
Submitted:
29 August 2023
Posted:
30 August 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The pathogenesis of coronavirus disease 2019 (COVID-19) is associated with a hyperinflammatory response. The mechanisms of SARS-CoV-2-induced inflammation are scantly known. Methylglyoxal (MG) is a glycolysis-derived by-product, endowed with a potent glycating action, leading to the formation of advanced glycation end products (AGEs), the major of which is MG-H1. MG-H1 exerts strong pro-inflammatory effects, frequently mediated by the receptor for AGEs (RAGE). Here, we investigated the involvement of MG-H1/RAGE axis as a potential novel mechanism in SARS-CoV-2-induced inflammation by resorting to human bronchial (BEAS-2B) and alveolar (A549) epithelial cells, expressing different levels of ACE2 receptor (R), exposed to SARS-CoV-2 spike protein 1 (S1). Interestingly, we found in BEAS-2B cells that do not express ACE2-R, that S1 exerted a pro-inflammatory action through a novel MG-H1/RAGE-based pathway. RAGE expression levels in nasopharyngeal swabs from SARS-Cov-2 positive and negative individuals, as well as glyoxalase 1 expression, the major scavenging enzyme of MG, seem to support the results obtained in vitro. Altogether, our findings reveal a novel mechanism involved in the inflammation triggered by S1, paving the way to the study of MG-H1/RAGE inflammatory axis in SARS-CoV-2 infection as a potential therapeutic target to mitigate COVID-19 associated pathogenic inflammation.
Keywords:
COVID-19
; SARS-CoV-2 spike protein 1 (S1)
; methylglyoxal
; MG-H1
; Glyoxalase 1
; RAGE
; inflammation
; Nrf2
; BEAS-2B
; A549
1. Introduction
COVID-19, short for "Coronavirus Disease 2019," is a highly contagious illness caused by the novel coronavirus SARS-CoV-2.
SARS-CoV-2 infection can lead to a range of clinical manifestations, from mild respiratory symptoms to severe respiratory distress and multi-organ failure, these latter frequently associated with an excessive and dysregulated immune response, leading to a state of hyperinflammation. This extreme release of pro-inflammatory cytokines, especially IL-1β, IL-6, IL-8, and TNF-α [1], in the context of COVID-19, is often associated with a phenomenon called the "cytokine storm”, representing a distinctive sign of the immunopathogenesis of SARS-CoV-2, in addition to being directly related to the severity and mortality of the disease. Even more critical is the fact that some COVID-19 survivors experience the so-called post-acute sequelae of COVID-19 (PASCA), also known as "long COVID" or "long-haul COVID," a range of persistent symptoms and health issues that continue for weeks or months after the acute phase of a COVID-19 infection has resolved. Even individuals who initially experienced mild cases of COVID-19 can develop these long-lasting symptoms [1]. It is important to note that researchers are still studying the exact mechanisms behind these long-lasting symptoms and how they relate to the initial COVID-19 infection. Some theories suggest that the virus might trigger an inflammatory response that continues even after the virus is no longer present in significant amounts in the body.
In consideration of the absence of any definitive therapeutic treatment for COVID-19, the identification of novel (and potentially selectable as targets) inflammatory pathways that can mitigate the aggressive and uncontrolled inflammatory response in COVID-19 patients, could be crucial to develop effective therapeutic strategies against SARS-CoV-2 infection [2]. Besides, currently, the exact mechanisms of SARS-CoV-2-induced inflammation are scantly known.
Methylglyoxal (MG) is an extremely reactive metabolite generated as a byproduct of various cellular processes, mainly glucose metabolism [3]. It is a potent inducer of a process called glycation, occurring when MG reacts with biological macromolecules (proteins, nucleic acids, lipids) in a non-enzymatic manner, leading to the formation of advanced glycation end products (AGEs). AGEs are stable structures that can accumulate in various tissues, a condition known as glycative stress, causing cellular damage and contributing to the development and progression of several diseases [4-6]. MG-H1 (methylglyoxal-derived hydroimidazolone 1), one of the specific products of protein glycation by MG [3], can activate inflammatory pathways, usually by binding to the receptor RAGE [4,7,8], which is mostly expressed in the lungs, and promote the release of pro-inflammatory molecules [4,8].
To counteract pro-inflammatory MG-H1-derived glycative stress, the body has a major defense mechanism, the GSH-dependent glyoxalase 1 (Glo1), that converts MG, the precursor of MG-H1, into the non-toxic D-lactate [9].
We herein investigated the involvement of MG-H1/RAGE axis in SARS-CoV-2-induced pathogenic inflammation, by exposing human bronchial (BEAS-2B) and alveolar (A549) epithelial cells to the SARS-CoV-2 spike 1 (S1) protein [1]. We found that S1 triggered inflammation in both cells. Interestingly, we found, specifically in BEAS-2B cells, that do not express ACE2-R, compared to A549 cells, that S1 exerted a pro-inflammatory action through a novel MG-H1/RAGE-based pathway. Moreover, RAGE and Glo1 expression levels in nasopharyngeal swabs from SARS-Cov-2 positive and negative subjects seems to support the results obtained in vitro. Altogether, our findings reveal a novel mechanism involved in the inflammation triggered by S1, paving the way to the study of MG-H1/RAGE inflammatory axis in SARS-CoV-2 infection as a potential therapeutic target to mitigate COVID-19 associated pathogenic inflammation.
2. Results
2.1. Effect of SARS-CoV-2 S1 spike protein on human bronchial BEAS-2B and alveolar A549 Cell Viability
Before studying the pro-inflammatory effect of SARS-CoV-2 S1 spike protein on BEAS-2B and A549 cells, viability was evaluated by MTT assay 6 and 24 h post-exposure at 25 and 100 ng/ml. As shown in Figure 1, S1 did not affect BEAS-2B and A549 viability, compared to untreated control cells. Morphological analysis, by phase-contrast imaging, confirmed the results obtained from cell viability (Figure S1). Altogether, these findings showed that at the considered doses and time course, S1 was not cytotoxic towards both epithelial cells.
2.2. SARS-CoV-2 S1 spike protein induces inflammatory cytokines in human bronchial BEAS-2B and alveolar A549 cells
To determine the pro-inflammatory effects of S1 spike protein on BEAS-2B and A549 cells, we measured the levels of IL-1β, TNF-α, IL-6 and IL-8, crucial inflammatory cytokines in COVID-19 [1]. We found that both 25 ng/ml (Figure 2) and 100 ng/ml (Figure S2) S1 protein induced a significant increase in all the considered cytokines compared with unexposed cells, at 6 and 24 h post-exposure. Since an inflammatory status was induced in both cell models with 25 ng/ml S1 and it was more evident at 24 h post-S1 treatment, we decided to perform subsequent experiments by exposing cells to these experimental conditions.
2.3. SARS-CoV-2 S1 spike protein affects MG-H1 levels and RAGE expression in human bronchial BEAS-2B cells
MG-H1, a major AGE derived from the spontaneous reaction of MG with the arginine residues of proteins [3], is a potent inducer of inflammatory pathways, usually through its binding to the receptor RAGE [4,7,8]. In the attempt to evaluate the potential involvement of both MG-H1 and RAGE in S1-induced inflammation, we detected MG-H1 levels and RAGE expression in BEAS-2B and A549 exposed to 25 ng/ml S1 for 24 h. We found that S1 induced a significant increase in MG-H1 (Figure 3a) and RAGE (Figure 3b) levels in BEAS-2B cells, while it induced a modest decrease in MG-H1 levels (Figure 3c), without affecting RAGE expression (Figure 3d), in A549 cells. These results suggested that MG-H1/RAGE axis could represent a novel S1-induced pro-inflammatory pathway in human bronchial BEAS-2B cells.
2.4. SARS-CoV-2 S1 spike protein induces inflammation in human bronchial BEAS-2B cells through MG-H1/RAGE axis
To demonstrate that MG-H1/RAGE axis was involved in S1-induced inflammation in human bronchial BEAS-2B cells, we pre-treated them with 10 mM aminoguanidine (AG), that specifically reacting with MG, prevents MG-derived MG-H1 formation [6,10]. As shown in Figure 4, AG pre-treatment completely abrogated the effects induced by S1 on RAGE expression (Figure 4a) and inflammatory cytokines (Figure 4 b-e), thus confirming in BEAS-2B cells, that do not express ACE2 receptor, a novel pathway involving MG-H1/RAGE through which S1 can orchestrate inflammation.
