
Submitted:
25 August 2023
Posted:
28 August 2023
You are already at the latest version
Abstract
Genetic engineering technology offers opportunities to improve many important agronomic traits in crops, including insect-resistance. However, genetically modified (GM) exogenous proteins in edible tissues of transgenic crops has become an issue of intense public concern. To advance the application of GM techniques in maize, a Cre/loxP-based strategy was developed for manipulating the transgenes in green tissues while locking it in non-green tissues. In the strategy, the site-specific excision can be used to switch on or off the expression of transgenes at specific tissues. In this work, two basic transgenic maize named KEY carrying the Cre gene and LOCK containing the Vip3A gene with a blocked element, were obtained based on their separate fusion gene cassettes. The expression level and concentration of Vip3A were observed with high specific accumulation in green tissues (leaf and stem), and only a small amount in root and kernel tissues in the KEY × LOCK hybrids. The insect-resistance of transgenic maize against two common lepidopteran pests, Ostrinia furnacalis and Spodoptera frugiperda, was assessed in the laboratory and field. The results indicated that the hybrids possessed high resistance levels against the two pests, with mortality rates above 73.6% and damage scales below 2.4 compared with the control group. Our results suggested that the Cre/loxP-mediated genetic engineering approach has a competitive advantage in GM maize. Overall, findings from this study are significant for providing a feasible strategy for transgenes avoiding expression in edible parts and exploring novel techniques toward the biosafety of GM plants.
Keywords:
genetically modified
; Cre/loxP
; site-specific excision
; transgenic maize
; insect-resistance
1. Introduction
Maize is one of the primary staple food crops and plays a pivotal role in sustaining global food security. In maize growing areas, many biotic stresses affect production capacity and safety, including the corn borer (Ostrinia furnacalis Guenée [Crambidae]) and the fall armyworm (Spodoptera frugiperda [J. E. Smith]), resulting in yield and quality loss [1,2,3]. Currently, general management for preventing the threat of pests relies primarily on chemical insecticides, which may lead to possible environmental and health hazards [4]. The use of genetically modified (GM) crops expressing Bacillus thuringiensis (Bt) proteins is the most common method for controlling herbivorous pests [5,6,7]. Transgenic maize expressing Vip3A (Vegetative insecticidal protein 3) protein from Bt are widely used in GM crops to control lepidopteran pests and have been planted for more than two decades [8,9,10]. Because of their high control efficacy, transgenic Bt crops have increased consistently since their initial commercialization in the USA during the 1990s[11]. Although Bt maize has been widely planted at multiple countries in recent decades, its biosafety is still controversial in certain countries [12,13]. One possible reason is that exogenous proteins of GM crops were expressed simultaneously in various tissues, including the edible parts, which would lead to potential risks for environment or consumer health [14].
To relieve public concerns about GM crops, several genetic engineering techniques have been developed to manipulate transgenes expression in confined tissues [15,16,17,18,19,20,21,22]. For instance, the Cre/loxP site-specific recombination system is the most-mentioned tool for precisely manipulating introduced DNA in transgenic plants due to its simplicity and efficiency [23,24]. In the system, Cre recombinase, responsible for specific excision of the sequences embedded by directly repeated loxP sites, has been applied to remove transgenes from the host genome [25]. Based on the system, a new strategy has been developed to enable expression of transgenes in desired tissues, depending on the site-specific excision reaction occuring at particular places. Here, two basic transgenic plants, individual Cre and loxP, are required for performing a site-specific excision reaction in their heterologous [26,27,28]. The loxP plants that carry transgenes blocked by a terminator can be crossed with Cre plants that express Cre activity, in which case the terminator can then be excised away in their F1 progeny. In this case, the blocked transgenes in the loxP plants can be activated and allowed to express after the excision reaction at specific places. Therefore, the presence of the Cre in particular tissues is a key role in determining the tissue-specific expression of transgenes. The Cre gene can be drived by diverse promoters and then expressed in specific tissues, and could prompt the excision reaction occurring in defined tissues, thereby enabling the expression of blocked transgenes in target tissues. This promising Cre/loxP-mediated system has previously been applied in transgenic tobacco, rice and oilseed rape for manipulating transgenes expression in desired tissues [26,27,29].
Here, we developed a comprehensive strategy based on the Cre/loxP-mediated system for manipulating transgene expression in maize green tissues outside edible parts to relieve public concerns. In this work, the transgene Vip3A was explicitly expressed in green tissues owing to the presence of the Cre in the specific tissues when the two basic transgenic maize were brought together. Moreover, the crossed hybrids were evaluated with high resistance against insect pests. These findings illustrate a feasible strategy for manipulating transgenes expression in specific tissues and also provide a valuable tool for GM safety in other transgenic plants.
2. Materials and Methods
2.1. Construction of fusion gene cassette
The green tissue-specific promoter Zm1rbcS was chosen according to previous relevant studies [26,30], given its excellent characteristics in driving specific expression in green tissues of the related genes. Under the control of the Zm1rbcS promoter, a cassette named pKEY was contructed to drive Cre gene expression in green tissues. The other cassette named pLOCK containing an inserted NosT between the pairwise lox sites following the transgene Vip3A, was driven by the strong constitutive promoter ZmUbi (Figure 1). Herein, the sequences of genes Cre and Vip3A, were kindly provided by Dr. Chen (Zhejiang University, China). The two fusion cassettes were made in plant vector pCAMBIA3300 and then transferred into Agrobacterium tumefaciens strain EHA105 [31]. The neomycin phosphotransferase gene (nptII) was a marker gene for the selection of transgenic plants during transformation. Vectors and primers are listed in Supplementary Materials Table S1 and Figure S1.
