
Submitted:
23 August 2023
Posted:
24 August 2023
You are already at the latest version
Abstract
Neuronal PAS domain protein 2 (NPAS2) is a hemeprotein comprising a basic helix-loop-helix domain (bHLH) and two heme-binding sites, the PAS-A and PAS-B domains. This protein acts as a pyridine nucleotide-dependent and gas-responsive CO-dependent transcription factor and is encoded by a gene whose expression fluctuates with circadian rhythmicity. NPAS2 can heterodimerize with ARNTL in the activating limb of the transcriptional/translational feedback loop operating the circadian clock circuitry. NPAS2 can functionally substitute for the core transcription factor and histone/protein acetylase CLOCK in the molecular clockwork endowing central oscillators in the suprachiasmatic nuclei of hypothalamus as well as in peripheral oscillators in the cell of other body tissues, such as the liver and kidney.
Keywords:
NPAS2
; circadian
; rhythmicity
; biological clock
; PAS-domain
1. Introduction
Fundamental processes bringing on cell, tissue and organ physiology are featured by rhythmic fluctuations with prevalent 24-hour periodicity, defined circadian and driven by molecular clockworks endowing every single cell in the body [1]. These autonomous peripheral oscillators are organized at tissue, organ and organ system level to arrange a hierarchical network structure defined circadian timing system and organized by dominant oscillators in the central nervous system [2,3,4,5,6]. In particular, a master pacemaker is located in the suprachiasmatic nuclei (SCN) of hypothalamus, which synchronize rhythmic activity in other brain regions and centers through direct monosynaptic projections connections and in peripheral tissue through autonomic nervous system fibers and neuroendocrine outputs, primarily cortisol and melatonin [2,3,4,5,6]. SCNs are located in the anterior part of the hypothalamus on each side of the third ventricle, in the area situated directly above the optic chiasm and consist of roughly 8000-10000 neurons in mus musculus and 80.000-100.000 neurons in Homo sapiens. SCN receive photic inputs through the retino-hypothalamic tract (RHT), which conveys signals from intrinsically photosensitive retinal ganglion cells (ipRGCs), a subpopulation representing less than 5% of ganglion cells. ipRGCs signal light stimuli synchronizing and entraining physiological circadian rhythmicity to the foremost environmental cue represented by light/darkness alternation (photoentrainment) and start off mainly involuntary non-image-forming visual reflexes, for example pupillary constriction and neuroendocrine regulation [2,3,4,5,6]. In the SCN were identified "core" and "shell" sub-regions expressing different neuropeptides crucial for circadian rhythmicity preservation: vasoactive intestinal peptide (VIP) and gastrin-releasing peptide (GRP) are mainly expressed in the retino-recipient core, whereas arginine vasopressin (AVP) is expressed in the shell. The activity of the core and shell VIP, GRP, and AVP-expressing neurons is stringently ordered in a time-qualified manner: VIP with its VPAC2receptor upholds SCN internal synchronization and increases in the core sub-region during darkness, while GRP is definitely stimulated by direct inputs from the photosensitive ganglion cells in the retina via the RHT and rises during the morning with peaks around midday. AVP-expressing neurons in the "shell" sub-region project to the paraventricular nucleus (PVN), coordinating feeding-fasting with sleep-wake and rest-activity cycles, and to thirst-controlling neurons in the organum vasculosum lamina terminalis (OVLT) [7,8]. In the hypothalamic SCN circadian fluctuations of gene expression and electrical activity can continue indeterminately in temporal isolation and this robust and resilient timekeeping is produced inherently and is largely considered related to the intrinsic connectivity of its neurons. Indeed, SCN circuit-level timekeeping is operated through mutually dependent astrocytic-neuronal signaling. SCN neurons are metabolically active during daytime, whereas SCN astrocytes are active during nighttime and drive extracellular glutamate levels, thus hindering the activity of SCN neurons. Neurons of the dorsal SCN gauge this glutamatergic gliotransmission by via specific pre-synaptic NMDA receptor assemblies containing NR2C subunits [9].
2. NPAS and the Molecular Clockwork
At the cellular level the molecular cogs that run the biological clock driving rhythmicity of cell functions and processes are embodied by a group of circadian proteins encoded by genes, so-called core clock genes, operating interlocking transcription-translation feedback loops (TTFL), completing one cycle in approximately 24 hours along with non-transcriptional loops [10,11,12,13,14]. The bHLH-PAS (basic helix-loop-helix–Period-Arnt-Single-minded) transcriptional activators CLOCK (circadian locomotor output cycles kaput), and its paralog NPAS2 (neuronal PAS domain protein 2), and ARNTL-2/BMAL1-2 (aryl-hydrocarbon receptor nuclear translocator-like/brain and muscle aryl-hydrocarbon receptor nuclear translocator-like) operate the positive limb of tbe TTFL, heterodimerizing and binding to enhancer (E)-box DNA consensus sequences of the target genes Cryptochrome (CRY 1-2) and Period (PER1-3). The TTFL negative limb is operated by the circadian proteins CRY1-2 and PER1-3, which in the cytoplasm accumulate and dimerize establishing repressor complexes that pass back to the nucleus and hinder the transcriptional activity of ARNTL:NPAS2 and ARNTL:CLOCK heterodimers [10,11,12,13,14]. The circadian proteins go through post-translational modifications (PTMs), such as phosphorylation, acetylation, sumoylation, O-GlcNAcylation, which allow activity modulation and sequential ubiquitination/proteasomal degradation: this is crucial for appropriate functioning of the biological clock [15,16,17]. The nuclear receptors (NRs) REV-ERBs and retinoic acid-related (RAR) orphan receptor (RORs) manage an auxiliary interconnected loop driving Arntl rhythmic transcription competing for a ROR specific response elements (RORE) in its promoter [18]. ROR-α physically interacts with peroxisome proliferator-activated receptor (PPAR)-γ coactivator-1α (PGC-1α) and acts as transcription activator recruiting chromatin-remodelling complexes to proximal ARNTL promoters and prompting Arntl transcription. Contrariwise, REV-ERB-α inhibits Atranscription interacting with the nuclear corepressor/histone deacetylase3 (NCoR-HDAC3) corepressor complex [18]. In addition to the NR operated feedback loop, binding to cognate D-box elements CLOCK: ARNTL heterodimers drive the expression of first order clock controlled gens, the PAR domain basic leucine zipper (bZIP) transcription factors DBP (albumin D-site binding protein), TEF (thyrotroph embryonic factor), HLF (hepatic leukaemia factor), which sequentially drive the rhythmic expression of thousands tissue specific (output) genes [19,20]. Moreover, REV-ERBs and DBP compete for response elements in the promoter and drive the rhythmic expression of the Nuclear factor, interleukin 3 regulated protein (NFIL3, also known as E4BP4), with opposed phase respect to DBP [19,20]. Other molecular cogs of the biological clock are the basic helix-loop-helix transcription factors Differentially expressed in chondrocytes protein 1 (DEC1) and 2 (DEC2), which interplay with the core clock proteins. In particular, DEC1 and DEC2 transcription is activated by ARNTL:NPAS2 and ARNTL:CLOCK heterodimer, but in sequence DEC1 and DEC2 proteins block their transcriptional activity, managing a feed-back steering circuit [19,20,21]. The functioning of the molecular clockwork is finely tuned through rhythmic chromatin-histone remodeling and epigenetic modifications principally geared up by acetylation/deacetylation and methylation/demethylation processes. Concerning the cogs of the biological clock, CLOCK, which has intrinsic protein and histone acetyltransferase capability, acetylates ARNTL [22]. Conversely, the type III histone/protein deacetylase SIRT1, opposes this process and its deacetylating activity relies on the intracellular levels of nicotinamide adenine dinucleotide (NAD+), a nutrient sensor produced from tryptophan through the enzymatic activity of nicotinamide phosphoribosyl-transferase (NAMPT/visfatin), which is expressed rhythmically driven by the biological clock [23,24,25,26].
