
Submitted:
30 August 2023
Posted:
01 September 2023
You are already at the latest version
Abstract
Streptococcus suis serotype 2 is an important swine bacterial pathogen causing sudden death, septic shock, and meningitis. However, serotype 2 strains are phenotypically and genotypically heterogeneous and composed by a multitude of sequence types (STs) whose distribution greatly varies worldwide. It has been previously shown that lipoprotein (LPP) maturation enzymes diacylglyceryl transferase (Lgt) and signal peptidase (Lsp) significantly modulate the inflammatory host response and play a differential role in virulence depending on the genetic background of the strain. Differently from Eurasian ST1/ST7 strains, the capsular polysaccharide of a North American S. suis serotype 2 ST25 representative strain only partially masks sub-capsular domains and bacterial wall components. Thus, our hypothesis is that since LPPs would be more surface exposed in ST25 strains than in their ST1 or ST7 counterparts, the maturation enzymes would play a more important role in the pathogenesis of the infection caused by the North American strain. Using isogenic Δlgt and Δlsp mutants derived from the wild-type ST25 strain, our studies suggest that these enzymes do not seem to play a role in the interaction between S. suis and epithelial and endothelial cells, regardless the genetics background of the strain used. However, a role in the formation of biofilms (also independently of the STs) has been demonstrated. Moreover, the involvement of LPP dendritic cell activation in vitro seems to be somehow more pronounced with the ST25 strain. Finally, the Lgt enzyme seems to play a more important role in the virulence of the ST25 strain. Although some differences between STs could be observed, our original hypothesis that LPPs would be significantly more important in ST25 strains due to a better bacterial surface exposition could not be confirmed.
Keywords:
Streptococcus suis
; Serotype 2
; North America
; Lipoprotein maturation enzymes
; Inflammation
1. Introduction
Streptococcus suis is one of the most important bacterial swine pathogens. It has been associated to a variety of infections including meningitis, septicemia, arthritis, and endocarditis. S. suis is also an emerging zoonotic agent causing mainly meningitis and septicemia with or without septic shock [1]. A total of 29 serotypes have been described and are defined based on the antigenicity of their capsular polysaccharide (CPS). Among them, serotype 2 is the most frequently type isolated from diseased pigs and humans worldwide [2]. However, evidence accumulated throughout the years has demonstrated a high level of genetic and phenotypic diversity within the S. suis species and serotypes. Therefore, serotype 2 strains are heterogeneous and belong to numerous sequence types (STs), as determined using multilocus sequence typing, with the highly virulent ST1 predominating in Eurasia, the epidemic virulent ST7 in China, and the virulent ST25 and lower virulent ST28 in Canada and USA [2-4]. Albeit these important differences, studies on the pathogenesis of the infection have dominantly used Eurasian strains, even though ST25 strains account for nearly 50% of serotype 2 isolates recovered from diseased pigs in Canada [5].
Although available information on the pathogenesis of the infection caused by S. suis has improved in recent years, knowledge about the mechanisms by which S. suis induces disease remains incomplete [1]. A variety of virulence factors have been proposed to be involved in the S. suis pathogenesis, including the CPS, cell-wall proteins, the suilysin, lipoteichoic acid modifications and lipoproteins (LPPs) [6]. The majority of these factors have been described using Eurasian ST1 or ST7 serotype 2 strains. LPPs, a major class of surface proteins of this and other bacterial pathogens [7], are mainly anchored in the outer leaflet of the cytoplasmic membrane. Bacterial LPPs have been shown to perform various roles, such as nutrient uptake, signal transduction as well as a participation in antibiotic resistance and transport systems (such as ABC transporters) [8]. In addition, they have been shown to play a direct role in virulence-associated functions, such as colonization, adhesion and invasion, evasion of host defense, and immunomodulation [7,9-12]. LPPs are first translated as preprolipoproteins, which possess an N-terminal signal peptide with typical characteristic features of the signal peptides of secreted proteins. A conserved sequence at the C region of the signal peptides, referred to as lipobox, is modified through the covalent attachment of a diacylglycerol moiety to the thiol group of the cysteine residue. This modification is catalyzed by the enzyme lipoprotein diacylglyceryl transferase (Lgt), resulting in a prolipoprotein. After lipidation, lipoprotein signal peptidase (Lsp) is responsible for cleaving the signal sequence of the lipidated prolipoprotein and leaves the cysteine of the lipobox as the new amino-terminal residue, resulting in mature LPPs [13].
A recent study suggests that LPP maturation in S. suis regulates dendritic cell activation in vitro and host activation after infection. In addition, these enzymes seem to play a differential role in virulence depending on the genetic background of the strain (ST1 vs. ST7) [7]. It has been reported that, differently from Eurasian strains, the CPS of a North American S. suis serotype 2 ST25 representative strain only partially masks sub-capsular domains and bacterial wall components [14,15]. Thus, our hypothesis is that LPP maturation enzymes would be more surface exposed in ST25 strains than in their ST1 or ST7 counterparts, playing a more important role in the pathogenesis of the infection. In the present work, the role of the Lgt and Lsp enzymes in bacterial adhesion/invasion of porcine epithelial and endothelial cells, biofilm formation, in vitro and in vivo induction of inflammatory mediators and virulence of S. suis serotype 2 ST25 strain have been studied.
2. Materials and Methods
2.1. Bacterial strains and growth conditions
The strains and plasmids used in this study are listed in Table 1. The wild-type S. suis serotype 2 strains P1/7 (ST1) and North American 89-1591 (ST25), previously used in our studies [7,15] and their lipoprotein maturation isogenic mutants previously obtained (P1/7) or those generated in the current study (89-1591) are also listed in Table 1. S. suis strains were cultured in Todd Hewitt broth (THB; Becton Dickinson, Mississauga, ON, Canada) as previously described [16]. The coagglutination test was performed with the standard technique [17]. Non-encapsulated CPS mutants (cspF) derived from strain P1/7 and 89-1591 were used as positive controls for some experiments [14]. Escherichia coli strains, and different plasmids used in this study are also listed in Table 1. For in vitro cell culture assays, bacteria were prepared as previously described and resuspended in cell culture medium. When needed, antibiotics (Sigma-Aldrich, Oakville, ON, Canada) were added to the media at the following concentrations: for S. suis, spectinomycin (Spc) at 100 μg/mL; for E. coli, kanamycin (Km) and spectinomycin at 50 μg/mL and ampicillin (Ap) at 100 μg/mL.
2.2. DNA manipulations
Genomic DNA was extracted from the S. suis wild-type strain 89-1591 using InstaGene Matrix solution (BioRad Laboratories, Hercules, CA, USA). Mini preparations of recombinant plasmids were carried out using the QIAprep Spin Miniprep Kit (Qiagen, Valencia, CA, USA). Restriction enzymes and DNA-modifying enzymes (Fisher Scientific, Ottawa, ON, Canada) were used according to the manufacturer’s recommendations. Oligonucleotide primers (Table 2) were obtained from Integrated DNA Technologies (Coralville, IA, USA) and PCRs carried out with the iProof proofreading DNA polymerase (BioRad Laboratories, Mississauga, ON, Canada) or the Taq DNA polymerase (Qiagen). Amplification products were purified using the QIAquick PCR Purification Kit (Qiagen) and sequenced using an ABI 310 Automated DNA Sequencer and ABI PRISM Dye Terminator Cycle Sequencing Kit (Applied Biosystems, Carlsbad, CA, USA).
2.3. Construction of the lipoprotein maturation isogenic mutants
Precise in-frame deletion of lgt and lsp genes from strains 89-1591 were constructed using splicing-by-overlap-extension PCRs as previously described [24,25]. Overlapping PCR products were cloned into pCR2.1 (Invitrogen, Burlington, ON, Canada), extracted with EcoRI, recloned into the thermosensitive E. coli–S. suis shuttle plasmid pSET4s, and digested with the same enzyme, giving rise to the knockout vector p4Δlgt or p4Δlps. Electroporation of wild-type strain 89-1591 and procedures for isolation of the mutants were previously described [22]. Allelic replacement was confirmed by PCR and DNA sequencing analyses. Amplification products were purified with the QIAgen PCR Purification Kit (Qiagen) and sequenced as described above.
2.4. Complementation of the mutants
pMX1 vector was used for the generation of recombinant plasmids for complementation (Table 1). This vector is a derivative of the E. coli- S. suis shuttle cloning vector pSET2 [25] and possesses the S. suis malX promoter for transgene expression in S. suis. The entire lgt and lsp genes were amplified from genomic DNA of S. suis 89-1591 strain and cloned into pMX1 via EcoRI and NcoI sites, generating complementation vectors pMX1-lgt, and pMX1-lsp. These plasmids were introduced into E. coli MC1061 for verification of the sequences and then into the respective deletion mutants derived from S. suis 89-1591 to construct lgt- and lsp-complemented mutants.
2.5. Growth analysis
Overnight cultures of wild type and mutant strains were diluted in fresh THB or plasma and bacterial growth was followed during 24 h of incubation at 37°C. The total number of CFU/mL was evaluated at different incubation times.
2.6. Bacterial surface hydrophobicity assay
Relative surface hydrophobicity of the S. suis wild-type strain and its non-encapsulated mutant (positive control) was determined by measuring adsorption to n-hexadecane as previously described [14].
2.7. Bacterial adhesion and invasion assays using porcine brain microvascular endothelial and tracheal epithelial cells
The porcine brain microvascular endothelial cell line (PBMEC) and the neonatal porcine tracheal epithelial cell line (NPTr) were used and cultured until confluence as previously described [26,27]. Cells were infected with 1 x 106 CFU/well (multiplicity of infection or MOI = 10) of the different S. suis strains and incubated for 2 h at 37 °C in 5% CO2. The adhesion assay, which quantifies total cell-associated bacteria (surface-adherent and intracellular bacteria), and invasion assay (using the antibiotic protection assay) were performed as previously described [26,27].
2.8. Biofilm assay
Biofilm assay was performed as previously described [28]. Briefly, strains were growth in basal broth medium that contained 0.5% glucose, 2% peptone (proteose peptone no. 3; BBL Microbiology Systems), 0.3% K2HPO4, 0.2% KH2PO4, 0.01% MgSO4·7H2O, 0.002% MnSO4·6H2O, and 0.5% NaCl. Overnight cultures of S. suis were diluted in fresh culture broth to obtain an optical density at 655 nm (OD655) of 0.2. Samples (100 μl) were added to the wells of a Falcon® 96-well polystyrene tissue culture plate (Corning, NY, USA) containing 100 µL of culture medium. Biofilm formation capacity was determined as previously described in the absence or presence of 5 mg/mL of porcine fibrinogen (Sigma-Aldrich). The plates were incubated for 24 h at 37°C, and bacterial growth was evaluated by determining the OD655 using a microplate reader. Medium and free-floating bacteria were then removed, and biofilms were stained with crystal violet dye as previously described [28].
2.9. Generation of bone marrow-derived dendritic cells (bmDCs
The femur and tibia from C57BL/6J mice (Jackson Research Laboratories, Bar Harbour, MA, USA) were used to generate bmDCs, as previously described [19]. Briefly, hematopoietic bone marrow stem cells were cultured in complete culture medium (RPMI-1640 supplemented with 5% heat-inactivated fetal bovine serum, 10 mM HEPES, 2 mM L-glutamine, and 50 µM 2-mercaptoethanol (Gibco, Burlington, ON, Canada) and complemented with 20% granulocyte-macrophages colony-stimulating factor from mouse-transfected Ag8653 cells [29]. Cell purity was confirmed to be at least 90% CD11c+ by flow cytometry as previously described [19]. Albeit this culture system cannot completely rule out the presence of other innate cells such as macrophages, it represents an enriched source of bmDCs [15].
