
Submitted:
30 August 2023
Posted:
01 September 2023
You are already at the latest version
Abstract
During the bioassay, on leaf sections of the A. roylei species, a significantly higher number of T. tabaci females was recorded compared to the cv. Tęcza of the A. cepa species and the cv. Kroll of the A. fistulosum species, in each observation period. Significantly more thrips settled on cv. Kroll compared to Alibaba and Bila. Regarding the results obtained on A. cepa, significantly fewer females were found on cv. Bila compared to cv. Tęcza. Opposite results were observed in a combination of cvs. Tęcza - Alibaba, where significantly more insects settled on leaves of cv. Alibaba. Statistically significant differences of cultivars/species were found in the number of hatched larvae on the leaves of the tested cultivars/species of onion. The significantly lowest number of larvae hatched from eggs laid on A. roylei compared to A. fistulosum and cultivars of A. cepa, except for Bila.
Keywords:
bioassay
; oviposition
; settling preference
; resistance
1. Introduction
Onion (common onion or bulb onion) (Allium cepa L.) is one of the most important vegetable crops worldwide with a global total production of 93,226,400 tones and it accounts for about 24% of the world’s total vegetable production [1]. Onion production and breeding encounter many challenges. Like other crops, onions are susceptible to insect, fungal, bacterial, viral, and nematode pests [2,3]. One of the main insect pests is the onion thrips (Thrips tabaci Lind.; Thysanoptera: Thripidae). It is a phytophagous and polyphagous, invasive, cosmopolitan and highly fecundating insect pest with a rapid development rate, and vector of several onion pathogens and tospoviruses [4-8]. Attack by T. tabaci not only leads to a complete loss of onion seedlings but also may cause damage to older crops by feeding on leaves as well as onion bulbs [9]. The poor effectiveness of insecticides at controlling thrips – and thrips’ increased resistance to them – leads to high losses in the cultivation of onions [10,11]. A reduction in yield of about 40-65% has been reported due to attacks by these pests [12,13]. Full resistance (or tolerance) of onion to T. tabaci would be very beneficial, however, in the available literature, we have not found information about the existence of such cultivars and breeding lines of onion. However, in recent years, some plant characteristics that are responsible for the partial resistance of onions to T. tabaci have been identified. The differences in resistant and susceptible cultivars have been associated with leaf color [14], amounts and types of epicuticular leaf wax [15,16], plant architecture and anatomy [17,18] and total phenol content [19,20]. Recently, it was also possible to select some onions genotypes that showed a lower number of thrips compared to susceptible ones [19,21], and those produced large bulb yields under thrips pressure [17,22]. Onion breeding programs currently focus mainly on the improvement of existing cultivars; however, wild Allium species possess many desirable traits such as disease and pest resistance [23].
Allium roylei Stearn and Allium fistulosum L. have been recognized as the most important gene pools of onion [24,25]. A. roylei is a wild species originating from the Indian subcontinent which possesses genes imparting resistance to various harmful fungal diseases and pests [26-29]. A. fistulosum (Welsh onion, Japanese bunching onion, spring onion) is widely cultivated in Japan, China and Korea. Its origin is unknown; it is believed to be of Chinese origin [30]. This Allium sp. also possesses many traits agronomically useful for onions, due to its resistance to onion leaf blight, pink root, anthracnose, and onion fly [23,31,32].
Our previous field studies, in which A. roylei and A. fistulosum were less colonized and damaged by T. tabaci compared to A. cepa cultivars and breeding lines [33], encouraged us to continue research on them. Therefore, this research under laboratory conditions was conducted to prove T. tabaci settlement and oviposition activity on leaves of A. roylei and A. fistulosum compared to three cultivars of A. cepa that differed in their susceptibility to colonization and feeding of thrips under field conditions [20].
2. Materials and Methods
2.1. Test plants
Three onion species were used in the laboratory experiments: Allium cepa L. (cvs. Alibaba, Bila and, Tęcza.), Allium fistulosum (cv. Kroll), and Allium roylei (ecotypes 333). All the A. cepa and A. fistulosum cultivars used in the trials are recommended for cultivation in central Europe and are commercially available. The seeds were obtained from Polish breeding companies, namely PlantiCo Zielonki in Stare Babice (cvs. Alibaba, Bila, and Kroll) and Spójnia in Nochowo (cv. Tęcza). A. roylei seeds we obtained from the bank Plant Genetic Resources Laboratory, Research Institute of Vegetable Crops in Skierniewice in Poland. The onion plants used in the experiments were grown in a standard substrate in trays within a plant growing room at 24 ± 1°C, 35 ± 5% relative humidity and a photoperiod of 16:8 h light:dark. All plants were watered regularly, only with tap water. Leaves from onion plants that were approximately four weeks old were used in all bioassays.
2.2. Test insects
Using a rearing method adapted from Loomans & Murai (1997) [34], a stock culture of a thelytokous T. tabaci strain was maintained on white cabbage leaves in 0·75-liter glass jars covered with a fine mesh to ensure ventilation. The rearing was kept in a climate chamber at 24 ± 1°C, 35 ± 5% relative humidity and a photoperiod of 16:8 h light:dark. White cabbage was purchased regularly, and fresh pieces of leaf were added two to three times a week.
To obtain groups of females of known age, thrips pupae were randomly collected from the rearing jars and transferred to Petri dishes (diameter 90 mm) with sections of leek (Allium ampeloprasum L.) leaves. The dishes were closed with lids with central holes covered with a fine mesh to allow air circulation and were sealed with sealing film to prevent the thrips from escaping. After 48 h, adult females were checked, and any remaining pupae were removed. Following an additional pre-oviposition period of 48 hours, single females were used in the bioassays.