3. NPAS2, hemeprotein and gas-responsive transcription factor
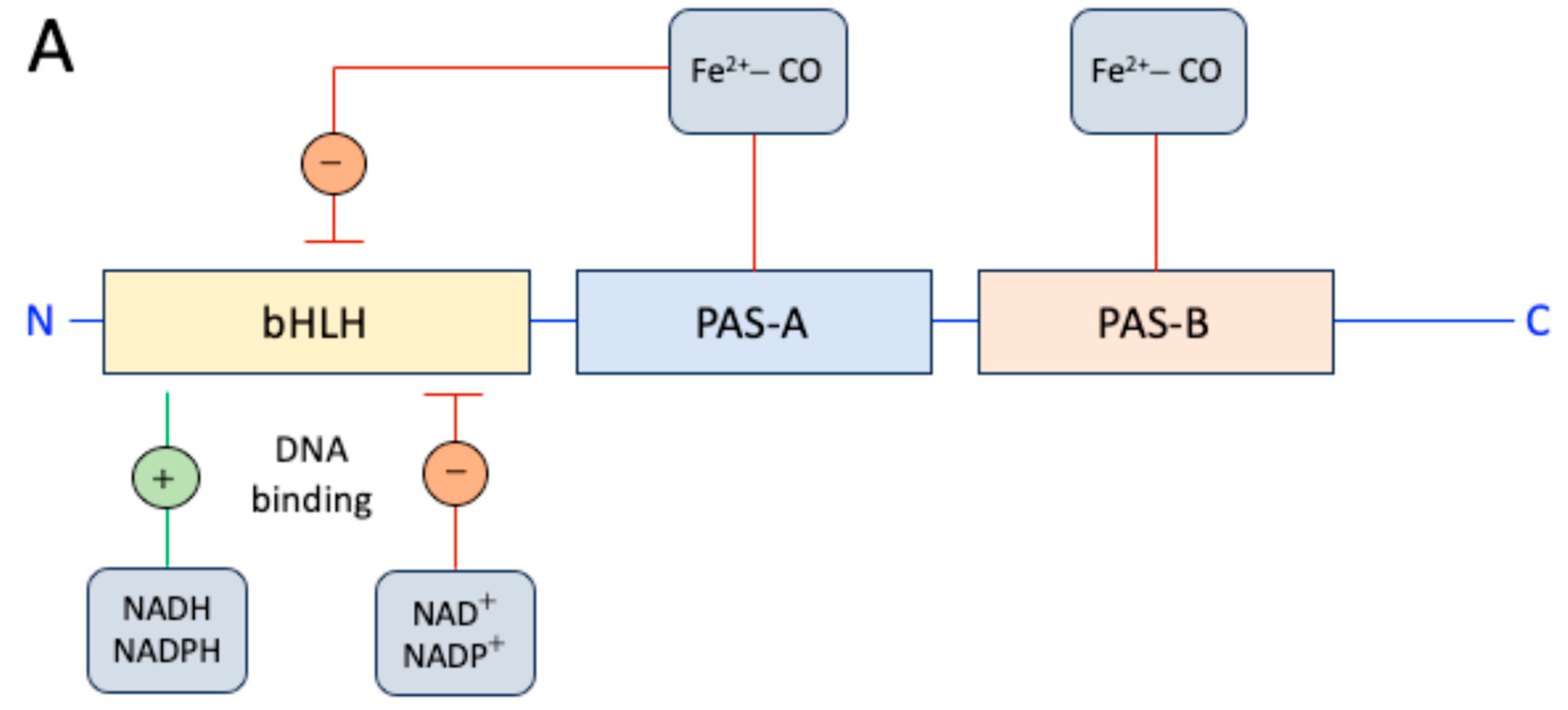
A protein in which a heme-binding domain controls the activity of another domain is defined as heme-based sensor. NPAS2 (in extensum Neuronal-PAS type signal-sensor protein-domain protein 2) is a pyridine nucleotide-dependent and carbon monoxide (CO)-dependent transcription factor consisting of a stabilizing and DNA binding basic helix-loop-helix domain (bHLH) and two heme-binding PAS (Period–Arnt–Single-minded) domains in the N-terminal region (Figure 1A). ARNTL:NPAS2 heterodimer establishment and DNA binding can be impeded by a heme-mediated mechanism or self-sufficiently repressed by an excessively low NADPH/NADP ratio (reduced and oxidized forms of NADP, respectively) and the unproductive ARNTL-ARNTL homodimer is formed if either or both signals that inhibit formation of the ARNTL:NPAS2 heterodimer are present [27,28]. PAS-A and PAS-B domains of NPAS2 bind heme as a prosthetic group, realizing a gas-regulated sensor that modifies DNA binding in vitro depending on heme-status and modulating DNA binding in response to CO, with CO concentrations above 3 µM hindering DNA binding by holo-NPAS2 [27,28]. Thus, the apo (heme-free) or holo (heme-loaded) states of ARNTL:NPAS2 heterodimers strongly bind DNA under favorably reducing ratios of NADPH/NADP, while micromolar concentrations of CO impede the DNA binding activity of holo-NPAS2 but not that of apo-NPAS2 [27,28].
Mouse NPAS2 and ARNTL truncated forms containing only the corresponding bHLH domains and no PAS domain, completely retained NADPH-dependent DNA binding [27]. Thus, i) the NAPDH binding site must reside on the bHLH domain, and ii) most likely NADPH does not undergo redox changes in the bound state and behaves as an allosteric ligand; alternatively, in situ NAPDH autooxidation could set the lifetime of the DNA-heterodimer complex.
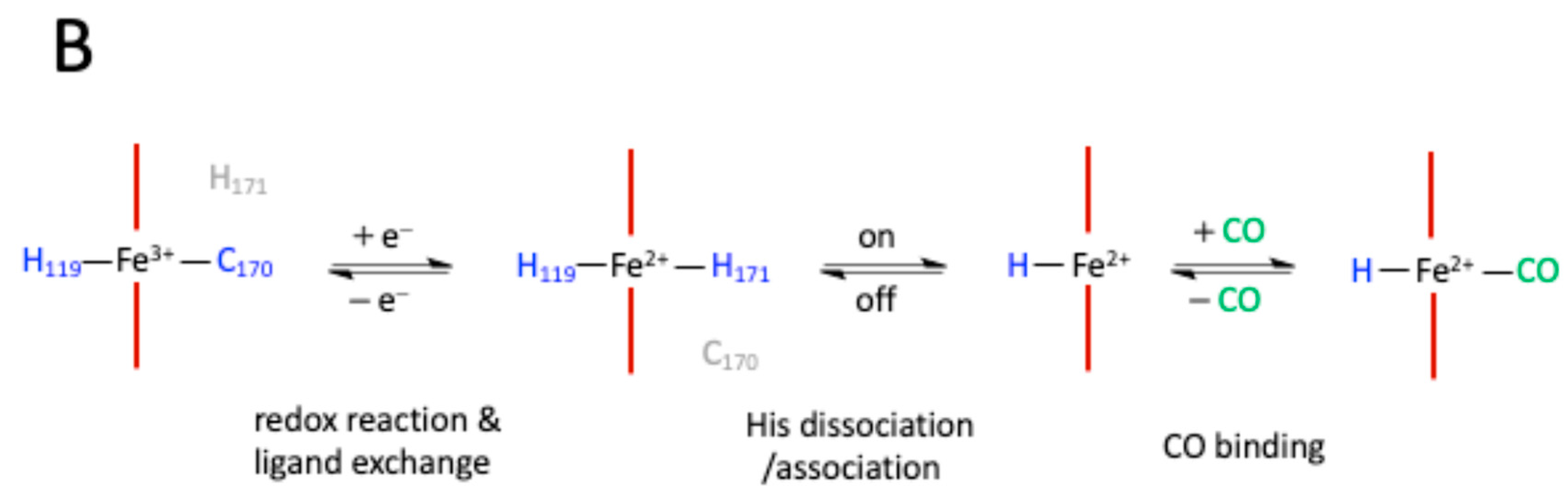
The presence of heme in NPAS2 is in order linked to heme metabolism. Indeed, haem biosynthesis is rhythmically controlled by the biological clock, which drives the circadian expression of Alas1, the gene coding for 5'-aminolevulinate synthase 1, the rate-limiting enzyme in the biochemical pathway of heme synthesis, bringing on a mutual regulation [29]. Elegant resonance Raman (RR) experiments coupled with mutational studies performed on a mouse isolated PAS-A domain [30,31] shed light on the iron coordination sphere of the ferric, ferrous and carbonylated states of the protein and on the spin state of the heme (Figure 1B). The overall conclusion of this work is that both the ferric and ferrous PAS-A domains consist of a mixture of five- and six-coordinate heme.
The resonance Raman spectra of the isolated PAS-A domain was measured upon excitation at 363.8 nm and a band assignable to Fe3+-S− stretching was detected at 334 cm-1 for the ferric protein, with Cys170 resulting an axial ligand of the ferric heme [30,31]. The Raman spectrum of the reduced form was mostly of six-coordinate low spin, and the ν11 band, which is sensitive to the donor strength of the axial ligand, was lower than that of reduced cytochrome c3, suggesting coordination of a strong ligand and thus a deprotonated His [30,31]. In the reduced forms of H119A and H171A, the five-coordinate species were more abundant, whereas no such changes were observed for C170A, indicating that His119 and His171, but not Cys170, are axial ligands in the ferrous heme [30,31]. This suggests that ligand replacement from Cys to His occurs upon heme reduction, while the νFe-CO versus νC-O correlation indicates that a neutral His is a trans ligand of CO [30,31]. In the isolated PAS-A domain His119 and Cys170 are the axial ligands of the ferric heme, while Cys170 is replaced by His171 upon reduction of heme. The coordination structure of the isolated PAS-A domain is in equilibrium between Cys-Fe-His and His-Fe-His coordinated species, but the interaction of the PAS-A domain with the bHLH domain shifts the equilibrium toward the second structure [30,31]. The fragment containing the N-terminal basic helix-loop-helix (bHLH) of the first PAS (PAS-A) domain of NPAS2 mostly occurs as a dimer in solution and the Soret absorption peak of the ferric complex for bHLH-PAS-A (421 nm) is red-shifted by 9 nm relative to isolated PAS-A (412 nm). Based on the RR spectra the axial ligand trans to CO in bHLH-PAS-A appears to be His. Furthermore, the rate constant for heme association with apo-bHLH-PAS is more than two orders of magnitude higher than for association with apo-PAS-A [32]. Optical absorption spectra of the PAS-B domain (residues 241-416) of mouse NPAS2 showed that the Fe(III), Fe(II) and Fe(II)-CO complexes are 6-coordinated low-spin complexes, while resonance Raman spectra showed that both the Fe(III) and Fe(II) complexes contain mixtures of 5-coordinated high-spin and 6-coordinated low-spin complexes [33]. Overall, the emerged picture of the heme environment is complex and most likely dynamic. Reduction of heme iron (by an as yet unknown electron donor) results in a unique endogenous ligand exchange reaction whereby the Cys170 is exchanged to His171 in the ferrous state. To bind exogenous ligands, one of the His ligands must dissociate from the ferrous ion (Figure 1B).
