
Submitted:
01 September 2023
Posted:
05 September 2023
You are already at the latest version
Abstract
In sub-Saharan Africa, sweetpotato weevils are the most devastating pests of cultivated sweetpotato, causing estimated losses of between 60% and 100%, primarily during dry spells. The predominantly cryptic feeding behavior of the Cylas spp within the roots makes their control difficult, thus host plant resistance is one of the most promising lines of protection against these pests. However, progress in breeding for weevil-resistant cultivars has been slow in part due to the complex hexaploid genome of sweetpotato, which complicates conventional breeding in addition to the scarcity of varieties with significant levels of resistance for use as sources of resistance. Pollen sterility, cross incompatibility and poor seed set and germination in sweetpotato are also common challenges to improving weevil resistance. Accurate phenotyping of sweetpotato weevil resistance to enhance efficiency of selection has been equally difficult. Genomics-assisted breeding though in its infancy stages in sweetpotato has the potential application in overcoming some of these barriers. However, it will require the development of more genomic infrastructure; particularly single nucleotide polymorphism markers (SNPs), robust next generation sequencing plat-forms together with relevant statistical procedures for analyses. With the recent advances in genomics, we anticipate that genomic breeding for sweetpotato weevil resistance will be expedited in the coming years. This review sheds light on Uganda's efforts to date to breed against the Cylas puncticollis (Boheman) and Cylas brunneus (Fabricius) species of African sweetpotato weevil.
Keywords:
sweetpotato
; weevils
; Cylas spp.
; resistance
; breeding
; Uganda
1. Introduction
Sweetpotato (Ipomoea batatas (L.) Lam.) (2n = 6x = 90) is the third most important root crop in the world after potato and cassava, with an estimated total global production of 88.7 million metric tons [1]. It is one of the major staple foods in sub-Saharan Africa (SSA), where it ranks as the second most important root crop after cassava. Africa is currently the 2nd largest sweetpotato producing continent, after Asia, with an estimated annual production of 28.6 million metric tons accounting for 32% of the total worldwide production [1]. This clonally propagated crop is part of the Convolvulaceae family and is the only economically important species of the Ipomoea genus [2].
Sweetpotato is predominantly cultivated by small holder farmers through subsistence farming in Uganda. Its importance in the region is mainly attributed to its contribution to food security and income generation for marginalized farmers [3]. Sweetpotato is known to possess a wide range of nutrients including carbohydrates (21%), fibre (3.3%), minerals (potassium, copper and manganese) and vitamins such as vitamin A, B6 and C [4]. The orange-fleshed genotypes are particularly known to be an important source of β-carotene, a precursor for vitamin A and are making a significant contribution towards alleviating vitamin A deficiency in SSA particularly among children and pregnant women [5,6]. Many farmers in Uganda and the region like it as a crop because of its ability to generate high yields in relatively poor soils with minimum inputs and its resistance to a variety of biotic and abiotic stresses [5,7].
Sweetpotato weevils (SPWs) Cylas spp., are the most destructive pests of sweetpotatoes in Uganda and the SSA region, causing estimated yield losses of between 60% and 100% [8,9,10], mainly during dry spells and under heavy infestations. Cylas puncticollis (Boheman) and Cylas brunneus (Fabricius) are the predominant species occurring in Africa [8,11]. They attack the leaves, stems, and storage roots of sweetpotatoes, resulting in poor quality roots with an unacceptable smell and bitter taste due to secretion of defense sesquiterpenes by the infested plants, that makes the roots unfit for human and livestock consumption [12,13].
The concealed feeding behavior of weevils within storage roots makes them quite challenging to control by methods such as chemical control. Host plant resistance, therefore, presents a more suitable alternative for the integrated pest management (IPM) of this pest. Sweetpotato breeding, however, remains a difficult task due to the large and complex nature of its genome, with additional limitations such as self and cross incompatibility, limited flowering ability and seed setting [14], which further complicate conventional breeding.
The development of SPW resistant varieties has been unsuccessful to date, owing to a lack of high heritable resistance in the sweetpotato germplasm evaluated in the past [3,15]. However, some landraces with moderate resistance have previously been identified and researchers are currently using these as parental materials in an effort to develop resistant varieties [16,17]. Genomic selection and marker assisted selection stand out as promising options for developing SPW resistant varieties in Uganda [17]. This review gives an insight into the progress that has so far been made in breeding for resistance to the African sweetpotato weevils, Cylas puncticollis (Boheman) and Cylas brunneus (Fabricius) in Uganda. It also highlights aspects relating to the management and control of the two Cylas species.
2. Taxonomy and Geographical Distribution of Cylas species
Sweetpotato weevils (Cylas spp.) belong to the Order Coleoptera, Brentidae family and Cyladinae subfamily [18]. They are divided into three groups; C. brunneus, C. puncticollis and C. formicarius, which vary in physical characteristics such as body shape, length of the hind femora, head structure, and genitalia features [8]. Cylas formicarius is typically an Asian species but occurs throughout the tropical and sub-tropical regions of the world including Asia, Europe, North America and Africa [19]. Cylas puncticollis and Cylas brunneus however, are restricted to Africa [8,20]. They are the most prevalent and commonly reported SPW species on the continent [16,21,22,23]. Cylas puncticollis in particular, is the most abundant in tropical Africa, reported in 24 African countries, including Uganda, Rwanda, Ghana, Nigeria, DR Congo, Kenya, Malawi, Cameroon and Ethiopia [13,24].
3. Description and Life cycle of Cylas species
Adult females of Cylas species usually lay their oval, yellowish eggs singly in a cavity excavated either in sweetpotato roots or stems, which they subsequently seal with faecal plug [25]. Roots are the most preferred oviposition sites and most eggs tend to be deposited near the crown of the root. Duration of the egg stage varies from 5 to 12 days, with eggs of C. puncticollis developing faster than those of C. brunneus [26]. The larvae of Cylas spp. are usually curved in shape and white in color, feeding and developing within the stems and roots of sweetpotato. Pupae are glabrous in shape, with those of C. puncticollis being relatively larger than C. brunneus [25] (Figure 1). Male and female adults of both species can be differentiated by the shape of the distal antennal segment, which is filiform in males and club like in females [18]. The two species Cylas spp. differ in morphology, coloration and general development (Table 1).
4. Host Range and Dispersal of Cylas species
Sweetpotato is the preferred host plant for C. puncticollis and C. brunneus. In Uganda, the demand for additional hosts for weevil survival may not be as great because most cropping systems allow for year-round sweetpotato production. On the other hand, other Ipomoea species do not produce swollen roots, which provide oviposition sites; hence, population build up may be very low. Sweetpotato weevils usually get dispersed by flying and crawling, with males being more agile and flying more than females [12,25]. Long-distance dispersal however, is mostly through the movement of infested sweetpotato storage roots and vines [13].
5. Damage Caused by Cylas species
Cylas spp damage the leaves, stems and storage roots of sweetpotato. Adults injure the vines and leaves by feeding on the epidermis, scraping and creating oval patches, which may lead to a significant reduction in plant vigor and poor establishment in case of early infestation [8]. Larvae usually damage the stem by feeding at the base, thereby affecting the vascular system, causing the vines to swell and eventually wilt or die [13]. However, the most significant damage is inflicted when weevils attack the underground storage roots. Adults feed on the external root surfaces causing feeding punctures, which can be distinguished from oviposition sites by their greater depth and the absence of faecal plug [27]. Larval feeding and tunneling creates irregular galleries in the inner part of the root or stem, further damaging the plant (Figure 2). In response to this, the roots produce sesquiterpenes in self-defense, which give the root an unacceptable odour and bitter taste, rendering them unsuitable for consumption either as food or animal feed [13,15]. The damage caused by Cylas spp therefore affects the quantity, quality and value of sweetpotato storage roots produced. Losses of commercial storage root yield as high as 60–100% have been reported under heavy infestations in SSA [8,28].
6. Management and Control Strategies
A number of agricultural practices have been suggested for the management and control of SPWs (Table 2). Because none of them is completely effective in the control of this elusive pest, an integrated pest management (IPM) approach is usually recommended. Control is made difficult because all immature developmental stages of Cylas spp. occur deep inside the storage roots and stems of the plant [13].
7. Mechanisms of resistance of sweetpotato to Cylas spp.
Different resistance mechanisms found existing in sweetpotato germplasm against the SPW have previously been reported including antibiosis, antixenosis, and escape [44]. Other studies demonstrated that SPW resistance is more than simply an escape mechanism through field phenology such as deep rooting plants, but rather an active mechanism that is quantifiable and potentially manageable for breeding purposes [3,15,23]. Hydroxycinnamic acid (HCA) esters in the root latex and root surface (epidermal and peridermal tissues) of sweetpotatoes were shown to deter the development of SPW larvae, reducing feeding damage and oviposition. The HCAs were identified as heptadecylcaffeic acid, hexadecylcaffeic acid, octadecylcaffeic, hexadecylcoumaric acid, octadecylcoumaric acid and 5-0-caffeoylquinic acids from Ugandan sweetpotato varieties “New Kawogo” and “Tanzania” [15,16,17]. These acids are a major group of phenolic compounds with bioactive properties that are produced by plants for protection against biotic and abiotic stress [45].
