
Submitted:
04 September 2023
Posted:
05 September 2023
You are already at the latest version
Abstract
Intestinal microbiota attracts daily attention of a growing number of study which have attempted to link gut dysbiosIs with a variety of disease states: irritable bowel syndrome (IBS), inflamed bowel disease (IBD), Crohn's disease (CD), leaky gut syndrome (LGS), food intolerance, diabetes, metabolic syndrome, cancer, etc..
In our study we analyzed how intestinal dysbiosis may be related to chronic fatigue syndrome (CFS) and depression through the exchange of information through the gut-brain axis (GBA).
We studied 33 subjects, 13 males and 20 females, who reported CFS or/and depression: we investigated their salivary cortisol levels, blood serotonin, omega 3/6 ratio, intestinal dysbiosis (calculated on the urinary levels of indoxyl sulfate and skatole), and we looked for the presence of Candida a. or mycetes in the stool; the data accumulated with this research show a correlation between the presence of Candida a./miceti, indoxyl sulfate urine values beyond the physiological and low serotonin levels. In addition, data analysis showed that the EPA/DHA values also show pro-inflammatory levels in case of dysbiosis and low serotonina levels.
The relationship, however, with cortisol levels requires further research although this study showed a statistically significant positive correlation between these values, measured at specific times, and serotonin levels.
Aims its connections with
We investigated the relationship between stress (evaluated through the measurement of salivary cortisol levels) and gastrointestinal efficiency measured as a function of intestinal fermentative and putrefactive dysbiosis, evaluating the levels of urinary indoxyl sulfate in the first case (a possible correlation with the presence of Candida spp or Mycetes in the subjects feces was investigated), urinary skatole levels in the second one, in patients with chronic fatigue syndrome (SFC) and depression. In these patients we also have studied omega 3/6 ratio, and finally we have analized the impact that the alteration of these parameters can have on the serotonin levels.
This research attemps to highlight the contact points, in some cases not so obvious, among these topics, contact points that, although they give us interesting indications, show the need to be further deepened by analyzing a larger amount of data.
Keywords:
stess
; chronic fatigue syndrome
; depression
; dysbiosis
; serotonin/cortisol
Introduction
All recent studies agree on the need to look at the intestine as a complex and interconnected structure with numerous systems starting with the nervous one in which the exchange of information and mutual influences is so strong that it has supported the need to identify a new axis: the gut-brain axis – GBA - (Gherson M.D, 1998; Hong-Xing Wang, Yu-Ping Wang, 2016). Thanks to this emerging vision, the microbiota now plays a very prominent role, so much so that the old description of the intestine as an "empty tube" now appears unreal in light of the impressive amount of bacteria that crowd the intestinal lumen: 1014 microorganisms belonging to about 400 different species (Koboziev I. et al., 2014; Eckburg P.B. et al., 2005) that literally crowd it with 9 million genes (Li, J. et al., 2014), whose collaboration is essential to keep us healthy, for this reason the concept of microbiota understood as a virtual organ (Evans J.M. et al., 2013) or even forgotten organ (O’Hara A.M., Shanahan F., 2006) has gained ground. This is an interesting aspect of intestinal microbiology that has once again put researchers in difficulty because of the classic definition of organ intended as completely differentiated functional unit, and the microbiota does not fall into this classification, despite being involved in different metabolic activities through the synthesis of molecules involved in numerous metabolic functions (Mayer E., 2016).
The composition of the microbiota is strongly influenced by a multiplicity of factors, including the intake of fiber with the diet (Figure 1), an element that has changed over the years and since the so-called western diet began to spread, it has known a drastic reduction in favor of refined carbohydrates (King D.E., Mainous A.G., Lambourne C.A., 2012), at the same time the chronic inflammatory pathologies increased in general (Bach J.-F. 2002; Eder W., Ege M.J., von Mutius E. 2006). Impressive, in this sense, is the amount of works that link the alteration of the microbiota to environmental and oxidative stress (Weiss G. Adrienne, Hennet Thierry, 2017), and to inflammatory diseases such as diabetes and colon-rectum cancer (Jurjus A. et al., 2016), irritable bowel syndrome - IBS - (Balsari et al., 1982; Bhattarai Y. et al., 2017) and inflamed bowel disease - IBD - (Rapozo et al., 2017; Ni J. et al., 2017), to Crohn’s disease of –CD- (Manichanh C., et al., 2006; Caparros E. et al., 2021), to Leaky Gut Syndrome (Jerrold R. Turner, 2009) in turn linked to asthma (Benard A. et al. , 1996; Hijazi Z. et al., 2004), to polycystic ovary syndrome (Lindheim L. et al., 2017), fibromyalgia (Othman M. et al., 2008; Erdrich S. et al., 2020), autism (Song Y. Et al., 2004; Mayer E.A. et al., 2014), autoimmune diseases (Yusuke K., Koji H., 2021), with heart failure (Mengchao J., Zhiyuan Q. et al., 2019) and hypertension (Yang T., Santisteban M.M. et al., 2015); we must not forget how nutrition (Lawrence A. et al., 2014), as well as the use/abuse of drugs and lifestyle, play a leading role in this sense (Charlie G. Buffie et al. al., 2011).
However, there is another factor capable of profoundly influencing the composition of the microbiota: stress (Bharwani A. et al., 2017) and its reverberation on the autonomous nervous system (ANS).
This study arises from analysis of data relating to patients who have come to the Istitute of Biological Medicine in Milan (IMBIO, Italy) to find relief from their disorders (depression and CFS).
My starting point was considering these pathologies as the expression of a condition of functional disregulation with respect to the reactivity of the biological system to stress (studied through analysis of the circadian levels of cortisol) as well as a functional imbalance that is realized through an inflammatory state (assessed through the omega ratio 3/6). The latter, in particular, linked to a condition of intestinal dysbiosis measured by urinary skatole (to evaluate putrefactive dysbiosis) and indoxyl sulfate levels (to evaluate fermentative dysbiosis) as well as in the analysis of the stool to determine the levels of Candida a. or mycetes among patients. Finally, all these parameters have been related to serum levels of serotonin in order to evaluate any correlations.
In short we wanted to analyze how a condition of intestinal dysbiosis can be related to CFS and depression by the exchange of information through the gut-brain axis (GBA) measured through serum serotonin levels, and how this exchange can be influenced by adrenal reactivity (intended as a measurement of salivary cortisol) and the omega3/6 ratio because of their link with depression (Liu J.J. et al., 2013) and chronic fatigue syndrome -CFS- (Puri B.K., 2007).
Systemic approach to Stress
The systemic approach applied to biology begins to make its way overwhelmingly thanks to the concept of System (Bertalanffy, 1967) and to Cybernetics (Wiener N., 1948), but already the experimentally measurable effects that emotions and stress produce on the organism in general and gastrointestinal efficiency in particular, had been tackled first with Cannon’s work (Cannon, 1932) and then with Selye’s (Selye H., 1976-1978). Selye, in fact, had shown that if an organism is subjected to a psychic stress, what we should expect is its manifestation in an organic key.
In short, attention is shifted from the single component of the system, to the relationships that the elements contract as a whole and how this interaction translates into a complex with characteristics different from the individual components of which it is made up (Figure 2).
This new point of view highlights another characteristic of living organisms clearly indicated by Bertalanffy: being open systems. Basically an open system, that is every living being (Burton, 1939), is a system that exchanges matter with the surrounding environment, and to work in such a way as to produce and decompose structures thanks to its own constitutional organic resources; however, the information we receive from the outside disturbs the system, producing changes in its operation. The bacteria that crowd our intestine are a fundamental part of these interactions and from the type of relationship we are able to establish with the forgotten organ, it follows incoming and outgoing signals capable of influencing the functioning of distant structures such as the brain (Clarke G. et al., 2014).
In relation to the effects that stress produces on our organism, the fundamental difference posed by Selye between eustress and distress should be noted. In both cases, in fact, the body responds in a non-specific and independent manner to the nature of the stressor but if the first is to be considered physiological, the second creates the conditions for the onset of the disease. From his studies it emerges that each phase of stress is accompanied by an increased energy demand, cortisol is a pivotal hormone in relation to stress reactivity: it is a molecule produced by the adrenal cortex (Marrama P., Angeli A., 1992; Larsen P.R., 2003) by activating the stress axis later to chronic stress of the organism.
Once the stressor, that is any internal or external factor capable of disturbing the biological balance, has been overcome, the organic system will move towards a physiological phase of recovery (Lazarev, 1947; Brekman, 1980, Selye, 1976) of those previously mobilized resources; on the other hand, if the stress extends well beyond the resistance limit of the system, the body would enter a phase of exhaustion, hence the disease (Figure 3).
The regulation of cortisol levels follows a fine regulation controlled by the stress axis that relates the hypothalamus, pituitary and adrenal glands via the HPA axis (Packard Amy E.B., Egan Ann E. et al., 2020); it should also be emphasized that the physiological levels of cortisol follow a pulsating trend in perfect agreement with our circadian rhythm (Androulakis P. Ioannis, 2021), therefore a maximum peak is recorded between 06:00 and 08:00 in the morning and then see its values gradually decrease until they touch a minimum between 21:00 and 23:00 in order to predispose the organism to night rest (Figure 5- Schmidt R.F. et al., 2003).
Cortisolemia is now commonly measured in saliva (Kirschbaum C. and Hellhammer D., 1994) in the four physiologically significant ranges: 7:00, 13:00, 17:00 and 23:00; thanks to these measurements we are able to trace a cortisolemic profile of the person in order to evaluate his reactivity to stress (Figure 4).
In conclusion, a subject subjected to a stressor will react through a first phase of alarm accompanied by the activation of the adrenal medulla and the consequent release of catecholamines, to then move on to the next phase of resistance with the activation of the adrenal cortex and consequent elevation of the levels of cortisolemia (Figure 3).
It is important to underline how the internal organs receive the simultaneous innervation of both systems (OS and PS), and that the effects of the activation of the two vegetative components on the aforementioned organs are antagonistic (Figure 5).
The two systems work together to allow the organism to tend to the maintenance of a condition of physiological equilibrium or homeostasis (Cannon W.B., 1932), a vision that has undergone a profound re-evaluation thanks to the work of Bertalanffy (Bertalanffy, 1967) on systems, with the introduction of the concept of steady state (Figure 6).
It is a more complex conception of equilibrium of the biological system, which includes the idea of equifinality. Open biological systems tend towards a time-independent state: the steady state, an equilibrium, without ever reaching it fully (and that is why they are capable of producing work). The tendency towards a steady state is independent of the initial conditions of the system, therefore unlike what occurs in closed systems, the final state is achieved thanks to equifinality, although starting from very different initial conditions and after the process has been perturbed in several ways. These concepts are applicable in many sectors of biology (Beier, 1962, 1965; Locker et al., 1964, 1966): in the human body, in fact, many tissues remain in a stationary state while their cells continuously go into apoptosis, or are in any case eliminated, to be continuously replaced.
In the light of what has been illustrated, we can affirm that both if the organism is in a situation of sympathotonic hyper-reactivity (valid for both acute and chronic), and hyper-parasympatheticotonia (with the mobilization of the mediators of inflammation), the biological system will, however, tend towards its stationary equilibrium over time. Obviously, all this is achieved as long as the system is able to restore balance from the autonomic and biochemical point of view; but what happens if the ANS finds itself stuck in one phase rather than in its opposite phase?
In case it were unable to interrupt the activation of the OS, if the stressor (physical or emotional) continues over time, the system will go through a first acute phase (alarm) which is characterized by the presence of catecholamines, followed by the subsequent chronic phase (resistance ) that will last as long as the organic resources will allow, in addition to this point the biological system will pass the point of no return, delivering itself to the disease phase understood as a biological standstill that allows the body to recover the resources already committed to the response to stress.
In case it was unable to interrupt the activation of the PS, the biological system is blocked in a phase predominantly dominated by the PS and characterized by intense vagotonia caused by insufficient vagolytic action (condition of chronic inflammation).
The intestinal mucosa, nutrition, bacteria and inflammation
The intestinal mucosa represents our real interface with the surrounding world since on average around 60 tons of food transit throughout our life cycle (Bengmark S., 1998); this is an extremely relevant detail since it is food the fundamental element to determine the well-being of the individual in a more general perspective, the composition and activity of the microbiota in a more particular one (Wu G.D., et al., 2011; Muegge B.D., et al., 2011; Duncan S.H., et al., 2007; Ley R.E. et al., 2006; Walker A.W., et al., 2011) however framed in a long-term perspective. We must always remember that it is thanks to a correct microbiota that we are able to effectively stimulate our immune system, produce vitamins (of group B and K), activate the motility and function of the gastrointestinal tract, perform digestive and absorption processes correctly, to prevent colonization and the consequent pathogenicity of any pathogens, the production of short-chain fatty acids (Figure 1) - SCFAs - (Holzapfel W.H. et al., 1998; Macfarlane G.T., Macfarlane S., 1997), in particular butyric acid a fundamental molecule for the trophism of the intestinal epithelium as well as performing a very important anti-inflammatory function (Lawrence A.D. et al., 2014).
Finally, we must remember that the products of bacterial metabolism represent 40% of the molecules in circulation (Mayer E., 2016).
As regards the composition of the microbiota, it should be emphasized that it is an extremely delicate balance, subject to many variables capable of profoundly influencing its fate, in particular in relation to the effect that antibiotic therapy shows on the intestinal bacterial composition (Hawrelak J.A., Myers S.P., 2004, see tables), even if limited to a single dose (Buffie C.G. et al., 2011) it is able to produce a very substantial increase in Enterobacteriaceae. An alteration of this balance leads to the onset of particularly dangerous conditions from an inflammatory point of view, both local and systemic, an alteration that manifests itself, for example, with the decrease in the presence of Faecalibacterium prausnitzii, an obligate anaerobic microorganism used as a marker of intestinal eubiosis, and the simultaneous increase in the proliferation of a facultative anaerobic bacteria such as E.coli and Enterobacteriaceae in general, rather than real aerobic bacteria; in this way the presence of bacteria belonging to these last two groups is a useful indicator for determining a condition of intestinal dysbiosis (Rigottier-Gois L., 2013). It is interesting to note that the increase in facultative anaerobic bacteria is recorded not only in the case of inflammation-induced dysbiosis, but also following treatment with antibiotics (Pettenkofer von M., 2015).
In relation to the effects that nutrition is able to produce on the composition of our microbiota, it has been shown that a few days of an unbalanced diet in favor of animal proteins, and devoid of fibers, are sufficient (Lawrence A.D. et al., 2014) to produce a drastic alteration of the intestinal microbiota. In fact, there is a net decrease in Faecalobacterium prausnitzii, therefore of dysbiosis considering that the eubiotic flora is anaerobic (Rigottier-Gois L., 2013), combined with the growth of Bilophila wadsworthia (microorganism producer of H2S, a powerful intestinal pro-inflammatory agent), has allowed us to understand how the bacterial composition present in the lumen of our intestine is extraordinarily sensitive to the type of foods that pass through it. In particular, the proliferation of Bilophila wadsworthia makes it possible to create a link between fats introduced with the diet, bile acids and the growth of bacteria capable of developing a condition of inflammation of the mucosa (Devkota S., et al., 2012). The considerations made so far clearly highlight how the changes in the Western lifestyle, which have occurred so rapidly in recent decades, characterized by a strong increase in the dietary consumption of fats and sugar, have altered the composition and metabolic activity of the human microbiota (Turnbaugh P.J. et al., 2009).
As just mentioned, the mucosa is continuously exposed to bacterial products - endotoxins (van Deventer S.J., 1988); hydrogen sulfide (Cummings J.H., 1997); phenols, ammonia and indoles (Macfarlane S., 1995) - which can produce harmful effects both on the mucosa itself and on our health (Macfarlane S., 1995). The presence of many of these toxic metabolites depends on the type of fermentation that develops in the intestine, which in turn depends on the type of bacteria that the intestine itself receives, as well as the substrates available for fermentation.
Dysbiosis
A diet rich in protein, and low in fiber, is able to determine the formation of highly toxic products with decidedly pro-inflammatory characteristics. Already in the early twentieth century, Metchnikoff postulated how diseases and biological aging itself were generated by the putrefaction of proteins in the intestine and how the intake of probiotics was recommended to favor detoxification and support a correct bacterial composition (Metchnikoff E., 1907; Tannock G.W., 1997). This is the toxicological theory of the intestine which soon turned into the term dysbiosis coined by Metchnikoff himself (Murray M., Pizzorno J., 1998) to describe a pathogenic bacterial alteration in the intestine; while others later referred to dysbiosis to indicate: "... qualitative and quantitative changes in the intestinal flora, their metabolic activity and their local distribution." (Holzapfel W.H. et al., 1998). At this point, dysbiosis must be considered as a state in which the microbiota produces dangerous and harmful effects through:
- -
- Qualitative and quantitative alterations of the microbiota itself;
- -
- Changes in the metabolic activity of intestinal bacteria;
- -
- Variations of their local distribution.
Starting from these assumptions, the proliferation of potentially pathogenic bacteria in the intestine is the cause of the release of numerous toxic products, previously mentioned, which end up playing a leading role in numerous chronic and degenerative diseases. In short, dysbiosis opens the door to possible interventions of great prospect in numerous chronic degenerative pathologies on an inflammatory basis.
Tryptophan, serotonin, melatonin and intestines
Brain serotonin levels are closely related to the intake of tryptophan in the diet. The enzyme most involved in its metabolism is tryptophan-hydroxylase which, to be activated, requires adequate amounts of folic acid, vitamin B6 and magnesium.
Tryptophan is an essential amino acid capable of crossing the blood-brain barrier (BBB) and reaching the brain; this is a very relevant detail since it is in the brain that its transformation into serotonin or melatonin takes place, a process linked to the light/dark, sleep/wake cycle. The mechanism illustrated in Figure 7 is the one which leads to the production of serotonin and melatonin in the brain under the control of the pineal (through the control of the serotonin-N-acetylase enzyme) thanks to the electrical activity of the suprachiasmatic nucleus (Foster R., Kreitzman L., 2007). We know how tryptophan is able to pass the BBB, however this passage is difficult if it must be carried out in the presence of other amino acids that preferably binding with the receptor involved, thus preventing the tryptophan from crossing the BBB. Wurtman, director of the clinical research center at Massachussetts Institute of Technology (MIT) was able to demonstrate how the characteristics of the meal, in terms of the prevalence of carbohydrates or proteins, is able to influence the amount of tryptophan available for the synthesis of serotonin in the brain (Wurtman R.J. et al., 2003). Thus, the amount of serotonin produced in the brain is directly linked to the availability of its precursor tryptophan, the latter being able to pass effectively to the brain if the meal is more based on carbohydrates than on proteins (Femstrom J.D., Wurtman R.J., 1971), the cause is to be found in the competition that is generated between the tryptophan itself and the other amino acids, to bind to the same receptor that allows the passage of the BEE (Bottaccioli F., 2007): amino acids such as tyrosine, valine, methionine, etc., called even large neutral amino acids (LNAA) show the tendency to preferentially bind to the receptor just mentioned, thus preventing the entry of tryptophan into the brain and the consequent synthesis of serotonin/melatonin; for this reason the ratio between tryptophan and the other amino acids is shifted in favor of the former when the meal is mainly composed of carbohydrates.