2.3. Oviposition rate
The oviposition rate of female onion thrips on Allium species/cultivars was evaluated using a no-choice test. For this bioassay, four-centimeter sections of onion leaves were cut from the middle part of the leaves of the respective test plant. To protect the leaf sections from desiccation and to prevent the thrips from getting inside the leaves, both ends of the leaf sections were briefly dipped in warm liquid paraffin wax. After the wax had solidified, the leaf sections were placed singly on a thin film of 1 % water agar (Agar – Agar, Kobe I, Carl Roth, Karlsruhe, Germany) in glass Petri dishes (60 mm diameter). Subsequently, single females of known age were transferred to each glass Petri dish and placed on the leaf section. To prevent the escape of the T. tabaci females, the dishes were covered with thin (14 µm) clear plastic film (Carl Roth, Karlsruhe, Germany), which was perforated (one hole per cm2 on average) using insect pins (0·4 mm diameter). The bioassay units were kept in a climate chamber at 24 ± 1°C, 35 ± 5% relative humidity and a photoperiod of 16:8 h light:dark. After 24 hours, the females were removed. The plant sections with eggs were kept in the climatic chamber for another five days. After that, the hatched larvae were counted under a stereoscopic microscope.
2.4. Settling preference
The settling preference of T. tabaci females for leaf sections of the different Allium species/cultivars was determined by means of a choice test. Eight pairs of species/cultivars were compared against each other: Tęcza × Alibaba; Tęcza × Bila; Tęcza × Kroll; Tęcza × A. roylei, Alibaba × Bila; Alibaba × Kroll; Bila × Kroll; and Kroll × A. roylei. Each pair consisted of two four-centimeter sections of onion leaves, sealed with wax at both ends (as described above). These sections were placed parallel and equidistant to each other in the center of the bottom of a 90 mm diameter glass Petri dish. Subsequently, ten females of unknown age were placed at the starting point in the center, between the parallel-lying onion leaf sections of the tested pair of cultivars. Each bioassay unit was covered with perforated plastic film and completely randomized under an artificial light source in a climate chamber at 24 ± 1°C and 35 ± 5% relative humidity. Ten, 30, 60, 120, and 180 minutes after the female thrips had dispersed from the starting point, the number of thrips were counted on each of the two sections of onion leaves of the tested cultivars, as well as in the surrounding space. The experiment was replicated 10 to 12 times resulting in preferences recorded for 100 to 120 female thrips for each pair of onion cultivars/species.
