Submitted:
02 September 2023
Posted:
06 September 2023
You are already at the latest version
Abstract
Defining the best conditions for the initial development of native species is one of the significant challenges to forest production. Therefore, this study aimed to evaluate andiroba's physiological behavior and growth (Carapa guianensis Aubl.) seedlings at different shading levels. In a nursery seedling, C. guianensis seedlings grew on 0, 30, 50, and 70% shading, obtained with polyethylene screens. We measured at six plants per treatment. For the physiological data we measurement four times the total chlorophyll content, chlorophyll a/b ratio, maximum photochemical efficiency of photosystem II (Fv/Fm), maximum potential photosynthesis (A), stomatal conductance (gs), transpiration (E), intrinsic water use efficiency (WUEi), and carbon use efficiency (CUE), besides the growth parameters. While for growth rates parameters we developed six evaluation for as leaf area ratio (LAF), relative growth rates (RGR), and net assimilation rate (NAR). Chlorophyll content and Fv/Fm were inversely proportional to the light intensity. The other physiological and growth parameters showed better behavior in shaded environments, emphasizing the 50% treatment, with no significant difference between the 30% shading treatment. Therefore, the C. guianensis seedlings present growth plasticity under different shading levels, and we recommended producing seedlings at 50 or 30% shading.
Keywords:
amazon tree
; growth
; physiology
; photoinhibition
; plasticity
; light levels
1. Introduction
Tropical forests are critical ecosystems for global carbon dynamics, acting as one of the main components to contain the rise in atmospheric CO2 concentration [1]. Among tropical forests, the participation of the Amazon is undeniable, as a critical ecosystem for the planet's climate regulation, contributing to global carbon storage, generating about 15% of terrestrial photosynthesis, harboring approximately 25% of worldwide biodiversity, and making a real contribution to global feedbacks from the water and carbon cycles [2,3,4,5].
This ecosystem has been threatened in recent decades, suffering numerous pressures caused by deforestation and the consequent effects of climate change that promote imbalances and enhance the possibilities of alternating the roles of these areas for net carbon sources and reduction of the genetic bases of species in this ecosystem [6,7]. In this context, and added to the growing environmental awareness, interest in recovering degraded areas and reforestation increased, increasing the need for production and studies of native species seedlings [8].
Carapa guianensis Aubl. (Meliaceae), popularly known as andiroba, stands out among the native species of the Amazon forest, as it occurs throughout the territory of the region with great abundance in flooded forests [9]. It is a species of multiple-use and socioeconomic value for extractive populations in the Amazon region, mainly due to the medicinal properties of the oil extracted from seeds and the high quality of its wood [10]. However, little is known about its initial growth and physiological responses to different light intensities despite its importance and economic potential.
Exploring the behavior of physiological parameters in response to environmental conditions, as light intensity, water quantity, and CO2 availability, generates a better understanding of primary resource use strategies, especially in the early stages of growth and, consequently, facilitates adaptation of species in nurseries [11]. According to a study by [12], the growth and physiological aspects of forest species seedlings respond more intensely to variations in light availability than nutritional stress due to the function of light as a primary source of energy and its influence on plant growth and development control. The duration and quality of incident light influence the processes of seed germination, inhibition of stem elongation, synthesis of chlorophyll and anthocyanins, leaf expansion, and flowering [13].
Plant responses to light are not homogeneous, with a wide range of strategies depending on the availability of this resource. For example, [14] observed hormonal strategies for adaptation under shading; [15] verified growth investment and flowering due to the quality of available light, while [16] found no effect of radiation on the growth and photosynthetic activity of Tabebuia aurea (Aubl.) Willd. In addiction Changes in plant light conditions result in distinct morphophysiological differentiation for each species to maximize phytomass gain under the new conditions [17]. Among these changes, we highlight the increase in chlorophyll concentration, leaf area, and specific leaf area ratio, as well as the increase in the root/shoot ratio for plants in abundantly shaded environments [18,19].
Due to the great diversity of the Amazon region, many native species still lack basic information about the ideal conditions for initial growth, especially the effects of light availability for this process [20].
This study aims to reduce these gaps by evaluating C. guianensis a seedlings' growth and physiological behavior at different shading levels and defining the best light adaptation conditions for their initial development.
2. Materials and Methods
2.1. Experiment location and installation
The present study was conducted from January 2016 to February 2017 in the nursery of the Federal University of Oeste do Pará – UFOPA. The region has climate Ami (humid tropical), with two well-defined annual seasons; the dry period starts in July and ends in November, and the rainy period, starting in December and ending in June [21].
The seeds were collected in a commercial plantation in Belterra – PA, and germinated in washed sand. After germination, the seedlings were transplanted in plastic vases with a capacity of 5 L with a substrate of forest soil, organic matter, and sand, in a 3:2:1 ratio, kept in a greenhouse, and watered daily. After two months of adaptation, bean the treatments of the seedlings in greenhouses, where the level of light interception occurred with the aid of polyethylene screens (somber).
Jointly evaluated the growth and physiological parameters, and the experimental design used was completely randomized, with four treatments of light levels, under shading of 0% (full sun), 30%, 50%, and 70%. According to the infrared gas analyzer (IRGA) model LICOR 6400 (LICOR, Inc, Lincon, NE, USA), the plants received 62.8%, 49.5%, and 24.1% of light in the 30% treatments, 50%, and 70% shading, respectively, determined from collections on physiological measurements days.