But these are not the only variables with respect to the conditions that can influence the levels of serotonin: subjects who face a condition of psycho-physical stress or who complain of emotional instability, record an increased consumption of tryptophan (Teradaira R. et al., 2007) measured as a decrease in its plasma levels. As proof of this evidence it has been shown (Walderhaug E. et al., 2007) that if the availability of tryptophan is reduced to healthy subjects, or in people who have recovered from depression, there is a decrease in mood and the reappearance of depressive symptoms which in males presented itself as anger associated with aggression and impulsive attitude, while in females it manifests itself as closure in itself.
It should also be emphasized that an excessive proliferation of E. coli is also capable of producing a decrease in serotonin levels since it uses tryptophan to produce indole and after hepatic metabolism it is eliminated as indoxyl sulfate in the urine (Smith T., 1897); this leads to a drastic decrease in intestinal serotonin levels which ends up reflecting on mood by virtue of the gut-brain axis (Gershon M.D, 1998; Hong Xing Wang, Yu Ping Wang, 2016).
However, there is a crucial aspect that draws our attention to the relationship between the intestine and serotonin, which consists in the fact that 95% of the serotonin in our body is synthesized in the intestine, while only 1% sees the light in the brain.
Gut-Brain Axis (GBA), stress and microbiota
I have already mentioned how compromising the delicate balance of our microbiota leads to a condition of dysbiosis, a condition that is evidenced by a deficit of intestinal function and mood disorders, both connected with the disruption of the GBA (Simrén M. et al., 2013; Berrill J.W. et al., 2013) as is shown in Figure 8.
In a work published by the Journal of Clinical Psychiatry (Lydiard R.B., 2001), the researchers divided mice into two groups: one control, while the other brought together mice previously infected in the intestine with Campylobacter jejuni, the latter group showed neither signs of infection or circulating inflammatory cytokines, however they were clearly more anxious than the controls. At this point the question was how communication between the intestine and the brain is achieved since no circulating mediators linked to infection and inflammation are measured. The answer to this question must be sought in the particular anatomy of the gastrointestinal system in general, in the intestine in particular, because the signal from the periphery to the center is transported by the viscero-sensory nerves that innervate the intestine, they are pathways that carry their signals to the brain, signals coming from the presence of bacteria in the intestinal lumen, which influence its functioning.
Vagal innervation, in particular, develops a close relationship with the immune cells of the submucosa (Williams et al., 1997; Stead R.H. et al, 2006), projecting its nerve endings up to the mucosa. The signals coming from the bacterial presence are able to influence brain functions through their transport through both the vagus nerve and the fibers of the spinal visceral sensory nerves (Lydiard RB., 2001). Furthermore, it seems that the stimulation of the intestinal peripheral vagal nerve fibers represents an extremely efficient immunosensory signal, a faster and more privileged way of communication in relation to the presence of bacterial infection than circulating cytokines (Goehler L.E. et al., 2005) than in these cases they are not even recorded.
Several studies (Kamiya et al., 2006; Liu et al., 2007) bring to our attention how the viscerosensory nerves provide a complex contribution to the dysfunctional response of the intestine associated with stress; in particular, the stimulation of the vagal intestinal component would produce anxious-depressive states in many patients.
Thanks to recent studies, data has been collected to support the thesis according to which regions of the brain involved in emotions are associated with autonomous viscerosensory signals (Craig A.D., 2003; Nauta, 1971; Price, 1999; Zagon, 2001); in other works which wanted to study the immune reactivity with respect to bacterial products (LPS) or cytokines (IL-1) that usually induce depressive symptoms, have shown how these immune stimuli are able to activate brain regions involved in the processing of viscerosensory stimuli (Goehler et al., 2000).
A very interesting data is that the risk of IBS increases 10 times when the subject reports a previous gastroenteritis (Rodriguez L.A.G., Ruigomez A., 1999), in many cases it is the consequence of a gastroenteritis acute (Spiller, 2002), a condition called post-infectious IBS; it should be noted that subjects in whom the diagnosis of IBS is more recent, show anxiety rather than depression, while when IBS is chronic or resistant to treatments, patients present anxiety and depression together; finally, IBS itself is often accompanied by chronic fatigue syndrome (Ballanger J.C. et al., 2001; Goehler L.E., 2007).
What has been said so far allows us to highlight the clear association of a specific symptomatology, in particular chronic fatigue syndrome as well as anxiety and depression, with gastrointestinal disorders, especially as a consequence of a state of inflammation or infection (Basso et al., 2003; Castex et al., 1995; Lacosta et al., 1999; Rossi-George et al., 2005).
We know that an alteration of the microbiota, because of its participation in the final processes of digestion (Gershon M.D., 1998), is able to result in an alteration of the availability of nutrients, we also know how there is a close relationship between the nutrients themselves and the secretion of peptides by enteroendocrine cells, peptides capable of interacting with GBA (Uribe A. et al., 1994).
Finally, it is known how a multiplicity of stressors are able to modify the composition and the overall biomass of the intestinal microbiota and this independently of its duration over time. In fact, it has been shown that exposing a subject to a social stress lasting only two hours, so as to activate the response of the HPA axis, is sufficient to determine a remodeling in the bacterial profile and in the relationships existing between the different phyla (Galley J.D. et al., 2014), including the genus Lactobacillus (Bailey M.T., Coe C.L., 1999; Mackos A.R. et al., 2013; Schreiber O. et al., 2009; Barouei J. Et al., 2012; Bailey et al., 2010).
To conclude, it must always be remembered that the ENS represents the privileged target of bacterial metabolites; one of the most significant products in this sense, as I have already mentioned, are the SCFAs (acetic, butyric, propionic acid), substances capable of producing a stimulation of sympathetic fibers (Kimura I. et al., 2011), the release of serotonin from the intestinal mucosa (Grider J.R., Piland B.E., 2007), as well as influencing the memorization and learning processes (Stefanko D.P., 2009; Vecsey C.G. et al., 2007).
Chronic Fatigue Syndrome
Chronic fatigue syndrome (CFS) is a severely debilitating condition in which patients complain of a varied range of symptoms including constant fatigue, immune alterations as well as musculoskeletal pain (Hudson J.I. et al., 1992), with a very similar picture to IBS (Gomborone J.E. et al., 1996).
Patients suffering from CFS are simultaneously suffering from neurological impairment, cognitive dysfunction (McGregor N.R. et al., 1996; Caldarini M.I., et al., 1996) and gastrointestinal disorders.
A significant increase in facultative anaerobic bacteria within the microbiota of these patients is clearly recorded, as well as a condition of acidosis characterized by a high level of lactic acid (Sheedy J.R., Wettenhall R. et al., 2009). We know that lactic acidosis is a condition accompanied by neurological weakening that includes ataxia, dysarthria, weakness and difficulty concentrating (Uribarri J. Et al., 1998). The condition of acidosis may be due to an excess of carbohydrate fermentation (Shah M. et al., 2001), an increase in E.coli and the proliferation of facultative anaerobic bacteria (Sheedy J.R. et al., 2009). As I have already had the opportunity to illustrate, it is precisely when this type of bacteria begins to make their way, to the detriment of obligate anaerobes, that the picture of intestinal dysbiosis begins to take shape.
Omega-3 / Omega-6 ratio
Fatty acids perform structural, energy and metabolic functions in living organisms. The fatty acids of the Omega-3 and Omega-6 series represent a group of substances essential for our organism for the production of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) in the case of the fatty acids belonging to the first series called PUFA altogether, arachidonic acid (AA) as regards the second.
From the study of Figure 9, it can be easily deduced that the quality of the diet will therefore be decisive in establishing the plasma ratios of the fats of the two series: an unbalanced diet towards foods rich in Omega-6 will support a physiological tendency to amplify inflammatory processes; on the contrary, a prevalence of Omega-3 will produce the opposite effect.
Because of their ability to carry out a natural anti-inflammatory action (Calder P.C., 2006), Omega-3s are studied and used in the therapy and prevention of many pathologies: neuropsychiatric (Mazza M. et al., 2007), cardiovascular, in gastrointestinal cancer, etc. (Eltweri A.M. et al., 2017). The key role played by these substances is particularly important to us since their link has also been demonstrated in depression (Liu J.J. et al., 2013). Connecting us to cancer we must remember that the diagnosis of this disease alone leads to stress and sadness in the patient, so as to lead him to a major depressive disorder (MDD); it is important to point out how the emotional impact of the diagnosis is not the only cause for MDD: a fundamental role is even played from pro-inflammatory cytokines cancer-related depression (Bortolato B., 2017), but at the same time it is important to emphatize how in these patients were recorded cases of depression about four times greater than in other subjects (Freitas R.D.S., Campos M.M., 2019).
It is currently calculated that in the Western diet the Omega-6 / Omega-3 ratio is equal to 20-25: 1 (Bottaccioli F., Carosella A., 2011), and that this imbalance affects 70% of the American population (U.S. Department of Health and Human Services, 2005) therefore clearly pro-inflammatory.
Materials and Methods
Candida and Fungi: Biological samples are collected and carried with specific transport medium (Amies Swab). with standardized culture medium: enriched media and differential media.
Used media are:
CHROMAgar Candida: chromogen medium useful for the growth and semiquantitative, immediate identification from mixed yeasts cultures (Candida spp.).
Saboraud Agar: a selective medium useful for the isolation of fungi and yeasts from sanitary and clinical samples. So inoculated media have been incubated for 48 hours at 37°C in the presence of air. The purpose of the sowing of the samples is to obtain cultures from which it is possible to produce pure colonies of known pathogens, in order to proceed with the certain reporting of the microorganism found.
Salivary cortisol: the cortisol values were measured on a saliva sample collected at four different times of the day (7AM – 1 PM – 5 PM – 11 PM), on an empty stomach and away from the intake of any food or drink for at least 1 hour, using as a means of collecting salivettes®.
Enzyme immunoassay method: the Diametra Cortisol Saliva ELISA kit was used; the kit includes a plate pre-treated with the anti-cortisol antibody. It works thanks to the competition between cortisol and horseradish peroxidase (hrp): if the saliva sample contains little cortisol, the hrp binds to the antibody giving a blue colour to the sample; the more cortisol is present, the less dark the sample is. The addition of the so-called STOP solution stops the reaction between cortisol, antibody and hrp by changing the various samples to yellow of different intensity. Spectrophotometric reading at 450nm follows. By comparing the samples with the known values of the calibration curve (0 – 0.5 – 1 – 5 – 10 – 20 – 100 pg/ml) we can trace the concentration of the tested samples. The reference values vary within the day.
Dysbiosis and serotonin: the quantification of skatole is done in HPLC (High Performance Liquid Chromatography): after having purified the sample (5 ml of morning urine) by filtration with 0.4 μm filter syringes, it is analysed with a RID (refraction index detector) placed at the end of a selection column (NH4) containing resin suitable for separating the tryptophan products. The sample reading is compared with a calibration straight line prepared previously by reading known skatole concentrations (10 – 20 – 40 μg/l) and the sample content is then determined. The procedure is then repeated for the indoxyl sulfate with the difference of the calibration line (10 – 20 – 40 mg/l).
Serotonin (5ht) is also dosed by HPLC, in this case, however, serum (1 ml) is used as a biological sample, which is purified by centrifugation and subsequent filtration in 0.4 µm filter syringes.
A 75% acetonitrile eluent solution diluted in water is used for all three analyses.
The concentration of serotonin is read thanks to an optical UV reader placed at the end of a column (C4) containing resin capable of separating the proteins, it is then compared with a calibration line prepared previously by reading known concentrations of serotonin (100 – 150 – 200 – 250 μg/l); the concentration of the sample under examination is then determined.
EPA and DHA: Fatty acid extraction: 10 μl of human serum were mixed with 10 ml of Heptadecanoic acid 50 mM as Internal Standard and 10 ml of CH3CN (acetonitrile); therefore, 30 ml of Perchloric acid was added to deproteinize, and 800 ml of chloroform, 100 ml of methanol, 100 ml of n-heptane and 100 ml of H2O were added to extract the fatty acids.
After the solution was centrifuged at 3000 rcf for 5 min, the organic layer (400ml) was transferred to a dark tube where the solvent evaporated. A residue is collected under reduced pressure using a centrifuge concentrator, VC-36N (TAITEC Co., Ltd., Saitama, Japan) without heating for 20 min.
The dry residue was dissolved in 50 mM DMF and mixed with 50 ml of 2 mM DBD-ED in CH3CN, 50 ml of 140 mM TPP in CH3CN and 50 ml of 140 mM DPDS in CH3CN to derivatize the fatty acids. After waiting at room temperature for 120 min, an aliquot of the solution was diluted 50 times with CH3CN – H2O (7: 3). The diluted solution was filtered and 10 mL was analysed with HPLC.
HPLC: The HPLC system used consisted of a pump, a column oven, a degasser and a fluorescence detector (1220 INFINITY II LC, Agilent). A CD-C18 column was used in a column oven at 40° C and the flow rate was constantly maintained at 1.0 mL/min. The excitation and emission wavelengths of the fluorescence detector were set to 450 and 560 nm, respectively. The mobile phases A, B and C used in the study were 0.1% TFA / CH3CN, 0.1% TFA / H2O, and 0.1% TFA / MeOH, respectively. The time schedules for the mobile phase were as follows: 0-25.00 min A% ¼ 60-70, B% ¼ 30-20 (linear gradient) and C% ¼ 10; 25.01-40.00 min A% ¼ 70, B% ¼ 20 and C% ¼ 10 (isocratic); 40.01-55.00 min A% ¼ 70-77, B% ¼ 20-13 (linear gradient) and C% ¼ 10; 55.01-65.00 min A% ¼ 77, B% ¼ 13 and C% ¼ 10 (isocratic) and 65.01-84.50 min A% ¼ 60, B% ¼ 30 and C% ¼ 10 (initialization). The resulting chromatogram was analysed using OpenLab software (Agilent). Calibration curves for ALA, PLA, AA, LA and OA were constructed in concentrations of 1-50, 1-200, 10-200, 25-800 and 25-800 mM, respectively. The peak area ratio of the Internal Standard was then plotted on each concentration (n ¼ 4).
Discussion
For the research, 33 subjects were recruited, 13 males between the ages of 26 and 77, and 20 females between the ages of 25 and 69. Each of them reported symptoms largely similar in many respects: low mood, abdominal swelling, fatigue, sleep disorders, anxiety, weight gain.
To highlight the link between HPA and dysbiosis (Farzi A. et al., 2018; Sonali S. et al., 2022) and between the latter and inflammation, the levels of indoxyl sulfate and skatole were evaluated and indicated urinary (skatole as a measure of putrefactive dysbiosis and indoxyl sulfate as a measure of fermentative dysbiosis), the possible presence of candida or fungi in the faeces, together with the plasma serotonin levels, and the ratio Omega-3/Omega-6 still evaluated in plasma.
Cortisolemia levels were also measured, but not in all subjects, assessed in the four physiologically significant moments, namely at 7 am and at 1 pm, 5 pm, and 11 pm.
Figure 10 shows the graph of the correlations between the main numerical variables considered in this research: the larger the colored circle, the stronger the correlation (the black color indicates a strong positive correlation, while the white ones a negative type). The same correlations are shown in the right table but expressed in numerical terms. To this end, it should be remembered that the calculated linear correlation can assume values between -1 and 1.
A correlation, in absolute value, equal to 1 indicates a strong linear dependence, while values close to zero reveal the absence of a linear link between the variables considered.
From the analysis of the above values it can be seen that a positive correlation is highlighted between the levels of serotonin and those of EPA and DHA, while a negative one emerges between these last two parameters and the number of fungi found.
Another interesting aspect that I want to emphasize is how the two fatty acids show a strong negative correlation (EPA to an even greater extent than DHA) with the age of the subjects considered in the study.
Finally, serotonin is negatively correlated with the number of fungi and with Cortisol-4 (measured at 11 pm). Cortisol-1 (measured at 7), on the other hand, shows a modest inversely proportional link with age, while Cortisol-4 (measured at 11pm) exhibits an equally modest but directly proportional link.
In order to identify the behavior of the numerical variables considered also with the variation of some observed parameters/variables, the following boxplots were then analyzed.
In Figure 11.1/11.2/11.3 the levels of serotonin, skatole and indoxyl sulfate are analyzed in relation to the presence or absence of Candida a.; in the first case (Figure 11.1) it is evident that the plasma levels of serotonin are on average higher in the 3rd and 4th quartile than in patients who do not present Candida a.. In relation to skatole (Figure 11.2) this evidence is not recorded since its levels appear substantially identical between the group of patients with Candida and those without.
We therefore wanted to verify how Candida a. can affect the levels of indoxyl sulfate, the result of our analysis is shown in the boxplot represented in Figure 11.3. In this case there is an opposite situation with respect to that shown in Figure 11.1 regarding the effects on serotonin levels: the values of indoxyl sulfate expressed both in the 2nd and in the 3rd and 4th quartiles, are clearly higher in the case in which it is excluding the presence of Candida a..
We therefore wanted to investigate the relationship between Candida a. and cortisolemia, evaluating the ratios in the four peaks that are physiologically expressed in our organism at four distinct times of the day: 7, 13, 17, 23; here they are shown in the following boxplots in Figure 12.1/12.2/12.3/12.4. In all four comparisons it is evident that in the presence of Candida a. higher cortisol levels are on average associated with all four measurements. In this sense, it is interesting to remember how high levels of cortisolemia correspond to a condition of metabolic acidosis which is one of the conditions favoring fungal proliferation in the intestine. We then wanted to analyze the data obtained through the relationships between the values relating to the levels of EPA and DHA in the presence or absence of Candida a., The results are summarized in Figure 13.1/13.2, in this case the data show exactly opposite indications: while the presence of Candida a. it is accompanied by a decrease in the concentration of EPA, in the case of DHA the opposite trend is observed.
Another aspect to be carefully evaluated is that in the relationship that emerged between the levels of Candida a. and those of the mycetes measured in the subjects included in the study (Figure 14.1). From the boxplot it is clearly observed in all four quartiles as in the presence of Candida a. higher levels of mycetes are associated with respect to subjects in which the fungus was absent. It is interesting to note that from the analysis of the data obtained it seems that high levels of fungi are mainly of female relevance (Figure 14.2); as well as the lower levels of EPA and DHA seem, once again, prevalent typical of the women included in the study (Figure 14.3/14.4).
Subsequently, we wanted to describe the relationship between the levels of serotonin with respect to the values of skatole, fungi, EPA and DHA (Figure 15.1/15.2/15.3/15.4) using as a discriminant of our analysis, the presence of low or high levels of the same serotonin (with a new reference interval).
From these data emerges strongly if on the one hand the skatole levels do not seem to influence those of serotonin in any way (Figure 15.1), on the other we can observe how the low levels of serotonin (Figure 15.2) are preferably associated with high values of mycetes (difference detectable in all quartiles represented in the two boxplots). Finally, low levels of EPA (Figure 15.3) are connected to low levels of serotonin, a bond that we do not observe with DHA (Figure 15.4) where in the 2nd and 3rd quartiles low values of the second correspond to high levels of the first.