2.10. Preparation of heat-killed S. suis
Heat-killed S. suis suspensions were prepared as previously described [30]. Briefly, S. suis was cultured to mid-log phase and then incubated at 60°C for 45 min. Cultures were confirmed to be killed following culture on blood agar plates at 37°C for 48 h. Heat-killed S. suis were re-suspended in cell culture medium at a concentration equivalent to 2 x109 CFU/mL prior to bone-marrow dendritic cell stimulation. All manipulations were performed using endotoxin-free material (lipopolysaccharide).
2.11. Preparation of bacterial supernatant
Bacterial supernatants were prepared as previously described [7]. Briefly, bacteria were grown to mid-log phase (OD600nm = 0.6), and growth was immediately stopped on ice. Bacterial cultures were appropriately diluted and plated on THB agar (THA) to accurately determine bacterial concentrations. Then, bacteria were centrifuged for 15 min at 3312 x g at 4°C to separate bacteria from medium. Supernatants were collected and filtered using 0.2 µm filters. Supernatants were then applied to an Amicon ® Ultra-15 10K centrifugal filter and resuspended in cell culture medium prior to bone-marrow dendritic cell stimulation. Absence of bacteria in supernatants was confirmed after culture on blood agar plates at 37°C for 48 h. All manipulations were performed using endotoxin (lipopolysaccharide )-free material.
2.12. S. suis activation of bmDCs
Cells were activated with either live bacteria, killed bacteria or bacterial-free supernatant. All experiments were performed in the absence of endotoxin contamination and under non-toxic conditions, the latter being evaluated by the lactate dehydrogenase release with the CytoTox 96® Non-Radioactive Cytotoxicity Assay (Promega, Madison, WI, USA) [16]. Prior to infection, cells were resuspended at 1 x 106 cells/mL in complete medium and stimulated with the different live S. suis strains (1 x 106 CFU/mL; initial MOI = 1). Conditions used were based on those previously published [7,16]. A higher MOI (corresponding to 100 = 1, before inactivation) was used with killed bacteria. Supernatants were collected at 4 h, 6 h, 8 h, 12 h and 16 h following infection with S. suis (either live, heat-killed or bacterial-free supernatant). [16]. Mock-infected cells served as negative controls. Secreted levels of tumor necrosis factor (TNF), interleukin (IL)-6, C-C motif chemokine ligand (CCL)3, and C-X-C motif chemokine ligand (CXCL)1 were quantified by sandwich ELISA using pair-matched antibodies from R&D Systems (Minneapolis, MN, USA) according to the manufacturer’s recommendations.
2.13. S. suis virulence mouse model of systemic infection
A C57BL/6J mouse model of infection was used [31,32]. These studies were carried out in strict accordance with the recommendations of and approved by the University of Montreal Animal Welfare Committee guidelines and policies, including euthanasia to minimize animal suffering using humane endpoints, applied throughout this study when animals were seriously affected since mortality was not an endpoint measurement (permit number Rech-1570). Sixty 6-week-old female C57BL/6J (Jackson Research Laboratories) were used for these experiments (15 mice per group). Mice were inoculated with 5 × 107 CFU via the intraperitoneal route and health and behavior monitored at least thrice daily until 72 h post-infection and twice thereafter until the end of the experiment (12 days post-infection) for the development of clinical signs of sepsis, such as depression, swollen eyes, rough hair coat, prostration, and lethargy. A second confirmatory trial with the wild-type strain and its Δlsp mutant strain was repeated to confirm results, using 20 mice/group. For bacteremia studies, blood samples were collected from the caudal vein of surviving mice 12, 24 and 48 h post-infection and plated as previously described [32].
2.13. Measurement of plasma (systemic) pro-inflammatory mediators
In parallel (a separate experiment), eight mice per group were intraperitoneally infected with 5 x 107 CFU and blood collected 12 h post-infection by intracardiac puncture following euthanasia and anti-coagulated with EDTA (Sigma-Aldrich) as previously described [31,32]. Plasma supernatants were collected following centrifugation at 10 000 x g for 10 min at 4 ºC and stored at -80 ºC. The 12 h post-infection time point was selected to obtain maximal pro-inflammatory mediator production in the absence of significant mouse mortality as previously shown [32]. Plasmatic concentrations of IL-6, G-CSF, CCL2, CCL3, CCL4, CCL5, CXCL2 and CXCL9 were measured using a custom-made cytokine Bio-Plex Pro™ assay (Bio-Rad, Hercules, CA, USA) according to the manufacturer’s instructions. Acquisition was performed on the MAGPIX platform (Luminex®) and data analyzed using the Bio-Plex Manager 6.1 software (Bio-Rad).
2.14. Statistical analyses
Normality of data was verified using the Shapiro-Wilk test. Accordingly, parametric (unpaired t-test) or non-parametric tests (Mann-Whitney rank sum test), where appropriate, were performed to evaluate statistical differences between groups. Log-rank test was used to compare survival rates between wild-type-infected mice and those infected mutant strains. Each in vitro test was repeated in at least three independent experiments. p < 0.05 was considered as statistically significant.
3. Results
3.1. Characteristics of the Δlgt and Δlsp mutants derived from the ST25 89-1591 strain
The Δlgt and Δlsp mutants presented general phenotypic characteristics similar to ST25 wild-type strains. Both mutants remained well encapsulated as shown by the presence of hydrophobicity values between 4.9% and 6.1%, similar to the wild–type strain (4.9%) (Figure 1A). In addition, both mutants remained similarly serotypable (serotype 2) by the co-agglutination test (data not shown). Finally, the ST25 wild-type and Δlgt and Δlsp strains cultured in plasma (Figure 1B) and in rich THB medium (not shown) presented similar good growth rate, as evaluated by bacterial counts.
3.2. Lack of lipoprotein maturation enzymes does not impair adhesion to and invasion of respiratory epithelial and brain microvascular endothelial swine cells regardless of the sequence type of the strain
We previously described that no differences were observed between the adhesion and invasion capacity of epithelial cells when ST1 and ST7 wild-type strains were compared to their respective lipoprotein maturation enzyme defective mutants [7]. However, since the cell-wall components of the ST25 strain are better surface exposed [14], LPPs may interplay differently with the host cells than ST1 and ST7 strains. Thus, the role of the Lgt and Lsp enzymes in the S. suis adhesion/invasion to NPTr and PBMEC was evaluated with the ST25 strain and its mutants. Interactions of the ST1 P1/7 strain (and the respective Δlgt and Δlsp mutants) with endothelial cells were also tested for the first time in the current study. For both cell types, and as expected, the non-encapsulated mutant strains (ST1 and ST25, positive controls) significantly adhered and invaded cells more efficiently than their respective wild-type strains (Figure 2A-F). Concerning the epithelial cells, no differences were observed between the adhesion and invasion capacity the ST25 wild type strain and defective Lgt or Lsp mutants (Figure 2C,F). Similar results were obtained with PBMEC cells (Figure 2A, B, D, E). The ST1 P1/7 strain and its derived Δlgt and Δlsp mutants behaved similarly to the ST25 strains (Figure 2A-F). These results indicate that the Lgt and Lsp enzymes do not play a critical role in bacterial cell adhesion/invasion regardless of strain background and cell types tested.
3.3. Lgt and Lsp enzymes are important for S. suis biofilm formation regardless of the sequence type of the strain
Biofilm formation by pathogenic microorganisms is a mechanism that allows them to become persistent colonizers and resist clearance by the innate and adaptive host immune system [28,33]. Since the involvement of LPPs in S. suis biofilm formation has been suggested [34], the ST1 and ST25 wild-type strains and their respective Δlgt and Δlsp mutant were tested using the crystal violet assay. Results confirmed that the wild-type ST1 strain produced more biofilm than the wild-type ST25 (Figure 3A,B) [35]. On the other hand, the Δlgt and Δlsp mutants of both strains showed a significantly lower capacity to form biofilm than their respective ST1 and ST25 wild-type strains (Figure 3A,B). The capacity to form biofilm was significantly restored in the complemented strains. Consequently, the LPP maturation enzymes play a role in the capacity to form biofilm regardless of strain background.
3.4. The diacyl motif and the peptide signal cleavage are important for the recognition by innate immune cells of periplasmic and/or secreted lipoproteins of S. suis serotype 2 ST25 strain
bmDC cells were used as an innate immune cell model given their critical role they play during S. suis pathogenesis; in addition, their strong inflammatory response to S. suis activation has been well characterized [16,19]. In our previous study with ST1 and ST7 strains, we noticed the importance of the diacyl motif for the recognitions of membranes and secreted LPPs, and the peptide signal cleavage were shown to be similarly important for the recognition of secreted LPPs [7]. To investigate if Lgt and Lsp enzymes of S. suis serotype 2 ST25 play a similar role, cells were activated for up to 16 h with either live bacteria (for evaluation of the role of periplasmic and possibly secreted LPPs) (Figure 4), heat-killed bacteria (for the evaluation of strict periplasmic LPPs) (Figure 5), or bacterial-free supernatant, to study the potential activation of strictly secreted LPPs (Figure 6).
For all experiments and all incubation times, control mock infected cells presented negligible cytokine level values < 300 pg/mL (not shown). When using live bacteria, the ST25 Δlgt mutant (but not the Δlsp mutant) induced lower levels of the different pro-inflammatory mediators TNF, IL6, CXCL1 and CCL3 during, at least, 16 h of activation (Figure 4). Similar results were observed using washed heat-killed bacteria (Figure 5). Moreover, both the diacyl motif and the peptide signal cleavage were shown to be similarly important for the recognition of secreted LPPs up to 16 h of incubation (Figure 6). In all cases, complemented mutants restored the activation capacity of the strains, confirming the influence of the gene deletion (Figure 4, Figure 5 and Figure 6).
3.5. The absence of Lgt enzyme, but not Lsp, significantly affects the virulence of S. suis serotype 2 ST25
To confirm the role of Lgt and/or Lps in the ST25 strain on virulence and development of clinical disease, a well-characterized C56BL/6 mouse model of infection was used. Mouse survival was better with mutant strains when compared to the wild-type strain. Surprisingly, only the absence of the Lgt, but not the Lsp, significantly affect the strain virulence (p < 0.05) (Figure 7A). Blood bacterial burden was evaluated at the early infection times of 12 h (Figure 7B), 24 h (Figure 7C) and 48 h (not shown) post-infection and no differences were observed in the acute blood bacterial burden between mice infected for the wild-type or mutant strains.
