
Submitted:
31 August 2023
Posted:
05 September 2023
You are already at the latest version
Abstract
Anthracnose (ANT) and angular leaf spot (ALS) are significant diseases in common bean, leading to considerable yield losses under specific environmental conditions. The California Dark Red Kidney (CDRK) bean cultivar is known for its resistance to multiple races of both pathogens. Previous studies have identified the CoPv01CDRK/PhgPv01CDRK resistance loci on chromosome Pv01. Here, we evaluated the expression levels of ten candidate genes near the CoPv01CDRK/PhgPv01CDRK loci and plant defense genes using quantitative real-time PCR in CDRK cultivar inoculated with races 73 of Colletotrichum lindemuthianum and 63-39 of Pseudocercospora griseola. Gene expression analysis revealed that the Phvul.001G246300 gene exhibited the most elevated levels, showing remarkable 7.8-fold and 8.5-fold increases for ANT and ALS, respectively. The Phvul.001G246300 gene encodes an abscisic acid (ABA) receptor pyrabactin resistance - PYR1-like (PYL) protein, which plays a central role in crosstalk between ABA and jasmonic acid responses. Interestingly, our results also showed that the other defense genes were initially activated. These findings provide critical insights into the molecular mechanisms underlying plant defense against these diseases and could contribute to the development of more effective disease management strategies in the future.
Keywords:
CoPv01CDRK/PhgPv01CDRK loci
; candidate gene expression
; common bean-anthracnose interaction
; common bean-angular leaf spot interaction
; plant defense genes
1. Introduction
Colletotrichum lindemuthianum (Sacc. & Magnus) Briosi & Cavara is a highly destructive pathogen that causes anthracnose (ANT) in common beans. In conditions of low temperature and high humidity, ANT can result in up to 100% yield losses. Furthermore, the pathogenic variability of C. lindemuthianum and the emergence of new races have resulted in the reduction or total loss of yield of previously resistant cultivars [1]. Angular leaf spot (ALS) is another disease that impacts common beans globally, caused by Pseudocercospora griseola (Sacc.) Crous & U. Braun. This disease can lead to yield losses of up to 70% [2-4]. Adopting genetically resistant cultivars provides a cost-effective, user-friendly, and environmentally conscious strategy for managing C. lindemuthianum and P. griseola infections in common beans [4]. Consequently, identifying and molecularly characterizing resistance genes is crucial for enhancing resistance efficacy and durability [5-8].
The Co and Phg loci associated with ANT and ALS, respectively, are often found in disease-resistance clusters on various chromosomes. Although several independent genes confer resistance to C. lindemuthianum, most resistance genes found in Andean cultivars have been mapped to the common bean chromosome Pv01. The Co-1, Co-12, Co-13, Co-14, Co-15 and Co-1HY alleles of the Co-1 genes are present in the cultivars Michigan Dark Red Kidney (MDRK), Kaboon, Perry Marrow, AND 277, Widusa and Hongyundou, respectively [9-12]. Other resistance genes were mapped to the end of Pv01: Co-x in Jalo EEP558, Co-AC in Amendoim Cavalo [13, 14]. Moreover, the CoPv01CDRK/PhgPv01CDRK gene in CDRK, which confers resistance to C. lindemuthianum races 73, 2047, and 3481, as well as race 63-39 of P. griseola, was recently fine-mapped to Pv01 [15].
A single dominant resistance gene primarily confers resistance to the ALS pathogen; however, recent studies have also identified quantitative resistance loci (QRLs) [16-17]. To date, five resistance loci have been mapped, including three independent loci, Phg-1, Phg-2, and Phg-3, located on chromosomes Pv01, Pv08, and Pv04, respectively [9, 18-19]. Additionally, two major QRLs, Phg-4, and Phg-5, have been found on Pv04 and Pv10 [16-17, 20-21].
Candidate genes for ANT resistance loci have been analyzed thorough gene expression analysis to infer functionality in resistant cultivars [11, 22-23]. Generally, examining the expression of the candidate and disease-resistance genes can reveal their roles and interactions, thereby contributing to our understanding of how these genes collaborate in effective resistance responses. Chen [11] evaluated the expression analysis of four candidate genes at the Co-1HY allele in the Hongyundou cultivar inoculated with C. lindemuthianum race 81. The authors observed significant induction of all genes at an early stage. However, expression levels decreased at 24 hours post inoculation (hpi) and beyond. In the susceptible cultivar, high expression was only observed at 120 hpi, suggesting that delayed gene expression might facilitate pathogen penetration and proliferation, ultimately leading to disease development.
To elucidate the precise timing and magnitude of expression related to resistance against C. lindemuthianum race 73 Mahiya-Farooq [22] analysed the expression of four candidate genes within the Co-1 locus using near-isogenic lines that differ in the presence of the Co-12 resistance allele. They observed that the Phvul.001G243800 gene exhibited substantially higher expression levels, nearly 144-fold, in the resistant near-isogenic line. The molecular basis of the ANT resistance Co-x locus was established by sequencing a 58-kb target region in the Jalo EEP558 cultivar. The KTR2/3 gene was identified as an additional gene within a CRINKLY4 kinase cluster between the candidate genes Phvul.001G243600 and Phvul.001G243700. Gene expression analysis demonstrated that KTR2/3 was upregulated in Jalo EEP558 at 24 hours post-inoculation in plants inoculated with strain 100 of C. lindemuthianum [13, 23].
Phaseolus vulgaris CDRK from the breeding program at the University of California Davis is a landrace collected around Sacramento, CA, USA [24]. CDRK is resistant to Andean races 2, 39, 55 and Mesoamerican races 9, 64, 65, 73, 89, 1545, 2047, and 3481 of C. lindemuthianum [15]. Through fine mapping, a previous study delimited CoPv01CDRK/PhgPv01CDRK in a genomic region of 33 Kb on chromosome Pv01 and detected five candidate genes: Phvul.001G246000 (ATP-dependent RNA helicase), Phvul.001G246100 (cation-dependent mannose-6-phosphate receptor), Phvul.001G246200 (protein trichome birefringence-like 33), Phvul.001G246300 (abscisic acid (ABA) receptor PYL5), and Phvul.001G246400 (SNF2 domain-containing protein class 1-related). Additionally, the candidate genes Phvul.001G245300 and Phvul.001G246800, encode putative leucine-rich repeat protein kinases, are close to the CoPv01CDRK/PhgPv01CDRK loci [15].
In summary, protein expression studies regarding the ANT resistance locus have identified the candidate genes Phvul.001G243800 for Co-12, KTR2/3 for Co-x and Phvul.001G243600 and Phvul.001G243700 for Co-1HY which showed high expression levels under inoculation conditions [11, 13, 22-23]. All of these loci have been mapped onto Pv01. However, allelism test revealed that CoPv01CDRK/PhgPv01CDRK is not allelic to Co-1 [15]. Moreover, the physical distances between CoPv01CDRK/PhgPv01CDRK and Co-1, Co-x, Co-1HY are 211 kb, 193 kb, and 181 kb, respectively. Given these considerations, the objective of this study was to identify specific candidate gene in the CDRK cultivar effective against C. lindemuthianum race 73 and P. griseola race 63-39 using gene expression analysis employing quantitative real-time PCR.
2. Results
2.1. Phenotypic evaluation of cultivars
The results of inoculation of C. lindemuthianum race 73 at 120 hpi onto the resistant cultivar CDRK and also the susceptible control cultivar Yolano are displayed in Figure 1. The disease in Yolano appears as small water-soaked lesions on the underside of the leaf and small sunken lesions on the stem and eventually leads to plant death. In contrast, no symptoms or hypersensitive response was observed in the resistant cultivar CDRK. Inoculations results of Pseudocercospora griseola race 63-39 onto CDRK resistant cultivar and Yolano susceptible cultivar 216 hpi are also displayed in Figure 1. The symptoms observed in Yolano cultivar were angular lesions in leaf area leading to early defoliation and plant death.
2.2. Differential expression of candidate and defense genes in the CDRK cultivar inoculated with race 73 of C. lindemuthianum
We conducted an in-depth exploration into the expression patterns of the following candidate genes: KTR2/3, Phvul.001G243800, Phvul.001G244300, Phvul.001G244400, Phvul.001G244500, Phvul.001G245300, Phvul.001G246000, Phvul.001G246100, Phvul.001G246200, and Phvul.001G246300. These genes are located in regions that overlap with Co-x, Co-12, Co-AC, and CoPv01CDRK/PhgPv01CDRK on the Pv01 chromosome. Additionally, defense genes PR1a, PR1b, and PR2, were evaluated to identify the molecular basis of CDRK resistance upon C. lindemuthianum race 73 inoculations.
Detailed information about the evaluated genes, accompanied by their functional annotations, can be found in Table 1. Table 2 shows the magnitude of the levels of gene expression within the candidate and defense genes. Particularly, the gene Phvul.001G246300 standout among all the candidate genes in the CDRK cultivar. This gene exhibited a remarkable 7.2-fold expression at 24 hpi in the resistant CDRK cultivar. As the interaction progressed to 72 hpi, the gene Phvul.001G246300 showed significantly heightened expression; 7.8-fold. Moreover, the expression of Phvul.001G246300 remained constant across multiple time points: 24, 48, 72, 96, and 120 hpi (Figure 2A, Figure 3 and Table 2).
Expression of the candidate gene Phvul.001G245300 revealed approximately 3.7-fold at 24, 72, 96 and 120 hpi (Figure 2B), and Phvul.001G246200 expression increased 2.7-fold at 24, 48 and 72 hpi (Figure 2C and Table 2). KTR2/3 expression exhibit higher expression only at 72 hpi, a 3.1-fold change (Figure 2D and Table 2). Although these genes exhibited some different expression levels in relation to the mock, they did not show large differences, as observed in Phvul.001G246300 or in pathogen-related genes.
The candidate genes Phvul.001G243800, Phvul.001G246100 and Phvul.001G246000 had the lowest levels of relative expression. The Phvul.001G243800 gene exhibited 2.0-fold change in relative expression at 72 and 96 hpi (Figure 2E and Table 2). Phvul.001G246100 and Phvul.001G246000 displayed an average of 1-fold change in relative expression (Figure 2F,G and Table 2). The candidate genes for resistance in Amendoim Cavalo, namely, Phvul.001G244500, Phvul.001G244400, and Phvul.001G244300, were not expressed in CDRK plants after inoculation of race 73 in relation to mock plants, indicating that these genes may not be involved in the CoPv01CDRK/PhgPv01CDRK response to C. lindemuthianum race 73 (Figure 2H–J, Figure 3 and Table 2).
Regarding the pathogenesis-related genes PR1b (Phvul.006G196900), PR2 (Phvul.009G256400), and PR1a (Phvul.003G109100), their reaction to the pathogen was high, with their expression surging by more than 6-fold and, in some cases, even up to 16.7-fold post-inoculation (Figure 4, Figure 5 and Table 2). Among them, PR1b (Phvul.006G196900) stood out as the most responsive to the pathogen, exhibiting a substantial 13.6-fold increase in expression from 24 to 96 hpi and a remarkable 16.7-fold increase at 120 hpi (Figure 4, Figure 5 and Table 2). The gene PR2 (Phvul.009G256400) showed a moderate level of expression and response upon exposure to the pathogen. Remarkably, there was a 10-fold increase in expression observed at both 24 and 48 hpi, alongside 11-fold at 72, 96, and 120 hpi, (Figure 4B). Similarly, the gene PR1a (Phvul.003G109100) exhibited a significant 6.2 to 8.1-fold increases in expression following inoculation (Figure 4C). These findings shed light on the distinct patterns of gene response to the pathogen, indicating that Phvul.009G256400 and Phvul.003G109100 may play crucial roles in plant defenses.
2.3. Differential expression of candidate and defense genes in CDRK cultivar inoculated with P. griseola race 63-39
Upon inoculating the CDRK cultivar with race 63-39 of P. griseola, significant alterations in the relative expression levels of candidate genes for the CoPv01CDRK/ PhgPv01CDRK loci and resistance genes were observed. Among the candidate genes assessed at the CoPv01CDRK/PhgPv01CDRK loci, the Phvul.001G246300 gene showcased the utmost responsiveness to race 63-39 P. griseola in the CDRK cultivar, as illustrated in Figures 6A and 7. Notably, this gene exhibited a substantial increase in expression at 72 hpi, reaching a fold change of up to 8.5-fold (Figure 6A). Furthermore, at 120 and 216 hpi, the gene maintained a consistent mean fold change of 6.1, indicating sustained response to the pathogen. These findings emphasize the potential significance of Phvul.001G246300 in the response of the CDRK cultivar to P. griseola race 63-39.
The gene Phvul.001G246200 showed induction only at 72 hpi, with an increase of nearly 1.5-fold (Figure 6B). As for the KTR2/3 gene, it exhibited induction at 72 and 120 hpi, with a fold change increase of 0.8, but was subsequently downregulated at 216 hpi (Figure 6C). The gene Phvul.001G246000 demonstrated minimal changes in expression levels at 72 and 120 hpi (Figure 6D), while Phvul.001G244300 exhibited similar patterns with few changes at 120 and 168 hpi (Figure 6E). The Phvul.001G246100 gene displayed induction from 72 to 168 hpi, albeit with a small increase of 0.3-fold (Figure 6F).