2.5. SARS-CoV-2 S1 spike protein controls MG-H1/RAGE proinflammatory pathway through the nuclear factor erythroid 2-related factor 2 (Nrf2)-dependent Glo1 down-regulation in human bronchial BEAS-2B cells
Intracellular MG and consequently MG-derived MG-H1 levels, are controlled by the enzyme Glo1 [9], which can be upregulated by Nrf2 [11]. It is also known that Nrf2 plays anti-inflammatory roles through several mechanisms including the suppression of pro-inflammatory genes such as IL-1β and IL-6 [12]. Based on these premises, in the attempt to understand the pathway acting upstream MG-H1/RAGE proinflammatory axis in BEAS-2B cells, we assumed that S1-triggered MG-H1 accumulation was dependent on Glo1 down-regulation, in turn mediated by Nrf2 signaling desensitization. To demonstrate our hypothesis, we first studied Glo1 and Nrf2 expression upon exposure to 25 ng/ml S1 for 24 h. As shown in Figure 5, S1 was able to inhibit Glo1 expression and activity (Figure 5a) as well as Nrf2 signaling (Figure 5b). More importantly, pretreatment with a Nrf2 activator (Nrf2-A) [11,13], rescued Glo1 expression and activity (Figure 5c), reduced MG-H1 intracellular accumulation (Figure 5d) and RAGE expression (Figure 5e) as well as inflammation (Figure 5f, g) compared with S1-challenged cells, thus confirming that SARS-CoV-2 S1 spike protein controls MG-H1/RAGE proinflammatory pathway through Nrf2-dependent Glo1 down-regulation in human bronchial BEAS-2B cells.
2.6. RAGE, Glo1 and IL-1β expression in nasopharyngeal swabs of SARS-CoV-2 infected patients at different clinical severity of Covid-19
To provide a potential clinical value to our in vitro data, we measured RAGE, Glo1 and IL-1β mRNA expression in swabs from the nasopharynx of SARS-CoV-2 infected patients, distributed on the basis of the clinical outcome into non-severe (not hospitalized: n = 20), and moderate/severe [total, n = 40: moderate (ordinary hospitalization), n = 20 - severe (intensive care), n = 20] disease groups. Mean age of the non-severe cohort was 64.75 years ± 19.3 (standard deviation, SD), median 65 years, range 32-89 years; mean age of the moderate/severe cohort was 76.05 years ± 16.0, median 80 years, range 29-96 years, p = 0.03. Nine females (F, 45%) and 11 males (M, 55%) were present in the non-severe group; while 22 F (55%) and 18 M (45%) were present in the moderate/severe group, p = 0.47. For comparison, a SARS-CoV-2 negative group (n = 19) was included (mean age 59.10 years ± 17.6, median 59 years, range 29-86 years; p = 0.34 vs the non-severe group, p = 0.0012 vs moderate/severe; 8 F (42%) and 11 M (58%). We found that patients with a moderate/severe disease presented higher levels of RAGE (Figure 6a) and IL-1β (Figure 6b), and lower levels of Glo1 (Figure 6c) expression, compared with the non-severe group, thus strongly suggesting that the mechanistic in vitro core results, showing MG-H1/RAGE pathway as a novel SARS-CoV-2 S1-driven pro-inflammatory axis, might occur during infection. The negative group presented no differences compared to the non-severe cohort with respect to the expression of all the considered genes (Figure 6).
3. Discussion
In the present research, we first observed, in line with the literature [1,14-17], that SARS-CoV-2 S1 spike surface protein is sufficient alone to activate the production of key COVID-19 pro-inflammatory cytokines (IL-1β, IL-6 and IL-8) in alveolar A549 cells, a major COVID-19 cell-type target. Moreover, we observed a comparable pro-inflammatory response also in human bronchial epithelial BEAS-2B cells, which was never investigated before. Altogether, these results are important in confirming the growing body of studies that show that the S1 spike protein, regardless of the whole virus, has a proinflammatory biological activity in lung epithelial cells. Despite epithelial cells, compared to innate immune cells, usually provide feebler inflammatory responses, they release cytokines and/or chemokines able to potently recruit monocytes, lymphocytes and neutrophils in the lungs infected by SARS-CoV-2, thus anyhow making a significant contribution to the “cytokine storm”-related immunopathology underpinning COVID-19 pathogenesis and progression [14]. Moreover, there is growing evidence supporting that sustained and protracted circulating levels of SARS-CoV-2 S1 spike1 protein are present in PASC [18-21]. Our findings suggest that these plasma levels, being S1 able to induce inflammation itself, may contribute to the sustained inflammatory-associated events, at least at lung level, that characterize post-acute COVID-19 syndrome [22]. Hence, our data contribute to focus on and strength the proinflammatory role of S1 and its implication in COVID-19 progression, including PACS, whose underlying mechanisms are entirely unknown.
Importantly, in this study we also demonstrated that in human bronchial epithelial BEAS-2B cells, SARS-CoV-2 S1 spike protein induces inflammation through a novel MG-H1/RAGE-based mechanism. MG-H1 is one of the specific advanced glycation end products originated by MG [3], able to initiate inflammatory pathways, frequently by binding to the receptor RAGE [4,7,8], that is mostly expressed in the lungs [4,8]. As previously reported [23,24], BEAS-2B cells expresses very low to undetectable levels of ACE2 receptor, to which S1 typically binds in order to allow endocytosis of the viral particle and begin its replicative cycle within the host cell. Hence, our MG-H1/RAGE axis would represent a novel mechanism for S1, and potentially a novel strategy for SARS-Cov-2, to start pathogenic inflammation in cells that do not express the surface ACE2 receptor. In support of this hypothesis, in A549 cells that express ACE2 [23], MG-H1/RAGE axis is not activated by S1.
Notably, aminoguanidine (AG), by specifically scavenging MG, thus preventing MG-derived MG-H1 formation [6,10], reverted RAGE-dependent inflammation. This result, in addition to demonstrate a causative role of MG-H1/RAGE axis in driving inflammation upon S1 challenge, indicates that AG could be promising in developing supportive therapeutics to prevent COVID-19-related pathogenic inflammation. Besides, AG has already proven potentiality in other inflammatory diseases [25-27]. Clearly, further investigation in this ambit is mandatory.
It is now consolidated that RAGE signaling produces pro-inflammatory responses via NF-kB activation [14,28] and, indeed, it has been recently described that SARS-CoV-2 spike protein is pro-inflammatory in pulmonary cells using NF-κB pathway activation [14,29-31]. Moreover, in pancreatic islets infected with SARS-CoV-2, elevated MG activates RAGE and NF-kB [32]. Finally, an upregulation of NF-kB has been observed in COVID-19 patients [33]. Hence, it is very likely that NF-kB may be involved down-stream RAGE activation also in our model and potentially upon virus infection.
With the aim of unveil how MG-H1 accumulation, leading to MG-H1/RAGE proinflammatory pathway activation, could occur in bronchial BEAS-2B cells, we found that this was ascribed to the down-regulation, at all expression levels, of the enzyme that modulates MG intracellular levels, named Glo1. Again, the mechanism seems to be specific for ACE2-independent BEAS-2B, since ACE2-dependent A549 cells showed increased Glo1 expression (Figure S3). It has been reported that glycolysis, the main route in the production of MG, is upregulated in SARS-CoV-2-infected cells to provide the substrate required for the virus replication [34,35]. Hence, it would be possible that also this metabolic route may contribute to S1-induced MG/MG-H1 accumulation in BEAS-2B cells, even though the fact that in A549 cells where S1 should still stimulate glycolysis, MG-H1 levels does not change upon S1 treatment, would make one conclude that Glo1 is indeed primarily responsible for it.
