
Submitted:
31 August 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Abstract: Down Syndrome (DS) is a common genetic disorder characterized by an extra copy of chromosome 21, leading to dysregulation of various metabolic pathways. Oxidative stress in DS is associated with neurodevelopmental defects, neuronal dysfunction, and the onset of dementia re-sembling Alzheimer's disease. Additionally, chronic oxidative stress contributes to cardiovascular diseases and certain cancers prevalent in DS individuals. This study investigates the impact of ageing on oxidative stress and liver fibrosis using a DS murine model (Ts2Cje mice). The liver in DS mice shows increased oxidative stress and impaired antioxidant defenses, as evidenced by reduced glutathione levels and increased lipid peroxidation. Furthermore, DS liver exhibits an altered in-flammatory response as measured by the expression of cytokines and heat shock proteins. DS liver also displays dysregulated lipid metabolism, indicated by altered expression of peroxisome prolif-erator-activated receptors and fatty acid transport proteins. Consistently, these changes might contribute to non-alcoholic fatty liver disease development, a condition characterized by liver fat accumulation. Finally, histological analysis of DS liver reveals increased fibrosis and steatosis, in-dicative of potential progression to liver cirrhosis. This finding highlights the increased risk of liver pathologies in DS individuals, particularly when combined with the higher prevalence of obesity and metabolic dysfunctions in DS patients. These results shed light on the liver's role in DS-associated pathologies and suggest potential therapeutic strategies targeting oxidative stress and lipid metabolism to prevent or mitigate liver-related complications in DS individuals.
Keywords:
down syndrome
; oxidative stress
; liver
; aging
1. Introduction
Down Syndrome (DS) is the human most frequent aneuploidy, characterized by an extra complete or segment of chromosome 21 (trisomy 21), eventually triggering the dysregulation of several factors [1]. Besides the already characterized neurodevelopmental defects and neuronal dysfunction, it has been recently reported how DS individuals also display a plethora of conditions related to unbalanced reactive oxygen species (ROS) production [2]. These outcomes are strictly related to chromosome 21 triplication, triggering the overproduction of several proteins involved in ROS balancing as superoxide dismutase 1 (SOD1), the transcription factor BTB and CNC homology 1 (BACH1), the Protein C-ets-2 (ETS2), carbonyl reductase (CBR), S100 calcium-binding protein B (S100B) [3]. Furthermore, it has been reported that the master regulator of antioxidant response elements (ARE) nuclear factor (erythroid-derived 2)-like 2 (NRF2) is transcriptionally activated in DS individuals [4]. This effect has been reported to be related to the hyperphosphorylation of mitogen-activated protein kinases (MAPKs) in turn mediating NRF2 phosphorylation thus promoting its dissociation from Kelch-like ECH-Associating protein 1 (Keap1) but preventing its nuclear translocation [5]. The establishment of a highly oxidative environment might be responsible for several of the outcomes characterizing DS, including accelerated aging.
Therefore, individuals affected by DS display an increased risk of developing a type of dementia mimicking the clinical and pathological course of Alzheimer’s disease (AD) already at the age of 40s [6]. This outcome is strictly related to the triplication of genes associated with AD such as amyloid precursor protein (APP), beta secretase 2 (BACE2), and S100B [7], which along with the oxidative context characterizing DS promote the accumulation of amyloid beta-peptide (Ab) [3].
The chronic exposure to oxidative stress also accounts for several health conditions, including cardiovascular disease, and certain types of cancer characterizing DS individuals [8,9]. In this context the liver might turn to be one of the major organs to be involved, despite liver function in DS patients is poorly described, and the molecular basis remain elusive.
To date, it has been reported that DS liver is more prone to the development of several pathologies. Corroborating this, a vascular portohepatic anomaly in DS individuals has been characterized to impair the direct communication between the right portal vein and the inferior vena cava [10], potentially linking congenital vascular malformation and hepatic vascular shunt [11]. Furthermore, DS individuals show a significant enhanced production of hepatitis B antigen which might be responsible for the occurrence of the autoimmune hepatitis, a chronic and progressive inflammation of the liver from an unknown cause [12]. In addition to this, the alternated levels of several amino acids, along with several metabolites involved in the methylation cycle have been detected in blood samples derived from DS individuals, pointing out metabolic dysfunctions as one of the drivers of DS liver pathologies [13]. In agreement with this, DS individuals, already in childhood, are typically obese typically presenting dyslipidemia and hyperinsulinemia [14].
In this context, a correlation between DS and the onset of non-alcoholic fatty liver disease (NAFLD), a pathology characterized by the accumulation of fat in the liver has been described in a cohort of patients, and strictly related to the enhanced insulin resistance reported in DS individuals [13-15]. Other key favors to consider are several single nucleotide polymorphisms (SNPs), such as PNPLA3, TM6SF2, and Klotho eventually promoting hepatic fat accumulation [16]. NAFLD spectra ranges from steatosis through non-alcoholic steatohepatitis (NASH), fibrosis and cirrhosis. The latter has been defined as the end stage of a wide number of chronic liver conditions displaying necroinflammation, fibrosis, and regenerative nodules, eventually modifying liver architecture and impairing its functional mass and vascular organization [17]. Given the correlation standing between DS and NAFL occurrence, further research is needed to fully understand the genetic mechanisms underlying this association.
2. Materials and Methods
2.1. Mouse colony
Ts2Cje (Rb(12.Ts171665Dn)2Cje) mice are an established DS murine model displaying a triple copy of a Robertsonian fusion chromosome where the distal end of Chr16 and Chr12 are located. Parental generations were purchased from Jackson Laboratories (Bar Harbour, ME, USA). The mouse colony was raised by a crossbreed of Ts2Cje trisomic females with euploid (B6EiC3SnF1/J) F1 hybrid males (Eu). The parental generations were purchased from Jackson Laboratories (Bar Harbour, ME, USA). These breeding pairs produce litters containing both trisomic (Ts2Cje) and euploid (Eu) offspring. Pups were genotyped to determine trisomy by standard PCR, using Reinoldth’s method [18,19]. Mice were housed in clear Plexiglas cages (20 × 22 × 20 cm) under standard laboratory conditions with a temperature of 22 ± 2 °C and 70% humidity, a 12-h light/dark cycle, and free access to food and water, as previously described [19]. All the experiments were performed in strict compliance with the Italian National Laws (DL 116/92), and the European Communities Council Directives (86/609/EEC). The experimental protocol was approved by the Italian Ministry of Health (#1183/2016-PR). All efforts were made to minimize the number of animals used in the study and their suffering. Immediately after isolation, samples were put into liquid nitrogen and then stored at −80 °C until utilization.
2.2. GSH evaluation
GSH levels were assayed on tissues homogenized performing a spectrophotometric assay based on the reaction of thiol groups with 2,2-dithio-bis-nitrobenzoic acid at a wavelength of 412 nm (εM=13,600 M-1 cm-1, where εM is a wavelength-dependent molar absorptivity coefficient) [20]. Measurements were performed quantified using Synergy H1 (Biotek; Milan; Italy) in quadruplicate per each sample.