On the contrary, the Phvul.001G244400 gene displayed downregulation at 216 hpi, with a reduction of 1-fold (Figure 6G). The Phvul.001G243800 gene exhibited downregulation at 24 hpi, with a reduction of 1-fold (Figure 6H). Phvul.001G245300 demonstrated downregulation at 24 and 168 hpi, followed by induction at 120 and 216 hpi, but with a slight increase of 0.4 and 0.7-fold (Figure 6I). The gene Phvul.001G244500 consistently showed downregulation at all evaluated time points, with a significant reduction of approximately 2.3-fold in response to the pathogen (Figure 6J). These observations provide valuable insights into the dynamic expression patterns of these genes in response to the pathogen, shedding light on their potential roles in the defense mechanisms of CDRK plants.
When the CDRK cultivar was inoculated with race 63-39 of P. griseola, pathogen-related genes exhibited high expression responses, as evidenced in Figure 8 and Figure 9. The PR2 gene displayed a significant increase in expression at 72 hpi, reaching peak induction levels at 120 and 216 hpi with fold increases around 5.3. At 24 and 168 hpi, there was a slight increase in the expression level of this gene, with an average fold change above 1.9 (Figure 8A and Table 2). The PR1a gene demonstrated an increase in gene expression starting from 24 hpi and peaking at 216 hpi, with a fold increase of nearly 6 in response to the pathogen (Figure 8B). The PR1b gene exhibited induction at 120 and 216 hpi, with fold changes above 3 and 4, respectively. However, at 24 and 168 hpi, the expression level of this gene decreased compared to 72, 120, and 216 hpi, but no differences were observed compared to the mock (Figure 8C). Overall, the PR1a and PR2 genes showed similar expression patterns and were more responsive to race 63-39 of P. griseola than the PR1b gene (Figure 9).
3. Discussion
This study discusses the cellular mechanisms employed by plants to combat unfavorable conditions caused by biotic factors. These responses are complex networks that involve changes in gene expression, regulation of metabolic processes, reinforcement of the plant cell wall, and hormone signaling pathways. In particular, the majority of resistance genes identified encode NBS-LRR proteins, which consist of an amino-terminal signaling domain, a nucleotide-binding site (NBS), and carboxy-terminal leucine-rich repeats (LRRs). These NBS-LRR proteins are capable of recognizing pathogen effectors through protein-protein interactions, subsequently triggering effector-triggered immunity [25-26]. However, it is noteworthy that the common bean chromosome Pv01 exhibits a low abundance of NBS-LRR genes. Instead, this chromosome harbors genes that encode other proteins involved in the resistance response, such as kinases functioning as pattern-recognition receptors (PRRs) that recognize pathogen-associated molecular patterns (PAMPs) and initiate PAMP-triggered immunity (PTI).
In this research we assessed the expression levels of candidate genes near the CoPv01CDRK/PhgPv01CDRK loci and plant defense genes in CDRK cultivar. Through this analysis, we identified genes that consistently manifested high expression in resistant plants under pathogen inoculation. We observed that the candidate gene Phvul.001G246300 as a potential candidate for the CoPv01CDRK/PhgPv01CDRK resistance loci in CDRK plants inoculated with C. lindemuthianum race 73, as it demonstrated the highest relative expression among the candidate genes. Among the tested candidate genes, the expression of Phvul.001G246300 demonstrated a significant 7.8-fold increase at 24, 48, 72, 96, and 120 hpi of race 73 of C. lindemuthianum. Moreover, it exhibited an 8.5-fold change at 72 hpi and over a 6-fold change at 120 and 216 hpi with race 63-39 of P. griseola, indicating its heightened responsiveness to both pathogens compared to the other candidate genes examined. This particular gene encodes an abscisic acid (ABA) receptor, PYL5 protein, which is known to play a crucial role in mediating the crosstalk between ABA and jasmonic acid (JA) responses [27]. ABA has been implicated in plant defense against pathogens and shows synergistic interactions with the ethylene (ET) signaling pathway. The upregulation of the PYR receptor during biotic stress suggests its involvement in perceiving ABA and initiating downstream signaling mediated by kinases [28].
The Phvul.001G245300 gene ranked second in terms of induction among the candidate genes for CoPv01CDRK/PhgPv01CDRK loci in plants inoculated with race 73 of C. lindemuthianum. However, its expression was only half that of Phvul.001G246300, and when the plants were inoculated with race 63-39 of P. griseola, this gene showed a less pronounced response, with higher expression observed only at 216 hpi. Hypothetically, this gene encodes a protein belonging to the protein kinase superfamily, which is predominantly composed of catalytic domains of serine/threonine-specific and tyrosine-specific protein kinases. Furthermore, the protein contains a Leucine-Rich Repeat (LRR) domain. Proteins containing LRRs, including tyrosine kinase receptors, are involved in diverse biological processes such as signal transduction, cell adhesion, DNA repair, recombination, transcription, RNA processing, apoptosis, disease resistance, and immune responses [29, 30].
The Phvul.001G246200 gene ranked third among the upregulated genes, mainly between 24 and 72 hpi with race 73 of C. lindemuthianum, but its induction was three times lower than that of Phvul.001G246300. When plants were inoculated with race 63-39 of P. griseola, this gene exhibited a modest upregulation at 72 hpi, five times less than Phvul.001G246300. In Arabidopsis thaliana, a homologous gene plays a crucial role in xylan acetylation and the proper deposition of secondary walls [31]. Acetylation of wall polymers is important for cell wall strength and disease resistance, as evidenced by several Arabidopsis mutants and overexpression lines [31, 32]. Therefore, the Phvul.001G246200 gene may contribute to the resistance response in the CDRK cultivar by modifying cell wall strength, thereby impeding pathogen infection and/or colonization of plant tissues during the biotrophic life stage.
The majority of the ANT resistance genes identified in Andean cultivars are located in a resistance cluster on the common bean chromosome Pv01, including Co-1, Co-12, Co-13, Co-14, Co-15, Co-1HY, Co-1X, Co-x, Co-AC, and CoPv01CDRK/PhgPv01CDRK (Figure 10). Interestingly, the resistance gene from the Jalo EEP558 cultivar, Co-x, conferring resistance against race 3993 of C. lindemuthianum, is located in close proximity to CoPv01CDRK/PhgPv01CDRK loci. The protein KTR2/3 has been identified as the controlling factor for resistance of Co-x in Jalo EEP558 cultivar [23]. In our study with the CDRK cultivar, we observed that the expression of the KTR2/3 gene was 2.6 times lower than that of Phvul.001G246300 at 72 hpi in response to race 73 of C. lindemuthianum. Furthermore, in response to race 63-39 of P. griseola, the KTR2/3 gene exhibited a remarkably lower expression level, approximately 10.6 times lower, compared to Phvul.001G246300. Taking together, the physical position where CoPv01CDRK/PhgPv01CDRK and Co-x were mapped, and different candidate genes significantly upregulated after inoculation in each cultivar, KTR2/3 for Jalo EEP558 and Phvul.001G246300 for CDRK its possible to conclude that different genetic resistance are involved in each cultivar against the same pathogen.
Using near-isogenic lines with differing resistance alleles, expression analysis of candidate genes for the Co-12 allele against C. lindemuthianum race 73 resistance revealed high levels of Phvul.001G243800 in the resistant NIL [22]. Furthermore, based on transcriptional analysis, it was observed that Phvul.001G243700, located near the Co-1 locus, was differentially upregulated in the resistant NIL at 72 and 96 hpi after race 73 inoculation [33]. In the present study, significant differences in the expression of the Phvul.001G246300 and Phvul.001G243800 genes in response to race 73 of C. lindemuthianum and race 63-39 of P. griseola were observed in CDRK cultivar. At 72 hpi with race 73 of C. lindemuthianum we observed a notable expression of the Phvul.001G246300 gene, which was approximately three times higher compared to Phvul.001G243800. Remarkably, within the context of P. griseola race 63-39, we observed that Phvul.001G243800 gene was repressed, on the other hand, the expression of Phvul.001G246300 gene exhibited a substantial increase of more than 8-fold.
Overall, our findings underscore the contrasting expression patterns of the Phvul.001G246300 and Phvul.001G243800 genes within the CDRK cultivar, shedding explanation on their potential roles in the defense mechanisms against C. lindemuthianum race 73 and P. griseola race 63-39. It is worth noting that the Co-1 and CoPv01CDRK/PhgPv01CDRK loci were accurately mapped to separate regions towards the terminal end of the common bean chromosome Pv01, positioned 211 kb apart. Consequently, it is of paramount importance to emphasize that the CDRK cultivar harbors a distinct and independent gene from the Co-1 locus.
The pathogenesis-related defense genes PR1a, PR1b, and PR2 exhibited significant responsiveness to the pathogen. Notably, PR1b displayed the highest level of responsiveness to pathogen race 73, with a pronounced increase in expression at 120 hpi, reaching up to 16.7-fold higher than the control. Both PR2 and PR1a displayed elevated expression levels, exhibiting 11-fold and 8-fold increases, respectively, compared to the mock. Notably, PR2 maintained consistently high expression levels from 24 hpi to 120 hpi in response to the pathogen. Mahiya-Farooq et al. [22] reported early expression of plant defense genes in the resistant NIL, with PR1b and PR2 showing accumulation at 24 hpi of C. lindemuthianum race 73. Similarly, Shams et al. [34] observed higher expression of PR2 in the Naz resistant bean cultivar upon inoculation with C. lindemuthianum race 2. Although to a lesser extent, these pathogenesis-related defense genes were also upregulated in plants inoculated with race 63-39 of P. griseola. PR1a and PR1b showed higher induction at 216 hpi, with nearly a 6-fold increase and over a 4-fold increase in gene expression, respectively. The PR2 gene exhibited a more than 5-fold increase at 120 and 216 hpi. Interestingly, at 168 hpi, all the genes displayed reduced or no responsiveness to the pathogen.
These results reveal that the ANT and ALS resistance genes in the CDRK cultivar are controlled independently from those previously identified at the Co-1 locus. This indicates that the robust resistance against ANT and ALS in CDRK is manifested through the heightened response of the candidate gene Phvul.001G246300 to the respective pathogens. These findings point out the complex nature of plant-pathogen interactions, emphasizing the significance of comprehending gene expression mechanisms. Our findings might contribute to an enhanced comprehension of novel and efficient strategies for the development of cultivars resistant to angular leaf spot and ANT.
4. Materials and Methods
4.1. Plant material and growth conditions
Two experiments were conducted using a completely randomized design. In Experiment I, seedlings from both the resistant (R) CDRK and the susceptible (S) Yolano cultivars were inoculated with race 73 of C. lindemuthianum. The isoline derived from the CDRK × Yolano cross, namely CY 19, was used as a positive control for disease scoring to ensure that the fungus was virulent on them and that the absence of disease on the resistant genotypes was not attributed to the fungus lacking virulence. Additionally, the CY 70 isoline was used as a resistant control. The relative expression levels of 13 specific genes only in CDRK cultivar were evaluated at multiple time points: 24, 48, 72, 96, and 120 hours post inoculation (hpi), as well as in the mock. In Experiment II, a parallel approach was adopted, involving the same genotypes as in Experiment I. However, the genotypes were subjected to inoculation with race 63-39 of P. griseola. Similarly, the relative expression of the same 13 genes only in the CDRK cultivar were evaluated at 24, 72, 120, 168, and 216 hpi, in addition to the mock condition. Each experimental condition was replicated across three separate biological replicates (plants). Within each biological replicate, the assessment was further reinforced by performing three technical replicates of quantitative polymerase chain reaction (qPCR) reactions for each experiment.Top of Form
The experiments were conducted at the Núcleo de Pesquisa Aplicada à Agricultura (Nupagri) at the Universidade Estadual de Maringá (UEM) in Maringá, Paraná, Brazil (latitude 23° 26’8” S, longitude 51° 53’42”). Briefly, seeds were planted in plastic trays filled with a commercial substrate, MecPlant (MEC PREC—Ind. Com Ltda, Telemaco Borba, Brazil), that had been previously sterilized and fertilized. The seedlings were grown in greenhouses under natural light at a temperature of 25 °C until the first trifoliate leaf growth stage [35].
4.2. Pathogenesis assays
Monosporic cultures of C. lindemuthianum and P. griseola were prepared according to the methodologies described by Mathur et al. [36] and Sanglard et al. [37], respectively. Inocula of the ANT pathogen races were produced by incubating them on a young green common bean pod medium [38] at 22°C for 14 days. The inoculum for the ALS pathogens was multiplied in Petri dishes with tomato medium [37, 39] containing 1.61% agar (m/v), 0.25% calcium carbonate (m/v), 61.94% distilled water (v/v), and 36.2% V8® vegetable juice [Campbell’s company soup (v/v)] and maintained in a bio-oxygen demand incubator at 24°C for 18 days. The concentration of fungal spores for the ANT pathogen was adjusted to 1.2 × 106 [38]. For the ALS pathogen, it was adjusted to 1.2 × 104 conidia ml−1 [40] using a hemacytometer (1/400 mm2, Hausser Scientific). Fourteen-day-old seedlings were inoculated with each race of the pathogen on the underside of their leaves. The inoculation process was carried out by using a manual pressurized pump sprayer for spraying. For the mock treatment, which served as the negative control, the seedlings were sprayed with only distilled water and Tween 20® (0.01%). After inoculation, the plants were maintained in a mist chamber at >95% relative humidity, at a temperature of 20 ± 2°C, with 12 hours of daily light (680 lux) for 72. Following the inoculation process, the seedlings were moved to benches and placed under the same conditions as before, except for a high-humidity environment. This environment was maintained until the end of the experiment when all the samples were collected. To evaluate the severity of ANT and ALS symptoms, we used disease severity scales ranging from 1 to 9, as proposed by Inglis et al. [41] and Pastor-Corrales et al. [35]. Based on these scales, plants with disease reaction scores between 1 and 3 were classified as resistant, while those with scores ranging from 4 to 9 were classified as susceptible.