3.6. Absence of the Lgt or Lsp enzymes significantly reduces the in vivo inflammatory response of mice infected with either the wild type S. suis serotype 2 strain ST25 or its respective Δlgt or Δlsp mutant
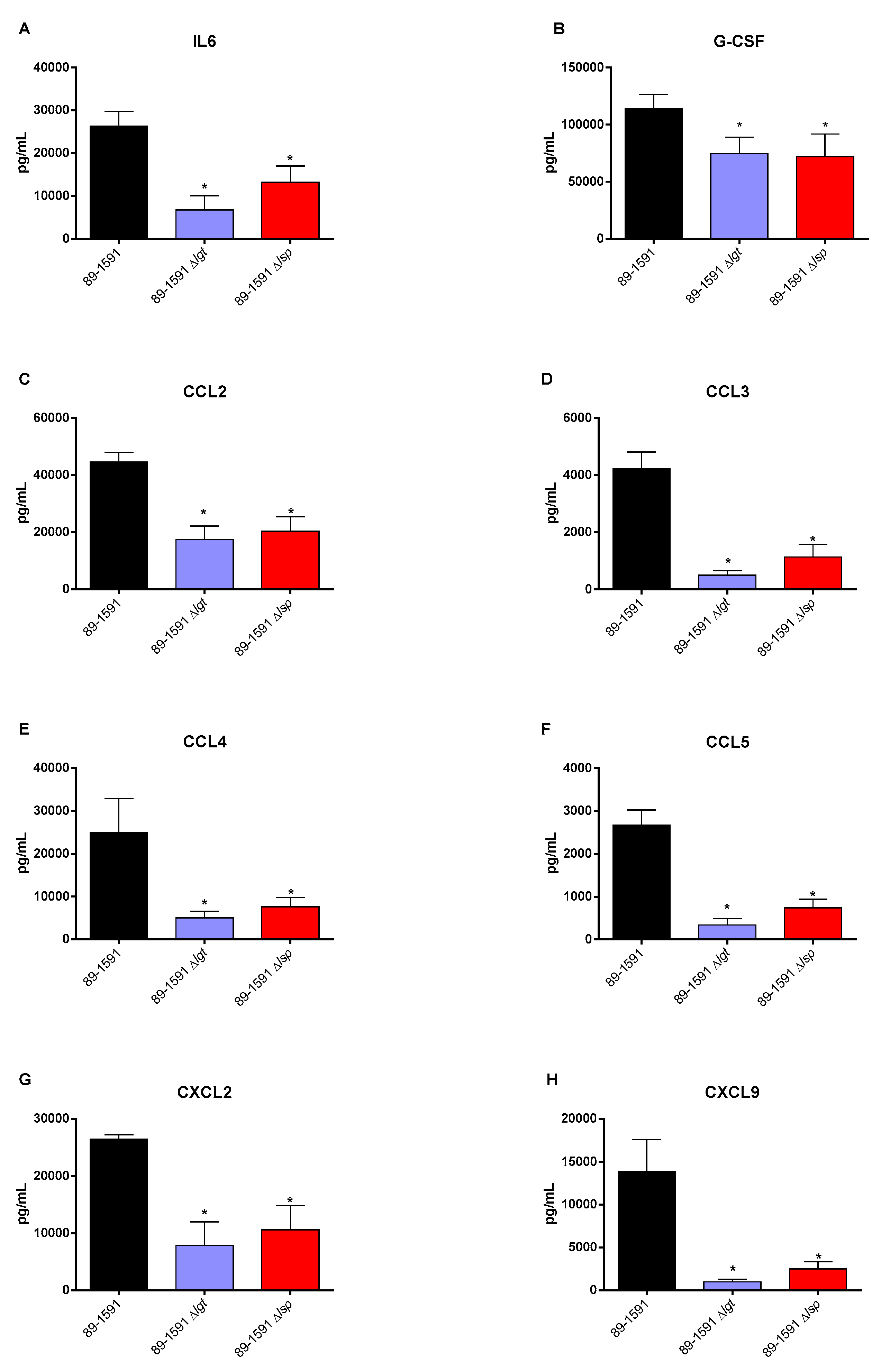
The deletion of LPP maturation enzymes genes was previously described to be implicated in S. suis recognition by the immune system that may result in an exacerbated systemic inflammatory response in Eurasian ST1 and ST7 strains [7]. Consequently, the effect of lgt and lsp gene deletion on the inflammatory response of animals infected with the ST25 wild-type and mutant strains, plasma mediators were evaluated after 12 h of infection. Concentrations of the all mediators tested (IL-6, IL-12p70, G-CSF, IFN-γ, CCL2, CCL3, CCL4, CCL5, CXCL2 and CXCL9) were significantly lower in mice infected with both mutants when compared to their respective ST25 wild-type strain.
Figure 8.
Lgt and Lsp enzymes are important for the recognition of lipoproteins and therefore for the establishment of the inflammatory response. Plasma levels of IL-6 (A), G-CSF (B), CCL2 (C), CCL3 (D), CCL4 (E), CCL5 (F), CXCL9 (G) and CXCL2 (H) in mice 12 h following intraperitoneal inoculation of S. suis serotype 2 wild-type ST25 strain 89-1591 (black) or its Δlgt (blue) and Δlsp (red) mutant strains. Data represent mean + SEM (n = 8 individuals). * p < 0.05 indicates a significant difference between plasma levels of mice infected with the mutant strains when compared to those infected with the wild-type strain.
Figure 8.
Lgt and Lsp enzymes are important for the recognition of lipoproteins and therefore for the establishment of the inflammatory response. Plasma levels of IL-6 (A), G-CSF (B), CCL2 (C), CCL3 (D), CCL4 (E), CCL5 (F), CXCL9 (G) and CXCL2 (H) in mice 12 h following intraperitoneal inoculation of S. suis serotype 2 wild-type ST25 strain 89-1591 (black) or its Δlgt (blue) and Δlsp (red) mutant strains. Data represent mean + SEM (n = 8 individuals). * p < 0.05 indicates a significant difference between plasma levels of mice infected with the mutant strains when compared to those infected with the wild-type strain.
4. Discussion
Evidence accumulated over the years demonstrated a high level of genetic diversity in the species S. suis [2]. However, most studies on the pathogenesis of the infection caused by S. suis serotype 2 have used Eurasian strains (virulent ST1 and/or ST7), which greatly differ from their North American counterparts, including ST25 strains. The pathogenesis of the infection seems to present marked difference between ST25 and ST1/ST7 strains. For example, the suilysin, a virulence factor present in ST1/ST7 strains [36], is not produced by ST25 strains [37]. In addition, mechanisms of production of type I interferon and interleukin-1 during S. suis infection is also strain-dependent (ST1 vs ST7 vs ST25). Interaction of ST25 strains with phagocytic cells also differs from those of other STs [14]. We recently studied the role of Lgt and Lsp in Eurasian strains (ST1 and ST7) and results suggest these enzymes also play a differential role in virulence depending on the genetic background of the strain [7]. Since it has been reported that the CPS only partially masks sub-capsular domains and bacterial wall components of North American S. suis serotype 2 ST25 strains, we evaluated the role of LPP maturation enzymes in different aspects of the pathogenesis of the infection caused by the North American type [14].
Since several LPPs are known for being substrate-binding proteins of ABC transporter systems responsible for the acquisition of multiple nutrients [13], the maturation enzymes may play a role on bacterial growth under different conditions. For example, Lgt has been shown to be important for Staphylococcus aureus and Streptococcus pneumoniae growth in poor medium or in vivo-like conditions [38,39]. Likewise, the growth of Lsp mutants was impaired in rich medium in S. pneumoniae and Listeria monocytogenes [40,41]. However, similarly to what has been described for ST1/ST7 strains, we showed that growth in both rich medium and in vivo-like conditions (plasma) of Δlgt and Δlsp in ST25 mutant strain was not significantly affected. These results indicate that LPP maturation enzymes are not important for growth of S. suis serotype 2 independently of the ST.
Results from our last study indicate that adhesion and invasion capacity of S. suis to epithelial cells of ST1 and ST7 mutants defective in LPP maturation enzymes was not affected [7]; similar results have been obtained with the ST25 strain in the current study. Likewise, the absence of these enzymes does not affect S. suis adhesion/invasion to brain endothelial cells and this, for the ST1 and ST25 strains. These results with different cells reinforce the concept that LPPs do probably not play important roles in the S. suis interactions with host cells. Another hypothesis includes the possibility that a lack of LPP maturation does not completely eliminate the functional activities of such proteins, as shown for Streptococcus equi [42]. Finally, since all our studies have been carried out with serotype 2 strains, a hypothetical role of the LPP maturation enzymes on bacterial-cell interactions in other S. suis serotypes cannot be ruled out. Previous studies showed that a single protein could play important or limited roles during the first steps of the pathogenesis of the infection depending on the serotype [43,44].
Alongside adhesion and invasion to host cells, the capacity to form biofilm has been described as being important for pathogenesis of the infection caused by different pathogens, including S. suis [26,45]. Studies on the role of Lgt and/or Lsp on biofilm formation by streptococci and other pathogens are scarce. It has been reported that LPPs (such as VacJ) plays an important role on biofilm formation by the swine pathogens Glaesserella parasuis and Actinobacillus pleuropneumoniae [46,47]. However, the role of LPPs in biofilm formation is not always clear. Although the LPP AdcA has been implicated in biofilm formation by Streptococcus gordonii [48], another study showed that Lgt negatively regulate biofilm formation for this bacterial species [49]. Similarly, in E. coli, Rcs pathway through the outer membrane LPP RcsF reduce biofilm formation [50]. Our results showed that the absence of the Lgt and Lsp leads to a decrease biofilm formation for both ST1 and ST25 strains, suggesting that LPPs are indeed involved in such process. Accordingly, AdcACB and Lmb, two zinc-binding LPPs, have already been described as implicated in S. suis biofilm formation [34].
Studies previously reported that Lgt and Lsp in S. suis ST1 and ST7 strains are critical for the induction of an inflammatory response in vitro and in vivo, especially Lgt [7,51]. The diacylglycerol provided by the Lgt enzyme is the main motif recognized by the immune system in Gram-positive bacteria [52]. Both enzymes have been described as being critical for cell activation in other streptococci [11,39,42,53]. In the current study, we confirmed that lack of LPP maturation enzymes affects cell activation by the North American ST25 strain. Indeed, the role of such enzymes seems to be enhanced with this strain. In the previous study with ST1 and ST7 strains, levels of the different pro-inflammatory mediators were affected only at early incubation times when the mutant strains were tested [7]. The reduction of cytokine expression using the ST25 Δlgt and/or Δlsp mutants was clearly observed at least up to 16h of incubation (maximum incubation time tested in the current study). Indeed, as mentioned, there are important phenotypic differences between ST1/ST7 and ST25 strains, and putative virulence protein factors, such as the suilysin, the muramidase-released protein and the extracellular protein factor, are present in the former strains only [44,54]. Even if their role in the pathogenesis of S. suis is not totally elucidated and they are not critical for the virulence, it is possible that these markers and other factors present in ST1/ST7 strains but absent in ST25 strains, compensate for further cell activation [36]. Indeed, other surface or secreted proteins not regulated by the Lgt and Lsp enzymes are also cell activators [43,55]. It is important to mention that differences in cell activation by the mutant strains tested in closed in vitro systems do not always represent in vivo activation. In fact, both mutants induced significantly less pro-inflammatory cytokines than the wild-type strain after in vivo infection.
