sucrose and amino acids, the products of plant carbon and nitrogen assimilation, are released from mature and aged leaves, packed into the phloem, and transported to the sink tissues by the sucrose transporters family (SUTs/SUCs/SWEETs) and amino acid transporters [44].
Nitrogen levels affect the expression levels of ZmSWEETs and ZmSUTs in different parts of the maize during the vegetative stage [55]. The expression levels of ZmSWEETs and ZmSUTs in the leaves are higher in the low nitrogen condition than in the sufficient nitrogen condition, while the expression levels of ZmSWEETs and ZmSUTs in the internodes are higher in the sufficient nitrogen condition than in the low nitrogen condition [55]. These results show that sucrose transport from source to sink is more efficient at sufficient nitrogen levels than that at low nitrogen levels [55]. Sucrose synthase (SS) is a regulatory enzyme for sucrose degradation. It facilitates the decomposition of sucrose transported from the source organ and provides the basic substances for the synthesis and accumulation of fats and oils, and proteins in grains [56]. The activity of sucrose synthetase (SS) in rapeseed seeds is related to nitrogen level, and the SS activity is low under low nitrogen treatment [57]. Phytohormones are a class of naturally occurring small organic molecules that play an important role in regulating plant growth and development [58]. Nitrogen signaling and metabolic networks were regulated by hormone signals in plants [60,61,62]. The gene Dehydration-ResponsiveElement-Binding Protein 1C (OsDREB1C) belongs to the APETALA2/ethylene-responsive element binding factor (AP2/ERF) family. Its overexpression can significantly increase nitrogen uptake, transport, and utilization efficiency in rice, as well as the number of grains per panicle, grain weight, yield, and harvest index [62].
NO3- and NH4+ can act as signaling molecules that interact with various plant hormone signals to regulate plant nitrogen uptake, transport, and assimilation. The regulatory factor DNR1 modulates nitrogen metabolism in rice by affecting auxin biosynthesis and signal transduction pathways [63]. The transcription factor GROWTH-REGULATING FACTOR 4 (OsGRF4) and the growth inhibitor DELLA confer co-regulation of the absorption, assimilation, and transport of NH4+, regulate the photosynthetic carbon fixation capacity of crops, and enable high nitrogen utilization efficiency, increasing the yield of rice, maintaining homeostasis [64]. In general, there have been numerous studies of the molecular mechanisms of the nitrogen effect on production, but most of them have focused on Arabidopsis, maize, rice, and other plants, and extremely little has been done in the field of rapeseed.

Table 1.
Research progress of carbon and nitrogen metabolism genes related to yield.
| Genes | Plants | Functions |
|---|---|---|
|
LBD gene family (LBD37, LBD38, LBD39) |
Arabidopsis | LBD transcription factors inhibition of anthocyanin biosynthesis and nitrogen availability signal [65] |
|
NRT1.1; NRT2.1; NRT2 |
Arabidopsis | Nitrate transport [66,67,68,69] |
| MYB | Soybean; Sugarcane ; Foxtail millet |
Regulate nitrate transporters [70,71,72] |
| NLP7 | Arabidopsis | Regulation of nitrate assimilation [73] |
| Nitrate sensors and transcriptional activators initiate nitrate-mediated transcriptome signaling [74] | ||
| NLP5 | Maize | Regulation of nitrate assimilation [75] |
| CLC gene family | Arabidopsis | Regulation of nitrate transport [76,77] |
| CBL, CIPK8 | Arabidopsis | Regulation of nitrate transport and assimilation [78] |
|
AMT1.3; AMT2.1; AMT2.2; AMT2.3; AMT3.1; AMT3.2; |
Rice | Regulation of ammonium salt absorption and transport [79] |
| SWEETs | Maize; Arabidopsis |
Sucrose transport; the expression affected by nitrogen [55,80,81] |
| SUTs | Arabidopsis; Maize |
Sucrose transport; expression is affected by nitrogen [55,82] |
| NAS1 /NAP1 | Soybean | Regulating the flow distribution of PEP to maintain carbon and nitrogen balance [83] |
| PBF1 | Maize | Regulate carbon and nitrogen metabolism in a nitrogen-dependent manner [5] |
| ANR1 | Arabidopsis; Chrysanthemum |
ANR1-mediated auxin response [9,84] |
| DNR1 | Rice | DNR1-mediated auxin response [63] |
| GRF4 | Rice | Regulation of nitrogen absorption, assimilation, and transport [64] |
| DREB1C | Rice | Regulation of nitrogen utilization; Ethylene-mediated [62] |
4. Effects of Nitrogen on the Oil Content of Rapeseed
Oil is the ultimate target product of oil crops. Protein is the basic material of the protoplasm. Nitrogen is a major component of protein in plants, and many intermediates of sugar metabolism are major sources of protein synthesis. At different levels of nitrogen, the oil content of rapeseed without nitrogen application is the highest, and the oil content gradually decreases and the protein content gradually increases with the increase of nitrogen application [1,57,85]. In the case of low nitrogen is adverse to the biosynthesis of protein and glucosinolate but is beneficial to the formation and accumulation of lipid in seed [61].