4.3. Total RNA extraction and cDNA synthesis
Leaf samples were collected from CDRK plants before inoculation (mock) and during incompatible reactions with race 73 of C. lindemuthianum, as well as with race 63-39 of P. griseola. Sampling was performed at specific time points critical for pathogens development: 24, 48, 72, 96, and 120 hpi for C. lindemuthianum, and 24, 72, 120, 168, and 216 hpi for P. griseola. Samples were obtained from three biological replicates for each pathogen. To ensure RNA integrity, the leaf samples were promptly frozen in liquid nitrogen for subsequent extraction [42].
Total RNA was extracted from 100 mg frozen and purified using GeneJet Plant RNA Purification Kit (Thermo Fisher Scientific, Waltham, EUA) following the manufacturer’s instructions. The integrity of the total RNA was assessed by electrophoresis on a 1% m/v agarose gel, run for 80 minutes at 80 volts, at 5°C, and in the absence of light. For the assessment of both the quality and quantity of total RNA, a spectrophotometer (FEMTO 700STM) was employed to measure absorbance at specific wavelengths: 230 nm, 240 nm, 260 nm, and 280 nm. The criteria for RNA purity were determined based on the following absorbance ratios: A260/A230 ranging from 1.9 to 2.4, A260/A240 of at least 1.4, and A260/A280 between 1.8 and 2.2. To compute the concentration of total RNA, the formula [RNA] (ng µL-1) = A260nm × 40 × 100, as outlined by Farrell [43], was utilized. Total RNA samples that met the purity and integrity criteria were treated with DNase ITM (Invitrogen™, Waltham, USA) to eliminate any possible genomic DNA contamination. The purification reaction involved 1 µg of total RNA, following the manufacturer's instructions.
To synthesize cDNA, the 'Superscript® IV First-Strand Synthesis System' kit (Invitrogen™, Waltham, USA) was used according to the manufacturer's instructions. The cDNA synthesis reaction was prepared with a total volume of 20 µL, containing the following constituents: 1 µg of total RNA, primer-oligo d(T) at a concentration of 2.5 µM, dNTP mix with each nucleotide at 0.5 mM, First-Strand Buffer at 1X concentration, DL-dithiothreitol at 5 mM, ribonuclease inhibitor with a concentration of 2 U µL-1, MMLV-RT at 10 U µL-1, and RNase-free water. The procedure began by combining total RNA, primer-oligo d(T), dNTP mix, and RNase-free water to reach a cumulative volume of 13 µL. The samples were incubated in a thermocycler (Applied Biosystems® Veriti® 96-Well Fast Thermal Cycler) at 65 °C for 5 minutes, followed by 4 °C for 1 minute. Then, the First-Strand Buffer, DL-dithiothreitol, ribonuclease inhibitor, and 1 MMLV-RT were added to the reaction. The samples were incubated at 55 °C for 10 minutes for cDNA synthesis activation, followed by 80 °C for 10 minutes to inactivate the reaction. To remove residual RNA after cDNA synthesis, 1 µL of Escherichia coli RNase H was added, and the samples were incubated at 37 °C for 20 minutes. The cDNA synthesis product (20 µL) was diluted 1:100 for qPCR analysis. To assess the cDNA synthesis efficiency, positive control was included, in which HeLa-S3 RNA (10 ng) was used instead of total RNA.
To verify the quality of the cDNA synthesis, a PCR reaction was performed on the positive control and a negative control (containing water instead of cDNA) using the following reaction mix: 5 µL PCR buffer (10 X), 2 µL MgCl (50 mM), 1 µL dNTP Mix (10 mM), 1 µL sense primer (10 µM), 1 µL antisense primer (10 µM), 2 µL of cDNA for the positive control, and 2 µL of ultrapure H2O for the negative control, 0.2 µL of Taq PlatinumTM DNA polymerase (Invitrogen™, Waltham, USA), and 37.8 µL of ultrapure H2O. The PCR reaction was performed for 35 cycles, with an initial denaturation at 94 °C for 2 minutes, denaturation at 94 °C for 15 seconds, annealing at 55 °C for 30 seconds, and synthesis at 68 °C for 1 minute. The PCR products were analyzed by electrophoresis on a 1.5% m/v agarose gel. The positive control showed a single band of approximately 353 bp, while no product was observed in the negative control, confirming the efficiency of the cDNA synthesis.
4.4. Target genes and primer design
Ten candidate genes were selected for expression analysis in CDRK plants inoculated with race 73 of C. lindemuthianum and race 63-39 of P. griseola. The genes evaluated in the CoPv01CDRK/PhgPv01CDRK resistance loci were Phvul.001G246000, Phvul.001G246100, Phvul.001G246200, and Phvul.001G246300. Additionally, the gene Phvul.001G245300 located near this locus was also assessed [15]. The genes proposed for the Co-AC locus, namely Phvul.001G244300, Phvul.001G244400, and Phvul.001G244500 [14], were also tested. The gene Phvul.001G243800 was evaluated as it was induced in the near isogenic line T9576R, which carries the Co-12 resistance allele, when inoculated with race 73 of C. lindemuthianum [22]. The KTR2/3 gene, a candidate gene for Co-x in Jalo EEP558, which was induced in response to isolate 100 of C. lindemuthianum, was also included [23]. Finally, three known plant defense genes, namely Phvul.003G109100 (PR1a), Phvul.006G196900 (PR1b), and Phvul.009G256400 (PR2) were evaluated [16, 22, 42]. To normalize gene expression levels, the reference genes Phvul.008G011000 (actin - ACT) and Phvul.001G133200 (insulin-degrading enzyme - IDE) were used as reference genes [44].
To design primers for qPCR, the coding sequences (CDS) and DNA sequences of the target genes were downloaded from the common bean (P. vulgaris L.) genome available at Phytozome 12 [45]. The 'Primer-BLAST web tool' [46] was used to design primers that met the following specifications for efficient qPCR: primer size 18-24 bp, melting temperature between 59-61°C, amplicon size between 80-160 bp, and whenever possible, at least one intron on the corresponding genomic DNA sequence was included between the primer pair. The primers for the KTR2/3 gene were obtained from Richard et al. [23].
To ensure the specificity and efficiency of the primers, dimers and secondary structures were checked using Gene Runner software (version 6.5.52), the 'Multiple Prime Analyzer' web tool (Thermo Fisher Scientific: https://bit.ly/34kZpnP), and 'The Sequence Manipulation Suite' web tool [47]. The amplicon secondary structure was also verified using the "The Mfold Web Server" platform [48] with coding sequences downloaded from Phytozome 12. All procedures used for primer design and in silico validation followed literature recommendations [49, 50]. The primer sequences for each candidate gene evaluated are listed in Table 3.
4.5. Quantitative PCR (qPCR) and data analysis
The determination of PCR efficiency for each primer involved establishing a standard curve through a fivefold serial dilution, utilizing the cDNA pool as the template. This process incorporated three replicates at every dilution point, following the methodologies outlined by Svec et al. [51] and Rasmussen [52]. The amplification efficiency was computed employing the equation E = [10(-1/slope)] - 1 [52], utilizing the slope values derived from linear regression analysis. This analysis encompassed the log10-transformed cDNA concentrations on the x-axis and corresponding Cq values on the y-axis. The calculated amplification efficiency for each primer pair ranged from 0.92 to 1.09, while maintaining a coefficient of determination (R2) for the linear regression of at least 0.97 (Table 3).
The cDNA quantification reactions were conducted in the StepOnePlus™ real-time PCR system (Applied Biosystems™; StepOnePlus™ Real-Time PCR Systems) using 96-well microplates [MicroAmp™ Fast 96 -well Reaction Plate (0.1 mL)] sealed with MicroAmp™ Optical Adhesive Film. The total reaction volume was 10 µL, consisting of 3.4 µL of cDNA, 1.6 µL of forward and reverse primer mix (800 nM), and 5 µL of PowerUp™ SYBR™ Green Master Mix (Applied Biosystems™). The thermocycling conditions included 50°C for 2 minutes, 95°C for 2 minutes, 40 cycles of 15 seconds at 95°C, and 30 seconds at 60°C.
After completion of the cDNA quantitation reaction, a dissociation curve was performed to verify target specificity using the manufacturer's standard continuous melt curve setup, and only samples exhibiting specificity based on the dissociation curve were used. Cq values were obtained using StepOnePlus™ Software v2.3 (Applied Biosystems™), with the baseline determined automatically and the threshold was determined manually in the exponential phase of amplification (0.7707 for all cDNA quantification reactions).
The genes Phvul.001G133200 (ACT) and Phvul.008G011000 (IDE) were used as reference genes [44], with the arithmetic mean of Cq values (quantification cycle) [53] calculated in each experimental condition evaluated. Relative expression was determined based on Cq values normalized with the reference genes, using the 2-ΔΔCT method [54, 55]. Mean Cq values were calculated for each gene at each experimental condition based on three biological repetitions and three replicates (n=3x3).
The relative expression of candidate genes to the CoPv01CDRK/PhgPv01CDRK loci and known disease resistance genes were investigated in response to race 73 of C. lindemuthianum at 24, 48, 72, 96, and 120 hpi, as well as in response to race 63-39 of P. griseola at 24, 72, 120, 168, and 216 hpi in CDRK cultivar. The calibrator condition for each gene was the relative expression in the mock (control, without pathogen). To analyze the data and show results, logarithmic base 2 transformation was performed before statistical analysis. The expression levels among experimental conditions were compared using the Alexander-Govern test with a significance level of 5%. Pairwise comparisons of relative expression mean among time points for each gene were assessed, and the significance level was adjusted using Bonferroni correction (p≤0.05). These statistical analyses were performed using the 'oneway-test' [56] and 'companion' R packages. All data wrangling and statistical analysis were performed using R software (version 4.0.3) (R Core Team), with plots generated using the package ggplot2 [57] and R base. Error bars represent the standard deviation of the means from three biological and three technical replicates (3x3). Heatmaps were generated with mean Cq values using the ‘heatmaply’ R package, and the dendrogram was based on the Euclidean distance measure and the average linkage function [58] among the relative expression of the genes.
5. Conclusions
In conclusion, our study provides valuable insights into the genomic organization of ANT and ALS resistance genes on Pv01, specifically in the context of the CDRK cultivar. We highlight the importance of closely situated candidate genes within resistance gene clusters, as they play a crucial role in conferring effective resistance against ANT and ALS pathogens. Furthermore, the CoPv01CDRK and PhgPv01CDRK resistance loci are concomitantly inherited and can be efficiently tracked using molecular markers. Our findings shed light on the fact that the Phvul.001G246300 gene develops a pleiotropic mechanism, demonstrating the highest responsiveness to both pathogens: C. lindemuthianum race 73 and P. griseola race 63-39. These discoveries carry significant practical implications for breeding endeavors aimed at developing bean cultivars resistant to ANT and ALS, facilitated by the implementation of marker-assisted selection.