Moreover, we found that Glo1 down-regulation was dependent on S1-driven Nrf2 desensitization. It is known that the promoter region of Glo1 has a functionally operating antioxidant response element (ARE) [3] that is bound by Nrf2 to induce Glo1 expression. Here, we found that Nrf2 desensitization causatively led to Glo1 down-regulation, since Nrf2 activation restored Glo1 expression and enzyme activity. In line with previous results [11,36,37], we here confirm Glo1 control by Nrf2. Interestingly, and in agreement with our findings, a very recent in vivo study has demonstrated that S1 induces Nrf2 down-regulation in rats exposed to this virus subunit [38]. It is also noteworthy that a very complex interplay exists between Nrf2 and NF-kB pathways [39]. Nrf2 can decrease the activation of NF-kB, while NF-kB transcription can inhibit Nrf2 activation [40,41]. The existence of such a mutual interaction would seem to strength our hypothesis of NF-kB involvement in our pro-inflammatory Nrf2/Glo1/MG-H1/RAGE-(NF-kB) pathway, where Nrf2 desensitization could very likely explain NF-kB activation. Concomitantly, NF-kB activation could inhibit Nrf2 in a circuitry nurturing S1-driven inflammation through Glo1/MG-H1/RAGE axis, which is very intriguing and needs further investigation.
Finally, we found that patients with a moderate/severe disease presented higher levels of RAGE and IL-1β, and lower levels of Glo1 expression, compared with the non-severe group, thus strongly suggesting that the mechanistic in vitro core results, showing MG-H1/RAGE pathway as a novel SARS-CoV-2 S1-driven pro-inflammatory axis, might occur also during infection. Besides, Alomar et al. have recently reported significantly higher MG and IL-1β plasma levels in patients who required intensive care unit (ICU) hospitalization compared to uninfected individuals and significantly lower Glo1 levels in patients who died compared to ICU patients that survived [42]. They also found strong inverse correlations between plasma MG and Glo1 and significant positive correlations between plasma MG and IL-1β. In the same study, authors state, as one limitation of the research, the need of additional methodology to measure MG, Glo1 and IL-1β plasma levels. By detecting MG, Glo1 and IL-1β levels in swabs from the nasopharynx of individuals, by RT-PCR our results appear to support those of Alomar et al.
4. Materials and Methods
4.1. Reagents
The chemicals used in the present study are analytical grade reagents from various sources. The SARS-CoV-2 S1 spike protein was purchased from Prodotti Gianni S.p.A (Milan, Italy). MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] and aminoguanidine bicarbonate (AG) were purchased from Merck Spa (Milan, Italy). Nrf2 activator (Nrf2-A) was from EMD Millipore Corporation (Billerica, MA, USA). Nrf2-A was solubilized in dimethyl sulfoxide (DMSO, Merck Spa, Milan, Italy) (final concentration in the essays of DMSO was 0.01%). Controls contained an identical volume of DMSO vehicle. Laemmli buffer was from Invitrogen (Milan, Italy), Roti-Block from Roth (Roth, Germany) and bicinchoninic acid (BCA) kit from Pierce (USA).
4.2. Cell cultures
4.3. Cell viability and morphology
4.4. RNA Isolation, Reverse Transcription, and Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) Analyses
TRIzol Reagent (ThermoFisher Scientific, Milan, Italy) was used to isolate total RNA. RevertAid™ H Minus First Strand cDNA Synthesis Kit (ThermoFisher Scientific, Milan, Italy) was used to produce cDNA, as previously described [9]. Gene expression of the studied genes versus GAPDH was evaluated by RT-PCR, using MX3000P Real-Time PCR System (Agilent Technology, Milan, Italy). The sequences of the oligonucleotide primers are reported in Table 1.
A total volume of 20 µL containing 25 ng of cDNA, 1X Brilliant II SYBR® Green QPCR Master Mix, ROX Reference Dye, and 600 nM of specific primers was employed in PCR reactions, thermal cycling conditions being: 1 cycle (95 °C, 5 min); 45 cycles (95 °C, 20 s and 60 °C, 30 s). Comparative analysis of gene expression was performed by the 2−(∆∆CT) method [6].
4.5. TNF-α, IL-6 and IL-8 detection
TNF-α (cod. KHC3011), IL-6 (cod. BMS213HS) and IL-8 (cod. KHC0081) were measured by ELISA (ThermoFisher Scientific, Milan, Italy).
4.6. Detection of Methylglyoxal (MG)-H1
MG-H1 was measured by ELISA (cat. STA-811, DBA Italia S.r.l., Milan, Italy).
4.7. Cell and nuclear lysis and Western blot
Cell lysis was performed in radioimmunoprecipitation assay (RIPA) lysis buffer [44]. FractionPREP Cell Fractionation kit (Biovision, Vinci-Biochem, Florence, Italy) was used for nuclear extraction [44]. Western blot was performed as previously described [46]. Samples were boiled in Laemmli buffer for 5 min, resolved on SDS-PAGE and blotted onto a nitrocellulose membrane. Roti-Block (room temperature, RT, 1 h) was used to block unspecific binding sites. Membranes incubation with the appropriate primary Abs: mouse anti-Glo1 (D-6) mAb (dilution 1:1000, Santa Cruz, cat. # sc-133144, DBA Italia Srl, Segrate, Italy), mouse anti-RAGE (A-9) mAb (dilution 1:1000, Santa Cruz, cat. # sc-365154, DBA Italia Srl, Segrate, Italy) and mouse anti-GAPDH (6C5) mAb (dilution 1:1000, Santa Cruz, cat. # sc- 32233, DBA Italia Srl, Segrate, Italy), as internal loading control, was performed overnight at 4 °C. Membranes were then incubated with the appropriate HRP-conjugated secondary Ab (RT, 1 h) and ECL was used as revealing system (Amersham Pharmacia, Milan, Italy).
4.8. Nrf2 activation detection
Nrf2 Transcription Factor Assay kit (Colorimetric) (ab207223) (DBA Italia srl, Milan, Italy), was employed to detect Nrf2 activation in nuclear extracts [44].
4.9. Total protein and enzyme specific activity of Glyoxalase 1 (Glo1) detection
BCA kit (cat. 23225, ThermoFisher Scientific) was used to quantify total protein concentration. Bovine serum albumin was used as a standard. Glo1 enzyme activity was assayed by an established method [6].
4.10. Patients
Nasopharyngeal swabs (NPS) from individuals referring to the Hospital of Perugia for SARS-CoV-2 molecular testing were collected from December 2022 to May 2023. The study was approved by the Ethical Committee of the Umbria Region (CER Umbria, approval number 3990/21) and of the University of Perugia (15/2023 del 15.03.2023) and carried out in accordance with the Declaration of Helsinki. Informed consent was obtained from the subjects involved in the study. The cohort of SARS-CoV-2 infection cases consisted of 60 patients, stratified into 2 groups, on the basis of clinical symptom severity into non-severe (asymptomatic: n = 20) and moderate/severe (total n = 40; ordinary hospitalization, n = 20 and intensive care, n = 20) groups. Once the specimens were tested for SARS-CoV-2 viral load, the residual sample was used for DNA extraction performed by using STARMag 96 x 4 Universal Cartridge Kit (Seegene, Korea) on automated STARLET (Seegene, Korea), according to the manufacturer's instructions.
4.11. Statistical analysis
Results were expressed as means ± standard deviation (SD) of three independent experiments. One-way analysis of variance with Dunnett’ s correction was employed to determine differences among groups. T-test or χ2 were also applied in the analysis of patients’ data where appropriate. Statistical significance was set at p < 0.05.
5. Conclusions
In summary, we demonstrated that SARS-CoV-2 Spike protein S1 induces MG-H1/RAGE activation to promote inflammation in human bronchial BEAS-2B cells, in a mechanism involving Nrf2-dependent Glo1 down-regulation, which is a novel finding. Our results improve knowledge for a better understanding of the mechanisms by which S1, and potentially SARS-Cov-2, activates inflammatory pathways, which is fundamental to find preventive strategies and/or better treatment regimens aimed at contrasting COVID-19-related inflammation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Effect of SARS-CoV-2 Spike protein S1 on BEAS-2B and A549 cell morphology; Figure S2: SARS-CoV-2 S1 Spike protein induces inflammatory cytokines in BEAS-2B and A549; Figure S3: Glyoxalase 1 (Glo1) mRNA expression in alveolar A549 epithelial cells exposed to SARS-CoV-2 S1 spike protein; Figure S4: whole original blots.
Author Contributions
Conceptualization, C.A.; methodology, D.M., M.P., S.B, and A.G.; formal analysis, C.A. and S.B.; data curation, C.A. and D.M.; writing - original draft preparation, C.A.; writing - review and editing, P.P., L.R., V.N.T, C.C.; funding acquisition, C.A., P.P. and V.N.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC were funded by Fondazione Cassa di Risparmio di Perugia, grant number 19645 (2020.0425) and 21060 (2022.0402).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethical Committee of the Umbria Region (CER Umbria, approval number 3990/21) and of the University of Perugia (15/2023 del 15.03.2023).