The human NPAS2 gene (alias MOP4) is the biggest core clock gene (176.68 kb) and was assigned to chromosome 2p11.2-2q13, while the mouse Npas2 gene was assigned to chromosome 1 at 21-22 centimorgans. The chromosomal regions to which human NPAS2 map are syntenic with those containing the mouse Npas2 genes, indicating that the mouse and human genes are true homologs [34]. Circadian transcription of NPAS2 gene, likewise ARNTL transcription, is regulated and synchronized through RORα and Rev-erbα competition at Rev-erb and retinoid-related orphan receptor response elements (Rev-REs and ROREs, respectively) in the upstream region of the transcription start site [35,36]. Additionally, a specific activating role is played by RORγ in the regulation of NPAS2 expression through direct binding onto two ROREs in its proximal promoter [37]. Experiments performed in a purified system showed that DNA-binding activity of the ARNTL:NPAS2 and ARNTL:CLOCK heterodimers is regulated by the redox state of NAD cofactors. NAD(H) and NADP(H), the reduced forms of the redox cofactors, powerfully enhance DNA binding of ARNTL:NPAS2 and ARNTL:CLOCK heterodimers, while NAD+ and NADP+, the oxidized forms, hinder DNA binding capability, suggesting a direct modulatory role played by cellular redox state on transcriptional activity in the circadian clock circuitry [27]. Modifications of cell pH from 7.0 to 7.5 in addition to additive enhancing activity of NADPH increases DNA binding capability of NPAS2, with its N-terminal amino acids 1-61 necessary to sense the change in both pH and NADPH [38]. NPAS2 can compensates for loss of the core transcription factor and histone/protein acetylase CLOCK in the molecular clockwork endowing SCN oscillators as well as peripheral tissues cells [39,40,41]. NPAS2 contains a heme-binding motif and heme controls the transcriptional activity of ARNTL:NPAS2 heterodimer [42]. Likewise its analog/paralog, NPAS2 can reversibly bind carbon monoxide (CO) in vitro and in vivo. CO binds the heme group in NPAS2 and inhibits the DNA-binding activity of the ARNTL:NPAS2 heterodimer. Endogenous CO is continuously produced during heme metabolism and modulates DNA binding patterns of NPAS2 and CLOCK onto the promoters of circadian genes. Experiments performed in a mouse model treated with hemoCD1, a supramolecular complex of iron(II)porphyrin with a per-O-methyl-β-cyclodextrin dimer with highly selective CO scavenging action, showed that endogenous CO removal leads to up-regulation of the E-box-controlled circadian genes Per1, Per2, Cry1, Cry2, and Rev-erbα in the liver, corroborating the role played by endogenous CO in the regulation of the mammalian circadian clock circuitry [43,44,45].
4. NPAS2 and Metabolic Pathways
Core circadian genes and tissue specific output genes drive rhythmic expression of hundreds transcripts enriching metabolic pathways involved in glucose and lipid metabolism as well as xenobiotics detoxification. Likewise, deregulation of circadian genes plays a crucial role in the pathophysiological mechanisms underlying metabolic derangements [46]. In the liver, Npas2 realizes with the nuclear receptor and transcriptional regulator small heterodimer partner (SHP) a feedback regulatory loop involved in triglyceride and lipoprotein homeostasis. SHP is a bona-fide transcriptional repressor of Npas2 and at the molecular level acts as a negative regulator of nuclear receptor-dependent signaling pathways, suppressing Rorγ transactivation and acting as a co-repressor of Rev-erbα. SHP enhances Rev-erbα inhibitory action on the positive transcriptional activity of Rorα at the Npas2 promoter, with subsequent inhibition of Npas2 transcription. On the contrary, NPAS2 binds to the Shp promoter and drives circadian Shp gene expression. In its side SHP, interplaying with RORα, RORγ, or REV-ERBα nuclear receptors-dependent signaling, modulates the regulatory role played by Npas2 in upholding 24-hour rhythms of lipid metabolism. As a result, Shp−/− mice are featured by severe derangement of time-qualified patterns of expression of fundamental genes implicated in cholesterol, fatty acid, bile acid, and lipid metabolism in the [47]. Experiments performed in vitro in hepatic cells (AML-12, Hepa1-6 and HepG2 cells) and in vivo in animal models (Npas2 double knockout mice) showed that NPAS2 drives 24-hour rhythms of expression and activity of hepatic CYP1A2 trans-activating its expression through specific binding to the -416 bp E-box-like element within the Cyp1a2 gene promoter [48]. CYP1A2 is a monooxygenase member of the cytochrome P450 enzyme superfamily that catalyzes numerous reactions involved in drug metabolism and the synthesis of cholesterol, steroids, and other lipids. CYP1A2 also intervenes in the metabolism of polyunsaturated fatty acids, transforming them into signaling molecules involved in physiological processes and pathophysiological mechanisms of disease. Likewise, NPAS2 was found crucial in determining time-qualified patterns of toxicity of brucine, one of the main bioactive and toxic constituents of the herb drug Semen Strychni. Brucine hepatotoxicity was evaluated by plasma alanine aminotransferase and aspartate aminotransferase measurements and histopathological analysis in wild type and Npas2-/- mice, and Npas2 double knock out evidently down-regulated Cyp3a11 expression at mRNA and protein levels as well as its enzymatic activity, with abolished circadian rhythmicity in brucine pharmacokinetics, liver distribution and hepatotoxicity [49].
5. NPAS2 and the Central Nervous System in Mammals
The biological clock plays a crucial role in affording neuroprotection and in restraining neuroinflammation. Experiments performed in double knockout mice showed that deletion of the transcriptional activator Arntl alone or concurrent ablation of Clock and Npas2 prompted severe age-dependent astrogliosis in the cortex and hippocampus, with degeneration of synaptic terminals, compromised cortical functional connectivity, decreased expression of numerous redox defense genes with consequent neuronal oxidative damage and striatal neurodegeneration [50]. Remarkably, NPAS2-deficient mice challenged with behavioral tests showed deficits in the long-term memory arm of the cued and contextual fear task, suggesting that NPAS2 may play a distinct regulatory role in the acquisition of specific types of memory [51]. NPAS2 is highly expressed in stress-related and reward-related brain regions. In this regard, Npas2-/- mice showed a smaller amount of anxiety-related behavior respect to controls when assessed through behavioral assays (Elevated Plus Maze, light/dark box and open field assay). Npas2 knockdown in the ventral striatum lead to a comparable decrease of anxiety-like behaviors, whereas acute or chronic stress increased striatal Npas2 expression. Furthermore, Npas2-/- mice showed reduced expression of genes coding for Gamma-Aminobutyric Acid (GABA) receptor subunits (principally Gabra1) and displayed decreased sensitivity to diazepam, a GABAa positive allosteric modulator in the ventral striatum, suggesting that Npas2 is crucially involve in the response to stress and the development of anxiety and takes part in the regulation of GABAergic neurotransmission in the ventral striatum [52]. Furthermore, Npas2-/- mice showed reduced sensibility to cocaine reward and NPAS2 knockdown specifically in the NAc recapitulated this effect, corroborating the important role played by NPAS2 in this region and especially in Drd1 expressing neurons [52]. Circadian rhythmicity and sleep homeostasis are modified by opioids and in turn sleep destructuration and circadian disruption conceivably reconcile the effects of opioids, exacerbating during opioid withdrawal and continuing during abstinence. In this context, NPAS2 is relevant in psychiatric disorders associated with deranged reward sensibility and was found to be enriched in reward-related brain regions, especially in the mammalian forebrain, comprising the nucleus accumbens (NAc), a main neural substrate of motivated and reward behavior and dopamine receptor 1 containing medium spiny neurons (D1R-MSNs) of the striatum. NPAS2 plays a role in reward regulation and substance self-administration and its downregulation in the NAc decreases the conditioned behavioral response to cocaine in mice [54]. NPAS2 drives time-qualified gene transcription in central nervous structures of the pain control system modulating opioids effects, with sex-specific effects in the regulation of fentanyl-induced tolerance, hyperalgesia and dependence. Experiments performed with male and female wild-type and NPAS2-/- mice showed that NPAS2 is pivotal in fentanyl analgesia, tolerance, hyperalgesia and physical dependence, with female NPAS2-/- animals showing increased analgesic tolerance and physical dependence to fentanyl [55]. Fentanyl impacts on sleep architecture and sleep-wake cycles, particularly during opioid withdrawal, and NPAS2 is involved in the modulation of sleep architecture and drug reward. In particular, NPAS2 and the NAD(+)-dependent deacetylase SIRT1 were found to cooperate and regulate reward in the mouse NAc, driving numerous reward-related and metabolic-connected pathways enriched through common gene targets [56]. Chronic fentanyl administration caused decreased non-rapid eye movement sleep, which persisted with progressive reduction during withdrawal and the alteration of sleep architecture was more evident in NPAS2-/- mice [57]. NAc-specific knockdown of Npas2 in male and female C57BL/6J mice modified accumbal excitatory synaptic transmission and strength. In addition, definite Npas2 knockdown in NAc modified the behavioral sensitivity to cocaine reward, through amplified excitatory drive on D1R-MSNs MSNs, but not to non-D1R-MSNs, obstructing cocaine-induced enhancement of synaptic strength and glutamatergic transmission specifically onto D1R-MSNs. These results suggest that NPAS2 regulates excitatory synapses of D1R-MSNs in the NAc and cocaine reward-related behavior synaptic adaptations and is essential for cocaine-induced plastic changes in MSNs expressing the dopamine 1 receptor (D1R), so that NPAS2 disruption in D1R-MSNs produces augmented cocaine preference [58]. Npas2 is involved also in the mechanisms underlying food reward. Npas2 and the transcription factor Egr1 mediate food reward effects and circadian pathways interplay in SCN, dorsomedial hypothalamus and prefrontal cortex, as evidenced in experiments performed with provision of small hedonic and caloric value food rewards to Winstar rats for 16 days 3 h after light phase onset [59]. NPAS2 was found involved in the regulation of sleep homeostasis and non-rapid eye movement sleep in particular. Experiments performed in Npas2-/- mice under conditions of augmented sleep necessity, such as at the end of the active period or after sleep deprivation, showed that Npas2 opens the way to sleep at the time-of-day when mice are customarily awake. Npas2 double knock out modified electroencephalografic activity of thalamo-cortical origin, lessening activity in the spindle range (10-15 Hz) and swinging activity in the delta range (1-4 Hz) toward faster frequencies during non-rapid eye movement sleep [60]. Remarkably, in female mice sleep need accumulated at a slower rate and rapid eye movement sleep loss was not recovered after sleep deprivation, suggesting a gender effect featuring the role played by Npas2 in sleep regulation [60]. An investigation of the allele distributions of a large database of DNA polymorphisms (SNPs genotyped in 2504 individuals, belonging to 26 worldwide populations from the 1000Genome browser) in human core clock, clock-controlled and clock-related genes and a reference data set of putatively neutral polymorphisms was performed using summary statistics to compare observed data with expectations generated under the null hypothesis of selective neutrality. Among the SNPs, 15 polymorphisms belonging to 11 clock genes were selected based on an association with human sleep disorders reported in the scientific literature, such as the advanced sleep phase syndrome and the delayed sleep phase syndrome, or with modifications in circadian phenotypes [61], at least in one world population. NPAS2 was recognize among loci showing evidence of positive selection with both local adaptation and clinical adaptation and in which populations living at different latitudes cuttingly differ from each other, hinting exposure to dissimilar patterns of seasonal changes of environmental variables, for example the photoperiod [62]. Regarding the role played in diagnosable psychiatric disorders, a series of genetic variants of NPAS2 gene were statistically associated with anxiety, mood, behavioral, eating, personality, psychotic and autistic spectrum as well as dementia-related disorders [63,64,65,66,67,68].