It has been previously described that the role of the LPP maturation enzymes on virulence was dependent of the strain background; indeed, the absence of the Lgt or Lsp enzymes reduced the virulence of a ST7 (but not that of a ST1) S. suis strain [7]. In the case of the S. suis serotype 2 ST25, only the Δlgt mutant was significantly less virulent when compared to the wild-type strain, although the Δlsp mutant present a clear tendency to also be less virulent (p = 0.07). Similar results were obtained when the infection was repeated (results not shown). Reasons for the differences observed between the STs are not completely understood, although the hypothesis that the three strains may use different pathogenic mechanisms can be postulated [15,56]. The reduced virulence could be attributed, as mentioned above, to a lesser inflammatory reaction, since animals infected with any of the mutant strains induce significantly less pro-inflammatory cytokines. In addition, similar to what has been observed for ST1 and ST7 strains, the absence of the LPP maturation did not seem to affect bacterial survival in blood, indicating that the better mouse survival was not due to a reduced bacteremia. It can be concluded that LPPs maturation is implicated in S. suis recognition by the immune system independently of the strain background. Finally, it should be considered that the infection used in the mouse model of systemic infection is five time higher with ST25 strain than that for ST1 and ST7 strains (due to a lower virulence potential). Although final mortality rates obtained are similar, the comparison with other STs may be somehow biased.
5. Conclusions
Overall, ours studies suggest that LPP maturation enzymes do not seem to play a role in the interaction between S. suis and epithelial and endothelial cells, regardless the genetics background of the serotype 2 strain used. However, a potential involvement of LPPs in the formation of biofilms (also independently of the STs) has been demonstrated. Moreover, the involvement of such enzymes in dendritic cell activation in vitro seems to be more pronounced with the ST25 strain. Finally, the Lgt enzyme seems to play a more important role in the virulence of the ST25 strain. Although some differences between STs could be observed, our original hypothesis that LPP maturation enzymes would be more important in ST25 strains due to a better bacterial surface exposition could not be confirmed.
6. Patents
Author Contributions
Conceptualization, M.G., S.P. and M.S.; methodology, S.P., M.G., D.R., M.O. and M.S.; writing—original draft preparation, S.P., M.G. and M.S.; writing—review and editing, S.P., M.G., and M.S.; supervision, M.G. and M.S.; project administration, M.G.; funding acquisition, M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) (04435 to M.G.).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of the University of Montreal (protocol code 1570, 12 December 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Sonia Lacouture (laboratory of M.G.) for technical help and advice.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? Future Microbiol 2010, 5, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg Microbes Infect 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg Infect Dis 2006, 12, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Berthelot-Herault, F.; Gottschalk, M.; Morvan, H.; Kobisch, M. Dilemma of virulence of Streptococcus suis: Canadian isolate 89-1591 characterized as a virulent strain using a standardized experimental model in pigs. Can J Vet Res 2005, 69, 236–240. [Google Scholar]
- Lacouture, S.; Olivera, Y.R.; Mariela, S.; Gottschalk, M. Distribution and characterization of Streptococcus suis serotypes isolated from January 2015 to June 2020 from diseased pigs in Quebec, Canada. Can J Vet Res 2022, 86, 78–82. [Google Scholar] [PubMed]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol 2012, 7, 259–279. [Google Scholar] [CrossRef]
- Payen, S.; Roy, D.; Boa, A.; Okura, M.; Auger, J.P.; Segura, M.; Gottschalk, M. Role of maturation of lipoproteins in the pathogenesis of the infection caused by Streptococcus suis serotype 2. Microorganisms 2021, 9, 2386. [Google Scholar] [CrossRef]
- Kovacs-Simon, A.; Titball, R.W.; Michell, S.L. Lipoproteins of bacterial pathogens. Infect Immun 2011, 79, 548–561. [Google Scholar] [CrossRef]
- Kohler, S.; Voss, F.; Gomez Mejia, A.; Brown, J.S.; Hammerschmidt, S. Pneumococcal lipoproteins involved in bacterial fitness, virulence, and immune evasion. FEBS Lett 2016, 590, 3820–3839. [Google Scholar] [CrossRef]
- Sander, P.; Rezwan, M.; Walker, B.; Rampini, S.K.; Kroppenstedt, R.M.; Ehlers, S.; Keller, C.; Keeble, J.R.; Hagemeier, M.; Colston, M.J.; et al. Lipoprotein processing is required for virulence of Mycobacterium tuberculosis. Mol Microbiol 2004, 52, 1543–1552. [Google Scholar] [CrossRef]
- Henneke, P.; Dramsi, S.; Mancuso, G.; Chraibi, K.; Pellegrini, E.; Theilacker, C.; Hubner, J.; Santos-Sierra, S.; Teti, G.; Golenbock, D.T.; et al. Lipoproteins are critical TLR2 activating toxins in group B streptococcal sepsis. J Immunol 2008, 180, 6149–6158. [Google Scholar] [CrossRef]
- Hashimoto, M.; Tawaratsumida, K.; Kariya, H.; Kiyohara, A.; Suda, Y.; Krikae, F.; Kirikae, T.; Gotz, F. Not lipoteichoic acid but lipoproteins appear to be the dominant immunobiologically active compounds in Staphylococcus aureus. J Immunol 2006, 177, 3162–3169. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Palmer, T.; Harrington, D.J.; Sutcliffe, I.C. Lipoprotein biogenesis in Gram-positive bacteria: knowing when to hold 'em, knowing when to fold 'em. Trends Microbiol 2009, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Dolbec, D.; Roy, D.; Segura, M.; Gottschalk, M. Role of the Streptococcus suis serotype 2 capsular polysaccharide in the interactions with dendritic cells is strain-dependent but remains critical for virulence. PLoS One 2018, 13, e0200453. [Google Scholar] [CrossRef] [PubMed]
- Lavagna, A.; Auger, J.P.; Girardin, S.E.; Gisch, N.; Segura, M.; Gottschalk, M. Recognition of lipoproteins by Toll-like receptor 2 and DNA by the AIM2 inflammasome is responsible for production of interleukin-1beta by virulent suilysin-negative Streptococcus suis serotype 2. Pathogens 2020, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Lecours, M.P.; Segura, M.; Fittipaldi, N.; Rivest, S.; Gottschalk, M. Immune receptors involved in Streptococcus suis recognition by dendritic cells. PLoS One 2012, 7, e44746. [Google Scholar] [CrossRef]
- Gottschalk, M.; Higgins, R.; Boudreau, M. Use of polyvalent coagglutination reagents for serotyping of Streptococcus suis. J Clin Microbiol 1993, 31, 2192–2194. [Google Scholar] [CrossRef]
- Slater, J.D.; Allen, A.G.; May, J.P.; Bolitho, S.; Lindsay, H.; Maskell, D.J. Mutagenesis of Streptococcus equi and Streptococcus suis by transposon Tn917. Vet Microbiol 2003, 93, 197–206. [Google Scholar] [CrossRef]
- Lecours, M.P.; Gottschalk, M.; Houde, M.; Lemire, P.; Fittipaldi, N.; Segura, M. Critical role for Streptococcus suis cell wall modifications and suilysin in resistance to complement-dependent killing by dendritic cells. J Infect Dis 2011, 204, 919–929. [Google Scholar] [CrossRef]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Dubreuil, D. Production and characterization of two Streptococcus suis capsular type 2 mutants. Vet Microbiol 1992, 30, 59–71. [Google Scholar] [CrossRef]
- Casadaban, M.J.; Cohen, S.N. Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J Mol Biol 1980, 138, 179–207. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 2001, 46, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Okura, M.; Osaki, M.; Fittipaldi, N.; Gottschalk, M.; Sekizaki, T.; Takamatsu, D. The minor pilin subunit Sgp2 is necessary for assembly of the pilus encoded by the srtG cluster of Streptococcus suis. J Bacteriol 2011, 193, 822–831. [Google Scholar] [CrossRef]
- Warrens, A.N.; Jones, M.D.; Lechler, R.I. Splicing by overlap extension by PCR using asymmetric amplification: an improved technique for the generation of hybrid proteins of immunological interest. Gene 1997, 186, 29–35. [Google Scholar] [CrossRef]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Construction and characterization of Streptococcus suis-Escherichia coli shuttle cloning vectors. Plasmid 2001, 45, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gagnon, C.A.; Savard, C.; Music, N.; Srednik, M.; Segura, M.; Lachance, C.; Bellehumeur, C.; Gottschalk, M. Capsular sialic acid of Streptococcus suis serotype 2 binds to swine influenza virus and enhances bacterial interactions with virus-infected tracheal epithelial cells. Infect Immun 2013, 81, 4498–4508. [Google Scholar] [CrossRef]
- Vanier, G.; Segura, M.; Friedl, P.; Lacouture, S.; Gottschalk, M. Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infect Immun 2004, 72, 1441–1449. [Google Scholar] [CrossRef]
- Bonifait, L.; Grignon, L.; Grenier, D. Fibrinogen induces biofilm formation by Streptococcus suis and enhances its antibiotic resistance. Appl Environ Microbiol 2008, 74, 4969–4972. [Google Scholar] [CrossRef]
- Segura, M.; Su, Z.; Piccirillo, C.; Stevenson, M.M. Impairment of dendritic cell function by excretory-secretory products: a potential mechanism for nematode-induced immunosuppression. Eur J Immunol 2007, 37, 1887–1904. [Google Scholar] [CrossRef]