Informed Consent Statement
Informed consent was obtained from the subjects involved in the study.
Data Availability Statement
Data are contained within the article or supplementary material.
Acknowledgments
We thank Roberta Frosini for her excellent technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Forsyth, C.B.; Zhang, L.; Bhushan, A.; Swanson, B.; Zhang, L.; Mamede, J.I.; Voigt, R.M.; Shaikh, M.; Engen, P.A.; Keshavarzian, A. The SARS-CoV-2 S1 Spike Protein Promotes MAPK and NF-kB Activation in Human Lung Cells and Inflammatory Cytokine Production in Human Lung and Intestinal Epithelial Cells. Microorganisms 2022, 10, 1996. [Google Scholar] [CrossRef] [PubMed]
- Hasankhani, A.; Bahrami, A.; Tavakoli-Far, B.; Iranshahi, S.; Ghaemi, F.; Akbarizadeh, M.R.; Amin, A.H.; Kiasari, B.A.; Shabestari, A.M. The role of peroxisome proliferator-activated receptors in the modulation of hyperinflammation induced by SARS-CoV-2 infection: A perspective for COVID-19 therapy. Front. Immunol. 2023, 14, 1127358. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Talesa, V.N. Glyoxalases in Urological Malignancies. Int. J. Mol. Sci. 2018, 19, 415. [Google Scholar] [CrossRef]
- Pariano, M.; Costantini, C.; Santarelli, I.; Puccetti, M.; Giovagnoli, S.; Talesa, V.N.; Romani, L.; Antognelli, C. Defective Glyoxalase 1 Contributes to Pathogenic Inflammation in Cystic Fibrosis. Vaccines 2021, 9, 1311. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.W.T.; Gonzalez, E.D.J.L.; Zoukari, T.; Ki, P.; Shuck, S.C. Methylglyoxal and Its Adducts: Induction, Repair, and Association with Disease. Chem. Res. Toxicol. 2022, 35, 1720–1746. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Mancuso, F.; Frosini, R.; Arato, I.; Calvitti, M.; Calafiore, R.; Talesa, V.N.; Luca, G. Testosterone and Follicle Stimulating Hormone–Dependent Glyoxalase 1 Up-Regulation Sustains the Viability of Porcine Sertoli Cells through the Control of Hydroimidazolone– and Argpyrimidine-Mediated NF-κB Pathway. Am. J. Pathol. 2018, 188, 2553–2563. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Matsui, T.; Nakamura, N.; Sotokawauchi, A.; Higashimoto, Y.; Yamagishi, S.-I. Methylglyoxal-derived hydroimidazolone-1 evokes inflammatory reactions in endothelial cells via an interaction with receptor for advanced glycation end products. Diabetes Vasc. Dis. Res. 2017, 14, 450–453. [Google Scholar] [CrossRef]
- Lim, J.-M.; Yoo, H.J.; Lee, K.-W. High Molecular Weight Fucoidan Restores Intestinal Integrity by Regulating Inflammation and Tight Junction Loss Induced by Methylglyoxal-Derived Hydroimidazolone-1. Mar. Drugs 2022, 20, 580. [Google Scholar] [CrossRef]
- Antognelli, C.; Ferri, I.; Bellezza, G.; Siccu, P.; Love, H.D.; Talesa, V.N.; Sidoni, A. Glyoxalase 2 drives tumorigenesis in human prostate cells in a mechanism involving androgen receptor and p53-p21 axis. Mol. Carcinog. 2017, 56, 2112–2126. [Google Scholar] [CrossRef]
- Antognelli, C.; Marinucci, L.; Frosini, R.; Macchioni, L.; Talesa, V.N. Metastatic Prostate Cancer Cells Secrete Methylglyoxal-Derived MG-H1 to Reprogram Human Osteoblasts into a Dedifferentiated, Malignant-like Phenotype: A Possible Novel Player in Prostate Cancer Bone Metastases. Int. J. Mol. Sci. 2021, 22, 10191. [Google Scholar] [CrossRef]
- Monache, S.D.; Pulcini, F.; Frosini, R.; Mattei, V.; Talesa, V.N.; Antognelli, C. Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway. Antioxidants 2021, 10, 716. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Patel, S.; Majumdar, A. Role of NRF2 and Sirtuin activators in COVID-19. Clin. Immunol. 2021, 233, 108879–108879. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kumar, S.; Hassan, M.; Wu, H.; Thimmulappa, R.K.; Kumar, A.; Sharma, S.K.; Parmar, V.S.; Biswal, S.; Malhotra, S.V. Novel Chalcone Derivatives as Potent Nrf2 Activators in Mice and Human Lung Epithelial Cells. J. Med. Chem. 2011, 54, 4147–4159. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Shafiei, M.S.; Longoria, C.; Schoggins, J.W.; Savani, R.C.; Zaki, H. SARS-CoV-2 spike protein induces inflammation via TLR2-dependent activation of the NF-κB pathway. eLife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Arjsri, P.; Srisawad, K.; Mapoung, S.; Semmarath, W.; Thippraphan, P.; Umsumarng, S.; Yodkeeree, S.; Dejkriengkraikul, P. Hesperetin from Root Extract of Clerodendrum petasites S. Moore Inhibits SARS-CoV-2 Spike Protein S1 Subunit-Induced NLRP3 Inflammasome in A549 Lung Cells via Modulation of the Akt/MAPK/AP-1 Pathway. Int. J. Mol. Sci. 2022, 23, 10346. [Google Scholar] [CrossRef]
- Chittasupho, C.; Srisawad, K.; Arjsri, P.; Phongpradist, R.; Tingya, W.; Ampasavate, C.; Dejkriengkraikul, P. Targeting Spike Glycoprotein S1 Mediated by NLRP3 Inflammasome Machinery and the Cytokine Releases in A549 Lung Epithelial Cells by Nanocurcumin. Pharmaceuticals 2023, 16, 862. [Google Scholar] [CrossRef]
- Semmarath, W.; Mapoung, S.; Umsumarng, S.; Arjsri, P.; Srisawad, K.; Thippraphan, P.; Yodkeeree, S.; Dejkriengkraikul, P. Cyanidin-3-O-glucoside and Peonidin-3-O-glucoside-Rich Fraction of Black Rice Germ and Bran Suppresses Inflammatory Responses from SARS-CoV-2 Spike Glycoprotein S1-Induction In Vitro in A549 Lung Cells and THP-1 Macrophages via Inhibition of the NLRP3 Inflammasome Pathway. Nutrients 2022, 14, 2738. [Google Scholar] [CrossRef]
- Suzuki, Y.J.; Gychka, S.G. SARS-CoV-2 Spike Protein Elicits Cell Signaling in Human Host Cells: Implications for Possible Consequences of COVID-19 Vaccines. Vaccines 2021, 9, 36. [Google Scholar] [CrossRef]
- Patterson, B.K.; Francisco, E.B.; Yogendra, R.; Long, E.; Pise, A.; Rodrigues, H.; Hall, E.; Herrera, M.; Parikh, P.; Guevara-Coto, J.; et al. Persistence of SARS CoV-2 S1 Protein in CD16+ Monocytes in Post-Acute Sequelae of COVID-19 (PASC) up to 15 Months Post-Infection. Front. Immunol. 2022, 12, 746021. [Google Scholar] [CrossRef]
- Files, J.K.; Sarkar, S.; Fram. , T.R.; Boppana, S.; Sterrett, S.; Qin, K.; Bansal, A.; Long, D.M.; Sabbaj, S.; Kobie, J.J.; et al. Duration of post-COVID-19 symptoms are associated with sustained SARS-CoV-2 specific immune responses. J. Clin. Investig. 2021, 6. [Google Scholar] [CrossRef]
- Swank, Z.; Senussi, Y.; Manickas-Hill, Z.; Yu, X.G.; Li, J.Z.; Alter, G.; Walt, D.R. Persistent Circulating Severe Acute Respiratory Syndrome Coronavirus 2 Spike Is Associated With Post-acute Coronavirus Disease 2019 Sequelae. Clin. Infect. Dis. 2022, 76, e487–e490. [Google Scholar] [CrossRef] [PubMed]
- Martins-Gonçalves, R.; Hottz, E.D.; Bozza, P.T. Acute to post-acute COVID-19 thromboinflammation persistence: Mechanisms and potential consequences. Curr. Res. Immunol. 2023, 4, 100058–100058. [Google Scholar] [CrossRef] [PubMed]