We wanted to deepen our evaluation of the results by dividing the serotonin values into low, normal, high (following the old reference range) compared to the values of mycetes (Figure 16.1) skatole (Figure 16.2), cortisolemia -measured at 7, 13, 17, 23- (Figure 17.1/17.2/17.3/17.4), EPA and DHA (Figure 18.1/18.2). The evidences that emerged (Figure 16.1) showed the scarce dependence between the serotonin values with respect to the influence of skatole highlight a dependence which in fact manifests itself weakly in case of low levels of the first compared to those of the second.
It is very interesting, however, to observe the link between serotonin and the levels of mycetes in Figure 16.2 which shows how low concentrations of the neurotransmitter are accompanied by high levels of the mycetes themselves.
We wanted to verify any correlations between the cortisol values, still measured in the four different and physiologically significant intervals of the day, and the serotonin values indicated in the reference intervals (Figure 17.1/17.2/17.3/17.4): we can see how there is an evident relationship between low serotonin levels and high concentrations of Cortisol-1/2/4.
Finally we evaluated the relationship between the values of EPA and DHA compared to those of serotonin and in this case we can observe an interesting fact: high levels of EPA and DHA are accompanied by normal concentrations of the neurotransmitter (Figure 18.1/18.2), in the case of the first fatty acid this is highlighted above all by observing the 2nd-3rd and 4th quartiles, while in the second this applies to all in and four quartiles represented in the boxplot.
In relation to what was previously illustrated and discussed about the sample examined, it is now desired to carry out a multivariate analysis that investigates, using a linear regression model, which are the main determinants that induce a variation in the levels of serotonin in the subjects taken into consideration (Figure 19). These variables are some of those discussed above. Their inclusion in the statistical model is, first of all, conditioned by the levels of correlation with serotonin - the variables with the highest correlation or the mutable ones are selected that induce classes of distinct values according to the relative modalities - and, at the same time, to avoiding errors in interpreting the model, independent variables that are highly correlated with each other are not entered jointly. Following the construction of multiple models and robustness analyzes, it was decided to present the model shown in Figure 19 formally expressed below:
where serotonin represents the dependent variable while all the others are defined as independent variables. The βi coefficients represent the effect of the i-th independent variable in the determination of serotonin. The greater the coefficient in absolute value, the stronger the link between serotonin and the i-th independent variable. The sign of the coefficient identifies directly proportional (if positive) or inversely proportional (if negative) bonds.
serotonin = β0 + β1 males + β2 cortisol1 + β3 cortisol2 + β4 cortisol3 + β5 cortisol4 + β6 indoxyl sulfate severe + β7 indoxyl sulfate medium + β6 indoxyl sulfate minor
Finally, remember that the categorical (or mutable statistical) variables -sex, candida/mycetes, indoxyl sulfate- should be read marginally with respect to the value that does not appear, as it is captured by the intercept of the model (that is the constant β0). Therefore, from the study of what is reported in Figure 19, the following emerges:
- -
- men have a higher level of serotonin than women examined;
- -
- in general, cortisol 1 and 3 have a positive and statistically significant effect on serotonin, while the increase in cortisol 4 there is a statistically significant reduction in serotonin values;
- -
- in the presence of Candida a. and/or mycetes there is a lower level of serotonin than in subjects who do not have Candida a. and/or mycetes;
- -
- all three levels of indoxyl sulfate reported contribute to the determination of changes in serotonin in an inversely proportional way, compared to subjects with eubiosis. These relations, statistically significant, have different magnitudes and in particular, the strongest effect is associated with the average indicate levels. Patients in our sample who have an average indoxyl sulfate level (according to reference to physiological levels) also have a much lower level of serotonin. The empirical evidence goes in the same direction for the other two levels (mild and severe), however with a lower magnitude.
Conclusions
An impressive number of elements, referring to both experiments and clinical data, indicate that the microbiota plays a leading role within the GBA and that this role does not only imply its interaction with the ENS, but rather an exchange of information with the CNS through neuroendocrine and metabolic pathways (Carabotti M. et al., 2015). In particular in humans, the most stimulating indication of the existence of an intestinal interaction between the microbiota and the brain emerged about twenty years ago after the improvements found in patients with hepatic encephalopathy, after administration of antibiotics (Morgan M.Y., 1991); at the same time, the link between microbiota and anxiety-depressive syndromes emerged (Naseribafrouei A. et al., 2014; Foster J.A. et al., 2013).
This study aimed to investigate the links between various parameters already associated with inflammatory diseases (such as the relationship between AA/EPA and AA/DHA) and others directly related to the stress axis (such as cortisol levels) with respect to intestinal dysbiosis (expressed through the indoxyl sulfate and skatole levels and the presence of Candida a. and/or mycetes) compared to the serotonin levels considered in relation to CFS and depression.
From the data collected, and summarized in Fig.-15, we can make a series of observations.
The research shows that of the 33 subjects included in the study, 88% have low levels of serotonin which are related to mood and chronic fatigue; of these 33, 82% complained of morning fatigue, while in 79% of cases, indoxyl sulfate levels characteristic of a state of dysbiosis were measured, with 39% of subjects reporting swollen abdomen and in 52% alterations of the bowel habits. Candida a. and/or mycetes were isolated in the faeces of 67% of the sample under investigation. A side note regarding muscle contraction which was reported by 39%, a typical manifestation of metabolic acidosis.
Although the results obtained are very interesting, and from many points of view encouraging specially for what emerged from the data analysis about the link between cortisol and serotonin levels (Fig.-12f: -13a/b/c) which highlight how there is a statistically significant link between these parameters, it is however necessary to collect a higher number of patients to obtain clinically meaningful indications. Even if the data shows the link between the levels of serotonin and the values of indoxyl sulfate with respect to the presence of Candida a. o mycetes as well as the ratio between omega-3 and omega-6 we need to accumulate more data; in particular, the first three parameters seem to present a relationship as a function of the CFS or depression.
Statistics
The choice of tools for the statistical data analysis relative to the illustrated study, has been made difficult by different factors such as the lack of a control group and the limited number of cases studied; despite the difficulties, we tried to find a statistical significance in the data at our disposal using the following methodological approach.
The analysis of the collected data was conducted using graphical tools (boxplot also conditional on the different modalities of the mutable statistics available) and dependency measures (correlation between all the numerical variables collected). In particular, it was decided to build a correlation chart to have a quick view of the possible presence of a linear link between the observed variables.
The numerical variables for which physiological levels are also available were used for the construction of further boxplots in order to highlight any stylized facts otherwise not identifiable with the sole study of the correlations.
Furthermore, the preliminary descriptive analysis allows to highlight some distinctive features of the sample examined and to guide the consequent inferential analysis. Specifically, it was decided to conduct a more in-depth study based on the construction of a linear regression model in order to highlight, where possible, the main determinants of serotonin. Particular attention was paid to the choice of variables in order to avoid violations of the assumptions of the model and, at the same time, problems of mispecification. In particular, this last aspect was taken care of avoiding the joint use of highly correlated variables in order to be able to interpret the marginal effect of each determinant on serotonin well. It should be remembered that the analysis conducted was carried out only on patients without missing values in the variables considered. Furthermore, this is a preliminary study, whose empirical evidence, currently based on a limited number of observations, suggests interesting results to be explored in future research, enriching the sample of patients examined.
The estimates made using the linear regression model were subjected to statistical tests to evaluate their significance. Specifically, to test the null hypothesis under which it is assumed that the estimated parameter βi is equal to zero (absence of an effect of the dependent variable with respect to the dependent) against the alternative that is non-zero, a t-test was conducted. The latter shows that most of the independent variables are statistically significant with p-value <0.01.
References
- American Psychiatric Association (1980) DSM-III:. Manuale diagnostico e statistico dei disturbi mentali. 3ª ed. Masson, Milano 1983.
- Aardal, E.; Holm A.. Cortisol in saliva. Eur J Clin Biochem 1995; 33; 927-932 .
- Abrams, G.D.; Bishop, J.E. Effect of the normal microbial ora on gastrointestinal motility. Proc Soc Exp Biol Med 1967;126:301-304.
- Androulakis, P.; Ioannis. Circadian rhythms and the HPA axis: a systems view. WIREs Mech Dis. 2021 July; 13(4): e1518.
- Bach, J-F. The effect of infections on susceptibility to autoimmune and allergic diseases. N Engl J Med 2002; 347: 911–920.
- Bhattarai, Y.; Pedrogo, D.A.; Kashyap, P.C. Irritable bowel syndrome: a gut microbiota-related disorder? Am J Physiol Gastrointest Liver Physiol, 2017 Jan 1;312(1): G52-G62.
- Bailey, M.T.; Coe, C.L. Maternal separation disrupts the integrity of the intestinal microflora in infant rhesus monkeys. Dev Psychobiol 1999, 35(2):146–155. .
- Bailey, M.T.; Dowd, S.E.; Parry ,N.M.; Galley, J.D.; Schauer, D.B.; Lyte, M. Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium. Infect Immun 2010, 78(4):1509–1519. [CrossRef]
- Ballanger, J.C.; et al. Consensus statement on depression, anxiety, and functional gastrointestinal disorders. J.Clini.Psychiatry 2001; 62 (supp.8).
- Balsari, A.; Ceccarelli, A.; Dubini, F.; Fesce, E.; Poli, G. The fecal microbial population in the irritable bowel syndrome. Microbiologica. 1982 Jul;5(3):185-94.
- Barbara, G.; Stanghellini, V.; Brandi, G.; et al. Interactions between commensal bacteria and gut sensorimotor function in health and disease. Am J Gastroenterol 2005;100:2560-2568. [CrossRef]
- Barouei, J.; Moussavi, M.; Hodgson, D.M. Effect of maternal probiotic intervention on HPA axis, immunity and gut microbiota in a rat model of irritable bowel syndrome. PLoS One 2012, 7(10):e46051. [CrossRef]
- Basso, A.; Costa Pinto, F.A.; Russo, M.; Giorgetti Britto, L.R.; de Sa-Rocha, L.C.; Palermo Neto, J. Neural correlates of IgE-mediated food allergy. J. Neuroimmunol 2003;140:69–77. [CrossRef]
- Benard, A.; Desreumeaux, P.; Huglo, D.; Hoorelbeke, A.; Tonnel, A.B.; Wallaert, B. Increased Intestinal Permeability in Bronchial Asthma. The Journal of Clinical Immunology, 1996, vol. 97-num 6, pag. 1173-1178. [CrossRef]
- Beier, W. Biophysic. 2 Aufl., Lipsia Thieme, 1962.
- Beier W. Einfuhrung in die theoretische biophysic. Stoccarda, Gustav Fisher, 1965.
- Bengmark S. Ecological control of the gastrointestinal tract. The role of probiotic flora. Gut 1998;42:2-7. [CrossRef]
- Berrill, J.W.; Gallacher, J.; Hood, K.; et al. An observational study of cognitive function in patients with irritable bowel syndrome and in ammatory bowel disease. Neurogastroenterol Motil 2013;25:918-e704.
- Bertalanffy von, L. General System Theory: Foundations, Development, Applications. 1967, G.Braziller Inc., USA.
- Bharwani, A.; et al. Oral treatment with Lactobacillus rhamnosus attenuates behavioural deficits and immune changes in chronic social stress. BMC medicine (2017). [CrossRef]
- Bonaz, B.; Bazin, T.; Pellissier, TS. The vagus nervea t the interface of the Microbiota-Gut-Brain Axis. Front.Neurosci., 2018 Feb;12:49.
- Bortolato, B.; Hyphantis,, T.N.; Valpione, S.; Perini,, G.; Maes, M.; Morris, G. Depression in cancer: The many biobehavioral pathways driving tumor progression. Cancer Treat Rev. 2017, 52, 58–70. [CrossRef]
- Bottaccioli F.; Carosella A. Mente inquieta. Ed. Tecniche Nuove, 2011.
- Bottaccioli F.; Carosella A. La saggezza del secondo cervello. Ed. Tecniche Nuove, 2007.
- Bottaccioli F.; Carosella A. Mente inquieta. Ed. Tecniche Nuove, 2011. .
- Bottaccioli F. PNEI. Ed. RED (1995).
- Bryant, R.A.; Harvey, A.G. Posttraumatic stress in volunteer fire-fighters. Predictors of distress. Journal of Nervous and Mental Disease, 1995, 183, 267-271. . [CrossRef]
- Brekhman, I.I. Man and Biologically Active Substances. Pergamon Press, Kronberg/Taunus, 1980. [CrossRef]
- Breuer, J.; Freud, S. Studies on hysteria: physical mechanism of hysteria. Tralated by Strachey J., New York, Penguin, 1973.
- Brodal, A. Neurological anatomy in relation to clinical medicine. Oxford University Press, V ed., Oxford (1992).
- Buffie, C.G.; Jarchum, I.; Equinda M.; Lipuma L.; et al. Profound Alterations of Intestinal Microbiota following a Single Dose of Clindamycin Results in Sustained Susceptibility to Clostridium difficile-Induced Colitis. Infection and Immunity, 2011, pag. 62-73. [CrossRef]
- Burton, A.C. The proterties of the steady state compared to those of equilibrium as shown in charateristic biological behavior. J. Cell. Comp. Physiol., 14 (1939), pp327-349.
- Cajochen, C.; Zeiter, J.M.; Czeisler, C.A; Dijk, D.J. Dose-response relationship for light intensity and ocular and electroencephaligraphic correlates of human alertness. Behav Brain Res., 2000, 115, 75-83. [PubMed]
- Caenepeel, P.; Janssens, J.; Vantrappen, G.; Eyssen, H.; Coremans, G. Interdigestive myoelectric complex in germ-free rats. Dig Dis Sci 1989;34:1180-1184. . [CrossRef]
- Caldarini, M.I.; Pons, S.; D’Agostino, D.; DePaula, J.A.; Greco, G.; Negri, G.; Ascione, A.; Bustos, D. Abnormal fecal flora in a patient with short bowel syndrome: An in vitro study on effect of pH on D-lactic acid production, Dig Dis Sci 41: 1649-1652, 1996. .
- Cannon, W.B. The wisdom of the body. Ed. Norton (1932), New York.
- Cannon, W.B. Organizationfor physiological homeostasis. Phys Rev, 9 (1929), 2nd edition, ed. Norton, New York.
- Cannon, W.B. Bodily changes in pain, hunger, fear and range. Ed. D. Applenton, 1932.
- Caparros, E.; Wiest, R.; Scharl, M.; Roglen, G.; et al. Dysbiotic microbiota interactions in Crohn’s disease. Gut Microbes, 2021, Vol.13, N.1. [CrossRef]
- Capra, F. La rete della vita. Ed. BUR, 2001.
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: interctions between enteric microbiota, central and enteric nervous systems. Annals of Gastroenterology 2015; 28, 203-209.
- Cassano, G.B.; D’Errico, A.; Pancheri, P.; Pavan, L.; Pazzagli, A.; Rossi, R.; Smeraldi, E.; Volterra, V. Trattato Italiano di Psichiatria. Ed. Masson, Milano, 1993, 1560-1585.
- Castex, N.; Fioramonti, J.; Fargeas, M.J.; Bueno, L. c-fos expression in specific rat brain nuclei after intestinal anaphylaxis:involvement of 5-HT3 receptors and vagal afferent fibers. Brain Res 1995;688:149–160. [CrossRef]
- Chadwick, V.S.; et al.. Activation of the mucosal immune system in irritable bowel syndrome. Gastroenterol 2002;120:1778–1783. [CrossRef]
- Clarke, G., Stillin R.M. et al.. Minireview: gut microbiota, the negletted endocrine organ. Mol Endocrinol, 2014 Aug;28(8): 1221-1238.
- Clarke, G.; Grenham, S.; Scully, P.; et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry 2013;18:666-673.
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat Rev Microbiol. 2012;10:735–42. [CrossRef]
- Colombo, P.P.; Mantua, V. Post-Traumatic Stress Disorder in ordinary life. Rivista di psichiatria, 2001, 36, 2.
- Craig, A.D. Interoception: the sense of the physiological condition of the body. Curr. Opin. Neurobiol 2003;13:500–505. [CrossRef]
- Crouzet, L.; Gaultier, E.; Del’Homme, C.; et al. The hypersensitivity to colonic distension of IBS patients can be transferred to rats through their fecal microbiota. Neurogastroenterol Motil 2013;25:e272-e282. [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: the impact of the gut microbiota on brain and behavior. Nat Rev Neurosci. 2012;13:701–12.
- Cummings, J.H.; Macfarlane, G.T. Role of intestinal bacteria in nutrient metabolism. JPEN J Parenter Enteral Nutr 1997;21:357- 365. [CrossRef]
- van Deventer, S.J.; Ten Cate, J.W.; Tytgat G.N. Intestinal endotoxemia. Clinical significance. Gastroenterology 1988;94:825-831.
- Devkota, S.; et al. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− mice. Nature 2012; 487:104–108. [CrossRef]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; et al. Normal gut microbiota modulates brain development and Behavior. Proc Natl Acad Sci USA 2011;108:3047-3052. [CrossRef]
- Duncan, S.H.; et al. Reduced dietary intake of carbohydrates by obese subjects results in decreased concentrations of butyrate and butyrate-producing bacteria in feces. Appl Environ Microbiol 2007; 73:1073–1078. [CrossRef]
- Dupont, H.L. Review article: evidence for the role of gut microbiota in irritable bowel syndrome and its potential in uence on therapeutic targets. Aliment Pharmacol Ther 2014;39:1033-1042.
- Eckburg ,P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; et al. Diversity of the human intestinal microbial flora Science 308:1635–1638; 2005.
- Eder, W.; Ege, M.J.; von Mutius, E. The asthma epidemic. N Engl J Med 2006; 355: 2226–2235.
- Erdrich, S.; Hawrelak, J.A.; Myers, P.; Harnett, J. Determining the association between fibromyalgia, the gut microbiome and its biomarkers: a systematic review. BMC Musculo Dis, (2020) 21:181. [CrossRef]
- Evans, J.M.; Morris, L.S.; Marchesi, J.R. The gut microbiome: the role of a virtual organ in the endocrinology of the host. J Endocrinol. 2013;218:R37–R47. [CrossRef]
- Farley ,M.; Barkan, H. Prostitution, violence, and posttraumatic stress disorder. Women Health 1998, 27, 37-49.
- Farzi, A.; Frohlich, E.E.; Holzer, P. Gut microbiota and the neuroendocrine system. Neurotherapeutics 2018 15:5-22. [CrossRef]
- Femstrom, J.D.; Wurtman, R.J. Brain serotonin content: increase following ingestion of carbohydrate diet. Science 1971, dec.3; 174(4013): 1023-5.
- Foster R.; Kreitzman, L. I ritmi della vita. Ed. Longanesi, 2007.
- Foster, J.A.; McVey-Neufeld, K.A. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci 2013;36:305-312. . [CrossRef]
- Freitas, R.D.S.; Campos, M.M. Protective effects of Omega-3 fatty acids in cancer-related complications. Nutrients 2019, 11, 945. [CrossRef]
- Furness, J.B.; Costa, M. The enteric nervous system. 1987, Churchill Livingstone, Edinburgh.