- Segura, M.; Stankova, J.; Gottschalk, M. Heat-killed Streptococcus suis capsular type 2 strains stimulate tumor necrosis factor alpha and interleukin-6 production by murine macrophages. Infect Immun 1999, 67, 4646–4654. [Google Scholar] [CrossRef]
- Lachance, C.; Gottschalk, M.; Gerber, P.P.; Lemire, P.; Xu, J.; Segura, M. Exacerbated type II interferon response drives hypervirulence and toxic shock by an emergent epidemic strain of Streptococcus suis. Infect Immun 2013, 81, 1928–1939. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Fittipaldi, N.; Benoit-Biancamano, M.O.; Segura, M.; Gottschalk, M. Virulence studies of different sequence types and geographical origins of Streptococcus suis serotype 2 in a mouse model of infection. Pathogens 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Xu, Y.; Dou, B.; Yang, F.; He, Q.; Liu, Z.; Gao, T.; Liu, W.; Yang, K.; Guo, R.; et al. The adcA and lmb genes play an important role in drug resistance and full virulence of Streptococcus suis. Microbiol Spectr 2023, 11, e0433722. [Google Scholar] [CrossRef]
- Auger, J.P.; Chuzeville, S.; Roy, D.; Mathieu-Denoncourt, A.; Xu, J.; Grenier, D.; Gottschalk, M. The bias of experimental design, including strain background, in the determination of critical Streptococcus suis serotype 2 virulence factors. PLoS One 2017, 12, e0181920. [Google Scholar] [CrossRef]
- Tenenbaum, T.; Asmat, T.M.; Seitz, M.; Schroten, H.; Schwerk, C. Biological activities of suilysin: role in Streptococcus suis pathogenesis. Future Microbiol 2016, 11, 941–954. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Fuller, T.E.; Teel, J.F.; Wilson, T.L.; Wolfram, T.J.; Lowery, D.E.; Gottschalk, M. Serotype distribution and production of muramidase-released protein, extracellular factor and suilysin by field strains of Streptococcus suis isolated in the United States. Vet Microbiol 2009, 139, 310–317. [Google Scholar] [CrossRef]
- Stoll, H.; Dengjel, J.; Nerz, C.; Gotz, F. Staphylococcus aureus deficient in lipidation of prelipoproteins is attenuated in growth and immune activation. Infect Immun 2005, 73, 2411–2423. [Google Scholar] [CrossRef]
- Chimalapati, S.; Cohen, J.M.; Camberlein, E.; MacDonald, N.; Durmort, C.; Vernet, T.; Hermans, P.W.; Mitchell, T.; Brown, J.S. Effects of deletion of the Streptococcus pneumoniae lipoprotein diacylglyceryl transferase gene lgt on ABC transporter function and on growth in vivo. PLoS One 2012, 7, e41393. [Google Scholar] [CrossRef]
- Pribyl, T.; Moche, M.; Dreisbach, A.; Bijlsma, J.J.; Saleh, M.; Abdullah, M.R.; Hecker, M.; van Dijl, J.M.; Becher, D.; Hammerschmidt, S. Influence of impaired lipoprotein biogenesis on surface and exoproteome of Streptococcus pneumoniae. J Proteome Res 2014, 13, 650–667. [Google Scholar] [CrossRef]
- Reglier-Poupet, H.; Frehel, C.; Dubail, I.; Beretti, J.L.; Berche, P.; Charbit, A.; Raynaud, C. Maturation of lipoproteins by type II signal peptidase is required for phagosomal escape of Listeria monocytogenes. J Biol Chem 2003, 278, 49469–49477. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Robinson, C.; Sutcliffe, I.C.; Slater, J.; Maskell, D.J.; Davis-Poynter, N.; Smith, K.; Waller, A.; Harrington, D.J. Mutation of the maturase lipoprotein attenuates the virulence of Streptococcus equi to a greater extent than does loss of general lipoprotein lipidation. Infect Immun 2006, 74, 6907–6919. [Google Scholar] [CrossRef] [PubMed]
- Chuzeville, S.; Auger, J.P.; Dumesnil, A.; Roy, D.; Lacouture, S.; Fittipaldi, N.; Grenier, D.; Gottschalk, M. Serotype-specific role of antigen I/II in the initial steps of the pathogenesis of the infection caused by Streptococcus suis. Vet Res 2017, 48, 39. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis virulence factors: are they all really critical? Trends Microbiol 2017, 25, 585–599. [Google Scholar] [CrossRef]
- Segura, M.; Calzas, C.; Grenier, D.; Gottschalk, M. Initial steps of the pathogenesis of the infection caused by Streptococcus suis: fighting against nonspecific defenses. FEBS Lett 2016, 590, 3772–3799. [Google Scholar] [CrossRef]
- Zhao, L.; Gao, X.; Liu, C.; Lv, X.; Jiang, N.; Zheng, S. Deletion of the vacJ gene affects the biology and virulence in Haemophilus parasuis serovar 5. Gene 2017, 603, 42–53. [Google Scholar] [CrossRef]
- Xie, F.; Li, G.; Zhang, W.; Zhang, Y.; Zhou, L.; Liu, S.; Liu, S.; Wang, C. Outer membrane lipoprotein VacJ is required for the membrane integrity, serum resistance and biofilm formation of Actinobacillus pleuropneumoniae. Vet Microbiol 2016, 183, 1–8. [Google Scholar] [CrossRef]
- Mitrakul, K.; Loo, C.Y.; Gyurko, C.; Hughes, C.V.; Ganeshkumar, N. Mutational analysis of the adcCBA genes in Streptococcus gordonii biofilm formation. Oral Microbiol Immunol 2005, 20, 122–127. [Google Scholar] [CrossRef]
- Park, O.J.; Jung, S.; Park, T.; Kim, A.R.; Lee, D.; Jung Ji, H.; Seong Seo, H.; Yun, C.H.; Hyun Han, S. Enhanced biofilm formation of Streptococcus gordonii with lipoprotein deficiency. Mol Oral Microbiol 2020, 35, 271–278. [Google Scholar] [CrossRef]
- Nepper, J.F.; Lin, Y.C.; Weibel, D.B. Rcs phosphorelay activation in cardiolipin-deficient Escherichia coli reduces biofilm formation. J Bacteriol 2019, 201, e00804–18. [Google Scholar] [CrossRef]
- Wichgers Schreur, P.J.; Rebel, J.M.; Smits, M.A.; van Putten, J.P.; Smith, H.E. Differential activation of the Toll-like receptor 2/6 complex by lipoproteins of Streptococcus suis serotypes 2 and 9. Vet Microbiol 2010, 143, 363–370. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Gotz, F. Lipoproteins of Gram-positive bacteria: key players in the immune response and virulence. Microbiol Mol Biol Rev 2016, 80, 891–903. [Google Scholar] [CrossRef]
- Petit, C.M.; Brown, J.R.; Ingraham, K.; Bryant, A.P.; Holmes, D.J. Lipid modification of prelipoproteins is dispensable for growth in vitro but essential for virulence in Streptococcus pneumoniae. FEMS Microbiol Lett 2001, 200, 229–233. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Xu, J.; Lacouture, S.; Tharavichitkul, P.; Osaki, M.; Sekizaki, T.; Takamatsu, D.; Gottschalk, M. Lineage and virulence of Streptococcus suis serotype 2 isolates from North America. Emerg Infect Dis 2011, 17, 2239–2244. [Google Scholar] [CrossRef] [PubMed]
- Baums, C.G.; Valentin-Weigand, P. Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim Health Res Rev 2009, 10, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Lavagna, A.; Auger, J.P.; Dumesnil, A.; Roy, D.; Girardin, S.E.; Gisch, N.; Segura, M.; Gottschalk, M. Interleukin-1 signaling induced by Streptococcus suis serotype 2 is strain-dependent and contributes to bacterial clearance and inflammation during systemic disease in a mouse model of infection. Vet Res 2019, 50, 52. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Bacterial hydrophobicity is not affected by the absence of Lgt or Lsp and both defective mutants grow as well as the wild type strain in plasma. Surface hydrophobicity using n-hexadecane (A) and bacterial growth in plasma (B) of S. suis serotype 2 wild-type 89-1591 (ST25) strain, Δlgt (blue) and Δlsp (red) mutants (n = 3 independent repetitions for each experiment). The non-encapsulated 89-1591 ΔcpsF was used as a positive control for the hydrophobicity test.
Figure 1.
Bacterial hydrophobicity is not affected by the absence of Lgt or Lsp and both defective mutants grow as well as the wild type strain in plasma. Surface hydrophobicity using n-hexadecane (A) and bacterial growth in plasma (B) of S. suis serotype 2 wild-type 89-1591 (ST25) strain, Δlgt (blue) and Δlsp (red) mutants (n = 3 independent repetitions for each experiment). The non-encapsulated 89-1591 ΔcpsF was used as a positive control for the hydrophobicity test.
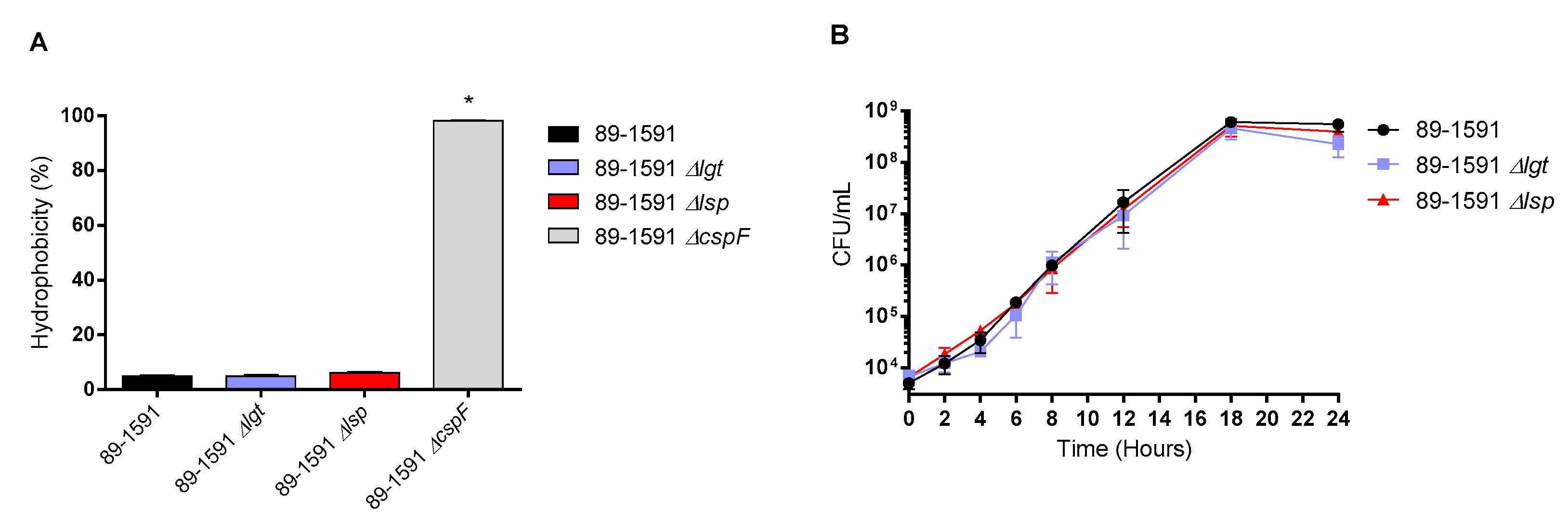
Figure 2.
The lack of lipoprotein maturation enzymes does not change the capacity of adhesion and invasion of S. suis to swine epithelial and endothelial cells. Adhesion (A) and invasion (B) of the S. suis 2 wild-type strain 89-1591 (ST25) as well as their respeective Δlgt (blue) and Δlsp (red) mutant strains to swine tracheal epithelial cells. Adhesion and invasion of the S. suis 2 wild-type P1/7 (ST1) strain (C, D) and 89-1591 (ST25) strain (E, F) as well as their respective Δlgt (blue) and Δlsp (red) mutant strains to brain microvascular endothelial cells. * Indicates a significant difference (p < 0.05). Each bar represents the mean bacterial concentration (CFU/mL) + SEM from at least three independent experiments.
Figure 2.
The lack of lipoprotein maturation enzymes does not change the capacity of adhesion and invasion of S. suis to swine epithelial and endothelial cells. Adhesion (A) and invasion (B) of the S. suis 2 wild-type strain 89-1591 (ST25) as well as their respeective Δlgt (blue) and Δlsp (red) mutant strains to swine tracheal epithelial cells. Adhesion and invasion of the S. suis 2 wild-type P1/7 (ST1) strain (C, D) and 89-1591 (ST25) strain (E, F) as well as their respective Δlgt (blue) and Δlsp (red) mutant strains to brain microvascular endothelial cells. * Indicates a significant difference (p < 0.05). Each bar represents the mean bacterial concentration (CFU/mL) + SEM from at least three independent experiments.
