Submitted:
29 August 2023
Posted:
30 August 2023
You are already at the latest version
Abstract
Clinical ketosis is a crucial metabolic disease in dairy cows that is often accompanied by severe lipolysis and inflammation in adipose tissue. Our previous study suggested that the fold change of calmodulin (CaM) was upregulated 2.401-fold in the adipose tissue of cows with clinical ketosis. Therefore, we hypothesize that CaM may regulate lipolysis and inflammatory responses in clinical ketosis cows. To verify the hypothesis, we conducted a thorough veterinary assessment of clinical symptoms and serum β-hydroxybutyrate (BHB) concentration before collecting subcutaneous adipose tissue samples from six healthy and six clinically ketotic Holstein cows at 17 ± 4 days postpartum. Commercial kits were used to test the abundance of BHB, Non-esterified fatty acid (NEFA), Liver function index (LFI), interleukin-6 (IL-6), interleukin-1β (IL-1β), and tumor necrosis factor-α (TNF-α). We found that Clinical ketosis cows had higher levels of BHB, NEFA, LFI, IL-6, IL-1β, TNF-α, and lower glucose levels compared to healthy cows. Furthermore, the abundance of CaM, toll-like receptor 4 (TLR4), inhibitor of nuclear factor κB kinase subunit β (IKK), phosphorylated nuclear factor κB/nuclear factor κB, (p-NF-κB/NF-κB), adipose triglyceride lipase (ATGL) and phosphorylated hormone-sensitive lipase/hormone-sensitive lipase (p-HSL/HSL) were increased, while that of perilipin-1 (PLIN1) was decreased in the adipose tissue of cows with clinical ketosis. To investigate the mechanistic underlying the responses, we isolated the primary bovine adipocytes from the adipose tissue of healthy cows and induced the inflammatory response mediated by TLR4/IKK/NF-κB with lipopolysaccharide (LPS), in addition to treating with CaM overexpression adenovirus and CaM small interfering RNA. In vitro, LPS upregulated the abundance of CaM, TLR4, IKK, NF-κB, ATGL, p-HSL/HSL, and CaM and downregulated PLIN1. CaM silencing downregulated the abundance of LPS-activated p-HSL/HSL, TLR4, IKK, and NF-κB and upregulated PLIN1 in bovine adipocytes, except for ATGL. By contrast, CaM overexpression upregulated the abundance of LPS-activated p-HSL/HSL, TLR4, IKK, and NF-κB and downregulated PLIN1 expression in bovine adipocytes. In summary, these data suggest that CaM promotes lipolysis in adipocytes through HSL and PINL1 while activating the TLR4/IKK/NF-κB inflammatory pathway to promote an inflammatory response. There is a positive feedback loop between CaM, lipolysis, and inflammation. Overall, inhibiting CaM may act as an adaptive mechanism to alleviate metabolic dysregulation in adipose tissue, contributing to relief from lipolysis and inflammatory response.
Keywords:
Calmodulin
; Ketosis
; Lipolysis
; Inflammation
; Adipocytes
; TLR4/IKK/NF-κB
1. Introduction
The introduction should briefly place the study in a broad context and highlight why it is important. It should define the purpose of the work and its significance. The current state of the research field should be carefully reviewed and key publications cited. Please highlight controversial and diverging hypotheses when necessary. Finally, briefly mention the main aim of the work and highlight the principal conclusions. As far as possible, please keep the introduction comprehensible to scientists outside your particular field of research. References should be numbered in order of appearance and indicated by a numeral or numerals in square brackets—e.g., [1] or [2,3], or [4,5,6]. See the end of the document for further details on references.
In 1995, the transition period in cows was defined as three weeks before and after calving. During this time, the cow's metabolic and endocrine status undergoes dramatic changes in preparation for parturient and lactation. Ketosis is a major metabolic disorder of high-yielding dairy cows during the transition period, characterized by elevated blood concentrations of β-hydroxybutyrate (BHB) and Non-esterified fatty acid (NEFA) resulting from intense lipolysis [1]. Triacylglycerol lipase (ATGL), hormone-sensitive lipase (HSL), and perilipin (PLIN1) is crucial in the lipolysis process in dairy cows. Triglycerides are sequentially metabolized to diacylglycerol and monoacylglycerol, glycerin by ATGL, HSL, and monoacylglycerol lipase, and fatty acids are generated [2,3]. PLIN1 usually covers the surface of lipid droplets to prevent excessive lipolysis. Lipolysis is initiated when comparative gene identification-58 on PLIN1 is transposed to ATGL, exposing lipid droplets [3,4]. Drastic lipolysis implies a large amount of NEFA production. The liver showed the characteristics of liver function injury, such as increased positive acute phase reactive protein, aspartate aminotransferase (AST) and alanine transaminase (ALT) levels, and decreased liver function index (LFI) [5]. On the other hand, NEFA can act on Toll-like receptor 4 (TLR4), activate nuclear factor κB (NF-κB), and promote the secretion of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), interleukin-6 (IL-6), and other inflammatory factors [6].
Calmodulin (CaM) is a highly conserved signal regulator protein that regulates cell division, development, proliferation, autophagy, and lipolysis by binding to different target proteins [7,8,9]. Furthermore, CaM has been confirmed to be associated with inflammation and lipolysis in many tissues [10,11,12]. Calmodulin kinase Ⅱ (CAMKⅡ) enhances lipolysis of fat and TNFα-induced inflammation [8]. In a study of post-traumatic osteoarthritis in mice, inhibition of CAMKⅡ attenuated chondrocyte inflammation [13]. In another study, imperatorin ameliorates mast cell-mediated allergic airway inflammation by inhibiting Mas-related G protein-coupled receptor-X2 and CaMKII/ extracellular regulated protein kinases (ERK) signaling pathways [14]. There are no reports on the relationship between CaM and the pathogenesis of ketosis in dairy cows. Our previous proteomic study found that the expression of CaM in adipose tissue of cows with ketosis was higher, and the difference was 2.401 times [15], suggesting a link between CaM, lipolysis, and the inflammatory response.
The present study assessed the association between CaM and lipolysis and inflammation in vitro and in vivo. We found that cows with ketosis exhibited enhanced lipolysis, inflammatory response, and adipose tissue CaM expression. Notably, our results demonstrated that CaM silencing and overexpression in primary adipocytes of dairy cows could inhibit and activate both lipolysis and inflammatory response. Furthermore, inflammatory cytokines produced by inflammatory response could promote CaM expression and activation of lipolysis in adipose tissue. This study provides new insights into regulating lipid metabolism and the inflammation mechanism in ketotic cows.
2. Materials and methods
2.1. Animals
This study selected 150 lactating Holstein cows with similar parity (median=3, range=2 – 4) and days in milk (median=17 d, range=12–21 d). Body condition was assessed by the same herd veterinarian by using a 5-point scale (1=emaciated to 5=obese) [16]. Based on clinical symptoms and serum BHB concentration, six clinical ketosis cows with serum BHB concentrations above 3.0 mmol/L were randomly selected as the clinical ketosis group (CK), and six cows with serum BHB concentrations below 1.20 mmol/L were randomly selected as the healthy control group (CON) [17]. The characteristics of parity, milk production, dry matter intake (DMI), BW, serum glucose, BHB, and NEFA concentrations of selected control and clinical ketosis cows are shown in Table 1. The total mixed ration of tested dairy cows complied for dairy cows. All cows were fed the same diet that complied with the NRC (2001) and Chinese feeding standards during the experiment. The diet composition of the cows during the experimental period is shown in Supplementary Table S1.
2.2. Collection of Blood
Blood samples were collected from the cows through a syringe through the coccygeal vein in the morning before feeding and then centrifuged at 3000 × g for 10 min to get the serum. The serum was recentrifuged at 12,000 × g for 10 min to collect supernatant and stored at −80℃.
2.3. Detection of serum indicators
Serum AST, ALT, glucose, NEFA, and BHB were measured using the Mindray BS-830 autoanalyzer (Mindray Laboratory Measurement, Shenzhen, China) and commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Infinite M200 PRO raster multifunctional microplate reader (Tecan Company, Switzerland) and commercial kits (XIN FAN BIOLOGY, Shenzhen, China) were used to detect IL-6, TNF-α, and IL-1β. Furthermore, the detection and calculation of LFI were conducted according to our previous study [18].
2.4. Adipose Tissue Collection and Processing
Subcutaneous adipose tissue samples were collected from the cows' tails on the same day at 17(±4) d postpartum by an experienced veterinarian using the methods previously described [19]. After collecting adipose tissue samples, they were stored in liquid nitrogen until protein extraction.
2.5. Immunohistochemistry technology
According to the method of Wang [20], fresh subcutaneous adipose tissue was cut into 1.5 cm × 1.5 cm × 0.3 cm and fixed in adipose tissue-specific fixative (Servicebio, Wuhan, China). The adipose tissue was dehydrated using alcohol at 85%, 95%, and 100%. Paraffin was dissolved in an incubator at 58°C to 60°C before embedding. After the paraffin was sufficiently cooled, the sections were made. The slices were then baked at 37°C overnight, followed by deparaffinization, and the sections were placed in xylene solution, followed by treatment with ethanol at 100%, 90%, 80%, and 70%, and finally gently rinsed with PBS. The dewaxing tissue was immersed in EDTA antigen repair buffer, heated continuously for 5 min, cooled, and then placed in PBS buffer (PH=7.4). The antigen repair was completed by shaking and washing thrice for 5 min each time. The slides were added with the working solution of the primary antibody drop (Rabbit CaM antibody, 1:1,000, Proteintech Group, Chicago, USA), incubated at 37°C for 1 h, and finally, rinsed with PBS. The slides were then added with the working solution of the secondary antibody drop (rabbit IgG, 1:10000, Proteintech Group, Chicago, USA), incubated at 37°C for 30 min, and rinsed with PBS. Subsequently, the sectioned tissues were counterstained using hematoxylin. Then dehydrated with 70%, 80%, and 90% absolute ethanol and sealed with neutral gum. Finally, the image information was observed and collected by microscope.