2.5. Statistical analysis
Analyses of the bioassay data were performed using Statistica 13 software (TIBCO Software Inc., 2017). The results of the settling preference tests were analysed with Student’s t-test with a significance level of P < 0.05. The data obtained from the oviposition tests with hatched thrips larvae were subjected to a one-way analysis of variance (ANOVA), with the factor onion cultivar/species. Residual plots were checked prior to data analysis. In cases where the data did not show a normal distribution, they were normalized by log(x+1) transformation. The Tukey test was used to compare means at a significance level of P < 0.05.
3. Results
In the no-choice experiments on the oviposition rate of female onion thrips, significant variability was observed among cultivars/species regarding the mean number of thrips larvae that had hatched from eggs laid inside the tissue of the onion leaves (F = 27.158; df = 4; p < 0.000). T. tabaci females laid significantly more eggs on leaves of A. cepa cultivars compared to A roylei (almost three times more) (Figure 1). Additionally, a significantly higher number of T. tabaci larvae hatched from eggs laid on cv. Tęcza and Bila compared to A. fistulosum (cv Kroll). Moreover, a significantly higher number of T. tabaci larvae hatched from eggs on cv Kroll than on A. roylei (Figure 1).
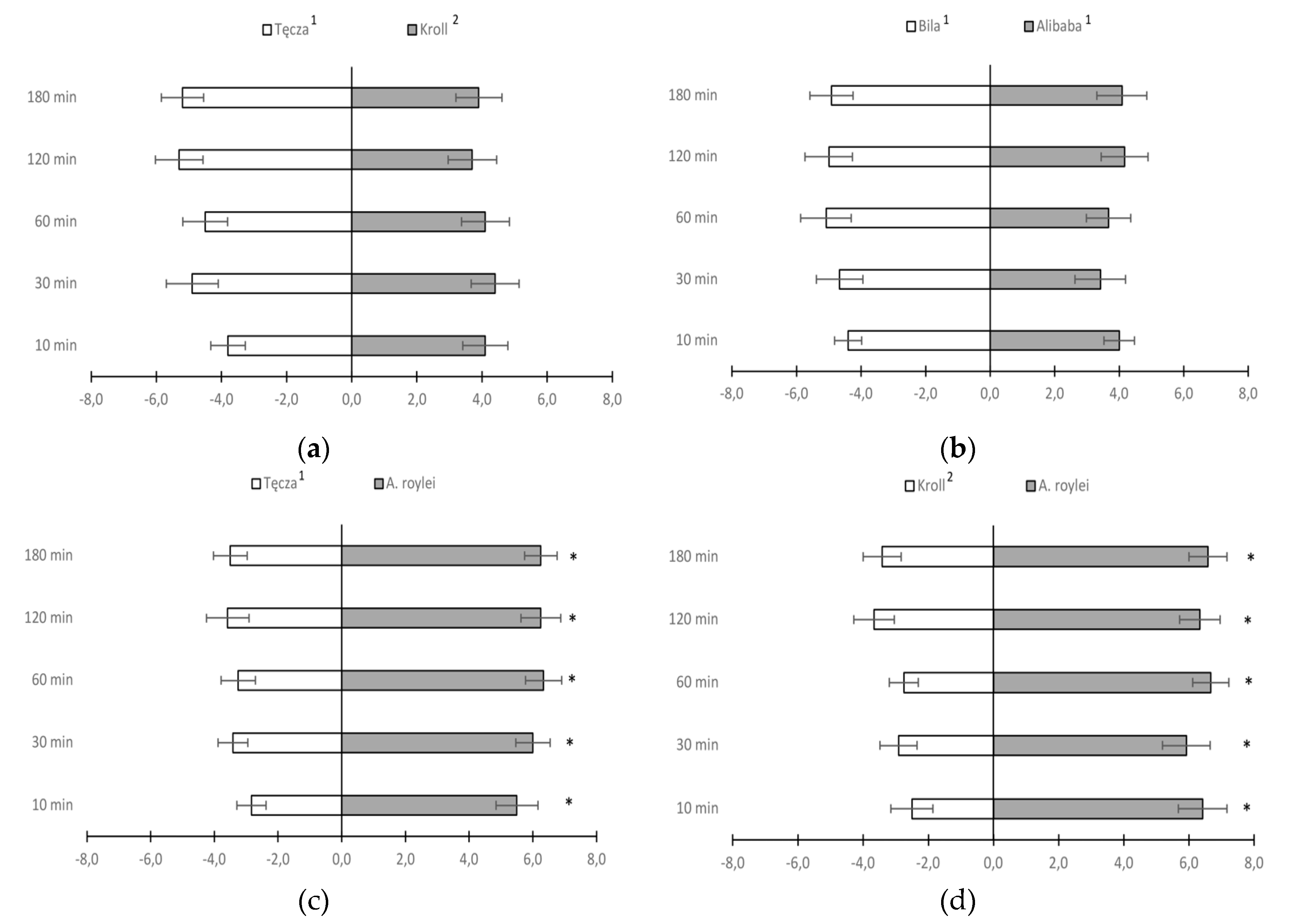
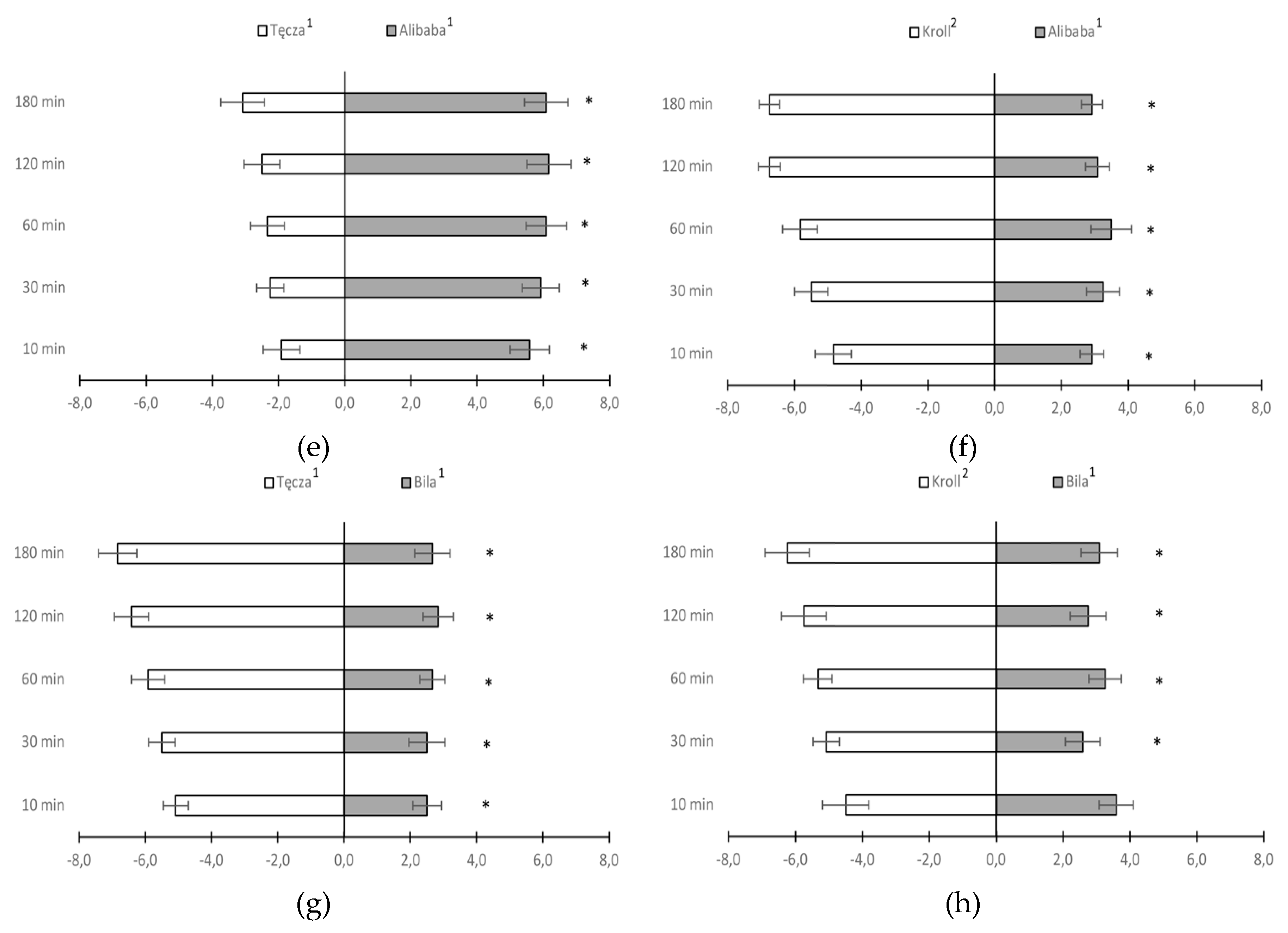
In a preference test for onion thrips settling on different species and cultivars of Allium sp., a significantly higher number of female onion thrips settled on leaf sections of A. roylei compared to cv. Tęcza (A. cepa) and cv. Kroll (A. fistulosum) during each observation period (Figure 2c, d). In both comparisons, almost twice as many T. tabaci females were observed on leaf sections of A. roylei. When A. cepa cultivars were paired with cv. Kroll (A. fistulosum), female T. tabaci demonstrated a significant preference for A. fistulosum over the cv. Alibaba throughout the test period and cv. Bila throughout the test period except for the first 10 minutes (Figure 2 f, h). When common onion cultivars Alibaba and Bila were paired with cv. Tęcza, the settling preference of T. tabaci females was different. A significantly higher number of thrips settled on cv. Alibaba compared to cv. Tęcza, whereas cv. Tęcza was preferred over Bila (Figure 2 e, g). For the pairs Alibaba x Bila (Figure 2 b) and Tęcza x Kroll (Figure 2 a) there were no significant differences in onion thrips’ preference.