Author Contributions
Conceptualization, M.L. and M.C.G-V.; methodology, M.L., M.C.G-V., A.C.C. and J.M.; data curation and formal analysis, M.L., A.C.C., J.M.; funding acquisition and project administration, P.S.V-F. and M.C.G-V.; supervision, M.C.G-V., P.S.V-F. and M.V-B.; writing—original draft preparation, M.L.; writing—review and editing, M.L., M.C.G-V., M.V-B., M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received financial support from the National Council for Scientific and Technological Development (CNPq). M. Lovatto and A.C. Calvi were supported by scholarships from CNPq and M. Vaz Bisneta by a scholarship from Capes. M.C. Gonçalves-Vidigal and P.S. Vidigal Filho are grateful for grants provided by CNPq and Capes, respectively.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Dr. Paul Gepts of the University of California Davis, USA, for generously providing the seeds of the CDRK cultivar.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Padder, B.A.; Sharma, P.N.; Awale, H.E.; Kelly, J.D. Colletotrichum lindemuthianum, the causal agent of bean anthracnose. J. Plant Pathol. 2017, 99, 317–330. [Google Scholar] [CrossRef]
- Crous, P.W.; Liebenberg, M.M.; Braun, U.; Johannes, Z.; Groenewald, J.Z. Re-evaluating the taxonomic status of Phaeoisariopsis griseola, the causal agent of angular leaf spot of bean. Stud. Mycol. 2006, 55, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Miklas, P.N.; Kelly, J.D.; Beebe, S.E.; Blair, M.W. Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Euphytica 2006, 147, 105–131. [Google Scholar] [CrossRef]
- Singh, S.P.; Schwartz, H.F. Breeding common bean for resistance to diseases: A review. Crop Sci. 2010, 50, 2199–2223. [Google Scholar] [CrossRef]
- Guzman, P. Characterization of variability in the fungus Phaeoisariopsis griseola suggests coevolution with the common bean (Phaseolus vulgaris). Phytopathology. 1995, 85, 600. [Google Scholar] [CrossRef]
- Pastor-Corrales, M.A. Jara, C.E. La evolución de Phaeoisariopsis griseola con el frijol común en América Latina. Fitop Colomb. 1995, 19, 15–24. [Google Scholar]
- Pastor-Corrales, M.A.; Jara, C.; Singh, S.P. Pathogenic variation in, sources of, and breeding for resistance to Phaeoisariopsis griseola causing angular leaf spot in common bean. Euphytica 1998, 103, 161–171. [Google Scholar] [CrossRef]
- Stenglein, S.A.; Balatti, P.A. Genetic diversity of Phaeoisariopsis griseola in Argentina as revealed by pathogenic and molecular markers. Physiol. Mol. Plant Pathol. 2006, 68, 158–167. [Google Scholar] [CrossRef]
- Gonçalves-Vidigal, M.C.; Cruz, A.S.; Garcia, A.; Kami, J.; Vidigal Filho, P.S.; Sousa, L.L.; McClean, P.; Gepts, P.; Pastor-Corrales, M.A. Linkage mapping of the Phg-1 and Co-14 genes for resistance to angular leaf spot and anthracnose in the common bean cultivar AND 277. Theor. Appl. Genet. 2011, 122, 893–903. [Google Scholar] [CrossRef]
- Zuiderveen, G.H.; Padder, B.A.; Kamfwa, K.; Song, Q.; Kelly, J.D. Genome-wide association study of anthracnose resistance in Andean beans (Phaseolus vulgaris). PLoS One 2016, 11, e0156391. [Google Scholar] [CrossRef]
- Chen, M.; Wu, J.; Wang, L.; Mantri, N.; Zhang, X.; Zhu, Z.; Wang, S. Mapping and genetic structure analysis of the anthracnose resistance locus Co-1HY in the common bean (Phaseolus vulgaris L.). PLoS One 2017, 12, e0169954. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.R.L.; Gonçalves-Vidigal, M.C.; Vaz Bisneta, M.; Valentini, G.; Vidigal Filho, P.S.; Martins, V.S.R., Souza, T.L.P.O. Genetic fine-mapping of anthracnose disease-resistance allele Co-14 present in the Andean common bean cultivar AND 277. Crop Sci. 2023, 1–14. [CrossRef]
- Richard, M.M.S.; Pflieger, S.; Sevignac, M.; Thareau, V.; Blanchet, S.; Li, Y.; Jackson, S.A.; Jackson, S.A.; Geffroy, V. Fine mapping of Co-x, an anthracnose resistance gene to a highly virulent strain of Colletotrichum lindemuthianum in common bean. Theor. Appl. Genet. 2014, 127, 1653–1666. [Google Scholar] [CrossRef] [PubMed]
- Gilio, T.A.S.; Hurtado-Gonzales, O.P.; Gonçalves-Vidigal, M.C.; Valentini, G.; Elias, J.C.F.; Song, Q.; Pastor-Corrales, M.A. Fine mapping of an anthracnose-resistance locus in Andean common bean cultivar Amendoim Cavalo. PLoS One 2020, 15, e0239763. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves-Vidigal, M.C.; Gilio, T.A.S.; Valentini, G.; Vaz Bisneta, M.; Vidigal Filho, P.S.; Song, Q.; Oblessuc, P.R.; Melotto, M. (2020). New Andean source of resistance to anthracnose and angular leaf spot: Fine-mapping of disease-resistance genes in California Dark Red Kidney common bean cultivar. PLoS One. 15, e0235215. [CrossRef]
- Oblessuc, P.R.; Baroni, R.M.; Garcia, A.A.F.; Chioratto, A.F.; Carbonell, S.A.M.; Camargo, L.E.A.; Benchimol, L.L. Mapping of angular leaf spot resistance QTL in common bean (Phaseolus vulgaris L.) under different environments. BMC Genet. 2012, 13, 50. [Google Scholar] [CrossRef]
- Keller, B.; Manzanares, C.; Jara, C.; Lobaton, J. D.; Studer, B.; Raatz, B. Fine-mapping of a major QTL controlling angular leaf spot resistance in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2015, 128, 813–826. [CrossRef]
- Sartorato, A.; Nietsche, S.; Barros, E.G.; Moreira, M.A. Inheritance of angular leaf spot resistance and RAPD markers linked to disease resistance gene in common beans. Ann. Rep. Bean Improv. Coop. 1999, 42, 21–22. [Google Scholar]
- Gonçalves-Vidigal, M.C.; Cruz, A.S.; Lacanallo, G.F.; Vidigal Filho, P.S.; Sousa, L. L.; Pacheco, C.M.N.A.; McClean, P.; Gepts, P.; Pastor-Corrales, M.A. Co-segregation analysis and mapping of the anthracnose Co-10 and angular leaf spot Phg-ON disease-resistance genes in the common bean cultivar Ouro Negro. Theor. Appl. Genet. 2013, 126, 2245–2255. [Google Scholar] [CrossRef]
- Oblessuc, P.R.; Perseguini, J.M.K.C.; Baroni, R.M.; Chiorato, A.F.; Carbonell, S.A. M.; Mondego, J.M.C.; Vidal, R.O.; Camargo, L.E.A.; Benchimol-Reis, L.L. Increasing the density of markers around a major QTL controlling resistance to angular leaf spot in common bean. Theor. Appl. Genet. 2013, 126, 2451–2465. [Google Scholar] [CrossRef]
- Nay, M. M.; Souza, T.L.P.O.; Raatz, B.; Mukankusi, C.M.; Gonçalves-Vidigal, M.C.; Abreu, A.F.B.; Melo, L.C.; Pastor-Corrales, M.A. A review of angular leaf spot resistance in common bean. Crop Sci. 2019, 59, 1376. [Google Scholar] [CrossRef] [PubMed]
- Mahiya-Farooq; Padder, B.A.; Bhat, N.N.; Shah, M.D.; Shikari, A.B.; Awale, H.E.; Kelly, J.D. Temporal expression of candidate genes at the Co-1 locus and their interaction with other defense related genes in common bean. Physiol. Mol. Plant Pathol. 2019, 108, 101424. [CrossRef]
- Richard, M.M.S.; Gratias, A.; Diaz, J.C.A.; Thareau, V.; Pflieger, S.; Meziadi, C.; Blanchet, S.; Marande, W.; Bitocchi, E.; Papa, R.; Miklas, P.N.; Geffroy, V. A common bean truncated CRINKLY4 kinase controls gene-for-gene resistance to the fungus Colletotrichum lindemuthianum. J. Exp. Bot. 2021, 72, 3569–3581. [Google Scholar] [CrossRef] [PubMed]
- McClean, P.E.; Myers, J.R.; Hammond, J.J. Coefficient of parentage and cluster analysis of North American dry bean cultivars. Crop Sci. 1993, 33, 190–197. [Google Scholar] [CrossRef]
- Boller, T.; Felix, G. A renaissance of elicitors, perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Vaz Bisneta, M.; Gonçalves-Vidigal, M.C. Integration of anthracnose resistance loci and RLK and NBS-LRR-encoding genes in the Phaseolus vulgaris L. genome. Crop Sci. 2020, 60, 2901–2918. [Google Scholar] [CrossRef]
- Aleman, F.; Yazaki. J.; Lee, M.; Takahashi, Y.; Kim, A.Y.; Li. Z.; Kinoshita, T.; Ecker, J.R.; Schroeder, J.I. ABA-increased interaction of the PYL6 ABA receptor with MYC2 transcription factor: a putative link of ABA and JA signaling. Sci. Rep. 2016, 6, 28941. [Google Scholar] [CrossRef]
- García-Andrade, J.; González, B.; Gonzalez-Guzman, M.; Rodriguez, P. L.; Vera, P. The role of ABA in plant immunity is mediated through the PYR1 receptor. Int. J. Mol. Sci. 2020, 21, 5852. [Google Scholar] [CrossRef]
- Ng, A.; Xavier, R.J. Leucine-rich repeat (LRR) proteins: integrators of pattern recognition and signaling in immunity. Autophagy. 2011, 7, 1082–1084. [Google Scholar] [CrossRef]
- Li, X.; Kapos, P.; Zhang, Y. NLRs in plants. Curr. Opin. in Immunol. 2015, 32, 114–121. [Google Scholar] [CrossRef]
- Yuan, Y.; Teng, Q.; Zhong, R.; Haghighat, M.; Richardson, E.A.; Ye, Z-H. Mutations of Arabidopsis TBL32 and TBL33 affect Xylan acetylation and secondary wall deposition. PLoS ONE. 2016, 11, e0146460. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Teng, Q.; Zhong, R.; Ye, Z-H. The Arabidopsis DUF231 Domain-Containing Protein ESK1 Mediates 2-O- and 3-O-Acetylation of Xylosyl Residues in Xylan. Plant Cell Physiol. 2013, 54, 1186–1199. [Google Scholar] [CrossRef] [PubMed]
- Padder, B.A.; Kamfwa, K.; Awale, H.E.; Kelly, J. D. Transcriptome profiling of the Phaseolus vulgaris - Colletotrichum lindemuthianum pathosystem. PLoS One. 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Shams, E.; Javan-Nikkhah, M.; Mirzadi Gohari, A. Dissecting molecular events and gene expression signatures involved in Colletotrichum lindemuthianum-Phaseolus vulgaris pathosystem in compatible and incompatible interactions. Eur. J. Plant Pathol. 2020, 156, 925–937. [Google Scholar] [CrossRef]
- Pastor-Corrales, M. A.; Otoya, M. M.; Molina, A.; Singh, S.P. Resistance to Colletotrichum lindemuthianum isolates from middle America and Andean South America in different common bean races. Plant Dis. 1995, 79, 63–67. [Google Scholar] [CrossRef]
- Mathur, R.S.; Barnett, H.l.; Lilly, V.G. Sporulation of Colletotrichum lindemuthianum in culture. Phytopathol. 1950, 40, 104–114. [Google Scholar]
- Sanglard, D.A.; Balbi, B.P.; Barros, E.G.; Moreira, M.A. An efficient protocol for isolation, sporulation and maintenance of Pseudocercospora griseola. Ann. Rep. Bean Improv. Coop. 2009, 52, 62–63. [Google Scholar]
- Cárdenas, F.; Adams, M.W.; Andersen, A. The genetic system for reaction of field beans (Phaseolus vulgaris L. ) to infection by three physiologic races of Colletotrichum lindemuthianum. Euphytica, 1964, 13, 178–186. [Google Scholar] [CrossRef]
- de Almeida, C. P.; Arruda, N.; de Carvalho Paulino, J. F.; de Freitas, G.M.; Bonfante, G.F.J.; Bajay, M.M. , Deus, B.C.; Patrício, F.R.A.; Carbonell, S.A.M.; Chiorato, A.F.; Benchimol-Reis, L.L. Genetic diversity of Pseudocercospora griseola resistance loci in common beans. Trop. Plant Pathol. 2021, 46, 129–138. [Google Scholar] [CrossRef]
- Castellanos, G.; Jara, C.; Mosquera, G. (2015). Bean pathogens: Practical guide for lab and greenhouse work. Cali, Colombia: Centro Internacional de Agricultura Tropical (CIAT).
- Inglis, D.A.; Hagedorn, J.; Rand, R.E. Use of dry inoculum to evaluate beans for resistance to anthracnose and angular leaf spot. Plant Dis. 1988, 72, 771–774. [Google Scholar] [CrossRef]
- Borges, A.; Melotto, M.; Tsai, S.M.; Caldas, D.G.G. Changes in spatial and temporal gene expression during incompatible interaction between common bean and anthracnose pathogen. J. Plant Physiol. 2012, 169, 1216–1220. [Google Scholar] [CrossRef] [PubMed]
- Farrell, R.E. RNA Methodologies: Laboratory guide for isolation and characterization, 5th ed.; Elsevier: Amsterdam, Netherlands, 2017; pp. 1–855. [Google Scholar]
- Borges, A.; Tsai, S.M.D.G.G. Validation of reference genes for RT-qPCR normalization in common bean during biotic and abiotic stresses. Plant Cell Rep. 2012, 31, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M. , Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; Rokhsar, D.S. Phytozome: a comparative platform for green plant genomics. Nucleic. Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. BioTechniques. 2000, 28, 1102–1104. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M. The MIQE Guidelines: minimum information for publication of quantitative real-time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Bustin, S.; Huggett, J. qPCR primer design revisited. Biomol. Detect. Quantif. 2017, 14, 19–28. [Google Scholar] [CrossRef]
- Svec, D.; Tichopad, A. ; Novosadova, V.; Pfaffl, M.W.; Kubista, M. How good is a PCR efficiency estimate: Recommendations for precise and robust qPCR efficiency assessments. Biomol. Detect. Quantif. 2015, 3, 9–16. [Google Scholar] [CrossRef]
- Rasmussen, R. Quantification on the LightCycler. In Rapid Cycle Real-Time PCR, Meuer, S., Wittwer, C., Nakagawara, K-I., Eds.; Springer: Heidelberg, Berlin, 2001; pp. 21–34. [Google Scholar] [CrossRef]
- Riedel, G.; Rüdrich, U. ; Fekete-Drimusz, N.; Manns, M.P.; Vondran, F.W.R.; Bock, M. An extended ΔCT-method facilitating normalisation with multiple reference genes suited for quantitative RT-PCR analyses of human hepatocyte-like cells. PLoS One. 2014, 9, e93031. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Mendiburu, F.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research. 2020, Available: {https://myaseen208.github.io/agricolae/},{https://cran.r-project.org/package=agricolae}.