- Gabella, G. Structure of autonomic nervous system. 1976, Chapman and Hall, London. [CrossRef]
- Galley, J.D.; Nelson, M.C.; Yu, Z.; et al. Exposure to a social stressor disrupts the community structure of the colonic mucosa-associated microbiota. BMC Microbiol 2014;14:189. [CrossRef]
- Gershon, M.D. The second brain: a groundbreaking new understanding of nervous disorders of the stomach and intestine. 1998, Ed. Harper.
- Gilbert, S.F. Biologia dello sviluppo. IV ed. italiana 2012, ed. Zanichelli.
- Goehler, L.E.; et al. Infection-induced viscerosensory signals from the gut enhance anxiety: implications for psychoneuroimmunology. Brain Behav Immun 2007 August ; 21(6): 721–726. [CrossRef]
- Goehler, L.E; et al. Activation in vagal afferents and central autonomic pathways: early responses to intestinal infection with Campylobacter jejuni. Brain Behav Immun 2005;19:334–344. [CrossRef]
- Goehler, L.E.; Gaykema, R.P.A.; Anderson, K.; Hansen, M.K.; Maier S.F.; Watkins L.R. Vagal immune-to-brain communication: a visceral chemoreceptive pathway. Auton Neurosci 2000;85:49–59.
- Gomborone, J.E.; Gorard, D.A.; Dewsnap, P.A.; Libby, G.W.; Farthing M.J. Prevalence of irritable bowel syndrome in chronic fatigue. J R Coll Physicians Lond 30: 512-513, 1996. .
- Grider, J.R.; Piland, B.E. The peristaltic re ex induced by short-chain fatty acids is mediated by sequential release of 5-HT and neuronal CGRP but not BDNF. Am J Physiol Gastrointest Liver Physiol 2007;292:G429-G437. .
- Grundy D. Neuroanatomy of visceral nociception: vagal and splanchnic afferent. Gut 2002;51(Suppl I):i2–i5. [CrossRef]
- Hijazi, Z.; Molla, A.M.; Al-Habashi, H.; Muawad, W.M.R.A.; Sharma, P.N. Intestinal Permeability is Increased in Bronchial Asthma. Arch Dis Child 2004; 89:227-229. [CrossRef]
- Hawrelak, J.A.; Myers, S.P. The causes of intestinal dysbiosis: a review. Altern Med Rev 2004;9:180–197.
- Holzapfel, W.H.; Haberer, P.; Snel, J.; et al. Overview of gut flora and probiotics. Int J Food Microbiol 1998;41:85-101. [CrossRef]
- Hong-Yan, Qin; Chung-Wha, Chenh; Xu-Dong, Tang; Zhao-Xiang, Bian. Impact of psycological stress on irritable bowel syndrome. WJG, Oct 21 (2014), Vol.20, Issue 39.
- Hong-Xing, Wang; Yu-Ping, Wang. Gut-microbiota-brain axis. Chinese Medical Journal, October 2016, Vol. 129, Issue 19.
- Hubbard, T.D.; Murray, I.A.; Perdew, G. Indole and tryptophan metabolism: endogenuos and dietary routes to Ah receptors activation. Drug Metab Dispos, oct 2015, 43:1522-1535.
- Hudson, J.I.; Goldenberg, D.L.; Pope, H.G. Jr; Keck, P.E. Jr; Schlesinger, L. Comorbidity of fibromyalgia with medical and psychiatric disorders. Am J Med 92: 363-367, 1992. .
- Husebye, E.; Hellström, P.M.; Sundler, F.; Chen, J.; Midtvedt, T. Influence of microbial species on small intestinal myoelectric activity and transit in germ-free rats. Am J Physiol Gastrointest Liver Physiol 2001;280:G368-G380.
- Iwai, H.; Ishihara, Y.; Yamanaka, J.; Ito, T. Effects of bacterial ora on cecal size and transit rate of intestinal contents in mice. Jpn J Exp Med 1973;43:297-305. .
- James, W. Principles of psychology. 1890, ed. eBooks@Adelaide.
- James, W. Che cos’è un’emozione, In James, W. L’uomo come esperienza. Identità, istinti, emozioni. 1999, ed. L’Ancora, Ancona.
- James, W. Esiste la coscienza. In James W. L’uomo come esperienza. Identità, istinti, emozioni. 1999, ed. L’Ancora, Ancona.
- Janet, P. The mental state of histericals: a study of mental stigmata and mental accidents. Washington DC: University Publications of America (1893, 1901, 1977).
- Jurjus, A.; et al. Inflammatory bowel disease, colorectal cancer and type 2 diabetes mellitus: the links. BBA Clinical (2016), 16-24. [CrossRef]
- Kamiya, T.; et al. Inhibitory effects of Lactobacillus reuteri on visceral pain induced by colorectal distension in Sprague-Dawley rats. Gut 2006;55:191–196. doi: 10.1136/gut.2005.070987 . [CrossRef]
- Kamphuis, J.H.; Emmelkamp, P.M. Crime-related trauma: psychological distress in victims of bankrobbery. Journal Anxiety Disorder, 1998, 12, 199-208. [CrossRef]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Irritable bowel syndrome: A microbiome-gut-brain axis disorder. World J Gastroenterol 2014;20:14105-14125. [CrossRef]
- Kimura, I.; Inoue, D.; Maeda, T.; et al. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc Natl Acad Sci USA 2011;108:8030-8035. . [CrossRef]
- Kinashi, Y.; Hase, K. Partners in Leaky gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Frontiers in Immunology, Apr 2021;12:673708. [CrossRef]
- King, D.E.; Mainous, A.G.; Lambourne, C.A. Trends in dietary fiber intake in the United States”, 1999-2008. J Acad Nutr Diet 2012; 112: 642–648.
- Kirschbaum, C.; Hellhammer, D. Salivary cortisol in psychoneuroendocrine resarch: recent developments and applications. Psychoneuroendocrinology, Vol. 19; 313-333, 1994. .
- Koboziev, I.; Reinoso-Webb, C.; Grisham, M.B. Role of the enteric microbiota in intestinal homeostasis and inflammation. Free Radic Biol Mol Med, Mar 2014, 68:122-33. [CrossRef]
- Konturek, P.C.; et al. Stress And The Gut: Pathophysiology, Clinical Consequences, Diagnostic Approach And Treatment Options. Journalnal of Physiology and Pharmacology 2011, 62, 6, 591-599.
- Lacosta, S.; Merali, Z.; Anisman, H. Behavioral and neurochemical consequences of lipopolysaccharide in mice: angiogenic-like effects. Brain Res 1999;818:291–303.
- Langley, J.N. The autonomic nervous system. Part I, 1921, Heffer, Cambridge. .
- Larsen, P.R. William’s textbook of endocrinology. 2003, ed. Saunders, Philadelphia. [CrossRef]
- David, L.A.; Maurice C.F.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature, 2014 January 23; 505(7484): 559–563. [CrossRef]
- Lazarev, N. 7° All-union Congr.Physiol.Biochem., Moscow, p.579-1947.
- Leigh-Gibson, E.; et al. Increased salivary cortisol reliably induced by a protein-rich meal. Psychosom Med. 1999 Mar-Apr;61(2):214-24). [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: human gut microbes associated with obesity. Nature 2006; 444:1022–1023. [CrossRef]
- Lindheim, L.; Bashir, M.; Munzker, J.; Trummer, C.; Zachhuber, V.; Leber, B.; et al. Alterations in Gut Microbiome Composition and Barrier Function Are Associated with Reproductive and Metabolic Defects in Women with Polycystic Ovary Syndrome (PCOS): A Pilot Study. PLoS ONE 2017, 12(1): e0168390. [CrossRef]
- Liu C.Y. et al. Vagal modulation of intestinal afferent sensitivity to systemic lipopolysaccharide in the rat. Am J Physiol Gastrointest Liver Physiol 2007 May;292(5):G1213-20. Epub 2007 Jan 4.
- Liu, J.J.; Galfavy, H.C.; Cooper, T.B.; Oquendo, M.A.; Grunebaum, M.F.; Mann, J.J.; Sublette, M.E. Omega-3 Polyunsaturated Fatty Acid Status in Major Depression with Comorbid Anxiety Disorders. J Clin Psychiatry 2013 July; 74(7): 732-738. [CrossRef]
- Locker, A.; et al. Elemente einer systemtheoretischen betrachtung des stoffwechsels. Helgolander Wissenschaftliche Meeruntersuchungen, 14 (1966a), pp. 4-24. [CrossRef]
- Luna, R.A.; Savidge, T.C.; Williams, K.C. The Brain-Gut- Microbiome Axis: What Role Does It Play in Autism Spectrum Disorder? Curr Dev Disord Rep 2016;3:75-81.
- Lydiard, R.B. Social Anxiety Disorder: Comorbidity and Its Implications. J. Of Clin. Psych. 2001; 62 (supp 1): 17-24.
- Lydiard R.B. Irritable bowel syndrome, anxiety, and depression: what are the links? J. Clin. Psychiatry 2001;62:38–45.
- Macfarlane, G.T.; Macfarlane, S. Human colonic microbiota: ecology, physiology and metabolic potential of intestinal bacteria. Scand J Gastroenterol Suppl 1997;222:3-9.
- Macfarlane, S.; Macfarlane, G.T. Proteolysis and amino acid fermentation. In: Gibson GR, Macfarlane GT, eds.: Human Colonic Bacteria: Role in Nutrition, Physiology, and Pathology. Boca Raton, FL: CRC Press; 1995:75-100.
- Macfarlane, G.T.; Blackett ,K.L.; Nakayama, T.; Steed, H.; Macfarlane, S. The gut microbiota in inflammatory bowel disease. Curr Pharm Design. 2009;15:1528–36. [CrossRef]
- McGregor, N.R.; Dunstan, R.H.; Zerbes, M.; Butt, H.L.; Roberts, T.K.; Klineberg, I.J. Preliminary determiantion of a molecular basis to chronic fatigue syndrome. Biochem Mol Med 57: 73-80, 1996. .
- Mackos, A.R.; Eubank, T.D.; Parry, N.M.; Bailey, M.T. Probiotic Lactobacillus reuteri attenuates the stressor-enhanced severity of Citrobacter rodentium infection. Infect Immun 2013, 81(9):3253–3263. . [CrossRef]
- Manichanh, C.; Rigottier-Gois, L.; Bonnaud, E.; et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. GUT; 2006;55:205–11. [CrossRef]
- Marazziti, D.; et al. New development on the serotonin hypothesis of depression: shunt of tryptophan. Riv. Psichiatr. 2013; 48(1): 23-34.
- Marmar, C.R.; Weiss, D.S.; Schlenger, W.E.; Fairbank, J.A.; Jordan, B.K.; Kulka, R.A.; Hough, R.L. Peritraumatic dissociation and post-traumatic stress in male Vietnam theater veterans. American Journal of Psychiatry 1994, 151, 902-907.
- Marrama, P.; Angeli, A. Manuale di endocrinologia. 1992, ed. Masson.
- Matthew A.; Odenwald and Jerrold R. Turner. Intestinal permeability defects: Is it time to treat? Clin Gastroenterol Hepatol, 2013 September ; 11(9): 1075–1083.
- Mayer, E. The mind-gut connection. Ed. Harper Wave (2016).
- Mayer, E.A.; Craske, M.; Naliboff, B.D. Depression, anxiety, and the gastrointestinal system. J. Clin. Psychiatry 2001;62:28–36.
- Mayer, E.A.; Padua, D.; Tillisch, K. Altered brain-gut axis in autism: comorbidity or causative mechanisms? Bioessays 2014;36:933-999. .
- Mengchao Jin, Zhiyuan Qian, et al. The role of intestinal microbiota in cardiovascular disease. J Cell Mol Med 2019;23:2343-2350.
- Metchnikoff, E. The Prolongation of Life: Optimistic Studies. London: William Heinemann; 1907:161-183. . [CrossRef]
- Miller-Fleming, L.; Olin-Sandoval, V.; et al. Remaining mysteries of molecular biology: the role of polyamines in the cell. J.Mol.Biol. 2015; 427:3389-3406.
- Morgan, M.Y. The treatment of chronic hepatic encephalopathy. Hepatogastroenterology 1991;38:377-387.
- Muegge, B.D.; et al. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science. 2011; 332:970–974. [CrossRef]
- Murray, M.; Pizzorno, J. Encyclopedia of Natural Medicine. Rocklin, CA: Prima Publishing; 1998:143.
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; et al. Correlation between the human fecal microbiota and depression. Neurogastroenterol Motil 2014;26:1155-1162. . [CrossRef]
- Nauta, W.J.H. The problem of the frontal lobe: A reinterpretation. J. Psychiatry Res 1971;8:167–187. [CrossRef]
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut microbiota and IBD: causation or correlation? Nat Rev Gastroenterol Hepatol, 2017 Oct;14(10): 573-584.
- Nord, C.E.; Heimdahl, A.; Kager, L. Antimicrobial agents and the human oropharyngeal and intestinal microflor. Ann Ist Super Sanita 1986;22:883-892.
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Reports 2006, 7 688–693. [CrossRef]
- Othman, M.; Aguero, R.; Lin, H.C. Alteration in intestinal microbial flora and human disease. Curr Opin Gastroenterol., 2008 Jan.; 24(1): 11-6. [CrossRef]
- Packard Amy, E.B.; Egan Ann, E.; et al. HPA axis- interaction with behavioral system. Compr. Physiol.; May 2020; 6(4): 1897-1934.
- Pettenkofer von, M. The roles of inflammation, nutrient availability and the commensal microbiota in enteric pathogen infection. Microbiology Spectrum, 2015, 3(3):MBP-008-2014.
- Pimentel, M.; Lembo, A; Chey, W.D.; et al. TARGET Study Group. Rifaximin therapy for patients with irritable bowel syndrome without constipation. N Engl J Med 2011;364:22-32. [CrossRef]
- Porges, S.W. The pocket guide to the polyvagal theory. The transformative power of feeling safe. Norton and Company Inc, 2017.
- Price, J.L. Prefrontal cortical networks related to visceral function and mood. Ann. NY. Acad. Sci 1999;877:383–396. [CrossRef]
- Puri, B.K. Long-chain polyunsatured fatty acids and the pathophysiology of myalgic encephalomyelitis (Chronic Fatigue Syndrome). J Clin Pathol 2007;60:122-124. [CrossRef]
- Rapozo, C.M.; Bernardazzi, C.; Siffert-Pereira de Souza, H. Diet and microbiota in inflammatory bowel disease: The gut in disharmony. WJG, 2017 March 28; 23(12): 2124-2140. .
- Rhee, S.H.; Pothoulakis, C.; Mayer, E.A. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat Rev Gastroenterol Hepatol 2009;6:306-314. [CrossRef]
- Rigottier-Gois, L. Dysbiosis in inflammatory bowel disease: the oxygen Hypothesis. International Society for Microbial Ecology, (2013) 7, 1256-1261. [CrossRef]
- Rodriguez, L.A.G.; Ruigomez, A. Increased risk of irritable bowel syndrome after bacterial gastroenteritis: cohort study. BMJ, volume 318, 27 february 1999. [CrossRef]
- Rooks, M.G.; Garret, W.S. Gut microbiota, metabolites and host immunity. Nat Rev Immunol 2016 May 27; 16(6): 341-352. .
- Rossi-George, A.; Urbach, D.; Colas, D.; Goldfarb, Y.; Kusnecov, A.W. Neuronal, endocrine and anorexic responses to the T-cell superantigen staphylococcal enterotoxin A: Dependence on tumor-necrosis factor-α. J. Neurosci 2005;25:5314–5322. [CrossRef]
- Rothbaum, B.O.; Foa, E.B. Subtypes of posttraumatic stress disorder and duration of symptoms, in Posttraumatic Stress Disorder: DSM IV and beyond. Edited by Davidson J.R.T., Foa E.B., Washington, DC, American Psychiatric Press, 1993, 23-35. .
- Ruigomez, A.; et al. One-year follow up of newly diagnosed IBS patients. Aliment Pharmacol Ther 1999; 13: 1097-1102.
- Schreiber, O.; Petersson, J.; Phillipson, M.; Perry, M.; Roos, S.; Holm, L. Lactobacillus reuteri prevents colitis by reducing P-selectin-associated leukocyte- and platelet-endothelial cell interactions”, Am J Physiol Gastrointest Liver Physiol 2009, 296(3):G534–G542.
- Selye, H. The stress of Life. McGraw-Hill (1976).
- Selye, H. Stress in health and disease. Butterworths, 1976. [CrossRef]
- Selye ,H. Endocronology. 21/2, 169-1937.
- Shah, M.; Beuerlein, M.; Danayan, K. An approach to the patient with a life-threatening acid-base disturbance: The acidemias. University of Toronto Medical Journal 78: 122-128, 2001.
- Sheedy, J.R.; Wettenhall, R.; et al. Increased D-lactiic acid intestinal bacteria in patients with chronic fatigue syndrome. In Vivo 2009, 23:621-628.
- Simrén, M.; Barbara, G.; Flint, H.J.; et al. Rome Foundation Committee. Intestinal microbiota in functional bowel disorders: a Rome foundation report. Gut 2013;62:159-176. [CrossRef]
- Smith, T. A modification of the method for determining the production of indol by bacteria. J Exp Med (1897) 2, 543–547. [CrossRef]
- Sonali, S; Ray, B.; et al. Mechanistic insights into the link between gut dysbiosis and major depression: an extensive review. Cells (2022), 11, 1362. [CrossRef]
- Song, Y.; Liu, C.; Finegold, S.M. Real-time PCR quantitation of clostridia in feces of autistic children. Appl Environ Microbiol 2004;70:6459-6465. [CrossRef]
- Spiller, R.C. Role of nerves in enteric infection. Gut 2002;51:759–762. [CrossRef]
- Spiller, R.; Lam, C. An Update on Post-infectious Irritable Bowel Syndrome: Role of Genetics, Immune Activation, Serotonin and Altered Microbiome. J Neurogastroenterol Motil 2012;18:258-268.
- Stanghellini, V. Relationship between upper gastrointestinal symptoms and lifestyle, psychosocial factors and comorbidity in the general population: results from the Domestic/International Gastroenterology Surveillance Study (DIGEST)”, Scand J Gastroenterol Suppl. 1999;231:29-37.
- Stanghellini, V. Three-month prevalence rates of gastrointestinal symptoms and the influence of demographic factors: results from the Domestic/International Gastroenterology Surveillance Study (DIGEST). Scand J Gastroenterol Suppl. 1999;231:20-8. [CrossRef]
- Stanghellini. V. International comparative study of the prevalence and impact of gastrointestinal symptoms: an introduction. Scand J Gastroenterol Suppl. 1999;231:1-2. [CrossRef]
- Stead, R.H.; et al. Vagal influences over mast cells. Auton. Neurosci., 2006 Apr 30; 125(1-2): 53-61. Epub 2006 Feb 24. [CrossRef]
- Stefanko, D.P.; Barrett, R.M.; Ly, A.R.; Reolon, G.K.; Wood, M.A. Modulation of long-term memory for object recognition via HDAC inhibition. Proc Natl Acad Sci USA 2009;106:9447-9452. [CrossRef]
- Stilling, R.M.; Dinan, T.G.; Cryan, J.F. Microbial genes, brain and behaviour - epigenetic regulation of the gut-brain axis. Genes Brain Behav 2014;13:69-86.