4. Discussion
Our experiments were short-term laboratory experiments that only provide information on thrips’ selection behavior at the first stage of colonization and acceptance of the plant for egg-laying. The observed differences in the numbers of hatched larvae indeed reflect differences in oviposition (numbers of eggs from which the larvae have hatched). Among the three onion species tested (A. cepa, A. fistulosum, and A. roylei), the most frequently colonized by T. tabaci females was A. roylei. While adult T. tabaci did not respond to A. cepa plant odor in a Y-tube olfactometer [14], plant volatiles from A. roylei leaf sections may have attracted T. tabaci during the 3 h period in this study. Olfactometer tests using T. tabaci and A. roylei plants could help to clarify this possible attraction. On the contrary, in the oviposition bioassay without choice over a 24-hour period, four times fewer larvae hatched on the leaves of A. roylei than on the common onion cv. Tęcza and almost two times fewer than on Welsh onion. It has been hypothesized by some authors that thrips and other herbivores females would prefer those leaves for oviposition on which they realize the highest reproductive success [35-37]. Lower fecundity of females on leaves of A. roylei could therefore result from less favorable conditions for feeding and development of offspring. In our earlier field studies we noticed that despite the presence of T. tabaci on the leaves of A. roylei, no damage caused by thrips feeding on them was observed, in contrast to the injured leaves of A. cepa and A. fistulosum [33]. The lower oviposition rate of females on A. roylei leaves under laboratory conditions and the lack of damage to the leaves under field conditions – when T. tabaci had a choice between many cultivars and the breeding lines of A. cepa and A. fistulosum – may suggest that this wild onion species has some traits of resistance to T. tabaci which discourage thrips from feeding and egg laying. A. roylei is a plant with narrow, drooping leaves which are sometimes described as filiform. Mature, full-sized leaves are fistulous, though smaller ones are sometimes solid. In turn, A. fistulosum (cv. Kroll) leaves are wide, erect and pointed hollow tubes and are more similar to the cylindrical, fleshy, and hollow leaves of A. cepa [38] (authors’ observations). Differences between resistance to thrips of the tested onion species may be due to certain morphological features of their leaves, which make it difficult for females to lay eggs. They may concern the thickness and rigidity of the cellular wall, the amount of epicuticular waxes, the structure of the epidermis, and the number of stomates [39,40]. Post-alighting host acceptance, and also subsequent feeding and reproduction, are strongly influenced by the plant’s nutritional quality and defenses such as secondary metabolites [41,42]. Many bioactive metabolites like cysteine, sulfoxides, flavanols, polyphenols, and saponins are synthesized by different organs of A. roylei to provide defense against a wide range of plant pathogens and herbivores [43,44]. Resistance against downy mildew (Perenospora destructor (Berk.) Casp. ex Berk.) was identified in A. roylei and successfully transferred to bulb onion [28]. This wild onion is partially resistant to leaf blight disease caused by Botrytis squamosa J. C. Walker and basal rot disease caused by Fusarium oxysporum f. sp. cepae [29,45]. Also, A. roylei has been proven to be partially resistant to beet armyworm Spodoptera exigua Hübner. Larval growth and survival of S. exigua proved to be significantly slower on A. roylei compared to the A. cepa, A. fistulosum, and A. galanthum Kar. et Kir. [46]. The prospect of using A. roylei as a source of resistance to T. tabaci in onion breeding is promising, but further research is needed on how many larvae can complete development, how quickly they will develop, and what will be the final condition and fertility of the next generation of females.
In our bioassay, A. fistulosum cv. Kroll was more frequently chosen for settling by female T. tabaci than the two cultivars of A. cepa i.e., Alibaba and Bila, but a significantly lower number of larvae hatched from eggs laid by females on leaves of A. fistulosum compared with leaves of two cultivars of A. cepa, i.e. Bila and Tęcza. In our field study, despite the very high number of thrips caught from the leaves of A. fistulosum, minor damage was recorded on them [33]. The leaves of A. fistulosum were damaged by feeding thrips only in 6.5% of cases in 2015 and 1.5% of cases in 2016 (unpubl. data) while the corresponding figures for leaves of A. cepa cvs. Alibaba, Bila, and Tęcza were, respectively, 13.5%, 13.9%, and 17.8% in 2015 and 5.3%, 6.8%, and 7.3% in 2016 [20]. This was also confirmed by a field study by Hudák and Pénzes [47], where A. fistulosum showed less damage than A. cepa under similar field conditions. It confirms that T. tabaci has shown higher affinity toward A. fistulosum phenotype during settlement and supports higher densities of thrips, but A. cepa creates better conditions for foraging and laying eggs. Ren et al. [48] proved that volatiles are important factors for thrips in host preference. In their study, one of the most attractive volatiles, along with Medicago sativa, for T. tabaci and Frankliniella occidentalis Pergande appeared to be volatiles of A. fistulosum in its vegetative and flowering stages. In turn, Jones et al. [49] showed that the Nebuka type of A. fistulosum has a similar low degree of thrips colonization as the resistant cv. White Persian of A. cepa. The authors noted that the leaves of both Allium sp. were circular, and they had a spreading growth habit and a long sheath region. Some authors report that A. fistulosum can be used for improvement of the common onion, especially for its resistance to pink root (Phoma terrestris E. M. Hans.) [50], Fusarium basal rot, T. tabaci, smut (Urocystis cepulae Frost) [51], and onion fly (Hylemya antiqua Bouche) [52]. Varietal resistance against Liriomyza chinensis (Kato) has been reported in A. fistulosum in Japan. Antibiosis studies revealed a significant difference in survival up to the pupal stage, in the forewing lengths of adults and in the development time from the egg to pupal stages among the resistant and susceptible varieties of A. fistulosum [53,54]. A. fistulosum, as well as wild Allium species like A. hookeri, A. altaicum, and A. angulosum, are a rich source of lectins and these compounds have recently been proven to have insecticidal activity against T. tabaci [55]. The authors suggest that the high lectin content of A. hookeri and A. fistulosum can be correlated with the low amount of thrips damage. Whole plant and detached leaf damage tests revealed that A. hookeri was resistant to T. tabaci. However, worse development of this pest was observed not only on A. hookeri, but also on A. fistulosum. There are many local and commercial cultivars of A. fistulosum with distinctive differences in morphological and other traits which are adapted to a variety of climatic conditions. The wide variety of A. fistulosum phenotypes that exists around the world [56] suggests that some of them will likely develop traits that will promote resistance or tolerance to T. tabaci, so further research in this direction appears to be justified.