- Bartolomeo, C.S.; Lemes, R.M.R.; Morais, R.L.; Pereria, G.C.; Nunes, T.A.; Costa, A.J.; Maciel, R.M.d.B.; Braconi, C.T.; Maricato, J.T.; Janini, L.M.R.; et al. SARS-CoV-2 infection and replication kinetics in different human cell types: The role of autophagy, cellular metabolism and ACE2 expression. Life Sci. 2022, 308, 120930–120930. [Google Scholar] [CrossRef]
- Tsou, H.-H.; Wang, P.-H.; Ting, T.-H.; Ping, Y.-H.; Liu, T.-Y.; Cheng, H.-W.; Wang, H.-T. Effect of heated tobacco products and traditional cigarettes on pulmonary toxicity and SARS-CoV-2-induced lung injury. Toxicology 2022, 479, 153318–153318. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Zhang, Y.; Zhang, L.; Song, Z. Synthesis and Biological Evaluation of Paclitaxel-aminoguanidine Conjugates for Suppressing Breast Cancer. Curr. Org. Synth. 2023, 20, 890–896. [Google Scholar] [CrossRef]
- Ma, Y.; Song, X.; Ma, T.; Li, Y.; Bai, H.; Zhang, Z.; Hu, H.; Yuan, R.; Wen, Y.; Gao, L. Aminoguanidine inhibits IL-1β-induced protein expression of iNOS and COX-2 by blocking the NF-κB signaling pathway in rat articular chondrocytes. Exp. Ther. Med. 2020, 20, 2623–2630. [Google Scholar] [CrossRef]
- Antognelli, C.; Moretti, S.; Frosini, R.; Puxeddu, E.; Sidoni, A.; Talesa, V.N. Methylglyoxal Acts as a Tumor-Promoting Factor in Anaplastic Thyroid Cancer. Cells 2019, 8, 547. [Google Scholar] [CrossRef]
- Tobon-Velasco, J.C.; Cuevas, E.; Torres-Ramos, M.A. Receptor for AGEs (RAGE) as Mediator of NF-kB Pathway Activation in Neuroinflammation and Oxidative Stress. CNS Neurol. Disord. Drug Targets 2014, 13, 1615–1626. [Google Scholar] [CrossRef]
- Schreiber, A.; Viemann, D.; Schöning, J.; Schloer, S.; Zambrano, A.M.; Brunotte, L.; Faist, A.; Schöfbänker, M.; Hrincius, E.; Hoffmann, H.; et al. The MEK1/2-inhibitor ATR-002 efficiently blocks SARS-CoV-2 propagation and alleviates pro-inflammatory cytokine/chemokine responses. Cell. Mol. Life Sci. 2022, 79, 1–18. [Google Scholar] [CrossRef]
- Paidi, R.K.; Jana, M.; Mishra, R.K.; Dutta, D.; Pahan, K. Selective Inhibition of the Interaction between SARS-CoV-2 Spike S1 and ACE2 by SPIDAR Peptide Induces Anti-Inflammatory Therapeutic Responses. J. Immunol. 2021, 207, 2521–2533. [Google Scholar] [CrossRef]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Bianchi, M.G.; Dall’asta, V.; Rotoli, B.M. Immune-Mediated Inflammatory Responses of Alveolar Epithelial Cells: Implications for COVID-19 Lung Pathology. Biomedicines 2022, 10, 618. [Google Scholar] [CrossRef] [PubMed]
- A Alomar, F. Methylglyoxal in COVID-19-induced hyperglycemia and new-onset diabetes. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 8152–8171. [Google Scholar] [CrossRef]
- Xia, J.; Tang, W.; Wang, J.; Lai, D.; Xu, Q.; Huang, R.; Hu, Y.; Gong, X.; Fan, J.; Shu, Q.; et al. SARS-CoV-2 N Protein Induces Acute Lung Injury in Mice via NF-ĸB Activation. Front. Immunol. 2021, 12, 791753. [Google Scholar] [CrossRef] [PubMed]
- Moolamalla, S.; Balasubramanian, R.; Chauhan, R.; Priyakumar, U.D.; Vinod, P. Host metabolic reprogramming in response to SARS-CoV-2 infection: A systems biology approach. Microb. Pathog. 2021, 158, 105114–105114. [Google Scholar] [CrossRef] [PubMed]
- Codo, A.C.; Davanzo, G.G.; Monteiro, L.d.B.; de Souza, G.F.; Muraro, S.P.; Virgilio-Da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; Junior, C.A.O.d.B.; Crunfli, F.; et al. Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020, 32, 498–499. [Google Scholar] [CrossRef]
- Antognelli, C.; Trapani, E.; Delle Monache, S.; Perrelli, A.; Daga, M.; Pizzimenti, S.; Barrera, G.; Cassoni, P.; Angelucci, A.; Trabalzini, L.; et al. KRIT1 loss-of-function induces a chronic Nrf2-mediated adaptive homeostasis that sensitizes cells to oxidative stress: Implication for Cerebral Cavernous Malformation disease. Free Radic. Biol. Med. 2018, 115, 202–218. [Google Scholar] [CrossRef]
- Antognelli, C.; Trapani, E.; Delle Monache, S.; Perrelli, A.; Fornelli, C.; Retta, F.; Cassoni, P.; Talesa, V.N.; Retta, S.F. Data in support of sustained upregulation of adaptive redox homeostasis mechanisms caused by KRIT1 loss-of-function. Data Brief 2018, 16, 929–938. [Google Scholar] [CrossRef]
- Sun, Q.; Li, L.; Jin, F.; Liu, Y.; Yang, B.; Meng, W.; Zhang, Z.; Qi, F. SARS-CoV-2 Spike Protein S1 Exposure Increases Susceptibility to Angiotensin II-Induced Hypertension in Rats by Promoting Central Neuroinflammation and Oxidative Stress. Neurochem. Res. 2023, 48, 3016–3026. [Google Scholar] [CrossRef]
- Casper, E. The crosstalk between Nrf2 and NF-κB pathways in coronary artery disease: Can it be regulated by SIRT6? Life Sci. 2023, 330, 122007. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; León, R.; López, M.G.; Oliva, B.; et al. Transcription Factor NRF2 as a Therapeutic Target for Chronic Diseases: A Systems Medicine Approach. Pharmacol. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef] [PubMed]
- Alomar, F.A.; Alshakhs, M.N.; Abohelaika, S.; Almarzouk, H.M.; Almualim, M.; Al-Ali, A.K.; Al-Muhanna, F.; Alomar, M.F.; Alhaddad, M.J.; Almulaify, M.S.; et al. Elevated plasma level of the glycolysis byproduct methylglyoxal on admission is an independent biomarker of mortality in ICU COVID-19 patients. Sci. Rep. 2022, 12, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Frosini, R.; Santolla, M.F.; Peirce, M.J.; Talesa, V.N. Oleuropein-Induced Apoptosis Is Mediated by Mitochondrial Glyoxalase 2 in NSCLC A549 Cells: A Mechanistic Inside and a Possible Novel Nonenzymatic Role for an Ancient Enzyme. Oxidative Med. Cell. Longev. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gambelunghe, A.; Giovagnoli, S.; Di Michele, A.; Boncompagni, S.; Dell’omo, M.; Leopold, K.; Iavicoli, I.; Talesa, V.N.; Antognelli, C. Redox-Sensitive Glyoxalase 1 Up-Regulation Is Crucial for Protecting Human Lung Cells from Gold Nanoparticles Toxicity. Antioxidants 2020, 9, 697. [Google Scholar] [CrossRef]
- Marinucci, L.; Balloni, S.; Fettucciari, K.; Bodo, M.; Talesa, V.N.; Antognelli, C. Nicotine induces apoptosis in human osteoblasts via a novel mechanism driven by H 2 O 2 and entailing Glyoxalase 1-dependent MG-H1 accumulation leading to TG2-mediated NF-kB desensitization: Implication for smokers-related osteoporosis. Free. Radic. Biol. Med. 2018, 117, 6–17. [Google Scholar] [CrossRef]
- Antognelli, C.; Mandarano, M.; Prosperi, E.; Sidoni, A.; Talesa, V.N. Glyoxalase-1-Dependent Methylglyoxal Depletion Sustains PD-L1 Expression in Metastatic Prostate Cancer Cells: A Novel Mechanism in Cancer Immunosurveillance Escape and a Potential Novel Target to Overcome PD-L1 Blockade Resistance. Cancers 2021, 13, 2965. [Google Scholar] [CrossRef]
Figure 1.