2.6. Isolation of bovine preadipocytes
Preadipocytes were isolated using the method published in previous studies [21]. In short, a healthy female one-day-old Holstein cow (40-50 kg) was selected and slaughtered after anesthesia. Adipose tissues of the greater omentum and mesentery of the calves were collected using aseptic procedures and washed three times with PBS containing penicillin (2,500 U/mL) and streptomycin (2,500 mg/mL) (Solarbio, Beijing, China). The collected adipose tissues were loaded in a sterile beaker. The blood vessels and fascia in the adipose tissue were removed, and the adipose tissue was cut into small uniform pieces; 0.2% type I collagenase (Sigma-Aldrich) was added at a volume ratio of 1:1 and digested in a 37℃-water bath with shaking table for 60 minutes. The digested tissue was filtered sequentially through 20-mesh and 200-mesh screens, and the filtrate was centrifuged at 1000 × g for 10 min. The supernatant was discarded, the precipitate was resuspended by adding the basal medium, centrifuged again at 1,000 × g for 10 min, and the supernatant was discarded. After resuspension with BCM (10% fetal bovine serum and 1% gentamycin) medium, the cells were inoculated in cell culture flasks and incubated at 37°C for 24 h under 5% CO2 in a cell incubator. The medium was replaced to remove the remaining tissue and non-adherent cells. Finally, the medium was changed every 48 h.
2.7. Cell culture and processing
According to the method previously published by [22]. To induce preadipocyte differentiation, we replaced the medium with basal medium containing 0.5 mM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), 1 μM dexamethasone (Sigma-Aldrich), and 1 μg/mL insulin (Sigma-Aldrich). The cells were cultured at 37℃ in 5% CO2 for 2 d and then replaced with basal medium containing 1 μg/mL insulin (Sigma-Aldrich) to further induce differentiation. After 2 d of culture, the culture was replaced with a basic culture medium for another 2 d.
After adipocyte differentiation, the medium was changed to BCM medium containing 4 μg/mL LPS (Solarbio, Beijing, China) for 3 h. In transfection experiments, the adipocytes were treated with CaM small interfering RNA (si-CaM) and CaM over-expression adenovirus (OC-CaM) for 48 h. The detailed grouping is described in the figure legends. The overexpression adenovirus vector (OC-CaM) was constructed by Hanbio (Shanghai, China). Similarly, siRNAs were designed and synthesized by Hanbio (Shanghai, China) based on bovine CaM mRNA sequences. siRNA sequences are Chain of Justice: GGUGAUGGCACCAUCACAATT, Antisense strand: UUGUGAUGGUGCCAUCACCTT.
2.8. Western Blotting
According to the manufacturer's instructions, adipose tissue and cells of the total protein were extracted using protein extraction kits (Beyotime Biotechnology, Shanghai, China). Protein concentrations were determined using the BCA Protein Assay kit (Beyotime Biotechnology, Shanghai, China). For adipose tissue, 70 mg of adipose tissue was taken, RIPA (Roche, Shanghai, China) tissue lysate was added, and adipose tissue was subsequently added to a grinder. The ground tissue was centrifuged using a centrifuge (3-16L, SIGMA, Germany) at 12,000 × g for 10 min at 4℃. Protein extraction reagent was added to the centrifuged supernatant, thoroughly mixed, and centrifuged again at 12,000 × g at 4℃ for 10 min to obtain total adipose tissue protein. For adipocytes, trypsin-EDTA (Solarbio, Beijing, China) was added to adipocytes; The adipocytes were subsequently digested in an incubator (Shanghai Boxun Industrial Co, Ltd, Shanghai, China) for 1-2 min. Centrifugation was performed using a centrifuge at 1,500 × g for 5 min. The centrifugation steps described above were repeated after the resuspension of the cells using PBS. After discarding the supernatant, the cells were resuspended in precooled lysis buffer and centrifuged at 12,000 × g for 10 min at 4°C to obtain total protein from the adipocytes. Total protein in adipose tissue and adipocytes was determined using the bicinchoninic acid (BCA) method. Typically, 30 μg of protein from each sample was separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) with known molecular weight markers, and the proteins were then transferred to a 0.45-μm PVDF membrane. The PVDFs were incubated with primary antibodies anti-β-actin (1:1,000, Proteintech Group, Chicago, USA), anti-Toll-like receptor 4 (TLR4, 1:1,000, Proteintech Group, Chicago, USA), anti-IKK (1:1,000, Proteintech Group, Chicago, USA), anti-NF-κB (1:4,000, Proteintech Group, Chicago, USA), anti-p-NF-κB (1:2,000, Cell Signaling Technology Danvers, Massachusetts, USA), anti-adipose triglyceride lipase (ATGL, 1;1,000, Proteintech Group, Chicago, USA), and anti-hormone sensitive lipase (HSL, 1:2,000, Proteintech Group, Chicago, USA). Anti-phosphorylated hormone-sensitive lipase (p-HSL, 1:1,000, ABclonal, Boston, USA), anti-perilipin-19 (PLIN1, 1:2,000, ABclonal, Boston, USA), and anti-calmodulin (CaM, 1:1,000, Proteintech Group, Chicago, USA) were incubated together and incubated overnight at 4°C. After washing the PVDF membrane with Tris-buffered saline-Tween, the membrane was incubated with the following secondary antibodies: HRP-labeled goat anti-mouse IgG (1:10000, Proteintech Group, Chicago, USA) and HRP-labeled goat anti-rabbit IgG (1:10000, Proteintech Group, Chicago, USA). The membrane was then washed using Tris-buffered saline-Tween. The immunoreactive bands were enhanced with a chemiluminescence solution (Biosharp, Beijing, China). The bands were imaged using a Protein Simple imager, and the gray-scale values were analyzed using Image-Pro Plus (Media Cybernetics).
2.9. Data analysis
IBM SPSS 23.0 (IBM Corp, Armonk, NY) software was used for statistical analysis, and data are expressed as mean ± standard error (X ± SEM). This study used an independent sample T-test to analyze the significance of differences in clinical background information, serum biochemical indicators, and adipose tissue lipolysis and inflammation-related protein abundance between healthy control cows and clinical ketosis cows. One-way analysis of variance and multiple comparisons were used to analyze the protein abundance of each group. Meanwhile, western blot results in this study were subjected to gray-scale analysis by using Image-J software (Image J 1.8.0; National Institutes of Health, Bethesda, MD, USA). Among them, P < 0.05 was considered statistically significant, and P < 0.01 was considered highly statistically significant. Graphs were drawn using GraphPad Prism software (Prism 9.2.0; GraphPad Software, San Diego, CA), * denotes P < 0.05, and ** denotes P < 0.01.
3. Results
3.1. Characteristics and blood variable of ketosis dairy cows
Table 2 shows that milk production, glucose, and DMI of CK were lower (P < 0.05), while BCS and BW of CK were higher (P < 0.05) compared with those of CON (Table 1). Additionally, CK had significantly higher (P < 0.01) serum concentrations of BHB, NEFA, IL-6, IL-1β, and TNF-α than CON. However, serum concentrations of glucose and LFI were significantly lower (P < 0.01) in CK than in CON.
3.2. The abundance of CaM, lipolysis, and inflammation-related proteins in dairy cow adipose tissue
Compared with CON, the protein abundance of ATGL and the phosphorylation of HSL was greater in adipose tissue of CK (P < 0.01, Figure 1A,B), but the abundance of PLIN1 was lower in CK compared with CON (P < 0.01, Figure 1A,B). Protein abundance of Inflammatory pathways (TLR4/IKK/NF-κB) proteins and CaM was greater in CK compared with CON (P < 0.01, Figure 1C,D).
3.3. Results of CaM immunohistochemistry in adipose tissue of dairy cows
The abundance of CaM was increased in CK adipose tissue. Compared with CON, the adipocytes in CK showed hypertrophy and irregular cell morphology (P < 0.01, Figure 1E,G).
3.4. Effect of LPS stimulation and CaM silencing on the abundance of lipolysis-related proteins
In this experiment, adding 30 nM si-RNA was the most effective in down-regulating CaM abundance. (P < 0.01, Figure 2A,B).
Compared with CON, LPS treatment up-regulated the abundance of CaM and ATGL and the phosphorylation of HSL but down-regulated the abundance of PLIN1 (P < 0.01, Figure 2C–G). Silencing of CaM down-regulated the abundance of CaM and the phosphorylation of HSL but up-regulated the abundance of PLIN1 (P < 0.01, Figure 2C,D; P < 0.05, Figure 2E,H; P<0.01, Figure 2E,G). Silencing of CaM could relieve the phosphorylation of HSL and the abundance of PLIN1 reduction by LPS when adipocytes were treated with LPS and CaM small interfering RNA simultaneously (P < 0.05, Figure 2E,H; P < 0.05, Figure 2E,G). However, silencing of CaM had no significant effect on ATGL (Figure 2E,F).
3.5. Effect of LPS stimulation and CaM silencing on the abundance of inflammation-related proteins
The abundance of TLR4 and IKK and the phosphorylation of p65 was upregulated by LPS treatment (P < 0.01, Figure 3A,B; P < 0.01, Figure 3A,C; P < 0.01; Figure 3A,D), while silencing of CaM downregulated the abundance of TLR4 and IKK and the phosphorylation of p65 (P < 0.01, Figure 3A,B; P < 0.01, Figure 3A,C; P < 0.05, Figure 3A,D) compared with CON. Silencing of CaM could relieve the abundance of TLR4 and IKK and the phosphorylation of p65 when adipocytes were treated with LPS and CaM small interfering RNA simultaneously (P < 0.01, Figure 3A,B; P < 0.01, Figure 3A,C; P < 0.05, Figure 3A,D).
3.6. Effect of LPS stimulation and CaM overexpression on the abundance of lipolysis-related proteins
In this experiment, CaM abundance was upregulated when the adenovirus reinfection index was 300 MOI (P < 0.01, Figure 4A,B), and notably, the addition of LPS modulated CaM abundance.