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistics Computing: Vienna, Austria. 2020. URL http://www.R-project.org/.
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. Heatmaply: an R package for creating interactive cluster heatmaps for online publishing. Wren, J., editor. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef]
Figure 1.
(A) Disease reaction of susceptible Yolano cultivar and no reaction in resistant California Dark Red Kidney cultivar at 120 h post inoculation with C. lindemuthianum race 73. (B) Disease reaction of susceptible Yolano cultivar and no reaction in resistant California Dark Red Kidney cultivar at 216 h post inoculation with P. griseola race 63-39.
Figure 1.
(A) Disease reaction of susceptible Yolano cultivar and no reaction in resistant California Dark Red Kidney cultivar at 120 h post inoculation with C. lindemuthianum race 73. (B) Disease reaction of susceptible Yolano cultivar and no reaction in resistant California Dark Red Kidney cultivar at 216 h post inoculation with P. griseola race 63-39.

Figure 2.
Relative expression of candidate genes: (A) Phvul.001G246300; (B) Phvul.001G245300; (C) Phvul.001G246200; (D) KTR2/3; (E) Phvul.001G243800; (F) Phvul.001G246000; (G) Phvul.001G246100; (H) Phvul.001G244500; (I) Phvul.001G244400; (J) Phvul.001G244300 in California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
Figure 2.
Relative expression of candidate genes: (A) Phvul.001G246300; (B) Phvul.001G245300; (C) Phvul.001G246200; (D) KTR2/3; (E) Phvul.001G243800; (F) Phvul.001G246000; (G) Phvul.001G246100; (H) Phvul.001G244500; (I) Phvul.001G244400; (J) Phvul.001G244300 in California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
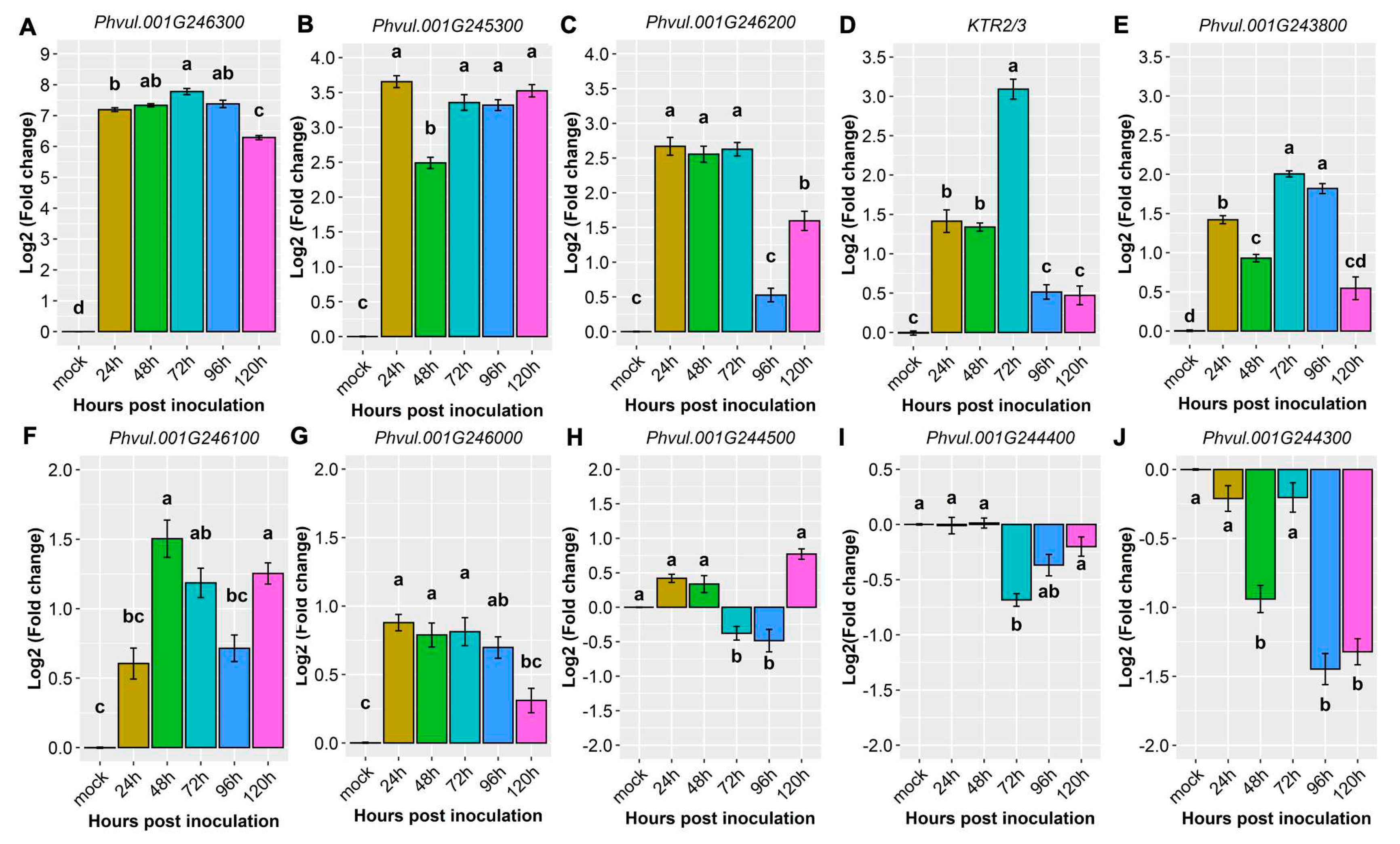
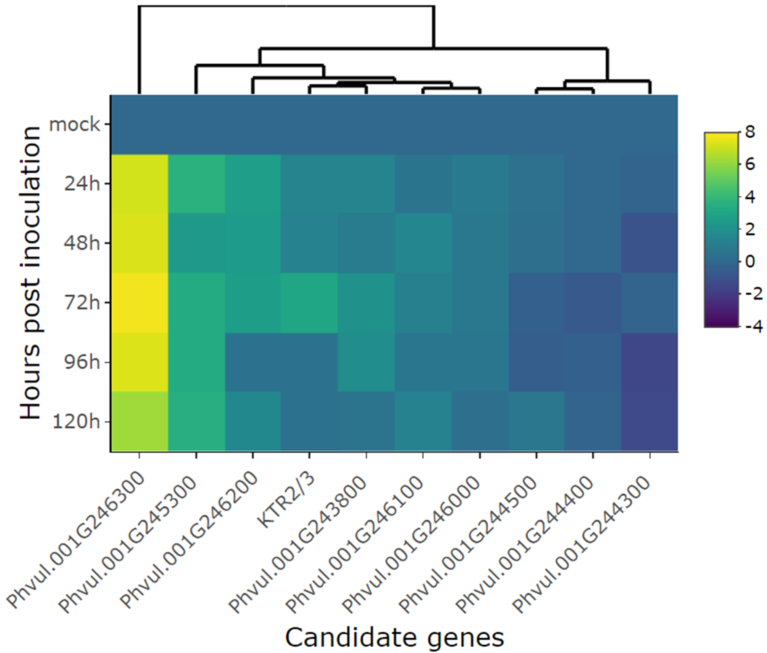
Figure 3.
Heatmap of relative expression of candidate genes for the CoPv01CDRK/PhgPv01CDRK and genes proximal to this loci in California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. The genes evaluated were Phvul.001G246300, Phvul.001G245300, Phvul.001G246200, KTR2/3, Phvul.001G243800, Phvul.001G246100, Phvul.001G246000, Phvul.001G244500, Phvul.001G244400, Phvul.001G244300. Yellow shading indicates higher expression and dark blue shading lower expression than that of reference genes.
Figure 3.
Heatmap of relative expression of candidate genes for the CoPv01CDRK/PhgPv01CDRK and genes proximal to this loci in California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. The genes evaluated were Phvul.001G246300, Phvul.001G245300, Phvul.001G246200, KTR2/3, Phvul.001G243800, Phvul.001G246100, Phvul.001G246000, Phvul.001G244500, Phvul.001G244400, Phvul.001G244300. Yellow shading indicates higher expression and dark blue shading lower expression than that of reference genes.
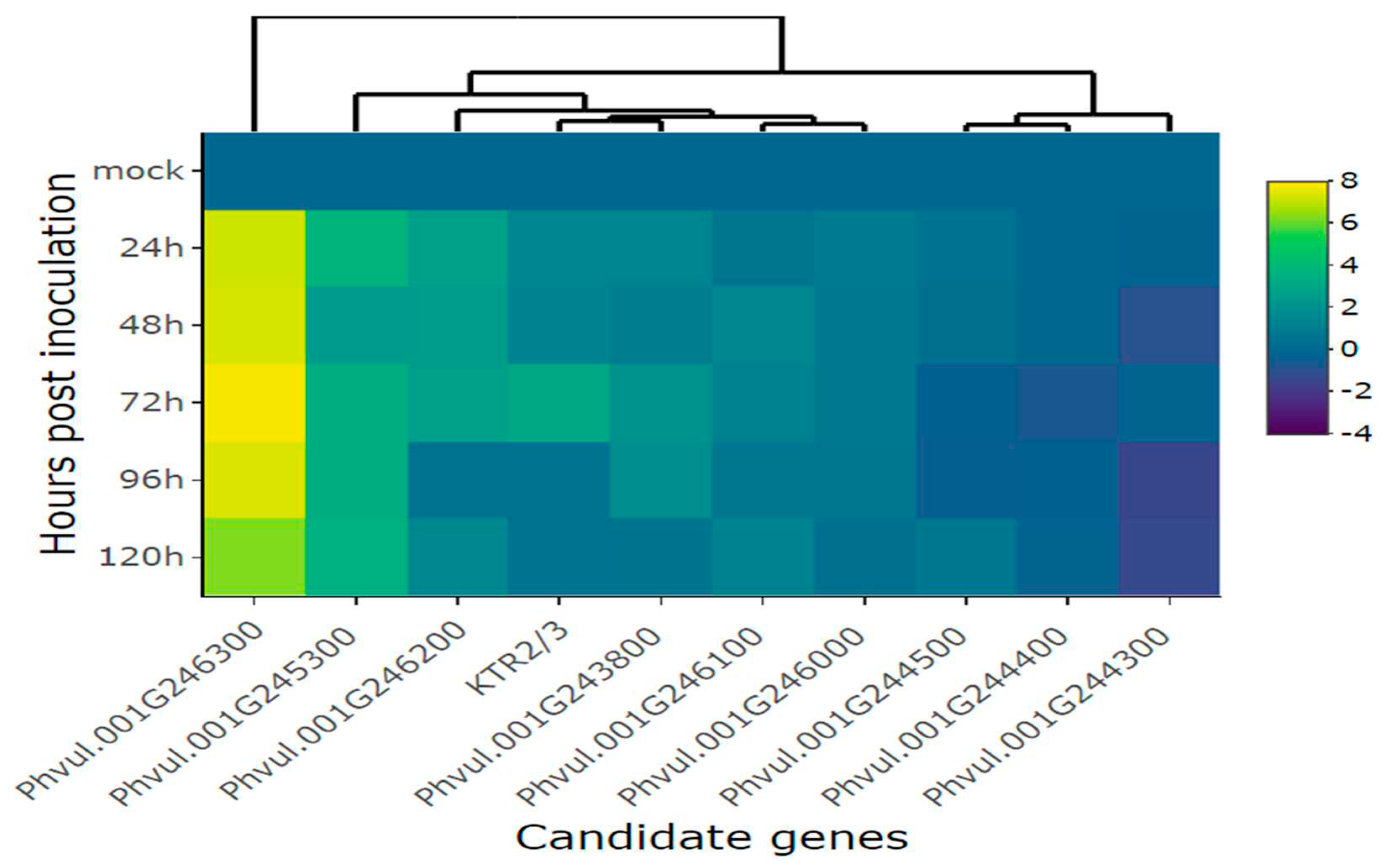
Figure 4.
Relative expression of plant defense genes: (A) Phvul.006G196900 (PR1b); (B) Phvul.009G256400 (PR2); (C) Phvul.003G109100 (PR1a) in the common bean cultivar California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
Figure 4.