Effect of SARS-CoV-2 S1 spike protein on human bronchial BEAS-2B and alveolar A549 cell viability. Viability of (a,b) BEAS and (c,d) A549 cells unexposed or exposed to S1 was measured by MTT assay. Data report the means of three separate experiments carried out in triplicate and error bars correspond to the standard deviation (SD) of the mean.
Figure 1.
Effect of SARS-CoV-2 S1 spike protein on human bronchial BEAS-2B and alveolar A549 cell viability. Viability of (a,b) BEAS and (c,d) A549 cells unexposed or exposed to S1 was measured by MTT assay. Data report the means of three separate experiments carried out in triplicate and error bars correspond to the standard deviation (SD) of the mean.
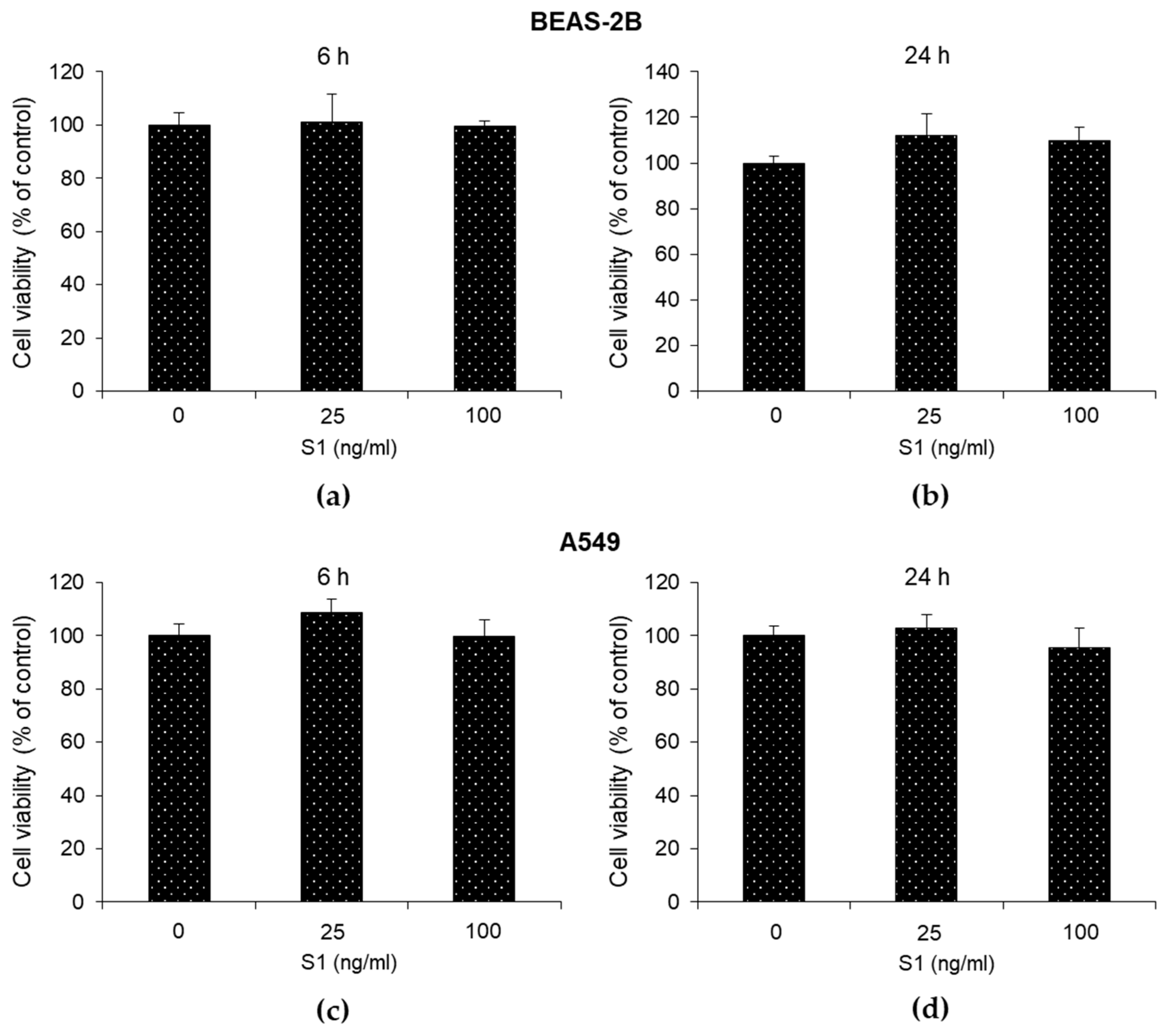
Figure 2.
SARS-CoV-2 S1 spike protein induces inflammatory cytokines in human bronchial BEAS-2B and alveolar A549 epithelial cells. BEAS-2B and A549 cells were stimulated with S1 at a concentration of 25 ng/ml. Six and 24 h post-stimulation, the expression of IL-1β was evaluated by real-time RT-PCR, while TNF-α, IL-6 and IL-8 levels, by ELISA. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 2.
SARS-CoV-2 S1 spike protein induces inflammatory cytokines in human bronchial BEAS-2B and alveolar A549 epithelial cells. BEAS-2B and A549 cells were stimulated with S1 at a concentration of 25 ng/ml. Six and 24 h post-stimulation, the expression of IL-1β was evaluated by real-time RT-PCR, while TNF-α, IL-6 and IL-8 levels, by ELISA. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001.
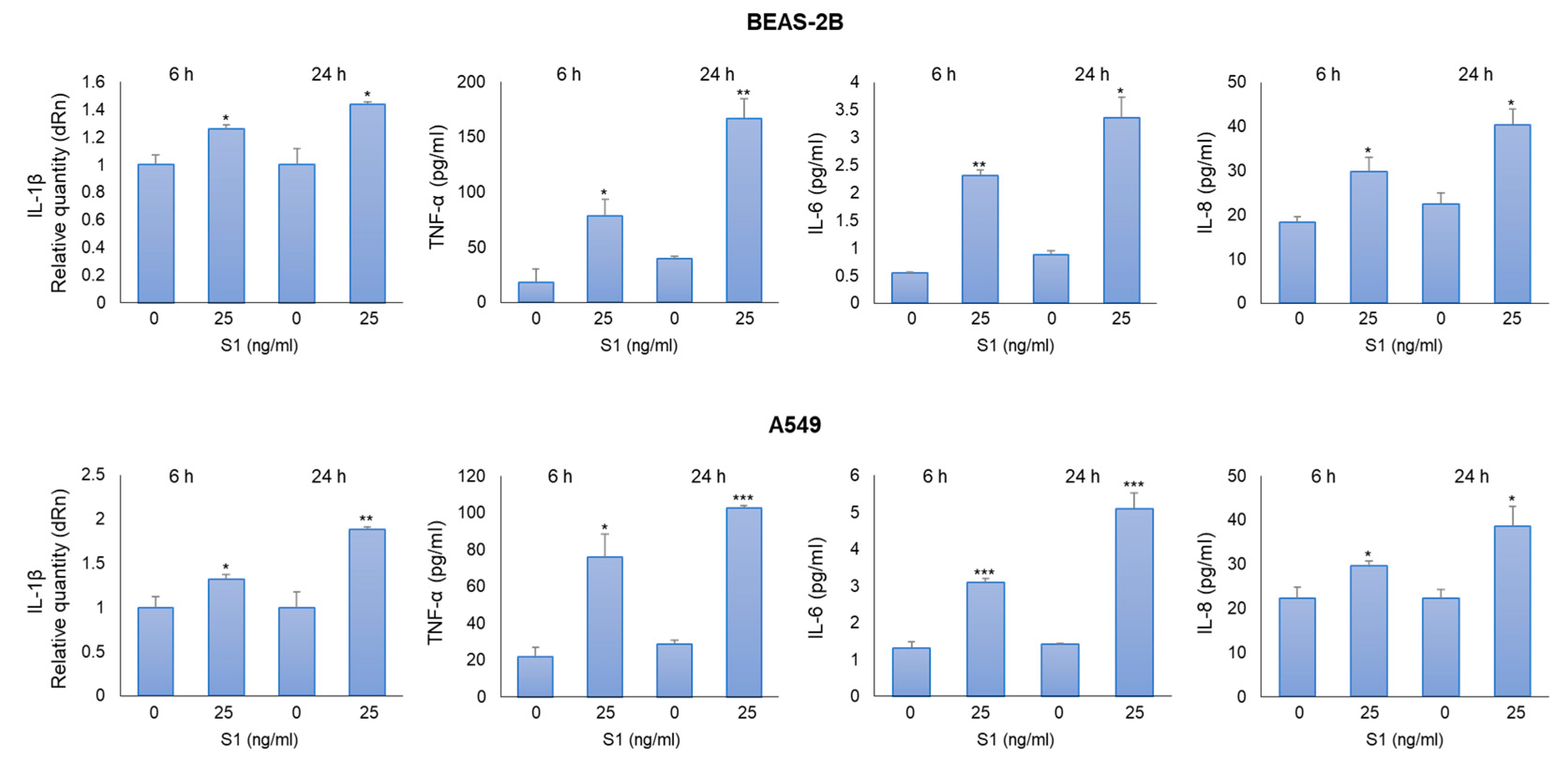
Figure 3.