2.2. Assessment of physiological responses
The subdivision of the plots for physiological responses was into four periods (30, 90, 180, and 300 days after induction to treatments - DAIT), always using leaves with good phytosanitary status for all analyses content was obtained. Of total chlorophyll and chlorophyll a/b ratio in all treatments, determined by the methodology cited by [22], from MgCO3 (0.5% v/v) to stabilize chlorophyll molecules. To determine the ratio of variable and maximum photochemical efficiency of photossystem II (PSII) (Fv/Fm) parameter was use a portable Plant Efficiency Analyzer (PEA - Plant Efficiency Analyzer, Hansatech Norfolk, UK), with dark adaptation for 30 minutes.
We measured the gas exchange parameters to understand the effect of light suppression on plants, including maximum potential photosynthesis (A), stomatal conductance (gs), transpiration (E), determined with IRGA, keeping light from constant saturation (1000 µmol m2 s-1), leaf temperature at 31ºC, water vapor at 30 mmol.mol-1 H2O in the chamber and CO2 concentration at 370 kPa. From the A/gs ratio got the intrinsic water use efficiency (WUEi), and the carboxylation use efficiency (CUE) was obtained from the ratio between e A/Ci (internal CO2 concentration).
2.3. Growth parameters
The quantification of plant production followed the same subdivision of plots to assess physiological responses, with six collection times (30, 60, 90, 120, 180, and 300 DAIT), a sample number of six plants per treatment. With the aid of a leaf area integrator (WinDIAS-C110-PC, Delta-T Devices Ltd, Cambridge, UK), we obtain the leaf area by three intact and fully expanded leaves from each individual. The calculate to leaf area ratio (LAR) is derived from the following equation:
Where: LAR = Leaf area ratio; LA = Leaf area; W = Total dry weight.
We separated the leaves from the stems and roots for each sample. These materials were over-dried at 65°C for 72 hours and weighed in analytical precision scales (Shimadzu, Mod. AUY220, ± 0.1mg), getting the total mass by the sum of the plant organs mass. Using phytomass, we calculate the relative growth rate (RGR) through the following equation:
Where: RGR = Relative growth rate; Ln = Natural logarithm; W1= Total dry weight at initial time; W2 = Total dry weight at final time; t1 and t2 = Time of initial and final measurements, respectively
The net assimilation rate (NAR) was calculated using the following equation from the leaf area and plant dry matter mass.
Where: NAR = Where: NAR = Net assimilation rate; W = Total dry mass; AF = Leaf area; t1 and t2 = Time of initial and final measurements, respectively.
2.4. Statistical analysis
Data were tested by analysis of variance at a 5% significance level, followed by Tukey's post hoc test to compare means using BioEstat 5.0 software.
3. Results and Discussion
Total chlorophyll concentrations in Carapa guianensis increased from 60 DAIT as an effect of reducing the amount of light. The 70% shading treatment had the highest total chlorophyll content, having about three times more total chlorophyll than the 50% and 30% shading treatments and five times more than the full sun irradiation treatment at the end of the experiment (Figure 1A).
The chlorophyll a/b ratio also responded to shading, increasing this ratio from 90 DAIT. However, the treatment under full sun showed a decrease in this parameter, reaching proportions below 1 (Figure 1B), meaning a higher content of chlorophyll b than chlorophyll a in this treatment.
The lack of shading also affected the efficiency of light utilization by C. guianensis seedlings, as indicated by the reduction in the maximum photochemical efficiency of photossystem II (PSII) (Fv/Fm) full sun irradiation treatment. After 90 DAIT, this treatment showed lower Fv/Fm and a more significant reduction than the other treatments, ranging from 0.79 to 0.70 until the end of the experiment. On the other hand, the shading level accentuate the differences in Fv/Fm up to 300 DAIT, presenting an inversely proportional relation to luminosity, being the treatment at 70% of shading more efficient in the use of light, keeping the values of Fv/Fm relatively constant Fm throughout the experiment (Figure 1C).
At the end of the experiment, the shading level promoted higher rates of photosynthetic assimilation (A), stomatal conductance (gs), and transpiration rate (E), with emphasis on treatments at 30% and 50% of shading, compared to complete sun plant (Figure 1D to F).
The dynamics in the photosynthetic parameters adjustments affected the intrinsic water use efficiency (WUEi), and all shaded treatments were more efficient for water consumption than the treatment in the full sun at 90 DAIT. In the 70% and 30% shading, there was an increase of 33% and 28% respectively in WUEi compared to plants exposed to the sun (Figure 2A), indicating that the shading allowed the optimization of the stomatal opening process and the production of photoassimilates.