Relative expression of plant defense genes: (A) Phvul.006G196900 (PR1b); (B) Phvul.009G256400 (PR2); (C) Phvul.003G109100 (PR1a) in the common bean cultivar California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
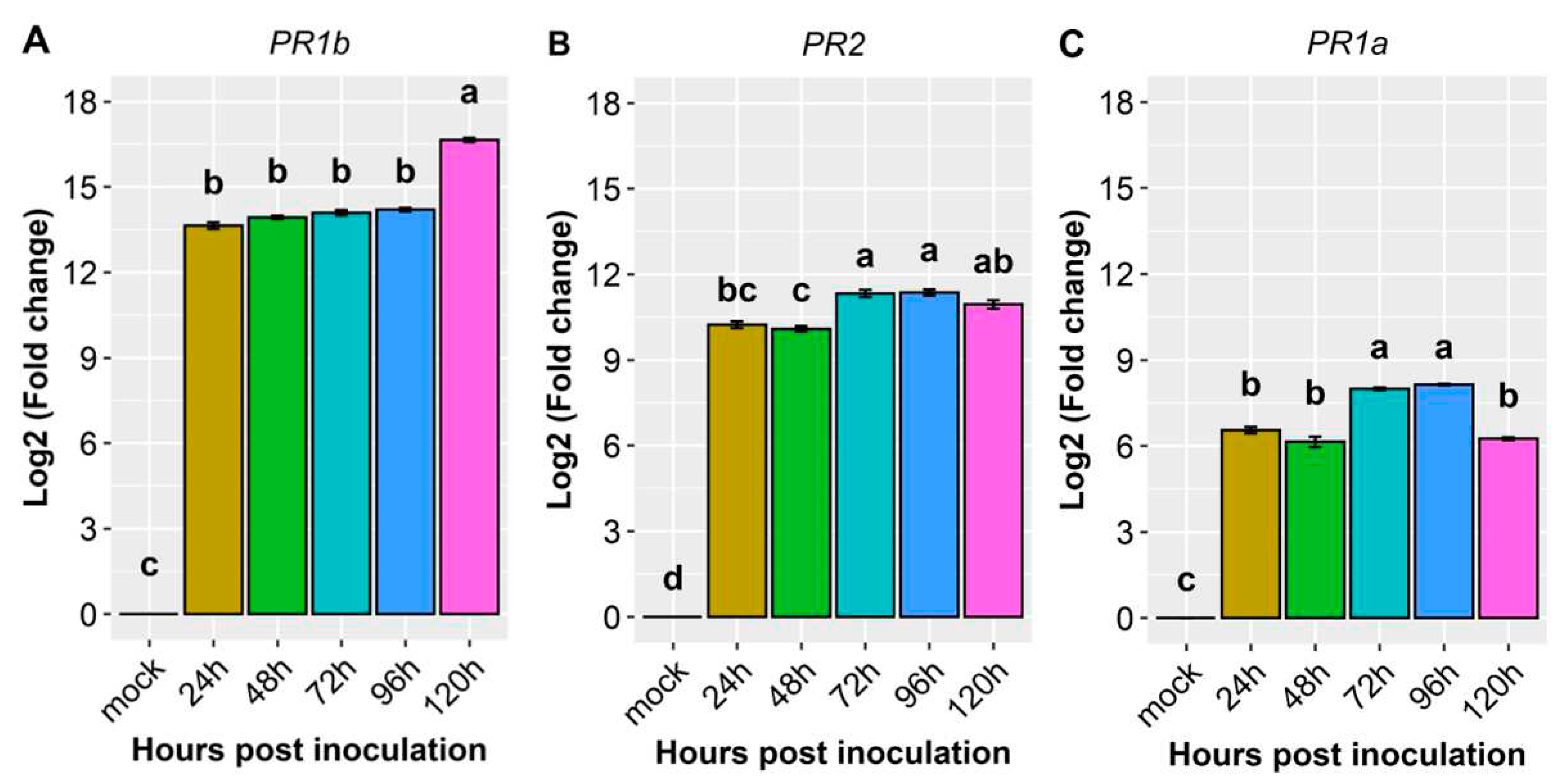
Figure 5.
Heatmap of relative expression of plant defense genes Phvul.006G196900 (PR1b), Phvul.009G256400 (PR2), and Phvul.003G109100 (PR1a) in the common bean cultivar California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. Yellow shading indicates higher expression and dark blue has lower expression than that of reference genes.
Figure 5.
Heatmap of relative expression of plant defense genes Phvul.006G196900 (PR1b), Phvul.009G256400 (PR2), and Phvul.003G109100 (PR1a) in the common bean cultivar California Dark Red Kidney at 24, 48, 72, 96, and 120 h post inoculation with race 73 of C. lindemuthianum and mock. Yellow shading indicates higher expression and dark blue has lower expression than that of reference genes.
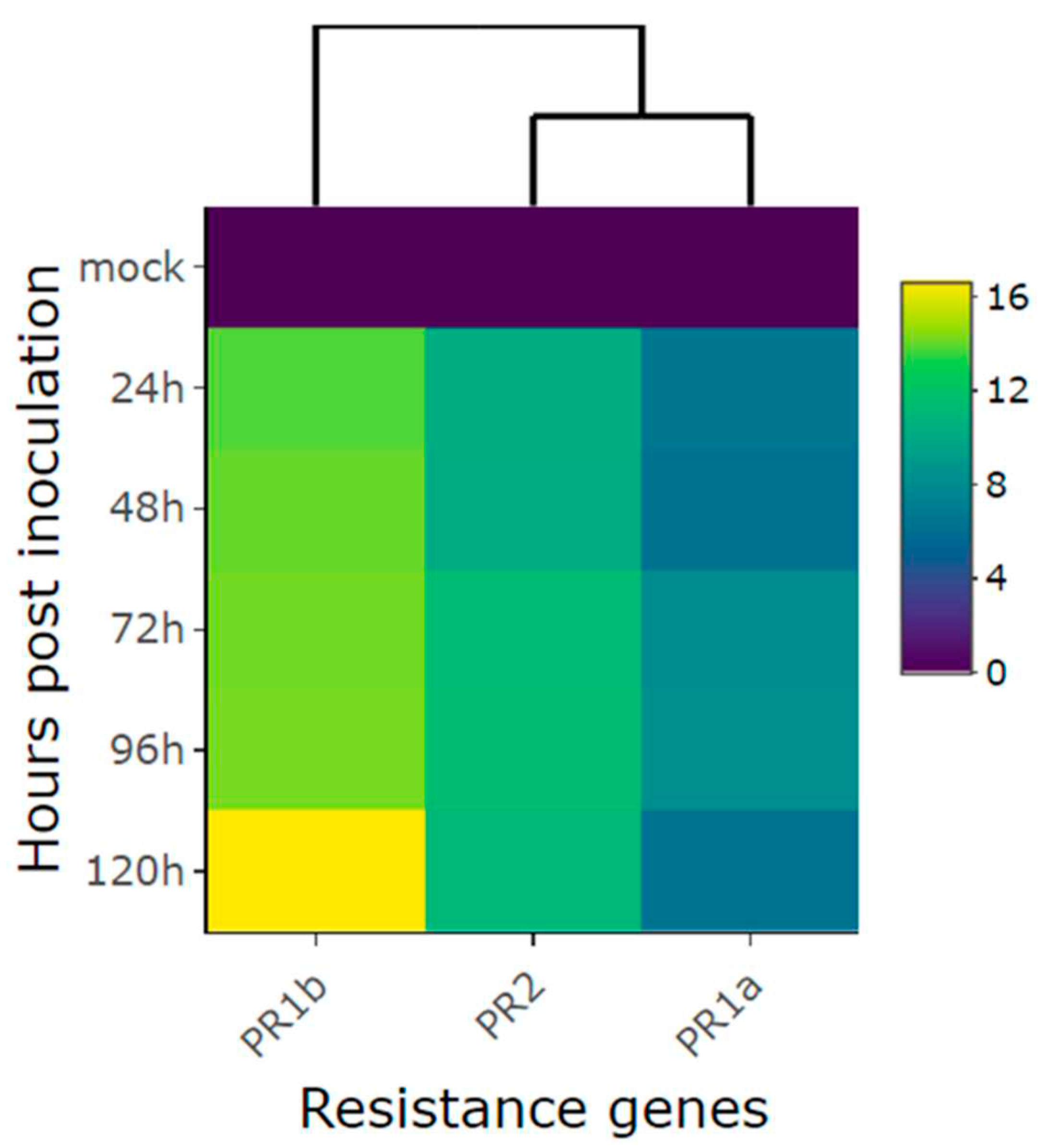
Figure 6.
Relative expression of candidate genes: (A) Phvul.001G246300; (B) Phvul.001G246200; (C) KTR2/3; (D) Phvul.001G246000; (E) Phvul.001G244300; (F) Phvul.001G246100; (G) Phvul.001G244400; (H) Phvul.001G243800; (I) Phvul.001G245300; (J) Phvul.001G244500 in California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
Figure 6.
Relative expression of candidate genes: (A) Phvul.001G246300; (B) Phvul.001G246200; (C) KTR2/3; (D) Phvul.001G246000; (E) Phvul.001G244300; (F) Phvul.001G246100; (G) Phvul.001G244400; (H) Phvul.001G243800; (I) Phvul.001G245300; (J) Phvul.001G244500 in California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
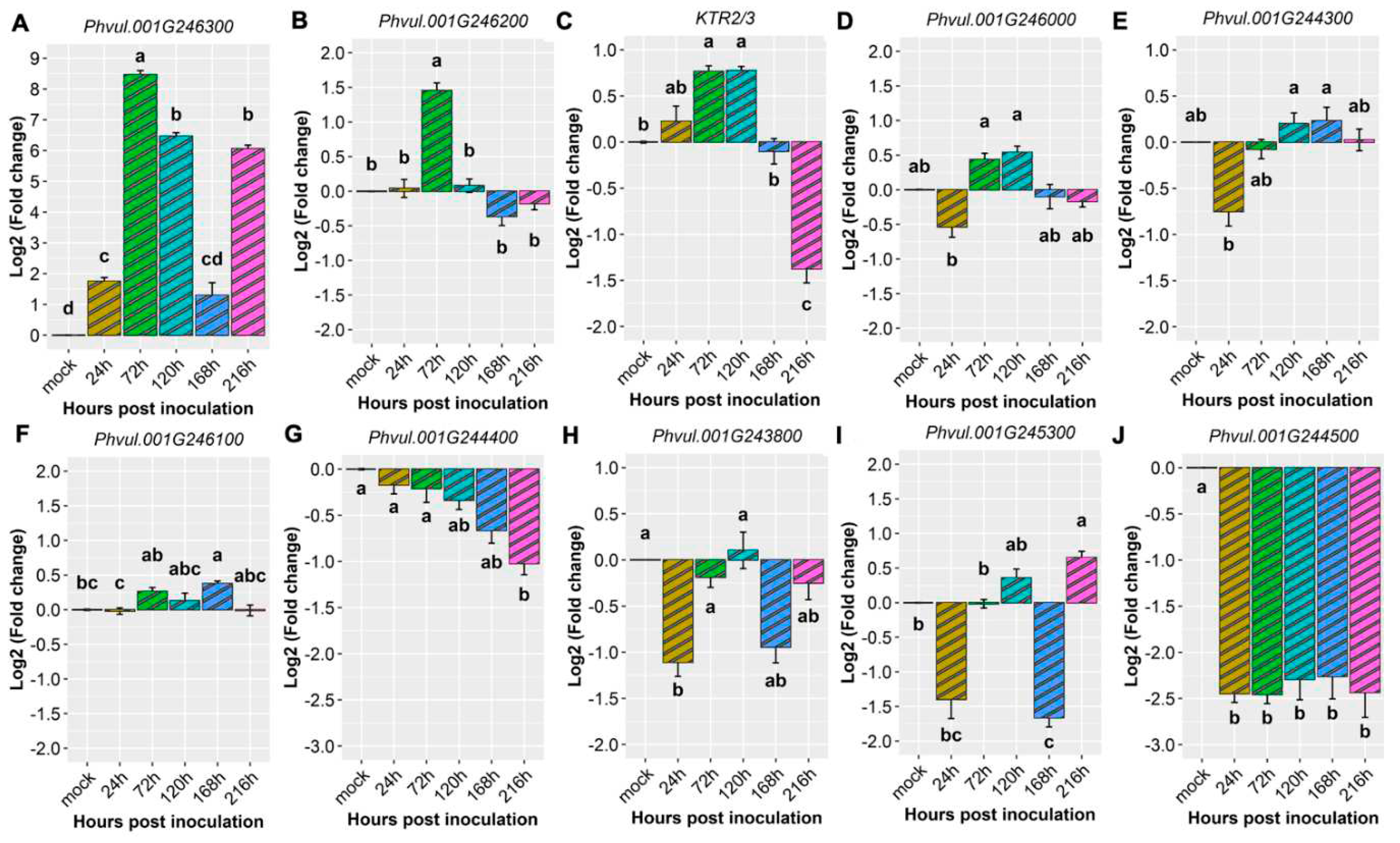
Figure 7.
Heatmap of relative expression of candidate genes for the CoPv01CDRK/PhgPv01CDRK and genes proximal to this loci in California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. The genes evaluated were Phvul.001G246300, Phvul.001G246200, KTR2/3, Phvul.001G246000, Phvul.001G244300, Phvul.001G246100, Phvul.001G244400, Phvul.001G243800, Phvul.001G245300 and Phvul.001G244500. Yellow shading indicates higher expression and dark blue shading lower expression than that of reference genes.
Figure 7.
Heatmap of relative expression of candidate genes for the CoPv01CDRK/PhgPv01CDRK and genes proximal to this loci in California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. The genes evaluated were Phvul.001G246300, Phvul.001G246200, KTR2/3, Phvul.001G246000, Phvul.001G244300, Phvul.001G246100, Phvul.001G244400, Phvul.001G243800, Phvul.001G245300 and Phvul.001G244500. Yellow shading indicates higher expression and dark blue shading lower expression than that of reference genes.