LPS treatment upregulated the abundance of CaM and ATGL as well as phosphorylation of HSL (P < 0.01, Figure 4C,D; P < 0.01, Figure 4E,F; P < 0.01, Figure 4E,H) and down-regulated the abundance of PLIN1 (P < 0.05, Figure 4E,G); overexpression of CaM upregulated CaM abundance, the phosphorylation of HSL, and downregulated PLIN1 abundance (P < 0.01, Figure 4C,D; P < 0.01, Figure 4E,H; P < 0.01, Figure 4E,G), compared with CON. Overexpression of CaM could enhance the phosphorylation of HSL, the abundance of ATGL increased by LPS (P < 0.05, Figure 4E,H; P < 0.05, Figure 4E,F), and overexpression of CaM promoted the abundance of PLIN1 decreased by LPS (P < 0.01, Figure 4E,G) when adipocytes were treated with LPS and adenovirus simultaneously.
3.7. Effect of LPS stimulation and CaM overexpression on the abundance of inflammation-related proteins
LPS treatment upregulated the abundance of TLR4 and IKK and the phosphorylation of p65 (P < 0.01, Figure 5A,B; P < 0.01, Figure 5A,C; P < 0.01, Figure 5A,D); overexpression of CaM upregulated TLR4 and IKK abundance and p65 phosphorylation (P < 0.01, Figure 5A,B; P < 0.05, Figure 5A,C; P < 0.01, Figure 5A,D), compared with CON. Similarly, overexpression of CaM could enhance the abundance of TLR4 and IKK and the phosphorylation of p65 (P < 0.01, Figure 5A,B; P < 0.01, Figure 5A,C; P < 0.01, Figure 5A,D) when adipocytes were treated with LPS and adenovirus simultaneously.
4. Discussion
To better understand the relationship between CaM, lipolysis, and inflammation in adipocytes during clinical ketosis in dairy cows, we constructed a regulatory network of CaM based on the results of our experiment and relevant literature on lipolysis and inflammation (Figure 6). The relationship between CaM, lipolysis, and inflammation is discussed as follows:
4.1. Relationship between CaM and lipolysis in adipocytes
CaM and its target protein CaMKⅡ play an important role in regulating lipolysis metabolism in adipocytes (Figure 6). There are reports that adipocyte lipolytic metabolism is enhanced by CaM via the cyclic adenosine monophosphate/Protein kinase A/HSL pathway [23]. Currently, there is no report on the effect of CaM on ATGL and PLIN1, but CaM can bind to its target protein CaMKⅡ to exert lipolysis. The Ca2+-CaMKⅡ-ERK-HSL pathway can enhance the expression of HSL independent of the traditional Protein kinase A pathway, thereby stimulating lipolysis in 3T3-L1 preadipocytes. Furthermore, ATGL is positively regulated by CaMKⅡ via the AMPK pathway, but the regulation of HSL by CaMKⅡ is uncertain under different circumstances [24,25]. However, the transcription and expression of ATGL remain controversial. Studies have found that ATGL mRNA transcription is up-regulated by leptin in porcine adipocytes, but its protein expression is down-regulated [24,26]. Thus, the inconsistency between ATGL protein transcription and expression suggests posttranslational regulation of the enzyme. Alternatively, ATGL activity is regulated by transcription and, more importantly, comparative gene identification-58 and G0/G1 switch gene 2 [27,28].
Moreover, lipolysis can be promoted by CaM-mediated inflammation. As shown in Figure 6, PLIN1 was downregulated by TNF-a via p42/p44 and C-Jun N-terminal kinase[29]. Alternatively, HSL phosphorylation is induced by TNF-a via the nitric oxide /cyclic guanosine monophosphate pathway [30]. The present study found no significant difference in ATGL expression between CaM silencing and overexpression conditions. However, upon LPS stimulation, the ATGL expression level was enhanced by CaM overexpression. In the presence or absence of LPS, the phosphorylation of HSL was attenuated, and the expression of PLIN1 was enhanced under CaM silencing conditions, whereas the opposite was observed when CaM was overexpressed. Moreover, both previous studies and our study consistently found that NEFA and BHB as products of lipolysis were upregulated in serum and CaM was upregulated in adipose tissue in cows with clinical ketosis, consistent with activation of the cell-tested pathway [31,32,33]. Therefore, our study suggests that in bovine adipocytes, lipolysis is promoted by CaM via enhancing p-HSL phosphorylation and attenuating PLIN1 expression, but ATGL’s lipolysis is not mediated by CaM. It is puzzling how LPS stimulation modifies the effect of CaM on ATGL, and more studies are required.
4.2. Relationship between CaM and inflammation in adipocytes
The TLR4/IKK/NF-κB pathway can be activated by CaM, and LPS can induce inflammatory responses while increasing CaM abundance (Figure 6). The TLR4/IKK/NF-κB inflammatory pathway was activated by LPS stimulation, and the secretion of its downstream inflammatory cytokines, such as TNF-α, IL-6, and IL-1β, was also up-regulated [34,35]. CaMKⅡ is one of the important target proteins of CaM [7] and is considered a key regulator of immunity and inflammation at different levels [10]. Moreover, TLR signaling also requires the interaction with the CaMKII pathway to be fully activated. In macrophages [36], T cells [37], and microglia [38], CaMKⅡ can indirectly activate NF-κB via IκB or Akt, thereby promoting the production of proinflammatory cytokines, such as TNF-α, IL-6, and type I interferon. Furthermore, CaMKII also has a central role in regulating inflammation in myocardial infarction [39]. Our assay found that cellular expression of TLR4, IKK, p-NF-κB/NF-κB, and CaM was enhanced by LPS. When CaM was silenced, the expression of TLR4, IKK, and p-NF-κB/NF-κB was attenuated, and when CaM was overexpressed, the results were the opposite. Furthermore, cows with clinical ketosis exhibited upregulation of TNF-α, IL-6, and IL-1β in their blood and increased CaM abundance in adipose tissue. These findings were consistent with the observed alterations in inflammatory pathways and CaM abundance in vitro. Therefore, our study demonstrated that CaM expression could be enhanced by LPS and TLR4/IKK/NF-κB inflammatory pathway in adipocytes was synergistically activated by CaM and LPS. Further investigation is required to elucidate the specific molecular mechanisms underlying the upregulation of CaM by LPS; the specific pathways by which CaM is upregulated by LPS require more investigation.
4.3. Relationship between inflammation and lipolysis in adipocytes
The inflammatory response within adipocytes can induce lipolysis, and excessive lipolysis may further exacerbate the inflammatory response (Figure 6). LPS-induced inflammation upregulates ATGL expression and stimulates HSL phosphorylation. LPS-induced inflammation increases ATGL and HSL expression, promoting HSL phosphorylation [34,35]. Rat ATGL can be directly activated by LPS and its core component lipid A [35]. When cells are exposed to LPS, ATGL is first activated, and subsequently, HSL is activated, so the breakdown of triglycerides by ATGL is thought to be part of the acute phase response to endotoxemia and whole-body energy expenditure is increased [22]. Furthermore, studies have shown that LPS can mediate lipolysis by activating IKK/NF-κB inflammatory pathway and cyclic adenosine monophosphate/ protein kinase A/HSL classical lipolysis pathway [40]. Inflammation can also enhance adipocyte lipolysis and reduce insulin sensitivity via multiple pathways, including TLR4/ERK1/2/HSL [35], inositol requires enzyme/HSL [41], TNF-α/nitric oxide synthase/nitric oxide/cyclic guanosine monophosphate [30], TNF-α/GTP-binding protein/inhibitory adenylate cyclase g protein [42], TNF-a/p42/p44 and c-Jun N-terminal kinase/PLIN1 [29], which subsequently induces HSL/PLIN phosphorylation [22]. Studies have demonstrated that inflammation can activate lipolysis via the c-Jun N-terminal kinase-1,4,5-Inositol triphosphate receptor-CaM/CaMKⅡ pathway and contribute to the positive feedback loop between inflammatory and metabolic signals in obesity [8]. Low levels of PLIN1, which implies stronger lipolysis, promote NF-κB pathway activation and inflammatory cytokine synthesis, such as TNF-α, IL-1β and IL-6, and increase fatty acid and triglyceride synthesis [21].
Moreover, NEFA produced by excessive lipolysis can directly act on TLR4 or IKK, further activating the NF-κB pathway and aggravating the inflammatory response [43]. Our assay found that the abundance of CaM, ATGL, and p-HSL was significantly enhanced in adipocytes after LPS stimulation, while the expression of PLIN1 was significantly attenuated, indicating enhanced lipolysis. Furthermore, adipose tissue lipolysis was enhanced in clinical ketosis cows, as characterized by the increased serum levels of NEFA and BHB. Simultaneously, TLR4/IKK/NF-κB pathway in adipose tissue was activated, and the levels of TNF-α, IL-1β, and IL-6 in serum were up-regulated. Therefore, our study demonstrated that LPS-induced inflammation could enhance lipolysis via multiple pathways on ATGL, HSL, and PLIN lipolysis-related proteins, while NEFA released from lipolysis could act on TLR4 further to activate TLR4/IKK/NF-κB pathway-mediated inflammation.
As shown in Figure 7, inflammatory cytokines released upon activation of NF-κB, such as TNF-α, could mediate lipolysis and up-regulate the abundance of CaM through ATGL, HSL, and PLIN1. NEFA and BHB released from excessive lipolysis aggravate the inflammatory response, creating a positive feedback loop of CaM, inflammation, and lipolysis signals.
5. Conclusions
We observed clinical ketosis cows to be in a state of negative energy balance, enhanced lipolysis, liver damage, and inflammation. This state is associated with increased expression of CaM, activation of the TLR4/IKK/NF-κB inflammatory pathway, and increased lipolysis of ATGL, p-HSL/HSL, and PINL1 in adipose tissue. Notably, we confirmed the positive regulation of CaM on lipolysis and inflammation in adipocytes. These findings suggest that CaM may play an important role in the pathogenesis of clinical ketosis in dairy cows. Reducing CaM abundance could help control the inflammatory response in the adipose tissue of dairy cows during the development of clinical ketosis.
Author Contributions
The experiment was designed by Cheng Xia. The experiment was carried out by Zhijie Wang, Jinshui Chang and Yu Hao. Experimental data processing was done by Yu Hao and Yuxi Song. The manuscript was completed by Jinshui Chang and Zhijie Wang, and the revision of the manuscript was completed by Cheng Xia. All authors read and approved the final version of the manuscript.