2.2. Plant material and transformation
Each contructed cassette was introduced into the wild-type maize inbred line KN5585. The individual transgenic maize containing Cre and Vip3A were called KEY and LOCK plants. Transgenic plants were either self-pollinated or crossed to generate KEY × LOCK hybrids. All plants were grown in the transgenic base of Sichuan Agricultural University (Chengdu, Sichuan Province, China) (30° 43′ 11.06″N; 103° 52′ 12.30″E). Each plant was sown in single 3-m row plots, with a distance of 50 cm between plants within a row and 75 cm between rows, following a randomized block design. No pesticides were applied during the maize growth period.
2.3. Tissue expression in transgenic plants
The expression levels of Cre and Vip3A were detected in various tissues (root, stem, leaf, sheath, husk, silk and kernel) of the different transgenic maize. Total RNAs from different maize tissues were extracted using TRIzol reagent (Invitrogen, USA) according to the manual. A quantitative real-time PCR (qRT-PCR) was performed, with 18S rRNA as the internal control. Thermal cycle conditions were as follows: 2 min at 95°C followed by 40 cycles of 15 s at 95°C, 15 s at 56-57°C, and 15 s at 72°C. Each PCR reaction was repeated with three technical replicates.
2.4. Concentration of the Vip3A protein
The concentration of Vip3A protein in the various tissues (root, stem, leaf, sheath, husk, silk and kernel) of the different transgenic maize was determined using Enzyme-linked immunosorbent assay (ELISA) (Agdia®, USA). Each sample was added to a 96-well ELISA microplate in triplicate. The protein concentration was calculated using a linear regression equation for the standard curve including only the triplicates with a coefficient of variation lower than 20%.
2.5. Bioassay in laboratory
We performed a diet-overlay bioassay to evaluate insect-resistance of transgenic maize against two common lepidopteran pests, Ostrinia furnacalis and Spodoptera frugiperda. In Southwest China, transgenic maize is not planted. The original populations of the two pests were collected from this area and have not previously been exposed to Bt proteins. The two pests were reared on an artificial diet in the laboratory and maintained under controlled conditions (27 ± 1°C, 14: 10 light/dark photoperiod, and 70–75% RH). Eggs of the two pests were commercially acquired (Keyun Biotechnology Co., Ltd., China) and reared in the laboratory. When plants reached the V6 growth stage, bioassays were started to evaluate the resistant response of transgenic maize. In the laboratory, fresh leaves of different maize were cut into pieces of approximately 3 cm2 and placed into a 24-well bioassay tray (Tsingke Biotechnology Co., Ltd., Beijing, China). Each well of the tray was added one first-instar larvael of a single pest for each maize. Three trays were set up for each maize in the insect-bioassay. The non-transgenic maize KN5585 and individual LOCK plants were served as negative and positive control, respectively. The number of living larvae, dead larvae, and their larval stages was observed and recorded after infestation. If a larva had not developed beyond the first instar, it would be counted as dead. Larval mortality was calculated as mortality (%) = 100 × number of dead larvae plus the number of surviving larvae still in the first instar divided by the total number of pests assayed [9]. The mortality rate was corrected for each kind of maize using Abbott’s formula [32].
2.6. Field tests for insect resistance
We also performed a field test in addition to characterizing the resistance of transgenic maize. The same two pests of O. furnacalis and S. frugiperda were inoculated into each maize when the plants reached the V6 stage under field conditions. For each maize, 30 plants were chosen randomly for a single pest implementation as a replicate, and three replicates were conducted for a single larval infestation in the field test. Each plant was infested with 60 first-instar larvae on the whorls, and the plant leaf damage scale of each maize was calculated as the mean score of the inoculated maize. The KN5585 and LOCK plants were controls. The damage index of the plants was surveyed 14 days after artificial infestation and calculated using a 0-9 whole-plant leaf damage scale according to the described study [33].
2.7. Statistical analyses
The data for Vip3A protein concentration was compared with the control using a Student’s t-test. The percentages of larval mortality and plant leaf damage scale were analyzed using a one-way analysis of variance (ANOVA) model to determine the insect-resistant response variable between the hybrid and controls. The data of insect-resistance in each maize were conducted using the Dunnett multiple comparison method for a significant difference test (P < 0.05). All statistical analyses were made using SPSS 21.0 (http://www.spss.com).
3. Results
3.1. Fusion gene cassette and plant transformation
We constructed two separate fusion gene cassettes to develop an alternative strategy to manipulate transgenes expression at specific tissues. The fusion gene cassettes were each built into plant vector pCAMBIA3300 (Figure S1). The green tissue-specific promoter Zm1rbcS was constructed into the pKEY cassette in front of the Cre gene to drive the recombinase expression specifically in green tissues (Figure 1). The other pLOCK cassette comprised the strong constitutive promoter ZmUbi and exogenous Vip3A gene, in which a NosT was inserted between the pairwise lox sites to block the expression of the transgene (Figure 1). The two cassettes were each transformed into the wild-type maize inbred line KN5585 background. Putative transgenic plants were initially screened for kanamycin resistance and then confirmed with molecular characterization. A total of seven individual events for pKEY and 140 events for pLOCK were produced, respectively. PCR analyses of transgenic events showed that the expected amplicons corresponding to genes Cre and Vip3A, respectively, were present in their transformed lines (Figure S2). Finally, these transgenic events were subsequently self-pollinated to obtain advanced generations. The transgenic maize containing of Cre and Vip3A were called KEY and LOCK plants.