8. Progress in breeding for resistance to Cylas spp.
Progress in breeding for sweetpotato weevil resistance has been rather slow due to factors such as the complex nature of the sweetpotato genome and the limited sources of genetic resistance to Cylas spp [3,15]. Genetic studies in sweetpotato are limited by high levels of self and cross-incompatibility, limited flowering ability, high ploidy levels and poor seed germination as also observed in Taiwan [46]. Each successful cross usually leads to the production of less than three botanical seeds [47]. This makes conventional breeding that relies only on hybridization to create genetic variability, a challenge.
Nevertheless, the breeding program at the National Crops Resources Research Institute (NaCRRI) in Uganda, has in the past and to-date, used conventional breeding and population improvement through recurrent mass selection for variety development in Uganda [48,49]. The phenotyping of SPW resistance was based on severity and incidence estimations. Sweetpotato severity was routinely assessed using standard protocols and a scale of 1-9 was employed where 1 = no weevil damage symptoms, 2 = unclear weevil damage symptoms, 3 = clear weevil damage symptoms < 5% of roots per plot, 4 = clear weevil damage symptoms at 6 to 15% of roots per plot, 5 = clear weevil damage symptoms at 16 to 33% of roots per plot, 6 = clear weevil damage symptoms in 34 to 66% of roots per plot (more than 1/3, less than 2/3), 7 = clear weevil damage symptoms in 67 to 99 % of roots per plot (2/3 to almost all), 8 = clear weevil symptoms in all roots per plot, 9 = severe weevil damage symptoms in all roots per plot (rotting) [96]. Incidence of sweetpotato weevil damage was measured as a percentage of number of storage roots showing weevil damage symptoms to the total number of roots in a plot.
In the past, sweetpotato morphological traits such as vine vigor, ground cover, foliage weight, storage root stalk (neck length), storage root cortex thickness, storage root size, storage root shape, storage root formation (arrangement), root latex production, and dry matter content were reported to play an important role in conferring field resistance to SPW [41]. But through correlation and path coefficient analysis, neck length was identified as the key morphological SPW resistance predictor [50]. This trait will be useful in selecting for SPW resistance among breeding clones during early stages of breeding. This could be done at a single site before advancing clones for multi-location evaluation and analysis of biochemical hydroxycinnamic acids, thereby minimizing research costs.
To date, the National Crops Resources Research Institute of the National Agricultural Research Organization (NARO) has released 29 sweetpotato varieties of varying levels weevil resistance in Uganda [97-102]. All the 29 sweetpotato varieties that have been released in Uganda so far are either moderately resistant or susceptible to sweetpotato weevils (Table 3). Varieties like “New Kawogo”, which express moderate resistance to Cylas spp. are currently deployed as parental material in SPW breeding and related studies in Uganda [15].
9. The genetic and biochemical basis for sweetpotato weevil resistance
The development of sweetpotato varieties with resistance to SPWs requires an understanding of the underlying biochemical and genetic mechanisms of resistance to this pest. Our understanding of the genetic and biochemical basis of resistance to SPWs has been improved in the past decade through a number of studies. Previous research conducted to determine the basis of weevil resistance in the Ugandan landrace, “New Kawogo”, to the African SPW C. puncticollis revealed that resistance in this variety was not only active, but also quantifiable and manageable for breeding [15]. In another study, 134 sweetpotato cultivars and landraces were evaluated for weevil resistance. Some sweetpotato cultivars, including “New Kawogo”, expressed resistance to Cylas spp. while others such as “Tanzania” and “NASPOT1” turned out to be susceptible [23].
In order to identify the biochemical basis of resistance to SPWs, the performance of seven resistant varieties of sweetpotato were compared with three susceptible varieties in field trials and laboratory bioassays [16]. Using liquid chromatography-mass spectrometry (LC-MS) of root surface and epidermal extracts, significant variation in the concentration of hexadecyl, heptadecyl, octadecyl, and quinic acid esters of caffeic and coumaric acid were exhibited, with higher concentrations of these compounds correlated with SPW resistance. “New Kawogo” recorded the highest concentration of these compounds. It was therefore concluded that the selection of sweetpotato varieties with higher levels of HCAs, particularly in the storage root surface, would probably contribute to the development of resistance to SPWs. However, this would necessitate the screening of hundreds of breeding lines using freeze-dried samples of roots and analysis by LC-MS in order to achieve long-term population improvement. Developing such a chemotyping platform, though desirable, would be quite costly [17].
In a further effort to understand the segregation of HCA esters that confer resistance to SPWs, Anyanga, et al [3] quantified these chemicals among 287 progenies of a segregating bi-parental population of the “New Kawogo” x “Beauregard” (NKB) cross. Beauregard is a USA-released cultivar, highly susceptible to SPWs. Results revealed that HCA esters conferred resistance to SPWs and segregate in the F1, suggesting that they are controlled quantitatively, and thus, might be useful as markers for resistance in sweetpotato breeding programs. The progeny that displayed more resistance to SPWs than “New Kawogo”, together with those that had higher concentrations of HCA esters, were selected as new candidates and are currently being utilized as parental genotypes in sweetpotato breeding.
Knowledge on the inheritance and heritability of resistance to the African SPWs is also apparently still limited. Some studies have been done in the past to establish the heritability of SPW resistance in which resistance to Cylas spp. was measured on the basis weevil severity of field storage roots and total HCA ester concentrations [17]. A moderately high broad sense heritability was estimated (H2=0.49). Another study revealed similar results (with H2=0.49%) and further showed that additive effects contributed the most to SPW resistance and HCA concentration [51]. More recently, a narrow sense heritability (h2=0.30) was reported for SPW resistance based on a relatively high population [52]. We hope that the adoption of modern experimental designs such as the row-column designs, which use more replications for field phenotyping, will enable estimation of higher heritability values for SPW resistance for future genomic analyses.
10. Marker assisted selection and Genomic selection for SPW resistance
The limited number of genomic resources has been a major bottleneck in breeding for SPW resistance. Over the years, sweetpotato genomic improvement has generally lagged behind other important crops such as maize [53], rice [54] and wheat [54]. This has been attributed to the scarcity of molecular markers to facilitate genetic analyses [55], lack of appropriate specific statistical procedures due to its polyploid nature, and limited user-friendly computer software to implement the statistical procedures for genetic analyses of important traits [56]. Sweetpotato was termed as an “orphan crop” with limited private investment in the development of its genomics infrastructure [56]. Nevertheless, to maximize the success rate in developing sweetpotato varieties, genomic tools are critical [57], particularly since most traits in sweetpotato are quantitatively inherited [55,58].
Molecular markers have hardly been used in sweetpotato genetic research compared to other staple crops yet they are critical in the genetic analysis of complex traits [55,59] and increase the efficiency of selection for low heritability traits through marker assisted selection (MAS) [55,59].
Sweetpotato weevil resistance, like most sweetpotato traits of agronomic and economic importance is quantitative in nature; controlled by many genes and with high environmental plasticity [46,51]. This has resulted in slow breeding progress through the use of conventional breeding approaches. Associating such a complex trait to genomic regions can be done through Quantitative Trait Loci (QTL) analysis [59]. However, there is currently a low number of published SSR and SNP markers for sweetpotato [17,51,60,61]. In the past, most molecular genetic studies in sweetpotato were based on dominant markers such as random amplified Polymorphic DNA (RAPDs) and amplified fragment length polymorphisms markers (AFLPs) that are relatively easy to score [62,63,64]. However, these QTLs have limited utility and cannot be used for sweetpotato improvement by MAS and genomic selection (GS). Simple sequence repeats (SSR) and single nucleotide polymorphism markers (SNPs), which are amenable to automation, are more suitable for sweetpotato breeding through MAS [17,65].
Some initial progress has been made in identifying SSR and SNP markers for use in breeding for SPW resistance. Yada et al [66] conducted a study in which he examined the genetic diversity in the NKB population using SSR markers. Sweetpotato SSR markers (405) were screened for polymorphism on the parents and progeny of the biparental cross. A total of 133 SSRs were identified as informative and recommended for use in further sweetpotato diversity and linkage analysis. The same population was later genotyped with these SSR markers to identify SPW resistance loci [17]. Five SSR markers were observed to be associated with field SPW severity whereas seven were linked to HCA ester concentration. Three markers showed significant association, suggesting their high potential for use in the development of SPW resistant cultivars [17]. This was the first report of co-dominantly inherited SSR markers being utilized to identify SPW resistance loci in sweetpotato in Uganda. Next-generation sequencing (NGS) technology and a genotyping by sequencing approach was later used to develop SNP markers and high-density genetic linkage maps using the NKB population [51]. This led to the identification of 4 QTL, explaining a total of 37.7% of the variation for SPW resistance and 15.9% of the variation for storage-root HCA ester concentration in the mapping population. This was the first report of a QTL mapping study for resistance to SPWs and HCA ester concentration [51].