4.1. Nitrogen Affects the Synthesis of Lipids and Proteins in Rapeseed
The reason for the negative correlation between oil content and protein content in plants is that the lipid and protein in plants come from the product of glycolysis-pyruvate (PRY), so there is substrate competition in the synthesis of the two products [86,87]. After entering reproductive growth, sucrose in the vegetative body is transported to the developing seed for protein and lipid synthesis [88]. Oil is a higher fatty acid glyceride synthesized by fatty acids and glycerol and is stored in seeds in the form of triacylglycerol [89]. The synthesis of lipids in plants is complex and is divided into three main stages: 1. Formation of fatty acids and fatty acid precursor acetyl-CoA: Sucrose is converted to hexose by glycolysis and hexose is oxidized to acetyl-CoA and one molecule of CO2 [11]. In this process, every two carbon atoms are converted to acetyl-CoA, and one carbon atom (one-third of carbon) is lost as CO2 [21]. Acetyl-CoA carboxylates to malonyl-CoA, which further synthesizes saturated fatty acids in the plastids, or desaturates to form unsaturated fatty acids [90]; 2. Production of 3-phosphoglycerol: In the cytoplasm, hydroxyacetone phosphate produced by the glycolytic pathway is reduced to 3-phosphoglycerol by the catalysis of 3-phosphoglycerol dehydrogenase [11]; 3. Assembly of fatty acyl-CoA and 3-phosphoglycerol: Fatty acyl-CoA and 3-phosphoglycerol were dehydrated and condensed in the endoplasmic reticulum to produce triacylglycerol [14,91]. In rapeseed, there are numerous enzymes and proteins involved in the synthesis of lipids and nitrogen is an important element in the formation of proteins. An adequate supply of nitrogen increases protein, and protein synthesis precedes fat synthesis, but in protein synthesis large amounts of photosynthetic products are consumed, resulting in a weakening of the library source allocated for lipid, and thus affecting oil content.
4.2. Molecular Mechanism of Nitrogen Level Affecting Lipid Synthesis in Rapeseed
The oil content is a complex quantitative trait influenced by multiple genes and subject to complex genetic interactions with the environment. The balance between lipid and protein synthesis in plants depends on the activity of key enzymes in their metabolism [88]. Pyruvate carboxylase (PEPC) is a key enzyme controlling the ratio of protein/oil content and is involved in the regulation of seed storage protein and fatty acid metabolism in plant seeds [56,86]. The activity of PEPC may be coordinated through post-translational regulation, with the overexpression of PEPC genes accelerated after nitrogen application [6,92]. Acetyl-CoA carboxylase (ACCase) is the rate-limiting enzyme in fatty acid biosynthesis [93]. The expression of the ACCase gene in rapeseed treated with different nitrogen changes significantly after 31 days of pollination [6]. After nitrogen application, the expression of the ACCase gene decreases in the high oil-content material, while it increases in the high protein-content material [6]. Phospholipid phosphatase (PPase) is the rate-limiting enzyme in the biosynthesis of triacylglycerol [56]. The oil content of seeds is related to the activity of the PPase gene. PPase activity in oilseeds is highest at 35 and 55 days after flowering under high nitrogen treatment, followed by medium and low nitrogen treatment [57]. Diacylglycerol acyltransferase (DGAT2) catalyzes the synthesis of triacylglycerol (TAG) from diacylglycerol (DAG) in plants and is the last step and the rate-limiting step of TAG synthesis [6]. Overexpression of the DGAT2 gene in tobacco can increase the oil content and the content of oleic acid and linoleic acid in leaves [6,94]. Similarly, the expression of the DGAT2 gene in the accession with high oil content was decreased by 12-fold after 31 days of pollination, compared to the case without nitrogen application [6]. Pyruvate kinase (PK) catalyzes phosphoenolpyruvate (PEP) to pyruvate (PYR) and is the last irreversible enzyme in the glycolysis pathway [95]. Glycerin-3-phosphate dehydrogenase (G3PDH) is involved in energy metabolic pathways such as glycolysis, gluconeogenesis, and the Calvin cycle, and plays an important role in promoting lipid synthesis [96]. The expression of the G3PDH and PK genes is found to be enhanced with increasing nitrogen application levels [97]. While many genes regulate lipid synthesis after nitrogen application, the regulatory network behind it is complex and huge, as oil content in rapeseed is a complex trait. Therefore, the molecular mechanism of nitrogen levels affecting the oil content of rapeseed still needs to be further investigated.