- Sudo, N.; Chida, Y.; Aiba, Y.; et al. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol 2004;558:263-275. . [CrossRef]
- Teradaira, R.; et al. Mental stress-induced changes in plasma serotonin, tryptophan, kynurenine concentration in healthy partecipants. International Congress Series 2007, 1304:175-179. [CrossRef]
- Turnbaugh, P.J.; et al. The effect of diet on the human gut microbiome: a metagenomic analysis in humanized gnotobiotic mice. Sci Transl Med. 2009; 1:6ra14. [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nature Reviews Immunology, nov. 2009, Vol.9: 799-899. [CrossRef]
- Uribe, A.; Alam, M.; Johansson, O.; Midtvedt, T.; Teodorsson, E. Microflora modulates endocrine cells in the gastrointestinal mucosa of the rat. Gastroenterology 1994;107: 1259-1269. [CrossRef]
- Uribarri, J.; Oh, M.S.; Carroll, H.J. D-lactic acidosis. A review of clinical presentation, biochemical features, and pathophysiologic mechanisms. Medicine (Baltimore) 77: 73-82, 1998.
- Van der Kolk, B.A.; McFarlane, A.C.; Weisaeth, L. Traumatic stressthe effect of overwhelming experience on mind, body and society. New York NY: Guilford Press, 1996.
- Van der Kolk, B.A.; Greenberg, M.S.; Pitman R.K. Endogenous opioids stress induced analgesia, and post-traumatic stress disorder. Psychopharmacology Bullettin (1989), 25, 417-421.
- Van der Kolk, B.A.; Fisler, R. Dissociation and the fragmentary nature of traumatic memories: overview and exploratory study. Journal of Traumatic Stress (1995), 8, 505-525. [CrossRef]
- Vecsey, C.G.; Hawk, J.D.; Lattal, K.M.; et al. Histone deacetylase inhibitors enhance memory and synaptic plasticity via CREB: CBP-dependent transcriptional activation. J Neurosci 2007;27:6128-6140. [CrossRef]
- Walderhaug; E.; et al. Interactive effects of sex and 5-HTTLPR on mood and impulsivity during tryptophan depletion in healthy people. Biological Psychiatry 2007; 62:593-599. [CrossRef]
- Walker, A.W.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011; 5:220–230. [CrossRef]
- Weiss, G.A.; Hennet, T. Mechanism and consequences of intestinal dysbiosis. Cellular and Molecular Life Sciences, 74(16):2959-2977. [CrossRef]
- Wiener, N. Cybernetics or Control and Communication in the Animal and the Machine. 1948.
- Williams, R.M.; Berthoud, H.R.; Stead, R.H. Vagal afferent nerve fibres contact mast cells in rat small intestinal mucosa. Neuroimmunomodulation 1997;5:266–270. [CrossRef]
- Wostmann, E.; Bruckner-Kardoss, E. Development of cecal distention in germ-free baby rats. Am J Physiol 1959;197:1345-1346.
- Wu, G.D.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science. 2011; 334:105–108. [CrossRef]
- Wurtman, R.J.; et al. Effects of normal meals rich in carbohydrates or proteins on plasma tryptophan and tyrosine ratios. American Journal of Clinical Nutrition 2003, 77: 128-132. [CrossRef]
- Xiao, L.; Liu, Q.; et al. Gut microbiota metabolites in Irritable Bowel Syndrome. Front.Cell.Infect.Microbiol., Sep.2021;11:729346. [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; et al. Gut dysbiosis is linked to hypertension. Hypertension, 2015;65:1331-1340. [CrossRef]
- Zagon, A. Does the vagus nerve mediate the sixth sense? TINS 2001;24:671–673.
Figure 1.
The fibers introduced with the diet are the essential nutritional substrate for the microbiota to produce short-chain fatty acids (SCFAs). The latter not only perform a fundamental trophic action at the level of the intestinal mucosa, but are also absorbed by passing into the circulation.
Figure 1.
The fibers introduced with the diet are the essential nutritional substrate for the microbiota to produce short-chain fatty acids (SCFAs). The latter not only perform a fundamental trophic action at the level of the intestinal mucosa, but are also absorbed by passing into the circulation.
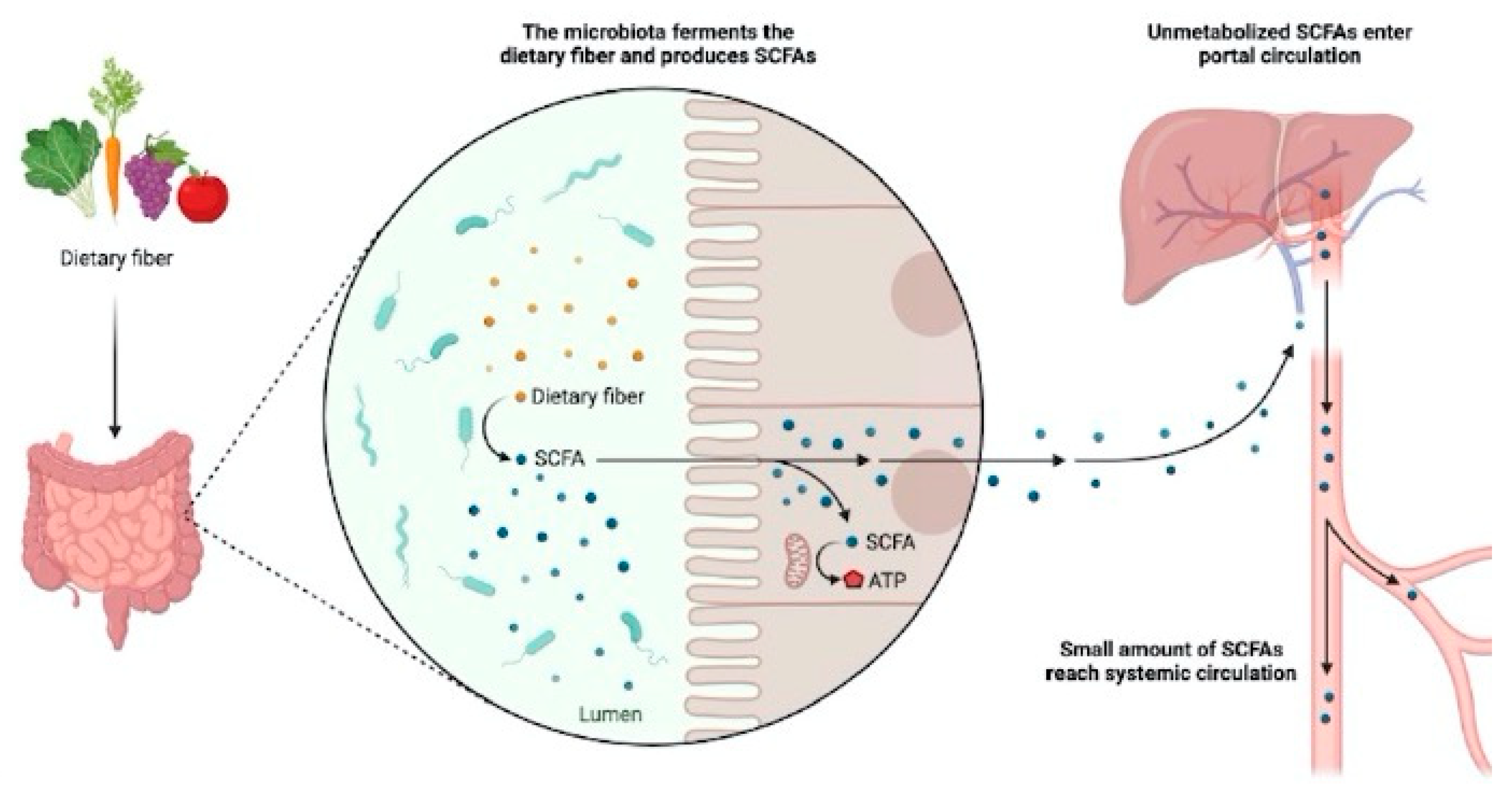
Figure 2.
The study of the biological system passes from the analysis of its individual components (on the left in blue) to the evaluation of the network of relationships between them (in yellow on the right) and the resulting effects on the functioning of the system.
Figure 2.
The study of the biological system passes from the analysis of its individual components (on the left in blue) to the evaluation of the network of relationships between them (in yellow on the right) and the resulting effects on the functioning of the system.
Figure 3.
Physiological reactivity of the organism to stress in relation to time. After an initial phase of alarm (RA) governed by catecholamines, a second phase known as resistance (SR) which involves cortisol follows, if the duration of the stress continues. The latter can be maintained as long as the body is able to deal with it with adequate cortisol secretion based on its individual reactive capacity, in addition to this capacity the body will end up in a condition of adrenal exhaustion (S.E.).
Figure 3.
Physiological reactivity of the organism to stress in relation to time. After an initial phase of alarm (RA) governed by catecholamines, a second phase known as resistance (SR) which involves cortisol follows, if the duration of the stress continues. The latter can be maintained as long as the body is able to deal with it with adequate cortisol secretion based on its individual reactive capacity, in addition to this capacity the body will end up in a condition of adrenal exhaustion (S.E.).
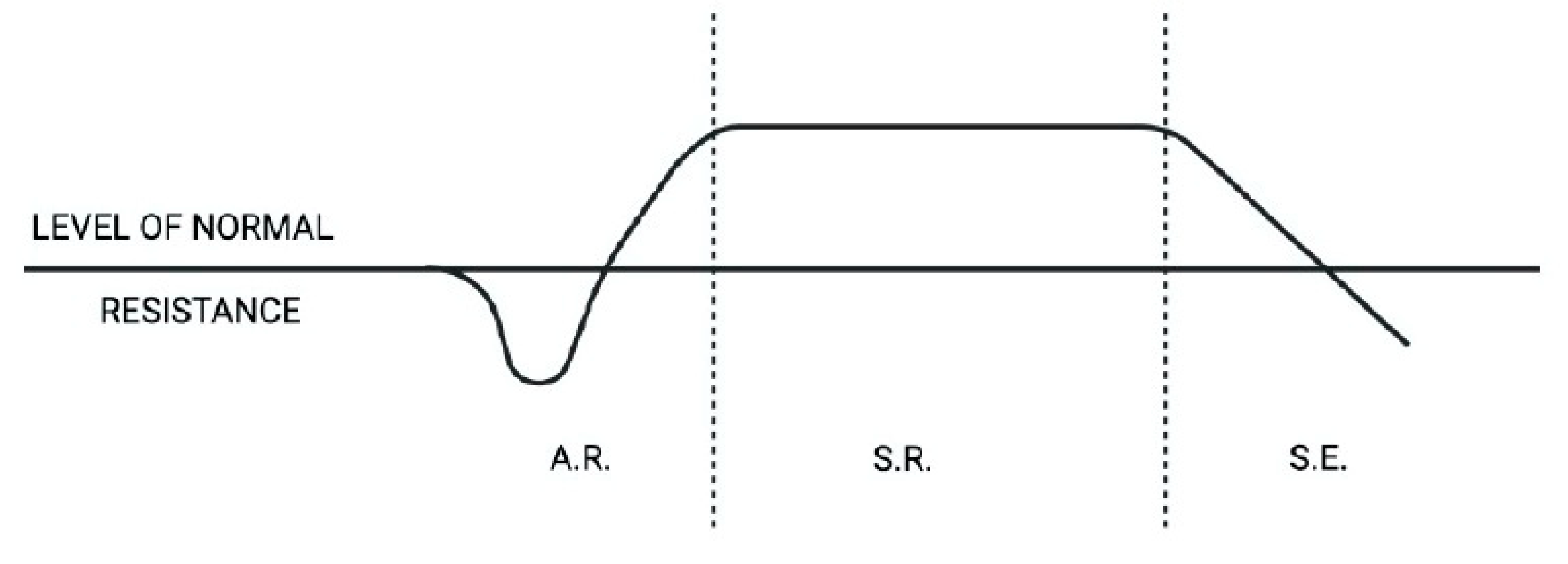
Figure 4.
Physiological levels of cortisol according with our circadian rhythm. A maximum peak is recorded between 06:00 and 08:00 in the morning and then see its values gradually decrease until they touch a minimum between 21:00 and 23:00 in order to predispose the organism to night rest. (Ore=hours).
Figure 4.
Physiological levels of cortisol according with our circadian rhythm. A maximum peak is recorded between 06:00 and 08:00 in the morning and then see its values gradually decrease until they touch a minimum between 21:00 and 23:00 in order to predispose the organism to night rest. (Ore=hours).
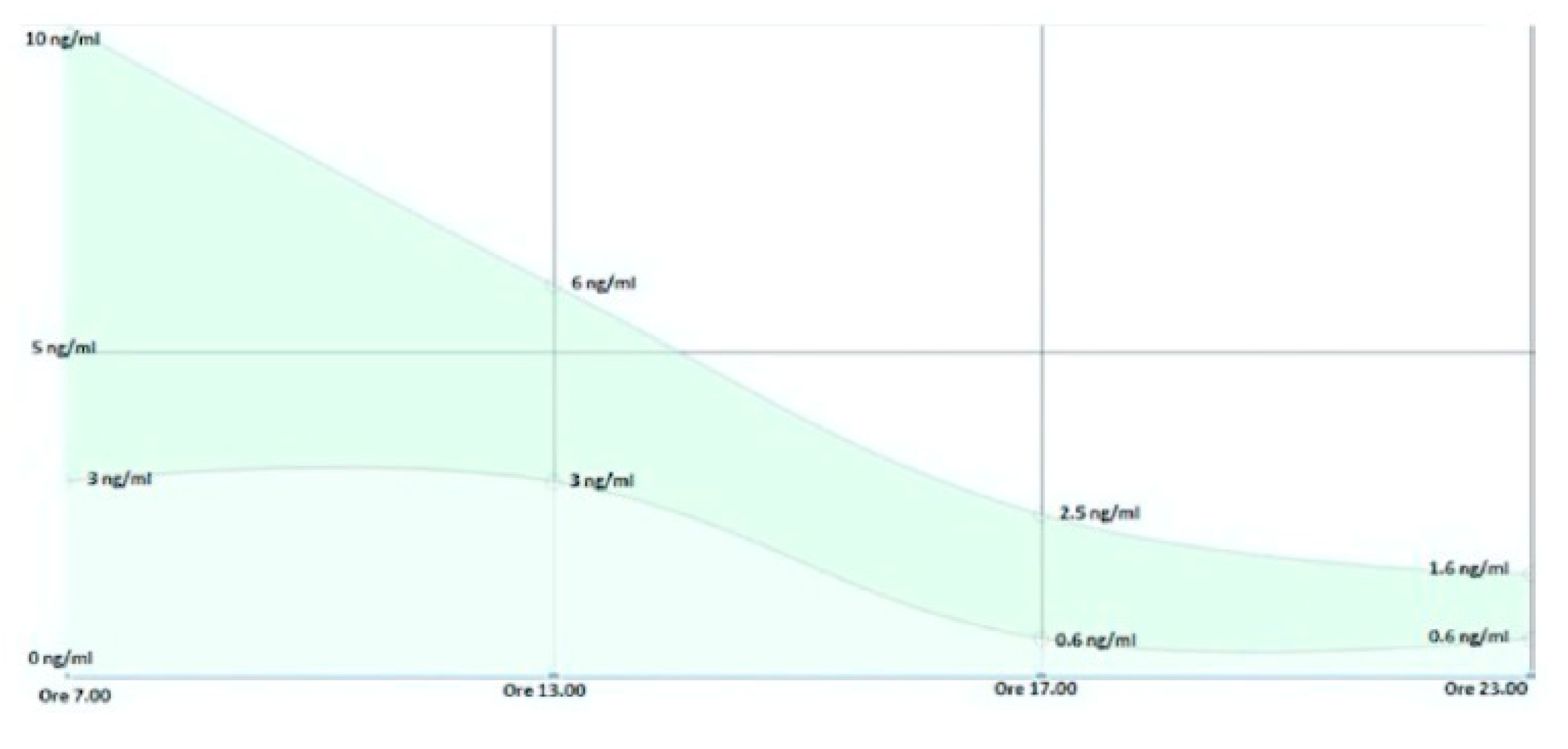
Figure 5.
Anatomy of the autonomic nervous system (ANS): in red the fibers of sympathetic system, in green those of the parasympathetic.
Figure 5.
Anatomy of the autonomic nervous system (ANS): in red the fibers of sympathetic system, in green those of the parasympathetic.
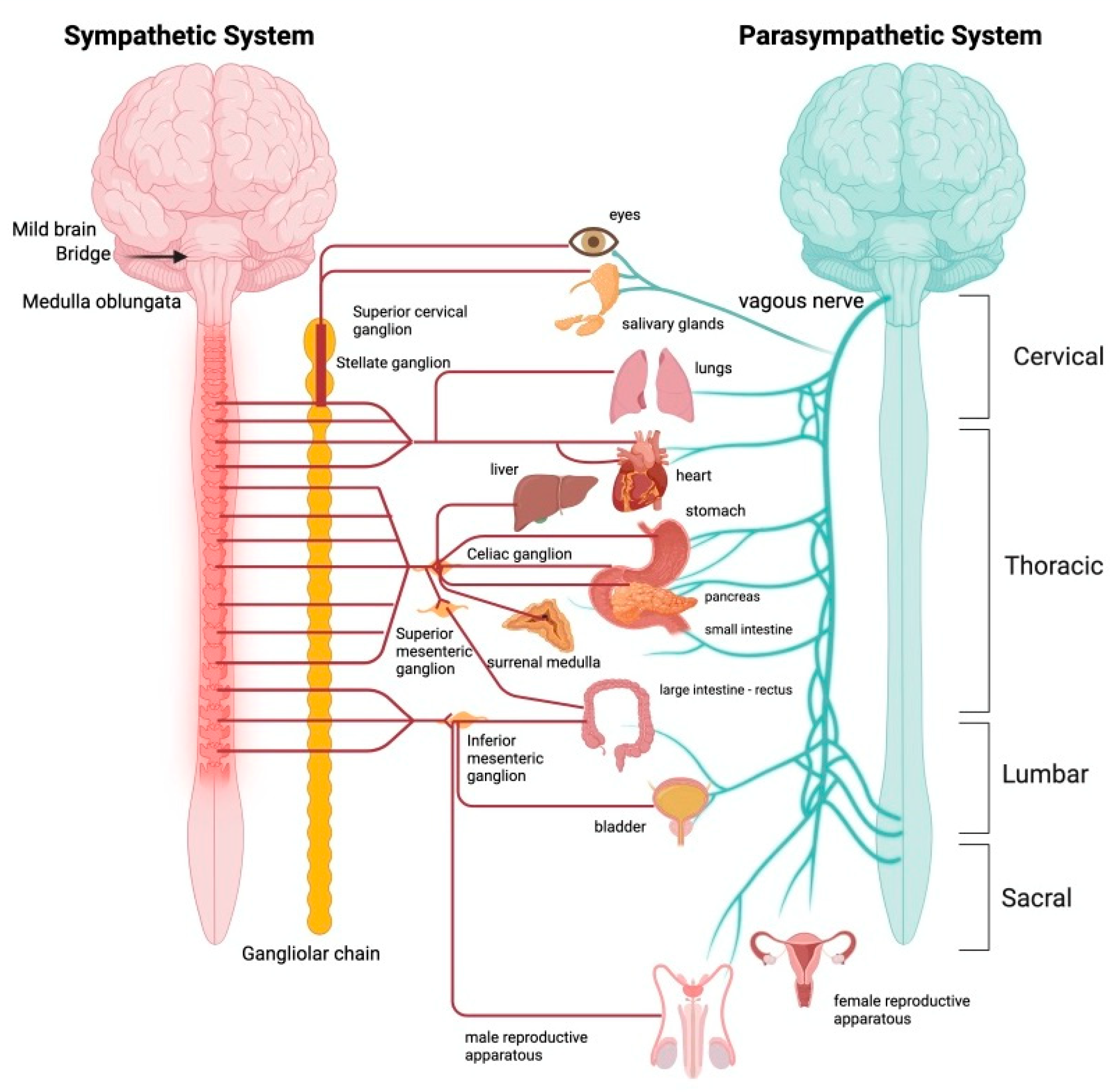
Figure 6.