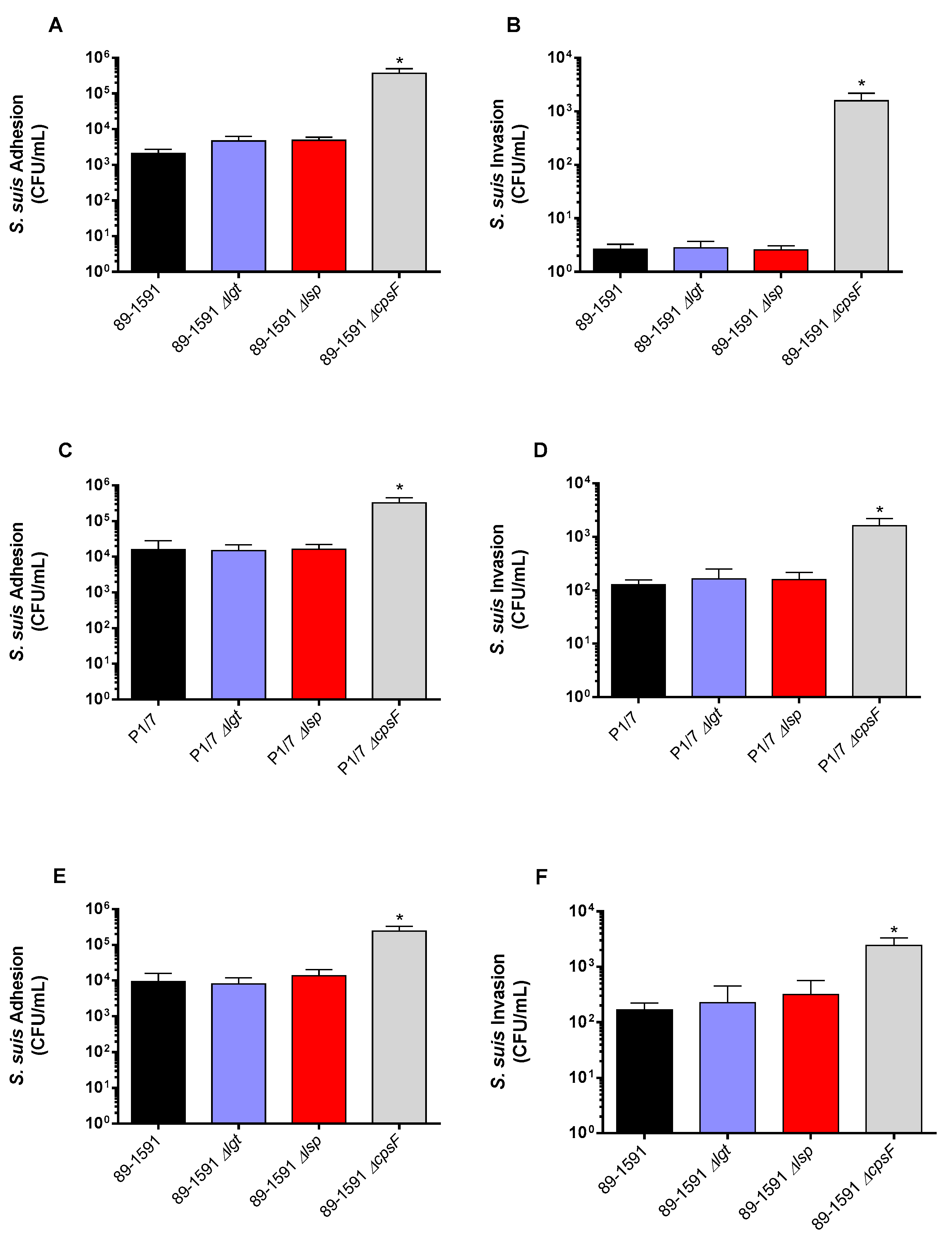
Figure 3.
The Lgt and Lsp enzymes are both implicated in biofilm formation by S. suis independently of the ST of the strain. Biofilm formation capacity in the presence of porcine fibrinogen after 24 h of incubation at 37 °C of S. suis serotype 2 wild-type strain P1/7 (ST1) (A) and strain 89-1591 (ST25) (B) as well as their respective Δlgt (blue), Δlsp (red) mutant, the 89-1591 comp Δlgt (dark blue) and 89-1591 comp Δlsp complemented strains (dark red). strains. Data represent the mean ± SEM from at least three independent experiments. * Indicates a significant difference with the respective wild-type strain (p < 0.05).
Figure 3.
The Lgt and Lsp enzymes are both implicated in biofilm formation by S. suis independently of the ST of the strain. Biofilm formation capacity in the presence of porcine fibrinogen after 24 h of incubation at 37 °C of S. suis serotype 2 wild-type strain P1/7 (ST1) (A) and strain 89-1591 (ST25) (B) as well as their respective Δlgt (blue), Δlsp (red) mutant, the 89-1591 comp Δlgt (dark blue) and 89-1591 comp Δlsp complemented strains (dark red). strains. Data represent the mean ± SEM from at least three independent experiments. * Indicates a significant difference with the respective wild-type strain (p < 0.05).
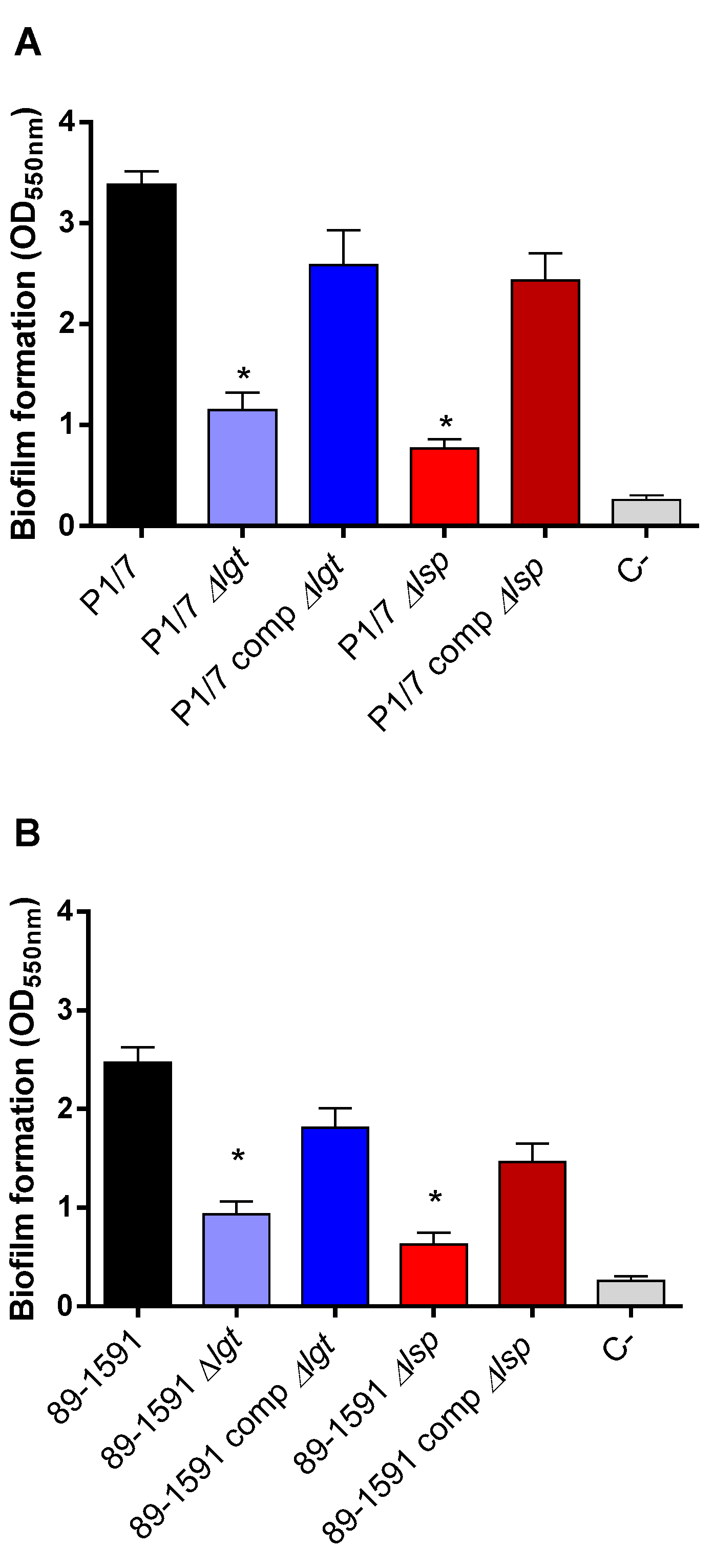
Figure 4.
The diacyl motif is important for recognition by innate immune cells of periplasmic and/or secreted S. suis serotype 2 ST25 lipoproteins. Pro-inflammatory mediator production by bmDCs following activation with live bacteria of the S. suis serotype 2 wild-type strain 89-1591 (ST25) (A–D) (black), as well as their respective Δlgt (blue) and Δlsp (red)) mutant strains and the Δlgt + pMX1-lgt complemented strain (dark blue). Production of TNF (A), IL-6 (B), CXCL1 (C) and CCL3 (D). Data represent the mean + SEM (n = 4 independent experiments). * (p < 0.05) indicates a significant difference between the wild-type and mutant strains. Mock-infected cells induced negligible cytokine values < 300 pg/mL (not shown).
Figure 4.
The diacyl motif is important for recognition by innate immune cells of periplasmic and/or secreted S. suis serotype 2 ST25 lipoproteins. Pro-inflammatory mediator production by bmDCs following activation with live bacteria of the S. suis serotype 2 wild-type strain 89-1591 (ST25) (A–D) (black), as well as their respective Δlgt (blue) and Δlsp (red)) mutant strains and the Δlgt + pMX1-lgt complemented strain (dark blue). Production of TNF (A), IL-6 (B), CXCL1 (C) and CCL3 (D). Data represent the mean + SEM (n = 4 independent experiments). * (p < 0.05) indicates a significant difference between the wild-type and mutant strains. Mock-infected cells induced negligible cytokine values < 300 pg/mL (not shown).
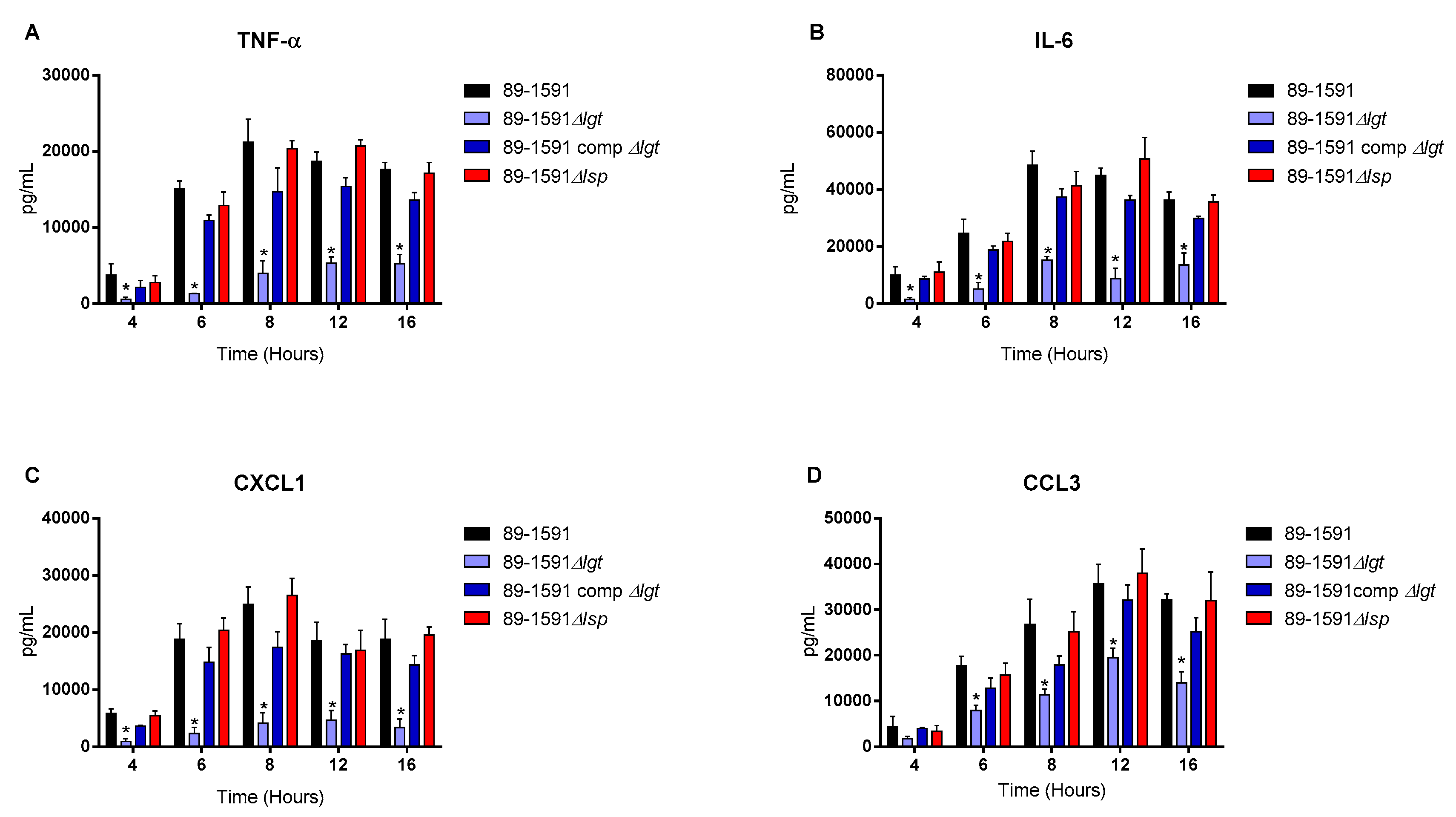
Figure 5.
