
Submitted:
04 September 2023
Posted:
06 September 2023
You are already at the latest version
Abstract
Conjugative transposons in Gram-negative bacteria have a significant role in the dissemination of antibiotic-resistance-conferring genes between bacteria. This study aims to genomically characterize plasmids and conjugative transposons carrying integrons in clinical isolates of Klebsiella pneumoniae. Genetic composition of conjugative transposons and phenotypic assessment of 50 multidrug-resistant K. pneumoniae isolates from Tertiary-care Hospital (SQUH), Muscat, Oman was investigated. Horizontal transferability was investigated by filter- mating conjugation experiments. Whole genome sequencing (WGS) was performed to determine the sequence type (ST), acquired resistome and plasmidome of integron-carrying strains. Class 1 integron was detected in 96% of isolates and among integron-positive isolates, 18 stains contained variable regions. Horizontal transferability by conjugation confirmed the successful transfer of integrons between cells and WGS confirmed their presence in conjugative plasmids. Dihydrofolate reductase (dfrA14) was the most prevalent (34.8%) gene cassettes in class 1 integrons. MLST analysis detected predominantly ST-231 and ST-395. BlaOXA-232 and blaCTX-M-15 were the most frequently detected carbapenems and beta-lactamases in sequenced isolates. This study highlighted the high transmissibility of MDR-conferring conjugative plasmids in clinical isolates of K. pneumoniae. Therefore, the wise use of antibiotics and the adherence to effective infection control measures are necessary to limit further dissemination of multidrug-resistant organisms.
Keywords:
mobile genetic elements
; Anti-microbial resistance
; Klebsiella pneumoniae
; Whole genome sequencing
; integrons
; plasmids
; sequence types
; phylogenetic analysis
1. Introduction
Klebsiella pneumoniae is an opportunistic pathogen that is associated with different serious nosocomial infections including pneumonia, septicemia, meningitis and urinary tract infections (UTIs) [1,2]. To treat the infections caused by extended-spectrum beta-lactamase (ESBL)-producing K. pneumoniae, carbapenem antibiotics are the drugs of choice however they are considered as last resort antibiotics [3]. Moreover, the number of K. pneumoniae carbapenemase (KPC) enzyme producers has been increasing [4,5]. The spread of resistance determinants have been facilitated by horizontal gene transfer mechanisms mainly via mobile genetic elements (MGEs). Integrons for example, are semi-mobile platforms that recognize and capture mobile gene cassettes and transform them to usable genes by ensuring their appropriate expression [6,7,8]. They are located on different MGEs such as plasmids, transposons and, pathogenicity islands (PAIs) which enable their movement between different bacteria [9,10]. A number of studies showed that most K. pneumoniae clinical isolates carried class 1 integron, whereas, class 2 integron was only present in 1-2% of the isolates and rarely harbored class 3 integron [11]. A significant association between integron positive isolates and antibiotic resistance for some drugs were observed including piperacillin-tazobactam, ciprofloxacin, cefotaxime, and ceftazidime [12,13]. Both classes 1 and class 2 integrons have the gene cassettes encoding resistance to trimethoprim (dfr) as a predominant gene, which may be due to long-term usage of this antibiotic [14].
Class I Integron is associated with transposons derived from Tn402 which can bind in a larger transposon like Tn21. Over 80 different antibiotic-resistance-encoding gene cassettes are associated with class I integrons, but most integrons have the gene aadA which encodes streptomycin-spectinomycin resistance [6]. Moreover, class 1 integrons have 3-conserved segment which contains a sulI gene encoding resistance to sulfonamides and a qacEΔ1 gene encoding resistance to quaternary ammonium compounds [15]. In Oman, there has been an increasing concern reporting the spread of MDR K. pneunomiae and in particular KPC producers in clinical settings [16,17,18]. However, molecular charecterization in are still lacking. Therefore, we aimed in this study to genomically analyse the MDR K. pneumoniae carrying Class 1 integrons, in terms of structure, sequence types (STs), and antimicrobial phenotype and genotype. In addition, we explored the association between antimicrobial susceptibility and integron carriage in clinical isolates of K. pneumoniae. Finally, we conducted conjugation experiments to evaluate the horizontal-transfer capability which is key to predict future dissemination and implementation of intervention strategies.
2. Materials and Methods
2.1. Bacterial strains
A total of fifty (n=50: ESBL (n=27), XDR (n=21), and PDR (n=2) strains K. pneumoniae isolates in this study were collected from the Diagnostic Microbiology and Immunology Laboratory in Sultan Qaboos University Hospital (SQUH), Muscat, Sultanate of Oman between July 2019 and October 2019. The isolates were mostly from urine (n= 25), respiratory (n= 10), wound (n= 9), bloodstream (n= 4), body fluid (n=1) and biopsies (n=1). All isolates were processed as per Clinical and Laboratory Standards Institute guidelines [19]. The isolates were cultured in a selective medium (Cefuroxime Cysteine Lactose Electrolyte Deficient agar, Oxoid, Basingstoke Hampshire, UK) prior to preservation into sterile CryoBeads (Mast Diagnostics, United Kingdom) at -80°C for further analysis (Table 1).
2.2. Antimicrobial susceptibility
The antimicrobial susceptibility profiles of the K. pneumoniae isolates were carried out using two methods: disk diffusion method and BD PhoenixTM automated system (Becton Dickinson Diagnostic Systems, Sparks, MD, USA). For both methods, three to five bacterial colonies from overnight pure culture were suspended in normal saline (Fisher Chemical, United Kingdom) and adjusted to a 0.5 McFarland standard (approximately 1–2 × 108 CFU/mL) using a CrystalSpec nephelometer (BD Diagnostics, USA), according to the manufacturer's recommendations. Within 15 minutes, the suspension was spread onto Mueller Hinton Agar (MHA) surface (Oxoid, UK) and left for 1-2 minutes at room temperature to be absorbed. Within 15 minutes, the selected antibiotic disks (BioMérieux and Liofilchem, Germany) were placed on the inoculated MHA plates using sterile forceps. The plates were incubated at 37°C for 18-24 hours. The antibiotics were selected according to CLSI standard as follows: ampicillin (AMP 10mg), piperacillin/tazobactam (TZP 110mg), cefepime (FEP 30mg), cefotaxime (CTX 30mg), cefoxitin (FOX 30mg), ceftazidime (CAZ 30mg), imipenem (IPM 10mg), meropenem (MEM 10mg), amikacin (AK 30mg), gentamicin (CN 10mg), and ciprofloxacin (CIP 5mg). The E. coli (ATCC 25922) and P. aeruginosa (ATCC 27853) were used as susceptible control strains. The interpretive categories and zone diameter breakpoints, nearest whole mm for each antibiotic are listed in Table 2. The BD PhoenixTM automated system was used to test the susceptibility of colistin and tigecycline, and to confirm the antimicrobial susceptibility of other antibiotics tested by disk diffusion method. For colistin, > 1 mg/L was considered resistant according to CLSI breakpoints [19]. European Committee on Antimicrobial Susceptibility Testing (EUCAST) interpretive criteria was used for tigecycline with ≤ 0.5 mg/L considered as sensitive [20]. The ESBL production was further confirmed using the disk diffusion method. Both cefotaxime and ceftazidime alone and in combination with clavulanic acid were used. An increase in the diameter of ≥ 5 mm with the clavulanic acid compared to the antibiotic alone was interpreted as positive for ESBL production. The XDR and PDR isolates were characterized genotypically to the carbapenemase genotype level using Xpert Carba-R (Cepheid, USA).
2.3. Genomic DNA extraction and purification
The DNA was extracted using Qiagen kit (QIAamp® genomic DNA kit, Hilden, Germany) as described in the manufacturer’s instructions with a slight modification. One to four colonies were suspended in 10 ml Mueller Hinton broth (Oxoid, United Kingdom) and left overnight on a shaking incubator set to 250 rpm at 37 °C. Then, bacterial suspension was centrifuged the next day for 15 minutes at 4000 x g. The amount of 20-40mg of pelleted bacterial cells was re-suspended in a previously prepared 100 μL of 0.1mg/mL pre-lysis buffer (100 μl TE buffer and 0.1 μl RNase A 100mg/mL) (Thermo Fisher Scientific, Winsford, UK). All cells were well resuspended by pipetting up and down several times. After that, the sample was incubated again at 37°C at a 400 rpm shaking incubator for 30-60 minutes (Innova 4000, New Brunswick Scientific, Hertfordshire, UK). A volume of 1μL Proteinase K (stock concentration = 20mg/mL) + 99 μL TE were added to the sample to get a final volume of 200 μL in 1.5 ml microcentrifuge tube. Then, 200 μl of the sample was mixed with 400 μl of lysis solution and incubated at 65°C for 15 minutes using a heat block (Eppendorf ThermoStat plus, Germany). The pellet was re-suspended in 50-100 μl of nuclease-free H2O (QIAamp® genomic DNA kit, Hilden, Germany). The extracted DNA was aliquoted into 2 vials which were stored at 4°C and also -80 °C for future use. The boiling method was used to extract the DNA from transconjugant colonies in the conjugation experiment. Three to five colonies were picked from the plate and resuspended in 30 μl of nuclease-free water. Then, samples were heated at 100 °C for 10 minutes in a heat block. Cells were then pelleted by centrifuging the samples at 16000 × g for 1 minute. The supernatant was used as a template for PCR.
2.4. Polymerase Chain Reaction (PCR)
2.4.1. Detection and characterization of class 1, 2 and 3 integrons
PCR assays were performed using Go-Taq DNA polymerase (Promega Ltd., USA). The cycle conditions were adjusted depending on the gene size and the primer’s melting temperature. The detection of class 1, 2 and 3 integrons was investigated by the amplification of integrase genes intl1, intl2, and intl3, respectively. The PCR reaction mixture consists of 5ml of 5X PCR green buffer; 0.5 ml of dNTPs ( 10mM); 1ml of both forward and reverse primers (10 mM); 0.1ml of Go-Tag DNA polymerase (5m/ml); and 1.5ml of template DNA. Then the nuclease-free water was added until the total volume reached 25 ml. A reaction mixture without a DNA template was used as a negative control. The same PCR reaction mixture was used to amplify variable region genes. For the DNA that was extracted by the boiling method, Dimethyl sulphoxide (DMSO) was added to encourage annealing of the primers to the template and further enhance the amplification. Thermocycler (Eppendorf master cycler®, MA, Germany) was used for PCR amplification and the cycling conditions for Go-Tag enzyme are mentioned in Table 3. Whereas, annealing temperatures which are different for each gene depending on the primer’s melting temperature are listed in Table 4. The specific primers for detecting integrase genes, variable regions, and capsules as previously described [14].
2.4.1. PCR purification
PCR products were purified using QIAquick PCR and Gel Cleanup Kit (Qiagen, Germany). Five volumes of Buffer PB were added to 1 volume of the PCR sample and then mixed. To bind DNA, the mixture was applied to the QIAquick column and centrifuged for 30–60 seconds at 17,900 x g at room temperature. After centrifugation, the column was placed in a clean 2 ml collection tube and the tube containing the filtrate was discarded. Then, 750 ml of Buffer PE was added to the QIAquick column and centrifuged again for 30-60 seconds at the same previous conditions. Again, the column was placed in a clean 2 ml collection tube and the tube containing the filtrate was discarded. Then, the QIAquick column was centrifuged once more for 1 minute to remove the residual wash buffer. Finally, the QIAquick column was placed in a clean 1.5 ml microcentrifuge tube to which 30ml of elution buffer (10 mM Tris·Cl, pH 8.5) was added. The tube was incubated at room temperature for 1 minute and then centrifuged for 30–60 seconds at 17,900 x g. The purified DNA was analyzed on 2% agarose gel electrophoresis by adding 1 volume of loading dye to 5 volumes of purified DNA.
2.5. Gel electrophoresis
All PCR products were visualized by using 2% agarose gel electrophoresis containing 1x TBE (ThermoFisher Scientific, USA) and MIDORIGreen Direct (NIPPON-genetics, Europe). The bands were visualized by using G:Box Chemi-XR5 device (Syngene, India). DNA fragments sizes were estimated by using the GeneRuler 1 kb DNA Ladder (ThermoFisher Scientific, USA).
2.6. Whole genome DNA Sequencing
Whole-genome sequencing (WGS) in this study was performed at microbesNG by Illumina next-generation sequencing at a minimum coverage of 30x (https://microbesng.co.uk/, Birmingham, accessed on 20th July 2022). The genomic DNA was extracted and prepared and then was sent for sequencing following the protocol provided by the sequencing facility. The assembled sequences are then retrieved from the websites and analyzed accordingly.
2.7. Bioinformatics analyses
WGS data of K. pneumoniae isolates files were analyzed using the center for genomic epidemiology server (CGE) (http://cge.cbs.dtu.dk/services, accessed on 02nd July 2020). At CGE, CSI phylogeny tool [22] was used to investigate the relatedness between the strains based on Single Nucleotide Polymorphisms (SNPs) identification. Assembled genome data for all samples were blooded to the server as input. The reference genome was K. pneumoniae subsp. pneumoniae HS11286 (GeneBank: CP003200). The SNPs were localized and filtered based on the default settings which include a minimum distance of ten bases between the SNPs, a sequencing depth of ten bases, and a minimum SNPs quality of 30. In addition, the Z-score for each SNP has to be above 1.96. Then the data file was visualized and managed by using an online tool; Interactive Tree Of Life (iTOL) [23]. ResFinder tool from the GCE server was used in this study to detect the acquired antimicrobial resistance genes and their specific location on the sequence [24]. MLST and PlasmidFinder were used to detect Multilocus sequence typing (MLST) and plasmids, respectively [25,26]. Along with ResFinder tool in CGE, Comprehensive Antibiotic Resistance Database (CARD) (https://card.mcmaster.ca/home, accessed on 20th July 2020) was used to detect the presence of putative antibiotic resistance genes by using Resistance gene identifier (RGI) tool. This tool in addition, can detect known point mutations within the resistance-conferring genes. Artemis software from the Sanger institute was used for visualization of WGS features. Moreover, Genbank from National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/genbank/, accessed on 03rd August 2020) was used to search in DNA database for similar DNA sequences and Basic Local Alignment Search Tool (BLAST) was used to search for similarities between DNA sequences (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 03rd August 2020).