3.2. Tissue-specific expression of transgenes
The homozygous T2 progenies of the individual KEY or LOCK plants and the F1 population of KEY × LOCK hybrids were used to detect specific expression levels of the Cre and Vip3A. Various tissues (root, stem, leaf, sheath, husk, silk and kernel) from different trangenic maize were checked using qRT-PCR. Results showed that the highest expression level of the Cre appeared in the leaf, followed by stem and sheath, and a small amount of expression in the root, silk, and kernel, indicating that the Cre gene under the control of the green tissue-specific promoter Zm1rbcS was activated specifically at green tissues in KEY plants and KEY × LOCK hybrids (Figure 2a). However, the expression of Vip3A was observed at a low level in various tissues of LOCK plants, which might be explained that the transgenes were inhibited by the inserted NosT (Figure 2b). Moreover, the expression of Vip3A in the KEY × LOCK hybrids was detected with high level in the leaf and stem. Still rare accumulation in other tissues including root and kernel, reflects that the expression level of Vip3A in the green tissues of the hybrids is extensively activated compared with those in individual LOCK plants.
3.3. The concentration of the insecticidal protein
We performed an ELISA method to track insecticidal protein in the various tissues (root, stem, leaf, sheath, husk, silk and kernel) of the trangenic maize. In the KEY × LOCK hybrids, the mean concentration of Vip3A protein reached 9.59 ng/g, 6.13 ng/g in leaf and stem, and only 0.68 ng/g, 0.71 ng/g in root and kernel, respectively (Table 1). Besides, the protein was detected to keep at a low level (ranging from 0.43 ng/g to 0.74 ng/g) in LOCK plant tissues. Expectedly, the content pattern of Vip3A protein in the hybrids was similar to the model of the above tissue expression. From these results, the leaf and stem tissues contained higher protein levels than those in the root and kernel parts, reflecting that the Vip3A protein is a specific accumulation in green tissues. After comparing the concentration of Vip3A protein in the different trangenic maize, the Vip3A protein was probably blocked at a low expression in the LOCK plants, whereas it might be activated in leaf and stem tissues when the KEY and LOCK plants were brought together.
3.4. The efficiency of insect resistance in transgenic maize
A bioactivity test was conducted using the two major insect pests O. furnacalis and S. frugiperda larvae to examine insect-resistant efficiency of the transgenic maize. The neonate larvae (1st instar) of the two insect pests were single manually fed in the laboratory with fresh leaves of the hybrids and control maize, and the mortality rate was evaluated after infestation via the diet-overlay bioassay. The leaf damage of the maize in the control trays became apparent after two days and was heavily damaged after three days of implementation (Figure 3(a,b), (d,e)). In detail, the mean mortality rates of O. furnacalis larvae fed with control plants were 8.3% and 10.6% (Table 2). Similarly, the mortality rates of S. frugiperda were 8.7% and 9.2% in the control trays, respectively (Table 2). No significant effects of the mean mortality rates were detected in the control maize. At the same time, the mean mortality of O. furnacalis and S. frugiperda larvae fed on the KEY × LOCK hybrids were overall high with rates of 81.2% and 73.6% after three days of implementation, respectively (Figure 3c,f, Table 2). Statistical analyses showed a significant difference in mean mortality rates between the hybrids and control maize (O. furnacalis: F2, 6 = 349.61, P < 0.001; S. frugiperda: F2, 6 = 278.69, P < 0.001) (Table 2). The results showed that the hybrids exhibited high resistant performance on protection of maize with almost no damage and were significantly better than those of controls.
To further confirm the insect resistance response of the transgenic maize, the same two pests, O. furnacalis and S. frugiperda larvae, were used in the field. The obvious damages were observed in the control plants after 14 days of implementation when the two instar larvae were introduced onto the maize whorls (Figure 4(a,b), (d,e)). Damaged scales from O. furnacalis infestation were 7.5 (KN5585) and 7.7 (LOCK), respectively, and there were no significant differences among them (Table 2). Similarly, the damaged scales caused by S. frugiperda showed no significant differences among the controls with 8.1 and 7.9, respectively (Table 2). In contrast, the damaged scales in the hybrids were only 2.4 for O. furnacalis and 1.6 for S. frugiperda, respectively, suggesting that the hybrids could increase the protective efficacy for the maize (Figure 4c,f). Statistical analyses showed a significant difference in plant leaf damage scale between the hybrids and control maize (O. furnacalis: F2, 6 = 117.58, P < 0.001. S. frugiperda: F2, 6 = 196.43, P < 0.001) (Table 2). The insect-resistant performance in the field followed the results of the diet-overlay bioassay in laboratory. High mortality rates and low damage scales were recorded during the infestation, implying that the hybrids may have sufficient insecticidal Vip3A protein to protect maize against the two lepidopteran pests.