The diacyl motif is important for recognition by innate immune cells of periplasmic S. suis serotype 2 ST25 lipoproteins. Pro-inflammatory mediator production by bmDCs following infection with heat-killed bacteria of the S. suis serotype 2 wild-type ST25 strain 89-1591 (ST25) (A–D) (black), as well as their respective Δlgt (blue) and Δlsp (red)) mutant strains and the 89-1591 comp Δlgt complemented strain (dark blue). Production of TNF (A), IL-6 (B), CXCL1 (C) and CCL3 (D). Data represent the mean + SEM (n = 4 independent experiments). * (p < 0.05) indicates a significant difference between the wild-type and mutant strains. Mock-infected cells induced negligible cytokine values < 300 pg/mL (not shown).
Figure 5.
The diacyl motif is important for recognition by innate immune cells of periplasmic S. suis serotype 2 ST25 lipoproteins. Pro-inflammatory mediator production by bmDCs following infection with heat-killed bacteria of the S. suis serotype 2 wild-type ST25 strain 89-1591 (ST25) (A–D) (black), as well as their respective Δlgt (blue) and Δlsp (red)) mutant strains and the 89-1591 comp Δlgt complemented strain (dark blue). Production of TNF (A), IL-6 (B), CXCL1 (C) and CCL3 (D). Data represent the mean + SEM (n = 4 independent experiments). * (p < 0.05) indicates a significant difference between the wild-type and mutant strains. Mock-infected cells induced negligible cytokine values < 300 pg/mL (not shown).
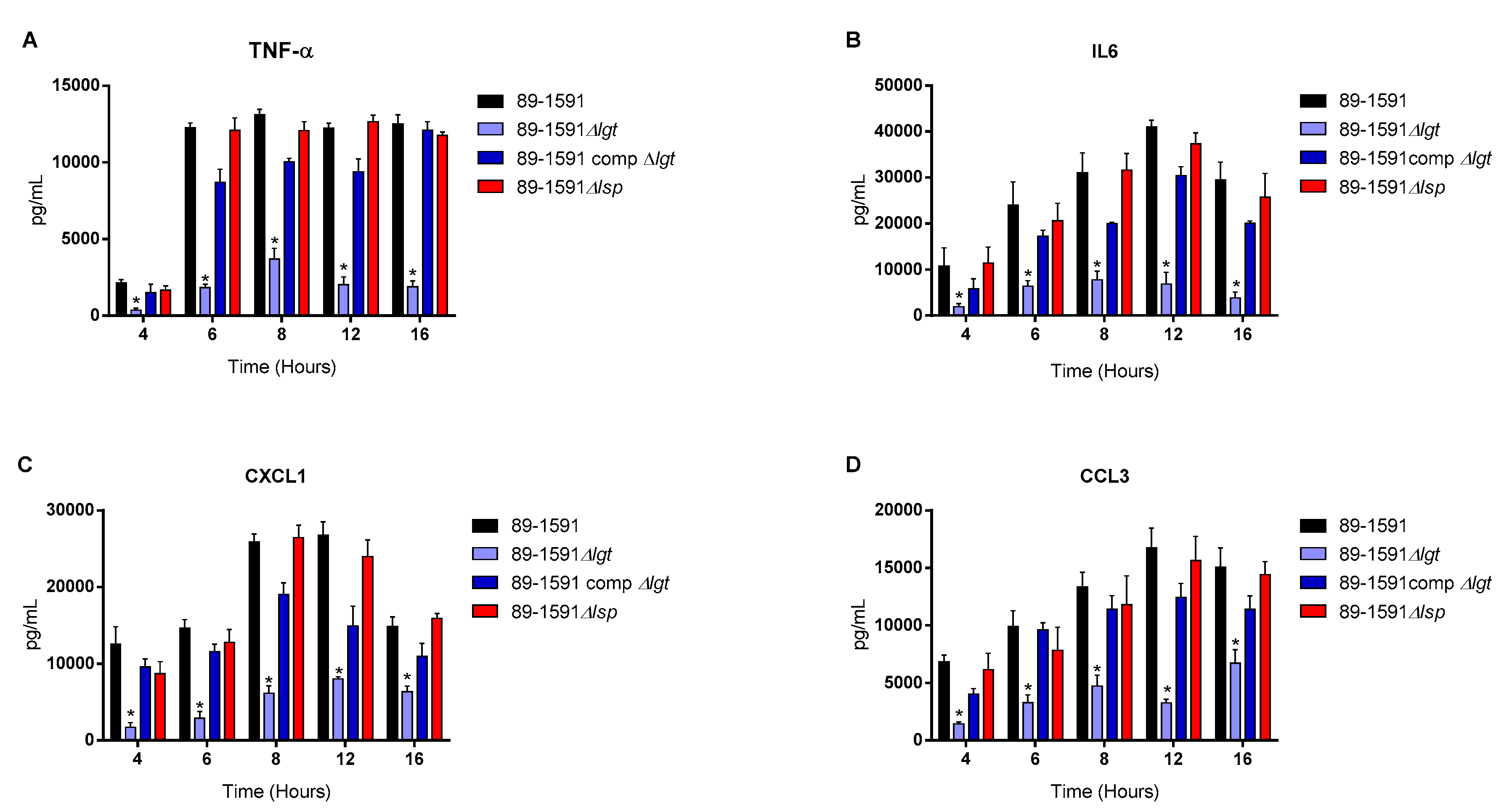
Figure 6.
Both the diacyl motif and the peptide signal cleavage are important for recognition by innate immune cells of secreted S. suis serotype 2 ST25 lipoproteins. Pro-inflammatory mediator production by bmDCs following infection with bacterial-free supernatant of S. suis serotype wild-type ST25 strain 89-1591 (black) as well as their respective Δlgt (blue) and Δlsp (red) mutant strains and the 89-1591 comp Δlgt (dark blue) or 89-1591 comp Δlsp complemented strains (dark red). Production of TNF (A), IL-6 (B), CXCL1 (C) and CCL3 (D). Data represent the mean + SEM (n = 4 independent experiments). * (p < 0.05) indicates a significant difference between wild-type and mutant strains. Mock-infected cells induced negligible cytokine values < 300 pg/mL (not shown).
Figure 6.
Both the diacyl motif and the peptide signal cleavage are important for recognition by innate immune cells of secreted S. suis serotype 2 ST25 lipoproteins. Pro-inflammatory mediator production by bmDCs following infection with bacterial-free supernatant of S. suis serotype wild-type ST25 strain 89-1591 (black) as well as their respective Δlgt (blue) and Δlsp (red) mutant strains and the 89-1591 comp Δlgt (dark blue) or 89-1591 comp Δlsp complemented strains (dark red). Production of TNF (A), IL-6 (B), CXCL1 (C) and CCL3 (D). Data represent the mean + SEM (n = 4 independent experiments). * (p < 0.05) indicates a significant difference between wild-type and mutant strains. Mock-infected cells induced negligible cytokine values < 300 pg/mL (not shown).
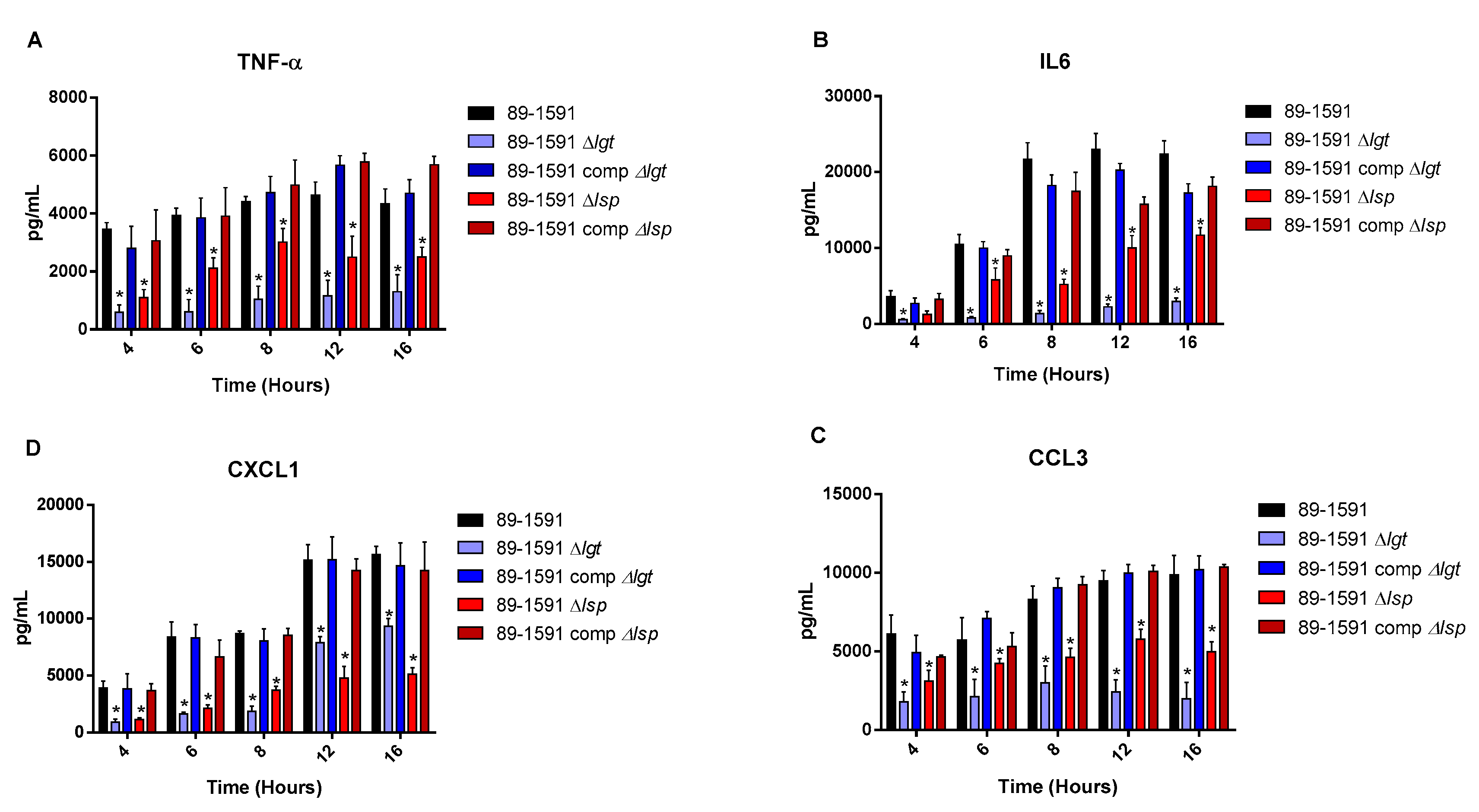
Figure 7.