Genomic selection stands out as a novel approach to accelerate genetic gains in sweetpotato [67]. In their pioneering work to explore the potential of genomic selection on sweetpotato, the predictive ability based on several factors was done for simple traits and yield traits for a single population. Though the predictive ability was low for complex traits, the use of multi trait models offered confidence for better prediction of such traits [67]. In addition, studies in other polyploids like octaploid strawberry, have reported high accuracy for lower heritability traits [68,69]. Despite the reports of lower heritability for insect pest resistance across crop species, the application of genomic prediction in maize showed moderate to high predictive ability for fall army worm and maize weevil [70]. Therefore, the application of genomic selection for weevil resistance is feasible and can be achieved after optimizing the size, composition, structure and phenotyping of training population and assembling sufficient diversity in the population.
11. Genomic resources for sweetpotato improvement
Through global collaborative research, high quality sequencing of the genomes of two diploid close relatives of sweetpotato, I. trifida and I. triloba, was successfully accomplished [71] and a genome sequence-based marker platform for sweetpotato improvement was developed. Ipomoea trifida is the closest wild species to sweetpotato, as evidenced by a number of cytology and molecular studies [72,73,74,75]. Though it’s the preferred reference for I. batatas, the I. triloba genome can also be used as a complementary reference sequence in sweetpotato breeding [71]. Although another sweetpotato reference genome was released by Yang et al [76] and later on improved, efforts are still underway to develop a high quality whole genome sequence for hexaploid sweetpotato [77,78]. Once completed, this will facilitate genomic selection and enable more efficient introgression of important traits such as SPW resistance, ultimately leading to the development of sweetpotato varieties with desirable attributes.
The advances in DNA sequencing methods through next-generation sequencing (NGS) technology and bioinformatics resources over the past decade have enabled large numbers of SNP markers to be developed and utilized [77,78]. Genome wide association studies (GWAS), which investigate the association between genotypes and phenotypes using the entire genome, have also been facilitated by the development of NGS techniques. Through GWAS, existing phenotypic and/or genotypic data can be utilized from larger population sizes to produce higher statistical power in analysis provided that a sufficient marker density is available [77].
12. Genetic transformation for weevil resistance
Research has been done in SSA to develop varieties that are resistant to SPWs through genetic transformation and crossing landraces with transgenic cultivars. Transformation for weevil resistance was among the first that were attempted in sweetpotato [57]. Sweetpotato cultivar ‘KB1’ was transformed with a delta endotoxin gene cry8Db, acquired from Bacillus thuringiensis via Agrobacterium mediated transformation. This gene codes for an insecticidal protein that is lethal to many insect larvae. The 21 transgenic lines that were regenerated from the transformed calli displayed lower SPW infestation compared to the untransformed lines [78]. Agrobacterium mediated transformation was similarly used to transform ‘Kyebandula’, a Ugandan sweetpotato landrace, by introducing two bacterial endotoxin genes cry7A1 and cry3Ca. The process was reportedly successful as evidenced by positive PCR results. However, transformed calli were unable to regenerate into whole plants [79].
Ekobu et al [80] conducted an experiment in which they evaluated the toxicity of Cry proteins to C. puncticollis and C. brunneus. The three most active Bt Cry proteins, with potential to confer resistance against C. puncticollis and C. brunneus, were Cry7Aa1, cryET33-34 and Cry3Ca1 with an LC50 value (mg/ml diet) against the first instar C. puncticollis and second instar C. brunneus below 1 ppm. However, they did not show efficacy against the weevil, a result partly ascribed to the low level of Cry protein expression in the root tissue. In another study, the feasibility of crossing transgenic and non-transgenic sweetpotato to obtain transgenic progeny was explored [7]. However, there were no significant differences in Cry protein expression among hybrid sweetpotato derived from the transgenic event crossed to different conventional sweetpotato varieties. Transgenic sweetpotato expressing Cry proteins did not perform with the desired levels of resistance to C. puncticollis. The study recommended evaluation of more transgenic events carrying Cry proteins to increase the possibility of finding those with higher Cry protein accumulation.
Recently, Anyanga et al [81] studied the effect of combining the anti-feedant effects of HCA esters with expression of Bt proteins in transformed plants. The study observed that although resistance was naturally conferred by HCA acids, and deterred oviposition by adults, these compounds were restricted to the roots and did not protect against the larvae, which feed in the cortex, causing the greatest damage. The authors evaluated the bioactivity of Cry7Aa1 protein and HCA esters both individually and in combination for causing mortality against SPW larvae. Larval mortality proved to be higher when a combination of HCA esters and Bt protein was used under laboratory bioassay conditions. Further research was recommended to establish the effect of the combination under field conditions.
13. Future Prospects
Further research on genetic transformation and genome editing: The evaluation of more transgenic events carrying cry genes would increase the likelihood of successfully identifying those with higher cry protein expression, which could potentially provide effective weevil control. Mitigating any existing bottlenecks in genetic transformation could provide a major breakthrough in genetic improvement of sweetpotato. The use of RNAi technology that has been tested on the African sweetpotato weevil species [82,83]offers a future potential for managing this pest in Uganda. Nevertheless, it is time to apply modern approaches such as gene editing in sweetpotato for sweetpotato weevil management.
Improved phenotypic strategies and analytics: As sequencing and genotyping costs continue to drop, phenotyping has been highlighted as one of the most expensive and major limitations to the accurate mapping of traits in plant breeding [84]. Novel next generation phenotyping platforms that facilitate accurate, low cost and timely acquisition of phenotypic data [85] will need to be developed and utilized for enhancing the quality of phenotypic data collected in Uganda and the region. High-throughput, near-infrared reflectance spectroscopy (NIRS) analysis of root chemistry composition, which has been implemented in many breeding programs [84], will need to be adopted and scaled out in the SSA region. Many imaging platforms and computer software have been developed for automated screening of attributes such as root and leaf architecture [86,87,88]. However, most of these modern phenotyping platforms have not been implemented in sweetpotato research in Uganda, where SPW severity is currently measured by visual assessment of damage symptoms on roots [89]. Additionally, the application of modern analytics is vital to maximizing phenotypic data outputs. For instance, the use of modern experimental designs such as the row column designs, which cater for more replications for field phenotyping, and analysis of data using mixed models would enhance heritability values for the genomic analyses.
Advances in plant bio-chemistry: Advances will need to be made in the use of plant biochemistry for revealing the basis of pest resistance in sweetpotato. Although plant biochemistry is now being used for screening for SPW resistance in Uganda breeding program, low-cost analysis throughput platforms such as near-infrared spectroscopy (NIRS) are needed since chemical profiling of resistance by wet chemistry platforms such as liquid-chromatography mass spectrometry (LC-MS) are still expensive for the breeders who screen large numbers of clones at the various breeding stage gates. The calibration of NIRS for analysis of total HCA ester concentrations in root samples would enhance throughput screening of breeding materials for population improvement [17]. This could be plausible considering that a similar method, fourier-infrared spectroscopy, has previously successfully been used for quantification of total HCA esters in forages [90].
Advances in genomics: It’s currently more important than ever that more molecular markers, particularly SNPs identified through NGS platforms since this would expedite genomic breeding for weevil resistance and other key traits in Uganda. This has been enabled by recent research advances such as the published genome sequence of diploid I. trifida, the closest relative and putative progenitor of sweetpotato [71] . Ultimately, the completion of the ongoing whole genome sequencing of cultivated hexaploid sweetpotato in the near future will further enable maximum exploitation of genomics-assisted improvement of this crop in Uganda and the region.
Exploring the potential of heterosis based breeding: In the past, sweetpotato breeding mainly relied on the ability of breeders to identify parental genotypes possessing desirable traits, and to combine these parents through hybridization schemes such as the polycross and controlled cross nurseries [48,91,92]. Preliminary studies have however shown that there are better genetic gains from heterosis based breeding compared to using the polycross [92]. Nonetheless, the potential limitation of this approach is the high level of self and cross-incompatibilities in some of the diverse parental genotypes [93] in addition to seed set abortions.
The application of molecular approaches is critical for exploiting heterosis for sweetpotato improvement. Genetic markers SNPs will be critical for diversity analysis among parental genotypes and selection of diverse parents in constituting heterotic groups for use in population improvement [94-95]. Because of self-incompatibility and severe inbreeding depression in sweetpotato, heterotic groups are currently separated based on the long-term geographic adaptations of breeding parental genotypes such as the African, Asian, and South and North American heterotic groups [48]. The application of heterosis based breeding, through selecting and genotyping the best performing SPW resistant clones and subsequently utilizing the diverse and superior parents for the next cycle of population improvement, would minimize chances of recombining closely related parents, eventually leading to higher genetic gains.