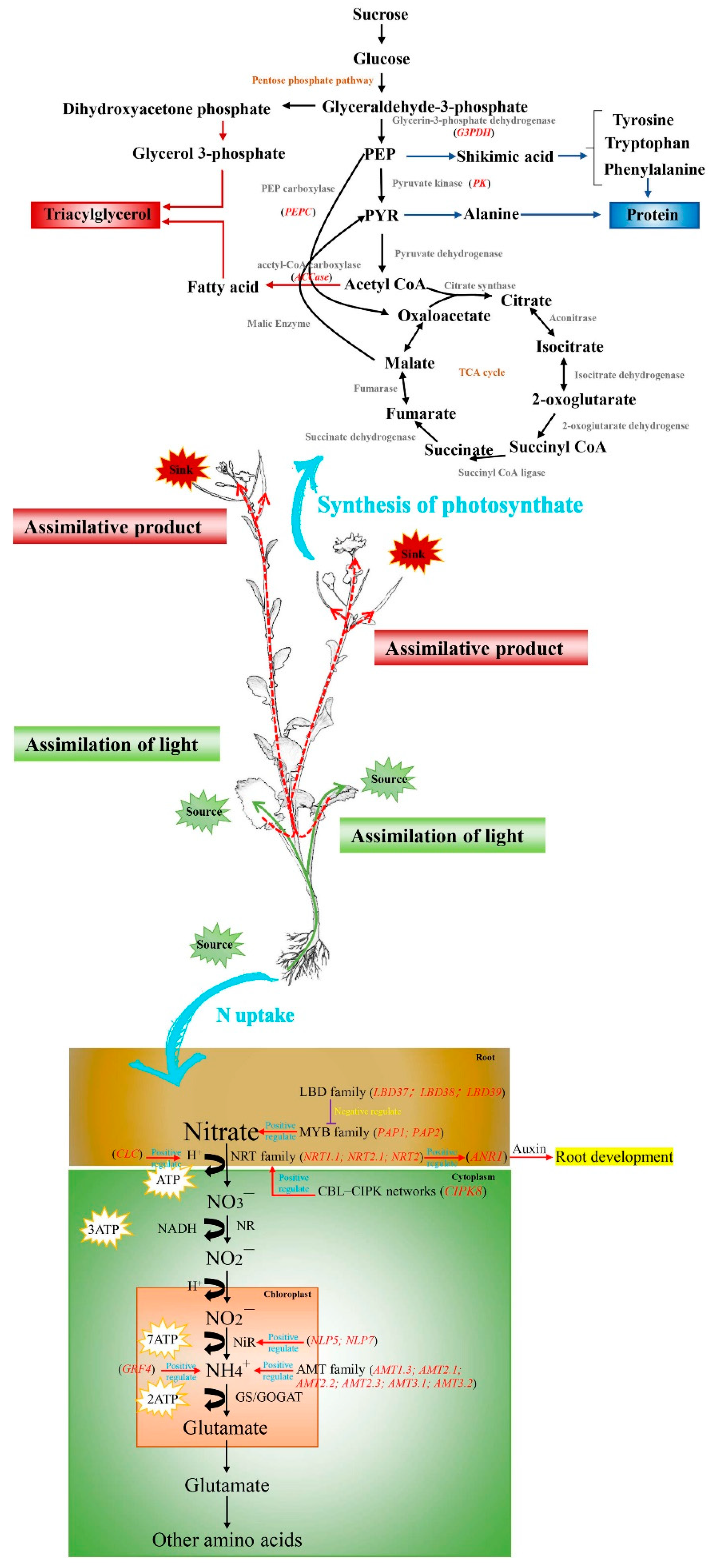
Figure 1.
Distribution of carbon and nitrogen source-sink nutrients in rapeseed. The sympathetic transporter (NRT) transports nitrate (NO3-) to mesophyll cells, and the nitrate reductase (NR) converts nitrate (NO3-) to nitrite (NO2-). Nitrite (NO2-) binding H+ is transferred to the stroma of chloroplasts and converted to ammonium (NH4+) by nitrite reductase (NiR). Ammonium (NH4+) is converted to glutamate by glutamine synthetase (GS) and glutamate synthetase (GOGAT). Glutamic acid is converted to other amino acids in the cytoplasm [29]. The root absorbs N and transports it to the leaves for photoassimilation, and the assimilated product is transported through the phloem to the developing leaves and growing roots. Nutrients from old leaves are remobilized into developing seeds for lipid and protein synthesis. The green line shows the path of nitrogen transfer and the red line shows the path of redistribution of photosynthetic products. PEP: phosphoenolpyruvate; PYR: pyruvate.
Figure 1.
Distribution of carbon and nitrogen source-sink nutrients in rapeseed. The sympathetic transporter (NRT) transports nitrate (NO3-) to mesophyll cells, and the nitrate reductase (NR) converts nitrate (NO3-) to nitrite (NO2-). Nitrite (NO2-) binding H+ is transferred to the stroma of chloroplasts and converted to ammonium (NH4+) by nitrite reductase (NiR). Ammonium (NH4+) is converted to glutamate by glutamine synthetase (GS) and glutamate synthetase (GOGAT). Glutamic acid is converted to other amino acids in the cytoplasm [29]. The root absorbs N and transports it to the leaves for photoassimilation, and the assimilated product is transported through the phloem to the developing leaves and growing roots. Nutrients from old leaves are remobilized into developing seeds for lipid and protein synthesis. The green line shows the path of nitrogen transfer and the red line shows the path of redistribution of photosynthetic products. PEP: phosphoenolpyruvate; PYR: pyruvate.
5. Discussion and Expectation
Rapeseed oil production is highest when nitrogen is applied at a rate of 180 kg·hm-2 [4,7,19]. Most of the current research has focused on the effect of nitrogen application on the yield of rapeseed or the effect of nitrogen application on the oil content, but little work has been done on the mechanisms by which nitrogen application affects the yield and oil content. It is necessary to investigate the mechanisms by which nitrogen affects yield and oil content, to balance the relationship between yield and oil content, and to achieve the goal of reducing nitrogen application without reducing oil production from rapeseed. In the future, to increase oil production by nitrogen application, the following suggestions have been made:
(1) Improving the cultivation level and the proper application of nitrogen. The relationship between protein and oil content should be balanced. Suitable varieties of rapeseed should be selected first, and then the amount of nitrogen should be applied to improve the yield and quality. The amount of nitrogen can be adjusted according to the needs in the range of 0 ~ 337.5 kg·hm-2 [7]. Studies have also shown that a nitrogen application rate maintained in the range of 90 ~ 180 kg·hm-2 can get higher oil content in the case of increased yield, but nitrogen agronomy efficiency, nitrogen partial productivity, nitrogen apparent efficiency, and nitrogen physiological efficiency of rapeseed are all at high values when the rate is 180 kg·hm-2 [19].