Funding
This research was supported by the Key Project of Heilongjiang Province Natural Science Foundation (ZD2021C006).
Institutional Review Board Statement
The experiment was conducted in a large intensive cattle farm with 6,000 Holstein cows located in the central region of Heilongjiang Province, China. All animal procedures were performed in accordance with the Guidelines for the Care and Use of Experimental Animals at Heilongjiang Bayi Agricultural University (Daqing, China) (DWKJXY2023064, Approval date: 1 January 2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
Special thanks to the Animal Metabolic and Toxic Disease Team of Heilongjiang Bayi Agricultural University for support of this study.
Conflicts of Interest
No conflicts of interest are declared by the authors.
References
- Loor, J.J.; Everts, R.E.; Bionaz, M.; Dann, H.M.; Morin, D.E.; Oliveira, R.; Rodriguez-Zas, S.L.; Drackley, J.K.; Lewin, H.A. Nutrition-induced ketosis alters metabolic and signaling gene networks in liver of periparturient dairy cows. Physiol Genomics 2007, 32, 105–116. [Google Scholar] [CrossRef]
- Schweiger, M.; Schreiber, R.; Haemmerle, G.; Lass, A.; Fledelius, C.; Jacobsen, P.; Tornqvist, H.; Zechner, R.; Zimmermann, R. Adipose Triglyceride Lipase and Hormone-sensitive Lipase Are the Major Enzymes in Adipose Tissue Triacylglycerol Catabolism. Journal of Biological Chemistry 2006, 281, 40236–40241. [Google Scholar] [CrossRef]
- Gruber, A.; Cornaciu, I.; Lass, A.; Schweiger, M.; Poeschl, M.; Eder, C.; Kumari, M.; Schoiswohl, G.; Wolinski, H.; Kohlwein, S.D.; et al. The N-terminal region of comparative gene identification-58 (CGI-58) is important for lipid droplet binding and activation of adipose triglyceride lipase. J Biol Chem 2010, 285, 12289–12298. [Google Scholar] [CrossRef]
- Brasaemle, D.L.; Rubin, B.; Harten, I.A.; Gruia-Gray, J.; Kimmel, A.R.; Londos, C. Perilipin A increases triacylglycerol storage by decreasing the rate of triacylglycerol hydrolysis. J Biol Chem 2000, 275, 38486–38493. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, G.; Trevisi, E. Use of the liver activity index and other metabolic variables in the assessment of metabolic health in dairy herds. Vet Clin North Am Food Anim Pract 2013, 29, 413–431. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.C.; LeBlanc, S.J.; Murphy, M.R.; Drackley, J.K. Prepartum nutritional strategy affects reproductive performance in dairy cows. J Dairy Sci 2013, 96, 5859–5871. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, M.W.; Antonio, V. The many faces of calmodulin in cell proliferation, programmed cell death, autophagy, and cancer. BBA - Molecular Cell Research 2014, 1843, 398–435. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Choubey, M.; Patel, S.; Singer, H.; Ozcan, L. Adipocyte CAMK2 deficiency improves obesity-associated glucose intolerance. Molecular metabolism 2021, 53, 101300. [Google Scholar] [CrossRef]
- Huang, S.H.; Shen, W.J.; Yeo, H.L.; Wang, S.M. Signaling pathway of magnolol-stimulated lipolysis in sterol ester-loaded 3T3-L1 preadipocyes. Journal of Cellular Biochemistry 2010, 91. [Google Scholar] [CrossRef]
- Lin, M.Y.; Zal, T.; Ch'En, I.L.; Gascoigne, N.R.J.; Hedrick, S.M. A Pivotal Role for the Multifunctional Calcium/Calmodulin-Dependent Protein Kinase II in T Cells: From Activation to Unresponsiveness. The Journal of Immunology 2005, 174, 5583–5592. [Google Scholar] [CrossRef]
- Hughes, K.; Edin, S.; Antonsson, A.; Grundstrm, T. Calmodulin-dependent kinase II mediates T cell receptor/CD3- and phorbol ester-induced activation of IkappaB kinase. The Journal of biological chemistry 2001, 276, 36008–36013. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Schaer, D.J.; Bachli, E.B.; Kurrer, M.O.; Schoedon, G. Wnt5A/CaMKII Signaling Contributes to the Inflammatory Response of Macrophages and Is a Target for the Antiinflammatory Action of Activated Protein C and Interleukin-10. Arteriosclerosis, Thrombosis, and Vascular Biology 2008, 28, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Mével, E.; Shutter, J.A.; Ding, X.; Mattingly, B.T.; Williams, J.N.; Li, Y.; Huls, A.; Kambrath, A.V.; Trippel, S.B.; Wagner, D.; et al. Systemic inhibition or global deletion of CaMKK2 protects against post-traumatic osteoarthritis. Osteoarthritis Cartilage 2022, 30, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, J.; Zhang, Y.; Zeng, Y.; Hu, S.; Bai, H.; Hou, Y.; Wang, C.; He, H.; He, L. Imperatorin ameliorates mast cell-mediated allergic airway inflammation by inhibiting MRGPRX2 and CamKII/ERK signaling pathway. Biochem Pharmacol 2021, 184, 114401. [Google Scholar] [CrossRef]
- Xu, Q.; Li, X.; Ma, L.; Loor, J.J.; Li, X. Adipose tissue proteomic analysis in ketotic or healthy Holstein cows in early lactation1. Journal of Animal Science 2019, 97. [Google Scholar] [CrossRef]
- Ferguson, J.D.; Galligan, D.T.; Thomsen, N. Principal Descriptors of Body Condition Score in Holstein Cows. Journal of Dairy Science 1994, 77, 2695–2703. [Google Scholar] [CrossRef]
- Vanholder, T.; Papen, J.; Bemers, R.; Vertenten, G.; Berge, A.C.B. Risk factors for subclinical and clinical ketosis and association with production parameters in dairy cows in the Netherlands. Journal of Dairy Science 2015, 98, 880–888. [Google Scholar] [CrossRef]
- Wang, Z.; Song, Y.; Zhang, F.; Zhao, C.; Fu, S.; Xia, C.; Bai, Y. Early warning for inactive ovaries based on liver function index, serum MDA, IL-6, FGF21 and ANGPTL8 in dairy cows. Italian Journal of Animal Science 2022, 21, 113–122. [Google Scholar] [CrossRef]
- Xu, Q.; Fan, Y.; Loor, J.J.; Liang, Y.; Sun, X.; Jia, H.; Zhao, C.; Xu, C. Adenosine 5′-monophosphate-activated protein kinase ameliorates bovine adipocyte oxidative stress by inducing antioxidant responses and autophagy. Journal of Dairy Science 2021, 104, 4516–4528. [Google Scholar] [CrossRef]
- Wang, L.; Johnson, J.A.; Chang, D.W.; Zhang, Q. Decellularized musculofascial extracellular matrix for tissue engineering. Biomaterials 2013, 34, 2641–2654. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, G.; Xu, C.; Liu, L.; Zhang, Q.; Xu, Q.; Jia, H.; Li, X.; Li, X. Perilipin 1 Mediates Lipid Metabolism Homeostasis and Inhibits Inflammatory Cytokine Synthesis in Bovine Adipocytes. Frontiers in immunology 2018, 9, 467. [Google Scholar] [CrossRef]
- Xu, Q.; Jia, H.; Ma, L.; Liu, G.; Xu, C.; Li, Y.; Li, X.; Li, X. All-trans retinoic acid inhibits lipopolysaccharide-induced inflammatory responses in bovine adipocytes via TGFβ1/Smad3 signaling pathway. BMC Veterinary Research 2019, 15. [Google Scholar] [CrossRef] [PubMed]
- Kawai, A. The role of calmodulin in hormone-stimulated lipolysis. Metabolism 1985, 34, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Roepstorff, C.; Vistisen, B.; Donsmark, M.; Nielsen, J.N.; Galbo, H.; Green, K.A.; Hardie, D.G.; Wojtaszewski, J.F.; Richter, E.A.; Kiens, B. Regulation of hormone-sensitive lipase activity and Ser563 and Ser565 phosphorylation in human skeletal muscle during exercise. J Physiol 2004, 560, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK Phosphorylates Desnutrin/ATGL and Hormone-Sensitive Lipase To Regulate Lipolysis and Fatty Acid Oxidation within Adipose Tissue. Mol Cell Biol 2016, 36, 1961–1976. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Zheng, X.L.; Liu, B.T.; Yang, G.S. Regulation of ATGL expression mediated by leptin in vitro in porcine adipocyte lipolysis. Molecular and Cellular Biochemistry 2010, 333, 121–128. [Google Scholar] [CrossRef]