14. Conclusions
Host plant resistance is the most effective approach for sweetpotato weevil management but to date, the development of sweetpotato weevil resistant varieties has not been so successful in Uganda. Nevertheless, some progress has been made in the past decades in the development of moderately resistant varieties being produced by the farmers in Uganda and the region. The underlying genetic and bio-chemical basis for resistance has been understood to some extent and molecular markers such as SNPs have been developed still not being fully utilized in breeding. More effort will be needed by sweetpotato breeders to develop more markers for mapping sweetpotato weevil resistance to facilitate marker assisted selection and genomic selection. With the on-going sequencing of the hexaploid sweetpotato genome, we anticipate that genomics-assisted breeding for sweetpotato weevil resistance will be expedited in the coming years. Further, the advances in sweetpotato phenomics and biochemistry globally will eventually pave way for the use of novel approaches for future improvement of sweetpotato for weevil resistance with resultant accelerated genetic gains for this trait in Uganda and the SSA region.
Author Contributions
Conceptualization, I.M. and B.Y.; writing—original draft preparation, I.M.; P.M., writing—review and editing, I.M., B.Y., D.M.C., B.M.O., R.T.S., M.A.O., F.O and A.K., project administration, B.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Bill & Melinda Gates Foundation (BMGF) through the PEARL I: Biochemistry-Based Selection and Development of Nutrient Rich, Weevil Resistant Sweetpotato Varieties in Uganda Sweetpotato project (Contract ID: OPP1112515).
Data Availability Statement
The raw data sets of the reviewed papers in this article are available for public and can be accessed online at: https://sweetpotatobase.org/.
Acknowledgments
The authors thank all the partners that have worked over the years in sweetpotato improvement in Uganda and SSA; International Potato Center (CIP), North Carolina State University (NCSU), and the National Crops Resources Research Institute for their contribution to the research reviewed in this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAOSTAT Food and Agricultural Organization of the United Nations. Statistical Database. Rome, FAO. 2023.
- Woolfe J.A Sweet Potato-Past and Present. In Sweet potato: an untapped food resource; Cambridge University Press: Cambridge, Great Britain, 1992.
- Anyanga, M.O.; Yada, B.; Yencho, G.C.; Ssemakula, G.N.; Alajo, A.; Farman, D.I.; Mwanga, R.O.M.; Stevenson, P.C. Segregation of Hydroxycinnamic Acid Esters Mediating Sweetpotato Weevil Resistance in Storage Roots of Sweetpotato. Front. Plant Sci. 2017, 8, 1011. https://doi.org/10.3389/fpls.2017.01011. [CrossRef]
- USDA National Nutrient Database for Standard Reference 1 Release April, 2018; United States Department of Agriculture, 2018;
- Yanggen, D; Nagujja, S The Use of Orange-Fleshed Sweetpotato to Combat Vitamin A Deficiency in Uganda. A Study of Varietal Preferences, Extension Strategies and Post-Harvest Utilization; International Potato Center (CIP): Lima (Peru). International Potato Center (CIP), 2006;
- Hotz, C.; Loechl, C.; Lubowa, A.; Tumwine, J.K.; Ndeezi, G.; Nandutu Masawi, A.; Baingana, R.; Carriquiry, A.; de Brauw, A.; Meenakshi, J.V.; et al. Introduction of β-Carotene–Rich Orange Sweet Potato in Rural Uganda Resulted in Increased Vitamin A Intakes among Children and Women and Improved Vitamin A Status among Children. The Journal of Nutrition 2012, 142, 1871–1880. https://doi.org/10.3945/jn.111.151829. [CrossRef]
- Rukarwa, R.J; Prentice, K.,; Ormachea, M.,; Kreuze, J.F.,; Tovar, J.,; Mukasa, S.B.,; Ssemakula, G.,; Mwanga, R.O.M.; Ghislain, M. Evaluation of Bioassays for Testing Bt Sweetpotato Events against Sweetpotato Weevils. Afr Crop Sci J 2013, 21, 235–244.
- Sorensen, K.A. Sweetpotato Insects: Identification, Biology and Management. In The Sweetpotato; Springer-Verlag New York Inc.: New York, 2009.
- Fuglie, K.O. Priorities for Sweetpotato Research in Developing Countries: Results of a Survey. horts 2007, 42, 1200–1206. https://doi.org/10.21273/HORTSCI.42.5.1200. [CrossRef]
- Capinera, J. Sweetpotato Weevil; Department of entomology and nematology. University of Florida, 2006;
- Stathers, T. E., Rees, D., Nyango, A., Kiozya, H., Mbilinyi, L., Jeremiah, S., Kabi, S. and Smit, N. Sweetpotato Infestation by Cylas Spp. in East Africa: II. Investigating the Role of Root Characteristics. International Journal of Pest Management 2003, 49, 141–146. [CrossRef]
- Musana, P; Okonya, J. S.; Mujica, N; Carhuapoma, P; Kroschel, J. Sweetpotato Weevil, Cylas Brunneus (Fabricius). In Pest Distribution and Risk Atlas for Africa. Potential Global and Regional Distribution and Abundance of Agricultural and Horticultural Pests and Associated Biocontrol Agents Under Current and Future Climates; International Potato Center (CIP)): eds. J. Kroschel, N. Mujica, P. Carhuapoma and M. Sporleder (Lima (Peru), 2016; pp. 63–73.
- Okonya, J. S.; Mujica, N.; Carhuapoma, P; Kroschel, J Sweetpotato Weevil, Cylas Puncticollis (Boheman 1883). In Pest Distribution and Risk Atlas for Africa. Potential Global and regional Distribution and Abundance of Agricultural and Horticultural Pests and Associated Biocontrol Agents Under Current and Future Climates; eds. J. Kroschel, N. Mujica, P. Carhuapoma and M. Sporleder (Lima (Peru), 2016; pp. 54–63.
- Ssali, R.T.; Sseruwu, G.; Yada, B.; Ssemakula, G.; Wasonga, C.; Grüneberg, W.J.; Eyzaguirre, R.; Low, J.W.; Mwanga, R.O.M. Efficiency of the Polycross and Controlled Hybridization Methods in Sweetpotato Breeding in Uganda. Journal of Agricultural Science (Toronto, Ont.) 2019, 11, 123. https://doi.org/10.5539/JAS.V11N17P123. [CrossRef]
- Stevenson, P.C.; Muyinza, H.; Hall, D.R.; Porter, E.A.; Farman, D.I.; Talwana, H.; Mwanga, R.O.M. Chemical Basis for Resistance in Sweetpotato Ipomoea Batatas to the Sweetpotato Weevil Cylas Puncticollis. Pure and Applied Chemistry 2009, 81, 141–151. https://doi.org/10.1351/PAC-CON-08-02-10. [CrossRef]
- Anyanga, M.O.; Muyinza, H.; Talwana, H.; Hall, D.R.; Farman, D.I.; Ssemakula, G.N.; Mwanga, R.O.M.; Stevenson, P.C. Resistance to the Weevils Cylas Puncticollis and Cylas Brunneus Conferred by Sweetpotato Root Surface Compounds. J. Agric. Food Chem. 2013, 61, 8141–8147. https://doi.org/10.1021/jf4024992. [CrossRef]
- Yada, B.; Alajo, A.; Ssemakula, G.N.; Brown-Guedira, G.; Otema, M.A.; Stevenson, P.C.; Mwanga, R.O.M.; Craig Yencho, G. Identification of Simple Sequence Repeat Markers for Sweetpotato Weevil Resistance. Euphytica 2017a, 213, 129. https://doi.org/10.1007/s10681-017-1917-1. [CrossRef]
- Wolfe, G.W. The Origin and Dispersal of the Pest Species of Cylas with a Key to the Pest Species Groups of the World. In: Jansson, R.K. AndRaman, K.V. (Eds.). In Sweet Potato Pest Management. A Global Perspective; Westview Press: Boulder, Colorado, 1991.
- Chalfant, R.; Jansson, R; Dakshina, R.; Schalk, J. Ecology and Management of Sweetpotato Insects. Annu. Rev. Entomol 1990, 17–180, doi:doi: 10.1146/ annurev.en.35.010190.001105. [CrossRef]
- Ames, T; Smit, N.E.J.M.; Braun, A.R; O’Sullivan, J.N.; Skoglund, L.G. Sweetpotato: Major Pests, Diseases, and Nutritional Disorders; International Potato Center (CIP): Lima, Peru., 1996.
- Nderitu, J.; Sila, M; Nyamasyo, G.; Kasina, M Effectiveness of Entomopathogenic Nematodes against Sweet Potato Weevil (Cylas Puncticollis Boheman (Coleoptera: Apionidae)] under Semi-Field Conditions in Kenya. J Entomol 2009, 6, 145–154. [CrossRef]
- Abidin, P.E. Sweetpotato Breeding for North Eastern Uganda: Farmer Varieties, Farmer-Articipatory Selection, and Stability of Performance. Ph.D. thesis, Wageningen University: Wageningen, Netherlands, 2004.