2.8. Conjugation experiment
Conjugation experiments were performed for K. pneumoniae strains with variable regions as donors, and E. coli HB101 as a recipient by using filter mating method [27]. To obtain the standard concentration of the required antibiotic [Amoxicillin and streptomycin (Sigma-Aldrich, USA)] solution, the powder was weighed on a calibrated analytical balance and dissolved in an appropriate solvent (as per manufacturer’s instructions). K. pneumoniae isolates were cultured in the Luria Agar (LA) (Lab M®, UK) plate with 100 mg/ml of amoxicillin. E. coli HB101 strain was cultured in the Luria agar (LA) plate with 50 mg/ml of streptomycin. Both strains were incubated overnight at 37°C. The next day, a single colony from previous plates was inoculated into 5 ml of Luria Broth (LB) (Lab M®, UK) with the same antibiotic selection and incubated overnight at 37°C on a shaking incubator. Then, 1:100 dilution of both cultures were inoculated into 5 ml fresh LB and incubated again on the shaking incubator at 37°C until the bacteria grown to their exponential phase (OD600 0.2) which was measured using Bio-Photometer (Eppendorf, Germany). After that, 1 ml of each culture was centrifuged at 8000 x g for 5 minutes in order to harvest the cells. The pellets were washed twice with 1X Phosphate Buffer Saline (PBS) (prepared from 10X PBS, pH 7.4, ThermoFisher Scientific) to remove the remaining antibiotics. Then, the pellets were re-suspended in 200 ml of PBS. Recipient, donor, and mixed bacterial suspension were spotted onto the center of 0.45 mm pore size membrane (Whatman Ltd. Sigma-Aldrich, St Louis, MO, USA) placed on LA without antibiotics. The spots were allowed to dry and then incubated overnight at 37°C. Subsequently, filter papers were washed in 1 ml 1X PBS to detach the bacterial cells. Serial dilutions up to 10-8 were performed for the donor, recipient, and mixed bacteria by using PBS as a diluent. The trans-conjugants are selected on an LA plate supplemented with both amoxicillin and streptomycin. For colony forming unit (CFU) counts, dilutions of the donor were plated on LA with 100mg/ml of amoxicillin and the recipient in 50 mg/ml of streptomycin. All plates were incubated at 37 ºC overnight. The efficiency of trans-conjugation is calculated using the following formulas:
Efficiency of transconjugation= nTransconjugants/nRecipient Recovered (CFU/ml)/
% Efficiency of transconjugation= nTransconjugants/ nRecipient Recovered (Cfu/ml) ×100
3. Results
Class 1 integron was detected in 92% of isolates (46/50) using IntI1F and IntI1R primers (Table 4). Screening of integron class 2 and 3 by detecting the presence of IntI2, and IntI3 genes, respectively, showed that these were not present in the collection (Gel electrophoresis supplementary files). The antimicrobial susceptibility patterns of 50 K. pneumoniae isolates was performed (Figure 1) . All clinical isolates were resistant to ampicillin (100%), most of the isolates were resistant to cefotaxime (98%), cefepime (92%) and ceftazidime (84%), and almost half of them showed resistance to piperacillin/tazobactam (48%). Moreover, 88% of the isolates showed resistance to ciprofloxacin (88%). Furthermore, these isolates showed intermediate resistance to gentamicin (54%), imipenem (52%), and a lesser extent to amikacin and colistin (30% and 6%), respectively.
According to the results obtained by phoenixBD semi-automated system, 54% of K. pneumoniae isolates were identified as ESBLs, whereas 42 % (n=21) of isolates were identified as XDR and two isolates (4%) as PDR. All XDRs were susceptible to colistin except 3 isolates (Kp 2, Kp 22, and Kp 50) (Table 5). In addition, 11.8% and 5.6% of the XDRs were susceptible to amikacin and gentamicin, respectively. Some of the XDR isolates were susceptible to amikacin, gentamicin, imipenem, and meropenem along with colistin (Table 5).
The conjugation was performed between variable region-positive isolates as donors and E. coli HB101 strain as a recipient. Seven representative samples were selected to be tested (Kp 5, Kp 21, Kp 27, Kp 37, Kp 42, Kp 49 and, Kp 50). The trans-conjugant PCR products are shown in (Supplementry file). All samples were positive for IntI1 gene, which confirmed the successful occurrence of conjugation via means of horizontal transfer. In the tested strains, the constituents of gene cassettes, plasmids, and sequencing types were analyzed by sequencing. WGS data showed that in our tested strains, plasmids were detected in all trans-conjugants which conforms that these conjugative plasmids are highly transmissible. The efficiency of trans-conjugation for each strain was calculated and ranged between 0.07% to 17.3% (Figure 2.). This exemplifies the role of these plasmids as the main vehicle for the transmission of integrons, and subsequently the transport of various types of gene cassettes from the donor to the recipient strains.
Out of the total 50 isolates of K. pneumoniae, 24 isolates were analysed by WGS, including the 7 isolates which were successful in conjugation. Sixteen of these isolates were positive for the variable regions (Kp 5, 7, 10, 11, 15, 16, 21, 28, 37, 40, 42, 43, 44, 45, 46, 49) and 2 isolates showed bands with unexpected size for intI3 gene (Kp 4 and KP 22). Among the 24 samples, 3 were integron-negative isolates by PCR method (Kp 4, Kp 27, and Kp 30). However, WGS showed that two of these isolates were positive for class 1 integron (Kp 27 and Kp 30), whereas Kp 4 was still negative. Therefore, Kp 4 isolate was excluded from the subsequent molecular analysis.
MLST of all sequenced isolates was determined by the online MLST database from the Center of Genomics Epidemiology (CGE) [25]. Nine of the isolates belong to sequence type (ST-231) (Kp 5, 6, 7, 10, 11, 15, 28, 30, and 45), six isolates were ST- 395 (Kp16, 22, 41, 43, 44, 50), and 2 isolates were ST-405 (Kp 42 and Kp 46). Only one isolate belongs to each of the following miscellaneous sequence types: ST- 37 (Kp 40), ST-45 (Kp 21), ST-147 (Kp 49), ST-280 (Kp 27), ST-1710 (Kp37) and ST-1741(Kp 25).
WGS data of 23 integron-positive isolates were used to construct a whole genome phylogenetic single nucleotide polymorphism (SNP) (Figure 3), and the output from the analysis showed three main clusters of strains. The first cluster is composed of 9 isolates belonging to ST-231. The second cluster of strains is in the ST-395 group (n=6). The clinical data of the above mentioned two STs, indicate frequent moving of patients from Intensive Care Unit (ICU) to the male medical ward or vice versa. However, the third cluster consisted of two strains only belonging to ST-405. The remaining strains belong to miscellaneous sequence types (ST-37, ST-147, ST-280, ST-1741, ST-45, ST-13, ST-17, and ST-10).
A close-up analysis of the two predominant STs (ST-231 and ST-395) is done to predict if there any possible recent outbreak through clustering (Figure 4.). SNP tree for ST-231 and ST-395 individually was created and it was observed that the PDR isolates (Kp6 and Kp11) were branching from the same clade suggesting possible vertical transmission among patients. Similarly, among the other ST-231 and ST-395 stains are very colsely related.
Figure 4.
SNPs trees of K. pneumoniae strains (ST-231 and ST-395).
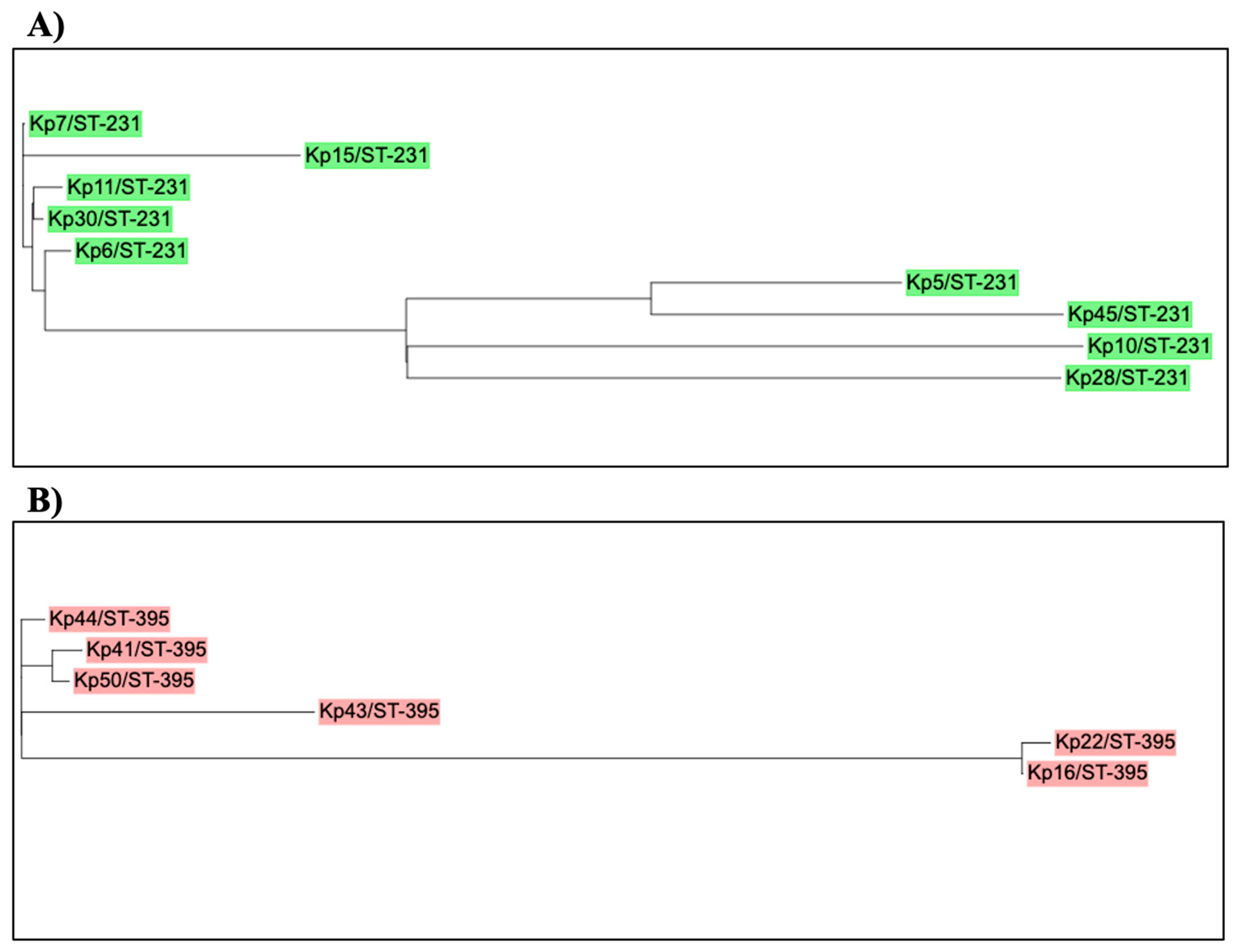
A phylogeny tree showing relatedness of K. pneumoniae strains that belonged to A) ST-231 (n=9) and, B) ST-395 (n=6) . Isolates are labeled according to their numbers and sequence type (ST). Green: ST-231 and red:ST-395. K. pneumoniae HS11286 was used as a reference isolate (accession number CP003200). The online tool (iTOL) was used to draw and edit the tree.
The demographic data for thirteen samples that belong to ST-231 (n=9) and ST-395 (n=4) which accounts for most samples were list in Table 6. Ninety-two percent of the patients were males with age more than 50 years (69.2%). The length of stay varied between patients with a range from 1 to 134 days. Most of the patients were admitted to the ICU (61.5%) and the rest of the patients were admitted to intermediate care wards (male medical wards). Highly critical ICU patients usually require broad spectrum antibiotics which exterts tremendous selection pressure, thus driving antimicrobial-resistance bacteria to thrive further in these settings.
At the level of ST, more than two-third (77.8%) of ST-231 and all ST-395 strains were isolated from patients admitted either in ICU or male medical wards or moved between the two places interchangeably. Almost all patients have various risk factors contributing to their likelihood of acquiring highly resistant K. pneumoniae strains including ICU admission, ventilation, urinary catheters, central venous catheters, and hospitalization for more than 7 days [28]. Unfortunately, the mortality rate among these patients is very high (84.6%).
All sequenced isolates carried a wide range of acquired antimicrobial resistance-conferring genes. 14 isolates harbored blaOXA-232 gene, all of which were XDR and PDR isolates except KP 7 which was the only XDR isolate that did not carry blaOXA-232 gene (Figure 3). The bla-NDM5 was detected in only one isolate (Kp 49). Based on the WGS data and Carba-R, 87% of XDR and PDR isolates (20/23) have blaOXA-48-like, 8.7% (2/23) have blaNDM, and one isolate (4.3%) has both blaNDM and blaOXA48-like genes. There was a high level of agreement between WGS and GeneXpert results for the tested samples (8/9). The blaOXA-1 allele was detected in eight isolates, most of which belong to ST-395 (n=6). The ESBL gene blaCTX-M-15 was detected in all isolates, while blaTEM-1B was seen in 15 isolates. Moreover, 10 isolates were positive for blaSHV-1, 9 isolates for blaSHV-11, 2 for blaSHV-76, and 2 for blaSHV-27.
A number of various genes conferring resistance against aminoglycosides were detected including aac(6')-Ib-cr, aac(6')-Ib Hangzhou, aadA1,2, aac(3)-lId, aph(3')-Ia, strA, strB, armA and rmtB. Fosfomycin-resistance conferring gene (fosA6) was detected in all isolates except Kp 49 which harbored fosA5. In addition, 6 different genes encoding quinolone resistance were detected in the isolates which are aac(6')-Ib-cr, oqxA, oqxB, qnrS1, B1and B66. Moreover, rifampicin (arr-2) and trimethoprim resistance encoding genes (dfr) were found in 10 and 11 isolates, respectively.
Tetracycline resistance gene, tetA, was detected in four ESBL isolates, whereas tetD in 2 XDR isolates. According to the phoenix automated system, all of these were tigecycline-resistant except one ESBL isolate that carried the tetA gene (Kp 46). The gene of acrAB efflux pumps’ regulator (marA), which contributes to the resistance against tigecycline and other antibiotic classes, was detected in all sequenced isolates. In addition, K. pneumoniae efflux pumps (Kpn E, F, G, H) that confer resistance to different antibiotic classes like macrolide, aminoglycoside, cephalosporin, tetracycline, rifamycin, and colistin, were detected in all isolates.
The four colistin-resistance isolates belonging to ST-231 and ST-395 (Kp 6, Kp 11, Kp 22, and Kp 50) were negative for plasmid-mediated mcr genes, therefore, further analysis of the SNPs in PhoPQ operons was performed. No chromosomal mutations were detected in the regulatory two-component systems (TCSs) PmrAB and crrAB, and in mgrB gene, a negative regulator gene of TCSs. However, all isolates harbored the regulatory TCSs (PhoPQ). Our analysis of the PhoPQ operons in the colistin-resistant isolates identified four novel undescribed SNP in the PhoP genes with the amino acid substitutions as follows: (Val129Glu), (Gln147His), (Gln131Glu), and (Pro129Thr) (Table 7.). The significance of these SNPs is yet to be determined as none of these substitutions have been described.
Most of the tested isolates showed an agreement between their resistance phenotypic patterns to different antibiotic families and the presence of resistance genes as shown in Table 8. Carbapenems, cephalosporin, and quinolone antibiotics showed a complete level of agreement between phenotype and genotype. However, aminoglycoside antibiotics (AK, CN) showed a low level of agreement with 21 isolates carrying the resistance genes and only 9 and 15 isolates with phenotypic resistance patterns to amikacin and gentamicin, respectively. This observation could be because these resistance-conferring genes might not be expressed. The expression of these genes needs to be further investigated which is not within the scope of this study.