Any system, in accordance with von Bertalanffy’s general systems theory, regardless of the starting conditions (a, b or c), will always tend towards a steady state.
Figure 6.
Any system, in accordance with von Bertalanffy’s general systems theory, regardless of the starting conditions (a, b or c), will always tend towards a steady state.
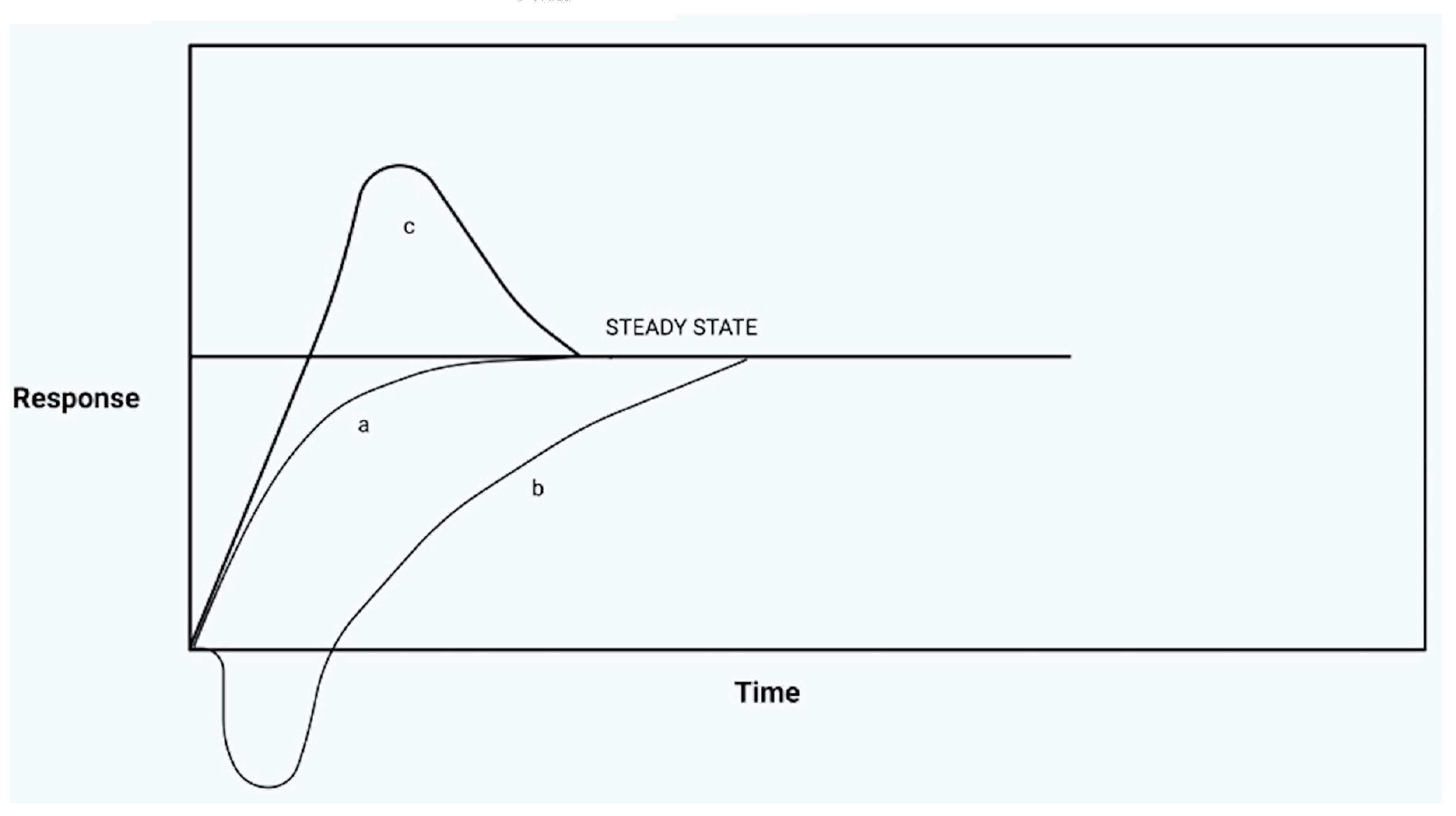
Figure 7.
Biosynthesis of serotonin from tryptophan.
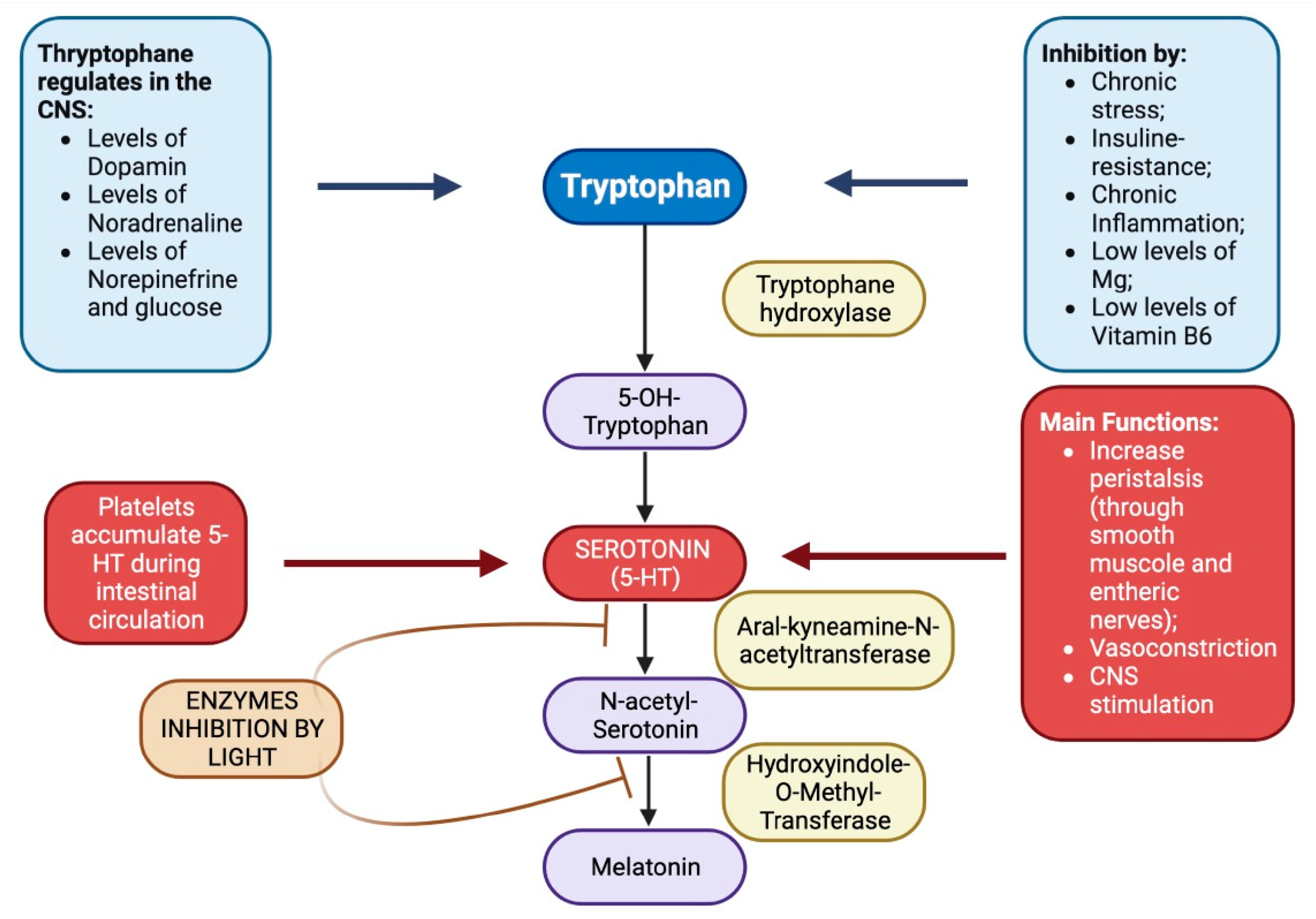
Figure 8.
The left side of the figure (Brain) shows the information exchange routes that reach the gastrointestinal system from the brain. It should be emphasized that this is a two-way path and that the information that travels upwards from the bottom is greater than what is recorded from the top to down. On the right side (Gut) we can observe how the presence of pathogens in the intestinal lumen is able to activate inflammatory signals, which mediators are involved and how the immune system reacts.
Figure 8.
The left side of the figure (Brain) shows the information exchange routes that reach the gastrointestinal system from the brain. It should be emphasized that this is a two-way path and that the information that travels upwards from the bottom is greater than what is recorded from the top to down. On the right side (Gut) we can observe how the presence of pathogens in the intestinal lumen is able to activate inflammatory signals, which mediators are involved and how the immune system reacts.
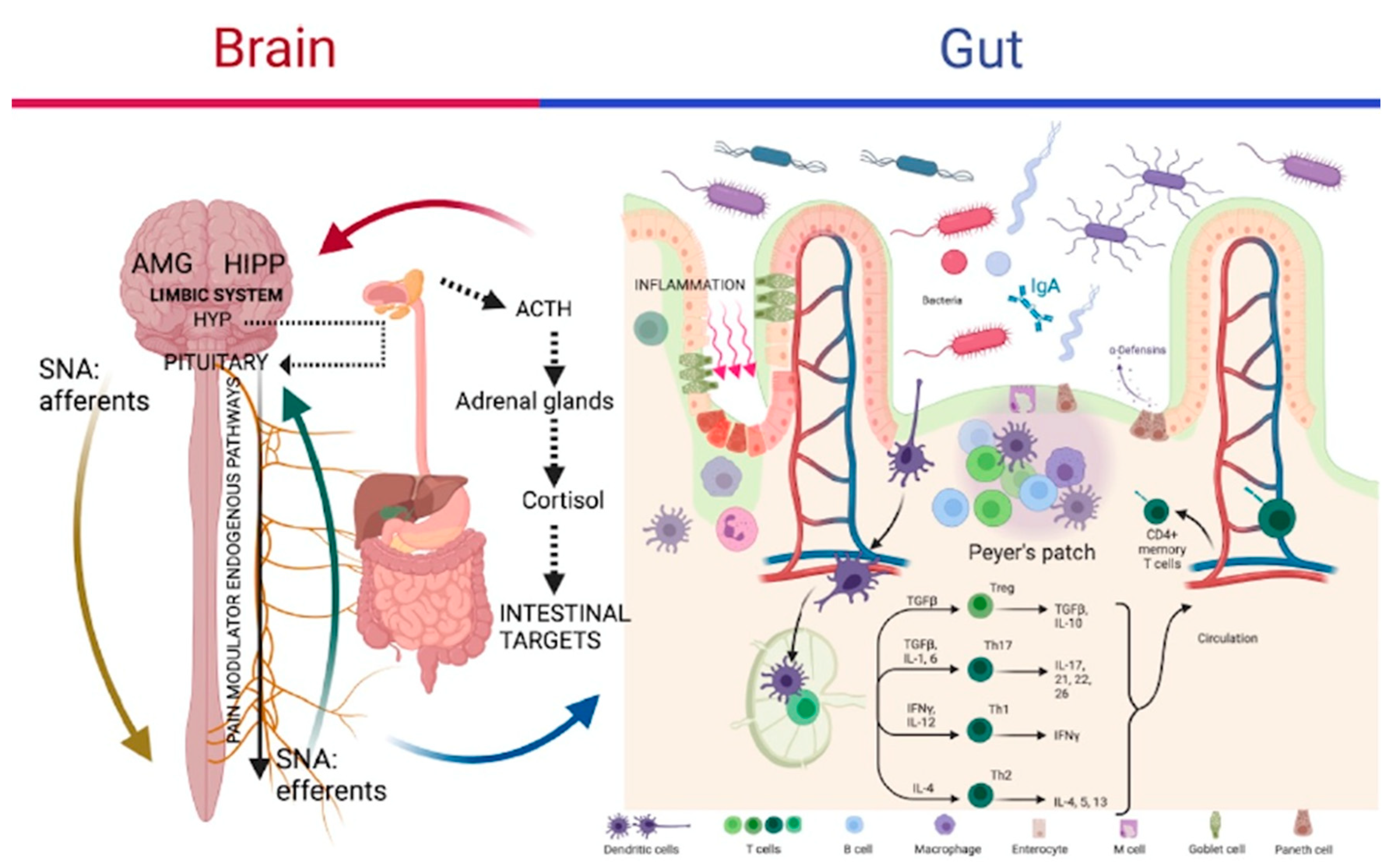
Figure 9.
From the study of the biochemical path involving omega 3/6 fatty acids it is clear that a diet that favors foods rich in omega 6 leads to the production of arachidonic acid and a consequent inflammatory state; conversely, a diet rich in omega 3 shifts the balance towards a greater presence of EPA and DHA with a consequent anti-inflammatory effect.
Figure 9.
From the study of the biochemical path involving omega 3/6 fatty acids it is clear that a diet that favors foods rich in omega 6 leads to the production of arachidonic acid and a consequent inflammatory state; conversely, a diet rich in omega 3 shifts the balance towards a greater presence of EPA and DHA with a consequent anti-inflammatory effect.
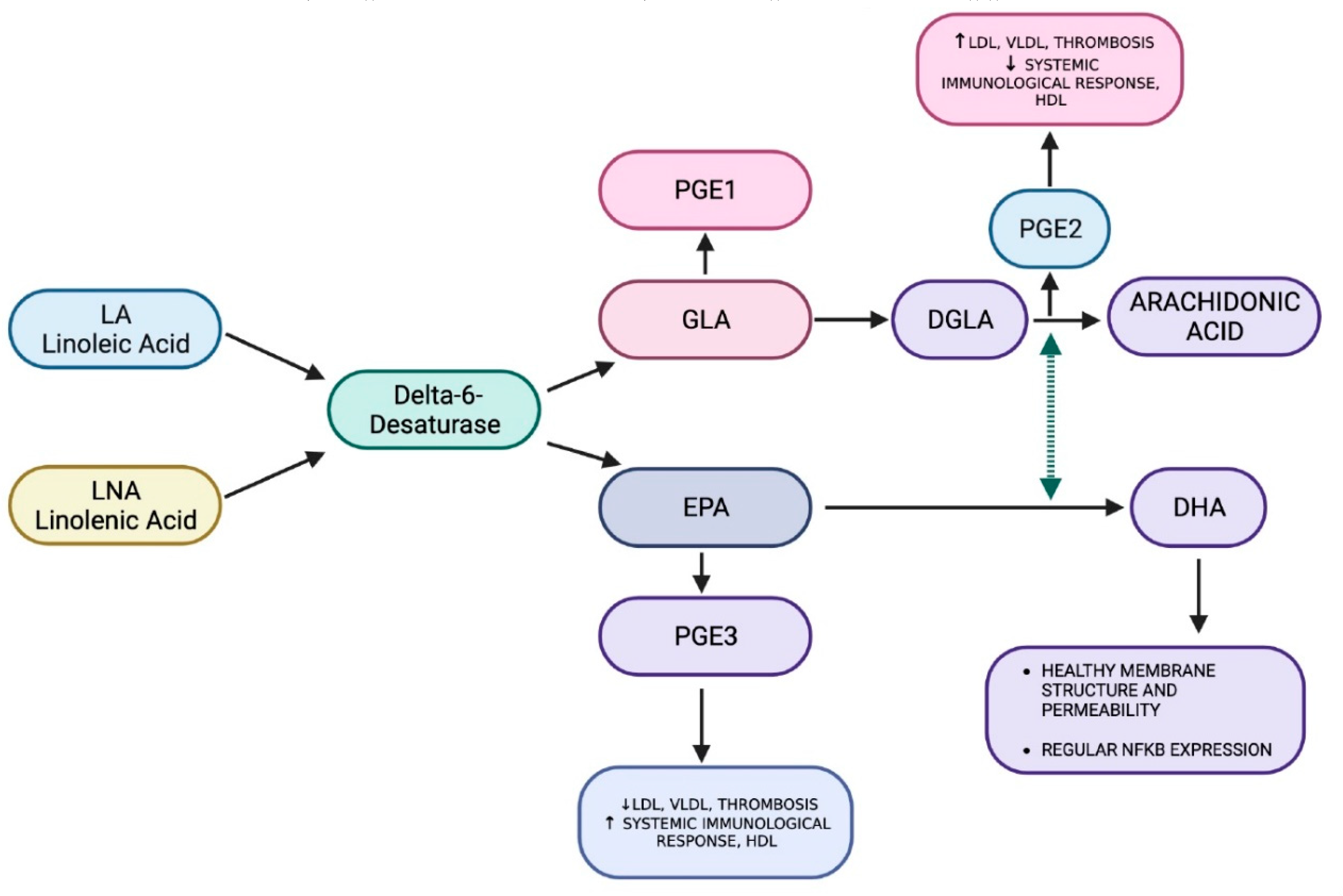
Figure 10.
Correlations between the main numerical variables considered in this research: the larger the colored circle, showed in the left table, the stronger the correlation (the black color indicates a strong positive correlation, while the white ones a negative type). The same correlations are shown in the right table but expressed in numerical terms.
Figure 10.
Correlations between the main numerical variables considered in this research: the larger the colored circle, showed in the left table, the stronger the correlation (the black color indicates a strong positive correlation, while the white ones a negative type). The same correlations are shown in the right table but expressed in numerical terms.
