
Submitted:
05 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
Background: The immunomodulatory role of 1,25-Dihydroxy vitamin D3 (1,25(OH)2D3) is exerted through its interaction with the Vitamin D receptor (VDR) present on pancreatic and immune cells. While a deficiency in Vitamin D has been linked to Type 1 Diabetes Mellitus (T1DM), the exact molecular mechanism driving this down-regulation in T1DM is yet to be fully understood. This study aimed to decipher differences in the expression of genes associated with Vitamin D metabolism in T1DM patients and to ascertain if there is a correlation between serum 1,25(OH)2D3 levels and the expression of these genes. We also sought to understand the influence of specific microRNAs (miRNAs) on the expression of Vitamin D metabolism genes in peripheral blood mononuclear cells (PBMCs) of T1DM patients. Furthermore, the study delved into the potential implications of altered Vitamin D metabolism genes and miRNAs on autoimmune processes. Methods: Utilizing real-time PCR, we assessed the expression profiles of genes encoding for 1-hydroxylases (CYP27B1) and 24-hydroxylases (CYP24A1), as well as related miRNAs, in PBMCs from 30 T1DM patients and 23 healthy controls. ELISA tests facilitated the measurement of 1,25(OH)2D3, GAD65, and IA-2 levels. Results: Our findings showcased down-regulated CYP27B1 mRNA levels, while CYP24A1 expression remained stable compared to healthy subjects (CYP27B1, p = 0.0005; CYP24A1, p = 0.205, respectively). In T1DM patients, the levels of has-miR-216b-5p were found to be increased, while the levels of has-miR-21-5p were decreased in comparison to the control group. Notably, no correlation was identified between the expression of CYP27B1 in T1DM patients and the levels of has-miR-216b-5p, has-miR-21-5p, and 1,25(OH)2D3. A significant negative correlation was identified between CYP27B1 mRNA levels in PBMCs of T1DM and IA2, but not with GAD65. Conclusions: The study highlights there were reduced levels of both CYP27B1 mRNA and has-miR-21-5p, along with elevated levels of has-miR-216b-5p in the PBMCs of T1DM. However, the absence of a correlation between the expression of CYP27B1, levels of has-miR-216b-5p, and the status of 1,25(OH)2D3 suggests the possible existence of other regulatory mechanisms. Additionally, the inverse relationship between IA2 autoantibodies and CYP27B1 expression in T1DM patients indicates a potential connection between this gene and the autoimmune processes inherent in T1DM.
Keywords:
Type 1 diabetes mellitus
; 1
; 25-Dihydroxy Vitamin D
; PBMCs
; 1α-hydroxylase (CYP27B1)
; 24-hydroxylase (CYP24A1)
; miRNA
; Ongoing Islet Autoimmunity
1. Introduction
Type 1 Diabetes Mellitus (T1DM) is a chronic autoimmune disease characterized by the degeneration of beta cells, resulting in insulin deficiency [1]. T1DM incidence is strongly associated with Vitamin D deficiency, and 1,25(OH)2D3 can potentially prevent islet cell death and enhance insulin production. Several studies suggest that low 1,25(OH)2D3 levels may affect beta cell regulation [2]. Numerous epidemiological studies have reported a high prevalence of Vitamin D deficiency among Saudi children with T1DM, indicating a strong correlation between these two factors [3]
The Vitamin D metabolism pathway is controlled by various genes, including CYP2R1, CYP27B1 (responsible for activation), 24-hydroxylase (CYP24A1) (responsible for inactivation), VDR (responsible for action), and GC. The concentration of 1,25(OH)2D3, the physiologically active form of Vitamin D, is tightly regulated by both 1-hydroxylase and the catabolic enzyme 24-hydroxylase (encoded by the CYP24A1 gene). CYP24A1 catalyzes the hydroxylation reaction leading to the degradation of 1,25(OH)2D3, resulting in the excretion of calcitroic acid and other metabolites in the bile. CYP24A1 expression is induced by both 1,25(OH)2D3 and 25(OH)D3, making it one of the most highly inducible genes in humans, capable of increasing its transcription by 20,000-fold [4].