Using WGS data, the variable region of 23 samples including the trans-conjugant strains (Kp 5, Kp 21, Kp 27, Kp 37, Kp 42, Kp 49 and Kp 50) were analyzed to detect gene cassettes (Table 9 and Figure 3.8). Ten different gene cassettes were identified in 16 strains with the varible region including those encoding resistance to aminoglycoside (aadA, aacA4, aac(6')-Ib , APH(3”)-Ia, ant1) trimethoprim (dfrA5, dfrA12, dfrA14), rifampin (arr2 and arr3), chloramphenicol (catB3, catB8 and cmlA1), macrolide (ereA2 and erm ) and quaternary ammonium compound (emrE). The most frequent gene cassette was composed of dfrA14 gene alone, which was present in 8 isolates (34.8%). However, 3 samples showed empty integrons (In0) without any gene cassette insertions (Kp 16, Kp 22, and Kp 25). One of them had a variable region (Kp 16), while the other two did not (Kp 22 and Kp 25).
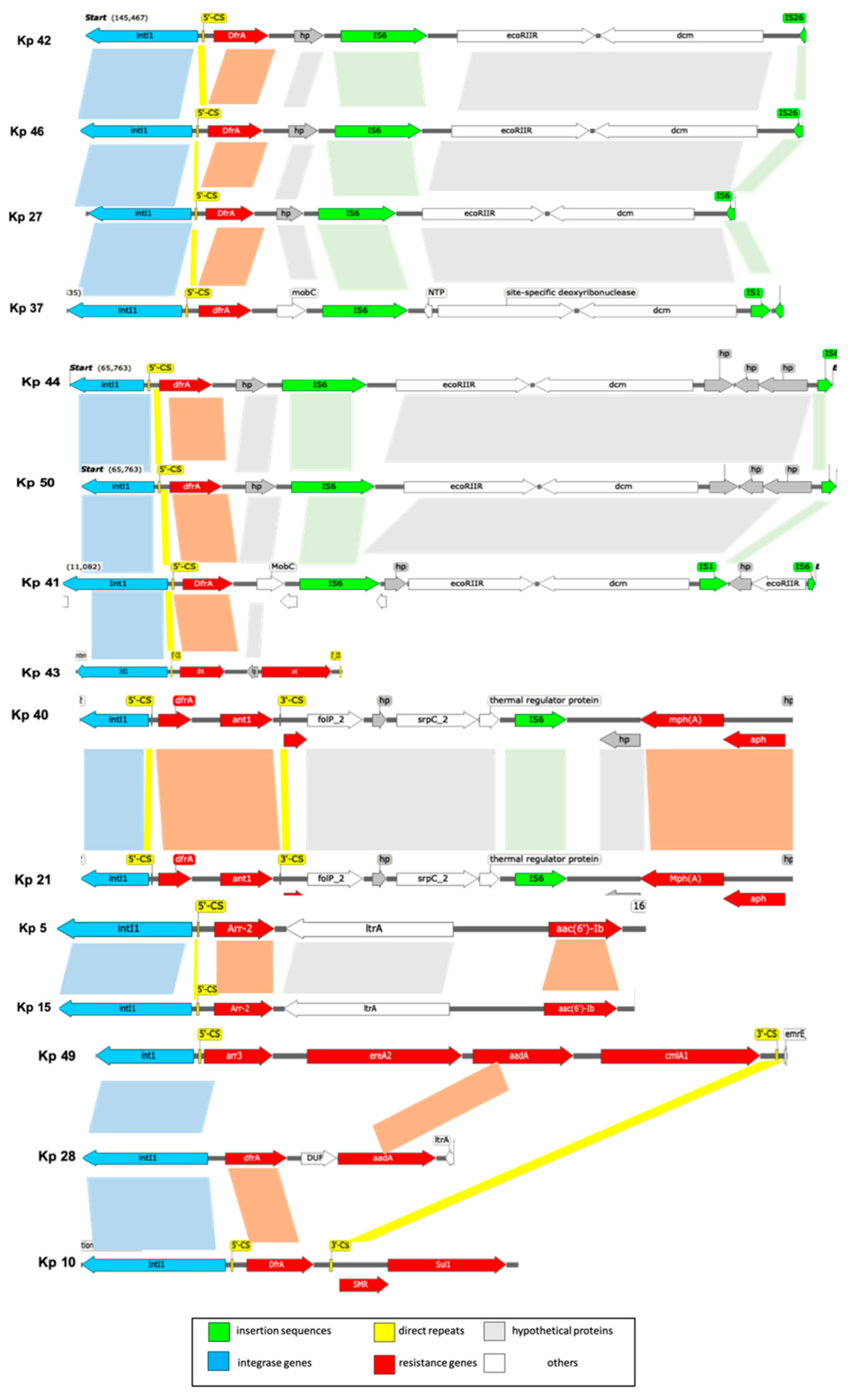
Comparative analysis of integrons was performed by locating the gene cassettes within the variable region. Mapping was done based on the genes between the primers of the variable region which varied among strains (Table 9). It was observed that there was a contig break in the middle of the integron cassettes in some strains and the downstream sequence (3’-end) was found in a separate contig. Therefore, the genetic maps of the integrons were drawn after manual reassembly of the various components (Figure 3).
Figure 4.
Schematic map of the composition of the integrons in K. pneumoniae strains. Genes and ORFs are denoted by arrowhead indicating the direction of transpcription and colored based on the gene function classifications as shown in the key in the lower part of the figure. Shaded areas denote regions of homology (>95% nucleotide sequence identity). Grouping of strains is based on the sequence similarity as follows: ST-231 strains (Kp 41, Kp 43, Kp 44, and Kp 50), (Kp 27, Kp 37, Kp 42, and Kp 46), (Kp 40, and Kp 21), (Kp 5, and Kp 15) and (Kp 49, Kp 28, and Kp10).
Figure 4.
Schematic map of the composition of the integrons in K. pneumoniae strains. Genes and ORFs are denoted by arrowhead indicating the direction of transpcription and colored based on the gene function classifications as shown in the key in the lower part of the figure. Shaded areas denote regions of homology (>95% nucleotide sequence identity). Grouping of strains is based on the sequence similarity as follows: ST-231 strains (Kp 41, Kp 43, Kp 44, and Kp 50), (Kp 27, Kp 37, Kp 42, and Kp 46), (Kp 40, and Kp 21), (Kp 5, and Kp 15) and (Kp 49, Kp 28, and Kp10).
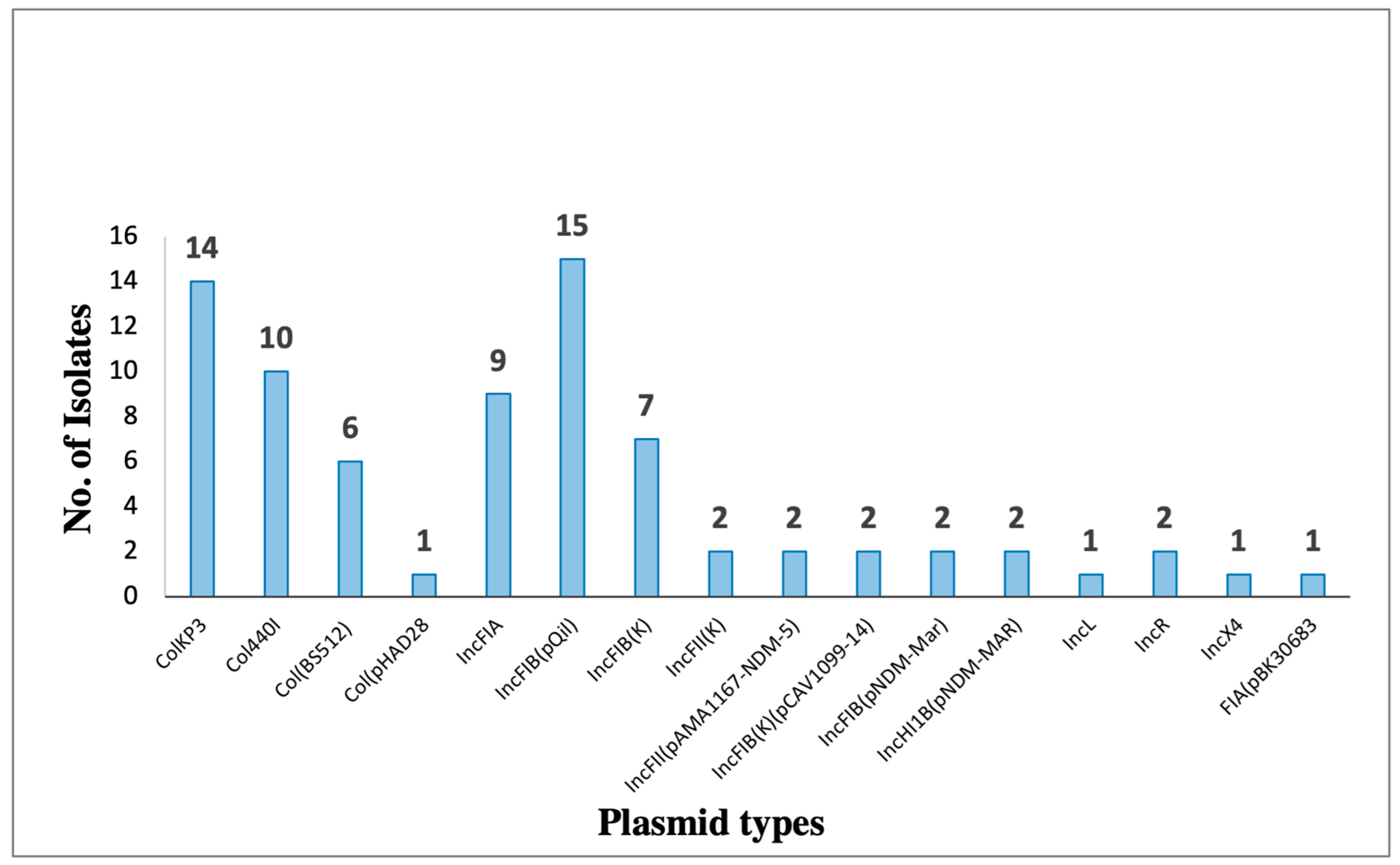
Among the detected plasmids, eight of them harbored antibiotic resistance genes (Figure 4. And Table 10). The most prevalent plasmid is pKPQIL-IT, which is present in 15 samples that belong to ST-395 and ST-231. It is a 115,300 bp in size IncFIB(QIL) replicon carrying genes associated with resistance to β-lactams (blaTEM-1, and blaKPC-3 ).
Figure 4.
Bar chart representing the distribution of plasmid replicon types identified among K. pneumoniae (n=23). Plasmid types were detected based on WGS data by using ResFinder online tool.
Figure 4.
Bar chart representing the distribution of plasmid replicon types identified among K. pneumoniae (n=23). Plasmid types were detected based on WGS data by using ResFinder online tool.
Plasmid pKP3-A is a 7,605 bp linear ColKP3 replicon, carrying blaOXA-181 gene. ST-231 and ST-405 sharing pKP3-A uniqueness that the integron is interrupted by insertion sequences (Figure 3.8). These isolates exhibited highly similar genotypes. All produced OXA-181, and the majority also have plasmid mediated- blaTEM-1, blaKPC-3 genes. Unlike Xpert Carba-R, the plasmid finder tool detected kpc gene from WGS data in all ST-231 and ST-405 isolates. This could suggest that the copy numbers of the plasmid might be low or the gene might have not been expressed and hence could not be detected by the GeneXpert system.
Isolates Kp 41 and Kp 50 are the only two strains carrying IncFIB (pNDM-Mar) and IncHI1B (pNDM-Mar) plasmids. In this isolate, blaNDM-1 was within pNDM-Mar plasmid. Whereas, Isolate Kp 49 (ST-147) had blaNDM-1 as part of (ble-blaNMD-1) operon where it was flanked by bleomycin resistance (ble) gene and N-(5'-phosphoribosyl) anthranilate isomerase (trpF) gene. Moreover, the plasmid composition of Kp 49 had IncR, IncFII, and pKP3-A. The plasmid pAMA1167-NDM-5 is present only in 7 of the ST-231 isolates. It is a 11,310 bp size IncFII(pAMA1167-NDM-5) replicon with genes encoding resistance against aminoglycosides [aph(3’’)- Ib, aph(6)-Id, aadA2, aadA5, aac(3)-IIa, aac(6’)-Ib-cr5 ], β-lactams (blaNDM-5, blaOXA-1, blaCTX-M-15-1, blaTEM-1), chloramphenicol (cat), sulfonamides (Sul1 and Sul2), trimethoprim (dfrA12 and dfrA17), tetracycline (tet(b) and tet(c) ), and macrolides (emrE and mp(A)).Sequence analysis of pKPN-IT plasmid revealed that it is a 208,191-bp IncFIB(K) replicon carrying (aadA2, cat, Mph(A), Sul1, dfrA12) genes. It is found in KP 10, 25, 27, 37, 42, and 46 isolates. However, plasmid pBK30683 is a 139,941-bp FIA replicon present only in one isolate (KP 37).

Table 11.
plasmids existent in K. pneumoniae isolates and there features.
| isolate (KP) | Plasmid | Size (bp) | Replicon | Resistance genes |
|---|---|---|---|---|
| 5, 6, 7, 10, 11, 15, 16, 22, 28, 30, 41, 43, 44, 45, 50 | pKPQIL-IT | 115300 | IncFIB(QIL) |
blaTEM-1, blaKPC-3 |
| 5, 6, 11, 15, 16, 22, 28, 30, 41, 43, 44, 45, 49, 50 | pKP3-A | 7605 | ColKP3 |
BlaOXA-181 |
| 5, 6, 7, 15, 28, 30, 45 | pAMA1167-NDM-5 | 11310 | IncFII(pAMA1167-NDM-5) |
aadA5, aadA2, aac(3)-IId, aph(6)-Id, aph(3’’)-Ib, aac(6’)-Ib-cr5, blaNDM-5,blaOXA-1, blaCTX-M-15-1, blaTEM-1 dfrA17, dfrA12, Mph(A), Sul1, Sul2, emrE, tet(b), tet(C), cat |
| 10, 25, 27, 37, 42, 46 | pKPN-IT | 208191 | IncFIB(K) | aadA2, cat, Mph(A), Sul1, dfrA12 |
| 21, 40 | pCAV1099-14 | 113992 | IncFIB(K)(pCAV1099-14) | dfrA19, APH(3’’)-Ib, APH(3’)-Ia, QnrB52 |
| 41, 50 | pNDM-MAR | 267242 | IncFIB (pNDM-Mar) IncHI1B (pNDM-Mar) |
aac(6’)-Ib, blaOXA-1 , blaNDM-1, cat, QnrB1 |
| 27, 49 | pK245 | 98264 | IncR | aacC2, strA, strB, dfrA14, catA2, Qnrs, blaSH2A , blaTEM |
| 49 | pC15-1a | 92353 | IncFII | aac(6’)-Ib, aac(3)-II, blaTEM-1 , blaOXA-1, blaCTX-M-15-1 , tet(A) |
| 37 | pBK30683 |
139941 | FIA(pBK30683) | dfrA14, StrA, StrB, blaTEM-1, blaOXA-9, , blaKPC-3 Sul2,ant(3’’)-Ia |
4. Discussion
The present study focused on molecular characterizations of integrons as a common class of mobile elements, and their significance in the dissemination of multi-drug resistance genes among K. pneumoniae. Whole genome sequence (WGS) data were analyzed to determine the sequence type as well as charecterisation of integron-carrying plasmids. In addition, WGS was used for studying the correlation between antimicrobial resistance genotype and phenotype, and the level of agreement was analyzed. Moreover, the phylogenetic relatedness of isolates was associated with the patient’s demographic data to explore the possible spread of mobile elements. Most of the studies in the region investigate the epidemiology of AMR with a few studies concerning the prevalence of mobile genetic elements such as genomic islands and their molecular characteristics in Gram negative bacteria (GNB) such as A. baumannii and K. pneumoniae [16,17,29,30,31,32,33]. Moreover, another study was conducted in Gram-negative bacteria isolates in Palestinian hospitals focused on integrons and their role in AMR dissemination [34]. To our knowledge, this is the first study in Oman that assesses that transmissibility of plasmids by examining the efficiency of conjugation in-vitro thus emphasizing the importance of this mechanism in the spontaneous transfer of integron-carried plasmids intra-specially.