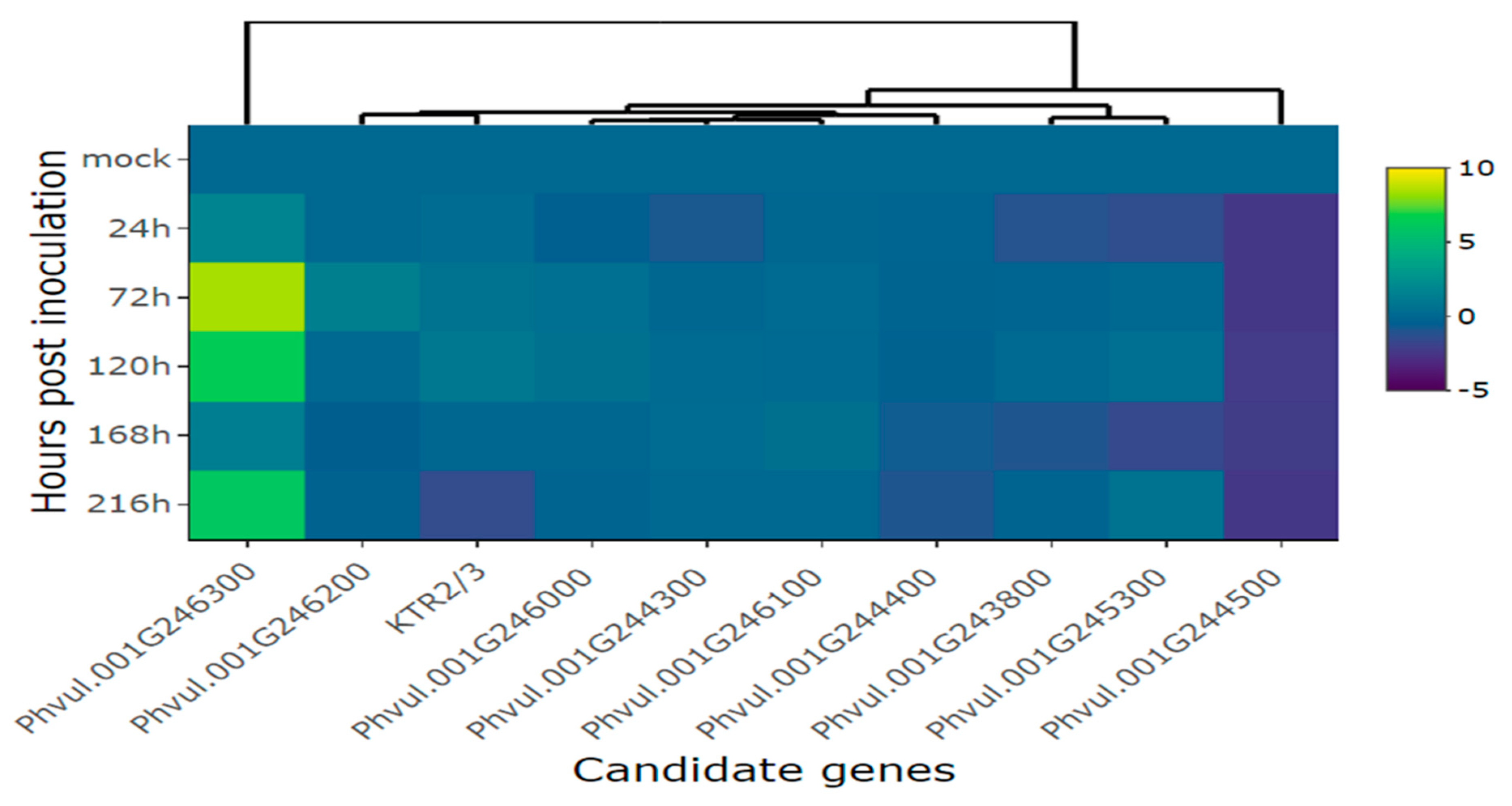
Figure 8.
Relative expression of plant defense genes: (A) Phvul.009G256400 (PR2); (B) Phvul.003G109100 (PR1a); (C) Phvul.006G196900 (PR1b) in the common bean cultivar California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
Figure 8.
Relative expression of plant defense genes: (A) Phvul.009G256400 (PR2); (B) Phvul.003G109100 (PR1a); (C) Phvul.006G196900 (PR1b) in the common bean cultivar California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. The results are presented as logarithmic base 2 of the fold change of gene expression. Error bars represent the standard deviation of the mean from three biological replicates and three technical replicates.
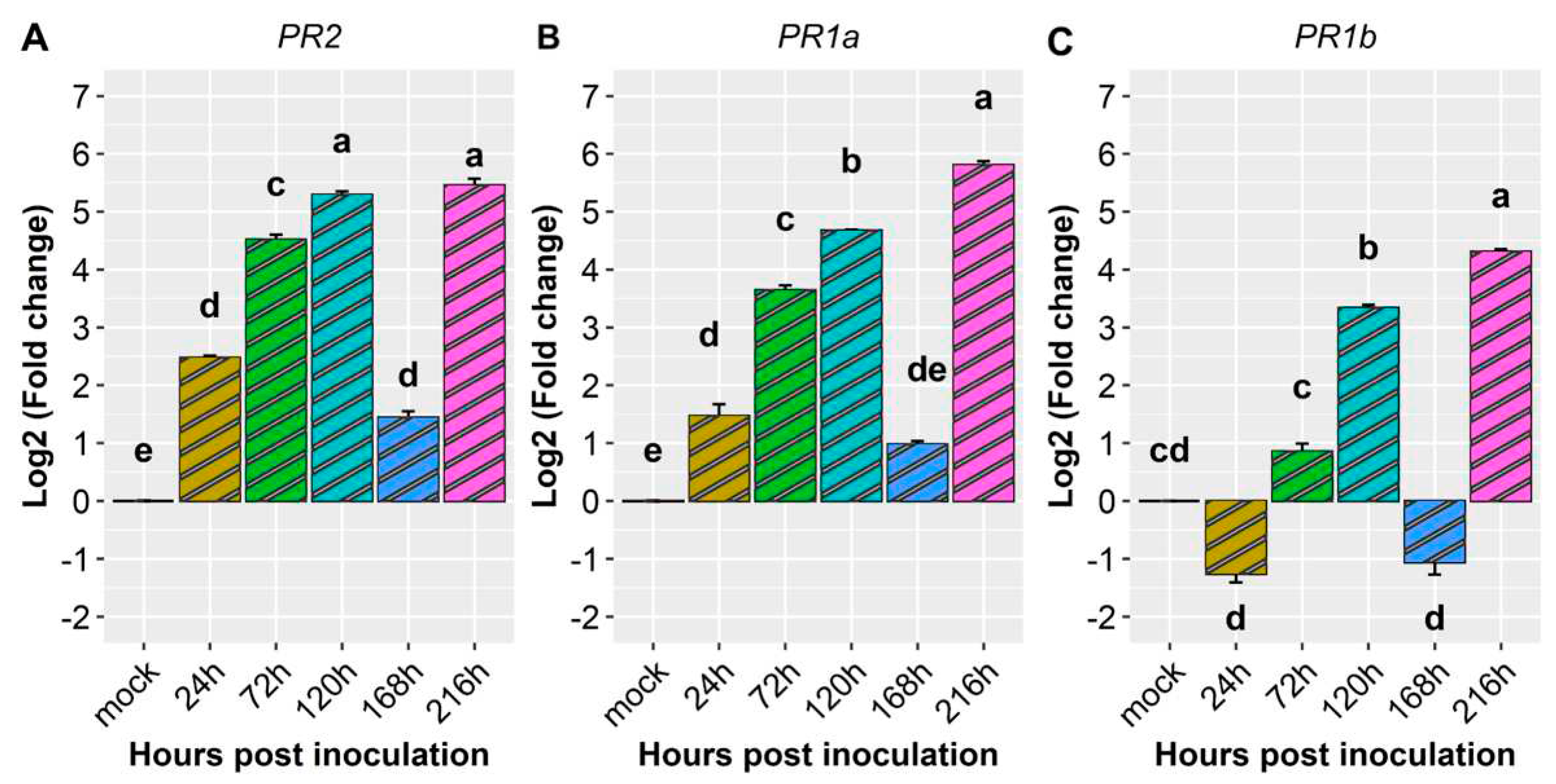
Figure 9.
Heatmap of relative expression of plant defense genes Phvul.003G109100, Phvul.006G196900, Phvul.009G256400 in the common bean cultivar California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. Yellow shading indicates higher expression and dark blue has lower expression than that of reference genes.
Figure 9.
Heatmap of relative expression of plant defense genes Phvul.003G109100, Phvul.006G196900, Phvul.009G256400 in the common bean cultivar California Dark Red Kidney at 24, 72, 120, 168 and 216 h post inoculation with race 63-39 of P. griseola and mock. Yellow shading indicates higher expression and dark blue has lower expression than that of reference genes.
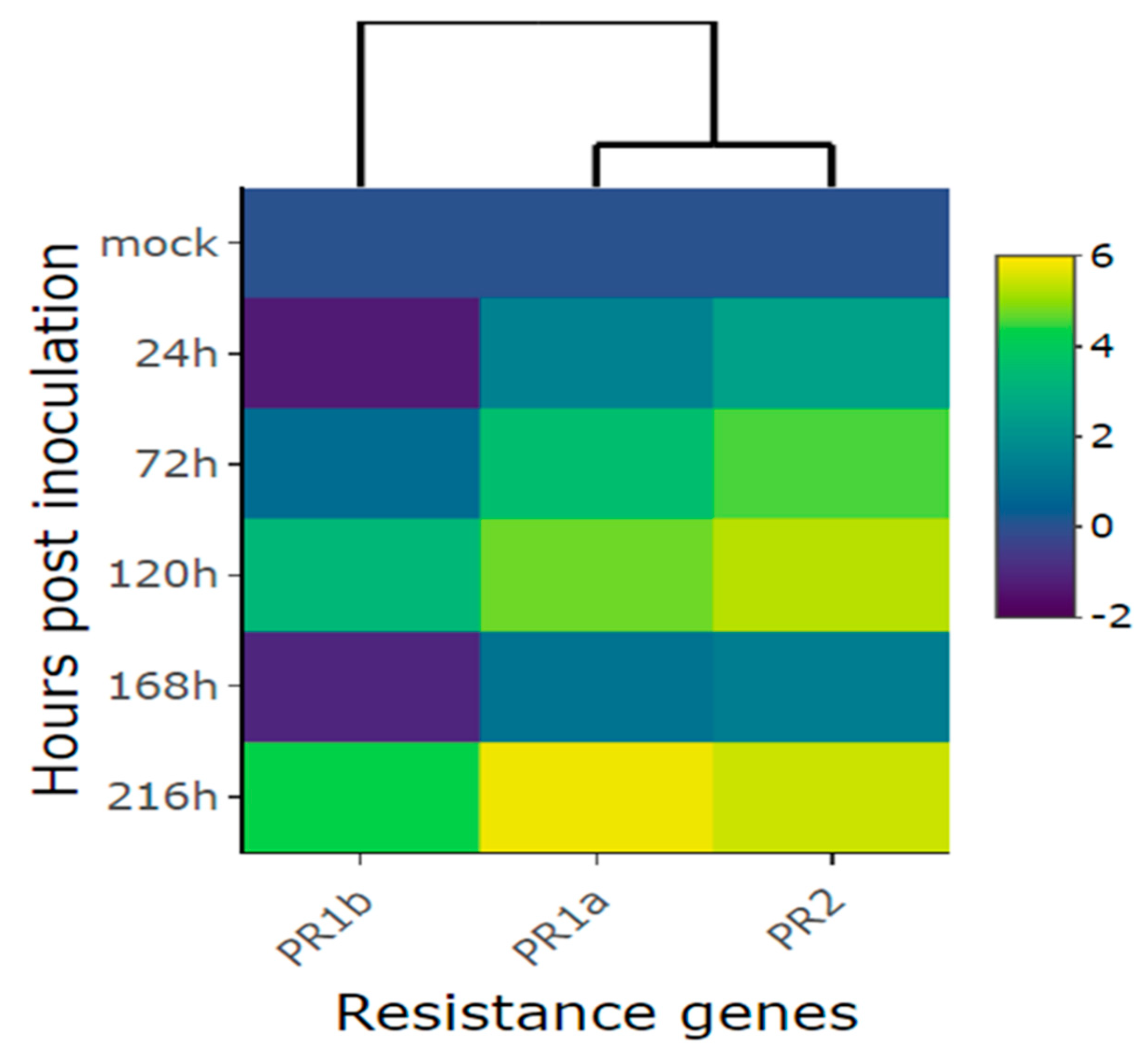
Figure 10.
Common bean chromosome Pv01 containing candidate genes for anthracnose resistance genes Co-x (KTR2/3), Co-1 (Phvul.001G243800), Co-AC (Phvul.001G244300, Phvul.001G244400, and Phvul.001G244500), and CoPv01CDRK/PhgPv01CDRK (Phvul.001G245300, Phvul.001G246000, Phvul.001G246100, Phvul.001G246200 and Phvul.001G246300).
Figure 10.
Common bean chromosome Pv01 containing candidate genes for anthracnose resistance genes Co-x (KTR2/3), Co-1 (Phvul.001G243800), Co-AC (Phvul.001G244300, Phvul.001G244400, and Phvul.001G244500), and CoPv01CDRK/PhgPv01CDRK (Phvul.001G245300, Phvul.001G246000, Phvul.001G246100, Phvul.001G246200 and Phvul.001G246300).