- Muyinza, H.; Talwana, H.L.; Mwanga, R.O.M.; Stevenson, P.C. Sweetpotato Weevil ( Cylas Spp.) Resistance in African Sweetpotato Germplasm. International Journal of Pest Management 2012, 58, 73–81. https://doi.org/10.1080/09670874.2012.655701. [CrossRef]
- Smit, N.E.J.M.; Matengo, L.O. Farmers’ Cultural Practices and Their Effects on Pest Control in Sweetpotato in South Nyanza, Kenya. International Journal of Pest Management 1995, 41, 2–7. https://doi.org/10.1080/09670879509371912. [CrossRef]
- Smit, N. The Effect of the Indigenous Cultural Practices of In-Ground Storage and Piecemeal Harvesting of Sweet Potato on Yield and Quality Losses Caused by Sweet Potato Weevil in Uganda. Agric Ecosyst Environ 1997, 191–200. [CrossRef]
- Smit, N. E. J. M.; Van Huis, A. Biology of the African Sweetpotato Weevil Species Cylas Puncticollis (Boheman) and C. Brunneus (Fabricius) (Coleoptera: Apionidae). Int. J. Trop. Insect Sci. 1998, 18, 93–100. [CrossRef]
- Stathers, T; Namanda, S; Mwanga, R. O. M; Khissa, G; Kapinga, R Manual for Sweetpotato Integrated Production and Pest Management Farmer Field Schools in Sub-Saharan Africa 2005.
- Smit, N.E.J.M.; Downham, M.C.A.; Laboke, P.O.; Hall, D.R.; Odongo, B. Mass-Trapping Male Cylas Spp. with Sex Pheromones: A Potential IPM Component in Sweetpotato Production in Uganda. Crop Protection 2001, 20, 643–651. https://doi.org/10.1016/S0261-2194(01)00026-6. [CrossRef]
- Talekar, N. S. Insect Factors in Breeding and Cultivation of Sweet Potato. In Sweet Potato Technology for the 21st Century. eds. W. Hill, C. Bonsai and P. A. Loretan; Tuskegee University: Tuskegee, USA, 1992.
- AVRDC Asian Vegetable Research and Development Center. Sweet Potato Breeding and Sweet Potato Entomology; 1987; pp. 163–198;
- Ebregt, E.; Struik, P.C.; Odongo, B.; Abidin, P.E. Pest Damage in Sweet Potato, Groundnut and Maize in North-Eastern Uganda with Special Reference to Damage by Millipedes (Diplopoda). J Life Sci. 2005, 53, 49–69. [CrossRef]
- Otto, N; Russel, M; Eric, C Sweetpotato Weevil. A Review of Recent Management Advances and Appraisal of Previous Research in Papua.; 2006;
- Allard, G. B. Integrated Control of Arthropod Pests of Root Crops; CAB International Institute of Biological Control: mid-term Report-November 1988-December 1989: Nairobi, Kenya, 1990;
- Hwang, J. S Integrated Control of Sweetpotato Weevil, Cylas Formicarius Fabricius, with Sex Pheromone and Insecticide 2000.
- Bassey, E. E. Field Evaluation of Yield and Resistances of Local and Improved Sweet Potato (Ipomoea Batatas (L) Lam) Accessions to Cylas Puncticollis and Meloidogyne Incognita in Southeastern Nigeria. Asian J. Agric. Sci. 2012.
- Jansson, R.K. Biological Control of Cylas Spp. In Jansson, R.K. and Raman, K.V. (eds.) Sweet Potato Pest Management: A Global Perspective.; Westview Press Inc.: Boulder, Colorado, 1991.
- Ondiaka, S.; Maniania, N.K.; Nyamasyo, G.H.N.; Nderitu, J.H. Virulence of the Entomopathogenic Fungi Beauveria Bassiana and Metarhizium Anisopliae to Sweet Potato Weevil Cylas Puncticollis and Effects on Fecundity and Egg Viability. Annals of Applied Biology 2008, 153, 41–48. https://doi.org/10.1111/j.1744-7348.2008.00236.x. [CrossRef]
- Kaya, H.K. Soil Ecology. In: Gaugler, R. and Kaya, H.K. (Eds.). In Entomopathogenic nematodes in biological control; CRC Press: Boca Raton, FL, 1990.
- Kaur, S. Molecular Approaches towards Development of Novel Bacillus Thuringiensis Biopesticides. World Journal of Microbiology and Biotechnology 2000, 16, 781–793. https://doi.org/10.1023/A:1008931207374. [CrossRef]
- Lacey, L.A; Frutos, R.; Kaya, H.K.; Vail, P. Insect Pathogens as Biological Control Agents: Do They Have a Future? Biological Control 2001, 21, 230–248. [CrossRef]
- Stathers, T.E.; Rees, D.; Kabi, S.; Mbilinyi, L.; Smit, N.; Kiozya, H.; Jeremiah, S.; Nyango, A.; Jeffries, D. Sweetpotato Infestation by Cylas Spp. in East Africa: I. Cultivar Differences in Field Infestation and the Role of Plant Factors. International Journal of Pest Management 2003, 49, 131–140. https://doi.org/10.1080/0967087021000043085. [CrossRef]
- Skoglund, L.G.; Smit, N.E. Major Diseases and Pests of Sweetpotato in Eastern Africa; International Potato Center (CIP): Peru, 1994; p. 67;
- Kreuze, K.F; Valkonen, J.P.T.; Ghislain, M. Genetic Engineering In: Loebenstein, G. and Thottappilly, G. (Eds.). In The Sweetpotato; Springer-Verlag New York Inc.: New York, 2009.
- Magira, P. Evaluation of Sweetpotato Clones from International Potato Center (CIP) for Resistance to the Sweetpotato Weevils, Cylas Puncticollis and C. Brunneus (Coleoptera: Curculionidae). MSc. Dissertation, Makerere University, 2003.
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivity of Phenolic Acids: Metabolites versus Parent Compounds: A Review. Food Chemistry 2015, 173, 501–513. https://doi.org/10.1016/j.foodchem.2014.10.057. [CrossRef]
- Lin, K.H; Lai, Y.C; Chang, K.Y; Chen, Y. F; Hwang, S.Y.; Lo, H.F Improving Breeding Efficiency for Quality and Yield of Sweetpotato. Bot. Stud. 2007, 283–292.
- Cervantes-Flores, J.C.; Sosinski, B.; Pecota, K.V.; Mwanga, R.O.M.; Catignani, G.L.; Truong, V.D.; Watkins, R.H.; Ulmer, M.R.; Yencho, G.C. Identification of Quantitative Trait Loci for Dry-Matter, Starch, and β-Carotene Content in Sweetpotato. Mol Breeding 2011, 28, 201–216. https://doi.org/10.1007/s11032-010-9474-5. [CrossRef]
- Grüneberg, W.J.; Mwanga, R.O.M.; Andrade, M.; Dapaah, H. Sweetpotato Breeding. In In: Andrade M, Barker I, Cole D, Dapaah H, Elliott H, Fuentes S, Gruneberg WJ, Kapinga R, Kroschel J, Labarta R, Lemaga B, Loechl C, Low J, Lynam J, Mwanga R, Ortiz O, Oswald A, Thiele G (eds). Unleashing the potential of sweetpotato in sub-Saharan Africa: current challenges and way forward Nairobi, CIP SSA; 2009.
- Mwanga, R.; Odongo, B.; Niringiye, C.; Kapinga, R.; Tumwegamire, S.; Abidin, P.; Carey, E.; Lemaga, B.; Nsumba, J.; Zhang, D. Sweetpotato Selection Releases: Lessons Learnt from Uganda. African Crop Science Journal 2010, 15. https://doi.org/10.4314/acsj.v15i1.54406. [CrossRef]
- Osaru, F.; Karungi, J.; Odama, R.; Chelangat, D.M.; Musana, P.; Otema, M.A.; Oloka, B.; Gibson, P.; Edema, R.; Ssali, R.T.; et al. Identification of the Key Morphological Sweetpotato Weevil Resistance Predictors in Ugandan Sweetpotato Genotypes Using Correlation and Path-coefficient Analysis. Crop Science 2023, csc2.20915. https://doi.org/10.1002/csc2.20915. [CrossRef]
- Oloka B.M. Genetic Linkage Map Construction and QTL Analysis of Important Pest and Agronomic Traits in Two Bi-Parental Sweetpotato SNP Mapping Populations. PhD Thesis, North Carolina State University: USA, 2019.
- Mugisa, I.; Karungi, J.; Musana, P.; Odama, R.; Alajo, A.; Chelangat, D.M.; Anyanga, M.O.; Oloka, B.M.; Gonçalves dos Santos, I.; Talwana, H.; et al. Combining Ability and Heritability Analysis of Sweetpotato Weevil Resistance, Root Yield, and Dry Matter Content in Sweetpotato. Front. Plant Sci. 2022, 13, 956936. https://doi.org/10.3389/fpls.2022.956936. [CrossRef]
- Gaffney, J.; Schussler, J.; Löffler, C.; Cai, W.; Paszkiewicz, S.; Messina, C.; Groeteke, J.; Keaschall, J.; Cooper, M. Industry-Scale Evaluation of Maize Hybrids Selected for Increased Yield in Drought-Stress Conditions of the US Corn Belt. Crop Science 2015, 55, 1608–1618. https://doi.org/10.2135/cropsci2014.09.0654. [CrossRef]
- Larkin; Lozada; Mason Genomic Selection—Considerations for Successful Implementation in Wheat Breeding Programs. Agronomy 2019, 9, 479. https://doi.org/10.3390/agronomy9090479. [CrossRef]
- Chang, K; Lo, H; Lai, Y; Yao, P; Lin, K; Hwang, S Identification of Quantitative Trait Loci Associated with Yield-Related Traits in Sweetpotato (Ipomoea Batatas). Bot. Stud. 2009, 43–50.