4. Discussion
As GM foods are starting to be present in our diet, the biosafety of GM crops has attracted significant public concern about whether these foods are harmful to their health [23,34]. We have focused on the insect-resistant transgenic maize since no accumulation of transgenes in edible parts may be more acceptable to the public. Therefore, we developed an available strategy based on the Cre/loxP-mediated system for manipulating Vip3A gene expression in defined green tissues outside kernel or seed. Consequently, two separate gene cassettes were constructed to generate two individual transgenic maize, KEY and LOCK (Figure 1). The comprehensive design here also appeared in previous studies. For instance, Luo et al. successfully constructed the Cre/loxP with pollen and seed-specific promoters system to remove functional transgenes from target tissues [28]. Another study used the Cre/loxP and seed-specific cruciferin C promoter resulting in 10% of marker-free transgenic tobacco plants [29]. Chen et al. developed a Cre/loxP gene switch system to limit Cry protein in rice green tissues [26]. Boszorádová et al. combined the Cre/loxP and the embryo-specific CRUC promoter to remove hazardous transgenes from the genome in commercial oilseed rape [27]. For this comprehensive strategy, diverse promoters are crucial regulatory elements and efficiently impact on the expression of transgenes in defined tissues [35,36,37]. In the current study, the green tissue-specific promoter Zm1rbcS with moderate drive capability was chosen to induce the expression of the simple gene Cre in KEY plants. However, the exogenous gene Vip3A required a powerful promoter to produce a sufficient quantity of insecticidal protein. Therefore, by taking advantage of the characteristics of the Cre/loxP-mediated system and these two types of promoters, the problem of tissue-specific expression compatible with insect resistance for GM maize can be resolved through the pyramiding of KEY and LOCK plants.
The presence of Cre protein in green tissues is necessary for the effective control of Vip3A protein in defined tissues. In our study, the Cre gene was expressed specifically in green tissues including leaf and stem, with low levels in root and kernel, suggesting that the gene should be specifically induced to express at tissues under the control of the green tissue-specific promoter Zm1rbcS (Figure 2a). Moreover, the expression of Vip3A in LOCK plants was observed at a low level. In contrast, a high level was detected in the KEY × LOCK hybrids leaf and stem. Still, rare accumulation in the root and kernel reflects that the Vip3A might be activated in target tissues of the hybrids (Figure 2b). The concentration of Vip3A protein in the hybrids harboring both Cre and Vip3A has been found with a very high accumulation in green tissues (Figure 3). It was speculated that the excision reaction might have occurred in target green tissues when the KEY and LOCK plants were brought together. Then, the inserted NosT of LOCK plants might be removed due to Cre presence, thereby allowing expression of the blocked Vip3A in the excision tissues. It should be noted that the Zm1rbcS promoter does not completely control the green tissue-specific expression of Vip3A due to its background expression in non-green tissues, thus leading to a trace amount of Vip3A protein observed in root and kernel tissues. These results are parallel to other similar studies in transgenic plants. For instance, the amount of GM protein detected in rbcS-driven Cry1C rice endosperm was 2.6 ng/g in the tested samples [38], and PEPC-driven Cry1Ab maize kernels were 15-18 ng/mg soluble protein [39]. A trace amount of transgene product was also observed in PNZIP-driven Bt-cotton seeds and PDX1-driven GUS-rice endosperm [40,41]. The Cre/loxP-mediated approach has significantly minimized the presence of exogenous Vip3A protein in edible parts, although it remains a slight imperfection. However, this was a preliminary step in manipulating exogenous Vip3A expression in maize green tissues. Further work is needed, including promoting the effectiveness of the green tissue-specific promoter, blocked capacity of the NosT, and excision efficiency of Cre. In brief, these results offered an alternative strategy to manipulate transgenes expression at specific maize tissues outside of edible parts by combing the two individual transgenic plants.
Referring to this Cre/loxP-mediated approach, insect-esistance of the transgenic plants against pest attact regarding the efficiency of the insecticidal protein was evaluated in the laboratory and field. High mortality rates and low damaged scales were recorded in the hybrids compared with the control group (Figure 3 and Figure 4). The results demonstrated that the hybrids showed a high degree of resistance against the two lepidopteran pests, implying that the Cre/loxP-mediated approach was effective for generating new transgenic maize with good resistance. These findings thus suggested that the insect-resistant transgenic maize hybrids could serve as elite germplasm resources for extensive Bt application in GM maize breeding, as well as for the research of plant genetic engineering [40], control of gene expression [26], and so on.
5. Conclusions
In the expected development and commercial exploitation of insect-resistant transgenic maize, the “excess” exogenous protein should preferably be outside edible parts, which might be more acceptable to the public. Tissue-specific expression of Vip3A protein in GM maize may help control the occurrence of pest attacks and limit the number of toxins in inedible tissues. Our results advocate that such Cre/loxP-mediated approach can be recommended in plant genetic engineering. The strategy has several advantages through combining the Cre/loxP and tissue-specific promoters to manipulate hazardous transgene expression in target tissues. First, Cre recombinase expression is mainly accumulated in the desired tissues under strict control of the tissue-specific promoters, leading the subsequent excision in the targeted tissues. Second, the strategy is just to shut down or turn on transgenes in only targeted tissues without removing DNA from the plant genome, which can be applied more widely. Third, tissue-specific promoter in KEY plants or blocked transgenes in LOCK plants can be replaced conveniently according to different demands for tissue-specific expression or desired traits. In this regard, the current Cre/loxP-mediated approach could provide more options for manipulating transgene expression in specific tissues and advance the plant genetic engineering concept closer to commercial implementation for GM maize.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Fusion gene cassettes pKEY and pLOCK were constructed in the Cre/loxP-mediated system; Figure S2: Photographs of transgenes with PCR products obtained on T2 transgenic maize plants. Table S1: Primers used for transgenes identification.