(2) Excavation of rapeseed for carbon and nitrogen-efficient germplasm resources. Most of the previous studies used a small number of accessions to analyze the relationship between nitrogen application rate and oil content but lacked representation. There are abundant germplasm resources of rapeseed with genetic diversity in China. It is necessary to ascertain whether any varieties produced an almost non-decreasing effect in the oil upon application of nitrogen. If such variety resources exist, high yield and oil content varieties with significantly increased oil yield after nitrogen application can be cultivated in the future.
(3) Studying the mechanism of carbon and nitrogen balance and identifying the important regulatory genes. Carbon and nitrogen partitioning in rapeseed growth and development is controlled by multiple genes and is involved in multiple metabolic pathways. A large number of previous studies have found that genes such as ACCase, PEPC, and DGAT2 control or affect carbon source partitioning [6,21,57]. 448 candidate genes related to acyl lipids and 11 genes related to storage proteins in rapeseed were identified [91]. However, the complex regulatory effects of nitrogen on oil and protein synthesis remain unclear. Functional genomics methods can be combined with transcriptome data to identify genes that control carbon and nitrogen partitioning, which is also an essential direction for future research.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (32072106), the Agricultural Science and Technology Innovation Project of the Chinese Academy of Agricultural Sciences (CAAS-ZDRW202105, CAAS-OCRI-ZDRW-202201, CAAS-ASTIP), National Crop Germplasm Resources Center (NCGRC2023-016) and the China Agriculture Research System (CARS-12).
References
- Yan, G.; Chen, B.; Xu, K.; Gao, G.; Lv, P.; Wu, X.; Li, F.; Li, J. , Differential gene expression profiles in developing seeds of Brassica napus L. under different nitrogen application levels. Acta Agron. Sin 2013, 38, 2052–2060. [Google Scholar] [CrossRef]
- Wang, H.; Yin, Y. , Analysis and strategy for oil crop industry in China. Chin. J. Oil Crop Sci 2014, 36, 414–421. [Google Scholar]
- Stahl, A.; Vollrath, P.; Samans, B.; Frisch, M.; Wittkop, B.; Snowdon, R. J. , Effect of breeding on nitrogen use efficiency-associated traits in oilseed rape. J Exp Bot 2019, 70, 1969–1986. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, J.; Gu, H.; Lu, Z.; Liao, S.; Li, X.; Cong, R.; Ren, T.; Lu, J. Effects of application of nitrogen on seed yield and quality of winter oilseed rape (Brassica napus L.). Acta Agron. Sin 2023, 2002-2011.
- Ning, L.; Wang, Y.; Shi, X.; Zhou, L.; Ge, M.; Liang, S.; Wu, Y.; Zhang, T.; Zhao, H. , Nitrogen-dependent binding of the transcription factor PBF1 contributes to the balance of protein and carbohydrate storage in maize endosperm. Plant Cell 2023, 35, 409–434. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Cheng, B.; Xu, K.; Gao, G.; Lv, P.; Wu, X.; Li, F.; Li, J. , Response of ACCase, DGAT2 and PEPC genes in developing seeds of Brassica napus to different nitrogen levels. Plant Nutr. Fert. Sci 2012, 18, 1370–1377. [Google Scholar]
- Zhou, N. The research on the nitrogen's effect on rapeseed yield and quality and relevant analysis. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2005. [Google Scholar]
- Huang, S.; Sun, G.; Jin, J.; He, P.; Wang, X.; Zhang, G.; Xie, J.; Zhang, K. , The effect of nitrogen levels on grain yield, protein, amino acid and fatty acid of high-oil maize. Chin. Agric. Sci 2004, 37, 250–255. [Google Scholar]
- Zhang, H.; Forde, B. G. , An Arabidopsis MADS box gene that controls nutrient-induced changes in root architecture. Science 1998, 279, 407–409. [Google Scholar] [CrossRef]
- Zhai, J.; Xue, J.; Zhang, Y.; Zhang, G.; Shen, D.; Wang, Q.; Liu, C.; Li, S. , Effect of nitrogen application rate on lodging resistance of spring maize stalks under integrated irrigation with water and fertilizer. J. Maize Sci 2021, 29, 137–144. [Google Scholar]
- Ni, F. Effect of different agronomic measures on carbon-nitrogen metabolism and oil accumulation in rapeseed pod. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2018. [Google Scholar]
- Remans, T.; Nacry, P.; Pervent, M.; Girin, T.; Tillard, P.; Lepetit, M.; Gojon, A. , A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation in Arabidopsis. Plant Physiol 2006, 140, 909–921. [Google Scholar] [CrossRef]
- Guan, C. , High-quality rapeseed physiological ecology and modern cultivation technology. first ed.; China Threegorges Press: Beijing, 2006. [Google Scholar]
- Pan, R. , Plant physiology. seventh ed.; Higher Education Press: Beijing, 2012. [Google Scholar]
- Zhang, X.; He, S.; Wang, W.; Guo, W.; Yu, L. , Effect of different nitrogen level on nitrogen metaboli characteristic, grain yield, and quality of strong gluten spring wheat. J. Triticeae Crops 2006, 130–133. [Google Scholar]
- Khan, S.; Anwar, S.; Kuai, J.; Noman, A.; Shahid, M.; Din, M.; Ali, A.; Zhou, G. , Alteration in yield and oil quality traits of winter rapeseed by lodging at different planting density and nitrogen rates. Sci Rep 2018, 8, 634. [Google Scholar] [CrossRef] [PubMed]
- Akhatar, J.; Singh, M. P.; Sharma, A.; Kaur, H.; Kaur, N.; Sharma, S.; Bharti, B.; Sardana, V. K.; Banga, S. S. , Association mapping of seed quality traits under varying conditions of nitrogen application in Brassica juncea L. czern & coss. Front Plant Sci 2020, 11, 744. [Google Scholar]
- Tang, W. Effects of nitrate and ammonium supply ratios on growth, physiology and gene expression of oilseed rape (Brassica napus). Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2019. [Google Scholar]
- Wu, Y.; Ma, N.; Huang, X.; Peng, H.; Li, Z.; Niu, Y.; Zhang, C. , Effect of nitrogen fertilizer on agronomic traits, yield, quality and nitrogen use efficiency in Brassica napus of "ZhongShuang 11" under Different Densities. J. Sichuan Agric. Univ 2014, 32, 260–264+282. [Google Scholar]
- Zhu, S.; Li, Y.; Yu, C.; Xie, L.; Hu, X.; Zhang, S.; Liao, X.; Liao, X.; Che, Z. , Effects of planting density and nitrogen application rate on rapeseed yield and nitrogen use efficiency. Chin. J. Oil Crop Sci. 2013, 35, 179–184. [Google Scholar]
- Plaxton, W. C. In Annual plant reviews volume 22 control of primary metabolism in plants, 2006.