2.3. LOOH measurement
Lipid peroxide assay was performed as already reported [20]. Briefly, the reaction is based on the oxidation of Fe2+ to Fe3+ in the presence of xylenol orange at 560 nm. Measurements were performed quantified using Synergy H1 (Biotek; Milan; Italy) in quadruplicate per each sample.
2.4. RNA extraction and cDNA preparation
Tissue sections were resuspended in 500mL of PRImeZOL Reagent (#AN1100, Canvax Biotech, Andalusia, Spain). RNA extraction was then performed as previously described [21] and resuspended in RNase-free water. RNA was quantified using Synergy H1 (Biotek; Milan; Italy). 2ug of RNA from each sample was retrotranscribed using High-Capacity cDNA Reverse Transcription Kit (#4368814; Applied biosystem; Massachusetts, USA) according to manufacturer instructions.
2.5. Real time PCR
2.6. Western blot analysis
Tissue sections were homogenized for protein extraction. Therefore, we performed western blot analysis as described in [23]. Rabbit anti HSP90 (ab59459), HSP60 (ab46798), and GAPDH (ab8245) were purchased from Abcam (Cambridge, UK). Rabbit anti CLPP (PA5-52722) was purchased from Thermofisher Scientific (Massachusetts, USA). Secondary antibody anti Rabbit-HRP (ab6721) was purchased from Abcam (Cambridge, UK).
2.7. Histopathological analysis
Liver sections were formalin-fixed, paraffin-embedded and treated for histological examination using a standard method [24]. Two pathologists (R.C. and G.B.) separately evaluated all histological slides, blinded to sample identity. The following histological features were assessed on 5 micron-thick sections stained with hematoxylin and eosin and Masson’s trichrome, as previously described [25]: fibrosis, inflammation, steatosis and hepatocellular ballooning. Fibrosis was graded on a 0-3 scale: 0, absence of fibrosis; 1, portal spaces expanded by fibrosis with or without fibrous septa; 2, portal areas expanded by fibrosis with formation of fibrous bridges; 3, numerous fibrous bridges with formation of nodules. The following score was used to grade inflammation: 0, lack of inflammation; 1, periportal inflammation; 2, mild/moderate portal inflammation; 3, marked portal inflammation. Hepatocellular ballooning was scored on a 0-2 scale: 0, absent; 1, mild; 2, moderate/severe. The following 0-3 score was used to quantify steatosis: 0, absent; 1, mild; 2, moderate; 3, diffuse. For each case, the final histological score was performed by summing the scores of fibrosis, inflammation, steatosis, and ballooning.
2.8. Statistics
Data are shown as mean ± standard deviation (SD). Statistical analysis was performed by using Prism 8.0.2. software (GraphPad Software, San Diego, CA, USA). Significant differences were assessed using a one-way ANOVA or the student test, when needed. A value of p < 0.05 was considered statistically significant and symbols used to indicate statistical differences are described in figure legends.
3. Results
3.1. DS liver display an enhanced exposure to oxidative stress
DS individuals display an impaired ROS scavenging system, which might be also hampering liver homeostasis [2]. To investigate this outcome also in our mice model, we therefore assayed the accumulation of glutathione (GSH) and lipid peroxide (LOOH) in wild type (Eu) and DS Ts2Cje liver homogenized derived from 3-, 6-, 9-, and 12-months old mice (Figure 1). Ts2Cje, but not Eu, were characterized by a deficit of GSH at 12-months. Any significant change was evident at any stage before the 12-months (Figure 1A). LOOH levels, on the other hand, were increased in 12-months old Ts2Cje liver homogenized compared to the Eu counterpart (Figure 1B). Similar to GSH, also LOOH levels were not significatively changed at any of the time points assayed before (Figure 1B). Given this evidence, we next sought to investigate HMOX1 expression in 3-, 6-, 9-, and 12-months old mice models by qPCR. Our results show that there is a trend in increasing HMOX1 expression at 3-, 6-, and 9- months, but it turns to be significatively downregulated at 12-months in Ts2Cje livers compared to Eu counterpart, overall highlighting an important increase in oxidative stress in DS mice model liver, and an impairment of the antioxidant defenses.
3.2. DS mice livers show an increase of inflammatory markers
An increase in oxidative stress as the one we reported in Ts2Cje livers, might be correlated with an increase in the overall inflammatory status [26]. To investigate this point, we decided to assess by qPCR the expression of two crucial cytokines as interleukin 1 β (IL-1β) and interleukin 10 (IL-10) (Figure 2A). Interestingly, while IL-1β expression is decreased (Figure 2A, left panel) IL-10 transcription is increased in Ts2Cje liver compared to the Eu counterpart (Figure 2A, right panel). The induction of an inflammatory status is strictly linked to the accumulation of the heat shock proteins (Hsps) [27]. For this reason, we sought to investigate the accumulation of Hsp90 and Hsp60 in our model. Western blot analysis unveiled a marked accumulation of both Hsp90 and Hsp60 in Ts2Cje liver protein extract compared to Eu counterpart (Figure 2B; Supplemental Figure 1).
The increased inflammatory status has been reported to also affect the mitochondrial dynamics [28]. In this context, the Caseinolytic Mitochondrial Matrix Peptidase Proteolytic Subunit (Clpp) has been described as one of the master regulators of mitochondrial quality control system [29]. Interestingly, western blot analysis highlighted a marked decrease in Clpp protein level in Ts2Cje liver homogenized compared to Eu counterpart (Figure 2B; Supplemental Figure 1). Overall, these data describe a scenario in which the increase in oxidative stress triggers an inflammatory status activating Hsp machinery and possibly impairing mitochondrial quality system.
3.3. DS liver increase lipid metabolism
The decreased level of Clpp has been described to be one of the NASH hallmarks, a pathology strictly associated to liver lipid accumulation [29]. To assay lipid accumulation in our model we assayed the expression of two members of peroxisome proliferator-activated receptors (PPARs) family. Interestingly, the expression of both PPARα and PPARγ in Ts2Cje liver were markedly decreased compared to the Eu counterpart (Figure 3). Furthermore, qPCR analysis also described a marked increase in fatty acid transport protein-5 (FATP5) and Carnitine palmitoyltransferase 2 (CTP2) in Ts2Cje, but not in Eu livers (Figure 3). Taken together these data we unveil an increase in fatty acids metabolism in DS mice livers.
3.4. Ds livers display an increased fibrosis
The accumulation of lipids within the hepatic tissue might drive the shift from NASH toward cirrhosis [30]. This process is characterized by the accumulation of collagen. To test this outcome, we performed a qPCR testing Col1a1 expression, which was significantly increased in Ts2Cje livers compared to the Eu counterparts (Figure 4A).
Furthermore, we also performed a histological analysis on Ts2Cje and Eu liver sections, evaluating the fibrosis percentage (Figure 4B-C). Histologically, Eu group showed lack of fibrosis and hepatocellular ballooning, along with low levels of steatosis and inflammation compared to Ts2Cje counterpart (Figure 4B-C). Conversely, the Ts2Cje group fibrosis formation with, at least focal, fibrous septa/bridging and a more widespread inflammation and steatosis than controls (Figure 4B-C). Taken together, these data report an increased fibrotic rate in DS liver compared to healthy controls.