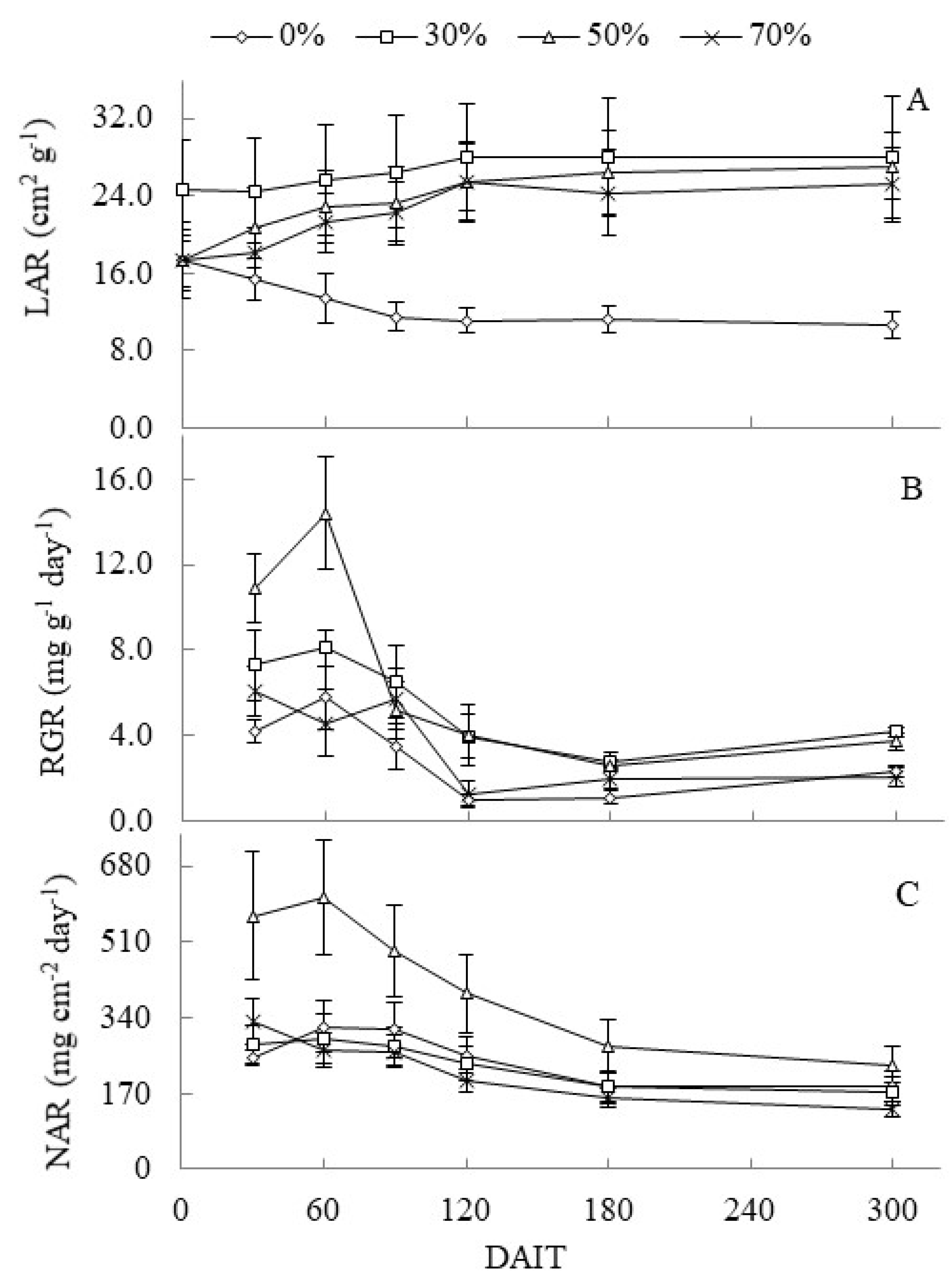
The leaf area ratio (LAR) in the complete sun treatment decreased during the experiment period, while shade treatments increased their functional leaf area for photosynthesis to 120 DAIT. At the end of the experiment the LAR of shady plants was approximately 60% higher than that observed in complete sun plants (Figure 3A). The RGR and net assimilation rate (NAR) showed similar dynamics. The peaks of these parameters occurred at 60 DAIT in the 50% treatment, followed by deceleration until 300 DAIT (Figure 3B and 3C).
3. Discussion
Another Amazonian species showed similar behavior, such as baru (Dipteryx alata Vog.) or macacaporanga (Aniba parviflora (Meisn.) Mez) seedlings, which increased chlorophyll contents when exposed to 70% shading [11,18,19]. Investment in chlorophyll and carotenoids in response to shading is expected [23], acting as a compensatory measure of the species to increase the efficiency of the use of available light [24]. Furthermore, under high irradiance, the degradation rate can surpass the chlorophyll regeneration and synthesis capacity, causing oxidative damage to photosystem II [25].
The reduction in total chlorophyll (Figure 1A) observed in this treatment mainly affected the concentration of chlorophyll a, possibly due to the slow degradation of chlorophyll b [24], or change in the process of converting chlorophyll b to chlorophyll a, strategy used for dissipation of absorbed energy in photosystem II and protection against photodamage under high radiation conditions [26].
In a study with Calendula officinalis L., the authors also observed higher Fv/Fm ratios in shaded plants compared to plants in full sun [27]. This result is possibly related to higher levels of chlorophyll since the Fv/Fm ratio reflects the proportion of energy from solar radiation absorbed by chlorophylls [24]. Furthermore, [26] also observed a reduced chlorophyll a/b ratio associated with reduced photosynthesis efficiency. According to [27], another mechanism for the photoprotection of plants in full sunlight is the increase in the carotenoid content in relation to chlorophyll, which may have happened to the plants in this study.
According to [28], light, when in excess, can act both as a subsidy and an inhibitor of photosynthesis, being associated with alteration or low acclimatization capacity of the photosystem in light receptors, which can cause photoinhibition in plants, reflecting on the Fv/Fm ratio. The plants in full sunlight in this experiment showed symptoms of photoinhibition stress (Figure 1C), as the Fv/Fm ratio below 0.8 is indicative of photoinhibition, and the lower, the more intense the effect [29]. The inverse situation would be the maintenance of the Fv/Fm ratio at high levels, which indicates the adaptation of the leaves to the solar radiation conditions to which they are submitted [30]. This characteristic was the behavior of seedlings exposed to 70% shading (Figure 1C), pointing to the best adaptability of this species in the initial stages in shaded environments.
According to [30], in the Amazon, seedlings, and trees in the juvenile stage are highly prone to undergo PSII photoinhibition or leaf photooxidation when subjected to intense solar radiation. In these cases, intense solar radiation possibly generates excessive energy production in the photosynthesis process, causing a high degree of stress and, consequently, damage to the photosynthetic apparatus [31]. This experiment characterized these consequences in complete sun treatment plants that showed low chlorophyll production, mainly with a reduction in chlorophyll a, and high symptoms of photoinhibition stress indicated by the low Fv/Fm ratio. These changes put pressure on the photosynthetic apparatus of these plants, reducing the gas exchange capacity of the exposed complete sun plants (Figure 1).