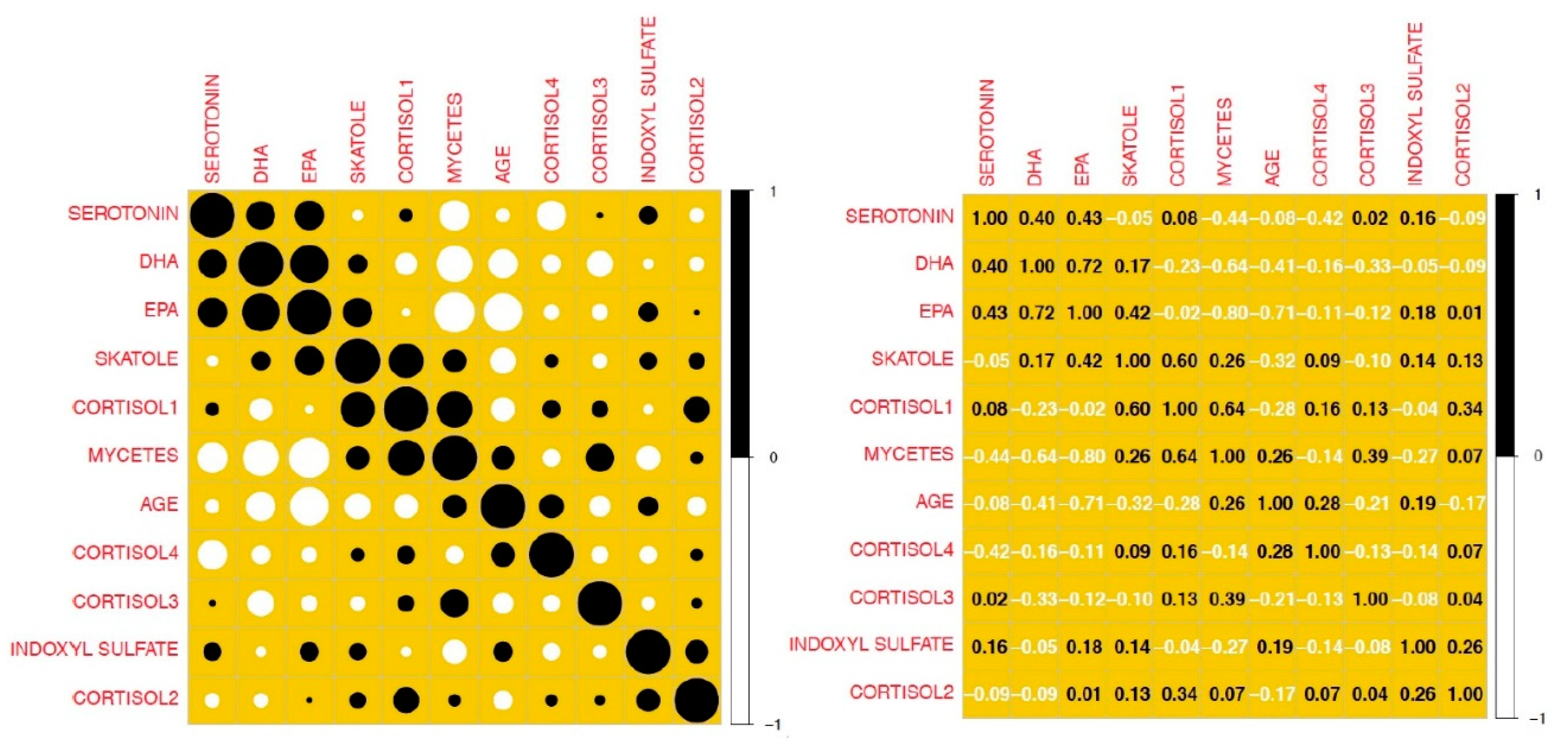
Figure 11.1/11.2/11.3.
Levels of serotonin, indoxyl sulfate and skatole analyzed in relation to the presence or absence of Candida a. Figure 11.1 shows that the plasma levels of serotonin are on average higher in the 3rd and 4th quartile than in patients who do not present Candida a.. In Figure 11.2 we can observe how this evidence is not recorded since skatole levels appear substantially identical between the group of patients with Candida and those without. At last, Figure 11.3 display how the levels of indoxyl sulfate, expressed both in the 2nd and in the 3rd and 4th quartiles, are clearly higher in the case in which it is excluding the presence of Candida a.
Figure 11.1/11.2/11.3.
Levels of serotonin, indoxyl sulfate and skatole analyzed in relation to the presence or absence of Candida a. Figure 11.1 shows that the plasma levels of serotonin are on average higher in the 3rd and 4th quartile than in patients who do not present Candida a.. In Figure 11.2 we can observe how this evidence is not recorded since skatole levels appear substantially identical between the group of patients with Candida and those without. At last, Figure 11.3 display how the levels of indoxyl sulfate, expressed both in the 2nd and in the 3rd and 4th quartiles, are clearly higher in the case in which it is excluding the presence of Candida a.
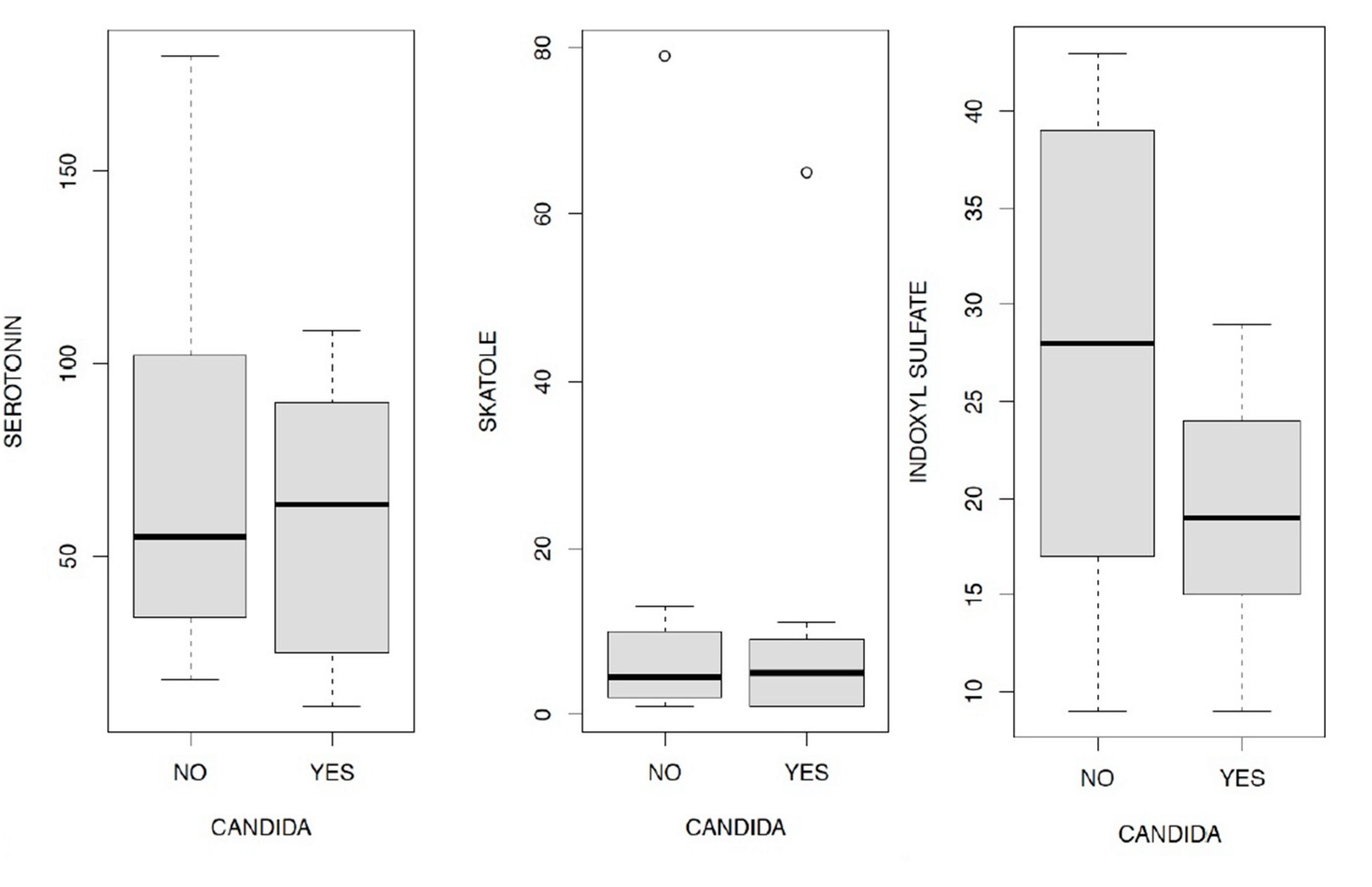
Figure 12.1/12.2/12.3/12.4.
Relationship between Candida a. and cortisolemia. We wanted to investigate the ratios in the four peaks physiologically expressed in our organism at four distinct times of the day: 7.00 (cortisol 1), 13.00 (cortisol 2), 17.00 (cortisol 3), 23.00 (cortisol 4). In all four comparisons it is evident that in the presence of Candida a. higher cortisol levels are on average associated with all four measurements.
Figure 12.1/12.2/12.3/12.4.
Relationship between Candida a. and cortisolemia. We wanted to investigate the ratios in the four peaks physiologically expressed in our organism at four distinct times of the day: 7.00 (cortisol 1), 13.00 (cortisol 2), 17.00 (cortisol 3), 23.00 (cortisol 4). In all four comparisons it is evident that in the presence of Candida a. higher cortisol levels are on average associated with all four measurements.
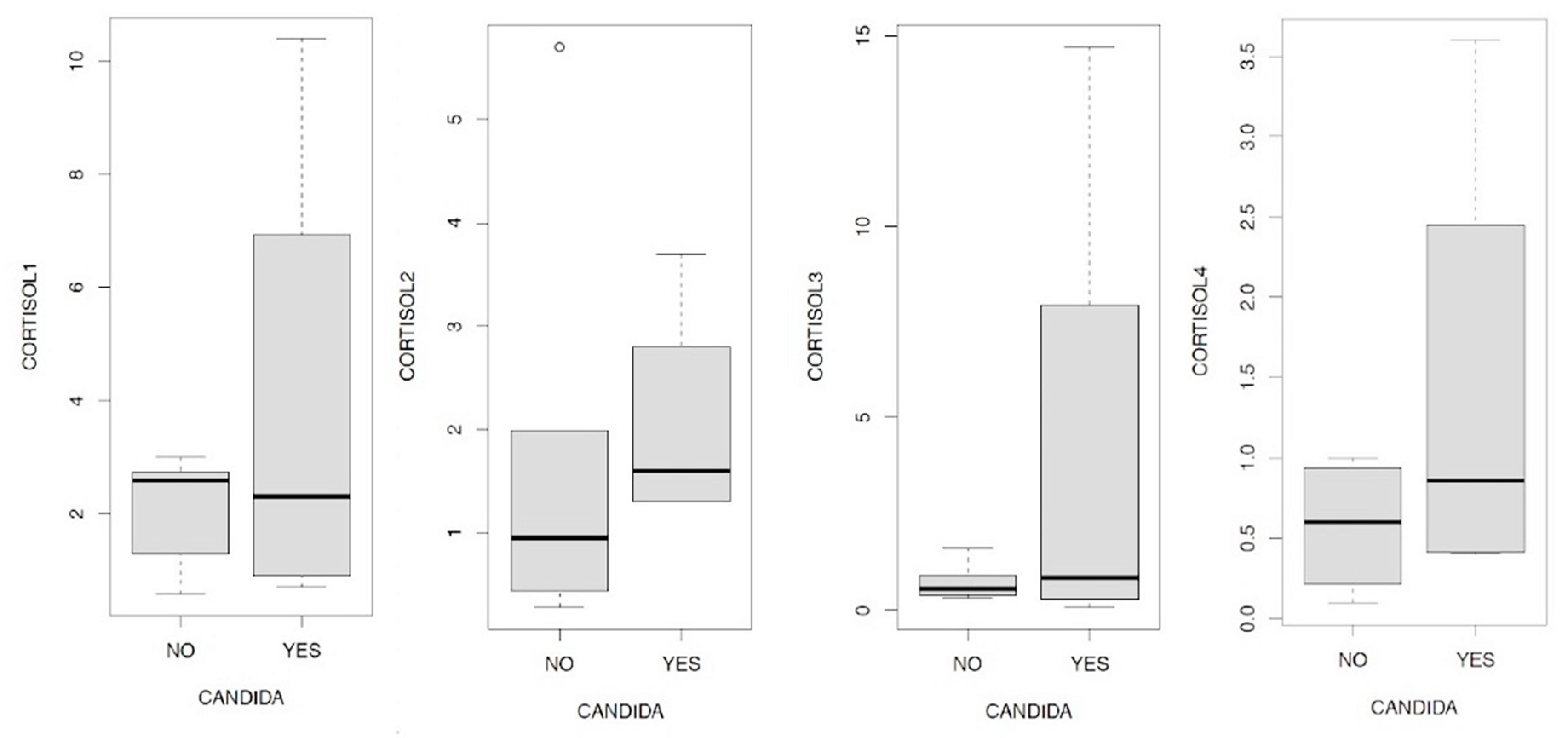
Figure 13.1/13.2.
Relationships between the values relating to the levels of DHA (Figure 13.1) and EPA (Figure 13.2) in the presence or absence of Candida a.. The presence of Candida a. it is accompanied by a decrease in the concentration of EPA, in the case of DHA the opposite trend is observed.
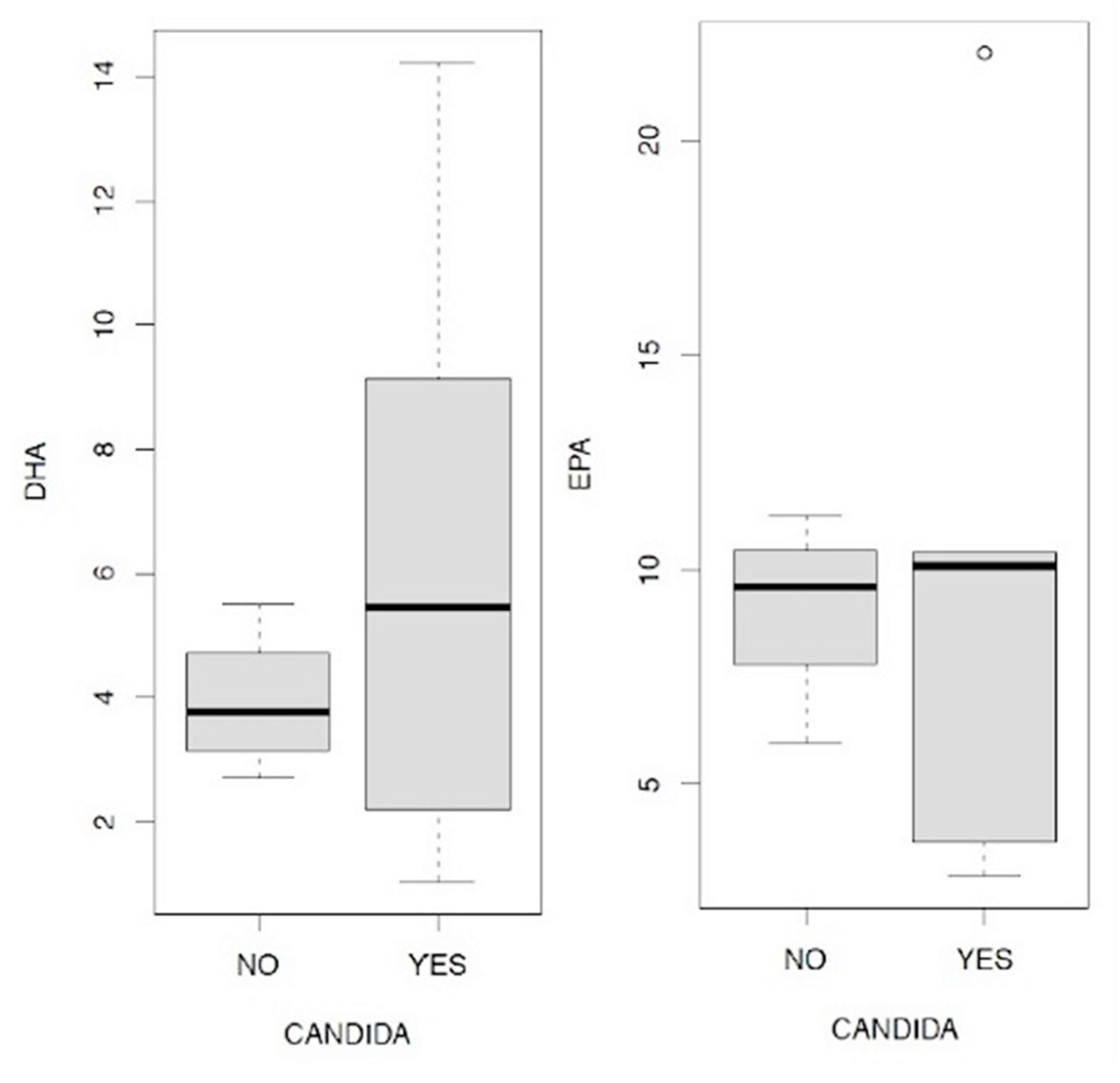
Figure 14.1/14.2/14.3/14.4.
Figure 14.1: relationship between the levels of Candida a. and mycetes. In all four quartiles in the presence of Candida a. higher levels of mycetes are associated with respect to subjects in which the fungus was absent. Figure 14.2: high levels of fungi are mainly of female relevance, as well as the lower levels of EPA and DHA seem prevalent typical of the women included in the study, as it shown in Figure 14.3/14.4.
Figure 14.1/14.2/14.3/14.4.
Figure 14.1: relationship between the levels of Candida a. and mycetes. In all four quartiles in the presence of Candida a. higher levels of mycetes are associated with respect to subjects in which the fungus was absent. Figure 14.2: high levels of fungi are mainly of female relevance, as well as the lower levels of EPA and DHA seem prevalent typical of the women included in the study, as it shown in Figure 14.3/14.4.