4. Discussion
Trisomy of chromosome 21 is the signature of DS, impairing the cognitive and intellectual abilities of individuals affected by this pathology. In the latest year it has come to attention that DS individuals are characterized by changes also on their metabolic profile [13] eventually affecting the homeostasis of several organs, including the liver, which has not been deeply investigated so far.
For this purpose, we decided to investigate the liver of DS mice model, which were previously generated and characterized by Barone and Perlugi’s lab [19][31-33]. Interestingly, we found that 12-months old mice liver were characterized by a decreased GSH concentration, a well-known antioxidant molecule [34]. The decreased level of antioxidant defenses prompted us to investigate the concentration of LOOH in this context. We therefore highlight an increased oxidative environment in DS 12-months old mice liver. This outcome has been reported to be related to the dysregulation of different factor occurring in the ROS balancing, as recently reviewed by our group [3]. Furthermore, we also reported a significant decrease in HMOX1 expression in 12-months old livers. This data correlates with the increased Bach1 protein level reported in the brain of DS subject, and further mirrored in Ts2Cje mice model, by Perluigi group [31,35]. Bach1 is a master transcription repressor having HMOX1 as one of the main targets [35,36]. In particular, the HO1 protein play a role in the complex pathophysiological cascade involved in insulin resistance mechanisms, oxidative stress, metabolic syndrome and cardiovascular diseases [36-38].The rewire of the oxidative balance in DS livers might also have a crucial role in rewiring the inflammatory response of this organ. Interestingly, we report here a repression of IL-1β transcription, along with an increase of IL-10 expression. As recently reported, an enhancement of IL-10 expression inhibits IL-1β production [39]. This outcome might be related to the increased susceptibility displayed by DS individuals in the development of several infections, given the lack of an inflammatory response [40].
To further assess the inflammatory response in our model, we evaluated the protein accumulation of Hsp90, which was decreased in DS-livers. Despite any significative change in Hsp90 accumulation within DS brains has been reported [41]. Furthermore, its downregulation might be related to an impaired inflammatory response, corroborating the previous data on IL-1β and IL-10 expression [42]. In addition to this, we also reported a decrease in the protein levels of the mitochondrial shock protein Hsp60. This data is in line with a previous report describing a deficit in Hsp60 on skin fibroblasts derived from DS individuals [43]. Interestingly, Hsp60 is the major mitochondrial heat shock protein, in charge for preventing protein aggregation following ROS unbalancing [44]. Therefore, we sought to assess the protein accumulation of one of the master regulators of mitochondrial fitness, Clpp, which we found to be upregulated in our DS model. It has been reported that under high fat diet, Clpp downregulation correlates with an increased protection against obesity and hepatic steatosis, also preventing insulin resistance [45,46]. Furthermore, Clpp is also part of the machinery preventing hepatocytes senescence [45]. Therefore, we speculate that the upregulation of this protein might be linked to the increased insulin resistance extensively described in DS individuals [13]. Furthermore, it might also be part of a compensatory effect preventing cellular senescence and liver cirrhosis, typical of DS individuals. To further investigate this point, we sought to perform a real-time PCR aiming to assess the expression level of PPARα and PPARγ, two ligand-activated transcription factors part of the nuclear hormone receptor superfamily, in turn in charge for regulating adipogenesis and insulin resistance [47]. Interestingly, the downregulation of PPARα and PPARγ, as the one we unveiled in our mode, might be correlated to the accumulation of lipid droplets within hepatocytes, also contributing to the insulin resistance reported in DS individuals [48,49]. The accumulation of lipid droplets in DS liver is further supported by our data showing an increased expression of FATP5 and CTP2 in 12-months old liver compared to the wild-type counterpart. The former is in charge for the uptake of long-chain fatty acids [50]. Corroborating this, its expression has been reported to be inversely correlated to NAFLD progression [51]. The increase in fatty acids uptake might also be correlated to increased CTP2 expression, in turn in charge for initiating fatty acids oxidation eventually promoting their clearance [52]. In this context, fatty acids accumulation might work as a driver for the onset of liver fibrosis. In agreement with this, we reported an enhanced expression of Col1a1 in 12-month-old DS mice liver. Therefore, we decided to assess the fibrotic rate of wild-type and DS livers by histopathology. Interestingly, our results corroborated a scenario in which DS livers display higher fibrosis and hepatocellular ballooning, together with increased steatosis and inflammation. Overall, these data unveil a significant increased risk for DS individuals to develop liver pathologies. Interestingly, liver failure has been reported to DS newborns showing transient abnormal myelopoiesis, a pathology characterized by transient appearance of blast cells and eventually also affecting liver homeostasis [53]. Furthermore, a diffuse lobular fibrosis around proliferating ductular elements and residual hepatocytes, as the one characterizing our DS models, was reported in DS newborns individuals presenting a severe liver disease [54]. These effects are further enhanced by the higher obesity rate reported in DS individuals, in turn promoting NAFLD onset by accumulation of hepatic fatty acids [55].
Overall, our work unveils a scenario in which DS liver is characterized by an impaired ROS scavenging system, locally unbalancing the oxidative stress level, and leading to an impaired response to inflammatory stimuli. This outcome correlates with the increased fibrotic rate reported in this context, along with the accumulation of fatty lipids, mirroring NAFLD profile. Given this evidence, the usage of antioxidants compounds might therefore represent a novel strategy in DS treatment to prevent liver-associated pathologies.
Author Contributions
Conceptualization, M.P., E.B., A.M.A. and I.A.B.; methodology, S.G., V.T., J.F., G.B., E.T.; software, S.G., R.C., G.B.; validation, S.G., V.T., J.F., E.T.; formal analysis, S.G., A.T., I.A.B; investigation, M.P., E.B., I.A.B. ; resources, A.M.A.; data curation, R.C., A.D.; writing—original draft preparation, S.G., R.C., G.B., A.T.; writing—review and editing, S.G., R.C., A.M.A., M.P.,E.B., I.A.B.; visualization, A.D., E.T.; supervision, M.P., E.B., I.A.B.; project administration, A.M.A, I.A.B.; funding acquisition, A.M.A. All authors have read and agreed to the published version of the manuscript.