SARS-CoV-2 S1 spike protein affects MG-H1 levels and RAGE expression in human bronchial BEAS-2B cells. BEAS-2B and A549 cells were stimulated with S1 at a concentration of 25 ng/ml. 24 h post-stimulation, the levels of MG-H1 (a,c) were evaluated by a specific ELISA kit, while RAGE mRNA and protein expression (b,d), by real-time RT-PCR and western blot (WB), respectively. GAPDH was used as internal loading control for WB normalization. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01.
Figure 3.
SARS-CoV-2 S1 spike protein affects MG-H1 levels and RAGE expression in human bronchial BEAS-2B cells. BEAS-2B and A549 cells were stimulated with S1 at a concentration of 25 ng/ml. 24 h post-stimulation, the levels of MG-H1 (a,c) were evaluated by a specific ELISA kit, while RAGE mRNA and protein expression (b,d), by real-time RT-PCR and western blot (WB), respectively. GAPDH was used as internal loading control for WB normalization. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01.
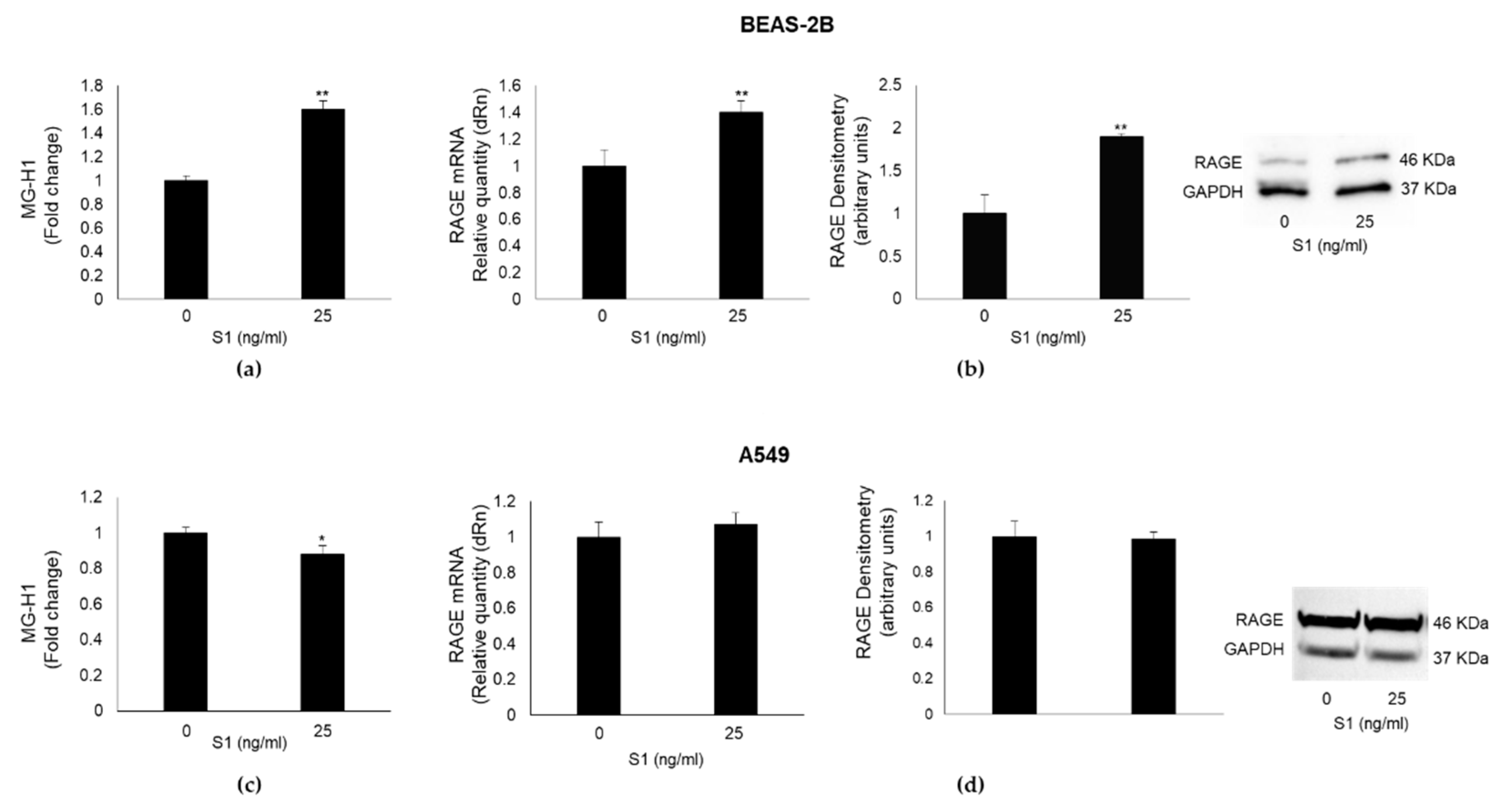
Figure 4.
SARS-CoV-2 S1 spike protein induces inflammation in human bronchial BEAS-2B cells through MG-H1/RAGE axis. BEAS-2B cells were pretreated with the specific MG scavenger aminoguanidine (AG) (10 mM for 24 h). 25 ng/ml S1 spike protein was then added and left for further 24 h. The expression of RAGE and IL-1β was evaluated by real-time RT-PCR, while TNF-α, IL-6 and IL-8 levels, by ELISA. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01.
Figure 4.
SARS-CoV-2 S1 spike protein induces inflammation in human bronchial BEAS-2B cells through MG-H1/RAGE axis. BEAS-2B cells were pretreated with the specific MG scavenger aminoguanidine (AG) (10 mM for 24 h). 25 ng/ml S1 spike protein was then added and left for further 24 h. The expression of RAGE and IL-1β was evaluated by real-time RT-PCR, while TNF-α, IL-6 and IL-8 levels, by ELISA. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01.
Figure 5.
SARS-CoV-2 S1 spike protein controls MG-H1/RAGE proinflammatory pathway through Nrf2-dependent Glyoxalase 1 (Glo1) down-regulation in human bronchial BEAS-2B cells. BEAS-2B cells were exposed to 25 ng/ml S1 for 24 h and (a) Glo1 mRNA, protein expression, specific activity, evaluated by real-time RT-PCR, western blot (WB) and spectrophotometric assay, respectively, and (b) Nrf2 nuclear expression, evaluated by a specific assay, were studied. Pretreatment with 10 µM Nrf2 activator (Nrf2-A) rescued (c) Glo1 expression and activity, reduced (d) MG-H1 intracellular accumulation, measured by a specific ELISA kit, (e) RAGE expression, evaluated by real-time RT-PCR, as well as inflammation, evaluated by IL-1β (f) and IL-6 levels (g), by real-time RT-PCR and ELISA, respectively, compared with S1-challenged cells. GAPDH was used as internal loading control for WB normalization. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 5.