In the current study, 96% (48/50) of our K. pneumoniae isolates carried class 1 integron, whereas none of the isolates contained either class 2 or class 3 integrons. This finding is similar to the results of previous studies that were conducted in Iran [35,36]. In addition, a previous study showed that 100% of the MDR isolates were found to be positive for class 1 integron, 36% for class 2, and none for class 3 integron [10]. However, other studies detected class 2 integron at a low prevalence level of 1.7% and 8.3% [27,34].
High prevalence of class 1 integron among our MDR isolates was observed and therefore it is the main focus of this study. The presence of class 1 integrons in most of the isolates suggest that these genetic elements confer a significant advantage to their hosts where exposure to antibiotic overuse creates a selective pressure in hospital environments [4,10]. The absence of both class 2 and class 3 integrons in our isolates is expected since these classes are relatively uncommon [6,37,38,39]. Moreover, this might be due to the fact that our selection is limited to representative MDR isolates which were unusually difficult to treat over a short period of time, which might not reflect the actual prevalence. Therefore, a larger sample size including screening specimens is more likely to reveal the other classes of integrons.
In our study, all isolates were resistant to ampicillin which corresponds to the report of previous studies [2,40,41]. Moreover, our findings showed a higher prevalence of carbapenem resistance (imipenem and meropenem) than other studies [1,16]. However, the resistance rate to aminoglycosides in our study was lower than previous reports where amikacin and gentamicin were resistant to 51.6% and 45.2%, respectively (Derakhshan et al., 2013). These results were expected since our isolates were selected as MDRO from the beginning of this study, whereas in the above-mentioned studies, isolates were chosen regardless of their antibiotic resistance profile and were mostly screening samples. This can explain the difference in the antimicrobial susceptibility profile of our isolates against some of the classes of antibiotics.
Our findings showed that 37.5% (18/48) of integron-positive isolates were carrying variable regions. By the analysis of WGS data, a total of 10 different gene cassettes were detected in these isolates. The latter isolates encode for aminoglycoside, trimethoprim, rifampin, chloramphenicol, macrolide, and quaternary ammonium compounds. Trimethoprim resistant gene cassettes (dfrA14), which encodes for dihydrofolate reductase enzymes were found to be predominant in our isolates (34.7%). A number of studies showed that dfr genes were the most frequent gene cassettes present in integrons [2,14,34,42]. The stability of this gene cassette in class 1 integrons might indicate the overuse of the trimethoprim for a long period of time specially to treat urinary tract infections [14]. Moreover, despite the initial results of the variable region which was detected by employing conventional PCR using primers that bind in the 5’ and 3’ conserved regions, further WGS analysis showed that 13% of isolates with positive variable regions carried empty gene cassettes. Moreover, previous studies reported the presence of sul1 at the 3’CS region of the integrons [43,44,45]. However, it was only observed in (3/20) isolates, despite the amplification of 5’CS-3’CS region by PCR. This has been described in Salmonella enterica [46,47]. The non-classical structure of integrons where intI1 and/or 5’ end of integrons is truncated were also observed in two strains (Kp 16 and Kp 25), which might suggest that amelioration of IntI1 confers a selective advantage for the host bacterium.
Simple PCR assays tend to overestimate the presence of integron cassettes, and cannot provide further information on the subsequent changes on the integron cassettes such as deletions and insertions. In fact, gene cassettes undergo continuous changes in their compositions which means that these cassettes can integrate antimicrobial resistant genes once conditions are favorable. Truncations and inversions within the integron structures are not uncommon, and could not be further detected by simple PCR. Therefore, WGS data were key in bridging the gap on the various components within these integrons.
The gene cassette arrays (dfrA12-ant1) and (dfra12-emrE-ant1) counted for 8.7%. Kp 49 was the only isolate that harbored a cassette composed of four genes (Arr3- ereA2- aadA- cmlA1). In addition, some of the gene cassettes like aac, cat, emerE, erm, arr, and ant occurred in combination with different cassettes, indicating that new gene cassettes can still be generated. In this study, an attempt was made to classify the integrons by looking for the integron types in the INTEGRALL database, and it was found that some isolates were assigned to the same number of gene cassettes in INTEGRALL despite having different compositions. Upon further manual analysis, it was found that some of the old deposited sequences in the INTEGRALL database have different annotations for some genes. To avoid confusion, INEGRALL classification numbers were omitted in our analysis.
Plasmids are considered as main carriers for antibiotic resistant genes through HGT mechanisms. Spontaneous plasmid conjugation is possible when the plasmid has a compatible mechanism of transfer. A plasmid is classified as conjugative when it helps other conjugative elements like integrons to move between different strains [48]. In the current study, the trans-conjugant colonies were positive for intI1 gene in all the representative samples with trans-conjugation efficiency ranged between 0.05% to 17.3%. This confirmed the successful transfer of integrons between cells via HGT. WGS data for the tested isolates supports our hypothesis, as plasmids were identified in all of the tested samples which is similar to the result of a previous study [14]. Being carried on plasmids, these integrons might have a significant fitness cost since more copies of plasmids can be present compared to chromosomally located integrons. This observation has been reported in previous studies on K. pneumoniae as well as other Gram negative organisms [49,50].
Most of our sequenced isolates showed a hierarchical relationship between different types of MGEs. The detected gene cassettes were embedded inside integrons which in turn inserted inside transposons which are flanked by insertion sequences (IS) that can confer mobility to transposons. Some of these composite transposons were incorporated into conjugative plasmids that offer transfer to other cells [51]. Most of the isolates in this study (Kp 5, 21, 27, 28, 37, 40, 41, 42, 44, 46, 49 and, Kp 50) have the integrons inserted in transposons that were flanked by IS (Figure 4.). All above-mentioned strains had similar IS which is IS6 and IS6100 (Kp 42 and Kp 46). IS6100 belongs to the IS6 family of transposable elements forming co-integrates as an endpoint of transposition, which was originally isolated from Mycobacterium fortuitum [52]. In general, different previous studies showed the same relationship, were the integrons were located in transposons that imbedded in a conjugative plasmid [53,54]. However, MGEs including Tns and ISs had disrupted the integron structure. IS are usually small MGEs which are ubiquitous within most bacterial genomes. These IS act as facilitators for horizontal gene transfer and play a major role in the evolution of microbes thus increasing their diversity and adaptive capacity.
The presence of IS6 on both sides of the Dcm-methylation operon which is a type II Restriction-Modification (RM) System and next to the integron denotes that RM systems are mobile and located in plasmids. These systems are involved in genome rearrangements and enhance virulence and resistance plasmids dissemination by carriage on other MGEs like in our study on integrons [55,56,57]. The existence of these IS6 in our isolates entails their importance in disseminating antibiotic resistance genes among various genera and species of bacteria [58,59]. In this study, the specific capture of resistance cassettes by the integrons signifies apparent bias driven by the selective pressure of antibiotic therapy regimes. Other studies have also shown that integron could carry catabolic genes which prove that integrons play a broader role in bacterial evolution [8,60].
According to the MLST results in this study, 23 isolates were classified into nine different STs. The most common sequence type was ST-231(39%) followed by ST-395 (26%). Moreover, two isolates belonged to ST-405 was observed in patients from the same ward which is also suggestive of possible spread, while only one isolate in each of the following STs: 37, 45, 147, 280, 1710, and 1741 which were reported in previous studies [61,62,63,64].
ST-231; the most abundant STs found in this study are widely distributed in South-West Asia with clonal dissemination in Singapore, Brunei, and Darussalam between 2013 and 2015 [65,66]. In addition, in India, ST-231 strains were reported as a predominant ST in 22 isolates (45%) [67]. In Europe, the first occurrence of the MDR K. pneumoniae ST-231 clone was confirmed in Switzerland [68]. This dissemination may represent a global public threat toward a new epidemic clone. ST-395 has been reported as the most common MDR K. pneumoniae clone (69%) in North-Eastern France as well as an outbreak in ICU in Italy [69,70].
In a previous study conducted in isolates from Arabian Peninsula, different ST types were detected in MDR K. pneumoniae in gulf countries including Oman, however, ST-231 was not among these STs [71]. Similarly, previous studies were conducted in Oman and Saudi Arabia which have concluded that no ST-231 strain was found [72,73]. The absence of ST-231 possibly indicates a recent emergence of ST-231 in Oman. However, ST-147 was one of the commonly detected STs from K. pneumoniae isolates in the two previous studies [71,72], whereas in the present study only one strain belongs to ST-147.
Both of these STs, ST-395 carrying strains and 77.8% of ST-231, were clustered in the same wards (mainly medical and ICU wards) with a significant mortality rate (84.6%). This observation highly suggests that there is a horizontal transfer of resistance-conferring genes, which is alarming. The heavy use of broad-spectrum antibiotics for critically-ill patients in high dependency areas and ICU patients selects for highly resistant strains and enhances the spread of resistance determinants. Furthermore, this may indicate inefficient infection control practices that allowed the dissemination of the plasmid-mediated resistance in the hospital. This finding corresponds to a study that was conducted in carbapenem-resistant GNB in SQUH, where 87% of infections were being healthcare-associated with a 62% mortality rate [17].
Over the last 15 to 20 years, carbapenem-hydrolyzing β-lactamases including OXA-48-like and NDM type carbapenemases disseminated and emerged in Enterobacteriaceae all over the world [74,75]. Carbapenemase OXA-48-like differs from the classical OXA-48 by one to five amino acids in which they hydrolyze carbapenems and penicillins but do not affect extended-spectrum cephalosporin [75].
OXA-232-producing K. pneumoniae (OXA232Kp) was identified for the first time on a 6.1-kb ColE-type non-conjugative plasmid in France in 2013 from a patient who returned from India and since then it has spread worldwide [74,75,76]. OXA-232 is considered as a point mutation derivative from OXA-181 with one amino acid difference [77]. In this study, WGS and Xpert Carba-R data detected the presence of the blaOXA-48-like gene in 87% (20/23) of our XDR and PDR isolates. These genes were identified by WGS as blaOXA-232 in 93.3% (14/15) of our XDR and PDR sequenced isolates. All of them belonged to ST-231 and ST-395. The level of agreement between WGS and GeneXpert tested samples was highly significant (n=8/9) with only one isolate that was negative for OXA gene at GeneXpert and positive in WGS (Kp 43). Illumina WGS has good coverage for sequencing the whole genome, whereas, GeneXpert is based on real-time PCR, even with 97% specificity might miss some allelic variants. Plasmid analysis of our strains showed that the OXA-181 gene (or OXA-232) was carried on pKP3-A (7605 bp) replicon type ColKP3. It was expressed only in the XDR and PDR OXA-232 positive strains. This is similar to a previous study where 33% and 100% of OXA-232 isolates were carried on ColKP3 plasmid in the USA and China respectively [77,78].
On the other hand, NDM was first detected in 2008 from a Swedish patient of Indian origin and then distributed all over the world [79]. In this study, Xpert Carba-R results showed that only 4% (2/50) of our isolates had the NDM gene. Whereas, only one isolate (4.3%) that belongs to ST-147 harbor both OXA-48-like (blaOXA-232) and NDM gene (blaNDM-5) according to WGS data. A study was conducted in UAE to characterize carbapenem-resistant Enterobacteriaceae in the Arabian Peninsula, showed that NDM and OXA-48-like are the most commonly detected genes with a rate of 46.5% and 32.5%, respectively. In the samples collected from Oman, 46% and 44.4% of the isolates were carrying NDM and OXA-48-like (blaOXA-181), whereas, only 1.6% carry both genes [71].
Along with the carbapenem genes, ESBL genes including bla(CTX-M), bla(TEM), and bla(SHV) were detected, and have been reported in different studies [66,67]. All bla(CTX-M) genes were identified including blaCTX-M-15, and it was the dominant ESBL amongst our strains since it was produced by all sequenced isolates. One main factor that contributes to the CTX-M-type ESBLs distribution in K. pneumoniae is the conjugative plasmids, particularly that belong to IncF with specific insertion sequences [80]. Moreover, all bla(TEM) genes belonged to blaTEM-1 and detected in 69.6%, whereas, the SHV types were characterized as blaSHV-1, blaSHV-11, blaSHV-27, and blaSHV-76. All ST-231 strains carrying blaSHV-1, while ST-395 strains carrying blaSHV-11. Besides, OXA-1 was positive in all ST-395 strains only with an association with other ESBLs genes (blaSHV-11, blaCTX-M-15, blaTEM-1). The association between OXA-1 and CTX-M-15 genes will render isolates resistant to β-lactam-β-lactamase inhibitor combinations. Note worthily, a large variety of GNB carried OXA-1 gene in plasmid and integron locations [81]. Plasmid analysis in this study found that blaOXA-1 gene was located along with blaCTX-M-15 in IncF plasmids IncFII and IncFII (pAMA1167-NDM-5).
Resistance to colistin, a drug that is used as the last line drug in the treatment of extensively resistant pathogens, is increasingly reported in Enterobacteriaceae, particularly K. pneumoniae [82]. Several mechanisms are involved in colistin resistance, mostly the emergence of mobilized colistin resistance (mcr) genes via plasmid and the mutations in the chromosomal gene (mgrB) and operons (PmrAB, and PhoPQ) which are associated with the biosynthesis and modification of lipopolysaccharide (LPS). The PhoPQ regulatory system is activated at a low concentration of Mg2+ or Ca2+ and acidic PH [83]. The PhoQ (sensor kinase) activates PhoP (regulator protein) by phosphorylation, which in turn activates pmrFHIJKLM operons. These operons lead to LPS modification by adding 4-amino-4-deoxy-L-arabinose (L-Ara4N) and phosphoethanolamine (PETN) to lipid A. This modification neutralizes the negative charge of LPS leading to the low affinity of LPS to positively charged colistin [83,84,85].