Vitamin D deficiency might enhance autoimmune responses in the context of T1DM. 1,25(OH)2D3 can exert its immunomodulatory effects through genomic responses and its ability to alter gene transcription, as most human cells contain Vitamin D receptors (VDR) [5]. Numerous immune cells express VDRs and CYP27B1 enzymes, the synthesis of which is influenced by various immune-specific signals [6]. In autoimmune diseases, the Vitamin D metabolite plays a crucial role in downregulating all mechanisms associated with adaptive immunity, inducing immunological tolerance, and promoting anti-inflammatory activity [7]. Therefore, understanding the molecular mechanism responsible for Vitamin D deficiency in T1DM patients is paramount.
miRNAs are small, non-coding RNA molecules that post-transcriptionally regulate gene expression, often by preventing or triggering the degradation of mRNAs [8]. miRNAs that target genes involved in Vitamin D metabolism can impact the circulating 1,25(OH)2D3 levels in individuals with T1DM. Several miRNAs have been predicted to target CYP24A1, but only has-miR-125b-5p has been experimentally validated [9].Studies conducted on ovarian granulosa and breast cancer cells have shown that overexpression or antisense knockdown of has-miR-125b-5p suppresses and enhances the expression of CYP24A1 protein, respectively [9]. Besides its role in Vitamin D catabolism, has-miR-125b-5p also targets the VDR. Reducing has-miR-125b-5p expression enhances the response to 1,25(OH)2D3 in melanoma cell lines [10]. Conversely, increased has-miR-125b-5p levels suppress endogenous VDR protein levels in MCF-7 breast cancer cells, contributing to resistance to 1,25(OH)2D [11]. While other miRNAs may potentially target VDR, only one, miR-326, has been validated in the peripheral blood lymphocytes of individuals with T1DM [12]. Considering the proposed role of 1,25(OH)2D3 in protecting against autoimmunity [13], it is reasonable to speculate that miR-326 may inhibit the immunomodulatory effects of 1,25(OH)2D3 in preventing inflammatory and autoimmune disorders. Moreover, studies have shown that miR-125a-5p is upregulated in Treg cells isolated from the lymph nodes draining the pancreas of T1DM patients. This upregulation potentially contributes to the reduced expression of its target gene, C-C Chemokine Receptor type-2 (CCR2)[14].
The Vitamin D -activating enzymes, CYP27B1 is predicted to be targeted by multiple miRNAs, although only one has been experimentally validated. Several studies have shown that miR-21 inhibits CYP27B1 expression in monocytes infected with Mycobacterium leprae (M. Leprae), thus inhibiting downstream antibacterial responses induced by Vitamin D intracrine signaling [15]. Another miRNA predicted to target CYP27B1 is has-miR-216b-5p. T1DM is associated with increased miR-216a expression in pancreatic islets, possibly as a compensatory mechanism [16]. Both miR-377 and miR-216a have been identified as early biomarkers of nephropathy in children with T1DM. Their correlation with carotid intimal thickness (CIMT) provides insights into the subclinical atherosclerotic processes in diabetic nephropathy [17].
Accumulating evidence suggests that miRNAs are involved in T1DM pathogenesis through multiple mechanisms, including the regulation of immune cell differentiation, development, activation, and the disruption of immune system equilibrium. Overexpression of miR-34a in diabetic mice reduces B lymphopoiesis capacity, disturbs pancreatic islet defense, and increases sensitivity to damage [18]. Differential miRNA expression also influences the production of specific T lymphocytes. Moreover, miR-26 and miR-101 have been associated with the differentiation of cells toward the T helper 1 (Th1) phenotype. Additionally, miR-21, miR-93, miR-326, and miR-31 are believed to alter T cell functions and play a role in T1DM autoimmunity [19,20].