- Yamaguchi, T. Crucial Role of CGI-58/α/β Hydrolase Domain-Containing Protein 5 in Lipid Metabolism. Biological & Pharmaceutical Bulletin 2010, 33, 342–345. [Google Scholar] [CrossRef]
- Yang, X.; Lu, X.; Lombès, M.; Rha, G.B.; Chi, Y.I.; Guerin, T.M.; Smart, E.J.; Liu, J. The G0/G1 Switch Gene 2 Regulates Adipose Lipolysis through Association with Adipose Triglyceride Lipase. Cell Metabolism 2010, 11, 194–205. [Google Scholar] [CrossRef]
- Rydén, M.; Arvidsson, E.; Blomqvist, L.; Perbeck, L.; Dicker, A.; Arner, P. Targets for TNF-alpha-induced lipolysis in human adipocytes. Biochem Biophys Res Commun 2004, 318, 168–175. [Google Scholar] [CrossRef]
- Lien, C.C.; Au, L.C.; Tsai, Y.L.; Ho, L.T.; Juan, C.C. Short-term regulation of tumor necrosis factor-alpha-induced lipolysis in 3T3-L1 adipocytes is mediated through the inducible nitric oxide synthase/nitric oxide-dependent pathway. Endocrinology 2009, 150, 4892–4900. [Google Scholar] [CrossRef]
- Dervishi, E.; Plastow, G.; Hoff, B.; Colazo, M. Common and specific mineral and metabolic features in dairy cows with clinical metritis, hypocalcaemia or ketosis. Res Vet Sci 2021, 135, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.K.; Jeong, J.K.; Choi, I.S.; Kang, H.G.; Hur, T.Y.; Jung, Y.H.; Kim, I.H. Relationships among ketosis, serum metabolites, body condition, and reproductive outcomes in dairy cows. Theriogenology 2015, 84, 252–260. [Google Scholar] [CrossRef]
- Kerwin, A.L.; Burhans, W.S.; Mann, S.; Nydam, D.V.; Wall, S.K.; Schoenberg, K.M.; Perfield, K.L.; Overton, T.R. Transition cow nutrition and management strategies of dairy herds in the northeastern United States: Part II-Associations of metabolic- and inflammation-related analytes with health, milk yield, and reproduction. J Dairy Sci 2022, 105, 5349–5369. [Google Scholar] [CrossRef] [PubMed]
- Chirivi, M.; Rendon, C.; Myers, M.; Prom, C.; Roy, S.; Sen, A.; Lock, A.; Contreras, G. Lipopolysaccharide induces lipolysis and insulin resistance in adipose tissue from dairy cows. Journal of dairy science 2021. [Google Scholar] [CrossRef] [PubMed]
- Zu, L.; He, J.; Jiang, H.; Xu, C.; Pu, S.; Xu, G. Bacterial Endotoxin Stimulates Adipose Lipolysis via Toll-Like Receptor 4 and Extracellular Signal-regulated Kinase Pathway. Journal of Biological Chemistry 2009, 284, 5915–5926. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yao, M.; Li, N.; Wang, C.; Zheng, Y.; Cao, X. CaMKII promotes TLR-triggered proinflammatory cytokine and type I interferon production by directly binding and activating TAK1 and IRF3 in macrophages. Blood 2008, 112, 4961–4970. [Google Scholar] [CrossRef]
- Singh, M.V.; Swaminathan, P.D.; Luczak, E.D.; Kutschke, W.; Weiss, R.M.; Anderson, M.E. MyD88 mediated inflammatory signaling leads to CaMKII oxidation, cardiac hypertrophy and death after myocardial infarction. Journal of Molecular & Cellular Cardiology 2012, 52, 1135–1144. [Google Scholar] [CrossRef]
- Jeon, S.; Kim, S.H.; Shin, S.Y.; Lee, Y.H. Clozapine reduces Toll-like receptor 4/NF-κB-mediated inflammatory responses through inhibition of calcium/calmodulin-dependent Akt activation in microglia. Prog Neuropsychopharmacol Biol Psychiatry 2018, 81, 477–487. [Google Scholar] [CrossRef]
- Rusciano, M.R.; Sommariva, E.; Douin-Echinard, V.; Ciccarelli, M.; Poggio, P.; Maione, A.S. CaMKII Activity in the Inflammatory Response of Cardiac Diseases. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Grisouard, J.; Bouillet, E.; Timper, K.; Radimerski, T.; Dembinski, K.; Frey, D.M.; Peterli, R.; Zulewski, H.; Keller, U.; Müller, B.; et al. Both inflammatory and classical lipolytic pathways are involved in lipopolysaccharide-induced lipolysis in human adipocytes. Innate Immun 2012, 18, 25–34. [Google Scholar] [CrossRef]
- Foley, K.P.; Chen, Y.; Barra, N.G.; Heal, M.; Schertzer, J.D. Inflammation promotes adipocyte lipolysis via IRE1 kinase. Journal of Biological Chemistry 2021, 100440. [Google Scholar] [CrossRef] [PubMed]
- Gasic, S.; Tian, B.; Green, A. Tumor necrosis factor alpha stimulates lipolysis in adipocytes by decreasing Gi protein concentrations. J Biol Chem 1999, 274, 6770–6775. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, X.; Ji, Y.; Lan, T.; Yan, W.; Xu, Y.; Gong, G. Anti-inflammatory Effect of a Limonin Derivative In Vivo and Its Mechanisms in RAW264.7 Cells. Inflammation 2023, 46, 190–201. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Abundance of ATGL, HSL, p-HSL, PLIN1, TLR4, IKK, p-NF-κB, NF-κB, and CaM in adipose tissue of healthy (n=6) and clinical ketosis cows (n=6). CON=Control group, CK=Clinical ketosis group, ATGL=Adipose triglyceride lipase, HSL=Hormone sensitive lipase, p-HSL=Phosphorylated hormone sensitive lipase, PLIN1=Perilipin-1, TLR4=Toll-like receptor 4, IKK=Inhibitor of nuclear factor κB kinase subunit β, p-NF-κB=Phosphorylated nuclear factor κB, NF-κB=Nuclear factor κB, CaM=Calmodulin. (A) representative western blots of ATGL, HSL, p-HSL, and PLIN1; (B) Relative protein abundance of ATGL, p-HSL/HSL, and PLIN1; (C) Representative western blots of TLR4, IKK, p-NF-κB, NF-κB, and CaM; (D) Relative protein abundance of TLR4, IKK, p-NF-κB/NF-κB (p-p65/p65), and CaM. (E) Picture under the microscope of adipose tissue from healthy dairy cows (200×); (F) Picture under the microscope of adipose tissue from clinical ketosis (200×); (G) Quantitative analysis of immunohistochemistry of CaM from adipose tissue of healthy and ketosis cows. Data are presented as mean ± SEM; *P < 0.05 and **P < 0.01.
Figure 1.
Abundance of ATGL, HSL, p-HSL, PLIN1, TLR4, IKK, p-NF-κB, NF-κB, and CaM in adipose tissue of healthy (n=6) and clinical ketosis cows (n=6). CON=Control group, CK=Clinical ketosis group, ATGL=Adipose triglyceride lipase, HSL=Hormone sensitive lipase, p-HSL=Phosphorylated hormone sensitive lipase, PLIN1=Perilipin-1, TLR4=Toll-like receptor 4, IKK=Inhibitor of nuclear factor κB kinase subunit β, p-NF-κB=Phosphorylated nuclear factor κB, NF-κB=Nuclear factor κB, CaM=Calmodulin. (A) representative western blots of ATGL, HSL, p-HSL, and PLIN1; (B) Relative protein abundance of ATGL, p-HSL/HSL, and PLIN1; (C) Representative western blots of TLR4, IKK, p-NF-κB, NF-κB, and CaM; (D) Relative protein abundance of TLR4, IKK, p-NF-κB/NF-κB (p-p65/p65), and CaM. (E) Picture under the microscope of adipose tissue from healthy dairy cows (200×); (F) Picture under the microscope of adipose tissue from clinical ketosis (200×); (G) Quantitative analysis of immunohistochemistry of CaM from adipose tissue of healthy and ketosis cows. Data are presented as mean ± SEM; *P < 0.05 and **P < 0.01.
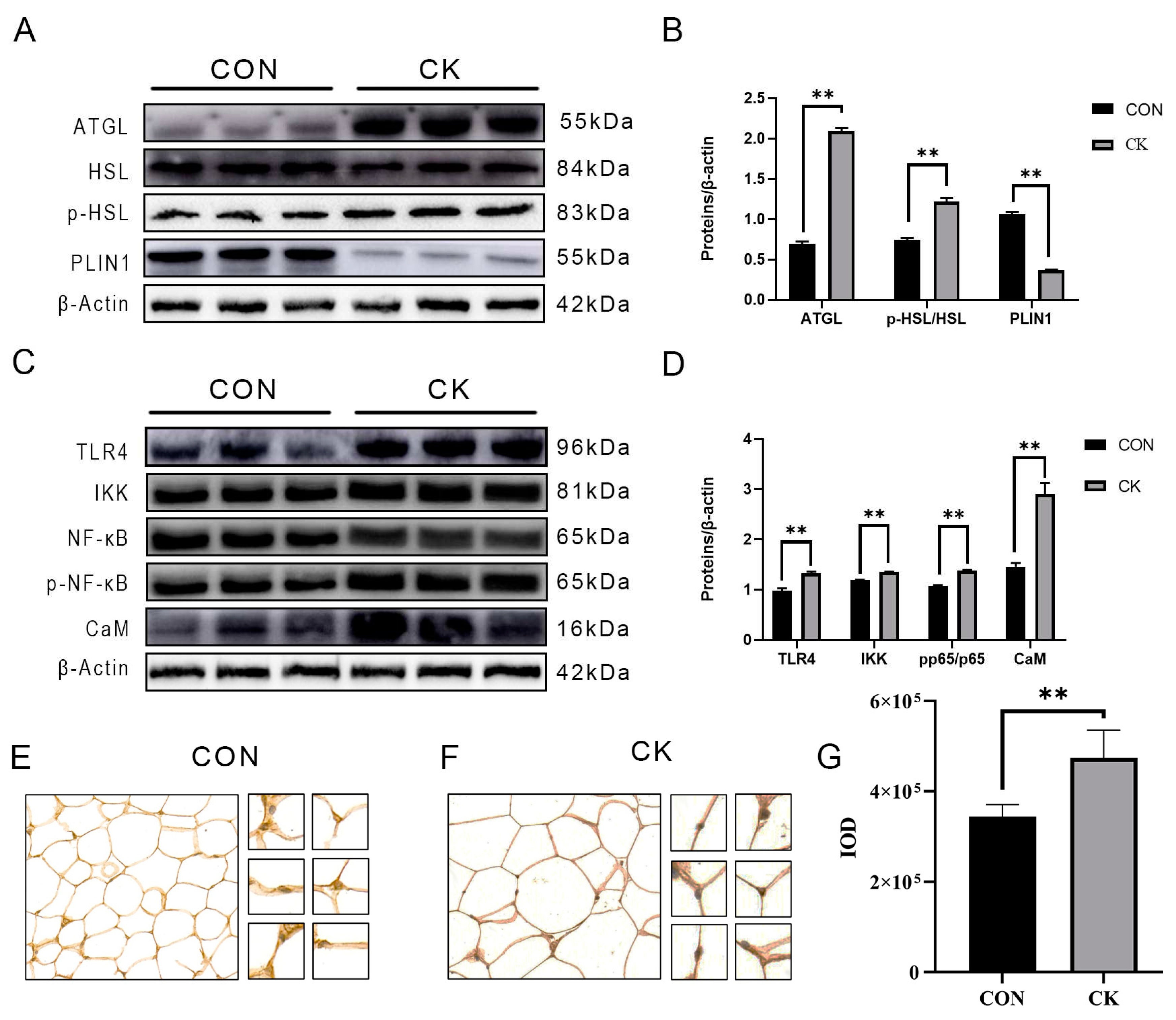
Figure 2.