Funding
This researcher was funded by the researchers supporting project number (RSP2023R261) King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Antonarakis, S.E.; Skotko, B.G.; Rafii, M.S.; Strydom, A.; Pape, S.E.; Bianchi, D.W.; Sherman, S.L.; Reeves, R.H. Down syndrome. Nat Rev Dis Primers 2020, 6, 9. [Google Scholar] [CrossRef]
- Campos, C.; Casado, A. Oxidative stress, thyroid dysfunction & Down syndrome. Indian J Med Res 2015, 142, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; Butterfield, D.A. Oxidative Stress and Down Syndrome: A Route toward Alzheimer-Like Dementia. Curr Gerontol Geriatr Res 2012, 2012, 724904. [Google Scholar] [CrossRef]
- Giallongo, S.; Rehakova, D.; Raffaele, M.; Lo Re, O.; Koutna, I.; Vinciguerra, M. Redox and Epigenetics in Human Pluripotent Stem Cells Differentiation. Antioxid Redox Signal 2021, 34, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; Tramutola, A.; Pagnotta, S.; Barone, E.; Butterfield, D.A. The BACH1/Nrf2 Axis in Brain in Down Syndrome and Transition to Alzheimer Disease-Like Neuropathology and Dementia. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Glasson, E.J.; Sullivan, S.G.; Hussain, R.; Petterson, B.A.; Montgomery, P.D.; Bittles, A.H. The changing survival profile of people with Down’s syndrome: implications for genetic counselling. Clin Genet 2002, 62, 390–393. [Google Scholar] [CrossRef]
- Giallongo, S.; Longhitano, L.; Denaro, S.; D’Aprile, S.; Torrisi, F.; La Spina, E.; Giallongo, C.; Mannino, G.; Lo Furno, D.; Zappala, A.; et al. The Role of Epigenetics in Neuroinflammatory-Driven Diseases. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, K.; Constantine, A.; Clift, P.; Condliffe, R.; Moledina, S.; Jansen, K.; Inuzuka, R.; Veldtman, G.R.; Cua, C.L.; Tay, E.L.W.; et al. Cardiovascular Complications of Down Syndrome: Scoping Review and Expert Consensus. Circulation 2023, 147, 425–441. [Google Scholar] [CrossRef]
- Rabin, K.R.; Whitlock, J.A. Malignancy in children with trisomy 21. Oncologist 2009, 14, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Kieran, M.W.; Vekemans, M.; Robb, L.J.; Sinsky, A.; Outerbridge, E.W.; Der Kaloustian, V.M. Portohepatic shunt in a Down syndrome patient with an interchange trisomy 47,XY,-2,+der(2),+der(21)t(2;21)(p13;q22.1)mat. Am J Med Genet 1992, 44, 288–292. [Google Scholar] [CrossRef]
- Burdall, O.C.; Grammatikopoulos, T.; Sellars, M.; Hadzic, N.; Davenport, M. Congenital Vascular Malformations of the Liver: An Association With Trisomy 21. J Pediatr Gastroenterol Nutr 2016, 63, e141–e146. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.P.; Kaye, G.; Clarke, A.C. Autoimmune hepatobiliary disease in trisomy 21. J Clin Gastroenterol 2000, 30, 330–332. [Google Scholar] [CrossRef]
- Dierssen, M.; Fructuoso, M.; Martinez de Lagran, M.; Perluigi, M.; Barone, E. Down Syndrome Is a Metabolic Disease: Altered Insulin Signaling Mediates Peripheral and Brain Dysfunctions. Front Neurosci 2020, 14, 670. [Google Scholar] [CrossRef] [PubMed]
- Valentini, D.; Alisi, A.; di Camillo, C.; Sartorelli, M.R.; Crudele, A.; Bartuli, A.; Nobili, V.; Villani, A. Nonalcoholic Fatty Liver Disease in Italian Children with Down Syndrome: Prevalence and Correlation with Obesity-Related Features. J Pediatr 2017, 189, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Valentini, D.; Mosca, A.; Di Camillo, C.; Crudele, A.; Sartorelli, M.R.; Scoppola, V.; Tarani, L.; Villani, A.; Raponi, M.; Novelli, A.; et al. PNPLA3 gene polymorphism is associated with liver steatosis in children with Down syndrome. Nutr Metab Cardiovasc Dis 2020, 30, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Crudele, A.; Dato, S.; Re, O.L.; Maugeri, A.; Sanna, P.; Giallongo, S.; Oben, J.; Panera, N.; De Rango, F.; Mosca, A.; et al. Pediatric Non-Alcoholic Fatty Liver Disease Is Affected by Genetic Variants Involved in Lifespan/Healthspan. J Pediatr Gastroenterol Nutr 2021, 73, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, C.; Zhan, Y.T. Nonalcoholic Fatty Liver Disease Cirrhosis: A Review of Its Epidemiology, Risk Factors, Clinical Presentation, Diagnosis, Management, and Prognosis. Can J Gastroenterol Hepatol 2018, 2018, 2784537. [Google Scholar] [CrossRef] [PubMed]
- Reinholdt, L.G.; Ding, Y.; Gilbert, G.J.; Czechanski, A.; Solzak, J.P.; Roper, R.J.; Johnson, M.T.; Donahue, L.R.; Lutz, C.; Davisson, M.T. Molecular characterization of the translocation breakpoints in the Down syndrome mouse model Ts65Dn. Mamm Genome 2011, 22, 685–691. [Google Scholar] [CrossRef]
- Tramutola, A.; Lanzillotta, S.; Aceto, G.; Pagnotta, S.; Ruffolo, G.; Cifelli, P.; Marini, F.; Ripoli, C.; Palma, E.; Grassi, C.; et al. Intranasal Administration of KYCCSRK Peptide Rescues Brain Insulin Signaling Activation and Reduces Alzheimer’s Disease-like Neuropathology in a Mouse Model for Down Syndrome. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Nibali, L.; Rizzo, M.; Li Volti, G.; D’Aiuto, F.; Giglio, R.V.; Barbagallo, I.; Pelekos, G.; Donos, N. Lipid subclasses profiles and oxidative stress in aggressive periodontitis before and after treatment. J Periodontal Res 2015, 50, 890–896. [Google Scholar] [CrossRef]