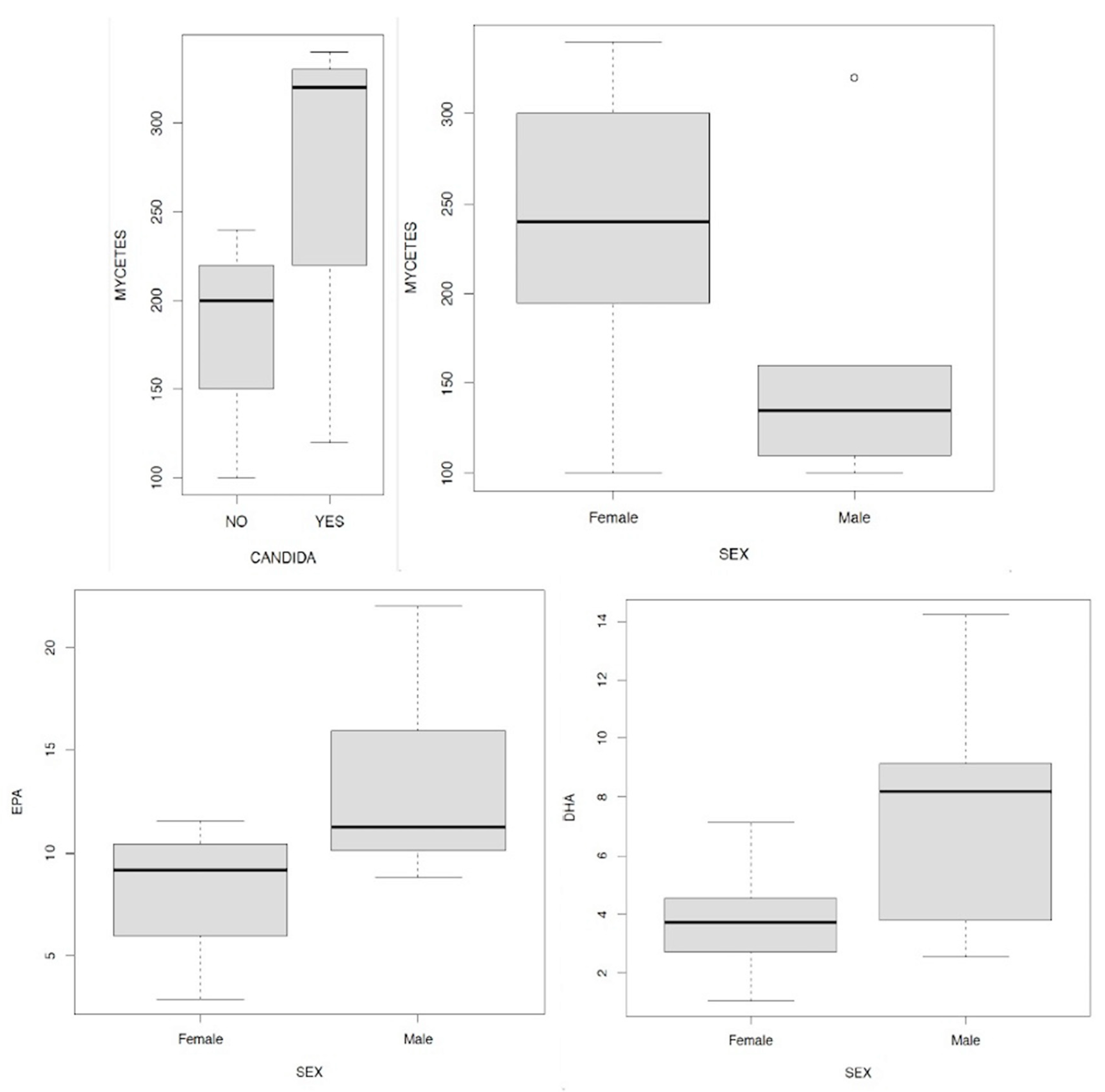
Figure 15.1/15.2/15.3/15.4.
Relationship between the levels of serotonin with respect to the values of skatole, mycetes, EPA and DHA using as a discriminant the presence of low or high levels of the same serotonin.
Figure 15.1/15.2/15.3/15.4.
Relationship between the levels of serotonin with respect to the values of skatole, mycetes, EPA and DHA using as a discriminant the presence of low or high levels of the same serotonin.
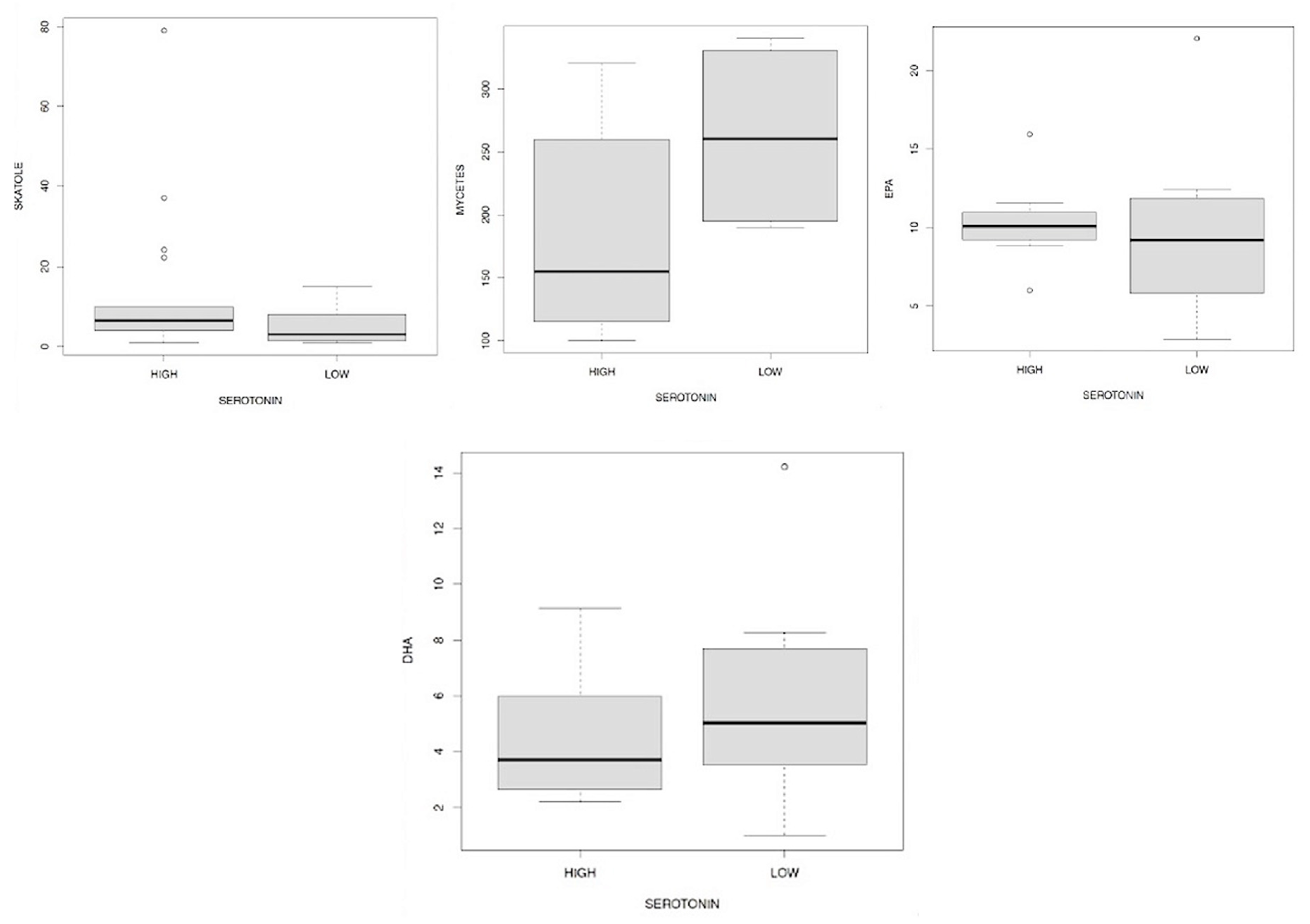
Figure 16.1/16.2.
Relationship between the levels of serotonin with respect to the values of fungi and skatole using as a discriminant the presence of high, normal or low levels of the same serotonin. As it is possible to observe in Figure 16.1, a weak dependence emerges between the presence of skatole and the serotonin values, while it is interesting to remark how low serotonin levels are found in the presence of high levels of mycetes as it is shown in Figure 16.2.
Figure 16.1/16.2.
Relationship between the levels of serotonin with respect to the values of fungi and skatole using as a discriminant the presence of high, normal or low levels of the same serotonin. As it is possible to observe in Figure 16.1, a weak dependence emerges between the presence of skatole and the serotonin values, while it is interesting to remark how low serotonin levels are found in the presence of high levels of mycetes as it is shown in Figure 16.2.
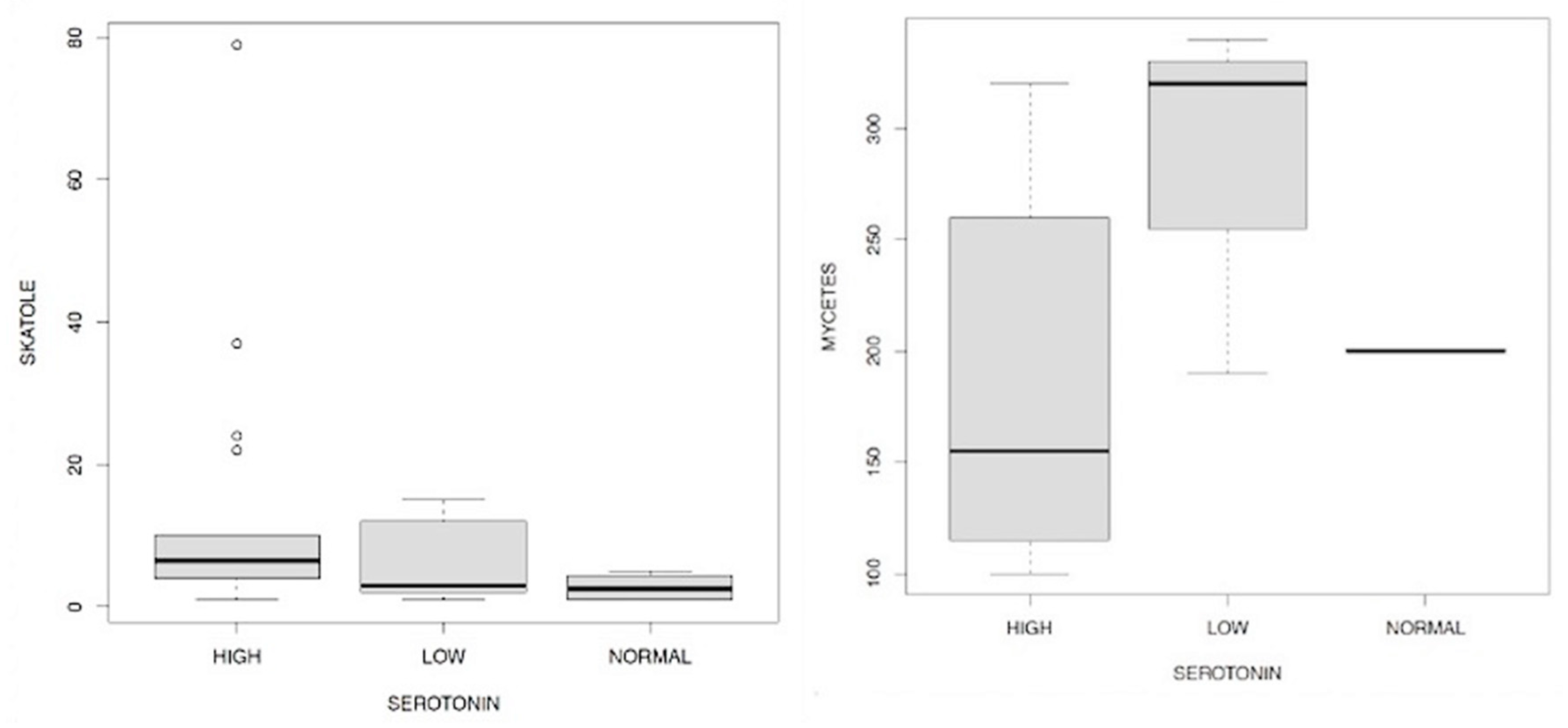
Figure 17.1/17.2/17.3/17.4.
Correlations between the cortisol values, measured in the four different and physiologically significant intervals of the day (7.00, 13.00, 17.00, 23.00), and the serotonin values indicated in the reference intervals. we can easily observe an evident relationship between low serotonin levels and high concentrations of Cortisol-1/2/4.
Figure 17.1/17.2/17.3/17.4.
Correlations between the cortisol values, measured in the four different and physiologically significant intervals of the day (7.00, 13.00, 17.00, 23.00), and the serotonin values indicated in the reference intervals. we can easily observe an evident relationship between low serotonin levels and high concentrations of Cortisol-1/2/4.
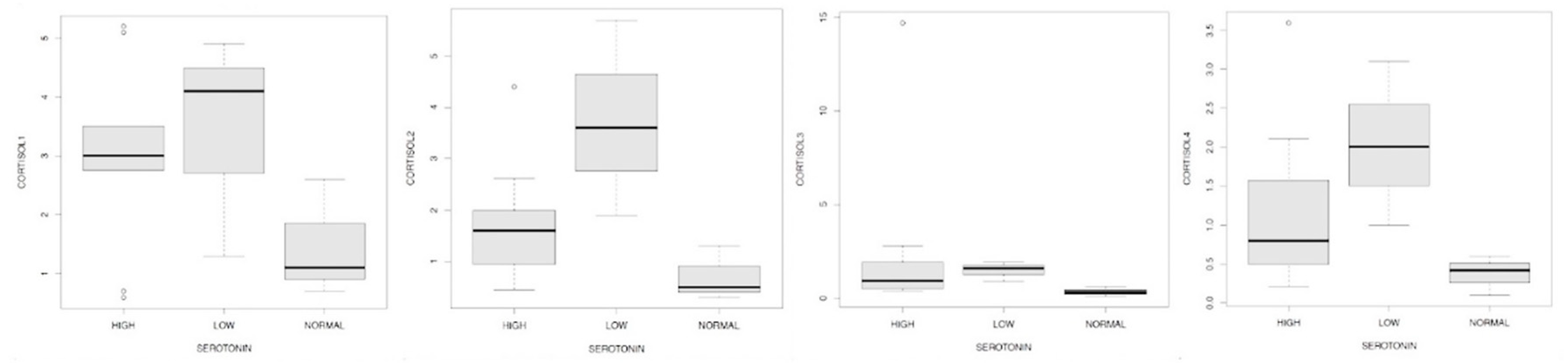
Figure 18.1/18.2.
Relationship between the values of EPA and DHA compared to those of serotonin. We can observe how high levels of EPA and DHA are accompanied by normal concentrations of the neurotransmitter.
Figure 18.1/18.2.
Relationship between the values of EPA and DHA compared to those of serotonin. We can observe how high levels of EPA and DHA are accompanied by normal concentrations of the neurotransmitter.
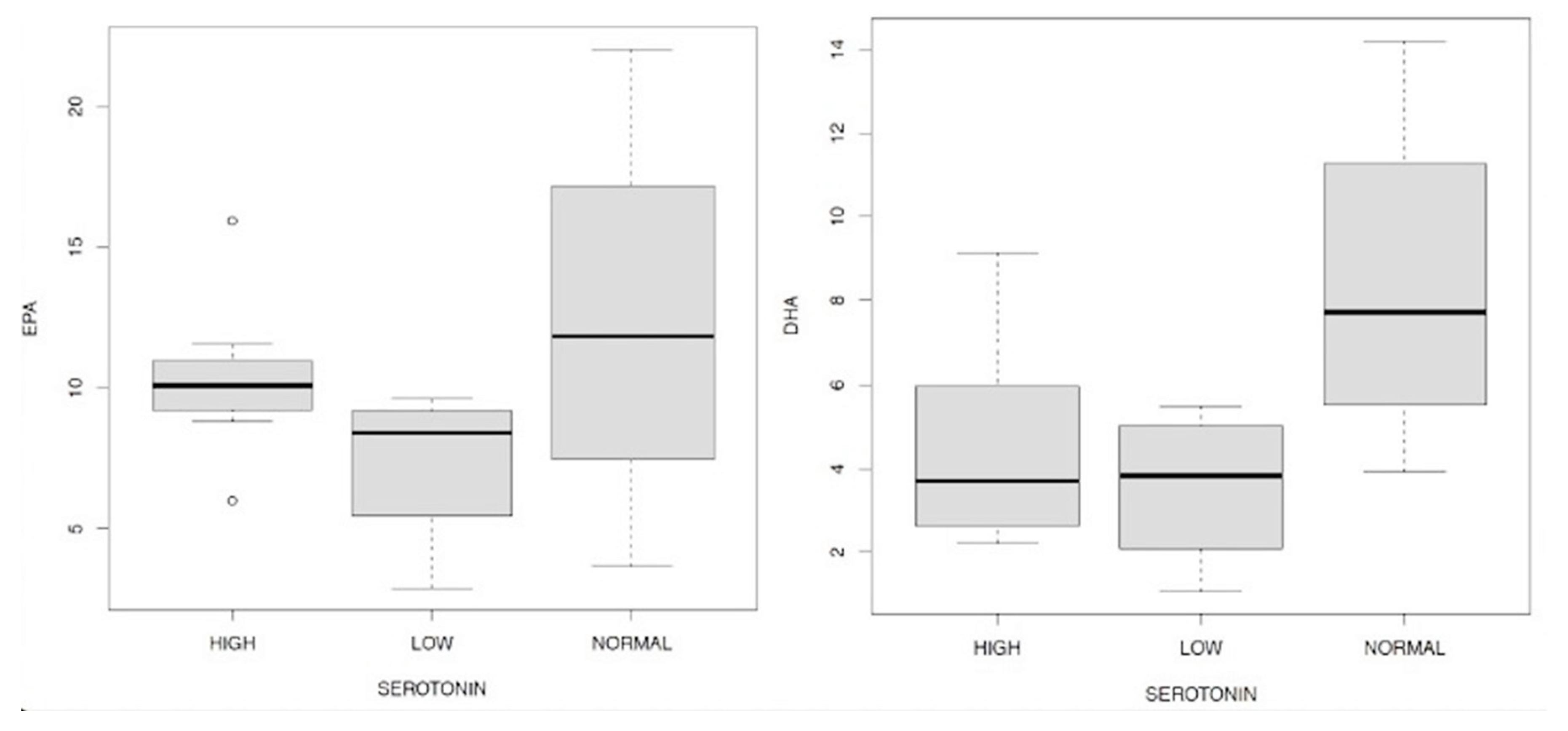
Figure 19.
Multivariate analysis that investigates the main determinants that induce a variation in the levels of serotonin, the inclusion of the variables is conditioned by the levels of correlation with serotonin. From the study of these data we observe how men have a higher level of serotonin than women examined, cortisol 1 (measured at 7.00) and 3 (measured at 17.00) have a positive and statistically significant effect on serotonin, while the increase in cortisol 4 (measured at 23.00) there is a statistically significant reduction in serotonin values; furthermore in the presence of Candida a. and/or mycetes we registered a lower level of serotonin than in subjects who do not have Candida a. and/or mycetes. In the end, all three levels of indoxyl sulfate reported contribute to the determination of changes in serotonin in an inversely proportional way.
Figure 19.
Multivariate analysis that investigates the main determinants that induce a variation in the levels of serotonin, the inclusion of the variables is conditioned by the levels of correlation with serotonin. From the study of these data we observe how men have a higher level of serotonin than women examined, cortisol 1 (measured at 7.00) and 3 (measured at 17.00) have a positive and statistically significant effect on serotonin, while the increase in cortisol 4 (measured at 23.00) there is a statistically significant reduction in serotonin values; furthermore in the presence of Candida a. and/or mycetes we registered a lower level of serotonin than in subjects who do not have Candida a. and/or mycetes. In the end, all three levels of indoxyl sulfate reported contribute to the determination of changes in serotonin in an inversely proportional way.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