- Yada, B Genetic Analysis of Agronomic Traits and Resistance to Sweetpotato Weevil and Sweet Potato Virus Disease in a Bi-Parental Sweetpotato Population. PhD Thesis, North Carolina State University (NCSU): USA, 2014.
- Mwanga, R.O.M; Ghislain, M; Kreuze, J; Ssemakula, G. N; Yencho, C Exploiting the Use of Biotechnology in Sweet Potato for Improved Nutrition and Food Security: Progress and Future Outlook. In AgroBiotechnology, Biosafety and Seed Systems in Developing Countries; 2011; pp. 25–31.
- Cervantes-Flores JC; Yencho GC; Kriegner A; Pecota KV; Faulk MA; Mwanga ROM; Sosinski B Development of a Genetic Linkage Map and Identification of Homologous Linkage Groups in Sweetpotato Using Multiple-Dose AFLP Markers. Mol Breeding 2008, 511–532. [CrossRef]
- Collard, B.C.Y.; Mackill, D.J. Marker-Assisted Selection: An Approach for Precision Plant Breeding in the Twenty-First Century. Phil. Trans. R. Soc. B 2008, 363, 557–572. https://doi.org/10.1098/rstb.2007.2170. [CrossRef]
- Buteler, M.I.; Jarret, R.L.; LaBonte, D.R. Sequence Characterization of Microsatellites in Diploid and Polyploid Ipomoea: Theor Appl Genet 1999, 99, 123–132. https://doi.org/10.1007/s001220051216. [CrossRef]
- Hu, J.; Nakatani, M.; Mizuno, K.; Fujimura, T. Development and Characterization of Microsatellite Markers in Sweetpotato. Breed. Sci. 2004, 54, 177–188. https://doi.org/10.1270/jsbbs.54.177. [CrossRef]
- Zhang, D.P; Cervantes, J; Huamán, Z; Ghislain, M Assessing Genetic Diversity of Sweetpotato (Ipomoea Batatas (L) Lam) Varieties from Tropical America Using AFLP. Genet Resour Crop Evol 2000, 659–665.
- Nakayama, H.; Tanaka, M.; Takahata, Y.; Matsui, K.; Iwahori, H.; Sano, Z.; Yoshinaga, M. Development of AFLP-Derived SCAR Markers Associated with Resistance to Two Races of Southern Root-Knot Nematode in Sweetpotato. Euphytica 2012, 188, 175–185. https://doi.org/10.1007/s10681-012-0678-0. [CrossRef]
- Kriegner, A.; Cervantes, J.C.; Burg, K.; Mwanga, R.O.M.; Zhang, D. A Genetic Linkage Map of Sweetpotato [Ipomoea Batatas (L.) Lam.] Based on AFLP Markers. Molecular Breeding 2003, 11, 169–185. https://doi.org/10.1023/A:1022870917230. [CrossRef]
- Oloka, B.M.; Da Silva Pereira, G.; Amankwaah, V.A.; Mollinari, M.; Pecota, K.V.; Yada, B.; Olukolu, B.A.; Zeng, Z.-B.; Craig Yencho, G. Discovery of a Major QTL for Root-Knot Nematode (Meloidogyne Incognita) Resistance in Cultivated Sweetpotato (Ipomoea Batatas). Theor Appl Genet 2021, 134, 1945–1955. https://doi.org/10.1007/s00122-021-03797-z. [CrossRef]
- Yada, B.; Brown-Guedira, G.; Alajo, A.; Ssemakula, G.N.; Mwanga, R.O.M.; Yencho, G.C. Simple Sequence Repeat Marker Analysis of Genetic Diversity among Progeny of a Biparental Mapping Population of Sweetpotato. horts 2015, 50, 1143–1147. https://doi.org/10.21273/HORTSCI.50.8.1143. [CrossRef]
- Gemenet, D.C.; Lindqvist-Kreuze, H.; De Boeck, B.; da Silva Pereira, G.; Mollinari, M.; Zeng, Z.B.; Craig Yencho, G.; Campos, H. Sequencing Depth and Genotype Quality: Accuracy and Breeding Operation Considerations for Genomic Selection Applications in Autopolyploid Crops. Theoretical and Applied Genetics 2020, 133, 3345–3363. https://doi.org/10.1007/S00122-020-03673-2/FIGURES/7. [CrossRef]
- Mahadevaiah, C.; Appunu, C.; Aitken, K.; Suresha, G.S.; Vignesh, P.; Mahadeva Swamy, H.K.; Valarmathi, R.; Hemaprabha, G.; Alagarasan, G.; Ram, B. Genomic Selection in Sugarcane: Current Status and Future Prospects. Frontiers in Plant Science 2021, 12, 708233. https://doi.org/10.3389/FPLS.2021.708233/BIBTEX. [CrossRef]
- Gezan, S.A.; Osorio, L.F.; Verma, S.; Whitaker, V.M. An Experimental Validation of Genomic Selection in Octoploid Strawberry. Horticulture Research 2017, 4. https://doi.org/10.1038/HORTRES.2016.70/42567762/41438_2017_ARTICLE_BFHORTRES201670.PDF. [CrossRef]
- Badji, A.; Machida, L.; Kwemoi, D.B.; Kumi, F.; Okii, D.; Mwila, N.; Agbahoungba, S.; Ibanda, A.; Bararyenya, A.; Nghituwamhata, S.N.; et al. Factors Influencing Genomic Prediction Accuracies of Tropical Maize Resistance to Fall Armyworm and Weevils. Plants 2021, Vol. 10, Page 29 2020, 10, 29. https://doi.org/10.3390/PLANTS10010029. [CrossRef]
- Wu, S.; Lau, K.H.; Cao, Q.; Hamilton, J.P.; Sun, H.; Zhou, C.; Eserman, L.; Gemenet, D.C.; Olukolu, B.A.; Wang, H.; et al. Genome Sequences of Two Diploid Wild Relatives of Cultivated Sweetpotato Reveal Targets for Genetic Improvement. Nat Commun 2018, 9, 4580. https://doi.org/10.1038/s41467-018-06983-8. [CrossRef]
- Komaki K; Regima HN; Katayama K; Tamiya S Morphological and RAPD Pattern Variations in Sweetpotato and Its Closely Related Species. Breeding Sci 1998, 281–286. [CrossRef]
- Huang, J.C.; Sun, M. Genetic Diversity and Relationships of Sweetpotato and Its Wild Relatives in Ipomoea Series Batatas (Convolvulaceae) as Revealed by Inter-Simple Sequence Repeat (ISSR) and Restriction Analysis of Chloroplast DNA: Theor Appl Genet 2000, 100, 1050–1060. https://doi.org/10.1007/s001220051386. [CrossRef]
- Srisuwan, S.; Sihachakr, D.; Siljak-Yakovlev, S. The Origin and Evolution of Sweet Potato (Ipomoea Batatas Lam.) and Its Wild Relatives through the Cytogenetic Approaches. Plant Science 2006, 171, 424–433. https://doi.org/10.1016/j.plantsci.2006.05.007. [CrossRef]
- Roullier, C.; Duputié, A.; Wennekes, P.; Benoit, L.; Fernández Bringas, V.M.; Rossel, G.; Tay, D.; McKey, D.; Lebot, V. Disentangling the Origins of Cultivated Sweet Potato (Ipomoea Batatas (L.) Lam.). PLoS ONE 2013, 8, e62707. https://doi.org/10.1371/journal.pone.0062707. [CrossRef]
- Yang, J.; Moeinzadeh, M.-H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W.; et al. Haplotype-Resolved Sweet Potato Genome Traces Back Its Hexaploidization History. Nature Plants 2017, 3, 696–703. https://doi.org/10.1038/s41477-017-0002-z. [CrossRef]
- Rosyara, U.R.; De Jong, W.S.; Douches, D.S.; Endelman, J.B. Software for Genome-Wide Association Studies in Autopolyploids and Its Application to Potato. The Plant Genome 2016, 9. https://doi.org/10.3835/plantgenome2015.08.0073. [CrossRef]
- Pham Bich Ngoc; Vu Thi Lan; Tran Thu Trang; Nguyen Hoai Thuong; Le Thu Ngoc; Chu Hoang Ha; Le Tran Binh Agrobacterium-Mediated Transformation of Cry8db Gene in Vietnam Sweet Potato Cultivar. JLS 2015, 10. https://doi.org/10.17265/1934-7391/2015.06.002. [CrossRef]
- Sefasi, A; Ssemakula, G; Ghislain, M; Prentice, K; Kiggundu, A; Mwanga, R.O.M Transient Expression of β -Glucoronidase in Recalcitrant Ugandan Sweetpotato and Putative Transformation with Two Cry Genes. Afr. Crop Sci. J. 2014, 215–227.