- Zhou, X.; Fei, Z.; Thannhauser, T. W.; Li, L. , Transcriptome analysis of ectopic chloroplast development in green curd cauliflower (Brassica oleracea L. var. botrytis). BMC Plant Biol 2011, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Urban, L.; Aarrouf, J.; Bidel, L. P. R. , Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Front Plant Sci 2017, 8, 2068. [Google Scholar] [CrossRef] [PubMed]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M. E.; Francia, F.; Danon, A.; Marchand, C. H.; Fermani, S.; Trost, P.; Lemaire, S. D. , Redox regulation of the Calvin-Benson cycle: something old, something new. Front Plant Sci 2013, 4, 470. [Google Scholar] [CrossRef]
- Chapman, S. P.; Trindade dos Santos, M.; Johnson, G. N.; Kritz, M. V.; Schwartz, J.-M. , Cyclic decomposition explains a photosynthetic down regulation for Chlamydomonas reinhardtii. Biosystems 2017, 162, 119–127. [Google Scholar] [CrossRef]
- Liu, T. Mechanisms underlying the effects of nitrogen supply on leaf nitrogen-forms partitioning and photosynthetic nitrogen use efficiency of Brassica napus. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2018. [Google Scholar]
- Xu, H. Nitrogen diagnosis of oilseed rape at different growth stages and leaf positions using chlorophyll fluorescence techniques. Ph.D. Thesis, Zhejiang university, Hangzhou, China, 2020. [Google Scholar]
- Bi, H.; Liu, P.; Jiang, Z.; Ai, X. , Overexpression of the rubisco activase gene improves growth and low temperature and weak light tolerance in Cucumis sativus. Physiol Plant 2017, 161, 224–234. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, H.; Song, J.; Wei, S.; Zhang, Y.; Wang, Q. , Effects of nitrogen fertilizer rate on the chlorophyll content and quality of kidney bean. Crops 2013, 81–87. [Google Scholar]
- Zhang, R. Effect of urea application rate on yield and quality of different soybeans. Ph.D. Thesis, Shenyang Agricultural University, Shenyang, China, 2008. [Google Scholar]
- Xiong, D.; Liu, X.; Liu, L.; Douthe, C.; Li, Y.; Peng, S.; Huang, J. , Rapid responses of mesophyll conductance to changes of CO2 concentration, temperature and irradiance are affected by N supplements in rice. Plant Cell Environ 2015, 38, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A. J. , Energetics of nitrogen acquisition. In Annual Plant Reviews Volume 42, 2010; pp 63-81.