Several miRNAs have been associated with T1DM in peripheral blood mononuclear cells (PBMCs). Patients with recently diagnosed T1DM exhibited significant downregulation of miR-21a and miR-93, which target NF-KB signaling to regulate apoptosis and inflammation [21]. Furthermore, miR-326 is overexpressed in PBMCs from T1DM patients [22,23], suggesting the involvement of miRNAs in T1DM autoimmunity as they target significant immune modulators—VDR and erythroblastosis virus E26 oncogenic homolog 1 (ETS-1). Additionally, miRNA signatures in PBMCs were correlated with autoantibodies in T1DM patients, with increased miR-326 levels correlated with antibodies against glutamic acid decarboxylase (GAD) and tyrosine phosphatase-like protein (IA2) [22,23], and reduced miR-146a levels correlated with antibodies against GAD [21]. Detecting miRNAs in PBMCs offers the advantage of using them as biomarkers for monitoring disease progression.
The cause of T1DM is not fully understood, but is believed to be the development of autoantibodies and autoreactive Th1 and cytotoxic T lymphocytes (CTLs), which cause the immune system to destroy insulin-producing pancreatic cells [1,24]. Vitamin D deficiency appears to contribute to increased activation of B cells and autoantibody production, and long-term supplementation with Vitamin D led to an increase in T-regulator cells in individuals with SLE [25,26].
Here, we hypothesize that irregular miRNA expression in PBMCs from patients with T1DM might influence the downstream target genes involved in Vitamin D metabolism. This could potentially play a significant role in the development of T1DM and 1,25(OH)2D3 insufficiency. Additionally, circulating miRNAs and Vitamin D-related genes may affect the pathogenesis of T1DM by modulating autoimmune responses.
In this study, we aimed to ascertain whether there are differential expressions of Vitamin D metabolism genes between patients with T1DM and healthy control. Additionally, we sought to identify any potential correlations between circulating serum 1,25(OH)2D3 concentrations and the expression levels of these metabolic genes in individuals with T1DM. We further investigated the potential influence of circulating miRNAs on the expression of vitamin D metabolism genes within the PBMCs of those with T1DM. Notably, alterations in vitamin D metabolism genes and associated miRNAs could hold significance in relation to the autoimmune aspect of T1DM.
2. Results
2.1. Comparison of Demographic Characteristics and 1,25(OH)2D3 Levels of Patients with Type 1 Diabetes and Healthy Controls
Table 1 displays the demographic characteristics and 1,25(OH)2D3 levels of patients with T1DM and healthy controls. The mean age of the T1DM patients was 10.71 ± 6.072 years, compared to 9.296 ± 3.006 years for the healthy controls. There were no significant differences in age, sex, and BMI between the T1DM patients and healthy controls.
1,25(OH)2D3 serum levels were significantly lower in T1DM patients than in healthy controls, with values of 24.37 ± 17.14 ng/ml and 43.18 ± 50.89 ng/ml, respectively. A notable 53.33% of T1DM patients were classified as having insufficient 1,25(OH)2D3 levels (<20 ng/ml), whereas no deficiency was observed among the healthy controls. Specifically, the rate of 1,25(OH)2D3 deficiency was 3.33% in T1DM patients compared to zero in healthy controls, with a p-value of 0.0045, indicating statistical significance (Table 1). Figure 1 provides a comprehensive overview of the variance in Vitamin D status between individuals with T1DM and those without, underscoring the differences in Vitamin D levels between the two groups.
Data are presented as either medians with interquartile ranges or as means ± standard deviations. Abbreviations: BMI, Body Mass Index; T1DM, Type 1 Diabetes Mellitus; HbA1c, Glycated Hemoglobin; GAD, Glutamic Acid Decarboxylase; IA2, Islet Antigen-2, a tyrosine phosphatase-like protein. Reference values are 5 IU/mL for GAD65, and 10 IU/ml for IA2; 1,25-Dihydroxy vitamin D3 (1,25(OH)2D3); p < 0.05 for comparison between T1DM and control groups.