Presence of Lgt, but not Lsp, is significantly important for S. suis ST25 virulence but it does not affect bacteremia levels. Survival (A) and blood bacterial burden at 12 and 24h post-infection (B, C) of C57BL/6 mice following intraperitoneal inoculation of the S. suis wild-type 89-1591 strain (ST25) (black), Δlgt (bleu) and Δlsp (red) mutant strains. Data represent survival curves (A) (n = 15) or geometric mean (C, B) (n = survived mice at each time point). * (p < 0.05) indicates a significant difference between survival of mice infected the wild-type and the Δlgt mutant strain.
Figure 7.
Presence of Lgt, but not Lsp, is significantly important for S. suis ST25 virulence but it does not affect bacteremia levels. Survival (A) and blood bacterial burden at 12 and 24h post-infection (B, C) of C57BL/6 mice following intraperitoneal inoculation of the S. suis wild-type 89-1591 strain (ST25) (black), Δlgt (bleu) and Δlsp (red) mutant strains. Data represent survival curves (A) (n = 15) or geometric mean (C, B) (n = survived mice at each time point). * (p < 0.05) indicates a significant difference between survival of mice infected the wild-type and the Δlgt mutant strain.
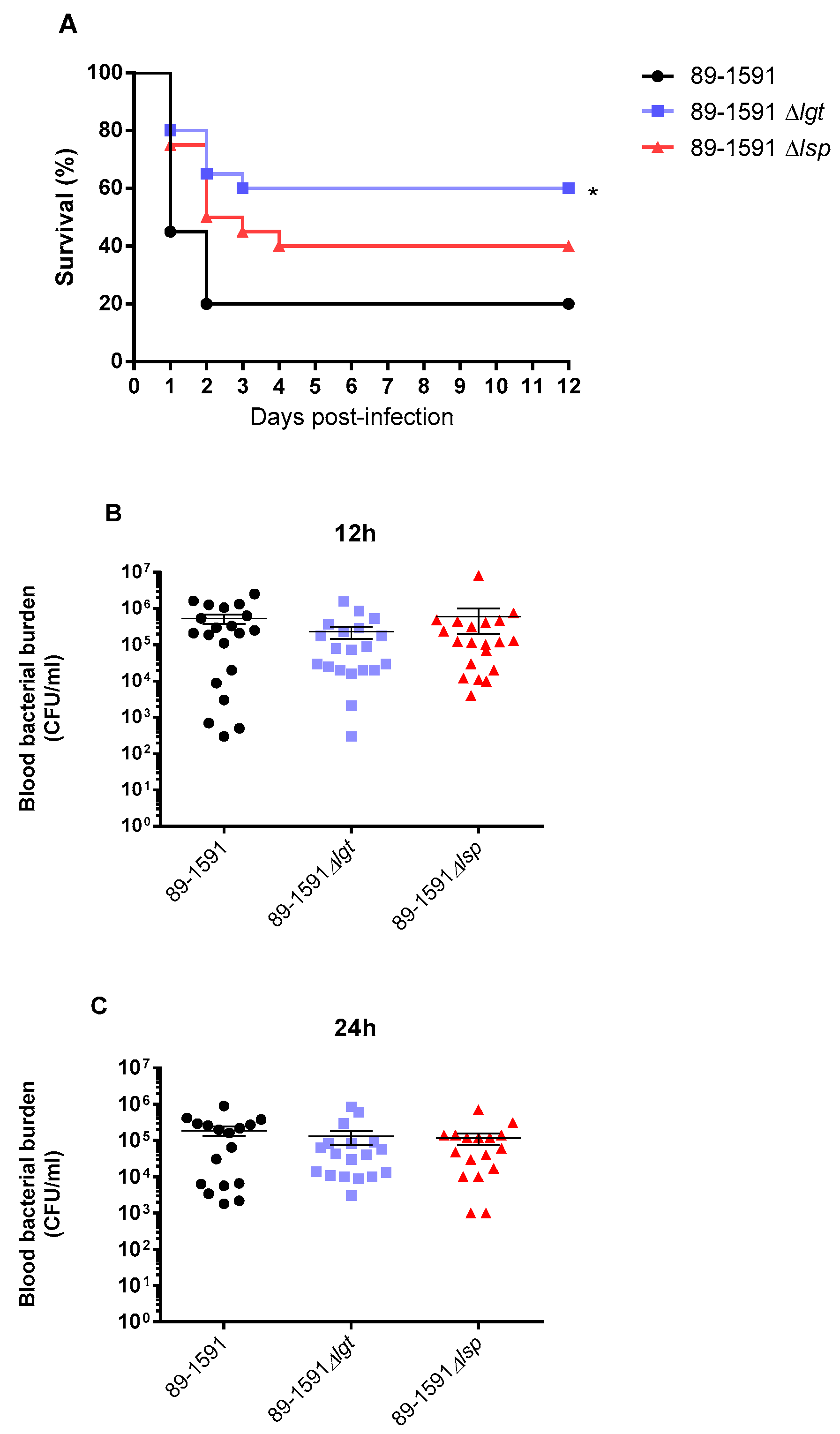

Table 1.
List of strains and plasmids used in this study.
| Strain or plasmid | Characteristics | Reference |
|---|---|---|
| Streptococcus suis | ||
| P1/7 | Virulent serotype 2 ST1 strain isolated from a case of pig meningitis in the United Kingdom | [18] |
| P1/7Δlgt | Isogenic mutant derived from P1/7; in frame deletion of lgt gene | [7] |
| P1/7 Δlsp | Isogenic mutant derived from P1/7; in frame deletion of lsp gene | [7] |
| P1/7 comp Δlgt | Mutant Δlgt complemented with pMX1-lgt complementation vector | [7] |
| P1/7 comp Δlsp | Mutant Δlsp complemented with pMX1-lsp complementation vector | [7] |
| P1/7 ΔcpsF | Isogenic mutant derived from P1/7; in frame deletion of cpsF | [19] |
| 89-1591 | Virulent North American ST25 strain isolated from a case of pig sepsis in Canada | [20] |
| 89-1591 Δlgt | Isogenic mutant derived from SC84; in frame deletion of lgt gene | This study |
| 89-1591 Δlsp | Isogenic mutant derived from SC84; in frame deletion of lsp gene | This study |
| 89-1591 comp Δlgt | Mutant Δlgt complemented with pMX1-lgt complementation vector | This study |
| 89-1591 comp Δlsp | Mutant Δlsp complemented with pMX1-lsp complementation vector | This study |
| 89-1591 ΔcpsF | Isogenic mutant derived from 89-1591; in frame deletion of cpsF | [14] |
| Escherichia coli | ||
| TOP10 | F- mrcA Δ(mrr-hsdRMS-mcrBC) φ80 lacZΔM15 ΔlacX74 recA1 araD139 Δ(araleu) 7697 galU galK rpsL (Strr) endA1 nupG | Invitrogen |
| MC1061 | F- Δ(ara-leu)7697 [araD139]B/r Δ(codB-lacI)3 galK16 galE15 λ- e14- mcrA0 relA1 rpsL150(StrR) spoT1 mcrB1 hsdR2(r-m+)Host for pMX1 derivatives | [21] |
| Plasmids | ||
| pCR2.1 | Apr, Kmr, pUC ori, lacZΔM15 | Invitrogen |
| pSET4s | Spcr, pUC ori, thermosensitive pG+host3 ori, lacZΔM15 | [22] |
| pMX1 | Replication functions of pSSU1, MCS pUC19 lacZ Spcr, malX promoter of S. suis, derivative of pSET2 | [22,23] |
| p4Δlgt | pSET-4s carrying the construct for lgt allelic replacement | This study |
| p4Δlsp | pSET-4s carrying the construct for lsp allelic replacement | This study |
| pMX1-lgt (P1/7) | pMX1 carrying intact lgt gene | [7] |
| pMX1-lsp (P1/7) | pMX1 carrying intact lsp gene | [7] |
| pMX1-lgt (89-1591) | pMX1 carrying intact lgt gene | This study |
| pMX1-lsp (89-1591) | pMX1 carrying intact lsp gene | This study |
Table 2.
List of oligonucleotide primers used in this study.
| Name | Sequence (5’ – 3’) | Construct |
|---|---|---|
| lgt-ID1 | GGAACGCTATGGAACAGGTC | p4Δlgt |
| lgt-ID2 | CACTCCATGAAAAGGCGACG | p4Δlgt |
| lgt-ID3 | CGTAGACGGCCAAAATTCC | p4Δlgt |
| lgt-ID4 | CGCTTATCTGCTGGATTCTCC | p4Δlgt |
| lgt-ID5 | GCCAATCGTCTGCATCAAGG | p4Δlgt |
| lgt-ID6 | GGGTTGATAGAATGGGATTGCATACCAACG | p4Δlgt |
| lgt-ID7 | CGTTGGTATGCAATCCCATTCTATCAACCC | p4Δlgt |
| lgt-ID8 | GACCGACTTGCTGGTCAAAC | p4Δlgt |
| lsp-ID1 | TGAGAAAACTGTTGTGGGTA | p4Δlsp |
| lsp-ID2 | AGAGCACCAGCAATCATCAA | p4Δlsp |
| lsp-ID3 | TTGATGATTGCTGGTGCTCT | p4Δlsp |
| lsp-ID4 | TAGACAGCGAACAGAGATAC | p4Δlsp |
| lsp-ID5 | TACGCTACGTTGTAGCCATTGC | p4Δlsp |
| lsp-ID6 | ACCTACACCAACTGTTAATACTACCATCAA | p4Δlsp |
| lsp-ID7 | TTGATGGTAGTATTAACAGTTGGTGTAGGT | p4Δlsp |
| lsp-ID8 | CGCGCTGCAGCCAAAGTGTAGTCACCAAAA | p4Δlsp |
| pMX1-lgt-F | CCGCCATGGACAGATGGGGTTTGATGCAAC | pMX1-lgt |
| pMX1-lgt-R | CGCGAATTCGGACAAGGCAATAATCAAGAC | pMX1-lgt |
| pMX1-lsp-F | GTGCCATGGACTTTATTGAAACCATGCAGG | pMX1-lsp |
| pMX1-lsp-R | ATCGAATTCAATACCACCAACCTCAACTCT | pMX1-lsp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