Four sequenced isolates were resistant to colistin via BMD, however, all of these strains were negative for the plasmid-mediated colistin resistance gene (mcr) and harbored a truncated mgrB gene. Bioinformatics analysis revealed the presence of four SNPs in the PhoP genes with the following amino acid substitutions: (Val77Glu), (Gln147His), (Gln131Glu), and (Pro129Thr). Further analysis of the promoter region of PhoPQ operon did not reveal any SNPs. A previous study reported different SNPs in PhoP (Val3Phe and Ser86Leu) and PhoQ (Leu26Pro) that were found to have a role in colistin resistance when tested by real-time PCR [86]. However, the detected SNPs in this study have not been described before, and therefore, further expression analysis by real-time PCR is needed to show their role a long with the truncated mgrB gene in colistin resistance. Moreover, colistin resistance might arise due to other resistance mechanisms such as capsule overproduction that causes a reduction in the interaction between colistin and its target site at K. pneumoniae (LPS) [83,87]. Furthermore, all isolates in this study carried K. pneumoniae efflux pumps (Kpn E, F, G, H) which might contribute to colistin resistance.
Tigecycline has broad-spectrum activity against Gram-positive and Gram-negative bacteria and has demonstrated efficacy and safety as salvage therapy for MDR/XDR bacteria [29,32,33,88,89]. However, the tigecycline resistance is becoming more common in ESBL-producing, MDR, XDR, and carbapenem-resistant isolates [90,91]. In this study, the majority of the sequenced isolates (17/19) were tigecycline-resistant, while the remaining four isolates were not tested. The WGS data showed that the efflux pumps’ regulator gene (marA) was present in all isolates (n=23), including the two sensitive ones (Kp37 and Kp46). In this context, a study reported that the marA gene, along with other regulatory pathways, could mediate the resistance to tigecycline through upregulating acrAB efflux pump even in the absence of ram A [92]. In another study, the development of tigecycline resistance was attributed to the tetA gene [93]. In our study, the tetA gene was detected in four ESBL isolates, from which only two were resistant. Thus, it’s more likely that marA regulator is engaged in the tigecycline resistance.
Different genes that confer resistance against aminoglycosides were detected including aac(6')-Ib-cr, aac(6')-Ib Hangzhou, aadA1,2, aac(3)-lId, aph(3')-Ia, strA, strB, armA and rmtB. The latter two genes are plasmid-encoded 16S rRNA methylase genes encoding for enzymes that have the ability to alter the antibiotic target [94]. Several studies were reported that armA gene was located in conjugative plasmids of the IncL/M or IncFIA plasmid, while rmtB in IncFI or IncFIA [94,95]. Interestingly, these two genes were only identified in Kp 49, which belongs to ST-147, and it is OXA-232, and NDM-5 producing isolate. In addition, IncFIA(HI1) and IncFII plasmids were uniquely present in this isolate only, and a further search of rmtB and armA genes by BLAST using WGS data confirmed the localization of these two genes in the (IncFII) plasmid.
Among carbapenems, cephalosporin, and quinolones, all isolates possessed expressed genes with a complete agreement between AMR genotypic findings and phenotypic expression. These results are close to what has been detected in a previous study [96]. In contrast, aminoglycosides antibiotics (Amikacin and gentamicin) had high non-expressed genes (57.1% and 28.6%). Since there are other mechanisms behind AMR, the presence or absence of certain genes might not be a sufficient indicator of the isolate’s resistance profile. Furthermore, genotypic analysis has to be correlated with phenotypic findings of various mechanisms involved.
The versatility and ubiquity of integrons in bacterial genomes indicate the key role of these mobile elements in bacterial adaptation. The abundance of repeat sequences of ISs and Tns within the integron structures in our WGS data pertains to the tremendous selective pressure of used antibiotic regimens in the hospital microbial environment. These integrons are very potent capture systems with a limitless capacity to exchange antibiotic gene cassettes as well as other genes that increase their fitness. Integron with combinations of various antibiotic cassettes had been termed as multi-resistance integrons (MRIs) in Gram negative bacteria [97].
The genomic plasticity was apparent in a few cases where there was a deletion of non-essential genes or empty integrons which confirms that these integron systems are performing adaptive rather than housekeeping functions. In support with this hypothesis, two of the isolates we examined had integrons with no antibiotic gene cassettes and the intI1 gene was partially truncated (Kp 16 and Kp25). The different attachment sites and the variety of open reading frames of hypothetical or unknown functions within these integron systems are compelling. Further detailed analysis is required to unravel the importance of integrons among other MGEs towards the adaptation of bacterial evolution.
5. Conclusions
In summary, this study demonstrates a high prevalence of class 1 integrons (96%) in MDR K. pneumoniae in SQUH and the absence of class 2 and class 3 integrons. The data of WGS confirmed the presence of diverse integron-carried gene cassettes, with dfrA as a predominant cassette. In addition, two main STs (ST-231 and ST-395) were detected with the dominance of OXA-232 carbapenemase while the NDM-5 type was identified in one isolate. Sequence typing and genotypic characterization of the isolates by WGS revealed a possible break in the infection control in the male medical ward (R3) and ICU causing intra-hospital transmission and spread of carbapenem-resistance. The dissemination of integrons by the horizontal transfer of the conjugative plasmids play a vital role in the spread and exchange of resistance genes between bacteria of the same or different species 80,98. This phenomenon imposes a tremendous threat to the currently available antibiotic regimens and necessitates the adoption of strict antimicrobial stewardship programs to prevent further dissemination of AMROs as well as improvement of infection control measures.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Z.J.; methodology, M.M. and Z.J.; software, M.M. and Z.J.; validation, M.R.., Z.M. and Z.J.; formal analysis, M.M. and M.Q.; investigation, M.M.; resources, Z.M.; data curation, Z.J. and M.R.; writing—original draft preparation, M.M.; writ-ing—review and editing, Z.J.; visualization, M.R.; supervision, Z.J and M.R..; project administration, Z.J. and Z. M..; funding ac-quisition, Z.J and M.R. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by Sultan Qaboos University Internal Grant, Muscat, Oman, grant number IG/MED/MIC/21/01
Institutional Review Board Statement
This study has been approved by the Medical Research Ethics Committee (MREC), College of Medicine and Health Sciences, Sultan Qaboos University, Oman. The MREC approval number 1896.
Data Availability Statement
All supporting data can be found in the supplemantry files. All whole genome sequencing data are deposited in Genbank at accession number PRJNA999478.
Acknowledgments
We would like to express our sincere thanks to the sequencing company pro- vided by MicrobesNG (Birmingham, UK) (https://microbesng.com (accessed on 4 July 2022) for performing WGS and bioinformatics analysis. We would like to thank supervising staff, College of Agriculture, Department of Food Microbiology for providing instrumental support to conduct some experiments for cultivating the bacterial isolates.
Conflicts of Interest
The authors declare no conflict of interest
References
- Najafgholizadeh Pirzaman, A.; Mojtahedi, A. Investigation of Antibiotic Resistance and the Presence of Integron Genes among ESBL Producing Klebsiella Isolates. Meta Gene 2019, 19. [Google Scholar] [CrossRef]
- Lima, A. M.d.S.; de Melo, M. E. S.; Alves, L. C.; Brayner, F. A.; Lopes, A. C. S. Investigation of Class 1 Integrons in Klebsiella Pneumoniae Clinical and Microbiota Isolates Belonging to Different Phylogenetic Groups in Recife, State of Pernambuco. Rev Soc Bras Med Trop 2014, 47. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R. L.; Da Silva, B. C. M.; Rezende, G. S.; Nakamura-Silva, R.; Pitondo-Silva, A.; Campanini, E. B.; Brito, M. C. A.; Da Silva, E. M. L.; De Melo Freire, C. C.; Da Cunha, A. F.; Da Silva Pranchevicius, M. C. High Prevalence of Multidrug-Resistant Klebsiella Pneumoniae Harboring Several Virulence and β-Lactamase Encoding Genes in a Brazilian Intensive Care Unit. Front Microbiol 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Hobson, C. A.; Pierrat, G.; Tenaillon, O.; Bonacorsi, S.; Bercot, B.; Jaouen, E.; Jacquier, H.; Birgy, A. Klebsiella Pneumoniae Carbapenemase Variants Resistant to Ceftazidime-Avibactam: An Evolutionary Overview. Antimicrobial Agents and Chemotherapy. 2022. [CrossRef]
- Bengoechea, J. A.; Sa Pessoa, J. Klebsiella Pneumoniae Infection Biology: Living to Counteract Host Defences. FEMS Microbiology Reviews. 2019. [CrossRef]
- Mazel, D. Integrons: Agents of Bacterial Evolution. Nature Reviews Microbiology. 2006. [CrossRef]
- Fluit, A. C.; Schmitz, F. J. Resistance Integrons and Super-Integrons. Clinical Microbiology and Infection. 2004. [CrossRef]
- Gillings, M. R. Integrons: Past, Present, and Future. Microbiology and Molecular Biology Reviews 2014, 78. [Google Scholar] [CrossRef]
- Tseng, C. H.; Huang, Y. T.; Mao, Y. C.; Lai, C. H.; Yeh, T. K.; Ho, C. M.; Liu, P. Y. Insight into the Mechanisms of Carbapenem Resistance in Klebsiella Pneumoniae: A Study on IS26 Integrons, Beta-Lactamases, Porin Modifications, and Plasmidome Analysis. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- Firoozeh, F.; Mahluji, Z.; Khorshidi, A.; Zibaei, M. Molecular Characterization of Class 1, 2 and 3 Integrons in Clinical Multi-Drug Resistant Klebsiella Pneumoniae Isolates. Antimicrob Resist Infect Control 2019, 8. [Google Scholar] [CrossRef]
- Shukla, S.; Joshi, P.; Trivedi, P.; Akinwotu, O.; Gajjar, D. Genomic Islands in Klebsiella Pneumoniae. In Microbial Genomic Islands in Adaptation and Pathogenicity; 2023. [CrossRef]
- Di Pilato, V.; Principe, L.; Andriani, L.; Aiezza, N.; Coppi, M.; Ricci, S.; Giani, T.; Luzzaro, F.; Rossolini, G. M. Deciphering Variable Resistance to Novel Carbapenem-Based β-Lactamase Inhibitor Combinations in a Multi-Clonal Outbreak Caused by Klebsiella Pneumoniae Carbapenemase (KPC)-Producing Klebsiella Pneumoniae Resistant to Ceftazidime/Avibactam. Clinical Microbiology and Infection 2023, 29. [Google Scholar] [CrossRef]
- Castillo-Polo, J. A.; Hernández-García, M.; Morosini, M. I.; Pérez-Viso, B.; Soriano, C.; De Pablo, R.; Cantón, R.; Ruiz-Garbajosa, P. Outbreak by KPC-62-Producing ST307 Klebsiella Pneumoniae Isolates Resistant to Ceftazidime/Avibactam and Cefiderocol in a University Hospital in Madrid, Spain. Journal of Antimicrobial Chemotherapy 2023, 78. [Google Scholar] [CrossRef]
- Xu, X.; Li, X.; Luo, M.; Liu, P.; Su, K.; Qing, Y.; Chen, S.; Qiu, J.; Li, Y. Molecular Characterisations of Integrons in Clinical Isolates of Klebsiella Pneumoniae in a Chinese Tertiary Hospital. Microb Pathog 2017, 104, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Drouin, F.; Mélançon, J.; Roy, P. H. The IntI-like Tyrosine Recombinase of Shewanella Oneidensis Is Active as an Integron Integrase. J Bacteriol 2002, 184. [Google Scholar] [CrossRef]
- Balushi, M. Al; Kumar, R.; Al-Rashdi, A.; Ratna, A.; Al-Jabri, A.; Al-shekaili, N.; Rani, R.; Sumri, S. Al; Al-Ghabshi, L.; Al-Abri, S.; Al-Jardani, A. Genomic Analysis of the Emerging Carbapenem-Resistant Klebsiella Pneumoniae Sequence Type 11 Harbouring Klebsiella Pneumoniae Carbapenemase (KPC) in Oman. J Infect Public Health 2022, 15. [Google Scholar] [CrossRef] [PubMed]
- Balkhair, A.; Al-Muharrmi, Z.; Al’Adawi, B.; Al Busaidi, I.; Taher, H. B.; Al-Siyabi, T.; Al Amin, M.; Hassan, K. S. Prevalence and 30-Day All-Cause Mortality of Carbapenem-and Colistin-Resistant Bacteraemia Caused by Acinetobacter Baumannii, Pseudomonas Aeruginosa, and Klebsiella Pneumoniae: Description of a Decade-Long Trend. International Journal of Infectious Diseases 2019, 85. [Google Scholar] [CrossRef]
- Balkhair, A.; Al-Farsi, Y. M.; Al-Muharrmi, Z.; Al-Rashdi, R.; Al-Jabri, M.; Neilson, F.; Al-Adawi, S. S.; El-Beeli, M.; Al-Adawi, S. Epidemiology of Multi-Drug Resistant Organisms in a Teaching Hospital in Oman: A One-Year Hospital-Based Study. The Scientific World Journal 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- 19. CLSI. M100 Performance Standards for Antimicrobial Susceptibility Testing, 32nd Edition. Clinical Laboratory Standard Institute, 2021.