- Longhitano, L.; Giallongo, S.; Orlando, L.; Broggi, G.; Longo, A.; Russo, A.; Caltabiano, R.; Giallongo, C.; Barbagallo, I.; Di Rosa, M.; et al. Lactate Rewrites the Metabolic Reprogramming of Uveal Melanoma Cells and Induces Quiescence Phenotype. Int J Mol Sci 2022, 24. [Google Scholar] [CrossRef] [PubMed]
- Longhitano, L.; Broggi, G.; Giallongo, S.; Failla, M.; Puzzo, L.; Avitabile, T.; Tibullo, D.; Distefano, A.; Pittala, V.; Reibaldi, M.; et al. Heme Oxygenase-1 Overexpression Promotes Uveal Melanoma Progression and Is Associated with Poor Clinical Outcomes. Antioxidants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Cambria, D.; Longhitano, L.; La Spina, E.; Giallongo, S.; Orlando, L.; Giuffrida, R.; Tibullo, D.; Fontana, P.; Barbagallo, I.; Nicoletti, V.G.; et al. IGFBP-6 Alters Mesenchymal Stromal Cell Phenotype Driving Dasatinib Resistance in Chronic Myeloid Leukemia. Life (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Broggi, G.; Lo Giudice, A.; Di Mauro, M.; Asmundo, M.G.; Pricoco, E.; Piombino, E.; Caltabiano, R.; Morgia, G.; Russo, G.I. SRSF-1 and microvessel density immunohistochemical analysis by semi-automated tissue microarray in prostate cancer patients with diabetes (DIAMOND study). Prostate 2021, 81, 882–892. [Google Scholar] [CrossRef]
- Mangano, K.; Cavalli, E.; Mammana, S.; Basile, M.S.; Caltabiano, R.; Pesce, A.; Puleo, S.; Atanasov, A.G.; Magro, G.; Nicoletti, F.; et al. Involvement of the Nrf2/HO-1/CO axis and therapeutic intervention with the CO-releasing molecule CORM-A1, in a murine model of autoimmune hepatitis. J Cell Physiol 2018, 233, 4156–4165. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front Physiol 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Moseley, P.L. Heat shock proteins and the inflammatory response. Ann N Y Acad Sci 1998, 856, 206–213. [Google Scholar] [CrossRef]
- Missiroli, S.; Genovese, I.; Perrone, M.; Vezzani, B.; Vitto, V.A.M.; Giorgi, C. The Role of Mitochondria in Inflammation: From Cancer to Neurodegenerative Disorders. J Clin Med 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.E.; Hwang, Y.; Lee, S.J.; Jung, H.; Shin, T.H.; Son, Y.; Park, S.; Han, S.J.; Kim, H.J.; Lee, K.W.; et al. Mitochondrial protease ClpP supplementation ameliorates diet-induced NASH in mice. J Hepatol 2022, 77, 735–747. [Google Scholar] [CrossRef]
- Pei, K.; Gui, T.; Kan, D.; Feng, H.; Jin, Y.; Yang, Y.; Zhang, Q.; Du, Z.; Gai, Z.; Wu, J.; et al. An Overview of Lipid Metabolism and Nonalcoholic Fatty Liver Disease. Biomed Res Int 2020, 2020, 4020249. [Google Scholar] [CrossRef]
- Pagnotta, S.; Tramutola, A.; Barone, E.; Di Domenico, F.; Pittala, V.; Salerno, L.; Folgiero, V.; Caforio, M.; Locatelli, F.; Petrini, S.; et al. CAPE and its synthetic derivative VP961 restore BACH1/NRF2 axis in Down Syndrome. Free Radic Biol Med 2022, 183, 1–13. [Google Scholar] [CrossRef]
- Zuliani, I.; Lanzillotta, C.; Tramutola, A.; Francioso, A.; Pagnotta, S.; Barone, E.; Perluigi, M.; Di Domenico, F. The Dysregulation of OGT/OGA Cycle Mediates Tau and APP Neuropathology in Down Syndrome. Neurotherapeutics 2021, 18, 340–363. [Google Scholar] [CrossRef] [PubMed]
- Lanzillotta, C.; Zuliani, I.; Tramutola, A.; Barone, E.; Blarzino, C.; Folgiero, V.; Caforio, M.; Valentini, D.; Villani, A.; Locatelli, F.; et al. Chronic PERK induction promotes Alzheimer-like neuropathology in Down syndrome: Insights for therapeutic intervention. Prog Neurobiol 2021, 196, 101892. [Google Scholar] [CrossRef] [PubMed]
- Averill-Bates, D.A. The antioxidant glutathione. Vitam Horm 2023, 121, 109–141. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, F.; Pupo, G.; Mancuso, C.; Barone, E.; Paolini, F.; Arena, A.; Blarzino, C.; Schmitt, F.A.; Head, E.; Butterfield, D.A.; et al. Bach1 overexpression in Down syndrome correlates with the alteration of the HO-1/BVR-a system: insights for transition to Alzheimer’s disease. J Alzheimers Dis 2015, 44, 1107–1120. [Google Scholar] [CrossRef]
- Barbagallo, I.; Marrazzo, G.; Frigiola, A.; Zappala, A.; Li Volti, G. Role of carbon monoxide in vascular diseases. Curr Pharm Biotechnol 2012, 13, 787–796. [Google Scholar] [CrossRef]
- Barbagallo, I.; Galvano, F.; Frigiola, A.; Cappello, F.; Riccioni, G.; Murabito, P.; D’Orazio, N.; Torella, M.; Gazzolo, D.; Li Volti, G. Potential therapeutic effects of natural heme oxygenase-1 inducers in cardiovascular diseases. Antioxid Redox Signal 2013, 18, 507–521. [Google Scholar] [CrossRef]
- Barbagallo, I.; Nicolosi, A.; Calabrese, G.; David, S.; Cimino, S.; Madonia, M.; Cappello, F. The role of the heme oxygenase system in the metabolic syndrome. Curr Pharm Des 2014, 20, 4970–4974. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, J.; Li, D.; Li, P.; Zhou, X.; Li, Y.; He, Z.; Qin, L.; Liang, L.; Luo, X. Interleukin-10 inhibits interleukin-1beta production and inflammasome activation of microglia in epileptic seizures. J Neuroinflammation 2019, 16, 66. [Google Scholar] [CrossRef] [PubMed]
- Broers, C.J.; Gemke, R.J.; Morre, S.A.; Weijerman, M.E.; van Furth, A.M. Increased production of interleukin-10 in children with Down syndrome upon ex vivo stimulation with Streptococcus pneumoniae. Pediatr Res 2014, 75, 109–113. [Google Scholar] [CrossRef]
- Yoo, B.C.; Vlkolinsky, R.; Engidawork, E.; Cairns, N.; Fountoulakis, M.; Lubec, G. Differential expression of molecular chaperones in brain of patients with Down syndrome. Electrophoresis 2001, 22, 1233–1241. [Google Scholar] [CrossRef]