2.2. CYP27B1 mRNA Expression is Downregulated in PBMCs from T1DM Patients Compared to Healthy Controls, While CYP24A1 Remains Unchanged
Given the decreased 1,25(OH)2D3 levels in T1DM patients compared to healthy controls, we explored the expression profile of Vitamin D-metabolism genes in PBMCs of T1DM. The levels of CYP27B1 mRNA (Figure 2A) were significantly downregulated in T1DM patients in comparison to healthy controls (p = 0.0005), whereas no significant changes were observed in the expression of CYP24A1(p = 0.205) (Figure 2B).
2.3. No Correlation Observed Between 1,25(OH)2D3 serum levels and Expression Levels of CYP24A1 and CYP27B1 mRNA
To assess whether 1,25(OH)2D3 status has an impact on the expression of CYP24A1 and CYP27B1 in PBMCs of T1DM patients and healthy control, we analyzed the correlation between serum 1,25(OH)2D3 levels and the expression levels of CYP24A1 and CYP27B1. However, no significant correlations between 1,25(OH)2D3 and the Vitamin D metabolism genes were identified in either the T1DM or the control group (Figure 3A–D).
2.4. Circulating Levels of has-miR-21-5p and has-miR-216b-5p, but not has-miR-125b-5p, are Differentially Expressed in PBMCs of T1DM Patients Compared with Healthy Controls
Based on our findings, which indicated no correlation between serum 1,25(OH)2D3 concentrations and CYP24A1 and CYP27B1 mRNA levels in PBMCs of T1DM patients, we further investigated whether epigenetic factors such as miRNAs might play a role in regulating CYP27B1 expression. We observed a significant upregulation in the expression of hsa-miR-216b-5p, while hsa-miR-21-5p was down-regulated in PBMCs of T1DM patients compared to healthy controls. However, no significant differences in the expression of hsa-miR-125b-5p were observed between T1DM patients and controls (Figure 4A–C).
2.5. CYP27B1 Not Correlated with hsa-miR-216b-5p and hsa-miR-21-5p in T1DM
Given the differential expression of hsa-miR-216b-5p, hsa-miR-21-5p, and CYP27B1 in T1DM, we performed a correlation analysis to determine whether these miRNAs were associated with CYP27B1 expression. In T1DM patients, we observed no significant correlations between CYP27B1 mRNA level and hsa-miR-216b-5p (P = 0.6, rs = 0.09, Figure 5A) or hsa-miR-21-5p (P = 0.2, rs = -0.23, Figure 5B).
2.6. Correlation Between miRNAs and CYP27B1 with Islet Autoantibodies
Given the differential expression of miRNAs and CYP27B1 in PBMCs of individuals with T1DM, we investigated whether these factors could correlate with islet autoantibodies such as GADA65 and IA2. Analysis of associations between hsa-miR-216b-5p and hsa-miR-21-5p expression and titers of islet autoantibodies (GADA65 and IA2A) revealed no significant correlations (Figure 6A,B and Figure 7A,B). IA2 titers negatively correlated with CYP27B1 (Figure 8A), whereas GADA65 titers showed no correlation with CYP27B1 (Figure 8B).
3. Discussion
Vitamin D is pivotal in modulating autoimmune diseases, including diabetes. Its deficiency has been implicated in the pathogenesis of T1DM. In this context, we analyzed whether serum 1,25(OH)2D3 concentrations and Vitamin D metabolic genes' expression correlate in T1DM patients. Additionally, we investigated if miRNAs might modulate the expression of these Vitamin D metabolism genes in PBMCs of T1DM, given that miRNAs targeting genes in the Vitamin D metabolism pathway could influence circulating 1,25(OH)2D3 levels in T1DM. To the best of our knowledge, this is the first study linking circulating miRNA expression with alterations in Vitamin D metabolism genes within PBMCs of T1DM, considering Vitamin D status and ongoing islet autoimmunity.