- Antimicrobial, E. C. on. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 10.0, 2020. Http://Www.Eucast.Org. Breakpoint tables for interpretation of MICs and zone diameters. Version 10.0, 2020. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing A CLSI Supplement for Global Application. Clinical and Laboratory Standards Institute 2020. [Google Scholar]
- Kaas, R. S.; Leekitcharoenphon, P.; Aarestrup, F. M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res 2021, 49. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R. S.; Ruppe, E.; Roberts, M. C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R. L.; Rebelo, A. R.; Florensa, A. F.; Fagelhauer, L.; Chakraborty, T.; Neumann, B.; Werner, G.; Bender, J. K.; Stingl, K.; Nguyen, M.; Coppens, J.; Xavier, B. B.; Malhotra-Kumar, S.; Westh, H.; Pinholt, M.; Anjum, M. F.; Duggett, N. A.; Kempf, I.; Nykäsenoja, S.; Olkkola, S.; Wieczorek, K.; Amaro, A.; Clemente, L.; Mossong, J.; Losch, S.; Ragimbeau, C.; Lund, O.; Aarestrup, F. M. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. Journal of Antimicrobial Chemotherapy 2020, 75. [Google Scholar] [CrossRef]
- Larsen, J.; Enright, M. C.; Godoy, D.; Spratt, B. G.; Larsen, A. R.; Skov, R. L. Multilocus Sequence Typing Scheme for Staphylococcus Aureus: Revision of the Gmk Locus. Journal of Clinical Microbiology 2012, 2538–2539. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M. V.; Lund, O.; Villa, L.; Aarestrup, F. M.; Hasman, H. PlasmidFinder and PMLST: In Silico Detection and Typing of Plasmid. Antimicrob Agents Chemother 2014, 58. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Davies, J.; Miao, V. Molecular Characterization of Class 3 Integrons from Delftia Spp. J Bacteriol 2007, 189. [Google Scholar] [CrossRef]
- Malek, M. M.; Amer, F. A.; Allam, A. A.; El-Sokkary, R. H.; Gheith, T.; Arafa, M. A. Occurrence of Classes I and II Integrons in Enterobacteriaceae Collected from Zagazig University Hospitals, Egypt. Front Microbiol 2015, 6. [Google Scholar] [CrossRef]
- Al-Jabri, Z.; Zamudio, R.; Horvath-Papp, E.; Ralph, J. D.; Al-Muharrami, Z.; Rajakumar, K.; Oggioni, M. R. Integrase-Controlled Excision of Metal-Resistance Genomic Islands in Acinetobacter Baumannii. Genes (Basel) 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Al-Jabri, Z. J.; Busaidi, B. A.; Muzahmi, M. A.; Shabibi, Z. A.; Rizvi, M.; Rashdi, A. A.; Al-Jardani, A.; Farzand, R. Diversity of the Sequence Type Determines the Unique Genetic Arrangement of K-Loci in Hypervirulent Klebsiella Pneumoniae Clinical Strains I n r e v i e w. www.frontiersin.org.
- Dandachi, I.; Chaddad, A.; Hanna, J.; Matta, J.; Daoud, Z. Understanding the Epidemiology of Multi-Drug Resistant Gram-Negative Bacilli in the Middle East Using a One Health Approach. Frontiers in Microbiology. 2019. [CrossRef]
- Quraini, M. AL; Jabri, Z. AL; Sami, H.; Mahindroo, J.; Taneja, N.; Muharrmi, Z. AL; Busaidi, I. AL; Rizvi, M. Exploring Synergistic Combinations in Extended and Pan-Drug Resistant (XDR and PDR) Whole Genome Sequenced Acinetobacter Baumannii. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Al-Quraini, M.; Rizvi, M.; Al-Jabri, Z.; Sami, H.; Al-Muzahmi, M.; Al-Muharrmi, Z.; Taneja, N.; Al-Busaidi, I.; Soman, R. Assessment of In-Vitro Synergy of Fosfomycin with Meropenem, Amikacin and Tigecycline in Whole Genome Sequenced Extended and Pan Drug Resistant Klebsiella Pneumoniae: Exploring A Colistin Sparing Protocol. Antibiotics 2022, 11. [Google Scholar] [CrossRef]
- Hussein, A. I. A.; Ahmed, A. M.; Sato, M.; Shimamoto, T. Characterization of Integrons and Antimicrobial Resistance Genes in Clinical Isolates of Gram-Negative Bacteria from Palestinian Hospitals. Microbiol Immunol 2009, 53. [Google Scholar] [CrossRef]
- Derakhshan, S.; Peerayeh, S. N.; Fallah, F.; Bakhshi, B.; Rahbar, M.; Ashrafi, A. Detection of Class 1, 2, and 3 Integrons among Klebsiella Pneumoniae Isolated from Children in Tehran Hospitals. Arch Pediatr Infect Dis 2014, 2. [Google Scholar] [CrossRef]
- Mohammadi, R.; Haddadi, A.; Harzandi, N. Prevalence of Integrons as the Carrier of Multidrug Resistance Genes among Clinical Isolates of Klebsiella; 2019; Vol. 8.
- Yang, L.; Zhang, G.; Zhao, Q.; Guo, L.; Yang, J. Molecular Characteristics of Clinical IMP-Producing Klebsiella Pneumoniae Isolates: Novel IMP-90 and Integron In2147. Ann Clin Microbiol Antimicrob 2023, 22. [Google Scholar] [CrossRef] [PubMed]
- Omar F., H.; A. H. Ibrahim. THE PREVALENCE OF INTEGRON CLASS I AND II AMONG MULTI-DRUG RESISTANCE PRODUCING KLEBSIELLA Pneumonia. IRAQI JOURNAL OF AGRICULTURAL SCIENCES 2023, 54. [Google Scholar] [CrossRef]
- Collis, C. M.; Kim, M. J.; Partridge, S. R.; Stokes, H. W.; Hall, R. M. Characterization of the Class 3 Integron and the Site-Specific Recombination System It Determines. J Bacteriol 2002, 184. [Google Scholar] [CrossRef] [PubMed]
- Samah, G.; Hatem, M. E. S.; El, K. A. T.; Nikhat, M. Antimicrobial Resistance Patterns of Klebsiella Isolates from Clinical Samples in a Saudi Hospital. Afr J Microbiol Res 2017, 11. [Google Scholar] [CrossRef]
- Ma, L.; Lin, C. J.; Chen, J. H.; Fung, C. P.; Chang, F. Y.; Lai, Y. K.; Lin, J. C.; Siu, L. K. Widespread Dissemination of Aminoglycoside Resistance Genes ArmA and RmtB in Klebsiella Pneumoniae Isolates in Taiwan Producing CTX-M-Type Extended-Spectrum β-Lactamases. Antimicrob Agents Chemother 2009, 53. [Google Scholar] [CrossRef]
- Li, B.; Hu, Y.; Wang, Q.; Yi, Y.; Woo, P. C. Y.; Jing, H.; Zhu, B.; Liu, C. H. Structural Diversity of Class 1 Integrons and Their Associated Gene Cassettes in Klebsiella Pneumoniae Isolates from a Hospital in China. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Levesque, C.; Piche, L.; Larose, C.; Roy, P. H. PCR Mapping of Integrons Reveals Several Novel Combinations of Resistance Genes. Antimicrob Agents Chemother 1995, 39. [Google Scholar] [CrossRef]
- Antunes, P.; Machado, J.; Sousa, J. C.; Peixe, L. Dissemination of Sulfonamide Resistance Genes (Sul1, Sul2, and Sul3) in Portuguese Salmonella Enterica Strains and Relation with Integrons. Antimicrob Agents Chemother 2005, 49. [Google Scholar] [CrossRef]
- Domínguez, M.; Miranda, C. D.; Fuentes, O.; De La Fuente, M.; Godoy, F. A.; Bello-Toledo, H.; González-Rocha, G. Occurrence of Transferable Integrons and Suland Dfrgenes among Sulfonamide-and/or Trimethoprim-Resistant Bacteria Isolated from Chilean Salmonid Farms. Front Microbiol 2019, 10. [Google Scholar] [CrossRef]
- Chuanchuen, R.; Koowatananukul, C.; Khemtong, S. Characterization of Class 1 Integrons with Unusual 3′ Conserved Region from Salmonella Enterica Isolates. Southeast Asian Journal of Tropical Medicine and Public Health 2008, 39. [Google Scholar]
- Evershed, N. J.; Levings, R. S.; Wilson, N. L.; Djordjevic, S. P.; Hall, R. M. Unusual Class 1 Integron-Associated Gene Cassette Configuration Found in IncA/C Plasmids from Salmonella Enterica. Antimicrob Agents Chemother 2009, 53. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; da Silva, G. J.; Nielsen, K. M. Integrons: Vehicles and Pathways for Horizontal Dissemination in Bacteria. Mob Genet Elements 2012, 2. [Google Scholar] [CrossRef]
- Huang, J.; Lan, F.; Lu, Y.; Li, B. Characterization of Integrons and Antimicrobial Resistance in Escherichia Coli Sequence Type 131 Isolates. Canadian Journal of Infectious Diseases and Medical Microbiology 2020, 2020, 3826186. [Google Scholar] [CrossRef] [PubMed]
- Ploy, M. C.; Chainier, D.; Thi, N. H. T.; Poilane, I.; Cruaud, P.; Denis, F.; Collignon, A.; Lambert, T. Integron-Associated Antibiotic Resistance in Salmonella Enterica Serovar Typhi from Asia. Antimicrob Agents Chemother 2003, 47. [Google Scholar] [CrossRef]
- Norman, A.; Hansen, L. H.; Sørensen, S. J. Conjugative Plasmids: Vessels of the Communal Gene Pool. Philosophical Transactions of the Royal Society B: Biological Sciences 2009. [Google Scholar] [CrossRef]
- Herron, P. R.; Evans, M. C.; Dyson, P. J. Low Target Site Specificity of an IS6100-Based Mini-Transposon, Tn1792, Developed for Transposon Mutagenesis of Antibiotic-Producing Streptomyces. FEMS Microbiol Lett 1999, 171. [Google Scholar] [CrossRef]
- Wang, D.; Hou, W.; Chen, J.; Yang, L.; Liu, Z.; Yin, Z.; Feng, J.; Zhou, D. Characterization of Novel Integrons, In1085 and In1086, and the Surrounding Genes in Plasmids from Enterobacteriaceae, and the Role for AttCaadA16 Structural Features during Atti1 × Attc Integration. Front Microbiol 2017, 8. [Google Scholar] [CrossRef]
- Martínez-García, S.; Rodríguez-Martínez, S.; Cancino-Diaz, M. E.; Cancino-Diaz, J. C. Extracellular Proteases of Staphylococcus Epidermidis: Roles as Virulence Factors and Their Participation in Biofilm. APMIS. Blackwell Munksgaard March 1, 2018, pp 177–185. [CrossRef]
- Oliveira, P. H.; Touchon, M.; Rocha, E. P. C. The Interplay of Restriction-Modification Systems with Mobile Genetic Elements and Their Prokaryotic Hosts. Nucleic Acids Res 2014, 42. [Google Scholar] [CrossRef]
- Greene, P. J.; Gupta, M.; Boyer, H. W.; Brown, W. E.; Rosenberg, J. M. Sequence Analysis of the DNA Encoding the Eco RI Endonuclease and Methylase. Journal of Biological Chemistry 1981, 256. [Google Scholar] [CrossRef]
- Shetty, V. P.; Akshay, S. D.; Rai, P.; Deekshit, V. K. Integrons as the Potential Targets for Combating Multidrug Resistance in Enterobacteriaceae Using CRISPR- Cas9 Technique. Journal of applied microbiology. NLM (Medline) July 4, 2023. [CrossRef]
- Wang, L.; Zhu, M.; Yan, C.; Zhang, Y.; He, X.; Wu, L.; Xu, J.; Lu, J.; Bao, Q.; Hu, Y.; Xu, T.; Liang, J. Class 1 Integrons and Multiple Mobile Genetic Elements in Clinical Isolates of the Klebsiella Pneumoniae Complex from a Tertiary Hospital in Eastern China. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Gai, Z.; Wang, X.; Liu, X.; Tai, C.; Tang, H.; He, X.; Wu, G.; Deng, Z.; Xu, P. The Genes Coding for the Conversion of Carbazole to Catechol Are Flanked by IS6100 Elements in Sphingomonas Sp. Strain XLDN2-5. PLoS One 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Qi, Y.; Zhang, X. X.; Ma, L. Xenogenetic Evolutionary of Integrons Promotes the Environmental Pollution of Antibiotic Resistance Genes — Challenges, Progress and Prospects. Water Research. 2023. [CrossRef] [PubMed]