6. NPAS2 in Cancer Onset and Progression
Disruption of the circadian clock circuitry plays a crucial role in cancer onset and progression [69,70]. Derangement of the biological clock and single polymorphisms of some circadian genes have been linked to cancer susceptibility [71]. NPAS2 is regarded as a promising predictor of clinical outcome in various malignancies and was reported to behave as tumor suppressor in colorectal carcinogenesis [72]. DNA CpG methylation profile and chromatin modification patterns represent fundamental constituents of the cell epigenome and chromatin regulators are major controllers of gene transcription in normal cells, directing histone modifications and chromatin remodeling. The biological clock drives cycles in spatial and temporal chromosomal organization controlling rhythmicity of transcriptional activation and repression [73]. During carcinogenesis genetic mutations and/or epigenetic alterations disrupt the functions of chromatin regulators initiating wide-ranging derangement of gene expression [74]. In this regard, NPAS2 was identified as component of a four chromatin regulators hub (CBX7, HMGA2, NPAS2 and PRC1) allowing risk stratification and outcome prediction in lung adenocarcinoma patients and providing a gene signature enriching PI3K/Akt/mTOR pathway and p53 signaling and associated to the infiltration percentages of macrophage M0, macrophage M2, resting NK cells, memory B cells, dendritic cells in tumor microenvironment [75]. Accordingly, NPAS2 was comprised in a five-gene signature (DKK1, CCL20, NPAS2, GNPNAT1 and MELTF) predicting prognosis (decreased overall survival rates and shorter progression-free survival) and immunotherapy response of lung adenocarcinoma patients [76]. Likewise, higher expression of the NPAS2 gene in tumor tissue was closely related with immune infiltration and overall survival of glioma patients [77] as well as to immune infiltration and poor prognosis in hepatocellular carcinoma patients [78,79]. In the setting of hepatocellular carcinogenesis, upregulation of NPAS2 expression, chiefly attributable to the downregulation of miR-199b-5p, was found capable to reprogram glucose metabolism through transcriptional activation of HIF-1α with upregulation of the glycolytic genes GLUT1, HK2, GPI, ALDOA, ENO2, PKM2 and MCT4 and downregulation of peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC-1α) with subsequent reduced mitochondrial biogenesis [80]. Interestingly, NPAS2 was found upregulated in hepatic stellate cells (HSCs) after fibrogenic injury, with ensuing HSCs activation, capable to play a role in liver fibrogenesis though direct transcriptional activation in HSCs of hairy and enhancer of split 1 (Hes1), a critical transcription factor involved in Notch signaling [81]. Additionally, in anaplastic thyroid carcinoma NPAS2 was found to be significantly up-regulated in tumor tissue and in vitro experiments showed that NPAS2 silencing in anaplastic thyroid carcinoma cell lines successfully obstructed cell proliferation, migration and invasion [82]. In the context of hormone-sensitive cancers, NPAS2 gene down-regulation was found associated with poorly differentiated tumors in comparison with well and moderately differentiated breast cancer [83] and rs2305160 polymorphism of NPAS2 gene (Ala394Thr) was found strongly associated with breast cancer risk and proposed as candidate breast cancer susceptibility locus [84,85,86]. Remarkably, Ala394Thr polymorphism in the NPAS2 gene was identified as modifier for the risk of non-Hodgkin's lymphoma, with robust association of the variant Thr genotypes (Ala/Thr and Thr/Thr) with reduced risk of lymphomagenesis [87]. Regarding other hormone-dependent tumors, NPAS2 dysregulation was found associated with poor prognosis and clinic-pathological characteristics of patients suffering from uterine corpus endometrial carcinoma (UCEC). NPAS2 upregulation in UCEC tissue samples compared to matched non-tumorous tissue was associated with higher clinical stage and tumor grade, estrogen receptor status, myometrial invasion, leading to decreased overall, disease free and relapse free survival in UCEC patients. Besides, NPAS2 showed significant negative correlations with miR-17-5p and miR-93-5p and positive correlations with miR106a-5p and miR-381-3p and NPAS2 knockdown or overexpression in vitro in UCEC cell lines impacted cell proliferation and apoptosis [88]. Among malignant genito-urinary tract neoplasms, NPAS2 expression in prostate neoplasia suggested tumor suppressor function and showed significant correlation with the immune components of the tumor microenvironment and was identified as part of a gene signature capable to predict probability of disease progression in prostate cancer patients [89]. In tumor tissue of prostate cancer patients NPAS2 expression was found upregulated respect to matched non-tumorous tissue. NPAS2 knockdown in prostate cancer cell lines hindered cell proliferation and upholded cell apoptosis in vitro and restrained tumor growth in vivo in xenotransplanted nude mice. At the molecular level, NPAS2 supported glycolysis and restrained oxidative phosphorylation in prostate cancer cell lines through amplification of hypoxia-inducible factor-1A (HIF-1A)-dependent signaling. Accordingly, NPAS2 knockdown decreased glucose uptake and lactate production, while increasing intracellular pH and oxygen consumption rate [90]. An association between NPAS2 expression and prostate cancer was also corroborated in the EPICAP study, a population-based case-control study with genotyped data that investigated the relationship between circadian genes polymorphisms and prostate cancer risk [91]. The non-synonymous mutations and genetic variants rs1369481, rs895521, and rs17024926 in NPAS2 gene were found significantly associated with susceptibility to prostate cancer, considering overall risk and risk of aggressive disease [92]. Moreover, the NPAS2 gene single nucleotide polymorphism rs6542993 A>T was found significantly associated with higher risk of disease progression in both localized and advanced prostate cancer cases, with decreased NPAS2 expression levels found in carriers of the T allele of rs6542993 compared with those carrying the A allele and associated with more aggressive prostate cancer and poor progression-free survival [93]. On the other hand, NPAS2 genetic variant rs895520 was found associated with a statistically significant higher predisposition to sarcoma and leiomyosarcoma [94]. Distinct expression of luminal and basal gene features muscle-invasive bladder cancers, practical biomarkers to predict disease progression and overall survival. NPAS2 was found to be clinically relevant and involved in the regulation of subtype-specific genes influencing cancer cell proliferation and migration in luminal bladder cancers [95]. As regards myeloproliferative neoplasms, NPAS2 was found upregulated in acute myeloid leukemia cells and experiments performed both in vivo and in vitro highlighted its fundamental role in supporting cell survival and proliferation as well as in restraining apoptosis at various stages of myeloid differentiation. NPAS2 knockdown obstructed CDC25A expression leading to G1/S cell cycle arrest, augmented caspase-3 cleavage and promoted cell death through Bcl2/Bax production modification [96]. A similar survival-promoting role was found for NPAS2 in HCC through transcriptional activation of the CDC25A phosphatase and consequent dephosphorylation of CDK2/4/6 and Bcl-2, which prompted cell proliferation and obstructed cell apoptosis, correspondingly [97]. MicroRNAs (miRNAs) are small, single-stranded, non-coding RNA molecules containing 21 to 23 nucleotides that post-transcriptionally regulate expression of coding genes in a sequence-specific manner. MicroRNAs modulate oscillating mRNA levels and are essential for generating a time delay critical for the molecular clockwork [98]. MicroRNAs impact cancer onset and progression as well as response to therapy modifying gene expression. RNA-seq and miRNA-omics analysis performed in radio-resistant nasopharyngeal cancer (NPC) cells pinpointed NPAS2-miR-20a-5p axis as crucially involved in NPC radio-resistance and this role was corroborated by experiments with down- or up-regulation of their levels in NPC cell lines [99]. Sequencing of putative promoter and 5' untranslated region of the NPAS2 promoter in patients suffering from melanoma identified several variants. Of particular interest was a microsatellite comprising a GGC repeat with different alleles ranging from 7 to 13 repeats located in the 5' untranslated exon. Homozygosity of an allele with nine repeats (9/9) was more prevalent in melanoma patients respect to control subjects, suggesting a role for NPAS2 variants in melanoma susceptibility [100].