Author Contributions
Conceptualization, G.Y. and Y.S.; methodology, C.Z and H.S.; formal analysis, Y.Y.; investigation, J.D.; data curation, C.Z.; writing—original draft preparation, G.Y.; writing—review and editing, L.M.; supervision, G.P. and Y.S.; project administration, all authors; funding acquisition, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Transgenic Major Project of China (2019ZX08010-002) and the National Natural Science Foundation of China (32272177).
Data Availability Statement
Data are available upon request.
Acknowledgments
We thank Dr. H. Chen and Prof. J. Tu (Zhejiang University, China) for providing the genes and much technical assistance. We would also like to thank Ms. H. He (South China Agricultural University, China) for contributing to this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ahmad, M.; Ali, Q.; Hafeez, M.; Malik, A. Improvement for biotic and abiotic stress tolerance in crop plants. Biol. Clin. Sci. Res. J. 2021, 2021, e004. [Google Scholar] [CrossRef]
- Huang, F. Resistance of the fall armyworm, Spodoptera frugiperda, totransgenic Bacillus thuringiensis Cry1F corn in the Americas: lessons and implications for Bt corn IRM in China. Insect Sci. 2021, 28, 574–589. [Google Scholar] [CrossRef]
- Yang. X.; Zhao, S.; Liu, B.; Gao, Y.; Hu, C.; Li, W.; Yang, Y.; Li, G.; Wang, L.; Yang, X.; et al. Bt maize can provide non-chemical pest control and enhance food safety in China. Plant Biotechnol. J. 2023, 21, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Terán-Santofimio, H.; Rodriguez-Maciel, J.C.; DiFonzo, C. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol 2019, 112, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 373, 22. [Google Scholar]
- Rostoks, N.; Grantiņa-Ieviņa, L.; Ieviņa, B.; Evelone, V.; Valciņa, O.; Aleksejeva, I. Genetically modified seeds and plant propagating material in Europe: potential routes of entrance and current status. Heliyon 2019, 5, e01242. [Google Scholar] [CrossRef]
- Li, G.P.; Feng, H.Q.; Ji, T.J.; Huang, J.R.; Tian, C.H. What type of Bt corn is suitable for a region with diverse lepidopteran pests: A laboratory evaluation. GM Crops Food 2021, 12, 115–124. [Google Scholar] [CrossRef]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Control 2019, 130, 136–154. [Google Scholar] [CrossRef]
- Yang, F.; González, J.C.S.; Williams, J.; Cook, D.C.; Gilreath, R.T. ; Kerns, D,L, Occurrence and ear damage of Helicoverpa zea on transgenic Bacillus thuringiensis maize in the field in Texas, US and its susceptibility to Vip3A protein. Toxins 2019, 11, 102. [Google Scholar] [CrossRef]
- He, K.; Wang, Z.; Wen, L.; Bai, S.; Ma, X.; Yao, Z. Determination of baseline susceptibility to Cry1Ab protein for Asian corn borer. J. Appl. Entomol. 2005, 129, 407–412. [Google Scholar] [CrossRef]
- Perry, E.D.; Ciliberto, F.; Hennessy, D.A.; Moschini, G. Genetically engineered crops and pesticide use in U.S. maize and soybeans. Sci. Adv. 2016, 2, e1600850. [Google Scholar] [CrossRef] [PubMed]
- Alderborn, A.; Sundström, J.; Soeria-Atmadja, D.; Sandberg, M.; Andersson, H.C. Hammerling U, Genetically modified plants for non-food or non-feed purposes: Straightforward screening for their appearance in food and feed. Food Chem. Toxicol. 2010, 48, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Hallerman, E.M.; Peng, Y.F. How can China prepare for the domestic cultivation of Bt maize? Trends Food Sci. Technol. 2018, 73, 87–88. [Google Scholar] [CrossRef]
- Domingo. JL.; Bordonaba, J.G. A literature review on the safety assessment of genetically modified plants. Environ. Int. 2011, 37, 734–742. [Google Scholar] [CrossRef]
- Gilbertson. L. Cre-lox recombination: Cre-ative tools for plant biotechnology. Trends Biotechnol. 2003, 21, 550–555. [Google Scholar] [CrossRef]
- Kamatham, S.; Munagapati, S.; Manikanta, K.N.; Vulchi, R.; Chadipiralla, K.; Indla, S.H.; Allam, U.S. Recent advances in engineering crop plants for resistance to insect pests. Egypt. J. Biol. Pest Control. 2021, 31, 120. [Google Scholar] [CrossRef]
- Bai, X.Q.; Wang, Q.Y.; Chu, C.C. Excision of a selective marker in transgenic rice using a novel Cre/loxP system controlled by a floral specific promoter. Transgenic Res. 2008, 17, 1035–1043. [Google Scholar] [CrossRef]
- Chen, Y.; Rice, P.A. New insight into site-specific recombination from Flp recombinase-DNA structures. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 135–159. [Google Scholar] [CrossRef]
- Li, Z.S.; Xing, A.Q.; Moon, B.P.; Burgoyne, S.A.; Guida, A.D.; Liang, H.L.; Lee, C.; Caster, C.; Barton, J.; Klein, T.; et al. A Cre/loxP-mediated self-activating gene excision system to produce marker gene free transgenic soybean plants. Plant Mol. Biol, 2007, 65, 329–341. [Google Scholar] [CrossRef]
- Maeser, S.; Kahmann, R. The Gin recombinase of phage Mu can catalyse site-specific recombination in plant protoplasts. Mol. Gen. Genet. 1991, 230, 170–176. [Google Scholar] [CrossRef]
- Mlynarova, L.; Conner, A.J.; Nap, J.P. Directed microspore-specific recombination of transgenic alleles to prevent pollen-mediated transmission of transgenes. Plant Biotechnol. J. 2006, 4, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Siebert, R.; Puchta, H. Efficient repair of genomic double-strand breaks by homologous recombination between directly repeated sequences in the plant genome. Plant Cell 2002, 14, 1121–1131. [Google Scholar] [CrossRef]