- Zhang, X.; Chen, D.; Xu, G.; Huang, W.; Wang, X. Molecular Epidemiology and Drug Resistant Mechanism in Carbapenem-Resistant Klebsiella Pneumoniae Isolated from Pediatric Patients in Shanghai, China. PLoS One 2018, 13. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X.; Torres, V. V. L.; Liu, H.; Rocker, A.; Zhang, Y.; Wang, J.; Chen, L.; Bi, W.; Lin, J.; Strugnell, R. A.; Zhang, S.; Lithgow, T.; Zhou, T.; Cao, J. An Outbreak of Carbapenem-Resistant and Hypervirulent Klebsiella Pneumoniae in an Intensive Care Unit of a Major Teaching Hospital in Wenzhou, China. Front Public Health 2019, 7. [Google Scholar] [CrossRef]
- Lascols, C.; Peirano, G.; Hackel, M.; Laupland, K. B.; Pitout, J. D. D. Surveillance and Molecular Epidemiology of Klebsiella Pneumoniae Isolates That Produce Carbapenemases: First Report of OXA-48-like Enzymes in North America. Antimicrob Agents Chemother 2013, 57. [Google Scholar] [CrossRef] [PubMed]
- Long, S. W.; Olsen, R. J.; Eagar, T. N.; Beres, S. B.; Zhao, P.; Davis, J. J.; Brettin, T.; Xia, F.; Musser, J. M. Population Genomic Analysis of 1,777 Extended-Spectrum Beta-Lactamase-Producing Klebsiella Pneumoniae Isolates, Houston, Texas: Unexpected Abundance of Clonal Group 307. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Abdul Momin, M. H. F.; Liakopoulos, A.; Phee, L. M.; Wareham, D. W. Emergence and Nosocomial Spread of Carbapenem-Resistant OXA-232-Producing Klebsiella Pneumoniae in Brunei Darussalam. J Glob Antimicrob Resist 2017, 9. [Google Scholar] [CrossRef]
- Teo, J. W. P.; Kurup, A.; Lin, R. T. P.; Hsien, K. T. Emergence of Clinical Klebsiella Pneumoniae Producing OXA-232 Carbapenemase in Singapore. New Microbes New Infect 2013, 1. [Google Scholar] [CrossRef]
- Shankar, C.; Mathur, P.; Venkatesan, M.; Pragasam, A. K.; Anandan, S.; Khurana, S.; Veeraraghavan, B. Rapidly Disseminating Bla OXA-232 Carrying Klebsiella Pneumoniae Belonging to ST231 in India: Multiple and Varied Mobile Genetic Elements. BMC Microbiol 2019, 19. [Google Scholar] [CrossRef]
- Mancini, S.; Poirel, L.; Tritten, M. L.; Lienhard, R.; Bassi, C.; Nordmann, P. Emergence of an MDR Klebsiella Pneumoniae ST231 Producing OXA-232 and RmtF in Switzerland. Journal of Antimicrobial Chemotherapy. 2018. [CrossRef] [PubMed]
- Muggeo, A.; Guillard, T.; Klein, F.; Reffuveille, F.; François, C.; Babosan, A.; Bajolet, O.; Bertrand, X.; de Champs, C. Spread of Klebsiella Pneumoniae ST395 Non-Susceptible to Carbapenems and Resistant to Fluoroquinolones in North-Eastern France. J Glob Antimicrob Resist 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Maida, C. M.; Bonura, C.; Geraci, D. M.; Graziano, G.; Carattoli, A.; Rizzo, A.; Torregrossa, M. V.; Vecchio, D.; Giuffrè, M. Outbreak of ST395 KPC-Producing Klebsiella Pneumoniae in a Neonatal Intensive Care Unit in Palermo, Italy. Infection Control and Hospital Epidemiology. 2018. [CrossRef]
- Sonnevend, Á.; Ghazawi, A. A.; Hashmey, R.; Jamal, W.; Rotimi, V. O.; Shibl, A. M.; Al-Jardani, A.; Al-Abri, S. S.; Tariq, W. U. Z.; Weber, S.; Pál, T. Characterization of Carbapenem-Resistant Enterobacteriaceae with High Rate of Autochthonous Transmission in the Arabian Peninsula. PLoS One 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Dortet, L.; Poirel, L.; Al Yaqoubi, F.; Nordmann, P. NDM-1, OXA-48 and OXA-181 Carbapenemase-Producing Enterobacteriaceae in Sultanate of Oman. Clinical Microbiology and Infection 2012, 18. [Google Scholar] [CrossRef]
- Zaman, T. uz; Alrodayyan, M.; Albladi, M.; Aldrees, M.; Siddique, M. I.; Aljohani, S.; Balkhy, H. H. Clonal Diversity and Genetic Profiling of Antibiotic Resistance among Multidrug/Carbapenem-Resistant Klebsiella Pneumoniae Isolates from a Tertiary Care Hospital in Saudi Arabia. BMC Infect Dis 2018, 18. [Google Scholar] [CrossRef]
- Mataseje, L. F.; Boyd, D. A.; Fuller, J.; Haldane, D.; Hoang, L.; Lefebvre, B.; Melano, R. G.; Poutanen, S.; Van Caeseele, P.; Mulvey, M. R. Characterization of OXA-48-like Carbapenemase Producers in Canada, 2011-14. Journal of Antimicrobial Chemotherapy 2018, 73. [Google Scholar] [CrossRef]
- Findlay, J.; Hopkins, K. L.; Loy, R.; Doumith, M.; Meunier, D.; Hill, R.; Pike, R.; Mustafa, N.; Livermore, D. M.; Woodford, N. OXA-48-like Carbapenemases in the UK: An Analysis of Isolates and Cases from 2007 to 2014. Journal of Antimicrobial Chemotherapy 2017, 72. [Google Scholar] [CrossRef]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like Carbapenemases: The Phantom Menace. Journal of Antimicrobial Chemotherapy 2012, 67. [Google Scholar] [CrossRef]
- Li, X.; Ma, W.; Qin, Q.; Liu, S.; Ye, L.; Yang, J.; Li, B. Nosocomial Spread of OXA-232-Producing Klebsiella Pneumoniae ST15 in a Teaching Hospital, Shanghai, China. BMC Microbiol 2019, 19. [Google Scholar] [CrossRef]
- Lutgring, J. D.; Zhu, W.; De Man, T. J. B.; Avillan, J. J.; Anderson, K. F.; Lonsway, D. R.; Rowe, L. A.; Batra, D.; Rasheed, J. K.; Limbago, B. M. Phenotypic and Genotypic Characterization of Enterobacteriaceae Producing Oxacillinase-48-like Carbapenemases, United States. Emerg Infect Dis 2018, 24. [Google Scholar] [CrossRef]
- Zarfel, G.; Hoenigl, M.; Leitner, E.; Salzer, H. J. F.; Feierl, G.; Masoud, L.; Valentin, T.; Krause, R.; Grisold, A. J. Emergence of New Delhi Metallo- β-Lactamase, Austria. Emerging Infectious Diseases. 2011. [CrossRef] [PubMed]
- Mathers, A. J.; Peirano, G.; Pitout, J. D. D. The Role of Epidemic Resistance Plasmids and International High- Risk Clones in the Spread of Multidrug-Resistant Enterobacteriaceae. Clin Microbiol Rev 2015, 28. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Naas, T.; Nordmann, P. Class D Ss-Lactamases : Diversity, Epidemiology and Genetics. Antimicrob Agents Chemother 2009. [Google Scholar]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes Laurent Poirela,b,c, Aurélie Jayola,b,c and Patrice Nordmann. Clin Microbiol Rev 2017, 30. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A. Z.; Mahmood, S. S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H. S. Molecular Mechanisms Related to Colistin Resistance in Enterobacteriaceae. Infection and Drug Resistance. 2019. [CrossRef]
- Jayol, A.; Nordmann, P.; Brink, A.; Poirel, L. Heteroresistance to Colistin in Klebsiella Pneumoniae Associated with Alterations in the PhoPQ Regulatory System. Antimicrob Agents Chemother 2015, 59. [Google Scholar] [CrossRef] [PubMed]
- Wright, M. S.; Suzuki, Y.; Jones, M. B.; Marshall, S. H.; Rudin, S. D.; Van Duin, D.; Kaye, K.; Jacobs, M. R.; Bonomo, R. A.; Adamsa, M. D. Genomic and Transcriptomic Analyses of Colistin-Resistant Clinical Isolates of Klebsiella Pneumoniae Reveal Multiple Pathways of Resistance. Antimicrob Agents Chemother 2015, 59. [Google Scholar] [CrossRef]
- Cheng, Y. H.; Lin, T. L.; Pan, Y. J.; Wang, Y. P.; Lin, Y. T.; Wang, J. T. Colistin Resistance Mechanisms in Klebsiella Pneumoniae Strains from Taiwan. Antimicrob Agents Chemother 2015, 59. [Google Scholar] [CrossRef]
- Kuhn, J. M.; Di, Y. P. Determination of Mutational Timing of Colistin-Resistance Genes through Klebsiella Pneumoniae Evolution. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Song, Y.; Hu, L.; Shu, Q.; Ye, J.; Liang, J.; Chen, X.; Tan, L. Tigecycline Salvage Therapy for Critically Ill Children with Multidrug-Resistant/Extensively Drug-Resistant Infections after Surgery. International Journal of Infectious Diseases 2018, 75. [Google Scholar] [CrossRef]
- Al-Jabri, Z.; Al-Shabibi, Z.; Al-Bimani, A.; Al-Hinai, A.; Al-Shabibi, A.; Rizvi, M. Whole Genome Sequencing of Methicillin-Resistant Staphylococcus Epidermidis Clinical Isolates Reveals Variable Composite SCCmec ACME among Different STs in a Tertiary Care Hospital in Oman. Microorganisms 2021, 9, 1824. [Google Scholar] [CrossRef] [PubMed]
- Pournaras, S.; Koumaki, V.; Spanakis, N.; Gennimata, V.; Tsakris, A. Current Perspectives on Tigecycline Resistance in Enterobacteriaceae: Susceptibility Testing Issues and Mechanisms of Resistance. International Journal of Antimicrobial Agents. 2016. [CrossRef] [PubMed]
- Wang, X.; Xie, Y.; Li, G.; Liu, J.; Li, X.; Tian, L.; Sun, J.; Ou, H. Y.; Qu, H. Whole-Genome-Sequencing Characterization of Bloodstream Infection-Causing Hypervirulent Klebsiella Pneumoniae of Capsular Serotype K2 and ST374. Virulence 2018, 9, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Veleba, M.; Schneiders, T. Tigecycline Resistance Can Occur Independently of the RamA Gene in Klebsiella Pneumoniae. Antimicrob Agents Chemother 2012, 56. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; He, F.; Shi, Q.; Zhao, F.; Xu, J.; Fu, Y.; Yu, Y. The Rapid Emergence of Tigecycline Resistance in BlaKPC-2 Harboring Klebsiella Pneumoniae, as Mediated in Vivo by Mutation in TetA during Tigecycline Treatment. Front Microbiol 2018, 9. [Google Scholar] [CrossRef]
- Bogaerts, P.; Galimand, M.; Bauraing, C.; Deplano, A.; Vanhoof, R.; De Mendonca, R.; Rodriguez-Villalobos, H.; Struelens, M.; Glupczynski, Y. Emergence of ArmA and RmtB Aminoglycoside Resistance 16S RRNA Methylases in Belgium. Journal of Antimicrobial Chemotherapy 2007, 59. [Google Scholar] [CrossRef]
- Ayad, A.; Drissi, M.; de Curraize, C.; Dupont, C.; Hartmann, A.; Solanas, S.; Siebor, E.; Amoureux, L.; Neuwirth, C. Occurence of ArmA and RmtB Aminoglycoside Resistance 16S RRNA Methylases in Extended-Spectrum β-Lactamases Producing Escherichia Coli in Algerian Hospitals. Front Microbiol 2016, 7. [Google Scholar] [CrossRef]
- Ragupathi, N. K. D.; Bakthavatchalam, Y. D.; Mathur, P.; Pragasam, A. K.; Walia, K.; Ohri, V. C.; Veeraraghavan, B. Plasmid Profiles among Some ESKAPE Pathogens in a Tertiary Care Centre in South India. Indian Journal of Medical Research 2019, 149. [Google Scholar] [CrossRef]
- Rowe-Magnus, D. A.; Guerout, A. M.; Ploncard, P.; Dychinco, B.; Davies, J.; Mazel, D. The Evolutionary History of Chromosomal Super-Integrons Provides an Ancestry for Multiresistant Integrons. Proc Natl Acad Sci U S A 2001, 98. [Google Scholar] [CrossRef]
- Lerminiaux, N. A.; Cameron, A. D. S. Horizontal Transfer of Antibiotic Resistance Genes in Clinical Environments. Can J Microbiol 2019, 65. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Bar chart representing the percentage of antimicrobial susceptibility of K. pneumoniae isolates against different antibiotics.
Figure 1.
Bar chart representing the percentage of antimicrobial susceptibility of K. pneumoniae isolates against different antibiotics.
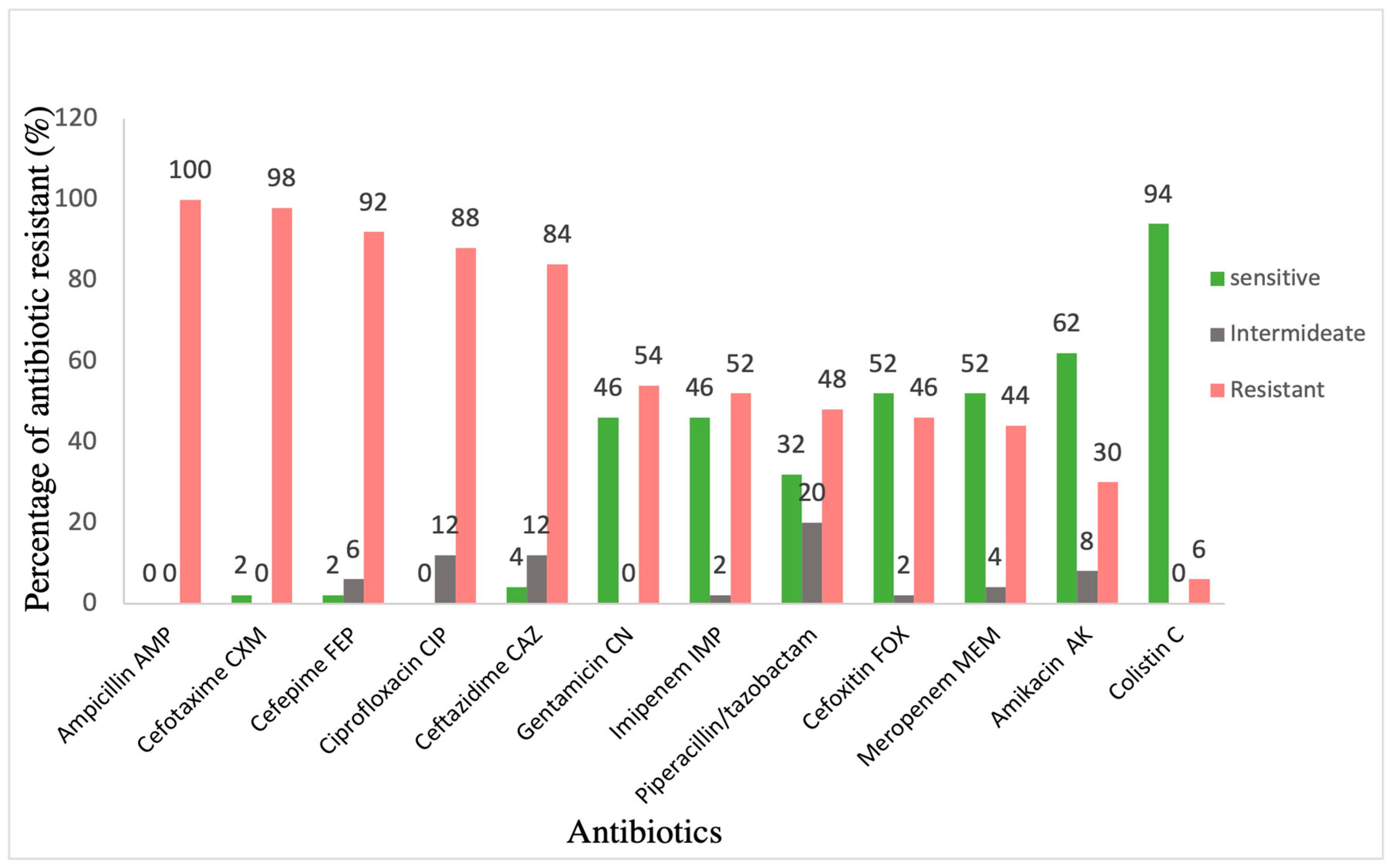
Figure 2.
Bar chart representing the percentage of trans-conjugation efficiency of K. pneumoniae isolates as donor and E. coli HB101 as a recipient. .
Figure 2.
Bar chart representing the percentage of trans-conjugation efficiency of K. pneumoniae isolates as donor and E. coli HB101 as a recipient. .