In a previous field study, cv. Tęcza was resistant to thrips abundance but susceptible to thrips feeding and was more heavily damaged than varieties susceptible to thrips infestation and foraging cv. Alibaba [20]. Although the laboratory test did not show significant differences in the number of hatched T. tabaci larvae between tested cultivars of A. cepa, the highest number of them was recorded on cv. Tęcza. Perhaps the cv. Tęcza, which stimulated T. tabaci individuals to feed more under field conditions, could also stimulate females to lay eggs more intensively during the bioassay test. In the settlement test in the Alibaba x Tęcza pairing, the most preference was exhibited by female onion thrips to the cv. Alibaba, but in pair of Bila x Tęcza, cv. Tęcza was preferred to cv. Bila. In a field study, Alibaba was also more populated by T. tabaci than cv. Tęcza but cv. Tęcza appeared to be less attractive than cv. Bila [20]. Laboratory tests do not always reflect the behavior of insects in the field, where they are influenced by many abiotic and biotic factors, and the results obtained must be interpreted with this in mind.
5. Conclusions
The choice between A. cepa, A. roylei and A. fistulosum by the females of T. tabaci in the tests on the preference for settlement and the rate of oviposition was the opposite. This confirms results obtained by other researchers that other plants’ characteristics attract females to colonize them, and others stimulate them to lay eggs [57]. Since the lowest number of larvae hatched on A. roylei, followed by A. fistulosum, it can be assumed that the leaves of these species have certain traits that discourage or inhibit females from laying eggs or hatching larvae. For this reason, these species should be studied in the future for the biology of T. tabaci on them and for the features, both morphological, anatomical, and biochemical, that may impede the development of thrips.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, M.O. and M.P.; methodology, E.K. and M.O.; software, M.P. and T.W.; validation, M.P. and M.O.; formal analysis, M.P.; investigation, M.O. and E.K.; resources, M.P and T.W.; data curation, M.O. and M.P.; writing—M.P. and M.O.; writing—review and editing, M.P.; M.O. and E.K.; visualization, T.W. and M.P.; supervision, M.P. and M.O.; project administration, M.P. and E.K.; funding acquisition M.P. and E.K. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was supported by the Ministry of Science and Higher Education of Poland as a part of a research subsidy to the University of Agriculture in Krakow (DS-3508/WBiO).
Institutional Review Board Statement
All animal work was conducted according to relevant national and international guidelines. For insect collection, no permits were required since the area where thrips were collected did not contain any strictly protected areas, and Thrips tabaci is not under protection in Europe. Also, no permits were required to use insects for experiments due to the observational nature of the data collection.
Data Availability Statement
The data presented in this study are openly available in Harvard Dataverse: https://doi.org/10.7910/DVN/5KCCM7 accessed date 16 July 2023.
Acknowledgments
We would like to thank to Joseph William Woodborn for proofreading this manuscript. Thanks to Ghita Della Pasqua for help with the analysis.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- FAOSTAT. Food and Agriculture Commodity Production Data; Food and Agriculture Organization: Rome, Italy, 2021. [Google Scholar]
- Mishra, R.K.; Jaiswal, R.K.; Kumar D.; Saabal P.R; Singh A. Management of major diseases and insect pests of onion and garlic: A comprehensive review. J. Plant Breed. Crop. Sci. 2014, 6, 160–170. [CrossRef]
- Schwartz, H.F; Mohan, S.K. Compendium of Onion and Garlic Diseases; APS Press: St. Paul, MN, USA, 1985.
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): a global pest of increasing concern in onion. J. Econ. Èntomol. 2011, 104, 1–13. [Google Scholar] [CrossRef]
- Gent, D.H.; du Toit, L.J.; Fichtner, S.F.; Mohan, S.K.; Pappu, H.R.; Schwartz, H.F. Iris yellow spot virus: An Emerging Threat to Onion Bulb and Seed Production. Plant Dis. 2006, 90, 1468–1480. [Google Scholar] [CrossRef]
- Grode, A.; Chen, S.; Walker, E.D.; Szendrei, Z. Onion thrips (Thysanoptera: Thripidae) feeding promotes infection by Pantoea ananatis in onion. J. Econ. Entomol. 2017, 110, 2301–2307. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.K.; Garg, H.; Gill, A.K.; Gillett-Kaufman, J.L.; Nault, B.A. Onion Thrips (Thysanoptera: Thripidae) Biology, Ecology, and Management in Onion Production Systems. J. Integr. Pest Manag. 2015, 6, 6–6. [Google Scholar] [CrossRef]
- Lewis, T. Pest thrips in perspective. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: New York, NY, USA, 1997; pp. 1–13. ISBN 9780851991788. [Google Scholar]
- Martin, N.A.; Workman, P.J. A new bioassay for determining the susceptibility of onion(Allium cepa)bulbs to onion thrips,Thrips tabaci(Thysanoptera: Thripidae). New Zealand J. Crop. Hortic. Sci. 2006, 34, 85–92. [Google Scholar] [CrossRef]