7. NPAS2 and the Cardiovascular System
The various cogs of the molecular clockwork show spatial-temporal and cell-specific patterns of expression and localization throughout the cardiovascular system. The biological clock controls rhythmic patterns of molecular signaling driving vascular remodeling, vascular morphology and tone modulating vascular resistance and blood pressure fluctuations [101]. Mouse models with deleted or mutated core clock genes, such as ARNTL-/-, Clock mutant, and Npas2 mutant mice, show disordered rhythms of fluctuation in blood pressure and heart rate [102]. In this regard, genetic variants in circadian genes comprising NPAS2 were found associated with the diurnal phenotype of hypertension, proposing a genetic association with diurnal blood pressure changes in essential hypertension. In particular, five tag SNPs within five loci, including rs3888170 in NPAS2, rs6431590 in PER2, rs1410225 in RORββ, rs3816358 in ARNTL and rs10519096 in RORα, were found significantly associated with the non-dipper phenotype in young hypertensive patients [103]. Cardiomyocytes hold an intrinsic biological clock that drives 24-hour rhythmic variations in myocardial biology as well [104]. Incidence of cardiovascular events such as myocardial infarction, stroke, and sudden death show circadian variation and genes involved in hemostasis and vascular integrity experience 24-hour rhythmic fluctuations influencing thrombotic events. In this regard, in a mouse model of thrombotic vascular occlusion following a photochemical injury NPAS2 deletion extended the time to thrombotic vascular occlusion and decreased blood pressure irrespective of time-of-day of prothrombotic challenge [105]. Heart failure is a condition caused by damage to the heart's structure and/or function lessening cardiac muscle capability to pump enough blood throughout the circulatory system to supply adequate oxygen and nutrients amount to body tissues. This syndrome represents one of the most common health problems world-wide and in particular in the western countries, is related to numerous pathological conditions, primarily metabolic, inflammatory, infective, degenerative, and manifests with multiple clinical symptoms. Circadian disruption could play a role in heart failure onset and worsening [106] and NPAS2 resulted down-regulated in heart failure patients when expression data of ischemic and dilated cardiomyopathy samples with or without heart failure from the GEO database were analyzed with bioinformatics tools [107]. Interestingly, NPAS2 was comprised in the set of genes found deregulated in myocardial transcriptome upon methamphetamine challenge. These genetic changes showed sex-specific pattern and the number of deregulated genes and the degree of the variation were significantly greater in female hearts respect to male hearts [108]. The role played by altered regulation of circadian genes in the pathogenesis of cardiovascular disease seems to be corroborated also in the case of cerebrovascular events. In this regard, the DNA methylation patterns of a set of core clock genes, comprising NPAS2, was found modified in patients suffering from ischemic stroke when previously exposed to particulate matter (PM2.5 exposure before the event).These epigenetic traits are impacted by environmental factors, such as pollutants exposure, and suggest the possibility that epigenetic changes in circadian genes could contribute to stroke development or alternatively could be used as prognostic marker of stroke [109]. The role of NPAS2 as gas-responsive transcription factor was explored in a mouse model of subarachnoid hemorrhage (SAH), which is featured by a time-qualified pattern of stroke incidence that was recapitulated through blood injection in subarachnoid space at different circadian time points. Various outcome were evaluated, comprising core clock gene expression, locomotor activity, vasospasm, neuro-inflammation and apoptosis markers. In addition, core clock gene expression was assayed in cerebrospinal fluid and peripheral blood leukocytes obtained from SAH patients and compared to control subjects [110]. Statistically significant upregulation of NPAS2, Per1 and Per2 was reported in the hippocampus, cortex, and SCN in mice subjected to SAH at zeitgeber time (ZT) 12 respect to ZT2. Heme oxygenase-1 (HO-1/Hmox1) was also significantly higher at ZT12 and correlated with expression amplitude of circadian genes. Interestingly, mice subjected to SAH at ZT 12 showed a significant decrease in cerebral vasospasm, neuronal apoptosis, and microglial activation respect to mice subjected to SAH at ZT 2 [110]. Besides, expression of NPAS2, Per1 and Per2 in the SCN was decreased while injury was augmented in animals with myeloid-specific HO-1 deletion. Low-dose CO rescued these alterations, suggesting that Npas2 and other core clock genes play a role in determining the severity of SAH and myeloid HO-1 activity is necessary to alleviate erythrocyte burden and lessen neuronal apoptosis [110].
8. NPAS2 and the Process of Wound Healing
Notwithstanding implementation of proper procedures of surgical wound closure scarring continues to represent a puzzling problem. NPAS2 is expressed in dermal fibroblasts and has been shown to play a critical role in regulating collagen synthesis during wound healing. Npas2 down-regulation plays a role for scarless wound healing and Npas2-/- mice showed improved healing of dermal excisional wounds [111]. Besides, high-throughput drug screening pinpointed a compound that downregulates Npas2 activity. Pharmacological inhibition of Npas2 expression by Dwn1 augmented murine dermal fibroblast cell migration and reduced collagen synthesis in vitro. When locally applied to iatrogenic full-thickness dorsal cutaneous wounds the expression of type I collagen, Tgfβ1, and α-smooth muscle actin was significantly reduced in a murine model and dermal wounds treated with Dwn1 healed more rapidly with satisfactory mechanical strength and were featured by a smaller amount of granulation tissue respect to controls [112].
9. Conclusions
NPAS2 is a core cog of the molecular clockwork in SCN and peripheral tissues and comprises two PAS domains and a basic-helix-loop-helix domain. The PAS-A and PAS-B domains support protein-protein interactions and heterodimerization with ARNTL, so as to functionally substitute for the core transcription factor and histone/protein acetylase CLOCK. The PAS domains are also capable to bind a heme moiety each, up keeping an additional feedback mechanism that fine tunes the biological clock through wavering in heme availability and gas-responsive CO-dependent modulation of transcriptional activity. On its side, the basic-helix-loop-helix domain provides a scaffold for specific DNA binding within target enhancer elements through which NPAS2 can take part in wide-ranging regulation of transcriptional circuits entailed in the control of biological processes and biochemical functions involved in a series of physiological events and pathogenic mechanisms of disease.
Author Contributions
G.M. and F.M. conceived the article; F.M. drew the figure; E.M, M.M.B., F.M., G.M. and T.C. reviewed the scientific literature, wrote the paper. All authors approved the final version of the manuscript and agreed to the published version of the manuscript.
Funding
This research was funded by the “5 × 1000” voluntary contribution and by a grant from the Italian Ministry of Health (Ricerca Corrente 2022-2024) to G.M.
Data Availability Statement
Not applicable.
Acknowledgments
We apologize to authors whose relevant publications were not cited due to space limitations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chaix A, Zarrinpar A, Panda S. The circadian coordination of cell biology. J Cell Biol. 2016 Oct 10;215(1):15-25. [CrossRef]
- Hastings MH, Reddy AB, Maywood ES. A clockwork web: circadian timing in brain and periphery, in health and disease. Nat Rev Neurosci. 2003 Aug;4(8):649-61. [CrossRef]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Boil. 2014, 24, 90–99.
- Maywood ES, O'Neill JS, Reddy AB, Chesham JE, Prosser HM, Kyriacou CP, Godinho SI, Nolan PM, Hastings MH. Genetic and molecular analysis of the central and peripheral circadian clockwork of mice. Cold Spring Harb Symp Quant Biol. 2007;72:85-94. [CrossRef]
- Ray S, Reddy AB. Cross-talk between circadian clocks, sleep-wake cycles, and metabolic networks: Dispelling the darkness. Bioessays. 2016 Apr;38(4):394-405. [CrossRef]
- Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science. 2010 Dec 3;330(6009):1349-54. [CrossRef]
- Hastings MH, Maywood ES, Brancaccio M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat Rev Neurosci. 2018;19(8):453-469. [CrossRef]
- Varadarajan S, Tajiri M, Jain R, et al. Connectome of the Suprachiasmatic Nucleus: New Evidence of the Core-Shell Relationship. eNeuro. 2018;5(5):ENEURO.0205-18.2018. Published 2018 Oct 2. [CrossRef]
- Brancaccio M, Patton AP, Chesham JE, Maywood ES, Hastings MH. Astrocytes Control Circadian Timekeeping in the Suprachiasmatic Nucleus via Glutamatergic Signaling. Neuron. 2017;93(6):1420-1435.e5. [CrossRef]
- Reick, M., Garcia, J. A., Dudley, C., & McKnight, S. L. (2001). NPAS2: an analog of clock operative in the mammalian forebrain. Science (New York, N.Y.), 293(5529), 506–509. [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179.
- Milev NB, Rhee SG, Reddy AB. Cellular Timekeeping: It's Redox o'Clock. Cold Spring Harb Perspect Biol. 2018 May 1;10(5):a027698. [CrossRef]
- Robinson I, Reddy AB. Molecular mechanisms of the circadian clockwork in mammals. FEBS Lett. 2014 Aug 1;588(15):2477-83. [CrossRef]
- Reddy AB. Genome-wide analyses of circadian systems. Handb Exp Pharmacol. 2013;(217):379-88. [CrossRef]
- Cardone, L.; Hirayama, J.; Giordano, F.; Tamaru, T.; Palvimo, J.J.; Sassone-Corsi, P. Circadian clock control by SUMOylation of BMAL1. Science 2005, 309, 1390–1394.
- Lee, J.; Lee, Y.; Lee, M.J.; Park, E.; Kang, S.H.; Chung, C.H.; Lee, K.H.; Kim, K. Dual modification of BMAL1 by SUMO2/3 and ubiquitin promotes circadian activation of the CLOCK/BMAL1 complex. Mol. Cell. Boil. 2008, 28, 6056–6065.
- Sahar, S.; Zocchi, L.; Kinoshita, C.; Borrelli, E.; Sassone-Corsi, P. Regulation of BMAL1 protein stability and circadian function by GSK3beta-mediated phosphorylation. PLoS ONE 2010, 5, e8561.
- Duez H, Staels B. Rev-erb-alpha: an integrator of circadian rhythms and metabolism. J Appl Physiol (1985). 2009 Dec;107(6):1972-80. [CrossRef]
- Hughes, M.E.; DiTacchio, L.; Hayes, K.R.; Vollmers, C.; Pulivarthy, S.; Baggs, J.E.; Panda, S.; Hogenesch, J.B. Harmonics of circadian gene transcription in mammals. PLoS Genet. 2009, 5, e1000442.
- Bozek, K.; Relogio, A.; Kielbasa, S.M.; Heine, M.; Dame, C.; Kramer, A.; Herzel, H. Regulation of clock-controlled genes in mammals. PLoS ONE 2009, 4, e4882.
- Noshiro, M., Furukawa, M., Honma, S., Kawamoto, T., Hamada, T., Honma, K., & Kato, Y. (2005). Tissue-specific disruption of rhythmic expression of Dec1 and Dec2 in clock mutant mice. Journal of biological rhythms, 20(5), 404–418. [CrossRef]
- Doi M, Hirayama J, Sassone-Corsi P. Circadian regulator CLOCK is a histone acetyltransferase. Cell. 2006 May 5;125(3):497-508. [CrossRef]
- Nakahata, Y.; Kaluzova, M.; Grimaldi, B.; Sahar, S.; Hirayama, J.; Chen, D.; Guarente, L.P.; Sassone-Corsi, P. The NAD+-dependent deacetylase SIRT1 modulates CLOCK-mediated chromatin remodeling and circadian control. Cell 2008, 134, 329–340.