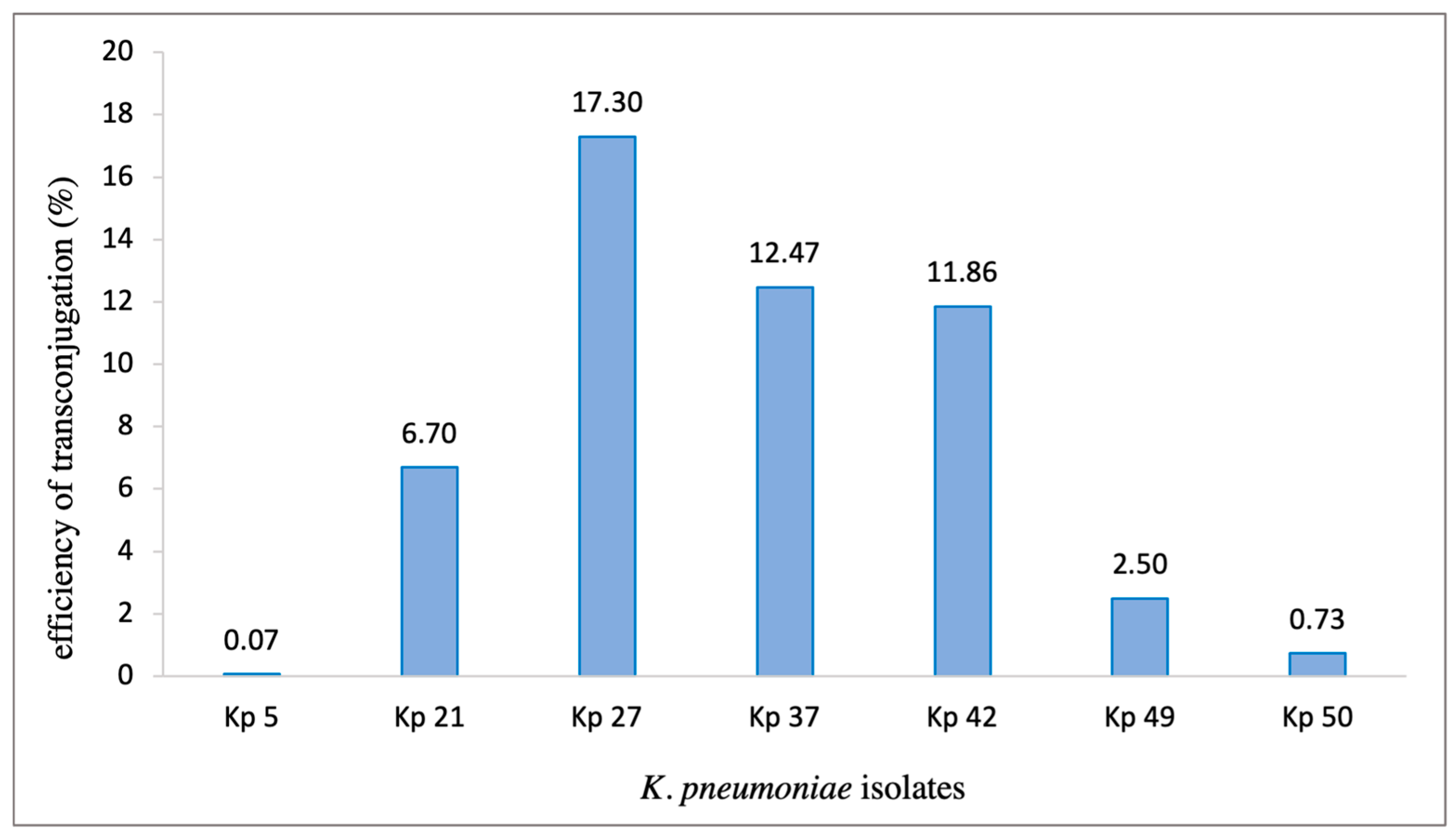
Figure 3.
Maximum likelihood phylogenetic tree constructed from WGS data for 23 K. pneumoniae isolates. The phylogenetic tree is annotated with the isolate’s number and sequence type (ST) as follows: Green: ST-231, purple: ST-405, and red:ST-395. Black boxes to the right of each strain number illustrates the distribution of antibiotic resistance genes as follows: Absent genes are shown in white boxes. K. pneumoniae HS11286 was used as a reference strain in this phylogenetic tree (accession number CP003200).
Figure 3.
Maximum likelihood phylogenetic tree constructed from WGS data for 23 K. pneumoniae isolates. The phylogenetic tree is annotated with the isolate’s number and sequence type (ST) as follows: Green: ST-231, purple: ST-405, and red:ST-395. Black boxes to the right of each strain number illustrates the distribution of antibiotic resistance genes as follows: Absent genes are shown in white boxes. K. pneumoniae HS11286 was used as a reference strain in this phylogenetic tree (accession number CP003200).

Table 1.
Clinical information of all K. pneumoniae isolates (n=50).
| Isolate | Year of isolation | Type of resistance | Specimen type | Hospital unit |
|---|---|---|---|---|
| Kp1 | 2019 | XDR | Tracheal aspirate | Emergency |
| Kp 2 | 2019 | XDR | Urine | Emergency |
| Kp 3 | 2019 | ESBL | Urine | Oncology |
| Kp 4 | 2019 | ESBL | Urine | Pediatrics |
| Kp 5 | 2019 | XDR | Urine | Male Medical |
| Kp 6 | 2019 | PAN | Tracheal aspirate | ICU |
| Kp 7 | 2019 | XDR | Wound | Male Medical |
| Kp 8 | 2019 | ESBL | Urine | Day Care |
| Kp 9 | 2019 | ESBL | Biopsy | Surgery |
| Kp 10 | 2019 | ESBL | Urine | Oncology |
| Kp 11 | 2019 | PAN | Tracheal aspirate | Male Medical |
| Kp 12 | 2019 | ESBL | Pus | Surgery |
| Kp 13 | 2019 | XDR | Blood culture | Emergency |
| Kp 14 | 2019 | ESBL | Wound | Male Medical |
| Kp 15 | 2019 | XDR | Catheter urine | Male Medical |
| Kp 16 | 2019 | XDR | Sputum | ICU |
| Kp 17 | 2019 | ESBL | Catheter urine | Pediatrics |
| Kp 18 | 2019 | ESBL | Urine | Emergency |
| Kp 19 | 2019 | ESBL | Urine | Pediatrics |
| Kp 20 | 2019 | ESBL | Urine | Pediatrics |
| Kp 21 | 2019 | ESBL | urine | Emergency |
| Kp 22 | 2019 | XDR | Wound | ICU |
| Kp 23 | 2019 | ESBL | Blood culture | Emergency |
| Kp 24 | 2019 | XDR | Skin | Emergency |
| Kp 25 | 2019 | ESBL | Peritoneal fluid | Male Medical |
| Kp 26 | 2019 | ESBL | wound | Surgery |
| Kp 27 | 2019 | ESBL | Urine | Emergency |
| Kp 28 | 2019 | XDR | Urine | Male Medical |
| Kp 29 | 2019 | XDR | Urine | Day Care |
| Kp 30 | 2019 | XDR | Wound | Male Medical |
| Kp 31 | 2019 | XDR | Sputum | ICU |
| Kp 32 | 2019 | XDR | bronchial wash | Emergency |
| Kp 33 | 2019 | ESBL | wound | Female Medical |
| Kp 34 | 2019 | ESBL | Urine | Pediatrics |
| Kp 35 | 2019 | ESBL | Blood culture | Female Medical |
| Kp 36 | 2019 | ESBL | Urine | Emergency |
| Kp 37 | 2019 | ESBL | Urine | Urology |
| Kp 38 | 2019 | ESBL | Urine | Emergency |
| Kp 39 | 2019 | ESBL | Urine | Emergency |
| Kp 40 | 2019 | ESBL | Tracheal aspirate | Neonatal unit |
| Kp 41 | 2019 | XDR | Wound | Male Medical |
| Kp 42 | 2019 | ESBL | Blood culture | Neonatal unit |
| Kp 43 | 2019 | XDR | Tracheal aspirate | Male Medical |
| Kp 44 | 2019 | XDR | Urine | Male Medical |
| Kp 45 | 2019 | XDR | Tracheal aspirate | Male Medical |
| Kp 46 | 2019 | ESBL | Tracheal aspirate | Neonatal unit |
| Kp 47 | 2019 | ESBL | Urine | Female Medical |
| Kp 48 | 2019 | XDR | Urine | Surgery |
| Kp 49 | 2019 | XDR | Urine | Male Medical |
| Kp 50 | 2019 | XDR | Urine | ICU |
Table 2.
Interpretive Categories and Zone Diameter Breakpoints, nearest whole mm [21.]
| Zone diameter breakpoints (mm) | ||||
| Antibiotic | Disk content | Susceptible | Intermediate | Resistant |
| Ampicillin | AMP 10mg | ≥17 | 14-16 | ≤13 |
| Piperacillin-tazobactam | TZP 110mg | ≥21 | 18–20 | ≤17 |
| Cefepime | FEP 30mg | ≥ 25 | 19– 24 | ≤ 18 |
| Cefotaxime | CTX 30mg | ≥26 | 23-25 | ≤22 |
| Cefoxitin | FOX 30mg | ≥18 | 15-17 | ≤ 14 |
| Ceftazidime | CAZ 30mg | ≥21 | 18-20 | ≤ 17 |
| Imipenem | IMP 10mg | ≥23 | 20-22 | ≤ 19 |
| Meropenem | MEM 10mg | ≥23 | 20-22 | ≤ 18 |
| Gentamicin | CN 30mg | ≥15 | 13-14 | ≤ 12 |
| Amikacin | AK 10mg | ≥17 | 15-16 | ≤ 14 |
| Ciprofloxacin | CIP 5mg | ≥31 | 21-30 | ≤ 20 |
Table 3.
PCR Cycling conditions for Go-Taq polymerases.
| Step | Temperature | Time |
|---|---|---|
| Initialization | 95 °C | 2 min |
| Denaturation | 95°C | 30 s |
| Annealing | 30 s | |
| Extension | 72°C | 1 min / kb |
| Final extension | 72°C | 10 min |
| Hold | 15°C |
Table 4.
List of specific primers used in this study. All primers were obtained from [14].
Table 4.
List of specific primers used in this study. All primers were obtained from [14].
| gene | Annealing temperature | Nucleotide sequence (5’ -3’ ) | Expected size | |
|---|---|---|---|---|
| Class 1 integrase gene | Intl1 | 56 °C | IntI1F (ACGAGCGCAAGGTTTCGGT) IntI1R (GAAAGGTCTGGTCATACATG) |
565 |
| Class 2 integrase gene | Intl2 | 52 °C | IntI2F (GTGCAACGCATTTTGCAGG) IntI2R (CAACGGAGTCATGCAGATG) |
403 |
| Class 3 integrase gene | Intl3 | 57 °C | IntI3F (CATTTGTGTTGTGGACGGC) IntI3R (GACAGATACGTGTTTGGCAA) |
717 |
| Variable regions | 52 °C | 5’ -CS (GGCATCCAAGCAGCAAG) 3’-CS (AAGCAGACTTGACCTGAT) |
Uncertain |
Table 5.
Antimicrobial susceptibility profile of the XDR (n=21), and PDR (n=2) isolates of K. pneumoniae.
Table 5.
Antimicrobial susceptibility profile of the XDR (n=21), and PDR (n=2) isolates of K. pneumoniae.
| Isolate | AMP | CTX | FEP | CIP | CAZ | TZP | FOX | IPM | MEM | CN | AK | CL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kp 1 | R | R | R | R | R | R | R | R | R | R | I | S |
| Kp 2 | R | R | R | R | R | R | R | R | R | S | I | R |
| Kp 5 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 6 PDR | R | R | R | R | R | R | R | R | R | R | R | R |
| Kp 7 | R | R | R | R | R | I | R | S | S | R | R | S |
| Kp 11 PDR | R | R | R | R | R | R | R | R | R | R | R | R |
| Kp 13 | R | R | R | R | R | R | R | I | R | R | R | S |
| Kp 15 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 16 | R | R | R | R | R | R | R | R | R | S | S | S |
| Kp 22 | R | R | R | R | R | S | R | R | R | S | S | R |
| Kp 24 | R | R | R | R | R | R | R | R | R | S | S | S |
| Kp 28 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 29 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 30 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 31 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 32 | R | R | R | R | I | R | R | R | R | R | R | S |
| Kp 41 | R | R | R | R | R | R | R | R | R | R | S | S |
| Kp 43 | R | R | R | R | R | R | R | R | R | R | S | S |
| Kp 44 | R | R | R | R | R | R | R | R | R | R | I | S |
| Kp 45 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 48 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 49 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 50 | R | R | R | R | R | R | R | R | R | R | S | R |
Abbreviations: Kp= Klebsiella pneumoniae, AMP= ampicillin, CTX= ceftriaxone, FEP= cefepime, CIP= ciprofloxacin, CAZ= ceftazidime, TZP= piperacillin tazobactam, FOX= cefoxitin, IPM= imipenem, MEM= meropenem, CN= gentamicin, AK= Amikacin, CL= colistin.
Table 6.
Demographic features and Clinical Characteristics of Patients.
| Demographic and Clinical Characteristics | Case Patients, n = 13 |
|---|---|
| Male, n (%) | 12 (92%) |
| Age | |
| Adults <=24 years, n (%) | 1 (7.7%) |
| From 25-50 years, n (%) | 3 (23.1%) |
| More than 50 years, n (%) | 9 (69.2%) |
| Age at first positive culture | |
| Mean age of adults, years (range) | 59.7 (20-86) |
| Length of stay | |
| Median length of stay to a first positive culture, days (range) | 48 (1–134) |
| Hospital location | |
| Intensive care unit (ICU), n (%) | 8 (61.5%) |
| Intermediate care ward, n (%) | 5 (83.5 %) |
Table 7.
The detected amino acids changes in PhoP and PhoQ in colistin-resistant strains (n=4).
| Isolate (KP) | PhoP | PhoQ |
|---|---|---|
| Kp 6 and Kp 11 | 3 SNPs | -Ve |
| (Gln147His) | ||
| (Gln131Glu) | ||
| (Pro129Thr) | ||
| Kp 22 and Kp 50 | 3 SNPs | -Ve |
| (Val129Glu) (Gln147His) | ||
| (Gln131Glu) |
Table 8.
Level of agreement of antimicrobial resistance genes and phenotypic resistance for K. pneumoniae isolates.
Table 8.
Level of agreement of antimicrobial resistance genes and phenotypic resistance for K. pneumoniae isolates.
| K. Pneumoniae (n=23) | Phenotypic resistance | Genes positive | Level of agreement- genotypic with phenotypic expression % | |
|---|---|---|---|---|
| Aminoglycosides | Amikacin | 9 | 21 | 42.90% |
| Gentamicin | 15 | 21 | 71.40% | |
| Quinolones | 23 | 23 | 100% | |
| Carbapenems | 14 | 14 | 100% | |
| Cephalosporins | 23 | 23 | 100% | |
Table 9.
The detected gene cassettes in K. pneumoniae isolates (n=23).
| Isolate | ST type | Gene cassette |
|---|---|---|
| Kp 40 | 37 | dfrA12,ant1 |
| Kp 49 | 147 | Arr3, ereA2, aadA, cmlA1 |
| Kp 21 | 45 | dfra12,ant1 |
| Kp 41 | 395 | dfrA14, |
| Kp 43 | 395 | dfrA12, APH(3”)-Ia, |
| Kp 44 | 395 | dfrA14 |
| Kp 50 | 395 | dfrA14 |
| Kp 16 | 395 | In0 |
| Kp 22 | 395 | In0 |
| Kp 5 | 231 | aac(6')-Ib, arr2, |
| Kp 6 | 231 | dfrA12, emrE, ant1 |
| Kp 7 | 231 | dfrA12, emrE, ant1 |
| Kp 10 | 231 | dfrA5 |
| Kp 11 | 231 | aacA4, cat1, ant1, |
| Kp 15 | 231 | aacA4, emrE, ant1 |
| Kp 28 | 231 | Ant1, erm, cat1 |
| Kp 30 | 231 | aacA4 |
| Kp 45 | 231 | dfrA14 |
| Kp 42 | 405 | dfrA14 |
| Kp 46 | 405 | dfrA14 |
| Kp 27 | 280 | dfrA14 |
| Kp 25 | 1741 | In0 |
| Kp 37 | 1710 | dfrA14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