- Ambade, A.; Catalano, D.; Lim, A.; Kopoyan, A.; Shaffer, S.A.; Mandrekar, P. Inhibition of heat shock protein 90 alleviates steatosis and macrophage activation in murine alcoholic liver injury. J Hepatol 2014, 61, 903–911. [Google Scholar] [CrossRef]
- Bozner, P.; Wilson, G.L.; Druzhyna, N.M.; Bryant-Thomas, T.K.; LeDoux, S.P.; Wilson, G.L.; Pappolla, M.A. Deficiency of chaperonin 60 in Down’s syndrome. J Alzheimers Dis 2002, 4, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Horwich, A.L.; Hartl, F.U. Prevention of protein denaturation under heat stress by the chaperonin Hsp60. Science 1992, 258, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Dabravolski, S.A.; Bezsonov, E.E.; Orekhov, A.N. The role of mitochondria dysfunction and hepatic senescence in NAFLD development and progression. Biomed Pharmacother 2021, 142, 112041. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, S.; Pharaoh, G.; Ranjit, R.; Murphy, A.; Matsuzaki, S.; Nair, B.C.; Forbes, B.; Gispert, S.; Auburger, G.; Humphries, K.M.; et al. Loss of mitochondrial protease ClpP protects mice from diet-induced obesity and insulin resistance. EMBO Rep 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Gervois, P.; Torra, I.P.; Fruchart, J.C.; Staels, B. Regulation of lipid and lipoprotein metabolism by PPAR activators. Clin Chem Lab Med 2000, 38, 3–11. [Google Scholar] [CrossRef]
- Wang, M.; Yan, Y.; Zhang, Z.; Yao, X.; Duan, X.; Jiang, Z.; An, J.; Zheng, P.; Han, Y.; Wu, H.; et al. Programmed PPAR-alpha downregulation induces inflammaging by suppressing fatty acid catabolism in monocytes. iScience 2021, 24, 102766. [Google Scholar] [CrossRef] [PubMed]
- Leonardini, A.; Laviola, L.; Perrini, S.; Natalicchio, A.; Giorgino, F. Cross-Talk between PPARgamma and Insulin Signaling and Modulation of Insulin Sensitivity. PPAR Res 2009, 2009, 818945. [Google Scholar] [CrossRef]
- Doege, H.; Baillie, R.A.; Ortegon, A.M.; Tsang, B.; Wu, Q.; Punreddy, S.; Hirsch, D.; Watson, N.; Gimeno, R.E.; Stahl, A. Targeted deletion of FATP5 reveals multiple functions in liver metabolism: alterations in hepatic lipid homeostasis. Gastroenterology 2006, 130, 1245–1258. [Google Scholar] [CrossRef] [PubMed]
- Enooku, K.; Tsutsumi, T.; Kondo, M.; Fujiwara, N.; Sasako, T.; Shibahara, J.; Kado, A.; Okushin, K.; Fujinaga, H.; Nakagomi, R.; et al. Hepatic FATP5 expression is associated with histological progression and loss of hepatic fat in NAFLD patients. J Gastroenterol 2020, 55, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Parimbelli, M.; Pezzotti, E.; Negro, M.; Calanni, L.; Allemano, S.; Bernardi, M.; Berardinelli, A.; D’Antona, G. Nutrition and Exercise in a Case of Carnitine Palmitoyl-Transferase II Deficiency. Front Physiol 2021, 12, 637406. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Sotomatsu, M.; Ohki, K.; Arai, K.; Maruyama, K.; Kobayashi, T.; Nishi, A.; Sameshima, K.; Takagi, T.; Hayashi, Y. Liver disease is frequently observed in Down syndrome patients with transient abnormal myelopoiesis. Int J Hematol 2014, 99, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Ruchelli, E.D.; Uri, A.; Dimmick, J.E.; Bove, K.E.; Huff, D.S.; Duncan, L.M.; Jennings, J.B.; Witzleben, C.L. Severe perinatal liver disease and Down syndrome: an apparent relationship. Hum Pathol 1991, 22, 1274–1280. [Google Scholar] [CrossRef] [PubMed]
- De Matteo, A.; Vajro, P. Down Syndrome and Pediatric Nonalcoholic Fatty Liver Disease: A Causal or Casual Relationship? J Pediatr 2017, 189, 11–13. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
DS mice are characterized by an increased oxidative stress. (A) GSH levels are decreased in Ts2Cje 12-months old mice liver. Representative scheme for GSH quantitation in Eu and Ts2Cje livers obtained from 3-, 6-, 9-, and 12- months old mice. (B) LOOH levels are increased in Ts2Cje 12-months old mice liver. Histograms representative of LOOH spectrophotometric evaluation on Eu and TS2Cje livers obtained from 3-, 6-, 9-, and 12- months old mice. (C) HMOX1 expression is downregulated in Ts2Cje 12-months old livers. Representative histogram of the real-time PCR against HMOX1 on the liver extract obtained from 3-, 6-, 9-, and 12- months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. Histograms are representative of four different experiments (*P≤0.05; ***P≤0.001).
Figure 1.
DS mice are characterized by an increased oxidative stress. (A) GSH levels are decreased in Ts2Cje 12-months old mice liver. Representative scheme for GSH quantitation in Eu and Ts2Cje livers obtained from 3-, 6-, 9-, and 12- months old mice. (B) LOOH levels are increased in Ts2Cje 12-months old mice liver. Histograms representative of LOOH spectrophotometric evaluation on Eu and TS2Cje livers obtained from 3-, 6-, 9-, and 12- months old mice. (C) HMOX1 expression is downregulated in Ts2Cje 12-months old livers. Representative histogram of the real-time PCR against HMOX1 on the liver extract obtained from 3-, 6-, 9-, and 12- months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. Histograms are representative of four different experiments (*P≤0.05; ***P≤0.001).
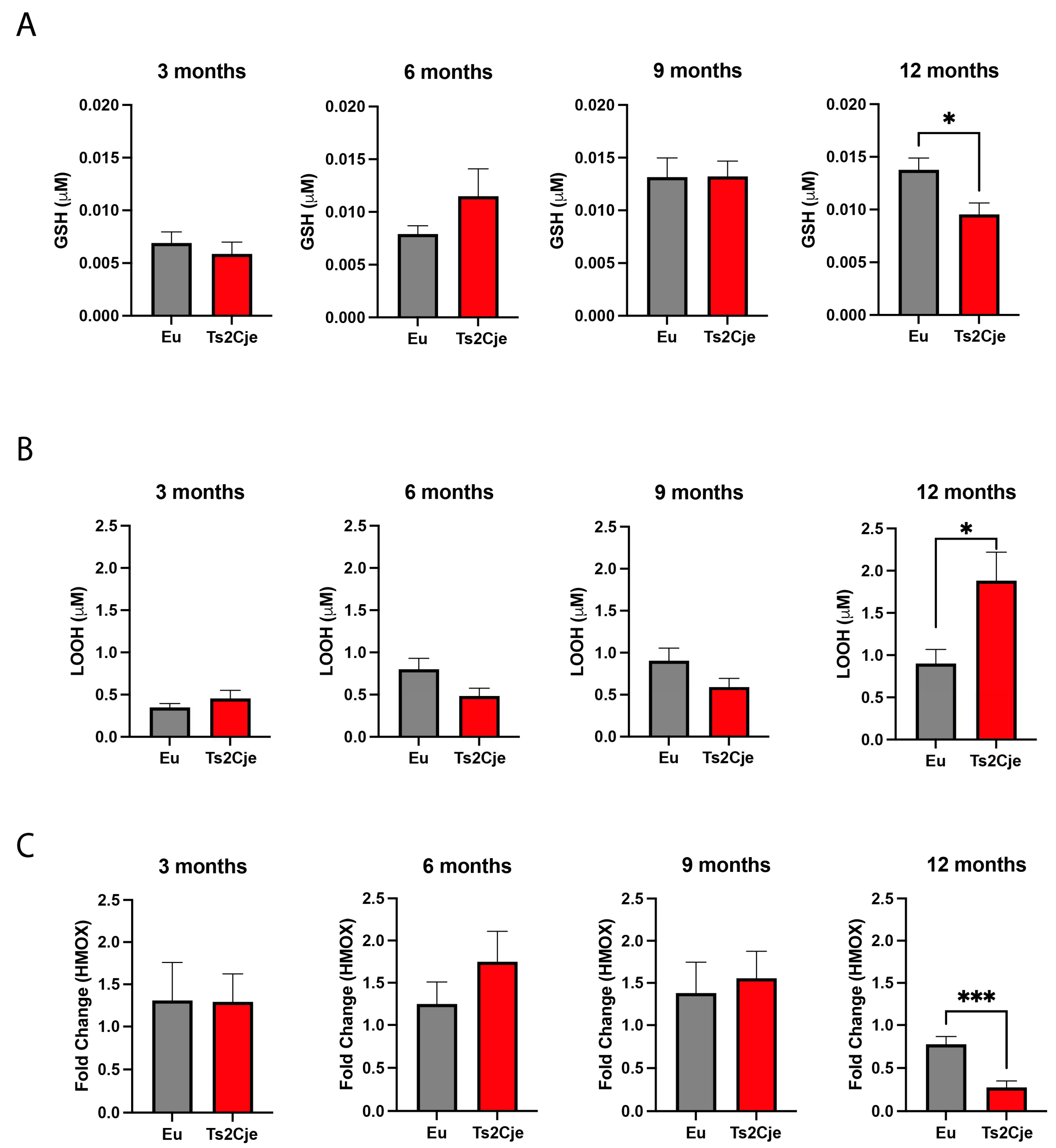
Figure 2.