- Asher, G.; Gatfield, D.; Stratmann, M.; Reinke, H.; Dibner, C.; Kreppel, F.; Mostoslavsky, R.; Alt, F.W.; Schibler, U. SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell 2008, 134, 317–328.
- Nakahata, Y.; Sahar, S.; Astarita, G.; Kaluzova, M.; Sassone-Corsi, P. Circadian control of the NAD+ salvage pathway by CLOCK-SIRT1. Science 2009, 324, 654–657.
- Ramsey, K.M.; Yoshino, J.; Brace, C.S.; Abrassart, D.; Kobayashi, Y.; Marcheva, B.; Hong, H.K.; Chong, J.L.; Buhr, E.D.; Lee, C.; et al. Circadian clock feedback cycle through NAMPT-mediated NAD+ biosynthesis. Science 2009, 324, 651–654.
- Rutter, J., Reick, M., Wu, L. C., & McKnight, S. L. (2001). Regulation of clock and NPAS2 DNA binding by the redox state of NAD cofactors. Science (New York, N.Y.), 293(5529), 510–514. [CrossRef]
- Dioum, E. M., Rutter, J., Tuckerman, J. R., Gonzalez, G., Gilles-Gonzalez, M. A., & McKnight, S. L. (2002). NPAS2: a gas-responsive transcription factor. Science (New York, N.Y.), 298(5602), 2385–2387. [CrossRef]
- Kaasik, K., & Lee, C. C. (2004). Reciprocal regulation of haem biosynthesis and the circadian clock in mammals. Nature, 430(6998), 467–471. [CrossRef]
- Uchida, T., Sato, E., Sato, A., Sagami, I., Shimizu, T., & Kitagawa, T. (2005). CO-dependent activity-controlling mechanism of heme-containing CO-sensor protein, neuronal PAS domain protein 2. The Journal of biological chemistry, 280(22), 21358–21368. [CrossRef]
- Uchida, T., Sagami, I., Shimizu, T., Ishimori, K., & Kitagawa, T. (2012). Effects of the bHLH domain on axial coordination of heme in the PAS-A domain of neuronal PAS domain protein 2 (NPAS2): conversion from His119/Cys170 coordination to His119/His171 coordination. Journal of inorganic biochemistry, 108, 188–195. [CrossRef]
- Mukaiyama, Y., Uchida, T., Sato, E., Sasaki, A., Sato, Y., Igarashi, J., Kurokawa, H., Sagami, I., Kitagawa, T., & Shimizu, T. (2006). Spectroscopic and DNA-binding characterization of the isolated heme-bound basic helix-loop-helix-PAS-A domain of neuronal PAS protein 2 (NPAS2), a transcription activator protein associated with circadian rhythms. The FEBS journal, 273(11), 2528–2539. [CrossRef]
- Koudo, R., Kurokawa, H., Sato, E., Igarashi, J., Uchida, T., Sagami, I., Kitagawa, T., & Shimizu, T. (2005). Spectroscopic characterization of the isolated heme-bound PAS-B domain of neuronal PAS domain protein 2 associated with circadian rhythms. The FEBS journal, 272(16), 4153–4162. [CrossRef]
- Zhou, Y. D., Barnard, M., Tian, H., Li, X., Ring, H. Z., Francke, U., Shelton, J., Richardson, J., Russell, D. W., & McKnight, S. L. (1997). Molecular characterization of two mammalian bHLH-PAS domain proteins selectively expressed in the central nervous system. Proceedings of the National Academy of Sciences of the United States of America, 94(2), 713–718. [CrossRef]
- Crumbley, C., Wang, Y., Kojetin, D. J., & Burris, T. P. (2010). Characterization of the core mammalian clock component, NPAS2, as a REV-ERBalpha/RORalpha target gene. The Journal of biological chemistry, 285(46), 35386–35392. [CrossRef]
- Matsumura, R., Matsubara, C., Node, K., Takumi, T., & Akashi, M. (2013). Nuclear receptor-mediated cell-autonomous oscillatory expression of the circadian transcription factor, neuronal PAS domain protein 2 (NPAS2). The Journal of biological chemistry, 288(51), 36548–36553. [CrossRef]
- Takeda, Y., Kang, H. S., Angers, M., & Jetten, A. M. (2011). Retinoic acid-related orphan receptor γ directly regulates neuronal PAS domain protein 2 transcription in vivo. Nucleic acids research, 39(11), 4769–4782. [CrossRef]
- Yoshii, K., Tajima, F., Ishijima, S., & Sagami, I. (2015). Changes in pH and NADPH regulate the DNA binding activity of neuronal PAS domain protein 2, a mammalian circadian transcription factor. Biochemistry, 54(2), 250–259. [CrossRef]
- DeBruyne, J. P., Weaver, D. R., & Reppert, S. M. (2007). CLOCK and NPAS2 have overlapping roles in the suprachiasmatic circadian clock. Nature neuroscience, 10(5), 543–545. [CrossRef]
- Bertolucci, C., Cavallari, N., Colognesi, I., Aguzzi, J., Chen, Z., Caruso, P., Foá, A., Tosini, G., Bernardi, F., & Pinotti, M. (2008). Evidence for an overlapping role of CLOCK and NPAS2 transcription factors in liver circadian oscillators. Molecular and cellular biology, 28(9), 3070–3075. [CrossRef]
- Landgraf, D., Wang, L. L., Diemer, T., & Welsh, D. K. (2016). NPAS2 Compensates for Loss of CLOCK in Peripheral Circadian Oscillators. PLoS genetics, 12(2), e1005882. [CrossRef]
- Itoh, R., Fujita, K., Mu, A., Kim, D. H., Tai, T. T., Sagami, I., & Taketani, S. (2013). Imaging of heme/hemeproteins in nucleus of the living cells expressing heme-binding nuclear receptors. FEBS letters, 587(14), 2131–2136. [CrossRef]
- Ascenzi, P., Bocedi, A., Leoni, L., Visca, P., Zennaro, E., Milani, M., & Bolognesi, M. (2004). CO sniffing through heme-based sensor proteins. IUBMB life, 56(6), 309–315. [CrossRef]
- Gilun, P., Stefanczyk-Krzymowska, S., Romerowicz-Misielak, M., Tabecka-Lonczynska, A., Przekop, F., & Koziorowski, M. (2013). Carbon monoxide-mediated humoral pathway for the transmission of light signal to the hypothalamus. Journal of physiology and pharmacology: an official journal of the Polish Physiological Society, 64(6), 761–772.
- Minegishi, S., Sagami, I., Negi, S., Kano, K., & Kitagishi, H. (2018). Circadian clock disruption by selective removal of endogenous carbon monoxide. Scientific reports, 8(1), 11996. [CrossRef]
- Mazzoccoli, G., De Cosmo, S., & Mazza, T. (2018). The Biological Clock: A Pivotal Hub in Non-alcoholic Fatty Liver Disease Pathogenesis. Frontiers in physiology, 9, 193. [CrossRef]
- Lee, S. M., Zhang, Y., Tsuchiya, H., Smalling, R., Jetten, A. M., & Wang, L. (2015). Small heterodimer partner/neuronal PAS domain protein 2 axis regulates the oscillation of liver lipid metabolism. Hepatology (Baltimore, Md.), 61(2), 497–505. [CrossRef]
- He, Y., Cen, H., Guo, L., Zhang, T., Yang, Y., Dong, D., & Wu, B. (2022). Circadian oscillator NPAS2 regulates diurnal expression and activity of CYP1A2 in mouse liver. Biochemical pharmacology, 206, 115345. [CrossRef]
- Zhou, Z., Lin, Y., Gao, L., Yang, Z., Wang, S., & Wu, B. (2019). Cyp3a11 metabolism-based chronotoxicity of brucine in mice. Toxicology letters, 313, 188–195. [CrossRef]
- Musiek, E. S., Lim, M. M., Yang, G., Bauer, A. Q., Qi, L., Lee, Y., Roh, J. H., Ortiz-Gonzalez, X., Dearborn, J. T., Culver, J. P., Herzog, E. D., Hogenesch, J. B., Wozniak, D. F., Dikranian, K., Giasson, B. I., Weaver, D. R., Holtzman, D. M., & Fitzgerald, G. A. (2013). Circadian clock proteins regulate neuronal redox homeostasis and neurodegeneration. The Journal of clinical investigation, 123(12), 5389–5400. [CrossRef]
- Garcia, J. A., Zhang, D., Estill, S. J., Michnoff, C., Rutter, J., Reick, M., Scott, K., Diaz-Arrastia, R., & McKnight, S. L. (2000). Impaired cued and contextual memory in NPAS2-deficient mice. Science (New York, N.Y.), 288(5474), 2226–2230. [CrossRef]
- Ozburn, A. R., Kern, J., Parekh, P. K., Logan, R. W., Liu, Z., Falcon, E., Becker-Krail, D., Purohit, K., Edgar, N. M., Huang, Y., & McClung, C. A. (2017). NPAS2 Regulation of Anxiety-Like Behavior and GABAA Receptors. Frontiers in molecular neuroscience, 10, 360. [CrossRef]
- Ozburn, A. R., Falcon, E., Twaddle, A., Nugent, A. L., Gillman, A. G., Spencer, S. M., Arey, R. N., Mukherjee, S., Lyons-Weiler, J., Self, D. W., & McClung, C. A. (2015). Direct regulation of diurnal Drd3 expression and cocaine reward by NPAS2. Biological psychiatry, 77(5), 425–433. [CrossRef]
- DePoy, L. M., Becker-Krail, D. D., Zong, W., Petersen, K., Shah, N. M., Brandon, J. H., Miguelino, A. M., Tseng, G. C., Logan, R. W., & McClung, C. A. (2021). Circadian-Dependent and Sex-Dependent Increases in Intravenous Cocaine Self-Administration in Npas2 Mutant Mice. The Journal of neuroscience : the official journal of the Society for Neuroscience, 41(5), 1046–1058. [CrossRef]
- Puig, S., Shelton, M. A., Barko, K., Seney, M. L., & Logan, R. W. (2022). Sex-specific role of the circadian transcription factor NPAS2 in opioid tolerance, withdrawal and analgesia. Genes, brain, and behavior, 21(7), e12829. [CrossRef]