- Ekobu, M.; Solera, M.; Kyamanywa, S.; Mwanga, R.O.M.; Odongo, B.; Ghislain, M.; Moar, W.J. Toxicity of Seven Bacillus Thuringiensis Cry Proteins Against Cylas Puncticollis and Cylas Brunneus (Coleoptera: Brentidae) Using a Novel Artificial Diet. ec 2010, 103, 1493–1502. https://doi.org/10.1603/EC09432. [CrossRef]
- Anyanga, M.O.; Farman, D.I.; Ssemakula, G.N.; Mwanga, R.O.M.; Stevenson, P.C. Effects of Hydroxycinnamic Acid Esters on Sweetpotato Weevil Feeding and Oviposition and Interactions with Bacillus Thuringiensis Proteins. J Pest Sci 2021, 94, 783–794. https://doi.org/10.1007/s10340-020-01297-5. [CrossRef]
- Prentice, K.; Pertry, I.; Christiaens, O.; Bauters, L.; Bailey, A.; Niblett, C.; Ghislain, M.; Gheysen, G.; Smagghe, G. Transcriptome Analysis and Systemic RNAi Response in the African Sweetpotato Weevil (Cylas Puncticollis, Coleoptera, Brentidae). PLoS ONE 2015, 10, e0115336. https://doi.org/10.1371/journal.pone.0115336. [CrossRef]
- Christiaens, O.; Prentice, K.; Pertry, I.; Ghislain, M.; Bailey, A.; Niblett, C.; Gheysen, G.; Smagghe, G. RNA Interference: A Promising Biopesticide Strategy against the African Sweetpotato Weevil Cylas Brunneus. Sci Rep 2016, 6, 38836. https://doi.org/10.1038/srep38836. [CrossRef]
- Cobb, J.N.; DeClerck, G.; Greenberg, A.; Clark, R.; McCouch, S. Next-Generation Phenotyping: Requirements and Strategies for Enhancing Our Understanding of Genotype–Phenotype Relationships and Its Relevance to Crop Improvement. Theor Appl Genet 2013, 126, 867–887. https://doi.org/10.1007/s00122-013-2066-0. [CrossRef]
- Cabrera-Bosquet, L.; Crossa, J.; Von Zitzewitz, J.; Serret, M.D.; Luis Araus, J. High-Throughput Phenotyping and Genomic Selection: The Frontiers of Crop Breeding ConvergeF: High-Throughput Phenotyping and Genomic Selection. Journal of Integrative Plant Biology 2012, 54, 312–320. https://doi.org/10.1111/j.1744-7909.2012.01116.x. [CrossRef]
- Armengaud, P.; Zambaux, K.; Hills, A.; Sulpice, R.; Pattison, R.J.; Blatt, M.R.; Amtmann, A. EZ-Rhizo: Integrated Software for the Fast and Accurate Measurement of Root System Architecture. The Plant Journal 2009, 57, 945–956. https://doi.org/10.1111/j.1365-313X.2008.03739.x. [CrossRef]
- Backhaus, A.; Kuwabara, A.; Bauch, M.; Monk, N.; Sanguinetti, G.; Fleming, A. leafprocessor : A New Leaf Phenotyping Tool Using Contour Bending Energy and Shape Cluster Analysis. New Phytologist 2010, 187, 251–261. https://doi.org/10.1111/j.1469-8137.2010.03266.x. [CrossRef]
- Clark, R.T.; MacCurdy, R.B.; Jung, J.K.; Shaff, J.E.; McCouch, S.R.; Aneshansley, D.J.; Kochian, L.V. Three-Dimensional Root Phenotyping with a Novel Imaging and Software Platform. Plant Physiology 2011, 156, 455–465. https://doi.org/10.1104/pp.110.169102. [CrossRef]
- Yada, B.; Tukamuhabwa, P.; Alajo, A.; Mwanga, R.O.M. Field Evaluation of Ugandan Sweetpotato Germplasm for Yield, Dry Matter and Disease Resistance. South African Journal of Plant and Soil 2011, 28, 142–146. https://doi.org/10.1080/02571862.2011.10640026. [CrossRef]
- Allison, G.G.; Thain, S.C.; Morris, P.; Morris, C.; Hawkins, S.; Hauck, B.; Barraclough, T.; Yates, N.; Shield, I.; Bridgwater, A.V.; et al. Quantification of Hydroxycinnamic Acids and Lignin in Perennial Forage and Energy Grasses by Fourier-Transform Infrared Spectroscopy and Partial Least Squares Regression. Bioresource Technology 2009, 100, 1252–1261. https://doi.org/10.1016/j.biortech.2008.07.043. [CrossRef]
- Yencho, G.C.; Pecota, K.V.; Schultheis, J.R.; VanEsbroeck, Z.-P.; Holmes, G.J.; Little, B.E.; Thornton, A.C.; Truong, V.-D. ‘Covington’ Sweetpotato. horts 2008, 43, 1911–1914. https://doi.org/10.21273/HORTSCI.43.6.1911. [CrossRef]
- Grüneberg, W.J.; Ma, D.; Mwanga, R.O.M.; Carey, E.E.; Huamani, K.; Diaz, F.; Eyzaguirre, R.; Guaf, E.; Jusuf, M.; Karuniawan, A.; et al. Advances in Sweetpotato Breeding from 1992 to 2012. In Potato and sweetpotato in Africa: transforming the value chains for food and nutrition security; Low, J., Nyongesa, M., Quinn, S., Parker, M., Eds.; CABI: UK, 2015; pp. 3–68 ISBN 978-1-78064-420-2.
- Gurmu, F; Hussein, S; Laing, M Self-and Cross Incompatibilities in Sweetpotato and Their Implications on Breeding. Aust. J. Crop Sci 2013, 7, 2074–2078.
- Buteler, M.I; LaBonte, D.R.; Jarret, R.L.; Macchiavelli, R .E. Microsatellite-Based Paternity Analysis in Polyploidy Sweetpotato. Journal of the American Society for Horticultural Science 2002, 392–396.
- Jarret RL; Bowen N Simple Sequence Repeats (SSRs) for Sweetpotato Germplasm Characterization. Plant Genet Res Newslett 1994, 9–11.
- Grüneberg WJ, Eyzaguirre R, Espinoza J, Mwanga ROM, Andrade M, Dapaah H, Tumwegamire S, Agili S, Ndingo-Chipungu FP, Attaluri S, Kapinga R, Nguyen T, Kaiyung X, Tjintokohadi K, Carey EE, Low J (2010) Procedures for the evaluation and analysis of sweetpotato trials. International Potato Center, Lima, Peru.
- Mwanga, R.O.M; Odongo, B.; Ocitti p’Obwoya, C.; Gibson, R.W.; Smit, N.E.J.M.; Carey. E.E. Release of five sweetpotato cultivars in Uganda. HortScience 2001, 36, 385-386, https://doi.org/10.21273/HORTSCI.36.2.385. [CrossRef]
- Mwanga, R.O.M; Odongo, B.; Turyamureeba, G.; Alajo, A.; Yencho, G.C.; Gibson, R.W.; Smit, N.E.J.M.; Carey, E.E. Release of six sweetpotato cultivars (‘NASPOT 1 to NASPOT 6’) in Uganda. HortScience 2003, 38, 475-476, https://doi.org/10.21273/HORTSCI.38.3.475. [CrossRef]
- Mwanga, R.O.M; Odongo, B.; Niringiye, C.; Alajo, A.; Abidin, P.E.; Kapinga, R.; Tumwegamire, S.; Lemaga, B.; Nsumba.; S; Carey, E.E. Release of two orange-fleshed sweetpotato cultivars, ‘SPK004’ (‘Kakamega’) and ‘Ejumula’ in Uganda. HortScience 2007, 42, 1728-1730. [CrossRef]
- Mwanga, R.O.M; Odongo, B.; Niringiye, C.N.; Alajo, A.; Kigozi, B.; Makumbi, R.; Lugwana, E.; Namakula, J.; Mpembe, I.; Kapinga, R.; Lemaga, B.; Nsumba, J.; Tumwegamire, S.; Yencho. C.G. ‘NASPOT 7’, ‘NASPOT 8’, ‘NASPOT 9 O’, ‘NASPOT 10 O’, and ‘Dimbuka-Bukulula’ Sweetpotato. HortScience 2009, 44, 828-832, https://doi.org/10.21273/HORTSCI.44.3.828. [CrossRef]
- Mwanga, R.O.M; Niringiye, C.; Alajo, A.; Kigozi, B.; Namakula, J.; Mpembe, I.; Tumwegamire, S.; Gibson, R.W.; Yencho, C.G. ‘NASPOT 11’, a sweetpotato cultivar bred by a participatory plant-breeding approach in Uganda. HortScience 2011, 46, 317-321. [CrossRef]
- Mwanga, R.O.M; Kyalo, G.; Ssemakula, G. N.; Niringiye, C.; Yada, B.; Otema, M. A.; Namakula, J.; Alajo, A.; Kigozi, B.; Makumbi, R.N.; Ball, A.M. ‘NASPOT 12 O’and ‘NASPOT 13 O’ Sweetpotato. HortScience 2016,.51, 291-295, https://doi.org/10.21273/HORTSCI.51.3.291. [CrossRef]
Figure 1.