- Yu, S.; Lo, S.; Ho, T. D. , Source-sink communication: regulated by hormone, nutrient, and stress cross-signaling. Trends Plant Sci 2015, 20, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Baxter, I. R.; Lahner, B.; Reinders, A.; Salt, D. E.; Ward, J. M. , Arabidopsis NPCC6/NaKR1 is a phloem mobile metal binding protein necessary for phloem function and root meristem maintenance. Plant Cell 2010, 22, 3963–3979. [Google Scholar] [CrossRef]
- Tilsner, J.; Kassner, N.; Struck, C.; Lohaus, G. , Amino acid contents and transport in oilseed rape (Brassica napus L.) under different nitrogen conditions. Planta 2005, 221, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Orsel, M. , Leaf nitrogen remobilisation for plant development and grain filling. Plant Biol 2008, 10 (Suppl. 1), 23–36. [Google Scholar] [CrossRef]
- Osorio, S.; Ruan, Y.; Fernie, A. R. , An update on source-to-sink carbon partitioning in tomato. Front Plant Sci 2014, 5, 516. [Google Scholar] [CrossRef]
- Gomes, E. N.; Moterle, D.; Biasi, L. A.; Koehler, H. S.; Kanis, L. A.; Deschamps, C. , Plant densities and harvesting times on productive and physiological aspects of Stevia rebaudiana Bertoni grown in southern Brazil. An. Acad. Bras. Cienc 2018, 90, 3249–3264. [Google Scholar] [CrossRef]
- Cholewa, E.; Griffith, M. , The unusual vascular structure of the corm of eriophorum vaginatum: implications for efficient retranslocation of nutrients. J Exp Bot 2004, 55, 731–741. [Google Scholar] [CrossRef]
- Yu, S. M.; Lo, S. F.; Ho, T. D. , Source-sink communication: Regulated by hormone, nutrient, and stress cross-signaling. Trends Plant Sci 2015, 20, 844–857. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.; Hölzer, R.; Feller, U. , Influence of nitrogen deficiency on senescence and the amounts of RNA and proteins in wheat leaves. Physiol. Plant 1998, 102, 192–200. [Google Scholar] [CrossRef]
- Uhart, S. A.; Andrade, F. H. , Nitrogen and carbon accumulation and remobilization during grain filling in maize under different source/sink ratios. Crop Sci 1995, 35. [Google Scholar] [CrossRef]
- Cao, L.; Wu, X.; Yang, R.; Tian, Y.; Chen, X.; Chen, B.; Li, Y.; Gao, Y. , Differences of nitrogen status between different N-uptake-efficiency rapeseed (Brassica napus L.) cultivars. Acta Agron 2013, 38, 887–895. [Google Scholar] [CrossRef]
- Lam, H.; Coschigano, K. T.; Oliveira, I. C.; Melo-Oliveira, R.; Coruzzi, G. , The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu Rev Plant Physiol Plant Mol Biol 1996, 47, 569–593. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, Y.; Chen, K.; Tsay, Y. , Nitrate transport, signaling, and use efficiency. Annu Rev Plant Biol 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Xing, J.; Cao, X.; Zhang, M.; Wei, X.; Zhang, J.; Wan, X. , Plant nitrogen availability and crosstalk with phytohormones signalings and their biotechnology breeding application in crops. Plant Biotechnol J 2022, 1320–1342. [Google Scholar]
- Khademi, S. O. C., Joseph 3rd; Remis, J.; Robles-Colmenares, Y.; Miercke, L. J. W.; Stroud, R. M. , Mechanism of ammonia transport by Amt/MEP/Rh: structure of AmtB at 1.3.5 angstrom. Science 2004, 305, 1587–1594. [Google Scholar] [CrossRef]
- Garai, S.; Tripathy, B. C. , Alleviation of nitrogen and sulfur deficiency and enhancement of photosynthesis in Arabidopsis thaliana by overexpression of uroporphyrinogen III methyltransferase (UPM1). Front Plant Sci 2018, 8. [Google Scholar]
- Meng, S.; Wang, S.; Quan, J.; Su, W.; Lian, C.; Wang, D.; Xia, X.; Yin, W. , Distinct carbon and nitrogen metabolism of two contrasting poplar species in response to different N supply levels. Int J Mol Sci 2018, 19, 2302. [Google Scholar] [CrossRef]
- Lea, P. J.; Azevedo, R. A. , Nitrogen use efficiency. 1. Uptake of nitrogen from the soil. Ann App Biol 2006, 149, 243–247. [Google Scholar] [CrossRef]
- Navarro-León, E.; Barrameda-Medina, Y.; Lentini, M.; Esposito, S.; Ruiz, J. M.; Blasco, B. , Comparative study of Zn deficiency in L. sativa and B. oleracea plants: NH4+assimilation and nitrogen derived protective compounds. Plant Sci 2016, 248, 8–16. [Google Scholar] [CrossRef]
- Tang, W.; He, X.; Qian, L.; Wang, F.; Zhang, Z.; Sun, C.; Lin, L.; Guan, C. , Comparative transcriptome analysis in oilseed rape (Brassica napus) reveals distinct gene expression details between nitrate and ammonium nutrition. Genes (Basel) 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tang, L.; Zhang, W.; Cao, W.; Zhu, Y. , Dynamic analysis on response of dry matter accumulation and partitioning to nitrogen fertilizer in wheat cultivars with different plant types. Acta Agron. Sin 2009, 35, 2258–2265. [Google Scholar]
- Song, X.; Liu, Q.; Song, H.; Guan, C.; Rong, x.; Wang, J.; Wang, S. , Changes of soluble sugar and free amino acids in stem and leaf and their effects on yield of rapeseed. Acta Agric. Boreali- Occident. Sin 2010, 19, 187–191. [Google Scholar]