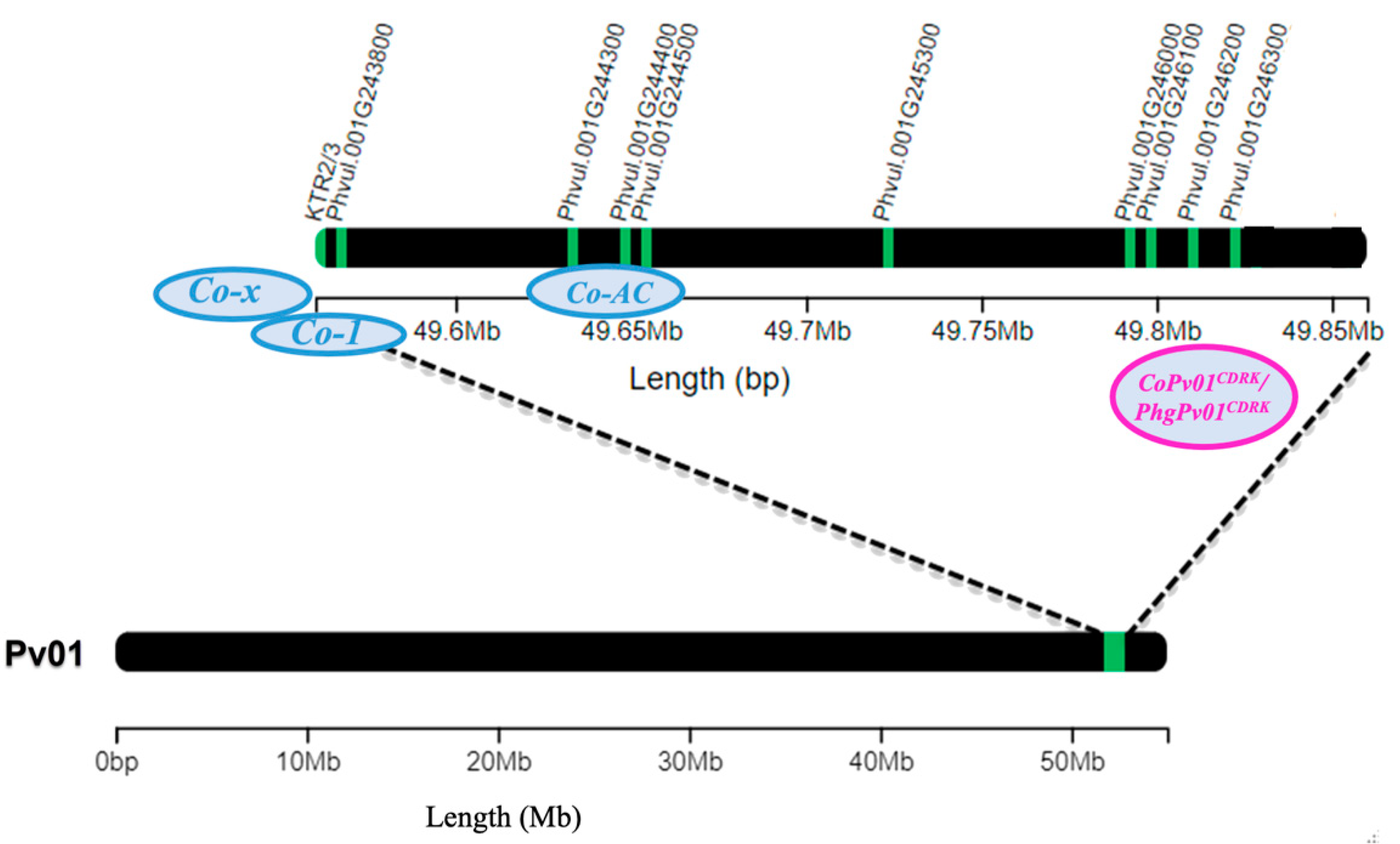

Table 1.
Gene model, previously mapped genes and functional annotation using Phytozome.
| Gene model | Gene | Functional annotation on Phytozome |
|---|---|---|
| Phvul.001G243800 | Co-1 | Serine/Threonine-protein kinase-like protein CCR3- related |
| KTR2/3 | Co-x | Serine/Threonine-protein kinase-like protein CCR3- related |
| Phvul.001G244300 | Co-AC | Clathrin Heavy Chain (CLTC) involved in plant defense signaling |
| Phvul.001G244400 | Co-AC | Unknown function |
| Phvul.001G244500 | Co-AC | Helix-loop-helix DNA-binding domain with possible transcription function |
| Phvul.001G245300 | CoPv01CDRK/ PhgPv01CDRK | Protein tyrosine kinase (pkinase_tyr) //leucine-rich repeat n-terminal domain (lrrnt_2) |
| Phvul.001G246000 | CoPv01CDRK/ PhgPv01CDRK | ATP-dependent RNA helicase ddx55/spb4 [ec:3.6.4.13] (ddx55, spb4) |
| Phvul.001G246100 | CoPv01CDRK/ PhgPv01CDRK | Cation-dependent mannose-6-phosphate receptor |
| Phvul.001G246200 | CoPv01CDRK/ PhgPv01CDRK | Protein trichome birefringence-like 33 |
| Phvul.001G246300 | CoPv01CDRK/ PhgPv01CDRK | Abscisic acid receptor pyl5 |
| Phvul.003G109100 | PR1a | Pathogenesis-related protein Bet v I family |
| Phvul.006G196900 | PR1b | Pathogenesis-related protein 1 |
| Phvul.009G256400 | PR2 | Pathogenesis-related protein 2 |
Table 2.
Summary table of mean relative gene expression [Log2(fold change)] of CoPv01CDRK/PhgPv01CDRK candidate genes and pathogenesis-related genes in response to ANT and ALS in CDRK cultivar.
Table 2.
Summary table of mean relative gene expression [Log2(fold change)] of CoPv01CDRK/PhgPv01CDRK candidate genes and pathogenesis-related genes in response to ANT and ALS in CDRK cultivar.
| Gene | Gene model | C. lindemuthianum race 73 (ANT) | P. griseola race 63-39 (ALS) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 24 hpi | 48 hpi | 72 hpi | 96 hpi | 120 hpi | 24 hpi | 72 hpi | 120 hpi | 168 hpi | 216 hpi | ||
| Co-x | KTR2/3 | 1.4 | 1.3 | 3.1 | 0.5 | 0.5 | 0.2 | 0.8 | 0.8 | -0.1 | -1.4 |
| Co-1 | Phvul.001G243800 | 1.4 | 0.9 | 2.0 | 1.8 | 0.6 | -1.1 | -0.2 | 0.1 | -0.9 | -0.3 |
| Co-AC | Phvul.001G244300 | -0.2 | -0.9 | -0.2 | -1.4 | -1.3 | -0.8 | -0.1 | 0.2 | 0.2 | 0.0 |
| Phvul.001G244400 | 0.0 | 0.0 | -0.7 | -0.4 | -0.2 | -0.2 | -0.2 | -0.3 | -0.7 | -1.0 | |
| Phvul.001G244500 | 0.4 | 0.3 | -0.4 | -0.5 | 0.8 | -2.4 | -2.5 | -2.3 | -2.3 | -2.4 | |
| CoPv01CDRK /PhgPv01CDRK | Phvul.001G245300 | 3.7 | 2.5 | 3.4 | 3.3 | 3.5 | -1.4 | 0.0 | 0.4 | -1.7 | 0.7 |
| Phvul.001G246000 | 0.9 | 0.8 | 0.8 | 0.7 | 0.3 | -0.5 | 0.4 | 0.5 | -0.1 | -0.2 | |
| Phvul.001G246100 | 0.6 | 1.5 | 1.2 | 0.7 | 1.3 | 0.0 | 0.3 | 0.1 | 0.4 | 0.0 | |
| Phvul.001G246200 | 2.7 | 2.6 | 2.6 | 0.5 | 1.6 | 0.0 | 1.5 | 0.1 | -0.4 | -0.2 | |
| Phvul.001G246300 | 7.2 | 7.3 | 7.8 | 7.4 | 6.3 | 1.7 | 8.5 | 6.5 | 1.3 | 6.1 | |
| Pathogenesis-related genes | PR1a | 6.6 | 6.2 | 8.0 | 8.1 | 6.3 | 1.5 | 3.6 | 4.7 | 1.0 | 5.8 |
| PR1b | 13.6 | 13.9 | 14.1 | 14.2 | 16.7 | -1.3 | 0.9 | 3.3 | -1.1 | 4.3 | |
| PR2 | 10.2 | 10.1 | 11.3 | 11.4 | 11.0 | 2.5 | 4.5 | 5.3 | 1.4 | 5.5 | |
Table 3.
Target genes, primers used, qPCR product size (amplicon), primer melting temperature (Tm), amplification efficiency (E) and coefficient of determination of linear regression (R²).
Table 3.
Target genes, primers used, qPCR product size (amplicon), primer melting temperature (Tm), amplification efficiency (E) and coefficient of determination of linear regression (R²).
| Gene Model a | Reference | Primers Forward (F) and Reverse (R) (5`-3`) | Amplicon(bp) | Tm(°C) | Eb | R² c |
|---|---|---|---|---|---|---|
| Phvul.001G133200 | IDE | F: AAGCAGGTATCTTGGCCATCTC | 126 | 60.16 | 1.04 | 0.99 |
| R: AAAGCAAACTCCAAGCTCCAATC | 59.99 | |||||
| Phvul.008G011000 | ACT | F: GAAGTTCTCTTCCAACCATCC | 154 | 59.67 | 1 | 0.98 |
| R: TTTCCT TGCTCATTCTGTCCG | 58.38 | |||||
| Phvul.001G243800 | Co-1 | F: CCTCAAGGTGGGGCTTTTGAG | 118 | 61.16 | 1.04 | 0.99 |
| R: TCACCGAGAAACTCCCATTGC | 60.61 | |||||
| KTR2/3 | Co-x | F: ATGCACAGGGGAATGGGATG | 279 | 60.11 | 1.04 | 0.98 |
| R: GCCATAGCGAGTGAGAGTGCG | 63.42 | |||||
| Phvul.001G244300 | Co-AC | F: GAAACGTCTCCGCAGAATAGTG | 150 | 59.4 | 1.03 | 0.99 |
| R: GTCTTGTGTTGTTCCTTGGAGTTG | 60.44 | |||||
| Phvul.001G244400 | Co-AC | F: TACAGCAAGAGAGCGGTTAAAGG | 121 | 60.62 | 1.01 | 0.99 |
| R: CCCTTTGTCACTTTGTTTTGAAGC | 59.67 | |||||
| Phvul.001G244500 | Co-AC | F: CAATGCACAGCTCGCAACTC | 141 | 60.45 | 1.09 | 0.97 |
| R: GGAACTGTGAAAGCTCTGCTAAC | 59.81 | |||||
| Phvul.001G245300 | CoPv01CDRK/ PhgPv01CDRK | F: TCTGCTGGAAGGGTGGTAGTC | 93 | 61.17 | 1.07 | 0.98 |
| R: GGACGTTATGTGAACAAGGTTTGC | 61.08 | |||||
| Phvul.001G246000 | CoPv01CDRK/ PhgPv01CDRK | F: ATGAAGCGGATGGATGTCTTG | 132 | 58.43 | 1.01 | 0.97 |
| R: TCTACGAAGCTTAGGCAATTGAG | 58.57 | |||||
| Phvul.001G246100 | CoPv01CDRK/ PhgPv01CDRK | F: CACGGTATCCTCAGCGAAGAC | 119 | 60.53 | 1.05 | 0.99 |
| R: CAGCAGTCAGCACATACTGGAG | 60.99 | |||||
| Phvul.001G246200 | CoPv01CDRK/ PhgPv01CDRK | F: GAAGGAGGCTGTGACGTGTTC | 150 | 61.2 | 1.04 | 0.99 |
| R: CCATCGCCACCGTTGATACTC | 61.13 | |||||
| Phvul.001G246300 | CoPv01CDRK/ PhgPv01CDRK | F: CTTCTTCCCTTCACTTCGATACC | 87 | 58.57 | 1.09 | 0.99 |
| R: GTTGAGAGTGTTTGTGGCAGT | 58.98 | |||||
| Phvul.003G109100 | PR1a | F: GTCCTAACGGAGGATCACTCA | 148 | 58.62 | 1.06 | 0.99 |
| R: CAGGGATTGGCCAGAAGGTAT | 59.5 | |||||
| Phvul.006G196900 | PR1b | F: GGTTTGCCTATGATCCCAATGC | 115 | 59.96 | 1.03 | 0.99 |
| R: TGTTGTGAGCGTTGAGGAAGTC | 61.06 | |||||
| Phvul.009G256400 | PR2 | F: CAGAGGTTCTCATTTGCTGCTTTC | 98 | 60.62 | 1.07 | 0.99 |
| R: ATGCCATAACACACCCCGATTTG | 61.75 |
a Based on the Phaseolus vulgaris genome available on the Phytozome 12 platform: https://phytozome.jgi.doe.gov/pz/portal.html#; * Reference genes; b Amplification efficiency obtained from the Equation E=[10(-1/slope)]- 1 (Rasmussen, 2001); c Coefficient of determination of linear regression.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