- Adesanya, A.W.; Waters, T.D.; Lavine, M.D.; Walsh, D.B.; Lavine, L.C.; Zhu, F. Multiple insecticide resistance in onion thrips populations from Western USA. Pestic. Biochem. Physiol. 2020, 165, 104553. [Google Scholar] [CrossRef]
- Shelton, A.M.; Zhao, J.-Z.; Nault, B.A.; Plate, J.; Musser, F.R.; Larentzaki, E. Patterns of Insecticide Resistance in Onion Thrips (Thysanoptera: Thripidae) in Onion Fields in New York. J. Econ. Èntomol. 2006, 99, 1798–1804. [Google Scholar] [CrossRef]
- Karar, H.; Abbas, G.; Hameed, A. ; Ahmad. G.; Ali, A. Losses in Onion (Allium cepa) Due to Onion Thrips (Thrips tabaci) (Thysanoptera: Thripidae) and Effect of Weather Factors on Population Dynamics of Thips. World Appl Sci J. 2014, 32 (11), 2250-2258. [Google Scholar] [CrossRef]
- Kendall, D.M.; Capinera, J.L. Susceptibility of Onion Growth Stages to Onion Thrips (Thysanoptera: Thripidae) Damage and Mechanical Defoliation. Environ. Èntomol. 1987, 16, 859–863. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Fail, J.; Deutschlander, M.; Nault, B.A.; Shelton, A.M. Characterization of Resistance, Evaluation of the Attractiveness of Plant Odors, and Effect of Leaf Color on Different Onion Cultivars to Onion Thrips (Thysanoptera: Thripidae). J. Econ. Èntomol. 2012, 105, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Damon, S.J.; Groves, R.L.; Havey, M.J. Variation for Epicuticular Waxes on Onion Foliage and Impacts on Numbers of Onion Thrips. J. Am. Soc. Hortic. Sci. 2014, 139, 495–501. [Google Scholar] [CrossRef]
- Munaiz, E.D.; Groves, R.L.; Havey, M.J. Amounts and Types of Epicuticular Leaf Waxes among Onion Accessions Selected for Reduced Damage by Onion Thrips. J. Am. Soc. Hortic. Sci. 2020, 145, 30–35. [Google Scholar] [CrossRef]
- Ferreira, G.d.O.; Santos, C.A.F.; Oliveira, V.R.; de Alencar, J.A.; da Silva, D.O.M. Evaluation of onion accessions for resistance to thrips in Brazilian semi-arid regions. J. Hortic. Sci. Biotechnol. 2017, 92, 550–558. [Google Scholar] [CrossRef]
- da Silva, V.C.P.; Bettoni, M.M.; Bona, C.; Foerster, L.A. Morphological and chemical characteristics of onion plants (Allium cepa L.) associated with resistance to onion thrips. Acta Sci. Agron. 2014, 37, 85. [Google Scholar] [CrossRef]
- Njau, G.M.; Nyomora, A.M.S.; Dinssa, F.F.; Chang, J.-C.; Malini, P.; Subramanian, S.; Srinivasan, R. Evaluation of onion (Allium cepa) germplasm entries for resistance to onion thrips, Thrips tabaci (Lindeman) in Tanzania. Int. J. Trop. Insect Sci. 2017, 37, 98–113. [Google Scholar] [CrossRef]
- Pobożniak, M.; Olczyk, M.; Wójtowicz, T.; Kamińska, I.; Hanus-Fajerska, E.; Kostecka-Gugała, A.; Kruczek, M. Anatomical and Biochemical Traits Associated with Field Resistance of Onion Cultivars to Onion Thrips and the Effect of Mechanical Injury on the Level of Biochemical Compounds in Onion Leaves. Agronomy 2022, 12, 147. [Google Scholar] [CrossRef]
- Kamal, N.; Nourbakhsh, S.S.; Cramer, C.S. Reduced Iris Yellow Spot Symptoms through Selection within Onion Breeding Lines. Horticulturae 2021, 7, 12. [Google Scholar] [CrossRef]
- Raut, A.; Pal, S.; Wahengbam, J.; Banu, A.N. Population dynamics of onion thrips (Thrips tabaci lindeman, Thysanoptera; Thripidae) and varietal response of onion cultivars against onion thrips. J. Èntomol. Res. 2020, 44, 547–554. [Google Scholar] [CrossRef]
- Kik, C. Exploitation of wild relatives for the breeding of cultivated Allium species. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CABI Publishing Oxon: Oxford, UK, 2002; pp. 81–100. ISBN 9780851995106. [Google Scholar]
- Chuda, A. Hybridization and Molecular Characterization of F1 Allium Cepa × Allium Roylei Plants. Acta Biol. Cracoviensia Ser. Bot. 2012, 54, 25–31. [Google Scholar] [CrossRef]
- Shigyo, M.; Kik, C. Onion. In Vegetables II: Fabaceae, Liliaceae, Solanaceae, and Umbelliferae; Prohens-Tomás, J., Nuez, F., Handbook of Plant Breeding, Vegetables, Eds.; Springer: New York, NY, USA, 2008; pp. 121–159. ISBN 978-0-387-74108-6. [Google Scholar]
- Kohli, B.; Gohil, R.N. Need to conserve Allium roylei Stearn: a potential gene reservoir. Genet. Resour. Crop. Evol. 2009, 56, 891–893. [Google Scholar] [CrossRef]
- Kofoet, A.; Kik, C.; Wietsma, W.A.; Vries, J.N. Inheritance of Resistance to Downy Mildew (Peronospora destructor [Berk.] Casp.) from Allium roylei Stearn in the Backcross Allium cepa L. x (A. roylei x A. cepa). Plant Breed. 1990, 105, 144–149. [Google Scholar] [CrossRef]
- Scholten, O.E.; van Heusden, A.W.; Khrustaleva, L.I.; Burger-Meijer, K.; Mank, R.A.; Antonise, R.G.C.; Harrewijn, J.L.; Van Haecke, W.; Oost, E.H.; Peters, R.J.; et al. The long and winding road leading to the successful introgression of downy mildew resistance into onion. Euphytica 2007, 156, 345–353. [Google Scholar] [CrossRef]
- de Vries, J.N.; Wietsma, W.A.; de Vries, T. Introgression of leaf blight resistance from Allium roylei Stearn into onion (A. cepa L.). Euphytica 1992, 62, 127–133. [Google Scholar] [CrossRef]
- Rian, V.; Ford-Lloyd, S.; Armstrong, J. 5 - Welsh onion: Allium fistulosum L., In: Genetic Improvement of Vegetable Crops; Kalloo, G., Bergh, B.O., Eds.; Pergamon, 1993, pp. 51-58.