- Zhao, Y.; Ning, P.; Feng, X.; Ren, H.; Cui, M.; Yang, L. , Characterization of stem nodes associated with carbon partitioning in maize in response to nitrogen availability. Int J Mol Sci 2022, 23, 4389. [Google Scholar] [CrossRef]
- Tang, X.; Guan, C. , Advances in the regulation of carbon, nitrogen and fat metabolism in crop yield and quality. J. Hunan Agric. Univ 1997, 96–106. [Google Scholar]
- Tang, X.; Guan, C. , Effects of nitrogen application on enzymatic activities of rape and its relationship with yield and quality. Chin. J. Oil Crop Sci 2001, 33–38. [Google Scholar]
- Mao, C.; He, J.; Liu, L.; Deng, Q.; Yao, X.; Liu, C.; Qiao, Y.; Li, P.; Ming, F. , OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol J 2020, 18, 429–442. [Google Scholar] [CrossRef]
- Wu, K.; Wang, S.; Song, W.; Zhang, J.; Wang, Y.; Liu, Q.; Yu, J.; Ye, Y.; Li, S.; Chen, J.; Zhao, Y.; Wang, J.; Wu, X.; Wang, M.; Zhang, Y.; Liu, B.; Wu, Y.; Harberd, N. P.; Fu, X. , Enhanced sustainable green revolution yield via nitrogen-responsive chromatin modulation in rice. Science 2020, 367, eaaz2046. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, K.; Harberd, N. P.; Fu, X. , Green Revolution DELLAs: From translational reinitiation to future sustainable agriculture. Mol. Plant 2021, 14, 547–549. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Fernie, A. R.; Stitt, M. , Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Wei, S.; Li, X.; Lu, Z.; Zhang, H.; Ye, X.; Zhou, Y.; Li, J.; Yan, Y.; Pei, H.; Duan, F.; Wang, D.; Chen, S.; Wang, P.; Zhang, C.; Shang, L.; Zhou, Y.; Yan, P.; Zhao, M.; Huang, J.; Bock, R.; Qian, Q.; Zhou, W. , A transcriptional regulator that boosts grain yields and shortens the growth duration of rice. Science 2022, 377, eabi8455. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, L.; Shen, C.; Ji, Z.; Zhang, H.; Zhang, T.; Li, Y.; Yu, J.; Yang, N.; He, Y.; Tian, Y.; Wu, K.; Wu, J.; Harberd, N. P.; Zhao, Y.; Fu, X.; Wang, S.; Li, S. , Natural allelic variation in a modulator of auxin homeostasis improves grain yield and nitrogen use efficiency in rice. The Plant Cell 2021, 33, 566–580. [Google Scholar]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; Harberd, N. P.; Fu, X. , Modulating plant growth-metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [PubMed]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.-R. , Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. The Plant cell 2009, 21, 3567–3584. [Google Scholar] [PubMed]
- Ho, C.; Lin, S.; Hu, H.; Tsay, Y. , CHL1 functions as a nitrate sensor in plants. Cell 2009, 138, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Tsay, Y.; Schroeder, J. I.; Feldmann, K. A.; Crawford, N. M. , The herbicide sensitivity gene CHL1 of Arabidopsis encodes a nitrate-inducible nitrate transporter. Cell 1993, 72, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Little, D. Y.; Rao, H.; Oliva, S.; Daniel-Vedele, F.; Krapp, A.; Malamy, J. E. , The putative high-affinity nitrate transporter NRT2.1 represses lateral root initiation in response to nutritional cues. PNAS 2005, 102, 13693–13698. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Okamoto, M.; Crawford, N. M.; Siddiqi, M. Y.; Glass, A. D. M. , Dissection of the AtNRT2.1: AtNRT2.2 inducible high-affinity nitrate transporter gene cluster. Plant Physiol 2007, 143, 425–433. [Google Scholar] [CrossRef]
- Miyake, K.; Ito, T.; Senda, M.; Ishikawa, R.; Harada, T.; Niizeki, M.; Akada, S. , Isolation of a subfamily of genes for R2R3-MYB transcription factors showing up-regulated expression under nitrogen nutrient-limited conditions. Plant Mol Biol 2003, 53, 237–245. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, S.; Su, Y.; Lin, Z.; Guo, J.; Li, M.; Wang, Z.; Que, Y.; Xu, L. , Transcripts and low nitrogen tolerance: regulatory and metabolic pathways in sugarcane under low nitrogen stress. Environ. Exp. Bot 2019, 163, 97–111. [Google Scholar]
- Qian, D. Q.; Ting, W. X.; Qin, H. L.; Xin, Q.; Hao, G. L.; Ya, X. W.; Shi, X. Z.; Bin, Z. Y.; Qing, J. G.; Min, D. X.; Hong, M. D.; Zhi, M. Y.; Ming, C. , MYB-like transcription factor SiMYB42 from foxtail millet (Setaria italica L.) enhances Arabidopsis tolerance to low-nitrogen stress. Hereditas 2018, 40. [Google Scholar]
- Castaings, L.; Camargo, A.; Pocholle, D.; Gaudon, V.; Texier, Y.; Boutet-Mercey, S.; Taconnat, L.; Renou, J.-P.; Daniel-Vedele, F.; Fernandez, E.; Meyer, C.; Krapp, A. , The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J. Cell Mol. Biol 2009, 57, 426–435. [Google Scholar]
- Liu, K.; Liu, M.; Lin, Z.; Wang, Z.; Chen, B.; Liu, C.; Guo, A.; Konishi, M.; Yanagisawa, S.; Wagner, G.; Sheen, J. , NIN-like protein 7 transcription factor is a plant nitrate sensor. Science 2022, 377, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Wang, Y.; Liu, Y.; Jiang, L.; He, B.; Ning, L.; Du, H.; Lv, Y.; Zhou, L.; Lin, F.; Zhang, T.; Liang, S.; Lu, H.; Zhao, H. , The NIN-like protein 5 (ZmNLP5) transcription factor is involved in modulating the nitrogen response in maize. Plant J 2020, 102, 353–368. [Google Scholar] [CrossRef]