Although generally, the highest photosynthetic rate is observed in plants in environments with greater luminosity [19], in cases of plants under shade, as pointed out in this study, the photosynthetic productivity can be compensated by maximizing the capture of available light rays. Thus, together with the greater use of thermal control provided by shading, it reflects on stomatal conductance and reduces water loss, decreasing the photorespiratory rate and increasing the photosynthetic rate [19].
The low photosynthetic rate observed in plants under the full sun may be a consequence of the lower concentration of chlorophyll-a in relation to chlorophyll b observed in this treatment (Figure 1B), as according to [26], to avoid photodamage from excess light, plants seek to reduce PS II light-absorbing antenna complexes by reducing the concentration of chlorophyll b, as its degradation depends on the lack of this component. However, in the case of this experiment, there was no reduction in the concentration of chlorophyll b, indicating that the light absorption rate most likely remained high, higher than its energy dissipation capacity, causing photodamage. In addition, high irradiance stress also causes a reduction in transient responses of chlorophyll a fluorescence, one of those responsible for the success of photosynthetic activity [32].
The low gs observed in complete sun treatment may occur due to a drop in the vapor pressure deficit, causing stomatal closure to avoid water loss [9] or also due to temperatures outside the ideal range for carrying out photosynthesis, which is one of the main factors responsible for the low gas exchange capacity of plants in full sun. High leaf temperature is associated with more significant photosynthetically active radiation (PAR). Leaves more exposed to light absorb more energy from the environment and, depending on the energy dissipation capacity, can generate an environment of low relative humidity, consequently generating higher leaf temperatures high, which acts directly on the increase in the deficit of vapor pressure and stomatal closure [30].
The relationships between the gas exchange parameters observed in C. guianensis seedlings (Figure 1D to F) are primarily in agreement with the close relationship between A and gs due to the feedback effect of the intercellular CO2 concentration with the stomatal opening [30] and secondly with the fact that by absorbing CO2 for the photosynthetic process, the stomatal opening allows more easily the stomatal opening. This fact determines the loss of water from plants, affecting transpiration rates [9].
However, from 180 DAIT, all treatments suffered a significant reduction in WUEi, in the proportion of 61% for the 70% shading treatment, more significant reduction among treatments, resulting in inequality between shading levels at the end of the experiment (Figure 2A). Shaded treatments also showed the highest carboxylation efficiency (CUE) compared to complete sun treatment (Figure 2B). The lowest CUE value observed in complete sun treatment is probably due to the photoinhibition observed from the Fm/Fv ratio (Figure 1C).
The water use efficiency for Enterolobium contortisiliquum (Vell.) was not affected the influence of light intensity, despite a higher photosynthetic rate for plants under full sun [24]. However, as this study observed at 90 DAIT, [25] verified the effect of luminosity on the WUEi, in young mahogany plants (Swietenia macrophylla King), in which there was higher WUEi in shade plants. In environments with intense solar radiation, water use efficiency can be reduced by stomatal close through a strategy carried out by the plant to avoid excessive water loss through transpiration [33]. However, the drop observed in the 70% treatment for this parameter is probably a reflection of the low rate of photosynthesis of this treatment, as, despite the high stomatal activity, the amount of light available probably did not compensate for the stomatal opening.
The low CUE observed in full sunlight is probably a reflection of the physiological performance of these plants under this condition, as according to [32], there is a relationship of photochemical efficiency of photosystem II in the energy dissipation capacity because while the photosynthetic apparatus suffers photodamage, its carbon fixation efficiency reduces. Therefore, the highest CUE was presented in shaded environments at 30 % and 50 % of shading, which reflected in a higher relative growth rate (RGR).
Seedlings of Swietenia macrophylla King present lower LAR under whole radiation, with averages 35% lower than that observed in plants under shade [25]. In the same way was observed that leaf area inversely proportional to the amount of light available for andiroba (Carapa guianensis Aubl.), sombrero (Clitoria fairchildiana R.A.Howard), and inga-bean (Inga edulis Mart.) seedlings [29].
The increase in leaf area in plants under low irradiance represents a morphological mechanism to maximize the use of available light [11], producing leaf structures that are more efficient in capturing light [24]. In these cases, there is a relationship with a higher concentration of chlorophyll per unit of leaf area, which also has the function of balancing the photo-destructive effects on plants under high light intensity [25]. The larger leaf area of C. guianensis seedlings in this study exposed to shading levels reinforces this mechanism. It corroborates the behavior observed by [9], who attribute this result to this species's more remarkable phenotypic plasticity.
A larger leaf area favors increased photosynthetic efficiency and allows for more significant biomass accumulation [15], reflected through the RCR and NAR values. There is a positive correlation between plant growth rate and photosynthesis [12]. In this study, the photosynthetic rate (A) influenced the leaf area dynamics and the growth rate, with higher LAR and NAR, both in treatments and periods with higher A. Consequently, influenced the RGR due to the importance of photosynthesis efficiency for the production of dry matter.
The characteristic of increasing biomass allocation in shaded plants has also been observed in other species [11]. As in this study, [23] found a higher RGR under the shade with a significant difference in complete sun treatment in ironwood plants (Caesalpinia ferrea Mart. ex. Tul. var. leiostachya Benth.), a species classified as early secondary. Tabebuia aurea seedlings also showed lower NAR in full sun environments compared to shaded environments, but without significant differences between them, indicating physiological plasticity of the species [16].
The high anatomical plasticity of C. guianensis was demonstrated under different light conditions [34]. This study indicates that this species also has physiological plasticity, showing high adaptability under shading. By providing shade, C. guianensis seedlings responded gradually to physiological adjustments that increased the carbon assimilation efficiency and conversion to growth, both in leaf area and in biomass gain.