- Becker-Krail, D. D., Parekh, P. K., Ketchesin, K. D., Yamaguchi, S., Yoshino, J., Hildebrand, M. A., Dunham, B., Ganapathiraju, M. K., Logan, R. W., & McClung, C. A. (2022). Circadian transcription factor NPAS2 and the NAD+-dependent deacetylase SIRT1 interact in the mouse nucleus accumbens and regulate reward. The European journal of neuroscience, 55(3), 675–693. [CrossRef]
- Gamble, M. C., Chuan, B., Gallego-Martin, T., Shelton, M. A., Puig, S., O'Donnell, C. P., & Logan, R. W. (2022). A role for the circadian transcription factor NPAS2 in the progressive loss of non-rapid eye movement sleep and increased arousal during fentanyl withdrawal in male mice. Psychopharmacology, 239(10), 3185–3200. [CrossRef]
- Parekh, P. K., Logan, R. W., Ketchesin, K. D., Becker-Krail, D., Shelton, M. A., Hildebrand, M. A., Barko, K., Huang, Y. H., & McClung, C. A. (2019). Cell-Type-Specific Regulation of Nucleus Accumbens Synaptic Plasticity and Cocaine Reward Sensitivity by the Circadian Protein, NPAS2. The Journal of neuroscience: the official journal of the Society for Neuroscience, 39(24), 4657–4667. [CrossRef]
- Herichová, I., Hasáková, K., Lukáčová, D., Mravec, B., Horváthová, Ľ., & Kavická, D. (2017). Prefrontal cortex and dorsomedial hypothalamus mediate food reward-induced effects via npas2 and egr1 expression in rat. Physiological research, 66(Suppl 4), S501–S510. [CrossRef]
- Franken, P., Dudley, C. A., Estill, S. J., Barakat, M., Thomason, R., O'Hara, B. F., & McKnight, S. L. (2006). NPAS2 as a transcriptional regulator of non-rapid eye movement sleep: genotype and sex interactions. Proceedings of the National Academy of Sciences of the United States of America, 103(18), 7118–7123. [CrossRef]
- Evans, D. S., Parimi, N., Nievergelt, C. M., Blackwell, T., Redline, S., Ancoli-Israel, S., Orwoll, E. S., Cummings, S. R., Stone, K. L., Tranah, G. J., Study of Osteoporotic Fractures (SOF), & Osteoporotic Fractures in Men (MrOS) Study Group (2013). Common genetic variants in ARNTL and NPAS2 and at chromosome 12p13 are associated with objectively measured sleep traits in the elderly. Sleep, 36(3), 431–446. [CrossRef]
- Dall'Ara, I., Ghirotto, S., Ingusci, S., Bagarolo, G., Bertolucci, C., & Barbujani, G. (2016). Demographic history and adaptation account for clock gene diversity in humans. Heredity, 117(3), 165–172. [CrossRef]
- Yeim, S., Boudebesse, C., Etain, B., & Belliviera, F. (2015). Biomarqueurs et gènes circadiens dans le trouble bipolaire [Circadian markers and genes in bipolar disorder]. L'Encephale, 41(4 Suppl 1), S38–S44. [CrossRef]
- Kim, H. I., Lee, H. J., Cho, C. H., Kang, S. G., Yoon, H. K., Park, Y. M., Lee, S. H., Moon, J. H., Song, H. M., Lee, E., & Kim, L. (2015). Association of CLOCK, ARNTL, and NPAS2 gene polymorphisms and seasonal variations in mood and behavior. Chronobiology international, 32(6), 785–791. [CrossRef]
- Milhiet, V., Boudebesse, C., Bellivier, F., Drouot, X., Henry, C., Leboyer, M., & Etain, B. (2014). Circadian abnormalities as markers of susceptibility in bipolar disorders. Frontiers in bioscience (Scholar edition), 6(1), 120–137. [CrossRef]
- Partonen T. (2012). Clock gene variants in mood and anxiety disorders. Journal of neural transmission (Vienna, Austria : 1996), 119(10), 1133–1145. [CrossRef]
- Soria, V., Martínez-Amorós, E., Escaramís, G., Valero, J., Pérez-Egea, R., García, C., Gutiérrez-Zotes, A., Puigdemont, D., Bayés, M., Crespo, J. M., Martorell, L., Vilella, E., Labad, A., Vallejo, J., Pérez, V., Menchón, J. M., Estivill, X., Gratacòs, M., & Urretavizcaya, M. (2010). Differential association of circadian genes with mood disorders: CRY1 and NPAS2 are associated with unipolar major depression and CLOCK and VIP with bipolar disorder. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology, 35(6), 1279–1289. [CrossRef]
- Johansson, C., Willeit, M., Smedh, C., Ekholm, J., Paunio, T., Kieseppä, T., Lichtermann, D., Praschak-Rieder, N., Neumeister, A., Nilsson, L. G., Kasper, S., Peltonen, L., Adolfsson, R., Schalling, M., & Partonen, T. (2003). Circadian clock-related polymorphisms in seasonal affective disorder and their relevance to diurnal preference. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology, 28(4), 734–739. [CrossRef]
- Yang, Y., Lindsey-Boltz, L. A., Vaughn, C. M., Selby, C. P., Cao, X., Liu, Z., Hsu, D. S., & Sancar, A. (2021). Circadian clock, carcinogenesis, chronochemotherapy connections. The Journal of biological chemistry, 297(3), 101068. [CrossRef]
- Sancar, A., & Van Gelder, R. N. (2021). Clocks, cancer, and chronochemotherapy. Science (New York, N.Y.), 371(6524), eabb0738. [CrossRef]
- Mocellin, S., Tropea, S., Benna, C., & Rossi, C. R. (2018). Circadian pathway genetic variation and cancer risk: evidence from genome-wide association studies. BMC medicine, 16(1), 20. [CrossRef]
- Rao, X., & Lin, L. (2022). Circadian clock as a possible control point in colorectal cancer progression (Review). International journal of oncology, 61(6), 149. [CrossRef]
- Aguilar-Arnal L., Hakim O., Patel V.R., Baldi P., Hager G.L., and Sassone-Corsi P.. 2013. Cycles in spatial and temporal chromosomal organization driven by the circadian clock. Nat. Struct. Mol. Biol. 20:1206–1213. [CrossRef]
- Shu, X. S., Li, L., & Tao, Q. (2012). Chromatin regulators with tumor suppressor properties and their alterations in human cancers. Epigenomics, 4(5), 537–549. [CrossRef]
- Shi, Q., Han, S., Liu, X., Wang, S., & Ma, H. (2022). Integrated single-cell and transcriptome sequencing analyses determines a chromatin regulator-based signature for evaluating prognosis in lung adenocarcinoma. Frontiers in oncology, 12, 1031728. [CrossRef]
- Tang, X., Qi, C., Zhou, H., & Liu, Y. (2022). A novel metabolic-immune related signature predicts prognosis and immunotherapy response in lung adenocarcinoma. Heliyon, 8(8), e10164. [CrossRef]
- Chai, R., Liao, M., Ou, L., Tang, Q., Liang, Y., Li, N., Huang, W., Wang, X., Zheng, K., & Wang, S. (2022). Circadian Clock Genes Act as Diagnostic and Prognostic Biomarkers of Glioma: Clinic Implications for Chronotherapy. BioMed research international, 2022, 9774879. [CrossRef]
- Zhang, Z., Liang, Z., Gao, W., Yu, S., Hou, Z., Li, K., & Zeng, P. (2022). Identification of circadian clock genes as regulators of immune infiltration in Hepatocellular Carcinoma. Journal of Cancer, 13(11), 3199–3208. [CrossRef]
- Liu, H., Gao, Y., Hu, S., Fan, Z., Wang, X., & Li, S. (2021). Bioinformatics Analysis of Differentially Expressed Rhythm Genes in Liver Hepatocellular Carcinoma. Frontiers in genetics, 12, 680528. [CrossRef]
- Yuan, P., Yang, T., Mu, J., Zhao, J., Yang, Y., Yan, Z., Hou, Y., Chen, C., Xing, J., Zhang, H., & Li, J. (2020). Circadian clock gene NPAS2 promotes reprogramming of glucose metabolism in hepatocellular carcinoma cells. Cancer letters, 469, 498–509. [CrossRef]
- Yang, T., Yuan, P., Yang, Y., Liang, N., Wang, Q., Li, J., Lu, R., Zhang, H., Mu, J., Yan, Z., & Chang, H. (2019). NPAS2 Contributes to Liver Fibrosis by Direct Transcriptional Activation of Hes1 in Hepatic Stellate Cells. Molecular therapy. Nucleic acids, 18, 1009–1022. [CrossRef]
- Xu, T., Jin, T., Lu, X., Pan, Z., Tan, Z., Zheng, C., Liu, Y., Hu, X., Ba, L., Ren, H., Chen, J., Zhu, C., Ge, M., & Huang, P. (2022). A signature of circadian rhythm genes in driving anaplastic thyroid carcinoma malignant progression. Cellular signalling, 95, 110332. [CrossRef]
- Lesicka, M., Jabłońska, E., Wieczorek, E., Seroczyńska, B., Siekierzycka, A., Skokowski, J., Kalinowski, L., Wąsowicz, W., & Reszka, E. (2018). Altered circadian genes expression in breast cancer tissue according to the clinical characteristics. PloS one, 13(6), e0199622. [CrossRef]
- Yi, C., Mu, L., de la Longrais, I. A., Sochirca, O., Arisio, R., Yu, H., Hoffman, A. E., Zhu, Y., & Katsaro, D. (2010). The circadian gene NPAS2 is a novel prognostic biomarker for breast cancer. Breast cancer research and treatment, 120(3), 663–669. [CrossRef]
- Monsees, G. M., Kraft, P., Hankinson, S. E., Hunter, D. J., & Schernhammer, E. S. (2012). Circadian genes and breast cancer susceptibility in rotating shift workers. International journal of cancer, 131(11), 2547–2552. [CrossRef]
- Wang, B., Dai, Z. M., Zhao, Y., Wang, X. J., Kang, H. F., Ma, X. B., Lin, S., Wang, M., Yang, P. T., & Dai, Z. J. (2015). Current evidence on the relationship between two common polymorphisms in NPAS2 gene and cancer risk. International journal of clinical and experimental medicine, 8(5), 7176–7183.