DS mice show a marked increase in liver inflammation. (A) IL-1β and IL10 are differently regulated in Ts2Cje livers. Representative histogram of the real-time PCR against IL-1β and IL-10 on the liver extract obtained from 12-months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. (B) Ts2Cje mice showed an increased Hsp90, Hsp60 levels, while Clpp accumulation is decreased. Western Blot analysis on 12-months old Eu and Ts2Cje liver extracts against Hsp90, Hsp60, and Clpp. GAPDH was used as housekeeping protein. Histograms are representative of four different experiments (*P≤ 0.05; **P≤0.01).
Figure 2.
DS mice show a marked increase in liver inflammation. (A) IL-1β and IL10 are differently regulated in Ts2Cje livers. Representative histogram of the real-time PCR against IL-1β and IL-10 on the liver extract obtained from 12-months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. (B) Ts2Cje mice showed an increased Hsp90, Hsp60 levels, while Clpp accumulation is decreased. Western Blot analysis on 12-months old Eu and Ts2Cje liver extracts against Hsp90, Hsp60, and Clpp. GAPDH was used as housekeeping protein. Histograms are representative of four different experiments (*P≤ 0.05; **P≤0.01).
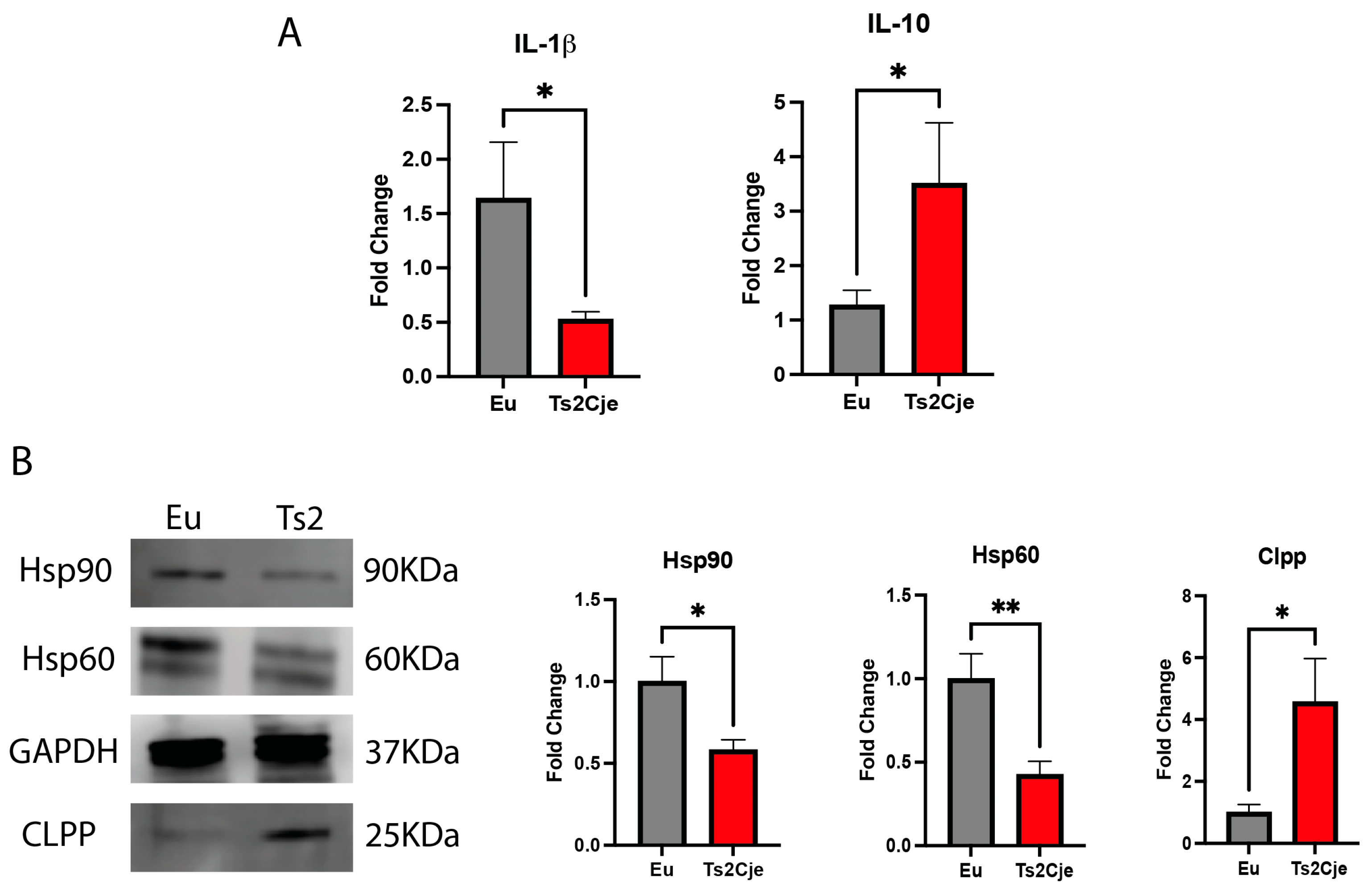
Figure 3.
DS mice have a marked increase in lipid metabolism. PPARα, PPARγ, FATP5, and CTP2 are differently regulated in Ts2Cje livers. Representative histogram of the real-time PCR against PPARα, PPARγ, FATP5, and CTP2 on the liver extract obtained from 12-months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. Histograms are representative of four different experiments (*P≤ 0.05).
Figure 3.
DS mice have a marked increase in lipid metabolism. PPARα, PPARγ, FATP5, and CTP2 are differently regulated in Ts2Cje livers. Representative histogram of the real-time PCR against PPARα, PPARγ, FATP5, and CTP2 on the liver extract obtained from 12-months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. Histograms are representative of four different experiments (*P≤ 0.05).