- Harada, H.; Kuromori, T.; Hirayama, T.; Shinozaki, K.; Leigh, R. A. , Quantitative trait loci analysis of nitrate storage in Arabidopsis leading to an investigation of the contribution of the anion channel gene, AtCLC-c, to variation in nitrate levels. J Exp Bot 2004, 55, 2005–2014. [Google Scholar] [CrossRef]
- De Angeli, A.; Monachello, D.; Ephritikhine, G.; Frachisse, J.-M.; Thomine, S.; Gambale, F.; Barbier-Brygoo, H. , CLC-mediated anion transport in plant cells. P Philos. Trans.: Biol. Sci 2009, 364, 195–201. [Google Scholar]
- Hu, H.; Wang, Y.; Tsay, Y. , AtCIPK8, a CBL-interacting protein kinase, regulates the low-affinity phase of the primary nitrate response. Plant J. Cell Mol. Biol 2009, 57, 264–278. [Google Scholar] [CrossRef]
- Wu, X.; Ding, C.; Baerson, S. R.; Lian, F.; Lin, X.; Zhang, L.; Wu, C.; Hwang, S.; Zeng, R.; Song, Y. , The roles of jasmonate signalling in nitrogen uptake and allocation in rice (Oryza sativa L. ). Plant Cell Environ 2019, 42, 659–672. [Google Scholar]
- Ning, P.; Yang, L.; Li, C.; Fritschi, F. B. , Post-silking carbon partitioning under nitrogen deficiency revealed sink limitation of grain yield in maize. J Exp Bot 2018, 69, 1707–1719. [Google Scholar]
- Chen, H.; Huh, J.; Yu, Y.; Ho, L.; Chen, L.; Tholl, D.; Frommer, W. B.; Guo, W. , The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection. Plant J. Cell Mol. Biol 2015, 83, 1046–1058. [Google Scholar]
- Slewinski, T. L.; Meeley, R.; Braun, D. M. , Sucrose transporter1 functions in phloem loading in maize leaves. J Exp Bot 2009, 60, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Xiao, H.; Peng, Y.; Wang, J.; Lv, Q.; Wang, X. , Phosphoenolpyruvate reallocation links nitrogen fixation rates to root nodule energy state. Science 2022, 378, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yu, J.; Duan, X.; Wang, J.; Zhang, Q.; Gu, K.; Hu, D.; Zheng, C. , The MADS transcription factor CmANR1 positively modulates root system development by directly regulating CmPIN2 in chrysanthemum. Hortic Res 2018, 5, 52. [Google Scholar] [CrossRef] [PubMed]
- Rathke, G. W.; Christen, O.; Diepenbrock, W. , Effects of nitrogen source and rate on productivity and quality of winter oilseed rape (Brassica napus L.) grown in different crop rotations. Field Crops Res. 2005, 94, 103–113. [Google Scholar] [CrossRef]
- Chen, J.; Lang, C.; Hu, Z.; Liu, Z.; Huang, R. , Antisense PEP gene regulates to ratio of protein and lipid content in Brassica napus seeds. J. Agric. Biotechnol 1999, 7, 316–320. [Google Scholar]
- Sangwan R S, Singh N, Plaxton W C. Phosphoenolpyruvate carboxylase activity and concentration in the endosperm of developing and germinating castor oil seeds. Plant Physiol 1992, 99(2): 445-9.
- Yu, S.; Du, S.; Yuan, J.; Hu, Y. , Fatty acid profile in the seeds and seed tissues of Paeonia L. species as new oil plant resources. Sci Rep 2016, 6, 26944. [Google Scholar] [CrossRef]
- Tang, M.; Guschina, I. A.; O'Hara, P.; Slabas, A. R.; Quant, P. A.; Fawcett, T.; Harwood, J. L. , Metabolic control analysis of developing oilseed rape (Brassica napus cv Westar) embryos shows that lipid assembly exerts significant control over oil accumulation. New Phytol 2012, 196, 414–426. [Google Scholar] [CrossRef]
- Nikolau, B. J.; Ohlrogge, J. B.; Wurtele, E. S. , Plant biotin-containing carboxylases. Arch. Biochem. Biophys 2003, 414, 211–222. [Google Scholar] [CrossRef]
- Chao, H.; Wang, H.; Wang, X.; Guo, L.; Gu, J.; Zhao, W.; Li, B.; Chen, D.; Raboanatahiry, N.; Li, M. , Genetic dissection of seed oil and protein content and identification of networks associated with oil content in Brassica napus. Sci Rep 2017, 7, 46295. [Google Scholar] [CrossRef]
- Champigny, M.-L.; Foyer, C. , Nitrate activation of cytosolic protein kinases diverts photosynthetic carbon from sucrose to amino acid biosynthesis: basis for a new concept. Plant Physiol 1992, 100, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, F. Cloning and functional analysis of key ACCase and PEPCase genes in lipid biosynthesis of rape seeds (Brasscia napus L.). Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2012. [Google Scholar]
- Bian, Z.; Chen, G.; Zhou, Q.; Lv, X.; Zhu, J.; Sun, L. , Cloning of a lipid synthesis-related gene DGAT2 from sunflower and transformation into tobacco. J. Shihezi Univ 2016, 34, 624–631. [Google Scholar]
- Mazzarino, R. C.; Baresova, V.; Zikánová, M.; Duval, N.; Wilkinson, T. G., 2nd; Patterson, D.; Vacano, G. N. , The CRISPR-Cas9 crADSL HeLa transcriptome: A first step in establishing a model for ADSL deficiency and SAICAR accumulation. MGM Rep 2019, 21, 100512. [Google Scholar] [CrossRef]
- Koga, Y.; Konishi, K.; Kobayashi, A.; Kanaya, S.; Takano, K. , Anaerobic glycerol-3-phosphate dehydrogenase complex from hyperthermophilic archaeon Thermococcus kodakarensis KOD1. J Biosci Bioeng 2019, 127, 679–685. [Google Scholar] [CrossRef]
- Zhou, X.; Fu, S. , Response of G3PDH and PK genes in Brassica napus L. to nitrogen fertilizer. J. Integr Agric 2019, 35, 110–114. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