Although the ecological classification presents C. guianensis as a late secondary species, which indicates low tolerance to shaded environments, including in the juvenile stage [35], in this study, we demonstrate that in certain circumstances, this species has the capacity to adapt to the shading, suggesting that the luminosity tolerance spectrum of this species is more remarkable than predicted. In view of these results, C. guianensis would not need to go through a hardening stage, which is generally recommended for forest seedlings in nurseries, as they have good levels of elasticity in terms of variation in the amount of light, being able to adapt from shaded to full sun, or from the nursery environment to lighted planting areas.
5. Conclusions
In this study, we demonstrate that C. guianensis seedlings have a broad spectrum of adaptation to light. In cultivation situations, the provision of 30 to 70% of shading promotes favorable physiological adjustments that increase the efficiency of light use with investment in photosynthetic process and higher plant productivity-increasing their growth rates, and these plants cultivated in different shading treatments have physiological development parameters at satisfactory levels from 180 days of nursery.
In nursery situations, cultivating C. guianensis seedlings in moderately shaded environments, providing 30 to 50% of shade, can intensify productivity and plant development. At the same time, the cultivation of this species in full sun is not recommended as the stimulus favors the photoinhibition stress and impair seedling growth
Author Contributions
Conceptualization, A.S.T and E.S.T; methodology, A.S.T and E.S.T; formal analysis, A.S.T, E.S.T, K.G.S and P.B.C; investigation, A.S.T and S.T.L.T; writing—original draft preparation, A.S.T, E.S.T, K.G.S, P.B.C, D.P and S.T.L.T; writing—review and editing, A.S.T, E.S.T, K.G.S, P.B.C, D.P and S.T.L.T; ; supervision, E.S.T.; project administration, E.S.T.; funding acquisition, E.S.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Pro-Rector of Research, Graduate Studies and Technological Innovation (Proppit) and Promotion Program for Course Completion Works – PROTCC of the Federal University of West Pará through public notices for research projects and quotas for scientific initiation scholarships. Additionally, the payment of article processing charges (APC) was funded through resources provided by the Call for Proposals 03/2022/PROPPIT/UFOPA-PAPCIQ, Program for Support of Qualified Scientific Publications.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Houghton, R. A.; Baccini, A.; Walker, W. S. Where is the residual terrestrial carbon sink?. Global change biology, 2018, 24(8), 3277-3279. [CrossRef]
- Field, C. B.; Behrenfeld, M. J.; Randerson, J. T; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science, 1998, 281(5374), 237-240. [CrossRef]
- Huntingford, C.; Zelazowski, P.; Galbraith, D.; Mercado, L. M.; Sitch, S.; Fisher, R., ... & Malhi, Y. Simulated resilience of tropical rainforests to CO 2-induced climate change. Nature Geoscience, 2013, 6(4), 268. [CrossRef]
- Levine, N. M.; Zhang, K.; Longo, M.; Baccini, A.; Phillips, O. L; Lewis, S. L.; ... & Feldpausch, T. R. Ecosystem heterogeneity determines the ecological resilience of the Amazon to climate change. Proceedings of the National Academy of Sciences, 2016, 113(3), 793-797. [CrossRef]
- Ahlström, A.; Canadell, J. G.; Schurgers, G.; Wu, M; Berry, J. A; Guan, K; &Jackson, R. B. Hydrologic resilience and Amazon productivity. Nature communications, 2017, 8(1), 387. [CrossRef]
- Mitchard, E. T. A. The tropical forest carbon cycle and climate change. Nature, 2018, v. 559, n. 7715, p. 527-534. [CrossRef]
- Boulton, C.A.; Lenton, T.M.; Boers, N. Pronounced loss of Amazon rainforest resilience since the early 2000s. Nat. Clim. Chang, 2022, 12, 271–278. [CrossRef]
- Dalanhol, S. J.; Nogueira, A. C.; Gaiad, S.; Kratz, D. Efeito de micorrizas e da fertilização no crescimento de mudas de Campomanesia Xanthocarpa (Mart.) O. Berg., produzidas em diferentes substratos. Ciência Florestal, 2017, v. 27, n. 3, p. 931-945. [CrossRef]
- Camargo, M. A. B.; Marenco, R. A. Growth, leaf and stomatal traits of crabwood (Carapa guianensis Aubl.) in central Amazonia. Revista Árvore, 2012, v.36, n. 1, p. 7-16. [CrossRef]
- Klimas, C. A.; Kainer, K. A.; De Oliveira Wadt, L. H. The economic value of sustainable seed and timber harvests of multi-use species: An example using Carapa guianensis. Forest Ecology and Management, 2012, v. 268, p. 81-91. [CrossRef]
- Mota, L. H. De S.; Scalon, S. De P. Q.; Heinz, R. Sombreamento na emergência de plântulas e no crescimento inicial de Dipteryx alata Vog. Ciência Florestal, 2012, v. 22, n. 3, p. 423-431. [CrossRef]
- Magalhães, N. S.; Marenco, R. A.; Camargo, M. A. B. Do soil fertilization and forest canopy foliage affect the growth and photosynthesis of Amazonian saplings?. Scientia Agricola, 2014, v. 71, n. 1, p. 58-65. [CrossRef]
- Dutra, T. R.; Massad, M. D.; Santana, R. C. Parâmetros fisiológicos de mudas de copaíba sob diferentes substratos e condições de sombreamento. Ciência Rural, 2012, v. 42, n.7, p. 1212-1218. [CrossRef]
- Colebrook, E. H.; Thomas, S. G.; Phillips, A. L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. Journal of experimental biology, 2014, v. 217, n. 1, p. 67-75. [CrossRef]
- Park, Y.; Runkle, E. S. Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environmental and Experimental Botany, 2017, v. 136, p. 41-49. [CrossRef]
- Souza Pinto, J. R.; Dombroski, J. L. D.; De Freitas, R. M. O. Crescimento e índices fisiológicos de Tabebuia aurea (Manso) Benth. & Hook., sob sombreamento no semiárido. FLORESTA, 2016, v. 46, n. 4, p. 465-472. [CrossRef]
- Fiorucci, A. S.; Fankhauser, C. Plant strategies for enhancing access to sunlight. Current Biology, 2017, v. 27, n. 17, p. R931-R940. [CrossRef]
- Felsemburgh, C. A.; Santos, K. J. S. Dos; Camargo, P. B. De; Carmo, J. B. Do; Tribuzy, E. S. Respostas ecofisiológicas de Aniba parviflora ao sombreamento artificial. Pesquisa florestal brasileira, 2016, v. 36, n.87, p. 201-210.