- Zhu, Y., Leaderer, D., Guss, C., Brown, H. N., Zhang, Y., Boyle, P., Stevens, R. G., Hoffman, A., Qin, Q., Han, X., & Zheng, T. (2007). Ala394Thr polymorphism in the clock gene NPAS2: a circadian modifier for the risk of non-Hodgkin's lymphoma. International journal of cancer, 120(2), 432–435. [CrossRef]
- Zheng, X., Lv, X., Zhu, L., Xu, K., Shi, C., Cui, L., & Ding, H. (2021). The Circadian Gene NPAS2 Act as a Putative Tumor Stimulative Factor for Uterine Corpus Endometrial Carcinoma. Cancer management and research, 13, 9329–9343. [CrossRef]
- Feng, D., Xiong, Q., Zhang, F., Shi, X., Xu, H., Wei, W., Ai, J., & Yang, L. (2022). Identification of a Novel Nomogram to Predict Progression Based on the Circadian Clock and Insights Into the Tumor Immune Microenvironment in Prostate Cancer. Frontiers in immunology, 13, 777724. [CrossRef]
- Ma, S., Chen, Y., Quan, P., Zhang, J., Han, S., Wang, G., Qi, R., Zhang, X., Wang, F., Yuan, J., Yang, X., Jia, W., & Qin, W. (2023). NPAS2 promotes aerobic glycolysis and tumor growth in prostate cancer through HIF-1A signaling. BMC cancer, 23(1), 280. [CrossRef]
- Wendeu-Foyet, M. G., Koudou, Y., Cénée, S., Trétarre, B., Rébillard, X., Cancel-Tassin, G., Cussenot, O., Boland, A., Bacq, D., Deleuze, J. F., Lamy, P. J., Mulot, C., Laurent-Puig, P., Truong, T., & Menegaux, F. (2019). Circadian genes and risk of prostate cancer: Findings from the EPICAP study. International journal of cancer, 145(7), 1745–1753. [CrossRef]
- Zhu, Y., Stevens, R. G., Hoffman, A. E., Fitzgerald, L. M., Kwon, E. M., Ostrander, E. A., Davis, S., Zheng, T., & Stanford, J. L. (2009). Testing the circadian gene hypothesis in prostate cancer: a population-based case-control study. Cancer research, 69(24), 9315–9322. [CrossRef]
- Yu, C. C., Chen, L. C., Chiou, C. Y., Chang, Y. J., Lin, V. C., Huang, C. Y., Lin, I. L., Chang, T. Y., Lu, T. L., Lee, C. H., Huang, S. P., & Bao, B. Y. (2019). Genetic variants in the circadian rhythm pathway as indicators of prostate cancer progression. Cancer cell international, 19, 87. [CrossRef]
- Benna, C., Rajendran, S., Spiro, G., Tropea, S., Del Fiore, P., Rossi, C. R., & Mocellin, S. (2018). Associations of clock genes polymorphisms with soft tissue sarcoma susceptibility and prognosis. Journal of translational medicine, 16(1), 338. [CrossRef]
- Iyyanki, T., Zhang, B., Wang, Q., Hou, Y., Jin, Q., Xu, J., Yang, H., Liu, T., Wang, X., Song, F., Luan, Y., Yamashita, H., Chien, R., Lyu, H., Zhang, L., Wang, L., Warrick, J., Raman, J. D., Meeks, J. J., DeGraff, D. J., … Yue, F. (2021). Subtype-associated epigenomic landscape and 3D genome structure in bladder cancer. Genome biology, 22(1), 105. [CrossRef]
- Song, B., Chen, Y., Liu, Y., Wan, C., Zhang, L., & Zhang, W. (2019). NPAS2 regulates proliferation of acute myeloid leukemia cells via CDC25A-mediated cell cycle progression and apoptosis. Journal of cellular biochemistry, 120(5), 8731–8741. [CrossRef]
- Yuan, P., Li, J., Zhou, F., Huang, Q., Zhang, J., Guo, X., Lyu, Z., Zhang, H., & Xing, J. (2017). NPAS2 promotes cell survival of hepatocellular carcinoma by transactivating CDC25A. Cell death & disease, 8(3), e2704. [CrossRef]
- Chen, R., D'Alessandro, M., & Lee, C. (2013). miRNAs are required for generating a time delay critical for the circadian oscillator. Current biology : CB, 23(20), 1959–1968. [CrossRef]
- Zhao, F., Pu, Y., Qian, L., Zang, C., Tao, Z., & Gao, J. (2017). MiR-20a-5p promotes radio-resistance by targeting NPAS2 in nasopharyngeal cancer cells. Oncotarget, 8(62), 105873–105881. [CrossRef]
- Franzoni, A., Markova-Car, E., Dević-Pavlić, S., Jurišić, D., Puppin, C., Mio, C., De Luca, M., Petruz, G., Damante, G., & Pavelić, S. K. (2017). A polymorphic GGC repeat in the NPAS2 gene and its association with melanoma. Experimental biology and medicine (Maywood, N.J.), 242(15), 1553–1558. [CrossRef]
- Anea, C. B., Merloiu, A. M., Fulton, D. J. R., Patel, V., & Rudic, R. D. (2018). Immunohistochemistry of the circadian clock in mouse and human vascular tissues. Vessel plus, 2, 16. [CrossRef]
- Curtis, A. M., Cheng, Y., Kapoor, S., Reilly, D., Price, T. S., & Fitzgerald, G. A. (2007). Circadian variation of blood pressure and the vascular response to asynchronous stress. Proceedings of the National Academy of Sciences of the United States of America, 104(9), 3450–3455. [CrossRef]
- Leu, H. B., Chung, C. M., Lin, S. J., Chiang, K. M., Yang, H. C., Ho, H. Y., Ting, C. T., Lin, T. H., Sheu, S. H., Tsai, W. C., Chen, J. H., Yin, W. H., Chiu, T. Y., Chen, C. I., Fann, C. S., Chen, Y. T., Pan, W. H., & Chen, J. W. (2015). Association of circadian genes with diurnal blood pressure changes and non-dipper essential hypertension: a genetic association with young-onset hypertension. Hypertension research: official journal of the Japanese Society of Hypertension, 38(2), 155–162. [CrossRef]
- Bray, M. S., Shaw, C. A., Moore, M. W., Garcia, R. A., Zanquetta, M. M., Durgan, D. J., Jeong, W. J., Tsai, J. Y., Bugger, H., Zhang, D., Rohrwasser, A., Rennison, J. H., Dyck, J. R., Litwin, S. E., Hardin, P. E., Chow, C. W., Chandler, M. P., Abel, E. D., & Young, M. E. (2008). Disruption of the circadian clock within the cardiomyocyte influences myocardial contractile function, metabolism, and gene expression. American journal of physiology. Heart and circulatory physiology, 294(2), H1036–H1047. [CrossRef]
- Westgate, E. J., Cheng, Y., Reilly, D. F., Price, T. S., Walisser, J. A., Bradfield, C. A., & FitzGerald, G. A. (2008). Genetic components of the circadian clock regulate thrombogenesis in vivo. Circulation, 117(16), 2087–2095. [CrossRef]
- Takeda, N., & Maemura, K. (2015). The role of clock genes and circadian rhythm in the development of cardiovascular diseases. Cellular and molecular life sciences : CMLS, 72(17), 3225–3234. [CrossRef]
- Sun, Q., Zhao, J., Liu, L., Wang, X., & Gu, X. (2023). Identification of the potential biomarkers associated with circadian rhythms in heart failure. PeerJ, 11, e14734. [CrossRef]
- Chavva, H., Brazeau, D. A., Denvir, J., Primerano, D. A., Fan, J., Seeley, S. L., & Rorabaugh, B. R. (2021). Methamphetamine-induced changes in myocardial gene transcription are sex-dependent. BMC genomics, 22(1), 259. [CrossRef]
- Cantone, L., Tobaldini, E., Favero, C., Albetti, B., Sacco, R. M., Torgano, G., Ferrari, L., Montano, N., & Bollati, V. (2020). Particulate Air Pollution, Clock Gene Methylation, and Stroke: Effects on Stroke Severity and Disability. International journal of molecular sciences, 21(9), 3090. [CrossRef]
- Schallner, N., Lieberum, J. L., Gallo, D., LeBlanc, R. H., 3rd, Fuller, P. M., Hanafy, K. A., & Otterbein, L. E. (2017). Carbon Monoxide Preserves Circadian Rhythm to Reduce the Severity of Subarachnoid Hemorrhage in Mice. Stroke, 48(9), 2565–2573. [CrossRef]
- Clements, A., Shibuya, Y., Hokugo, A., Brooks, Z., Roca, Y., Kondo, T., Nishimura, I., & Jarrahy, R. (2023). In vitro assessment of Neuronal PAS domain 2 mitigating compounds for scarless wound healing. Frontiers in medicine, 9, 1014763. [CrossRef]
- Shibuya, Y., Hokugo, A., Okawa, H., Kondo, T., Khalil, D., Wang, L., Roca, Y., Clements, A., Sasaki, H., Berry, E., Nishimura, I., & Jarrahy, R. (2022). Therapeutic downregulation of neuronal PAS domain 2 (Npas2) promotes surgical skin wound healing. eLife, 11, e71074. [CrossRef]
Figure 1.
Molecular structure and biochemistry of the circadian protein NPAS2, an important hemeprotein and gas-responsive transcription factor. A. Schematic of the NPAS2 structure (not to scale) and its regulation. B. Redox, endogenous ligand dissociation/association and CO binding scheme of the PAS-A domain.
Figure 1.
Molecular structure and biochemistry of the circadian protein NPAS2, an important hemeprotein and gas-responsive transcription factor. A. Schematic of the NPAS2 structure (not to scale) and its regulation. B. Redox, endogenous ligand dissociation/association and CO binding scheme of the PAS-A domain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