Developmental stages of sweetpotato weevils, Cylas puncticollis (A-egg; B-larvae; C-pupa; D-adult) and Cylas brunneus (E-egg; F-larvae; G-pupa; H-adult).
Figure 1.
Developmental stages of sweetpotato weevils, Cylas puncticollis (A-egg; B-larvae; C-pupa; D-adult) and Cylas brunneus (E-egg; F-larvae; G-pupa; H-adult).
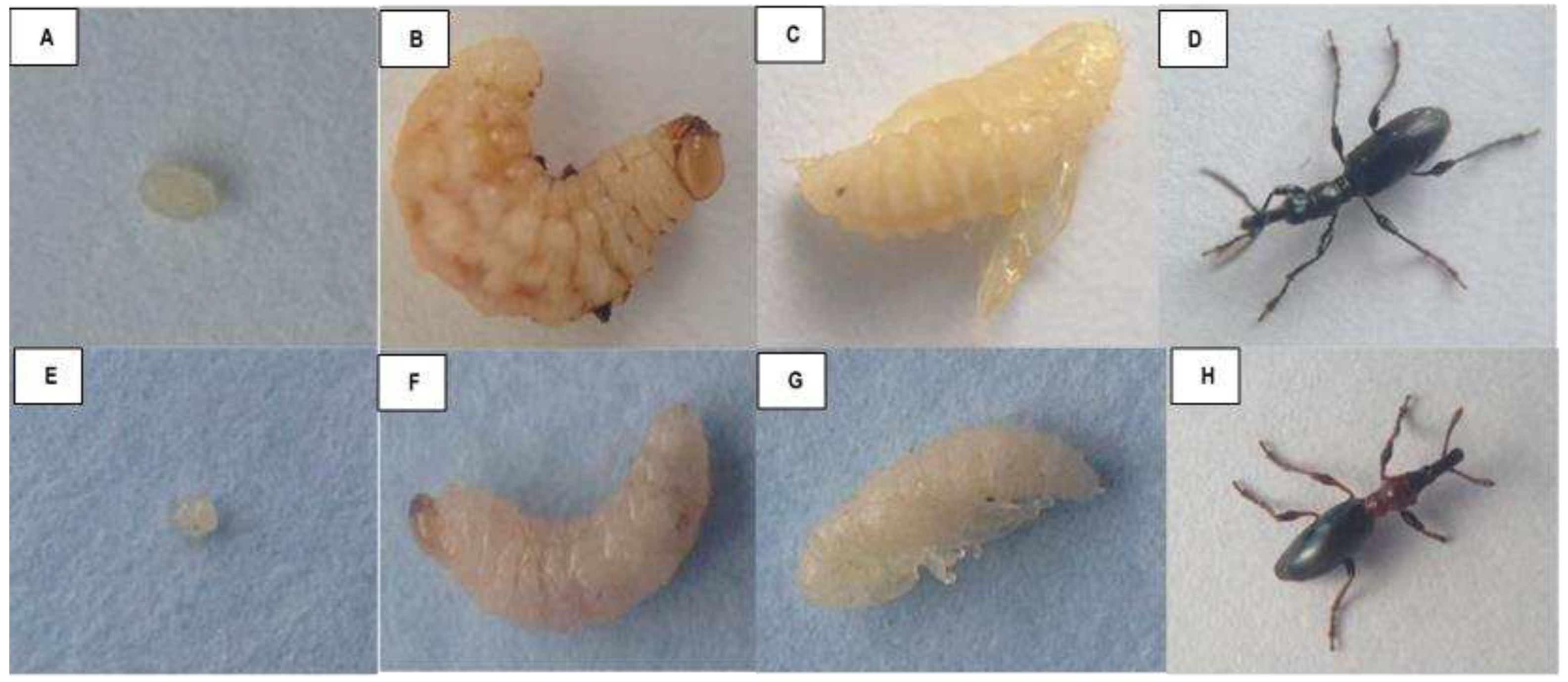
Figure 2.
Sweetpotato storage roots shortly after harvest: A- Clean (no weevil damage); B-Weevil infested (visible damage).
Figure 2.
Sweetpotato storage roots shortly after harvest: A- Clean (no weevil damage); B-Weevil infested (visible damage).


| Trait | Cylas puncticollis | Cylas brunneus |
|---|---|---|
| Morphology |
Eggs:0.45 x 0.30 mm in size Larvae:5–10 mm long Pupae:6–7 mm long; creamy-white cuticle Adults: 5–8 mm long; eyes narrowly separated |
0.7 X 0.5 mm in size 7–8 mm in length 4–5 mm long; white in colour 5–7 mm long; eyes widely separated |
| Oviposition | Females take 2–24 days before laying eggs | Females start laying eggs a day or so after becoming sexually active |
| Development | Takes place between 17.5°–35°C | Is possible between 17.5°–32°C |
| Coloration | Initially creamy white, but later change to gray and black | Turn from creamy white to brown and finally black, some are bi-colored (brown and black) |
| Host range | Has a wider host range including morning glory, cotton, sesame and maize | Has a smaller host range including morning glory and water spinach |
| Distribution | Reported in 24 African countries | Recorded in 9 African countries |
| Dispersion | Adults can fly for longer distances (up to 1000m) | Adults fly for short distances (up to 80m) |
Table 2.
An overview of the management and control strategies for Cylas species.
|
Cultural practices Use of clean planting material, crop rotation [29], hilling up and mulching, removal of alternate crops and wild plants, field sanitation [27,30], use of barrier crops, intercropping, planting of new crop away from weevil-infested fields, timely or early planting and harvesting before onset of the dry season [24,31], and flooding [32] |
|
Chemical practices Insecticides as foliar sprays controls weevils to some extent [33,34]. They can reduce the populations of adult weevils, but may not adequately control immature larvae due to their cryptic nature or when they have already infested roots and vines [8]. Dipping plant material into a synthetic pesticide before planting, can delay pest infestation for several months [27]. Insecticides are expensive and at times inaccessible to growers in SSA making their use impractical and unsustainable[35] |
|
Biological Natural enemies such as ants, maggots and wasps attack weevils, but most of them seem to be ineffective at suppressing SPW populations under field conditions [25,36]. Entomopathogenic fungi such as Beauveria bassiana have successfully been used to control SPWs in combination with other control methods. They caused a reduction in feeding ability, fecundity and egg viability in C. puncticollis [37]. Entomopathogenic nematodes such as Steinernema carpocapsae and Heterorhabditis bacteriophora, have shown potential for practical biological suppression of Cylas spp but are not readily available and small-scale farmers may not have the required purchasing power [21,38]. Bacteria such as Bacillus thuringiensis Berliner (Bt), have been developed to confer inherent pest resistance against SPWs [39,40]. Bt sweetpotato events against SPWs have been tested in SSA [7]. |
|
Host Plant Resistance Involves the use of sweetpotato weevil resistant or tolerant clones [15,41]. Deep-rooting and early maturing varieties are less susceptible to infestation than shallow rooting and late maturing varieties [42].Efforts to develop SPW resistant cultivars are still ongoing in SSA Transgenic sweetpotato plants expressing the Bt cry genes are also currently in use in some countries [43]. |
Table 3.
Reaction of sweetpotato varieties released in Uganda to Cylas spp.
| Year of release | Variety | Reaction to SPW |
| 1995 | “Bwanjule” | MR |
| “New Kawogo” | MR | |
| “Sowola” | MR | |
| “Tanzania” | S | |
| “Wagabolige” | MR | |
| “Tororo 3” | MR | |
| 1999 | NASPOT 1 | S |
| NASPOT 2 | S | |
| NASPOT 3 | MR | |
| NASPOT 4 | MR | |
| NASPOT 5 | MR | |
| NASPOT 6 | MR | |
| 2004 | “Ejumula” | S |
| SPK004 (Kakamega) | S | |
| 2007 | NASPOT 7 | S |
| NASPOT 8 | S | |
| NASPOT 9 O | S | |
| NASPOT 10 O | S | |
| “Dimbuka Bukulula” | S | |
| 2010 | NASPOT 11 | S |
| 2013 | NASPOT 12 O | S |
| NASPOT 13 O | S | |
| 2017 | NAROSPOT 1 | MR |
| NAROSPOT 2 | Low | |
| NAROSPOT 3 | Low | |
| NAROSPOT 4 | Low | |
| NAROSPOT 5 | Low | |
| 2023 | NAROSPOT 6 | MR |
| NAROSPOT 7 O | MR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