- Tribuzy, A.S.; Tabaldi, L.A.; Tribuzy, E.S.; Soares, J.C.R.; Finger, C.A.G. Crescimento e acúmulo de biomassa de Luehea divaricate Mart. & Zucc. Submetida ao sombreamento. Revista Ibero-Americana de Ciências Ambientais, 2022, v. 13, n. 3, 250-259. [CrossRef]
- Pinho, E. K. C.; Lopes, A. N. K.; Costa, A. C.; Silva, A. B. V.; Vilar, F. C. M.; Reis, R. G. E. Substratos e tamanhos de recipiente na produção de mudas de baruzeiro (Dipteryx alata Vog.). Ciência Agrícola, 2018, v. 16, n. 1, p. 11-19. [CrossRef]
- Alvares, C. A.; Stape, J. L.; Sentelhas, P. C.; Gonçalves, J. L. De M.; Sparovic, G. Köppen’s climate classification map for Brazil. Meteorologische, 2013, v. 22, n.6, p. 711–728. [CrossRef]
- Aquino, A. C. M. D. S.; Silva, M. H. M. D.; Rocha, A. K. S.; Castro, A. A.; Azevedo, G. F. C. Estudo da influência de diferentes tempos e métodos de cocção na estabilidade dos teores de clorofila e ácido ascórbico em brócolis (Brassica oleraceae). Scientia Plena, 2011, v. 7, n.1, p. 1-6.
- Lenhard, N. R.; Paiva Neto, V. B. de; Scalon, S. de P. Q.; Alvarenga, A. A. de. Crescimento de mudas de pau-ferro sob diferentes níveis de sombreamento. Pesquisa Agropecuária Tropical, 2013, v. 43, n.2, p. 178-186. [CrossRef]
- Souza, J. P.; Melo, N. M. J.; Halfeld, A. D.; Reis, J. N. Shading effects on leaf life span and functional traits in the widespread species Enterolobium contortisiliquum (Vell.) Morong. Acta Scientiarum. Biological Sciences, 2017, v. 39, n. 1, p. 113-122. [CrossRef]
- Gonçalves, J. F. De C.; Silva, C. E. M. da; Justino G. C.; Nina Jr A. da R. Efeito do ambiente de luz no crescimento de plantas jovens de mogno (Swietenia macrophylla King). Scientia Forestalis, 2012, v. 40, n. 95, p. 337-344.
- SATO, R.; ITO, H.; TANAKA, A. Chlorophyll b degradation by chlorophyll b reductase under high-light conditions. Photosynthesis research, 2015, v. 126, n. 2, p. 249-259. [CrossRef]
- Casierra-Posada, F.; Ávila-León, O. F. Shade tolerance of marigold plants (Calendula officinalis). Revista UDCA Actualidad & Divulgación Científica, 2015, v. 18, n. 1, p. 119-126. [CrossRef]
- Li, L.; Aro, E. M..; Millar, A. H. Mechanisms of photodamage and protein turnover in photoinhibition. Trends in plant science, 2018, v. 23, n. 8, p. 667-676. [CrossRef]
- Moura-Silva, C. E.; Andrade, P. C. R.; Dos Santos, W. N. Influence of luminosity on the initial growth of Carapa guianensis, Clitoria fairchildiana and Inga edulis tree seedlings. Científica, 2017, v. 45, n. 4, p. 422-429. [CrossRef]
- Marenco, R. A.; Vera, S. A. A.; Gouvêa, P. R. Dos S.; Camargo, M. A. B.; Oliveira, M. F. De; Santos, J. K. Da S. Fisiologia de espécies florestais da Amazônia: Fotossíntese, respiração e relações hídricas. Ceres, 2014, v. 61, n. 7, p. 786-799. [CrossRef]
- Tian, Y.; Yuan, H.; Xie, J.; Zheng, Y. Shade tolerance and suitability of tree species for planting in rubber plantations. Southern Forests: A Journal of Forest Science, 2016, v. 78, n. 1, p. 11-18. [CrossRef]
- Solovchenko, A.; Solovchenko, O.; Khozin-Goldberg, I.; Didi-Cohen, S.; Pal, D.; Cohen, Z.; Boussiba, S. Probing the effects of high-light stress on pigment and lipid metabolism in nitrogen-starving microalgae by measuring chlorophyll fluorescence transients: Studies with a Δ5 desaturase mutant of Parietochloris incisa (Chlorophyta, Trebouxiophyceae). Algal Research, 2013, v. 2, n. 3, p. 175-182. [CrossRef]
- Matthews, J. S.; Vialet-Chabrand, S. R.; Lawson, T. Diurnal variation in gas exchange: The balance between carbon fixation and water loss. Plant Physiology, 2017, v. 174, n. 2, p. 614-623. [CrossRef]
- Aragão, D. de S.; Lunz, A. M. P.; Oliveira, L. C. de; Raposo, A.; Fermino Jr., P. C. P. Efeito do sombreamento na anatomia foliar de plantas jovens de andiroba (Carapa guianensis Aubl.). Revista Árvore, Viçosa, 2014, v. 38, n. 4, p. 631-639. [CrossRef]
- Lima, R. B. de A.; Silva, J. A. A. da; Marangon, L. C.; Ferreira, R. L. C.; Silva, R. K. S. Da. Sucessão ecológica de um trecho de floresta ombrófila densa de terras baixas, Carauari, Amazonas. Pesquisa Florestal Brasileira, 2011, v. 31, n. 67, p. 161.