Effect of different siRNA concentrations on CaM abundance in adipocytes. Adipocytes were transfected with CaM small interfering RNA (SC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, SC=Calmodulin silencing, SC + LPS=Calmodulin silenced and lipopolysaccharide treatment, CaM=Calmodulin, ATGL=Adipose triglyceride lipase, HSL=Hormone sensitive lipase, p-HSL=Phosphorylated hormone sensitive lipase, PLIN1=Perilipin-1. (A) Representative western blots of CaM silenced by different si-RNA concentrations; (B) Relative protein abundance of CaM after silencing CaM with different concentrations of si-RNA; (C) Representative western blots of CaM abundance in the case of silenced CaM and LPS stimulation: (D) Relative protein abundance of CaM; (E) Representative western blots of ATGL, HSL, p-HSL, and PLIN1; (F) Relative protein abundance of ATGL; (G) Relative protein abundance of PLIN1; (H) Relative protein abundance of p-HSL/HSL. Data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
Figure 2.
Effect of different siRNA concentrations on CaM abundance in adipocytes. Adipocytes were transfected with CaM small interfering RNA (SC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, SC=Calmodulin silencing, SC + LPS=Calmodulin silenced and lipopolysaccharide treatment, CaM=Calmodulin, ATGL=Adipose triglyceride lipase, HSL=Hormone sensitive lipase, p-HSL=Phosphorylated hormone sensitive lipase, PLIN1=Perilipin-1. (A) Representative western blots of CaM silenced by different si-RNA concentrations; (B) Relative protein abundance of CaM after silencing CaM with different concentrations of si-RNA; (C) Representative western blots of CaM abundance in the case of silenced CaM and LPS stimulation: (D) Relative protein abundance of CaM; (E) Representative western blots of ATGL, HSL, p-HSL, and PLIN1; (F) Relative protein abundance of ATGL; (G) Relative protein abundance of PLIN1; (H) Relative protein abundance of p-HSL/HSL. Data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
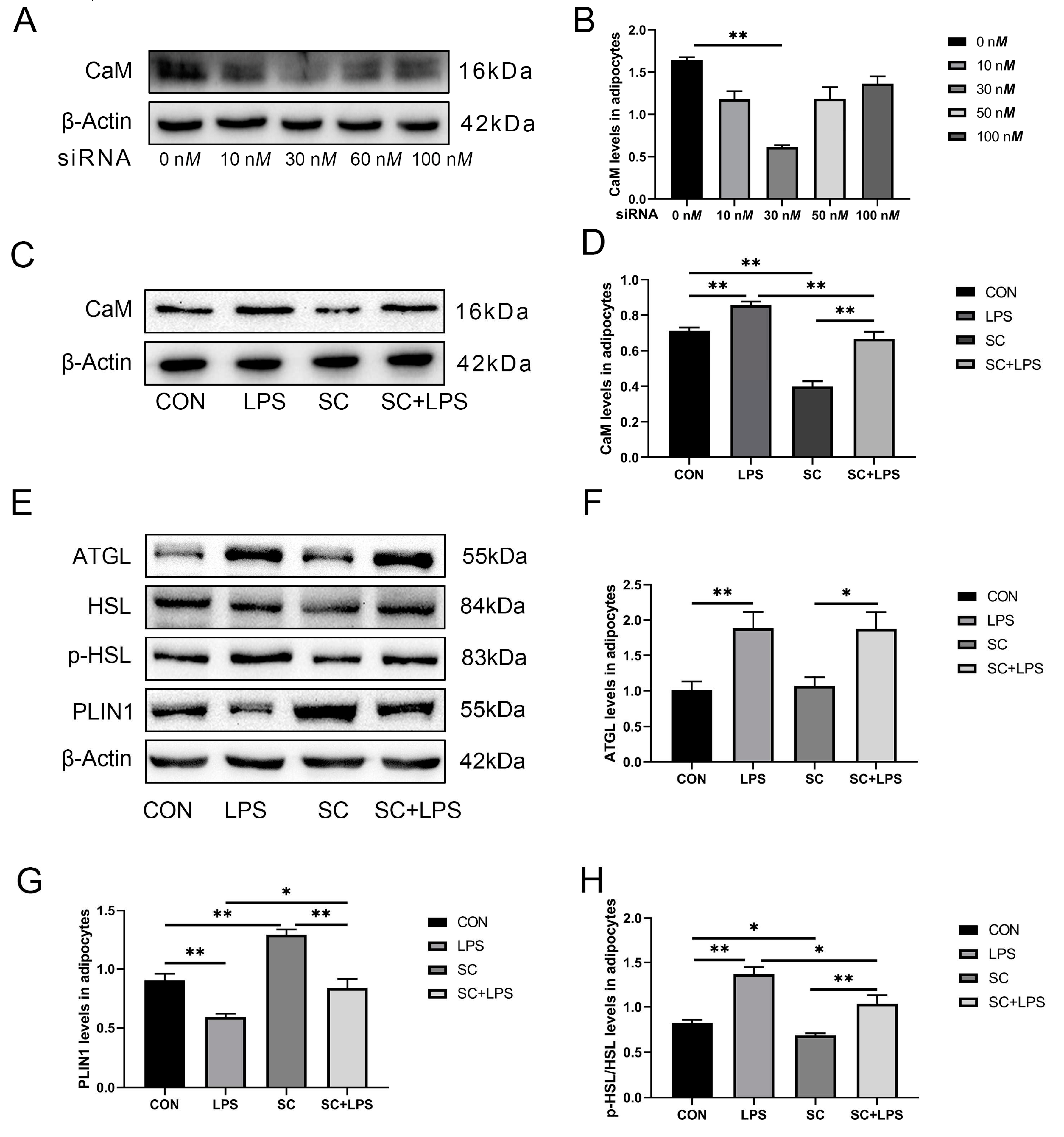
Figure 3.
CaM silencing reduced LPS-induced inflammation-related proteins. Adipocytes were transfected with CaM small interfering RNA (SC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, SC=Calmodulin silencing, SC + LPS=Calmodulin silenced and lipopolysaccharide treatment, TLR4=Toll-like receptor 4, IKK=Inhibitor of nuclear factor κB kinase subunit β, p-NF-κB=Phosphorylated nuclear factor κB, NF-κB=Nuclear factor κB. (A) Representative western blots of TLR4, IKK, p-NF-κB, and NF-κB; (B) Relative protein abundance of TLR4, IKK, p-NF-κB, and NF-κB; (C) Relative protein abundance of IKK; (D) Relative protein abundance of p-NF-κB/NF-κB (p-p65/p65). The data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
Figure 3.
CaM silencing reduced LPS-induced inflammation-related proteins. Adipocytes were transfected with CaM small interfering RNA (SC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, SC=Calmodulin silencing, SC + LPS=Calmodulin silenced and lipopolysaccharide treatment, TLR4=Toll-like receptor 4, IKK=Inhibitor of nuclear factor κB kinase subunit β, p-NF-κB=Phosphorylated nuclear factor κB, NF-κB=Nuclear factor κB. (A) Representative western blots of TLR4, IKK, p-NF-κB, and NF-κB; (B) Relative protein abundance of TLR4, IKK, p-NF-κB, and NF-κB; (C) Relative protein abundance of IKK; (D) Relative protein abundance of p-NF-κB/NF-κB (p-p65/p65). The data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
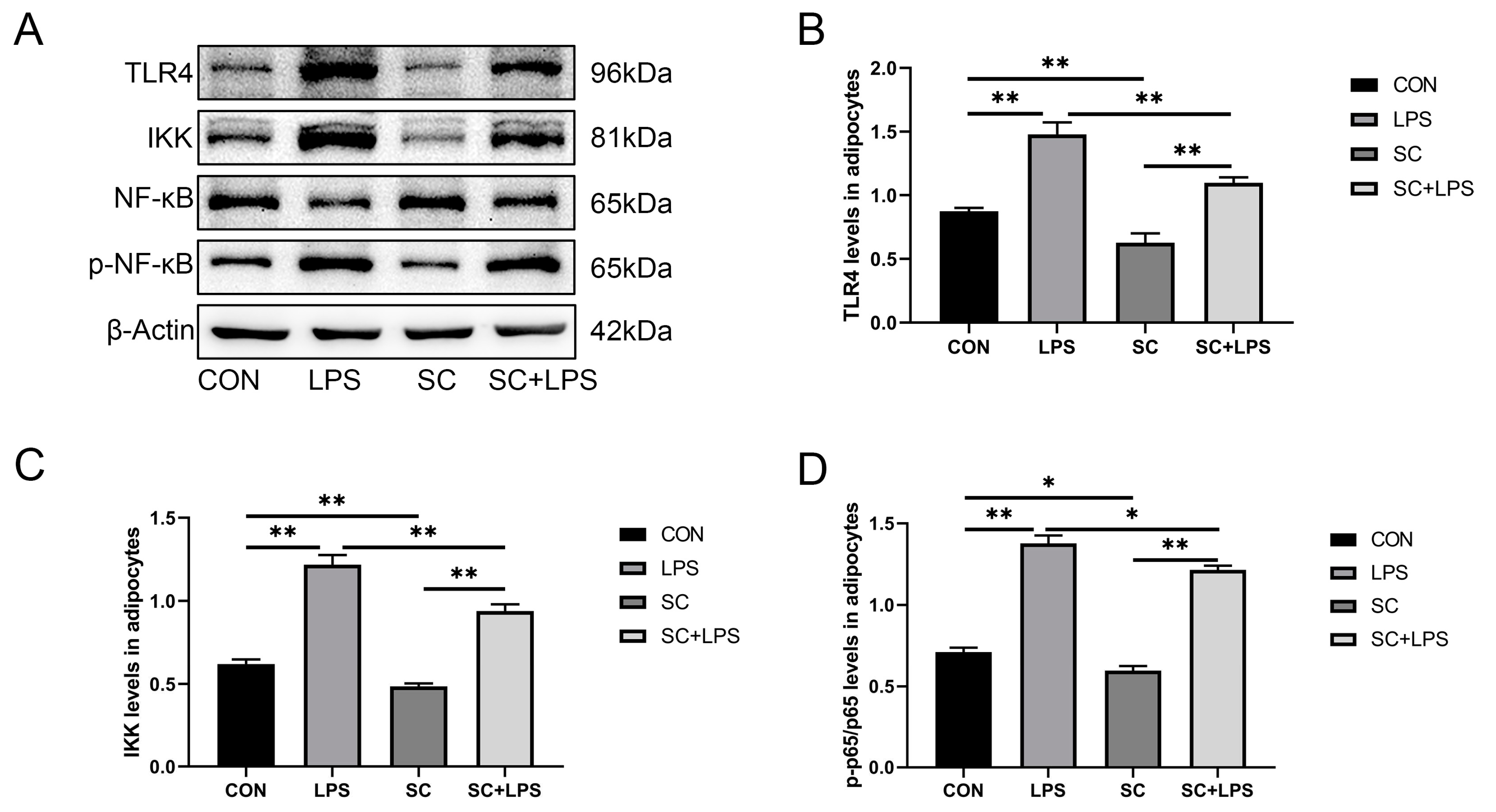
Figure 4.