- Pradhan, S.; Chakraborty, A.; Sikdar, N.; Chakraborty, S.; Bhattacharyya, J.; Mitra, J.; Manna, A.; Gupta, S.D.; Sen, S.K. Marker-free transgenic rice expressing the vegetative insecticidal protein (Vip) of Bacillus thuringiensis shows broad insecticidal properties. Planta 2016, 244, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N.; Verma, S.; Sahoo, R.K.; Raveendar, S.; Reddy, B.L. Recent advances in development of marker-free transgenic plants: regulation and biosafety concern. J. Biosci. 2012, 37, 167–97. [Google Scholar] [CrossRef]
- Hoa, T.T.C.; Huq, E.; Vincent, J.R.; Hodges, H.K.; Bong, B.B.; Hodges, T.K. Cre/lox site-specific recombination controls the excision of a transgene from the rice genome. Theor. Appl. Genet. 2002, 104, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Luo, J.; Zheng, P.; Zhang, X.; Zhang, C.; Li, X.; Wang, M.; Huang, Y.; Liu, X.; Jan, M.; et al. Application of Cre-lox gene switch to limit the Cry expression in rice green tissues. Sci. Rep. 2017, 7, 14505. [Google Scholar] [CrossRef]
- Boszorádová, E.; Matušíková, I.; Libantová, J.; Zimová, M.; Moravčíková, J. Cre-mediated marker gene removal for production of biosafe commercial oilseed rape. Acta Physiol. Plant 2019, 41, 73. [Google Scholar] [CrossRef]
- Luo, K.; Duan, H.; Zhao, D.; Zheng, X.; Deng, W.; Chen, Y.; Stewart Jr, C. N.; McAvoy, R.; Jiang, X.; Wu, Y.; et al. ‘GM-gene-deletor’: fused loxP-FRT recognition sequences dramatically improve the efficiency of FLP or CRE recombinase on transgene excision from pollen and seed of tobacco plants. Plant Biotechnol. J. 2007, 5, 263–274. [Google Scholar] [CrossRef]
- Moravcikova, J.; Vaculkova, E.; Bauer, M.; Libantova, J. Feasibility of the seed specific cruciferin C promoter in the self excision Cre/loxP strategy focused on generation of marker-free transgenic plants. Theor. Appl. Genet. 2008, 117, 1325–1334. [Google Scholar] [CrossRef]
- Schaffner, A.R.; Sheen, J. Maize rbcS promoter activity depends on sequence elements not found in dicot rbcS promoters. Plant Cell 1991, 3, 997–1012. [Google Scholar]
- Frame, B.R.; Shou, H.; Chikwamba, R.K.; Zhang, Z.; Xiang, C.; Fonger, T.M.; Pegg, S.E.K.; Li, B.; Nettleton, D.S.; Pei, D.; et al. Agrobacterium tumefaciens-mediated transformation of maize embryos using a standard binary vector system. Plant Physiol, 2002, 129, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Toepfer, S.; Fallet, P.; Kajuga, J.; Bazagwira, D.; Mukundwa, I.P.; Szalai, M.; Turlings, T.C. Streamlining leaf damage rating scales for the fall armyworm on maize. J. Pest Sci. 2021, 94, 1075–1089. [Google Scholar] [CrossRef]
- Xiao, Z.; Kerr, W.A. Biotechnology in China-regulation, investment, and delayed commercialization. GM Crops Food 2022, 13, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Wei, J.; Li, X.; Xu, C.; Wang, S. A rice promoter containing both novel positive and negative cis-elements for regulation of green tissuespecific gene expression in transgenic plants. Plant Biotechnol. J. 2007, 5, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh, S.; Motallebi, M.; Zamani, M.R. Efficient seed-specifically regulated autoexcision of marker gene (nptII) with inducible expression of interest gene in transgenic Nicotiana tabacum. Turk. J. Biol. 2016, 40, 1–11. [Google Scholar] [CrossRef]
- Sugita, K.; Kasahara, T.; Matsunaga, E.; Ebinuma, H. A transformation vector for the production of marker-free transgenic plants containing a single copy transgene at high frequency. Plant J. 2000, 22, 461–469. [Google Scholar] [CrossRef]
- Ye, R.; Huang, H.; Yang, Z.; Chen, T.; Lin, Y. Development of insect resistant transgenic rice with Cry1C-free endosperm. Pest Manag. Sci. 2009, 65, 1015–1020. [Google Scholar] [CrossRef]
- Koziel, M.G.; Beland, G.L.; Bowman, C.; Carozzi, N.B.; Crenshaw, R.; Crossland, L.; Dawson, J.; Desai, N.; Hill, M.; Kadwell, S.; et al. Field performance of elite transgenic maize plants expressing an insecticidal protein derived from bacillus thuringiensis. Nat. Biotechnol. 1993, 11, 194–200. [Google Scholar] [CrossRef]
- Wang, Q.; Zhu, Y.; Sun, L.; Li, L.; Jin, S.; Zhang, X. Transgenic Bt cotton driven by the green tissue-specific promoter shows strong toxicity to lepidopteran pests and lower Bt toxin accumulation in seeds. Sci. China Life Sci. 2016, 59, 172–182. [Google Scholar] [CrossRef]
- Ye, R.; Zhou, F.; Lin, Y. Two novel positive cis-regulatory elements involved in green tissue-specific promoter activity in rice (Oryza sativa L ssp.). Plant Cell Rep. 2012, 31, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Construction of gene cassettes and schematic diagram of Cre/loxP-mediated strategy. In the pKEY cassette, the Cre gene was driven by the green tissue-specific promoter Zm1rbcS. In the other pLOCK cassette, the exogenous Vip3A gene was blocked by an inserted NosT between the pairwise lox sites, driven by the strong constitutive promoter ZmUbi. The two fused cassettes were each transformed into the wild-type inbred line KN5585 to generate individual transgenic maize called KEY and LOCK plants, respectively. By crossing the KEY and LOCK plants, a site-specific excision reaction could occur in specific green tissues in their F1 progeny. The NosT should then be excised away, thereby enabling the expression of the blocked Vip3A in defined tissues.