Figure 1.
Evolution in the evaluation period of total chlorophyll (Total Chlr) expressed in mg.g-1 (figure 1A), chlorophyll a/b ratio (Chlr a/b) (figure 1B), maximum photochemical efficiency of photosystem II (Fv/Fm) (figure 1C ), maximum photosynthetic rate (A) in .mol.m-2.s-1 (figure 1D), stomatal conductance (gs) in mol.m-2.s-1 (figure 1E) and transpiration (E) in mmol.m-2.s-1 (figure 1F) in C. guianensis seedlings under different shading levels as a function of days after treatment induction (DAIT). The bars indicate the standard deviation of the mean.
Figure 1.
Evolution in the evaluation period of total chlorophyll (Total Chlr) expressed in mg.g-1 (figure 1A), chlorophyll a/b ratio (Chlr a/b) (figure 1B), maximum photochemical efficiency of photosystem II (Fv/Fm) (figure 1C ), maximum photosynthetic rate (A) in .mol.m-2.s-1 (figure 1D), stomatal conductance (gs) in mol.m-2.s-1 (figure 1E) and transpiration (E) in mmol.m-2.s-1 (figure 1F) in C. guianensis seedlings under different shading levels as a function of days after treatment induction (DAIT). The bars indicate the standard deviation of the mean.
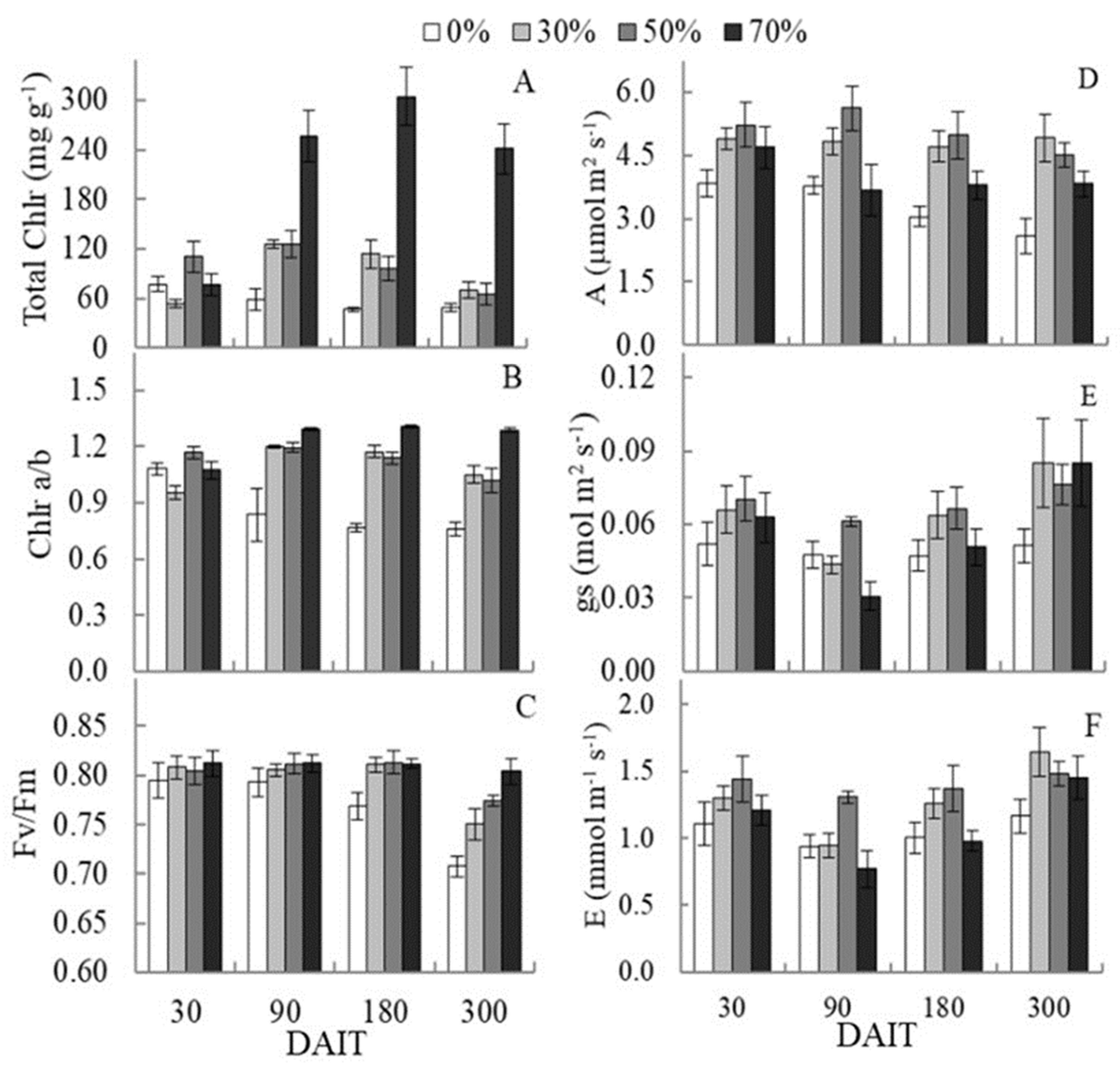
Figure 2.