Effect of adenoviruses with different infection indices on CaM abundance. Adipocytes were transfected with OC-CaM (OC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, OC=Calmodulin overexpression, OC + LPS=Calmodulin overexpression and lipopolysaccharide treatment, CaM=Calmodulin, ATGL=Adipose triglyceride lipase, HSL=Hormone sensitive lipase, p-HSL=Phosphorylated hormone sensitive lipase, PLIN1=Perilipin-1. (A) Representative western blots of CaM silenced by different adenovirus concentrations; (B) After overexpression of CaM by adenovirus at various concentrations relative protein abundance of CaM; (C) Representative western blots of CaM abundance under CaM overexpression and LPS stimulation conditions; (D) Relative protein abundance of CaM; (E) Representative western blots of ATGL, HSL, p-HSL, and PLIN1; (F) Relative protein abundance of ATGL; (G) Relative protein abundance of PLIN1; (H) Relative protein abundance of p-HSL/HSL. (E) Representative western blots of TLR4, IKK, p-NF-κB, and NF-κB; (F) Relative protein abundance of TLR4, IKK, p-NF-κB/NF-κB (p-p65/p65); (G) Relative protein abundance of IKK; (H) Relative protein abundance of p-NF-κB/NF-κB (p-p65/p65). Data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
Figure 4.
Effect of adenoviruses with different infection indices on CaM abundance. Adipocytes were transfected with OC-CaM (OC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, OC=Calmodulin overexpression, OC + LPS=Calmodulin overexpression and lipopolysaccharide treatment, CaM=Calmodulin, ATGL=Adipose triglyceride lipase, HSL=Hormone sensitive lipase, p-HSL=Phosphorylated hormone sensitive lipase, PLIN1=Perilipin-1. (A) Representative western blots of CaM silenced by different adenovirus concentrations; (B) After overexpression of CaM by adenovirus at various concentrations relative protein abundance of CaM; (C) Representative western blots of CaM abundance under CaM overexpression and LPS stimulation conditions; (D) Relative protein abundance of CaM; (E) Representative western blots of ATGL, HSL, p-HSL, and PLIN1; (F) Relative protein abundance of ATGL; (G) Relative protein abundance of PLIN1; (H) Relative protein abundance of p-HSL/HSL. (E) Representative western blots of TLR4, IKK, p-NF-κB, and NF-κB; (F) Relative protein abundance of TLR4, IKK, p-NF-κB/NF-κB (p-p65/p65); (G) Relative protein abundance of IKK; (H) Relative protein abundance of p-NF-κB/NF-κB (p-p65/p65). Data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
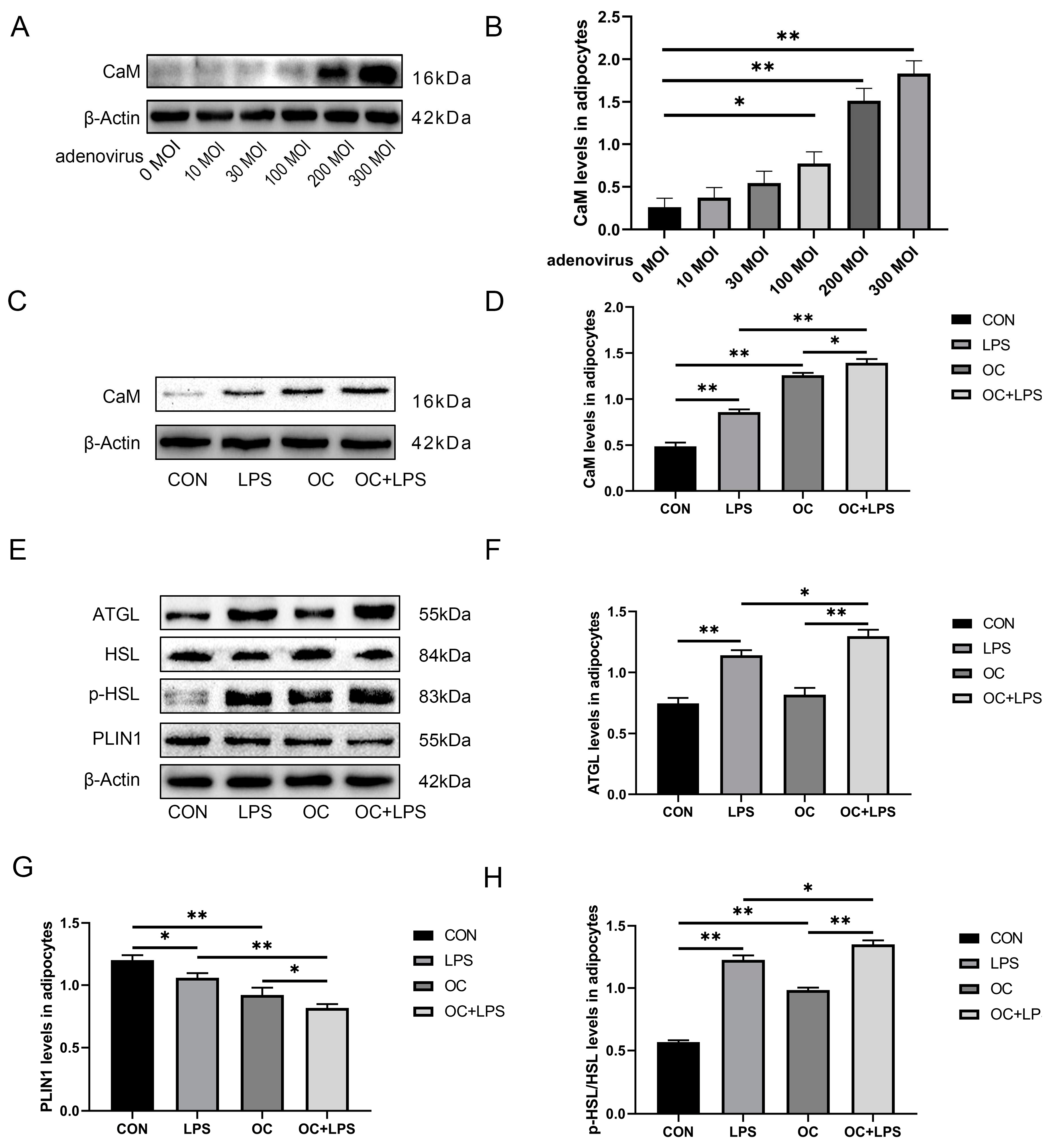
Figure 5.
CaM overexpression upregulated LPS-induced Abundance of inflammation-related proteins. Adipocytes were transfected with OC-CaM (OC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, OC=Calmodulin overexpression, OC + LPS=Calmodulin overexpression and lipopolysaccharide treatment, TLR4=Toll-like receptor 4, IKK=Inhibitor of nuclear factor κB kinase subunit β, p-NF-κB=Phosphorylated nuclear factor κB, NF-κB=Nuclear factor κB. (A) Representative western blots of TLR4, IKK, p-NF-κB and NF-κB; (B) Relative protein abundance of TLR4, IKK, p-NF-κB/NF-κB (p-p65/p65); (C) Relative protein abundance of IKK; (D) Relative protein abundance of p-NF-κB/NF-κB (p-p65/p65). Data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
Figure 5.
CaM overexpression upregulated LPS-induced Abundance of inflammation-related proteins. Adipocytes were transfected with OC-CaM (OC) for 48 h and cultured for 3 h in the presence or absence of LPS (4 μg/mL). CON=Control, LPS=Lipopolysaccharide treatment, OC=Calmodulin overexpression, OC + LPS=Calmodulin overexpression and lipopolysaccharide treatment, TLR4=Toll-like receptor 4, IKK=Inhibitor of nuclear factor κB kinase subunit β, p-NF-κB=Phosphorylated nuclear factor κB, NF-κB=Nuclear factor κB. (A) Representative western blots of TLR4, IKK, p-NF-κB and NF-κB; (B) Relative protein abundance of TLR4, IKK, p-NF-κB/NF-κB (p-p65/p65); (C) Relative protein abundance of IKK; (D) Relative protein abundance of p-NF-κB/NF-κB (p-p65/p65). Data are presented as mean ± SEM; *P < 0.05, **P < 0.01.