Figure 1.
Construction of gene cassettes and schematic diagram of Cre/loxP-mediated strategy. In the pKEY cassette, the Cre gene was driven by the green tissue-specific promoter Zm1rbcS. In the other pLOCK cassette, the exogenous Vip3A gene was blocked by an inserted NosT between the pairwise lox sites, driven by the strong constitutive promoter ZmUbi. The two fused cassettes were each transformed into the wild-type inbred line KN5585 to generate individual transgenic maize called KEY and LOCK plants, respectively. By crossing the KEY and LOCK plants, a site-specific excision reaction could occur in specific green tissues in their F1 progeny. The NosT should then be excised away, thereby enabling the expression of the blocked Vip3A in defined tissues.
Figure 2.
Expression pattern analyses of Cre and Vip3A genes in various tissues of different transgenic maize by qRT-PCR. (a) Tissue-specific expression profiles of Cre in multiple tissues (root, stem, leaf, sheath, husk, silk, and kernel) at the mature stage of the KEY × LOCK hybrids and KEY plants. (b) Expression profiles of Vip3A in various tissues of the KEY × LOCK hybrids and LOCK plants. Data are given as the mean ± SE (standard error) of three biological replicates.
Figure 2.
Expression pattern analyses of Cre and Vip3A genes in various tissues of different transgenic maize by qRT-PCR. (a) Tissue-specific expression profiles of Cre in multiple tissues (root, stem, leaf, sheath, husk, silk, and kernel) at the mature stage of the KEY × LOCK hybrids and KEY plants. (b) Expression profiles of Vip3A in various tissues of the KEY × LOCK hybrids and LOCK plants. Data are given as the mean ± SE (standard error) of three biological replicates.
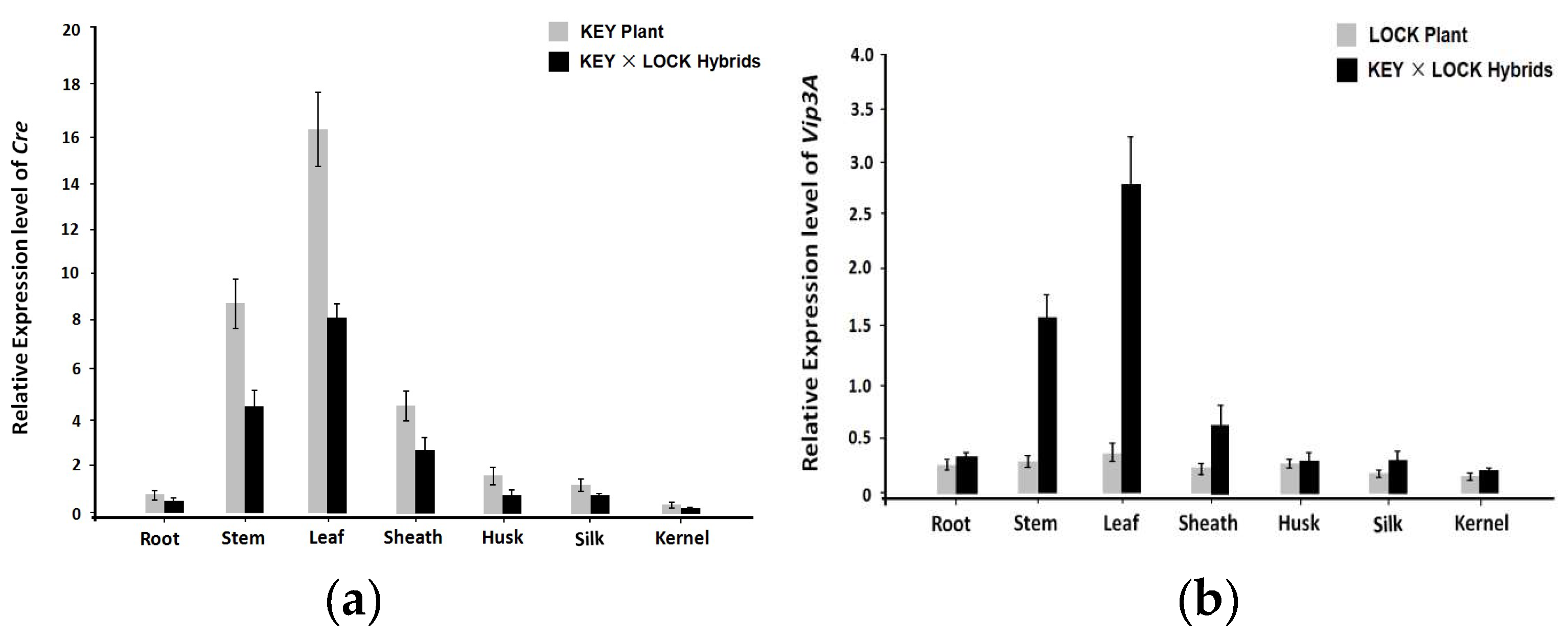
Figure 3.