Variation of intrinsic water use efficiency (WUEi) expressed in (µmolCO2.m-2.s-1)(mmolH2O.m-2.s-1)-1 (figure 2A) and carbon use efficiency (CUE) in (µmol.m-2.s-1)(μmol.mol-1)-1 (figure 2B) in C. guianensis seedlings under different light levels throughout the experiment period. The bars indicate the standard deviation of the mean.
Figure 2.
Variation of intrinsic water use efficiency (WUEi) expressed in (µmolCO2.m-2.s-1)(mmolH2O.m-2.s-1)-1 (figure 2A) and carbon use efficiency (CUE) in (µmol.m-2.s-1)(μmol.mol-1)-1 (figure 2B) in C. guianensis seedlings under different light levels throughout the experiment period. The bars indicate the standard deviation of the mean.
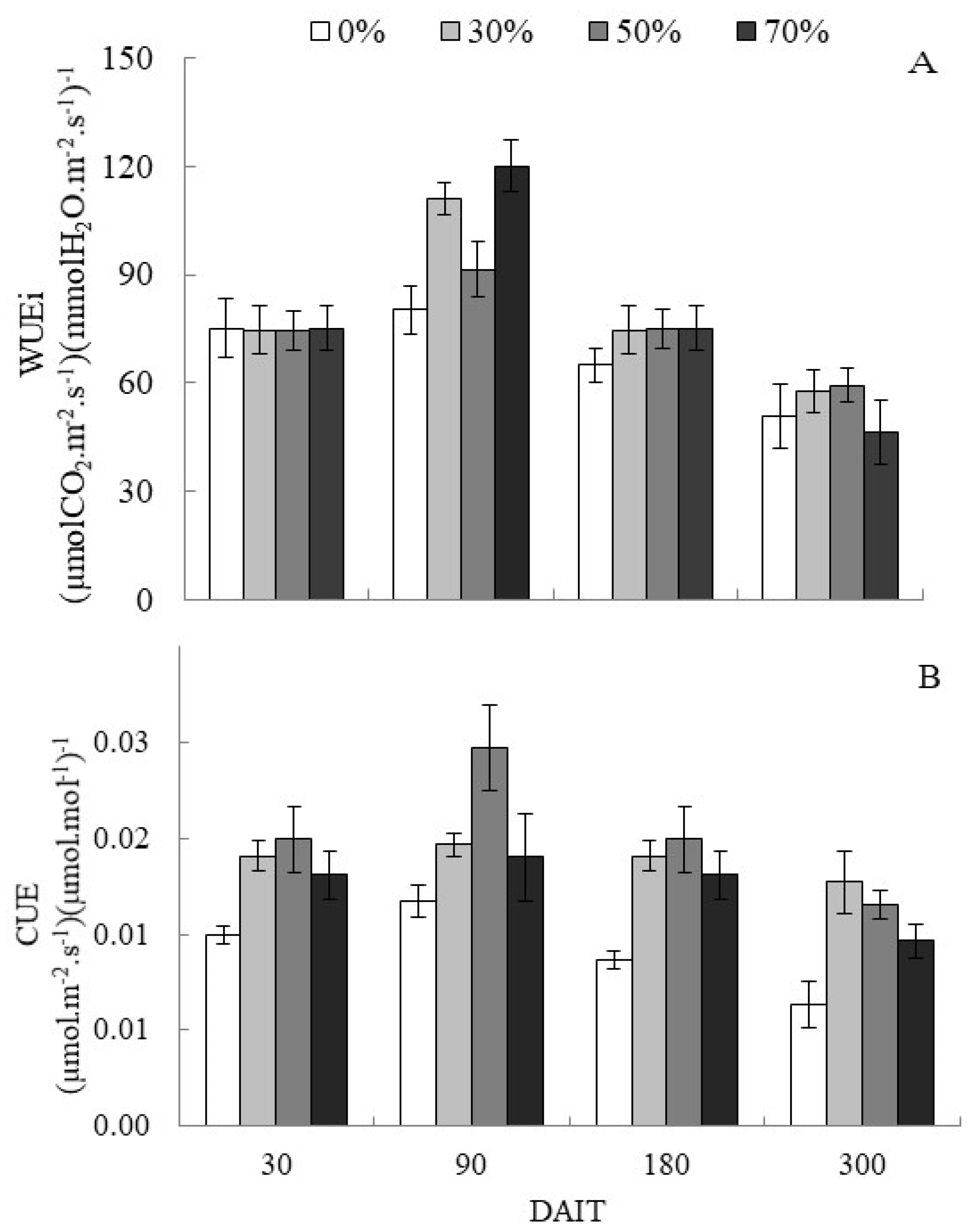
Figure 3.
Mean values of leaf area ratio (LAR) expressed in (cm2 g-1) (figure 3A), relative growth rate (RCR) in (mg.g-1.day-1) (figure 3B) and net assimilation rate (NAR) in (mg.cm-2 day-1) (figure 3C) of C. guianensis seedlings under different levels of shading as a function of days after induction to treatments (DAIT).
Figure 3.
Mean values of leaf area ratio (LAR) expressed in (cm2 g-1) (figure 3A), relative growth rate (RCR) in (mg.g-1.day-1) (figure 3B) and net assimilation rate (NAR) in (mg.cm-2 day-1) (figure 3C) of C. guianensis seedlings under different levels of shading as a function of days after induction to treatments (DAIT).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