- Peffley, E.B.; Hou, A. Bulb-type onion introgressants posessing Allium fistulosum L. genes recovered from interspecific hybrid backcrosses between A. cepa L. and A. fistulosum L. Theor. Appl. Genet. 2000, 100, 528–534. [Google Scholar] [CrossRef]
- Yamashita, K.-I.; Takatori, Y.; Tashiro, Y. Chromosomal location of a pollen fertility-restoring gene, Rf, for CMS in Japanese bunching onion (Allium fistulosum L.) possessing the cytoplasm of A. galanthum Kar. et Kir. revealed by genomic in situ hybridization. Theor. Appl. Genet. 2005, 111, 15–22. [Google Scholar] [CrossRef]
- Pobożniak, M.; Leśniak, M.; Chuda, A.; Adamus, A. Field assessment of the susceptibility of onion cultivars to thrips attack – preliminary results. Pol. J. Èntomol. 2016, 85, 121–133. [Google Scholar] [CrossRef]
- Loomans, A. J. M.; Murai, T. Culturing thrips and parasitoids. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Wallingford, Oxon, UK, 1997; pp. 477–503. [Google Scholar]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Shelton, A.M. Evaluation of Onion Cultivars for Resistance to Onion Thrips (Thysanoptera: Thripidae) and Iris Yellow Spot Virus. J. Econ. Èntomol. 2010, 103, 925–937. [Google Scholar] [CrossRef]
- Mayhew, P.J. Adaptive Patterns of Host-Plant Selection by Phytophagous Insects. Oikos 1997, 79, 417. [Google Scholar] [CrossRef]
- Thompson, J.N. Evolutionary ecology of the relationship between oviposition preference and performance of offspring in phytophagous insects. Èntomol. Exp. Appl. 1988, 47, 3–14. [Google Scholar] [CrossRef]
- Mc Collum, G.D. Experimental Hybrids between Allium fistulosum and A. roylei. Bot. Gaz. 1982, 143, 238–242. [Google Scholar] [CrossRef]
- Hanafy, A.R.I.; Tahany, R.A.; Nowar, E.E.; Hasan, S.M. Effect of anatomical and phytochemical diversity of two onion cultivars on the infestation with onion thrips (Thysanoptera: Thripidae). Middle East J. Appl. Sci. 2016, 6, 941–948. [Google Scholar]
- da Silva, V.C.P.; Bettoni, M.M.; Bona, C.; Foerster, L.A. Morphological and chemical characteristics of onion plants (Allium cepa L.) associated with resistance to onion thrips. Acta Sci. Agron. 2014, 37, 85. [Google Scholar] [CrossRef]
- Brown, A.S.S.; Simmonds, M.S.J.; Blaney, W.M. Relationship between nutritional composition of plant species and infestation levels of thrips. J. Chem. Ecol. 2002, 28, 2399–2409. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, M.; Abd-El-Haliem, A.; Bleeker, P.; Dicke, M.; Escobar-Bravo, R.; Cheng, G.; A Haring, M.; Kant, M.R.; Kappers, I.; Klinkhamer, P.G.L.; et al. Thrips advisor: exploiting thrips-induced defences to combat pests on crops. J. Exp. Bot. 2018, 69, 1837–1848. [Google Scholar] [CrossRef]
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K.; et al. Plant Secondary Metabolites as Defense Tools against Herbivores for Sustainable Crop Protection. Int. J. Mol. Sci. 2022, 23, 2690. [Google Scholar] [CrossRef]
- Khalid, M.; Rahman, S.U.; Bilal, M.; Huang, D. Role of flavonoids in plant interactions with the environment and against human pathogens — A review. J. Integr. Agric. 2019, 18, 211–230. [Google Scholar] [CrossRef]
- Taylor, A.; Vagany, V.; Barbara, D.J.; Thomas, B.; Pink, D.A.C.; Jones, J.E.; Clarkson, J.P. Identification of differential resistance to sixFusarium oxysporumf. sp. cepaeisolates in commercial onion cultivars through the development of a rapid seedling assay. Plant Pathol. 2012, 62, 103–111. [Google Scholar] [CrossRef]
- Zheng, S. , Henken, B. , Wietsma, W. et al. Development of bio-assays and screening for resistance to beet armyworm (Spodoptera Exigua Hübner) in Allium cepa L. and its wild relatives. Euphytica 2000, 114, 77–85. [Google Scholar] [CrossRef]
- Hudák, K.; Pénzes, B. Factors influencing the population of the onion thrips on onion. Acta Phytopathol. et Èntomol. Hung. 2004, 39, 193–197. [Google Scholar] [CrossRef]
- Ren, X.; Wu, S.; Xing, Z.; Gao, Y.; Cai, W.; Lei, Z. Abundances of thrips on plants in vegetative and flowering stages are related to plant volatiles. J. Appl. Èntomol. 2020, 144, 732–742. [Google Scholar] [CrossRef]
- Jones, H.A.; Bailey, S.F.; Emsweller, S.L. Field Studies of Thrips tabaci Lind. With Especial Reference to Resistance in onions. J. Econ. Èntomol. 1935, 28, 678–680. [Google Scholar] [CrossRef]
- Porter, D.R. , Jones, H., A. Resistance of some cultivated species of Allium to pink root (Phoma terrestrs). Phytopatology 1933, 23(29), 298.
- Jones, H.A.; Mann, L.K. Onions and Their Allies; London Leonard Hill [Books] Limited Interscience Publishers, Inc.: New York, NY, USA, 1963; p. 284. [Google Scholar]
- Cryder, C.M. A study of the associations of heritable traits in progeny from the interspecific backcross (Allium fistulosum x Allium cepa L.) x Allium cepa L. Ph.D. Thesis, New Mexico State University, Las Cruces, USA, 1988. [Google Scholar]
- Sueyoshi, T.; Shimomura, K.; Koga, T.; Yamamura, Y.; Takemoto, H. The varietal difference in resistance to stone leek leaf miner in Welsh onions. Bull. Fukuoka Agric. Res. Cent. 2006, 25, 37–41. [Google Scholar]
- Takeda, M.; Kawai, A.; Mitsunaga, T.; Tsukazaki, H.; Yamashita, K.-I.; Wako, T. A novel method for evaluating the egg killing defenses and varietal resistance of the bunching onion against Liriomyza chinensis (Diptera: Agromyzidae) via the artificial inoculation of eggs. Appl. Èntomol. Zoöl. 2019, 55, 93–103. [Google Scholar] [CrossRef]
- Khandagale, K.; Roylawar, P.; Randive, P.; Karuppaiah, V.; Soumia, P.S.; Shirsat, D.; Gedam, P.; Ade, A.; Gawande, S.; Singh, M. Isolation and Expression Profiling of Insecticidal Lectins from Wild Alliums Against Onion Thrips (Thrips tabaci Lindeman). Proc. Natl. Acad. Sci., India, Sect. B Biol. Sci. 2020, 92(2), 451–459. [CrossRef]
- Tsukazaki, H.; Honjo, M.; Yamashita, K.-I.; Ohara, T.; Kojima, A.; Ohsawa, R.; Wako, T. Classification and identification of bunching onion (Allium fistulosum) varieties based on SSR markers. Breed. Sci. 2010, 60, 139–152. [Google Scholar] [CrossRef]
- Basri, R.; Ansari, M.S. Analytical study of phenotypic and biochemical attributes of onion cultivars in relation to infestation of onion thrips, Thrips tabaci. J. Asia-Pacific Èntomol. 2021, 24, 529–535. [Google Scholar] [CrossRef]
Figure 1.