Insect-resistance of different maize leaf against O. furnacalis and S. frugiperda using a diet-overlay bioassay in the laboratory. (a-c) Typical performance of different maize leaves damaged by the pest of O. furnacalis after three days of implementation. (a) Negative control KN5585. (b) Positive control LOCK. (c) Low damaged KEY × LOCK hybrids leaf. (d-f) Typical performance of different maize leaves damaged by another pest of S. frugiperda after three days of implementation. (d) KN5585. (d) LOCK. (f) Low damaged KEY × LOCK hybrids leaf.
Figure 3.
Insect-resistance of different maize leaf against O. furnacalis and S. frugiperda using a diet-overlay bioassay in the laboratory. (a-c) Typical performance of different maize leaves damaged by the pest of O. furnacalis after three days of implementation. (a) Negative control KN5585. (b) Positive control LOCK. (c) Low damaged KEY × LOCK hybrids leaf. (d-f) Typical performance of different maize leaves damaged by another pest of S. frugiperda after three days of implementation. (d) KN5585. (d) LOCK. (f) Low damaged KEY × LOCK hybrids leaf.

Figure 4.
Field performance of different maize against O. furnacalis and S. frugiperda after manual infestation. (a–c) Insect-resistant performance of different maize against the pest O. furnacalis infestation. (a) Negative control KN5585. (b) Positive control LOCK. (c) Low damaged KEY × LOCK hybrids. (d–f) Insect-resistant performance of different maize against another pest, S. frugiperda infestation. (d) KN5585. (e) LOCK. (f) Low damaged KEY × LOCK hybrids. Scale bar = 10 cm.
Figure 4.
Field performance of different maize against O. furnacalis and S. frugiperda after manual infestation. (a–c) Insect-resistant performance of different maize against the pest O. furnacalis infestation. (a) Negative control KN5585. (b) Positive control LOCK. (c) Low damaged KEY × LOCK hybrids. (d–f) Insect-resistant performance of different maize against another pest, S. frugiperda infestation. (d) KN5585. (e) LOCK. (f) Low damaged KEY × LOCK hybrids. Scale bar = 10 cm.


Table 1.
The concentration of Vip3A protein in the LOCK plants and the KEY × LOCK hybrids a.
| Plant material | Root (ng/g) |
Stem (ng/g) |
Leaf (ng/g) |
Sheath (ng/g) |
Husk (ng/g) |
Silk (ng/g) |
Kernel (ng/g) |
|---|---|---|---|---|---|---|---|
| LOCK | 0.73 ± 0.12 | 0.51 ± 0.07 | 0.43 ± 0.09 | 0.74 ± 0.13 | 0.62 ± 0.07 | 0.68 ± 0.11 | 0.59 ± 0.08 |
| KEY × LOCK | 0.68 ± 0.23 | 6.13 ± 0.41** | 9.59 ± 0.38** | 1.08 ± 0.29 | 0.91 ± 0.35 | 0.74 ± 0.15 | 0.71 ± 0.12 |
a Values are given as the mean ± SE (standard error). LOCK transgenic maize is a positive control. Multiple comparisons of the KEY × LOCK hybrids with the control maize using Student’s t-test. **Means within a column significantly differed from LOCK transgenic maize at P < 0.01.
Table 2.
The mean mortality rate of O. furnacalis and S. frugiperda larvae via the diet-overlay bioassay in the laboratory and the plant leaf damage scale caused by the two pests in the field test a.
Table 2.
The mean mortality rate of O. furnacalis and S. frugiperda larvae via the diet-overlay bioassay in the laboratory and the plant leaf damage scale caused by the two pests in the field test a.
| Plant material | Larval mortality (%) | Plant leaf damage scale | ||
|---|---|---|---|---|
| O. furnacalis | O. furnacalis | O. furnacalis | O. furnacalis | |
| KN5585 | 8.3 ± 0.9b | 8.7 ± 0.7b | 7.5 ± 0.4b | 8.1 ± 0.5b |
| LOCK | 10.6 ± 1.4b | 9.2 ± 1.2b | 7.7 ± 0.5b | 7.9 ± 0.3b |
| KEY × LOCK | 81.2 ± 3.7a | 73.6 ± 2.9a | 2.4 ± 0.1a | 1.6 ± 0.1a |
a Data are given as the mean ± SE (standard error). Wild-type KN5585 was the negative control, and individual LOCK transgenic maize were the positive control. Means within a column for each kind of maize followed by the different letters are significantly different according to the one-way ANOVA and Dunnett comparison (P < 0.05). Each implementation included three replicates. For larval mortality, the means of the KEY × LOCK hybrids were significantly higher compared with the control maize group (O. furnacalis: F2, 6 = 349.61, P < 0.001; S. frugiperda: F2, 6 = 278.69, P < 0.001). For the plant leaf damage scale, the means of the hybrids were significantly lower compared with the control maize group (O. furnacalis: F2, 6 = 117.58, P < 0.001. S. frugiperda: F2, 6 = 196.43, P < 0.001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