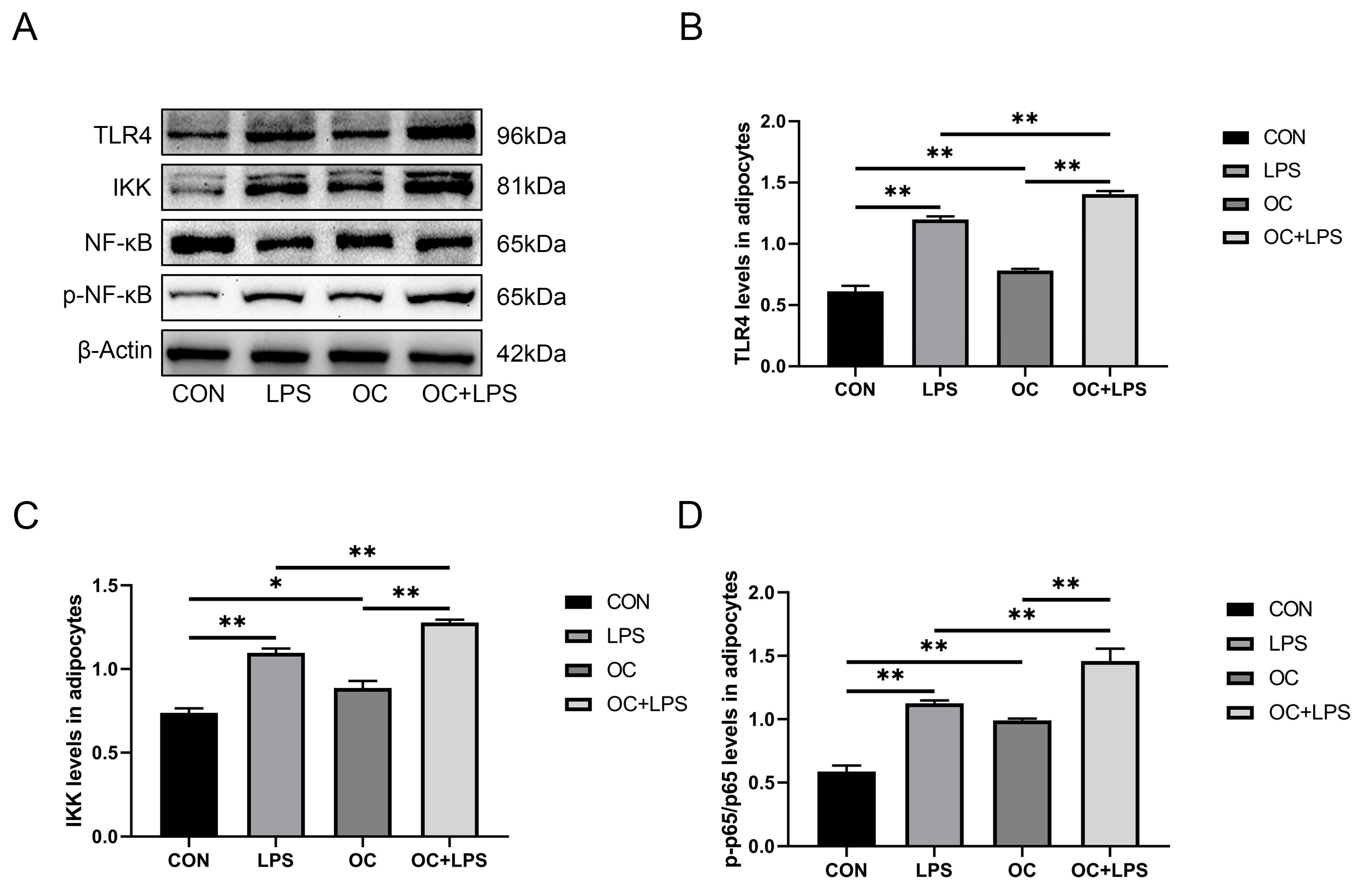
Figure 6.
.
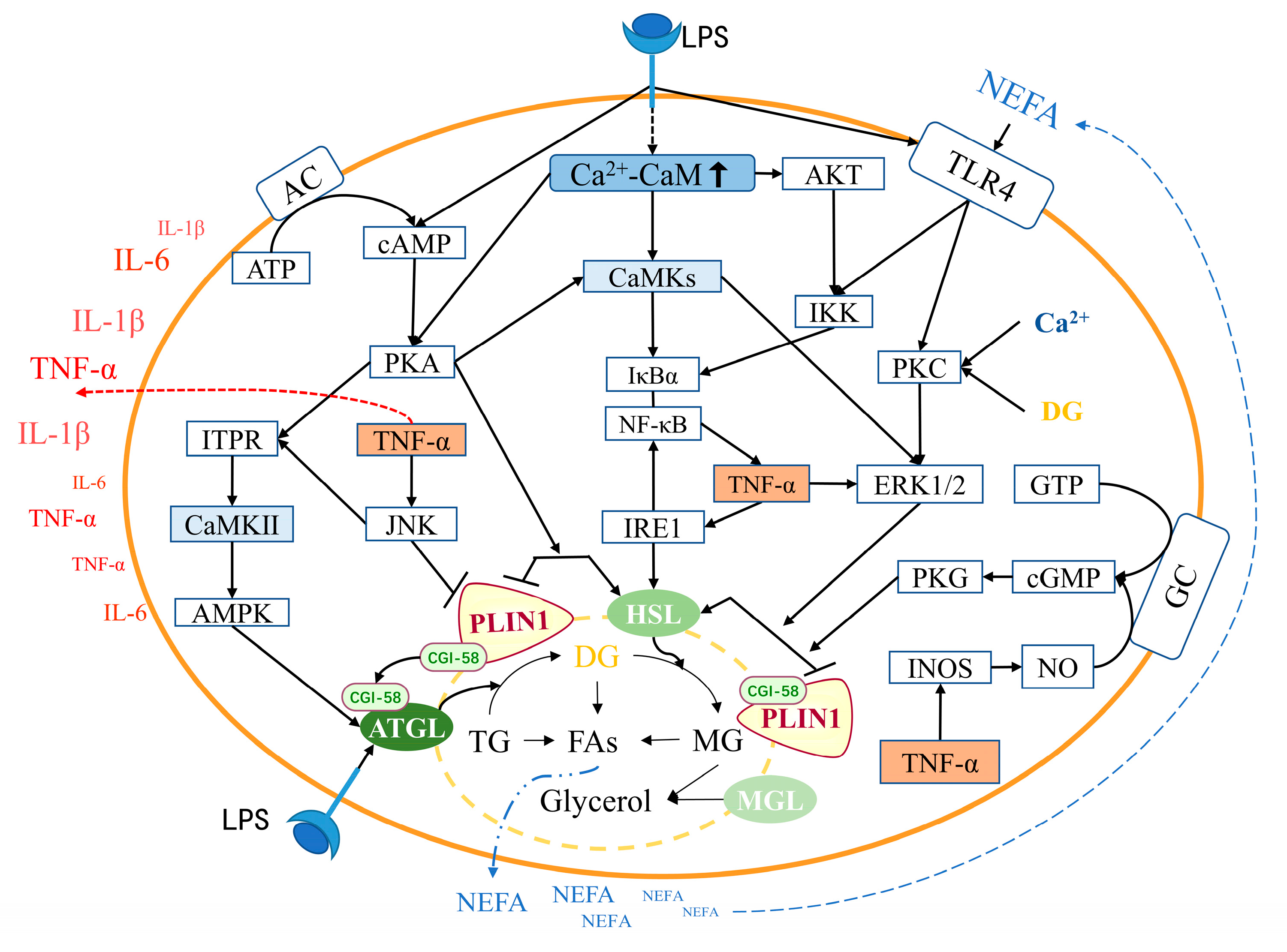
Figure 7.
.

Table 1.
The basal diet formulation %.
| Item (%) | Postpartum |
|---|---|
| Corn silage | 40.00 |
| Corn | 35.00 |
| Wheat bran | 8.00 |
| Soybean meal | 5.00 |
| Sunflower | 8.00 |
| NaCl | 1.00 |
| Premix* | 1.80 |
| NaHCO3 | 1.20 |
| Total | 100.00 |
| Nutrient composition (% of DM) | |
| NEL (MJ/Kg) | 6.70 |
| CP | 15.20 |
| NDF | 33.45 |
| ADF | 17.20 |
| NFC | 40.40 |
| Ca | 0.70 |
| P | 0.50 |
*The premix provided the following per kg of diets: VA 200,000 IU, VD 70,000 IU, VE 1,000 IU, Fe 2,000 mg, Cu 600 mg, Zn 2,400 mg, Mn 1,300 mg, I 6 mg, Co 7 mg. DM, Dry Matter; NEL, Net energy for lactation; CP, Crude protein; NDF, Neutral detergent fibre; ADF, Acid detergent fibre; NFC, Non-fibre carbohydrate.
Table 2.
Characteristics and blood variable of selected dairy cows1.
| Item | CON (n=6) | CK (n=6) | P-value |
|---|---|---|---|
| Parity | 3 | 3 | |
| Milk production (kg/d) | 38.34; 0.27 | 27.50; 0.21 | < 0.001 |
| DMI (kg/d) | 21.32; 0.47 | 19.75; 0.40 | 0.028 |
| BW (kg) | 613.29; 6.24 | 643.70; 7.18 | 0.010 |
| BCS | 2.67; 0.17 | 3.32; 0.22 | 0.041 |
| Glucose (mmol/L) | 4.03; 0.07 | 2.23; 0.03 | < 0.001 |
| BHB (mmol/L) | 0.38; 0.05 | 3.29; 0.11 | < 0.001 |
| NEFA (mmol/L) | 0.32; 0.06 | 1.18; 0.11 | < 0.001 |
| AST(U/L) | 72.00; 1.59 | 156.40; 3.52 | < 0.001 |
| ALT(U/L) | 18.80; 1.18 | 35.53; 1.87 | < 0.001 |
| LFI | 1.08; 0.86 | -7.53; 1.24 | < 0.001 |
| IL-6(ng/L) | 0.46; 0.02 | 1.26; 0.05 | < 0.001 |
| IL-1β(ng/L) | 1.50; 0.31 | 6.08; 0.59 | < 0.001 |
| TNF-α(ng/L) | 69.24; 3.95 | 102.49; 4.29 | < 0.001 |
1Data were analyzed using the T-test. Data are presented as median and interquartile range. CON=Control group, CK=Clinical ketosis group, BW=Body weight, DMI=Dry matter intake, BCS=Body condition score, AST=Aspartate aminotransferase, ALT=Alanine transaminase, LFI=liver function index. IL-6=Interleukin-6, IL-1β=Interleukin-1β, TNF-α=Tumor necrosis factor-α. Data are presented as mean ± SEM; *P < 0.05 and **P < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