Mean (± SE) number of hatched larvae of Thrips tabaci on cvs. Tęcza1, Alibaba1, Bila1, Kroll2 and Allium roylei. 1Allium cepa, 2 A. fistulosum. Means with the same letters on each bar do not differ significantly (Tukey HSD test, p< 0.05).
Figure 1.
Mean (± SE) number of hatched larvae of Thrips tabaci on cvs. Tęcza1, Alibaba1, Bila1, Kroll2 and Allium roylei. 1Allium cepa, 2 A. fistulosum. Means with the same letters on each bar do not differ significantly (Tukey HSD test, p< 0.05).
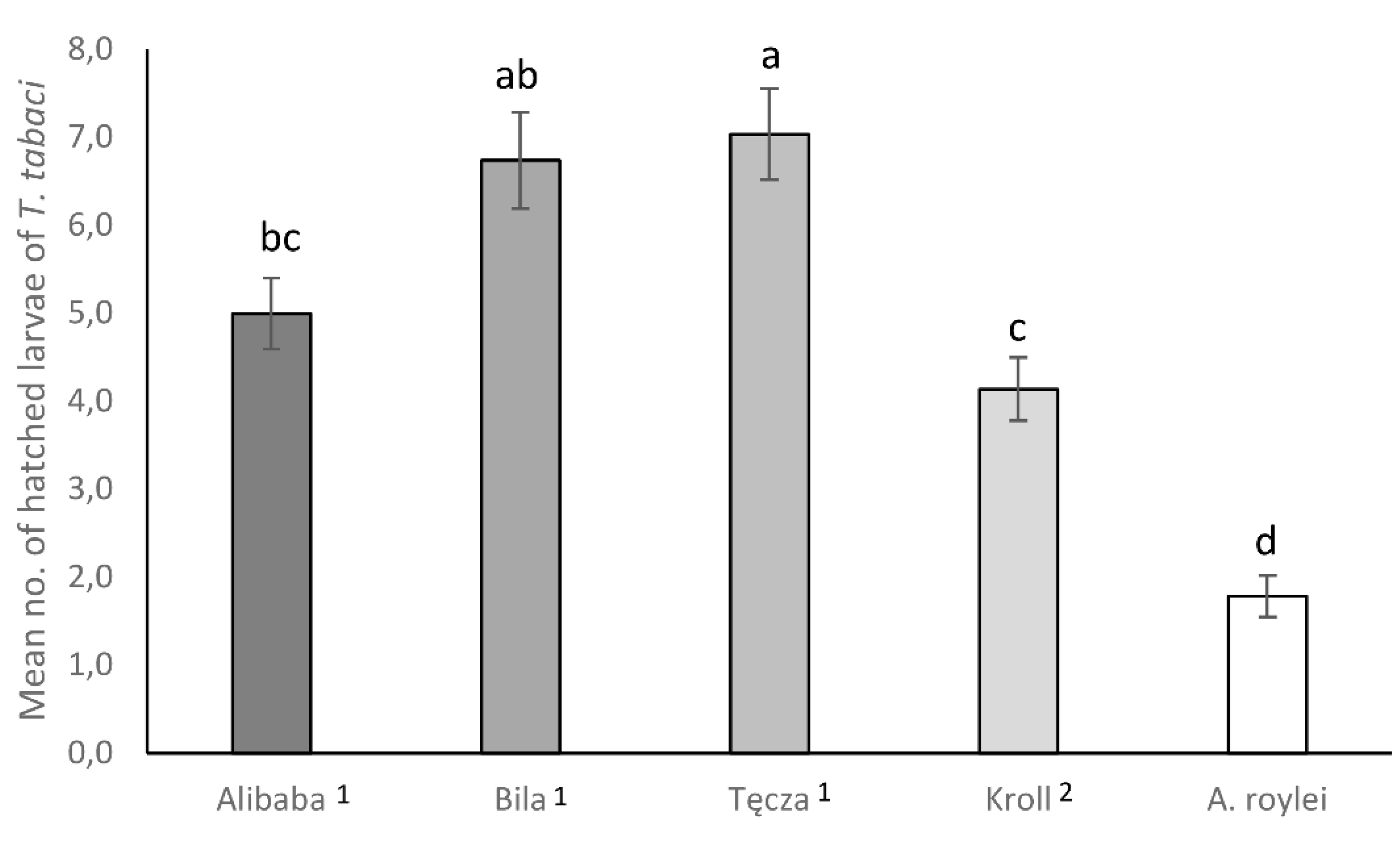
Figure 2.
Mean number (± SE) of Thrips tabaci females settled on leaf sections from two compared onion cultivars/species 10 min, 30 min and 1, 2, and 3 h after their release (* bars showing the significant difference in thrips settlement on the compared onion cultivars - Student t test at p <0.05); 1Allium cepa, 2A. fistulosum.
Figure 2.
Mean number (± SE) of Thrips tabaci females settled on leaf sections from two compared onion cultivars/species 10 min, 30 min and 1, 2, and 3 h after their release (* bars showing the significant difference in thrips settlement on the compared onion cultivars - Student t test at p <0.05); 1Allium cepa, 2A. fistulosum.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