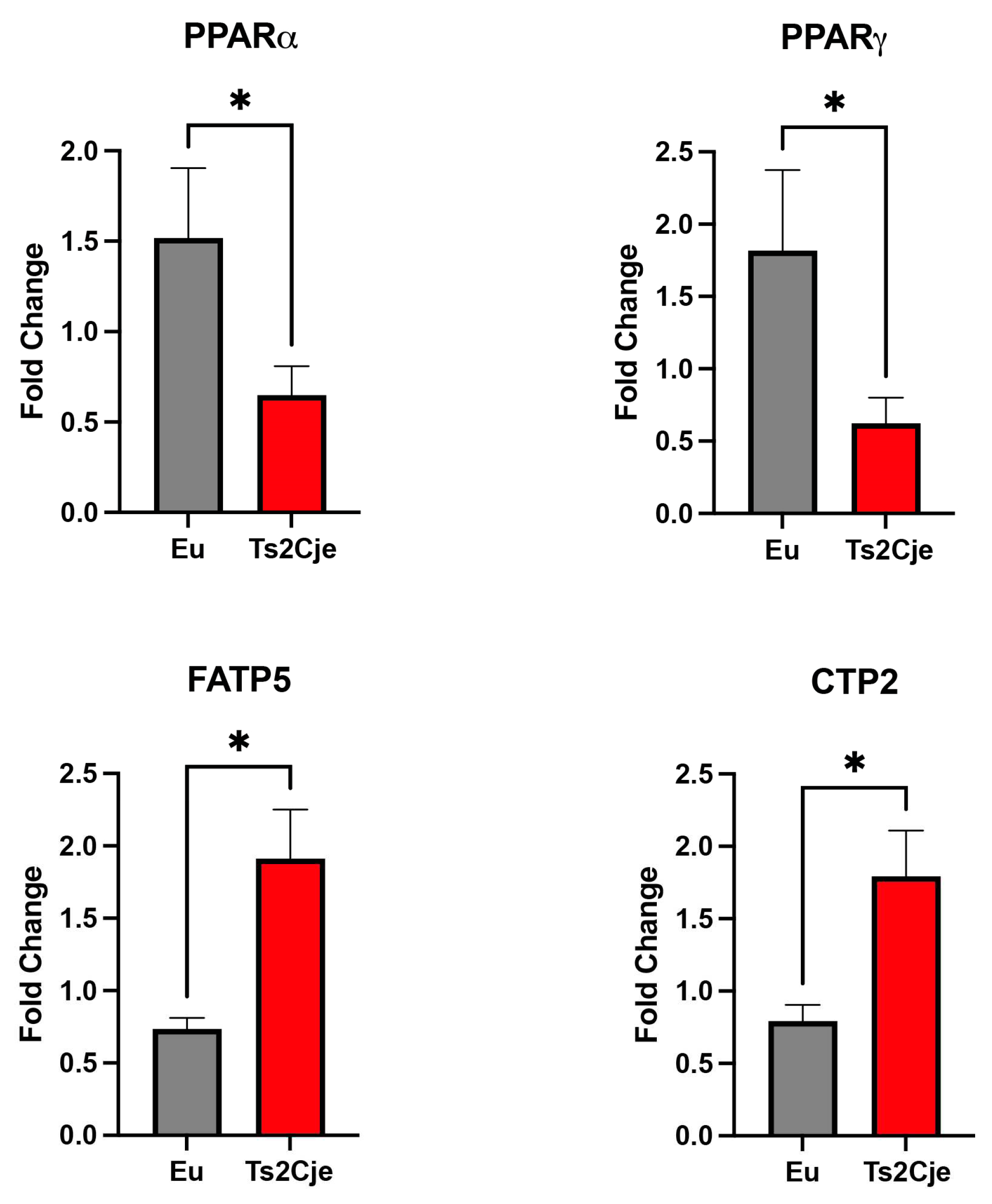
Figure 4.
DS mice show a marked increase in fibrotic markers. (A) Col1a1 expression level is increased in Ts2Cje livers. Representative histogram of the real-time PCR against Col1a1 on the liver extract obtained from 12-months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. (B) Ts2Cje liver section show an increased fibrotic level. Representative histological images of the rat livers from control and Ts2Cje group. i) Mouse liver from control group showing absence of steatosis (score 0) and mild periportal inflammation (score 1) (hematoxylin and eosin; original magnification 100x). ii) Mouse liver from Ts2Cje group exhibiting moderate steatosis (score 2) and moderate portal inflammation (score 2) (hematoxylin and eosin; original magnification 150x). iii) Mice liver from control group showing absence of fibrosis (score 0) (Masson’s trichrome; original magnification 50x). iv) Mice liver from Ts2Cje group exhibiting diffuse fibrosis (score 3) with fibrous bridging (arrows) (Masson’s trichrome; original magnification 50x). (C) Histological score quantified as in B. Histograms are representative of four different experiments (*P≤ 0.05; ***P≤ 0.001).
Figure 4.
DS mice show a marked increase in fibrotic markers. (A) Col1a1 expression level is increased in Ts2Cje livers. Representative histogram of the real-time PCR against Col1a1 on the liver extract obtained from 12-months old Eu and Ts2Cje mice. GAPDH was used as housekeeping gene. (B) Ts2Cje liver section show an increased fibrotic level. Representative histological images of the rat livers from control and Ts2Cje group. i) Mouse liver from control group showing absence of steatosis (score 0) and mild periportal inflammation (score 1) (hematoxylin and eosin; original magnification 100x). ii) Mouse liver from Ts2Cje group exhibiting moderate steatosis (score 2) and moderate portal inflammation (score 2) (hematoxylin and eosin; original magnification 150x). iii) Mice liver from control group showing absence of fibrosis (score 0) (Masson’s trichrome; original magnification 50x). iv) Mice liver from Ts2Cje group exhibiting diffuse fibrosis (score 3) with fibrous bridging (arrows) (Masson’s trichrome; original magnification 50x). (C) Histological score quantified as in B. Histograms are representative of four different experiments (*P≤ 0.05; ***P≤ 0.001).

Table 1.
Real time PCR primers’ list.
| Gene | Forward 5′ --> 3′ | Reverse 5′ --> 3′ | Accession Number |
|---|---|---|---|
| Hmox1 | TGACACCTGAGGTCAAGCAC | CAGCTCCTCAAACAGCTCAATG | NM_010442.2 |
| Il1β | TGCCACCTTTTGACAGTGATG | CGTCACACACCAGCAGGTTA | NM_008361.4 |
| Il10 | GTAGAAGTGATGCCCCAGGC | GACACCTTGGTCTTGGAGCTTATT | NM_010548.2 |
| Ppar𝛂 | TGCCTTCCCTGTGAACTGAC | CACAGAGCGCTAAGCTGTGA | NM_001113418.1 |
| Ppar𝛄 | GGTCAGTCATGGAACAGCCA | TTCTGGGAGAGGTCTGCAC | NM_001411509.1 |
| Fatp5 | TGTAACGTCCCTGAGCAACC | TAAGCCCACATTGCCCTCTG | NM_009512.2 |
| Cpt2 | GAATGACAGCCAGTTCAGGAAG | GCATGCAGCTCCTTCCCAAT | NM_009949.2 |
| Col1a1 | CCCTGGTCCCTCTGGAAATG | GGACCTTTGCCCCCTTCTTT | NM_007742.4 |
| Gapdh | AACCCTTAAGAGGGATGCTGC | TCTACGGGACGAGGAAACAC | NM_001289726.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