SARS-CoV-2 S1 spike protein controls MG-H1/RAGE proinflammatory pathway through Nrf2-dependent Glyoxalase 1 (Glo1) down-regulation in human bronchial BEAS-2B cells. BEAS-2B cells were exposed to 25 ng/ml S1 for 24 h and (a) Glo1 mRNA, protein expression, specific activity, evaluated by real-time RT-PCR, western blot (WB) and spectrophotometric assay, respectively, and (b) Nrf2 nuclear expression, evaluated by a specific assay, were studied. Pretreatment with 10 µM Nrf2 activator (Nrf2-A) rescued (c) Glo1 expression and activity, reduced (d) MG-H1 intracellular accumulation, measured by a specific ELISA kit, (e) RAGE expression, evaluated by real-time RT-PCR, as well as inflammation, evaluated by IL-1β (f) and IL-6 levels (g), by real-time RT-PCR and ELISA, respectively, compared with S1-challenged cells. GAPDH was used as internal loading control for WB normalization. Data represent mean ± SD (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001.
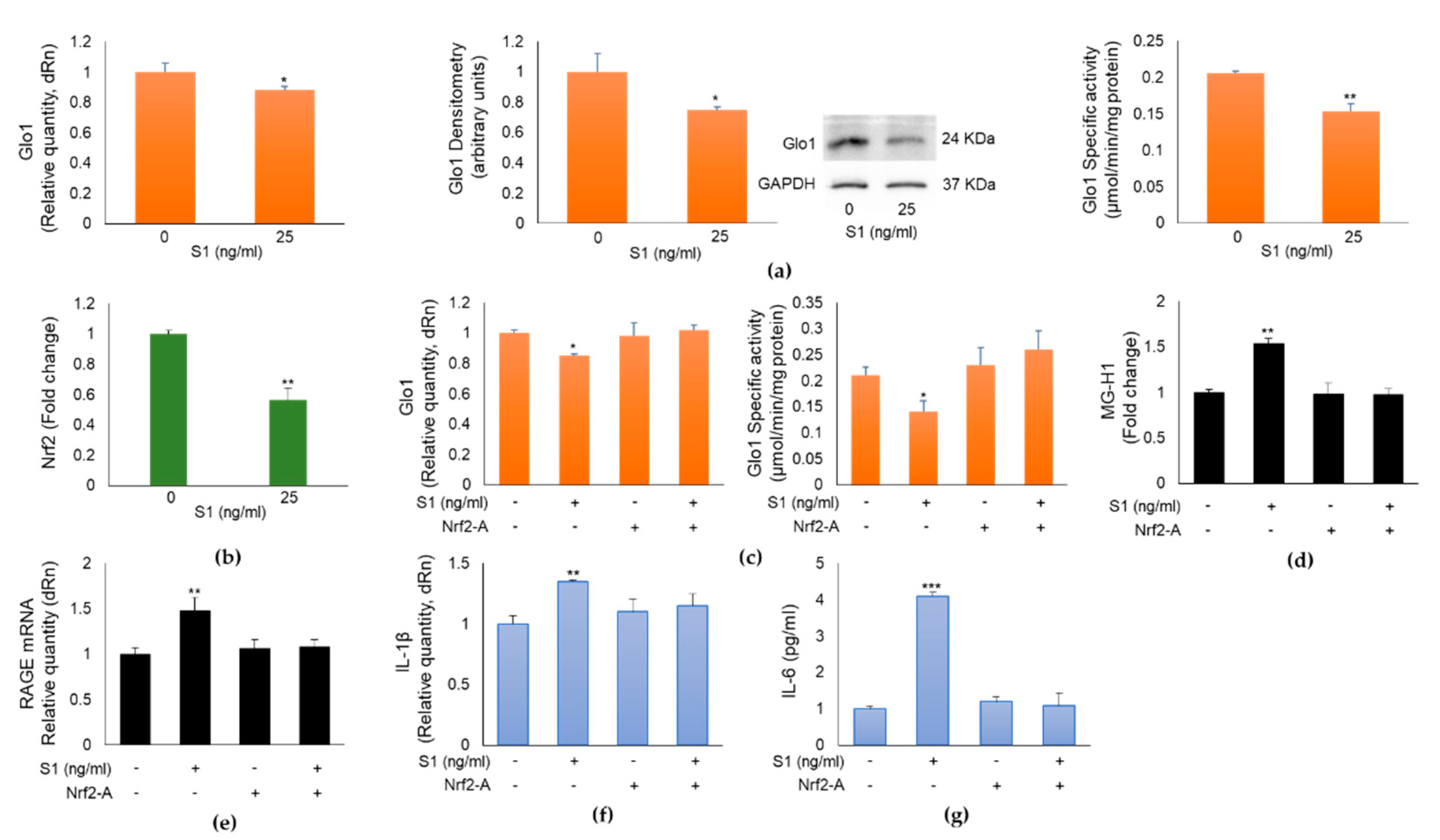
Figure 6.
RAGE, IL-1β and Glo1 expression in nasopharyngeal swabs of SARS-CoV-2 infected patients at different clinical severity of Covid-19. Gene expression of (a) RAGE, (b) IL-1β and (c) Glo1was evaluated by real-time RT-PCR. N: SARS-Cov-2 negative group; NS: SARS-Cov-2 positive with non-severe Covid-19 disease; M/S: SARS-Cov-2 positive with moderate/severe Covid-19 disease. *p < 0.05, **p < 0.01.
Figure 6.
RAGE, IL-1β and Glo1 expression in nasopharyngeal swabs of SARS-CoV-2 infected patients at different clinical severity of Covid-19. Gene expression of (a) RAGE, (b) IL-1β and (c) Glo1was evaluated by real-time RT-PCR. N: SARS-Cov-2 negative group; NS: SARS-Cov-2 positive with non-severe Covid-19 disease; M/S: SARS-Cov-2 positive with moderate/severe Covid-19 disease. *p < 0.05, **p < 0.01.
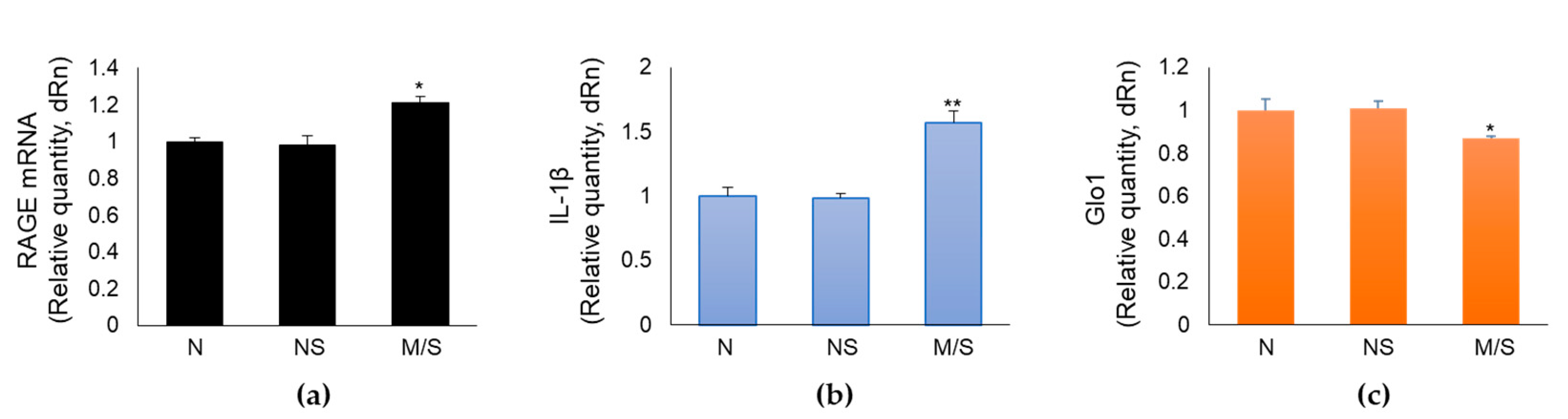

Table 1.
Sequences of oligonucleotide primers.
| Gene | Sense primer (5’-3’) | Antisense primer (5’-3’) |
|---|---|---|
| Glo1 | AGAAAGCACGGGGTGAAACTG | TACACCTTCAGTCCCGACTCC |
| IL-1β | GGACCTGGACCTCTGCCCTCTGG | GCCTGCCTGAAGCCCTTGCTGTAG |
| RAGE | TGAAGGAACAGACCAGGAGACAC | GCACAGGCTCCCAGACAC |
| GAPDH | CAAGGTCATCCATGACAACTTTG | GTCCACCACCCTGTTGCTGTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