There is a robust association between T1DM incidence and Vitamin D deficiency [27]. Many epidemiological studies have underscored the prevalence of Vitamin D deficiency among Saudi children diagnosed with T1DM, alluding to a potent connection between the two [3]. Our research findings revealed a heightened 1,25(OH)2D3 deficiency among T1DM patients compared to non-diabetic controls, with 53.33% of T1DM patients exhibiting reduced 1,25(OH)2D3 levels. This is congruent with prior studies highlighting the pervasive nature of 1,25(OH)2D3 deficiency among T1DM patients [28]. The relationship between Vitamin D deficiency and T1DM onset—whether causative or consequential—remains elusive.
Our investigations into a potential link between Vitamin D deficiency in T1DM and Vitamin D metabolism genes led us to compare gene expressions in T1DM patients and healthy individuals. We noted a significant decline in CYP27B1 mRNA levels among T1DM patients, with no observable changes for CYP24A1. Our results are coherent with previous findings showing attenuated CYP27B1 mRNA levels in T1DM patients relative to controls. The implications of the CYP27B1 gene on T1DM pathogenesis are evident as it modulates mRNA expression and influences 1,25(OH)2D3 serum levels, potentially via the −1260 C/A polymorphism [29]. However, our data did not showcase a direct correlation between CYP27B1 mRNA levels and serum 1,25(OH)2D3. There is speculation that a correlation might exist at the protein level. Further research is needed to uncover these connections, suggesting the possibility of significant breakthroughs ahead.
Given that CYP7B1 expression didn't correlate with 1,25(OH)2D3 status in T1DM, we explored other factors potentially influencing its expression. Prior research suggests epigenetic modifications, such as miRNAs, might impact CYP27B1 gene expression. Given the significant role of miRNAs in T1DM pathogenesis and CYP27B1 expression regulation in other tissues [30], we probed the potential of miRNA in modifying CYP27B1 expression. Bioinformatic predictions posited that has-miRNA-216b-5p and has-miRNA-21-5p could target the 3' UTR of CYP27B1, while has-miRNA-125b-5p might target CYP24A1's 3' UTR. The expressions of influencing miRNAs, governed by factors like BMI and age, were consistent across both groups in our study. Notably, our findings indicated an upregulation of has-miR-216b-5p and a downregulation of has-miR-21-5p, with no alterations for has-miR-125b-5p. Prior research corroborates the differential expression of has-miR-21-5p and has-miR-216b-5p in T1DM [16,21,31,32]. Interestingly, while our results showcased differential expression of specific miRNAs, consistent with previous studies, they did not directly correlate with CYP27B1 in PBMCs of T1DM. This raises the tantalizing possibility of other underlying mechanisms, perhaps genetic polymorphism or insulin resistance, playing a role in modulating CYP27B1 expression.
While the crucial role of miRNAs in preserving immune equilibrium is apparent, the perturbed expression of specific miRNAs and its subsequent contribution to autoimmune diseases introduces new avenues of exploration [33]. The accumulating evidence pointing towards miRNAs' involvement in T1DM pathogenesis—particularly in domains like immune responses and beta-cell metabolism—poses thought-provoking questions [34]. Might other unidentified miRNAs be just as critical, if not more so, in influencing T1DM onset? The prevailing research, emphasizing miRNAs' role in immune stability and the negative impact of their anomalous expression on autoimmunity, raises queries about potential therapeutic interventions harnessing miRNA regulation [33]–[35]. With multiple studies documenting shifts in miRNA expression linked to T1DM pathology, one cannot help but speculate if reversing these alterations could mitigate disease progression [21] [22,23]. Remarkably, the diminished expression of miR-21 and miR-93 in T1DM patient PBMCs, vital to inflammatory and apoptotic pathways, hints at potential therapeutic targets [21]. Given the consistent reduction of has-miR-21-5p in our study and others, can enhancing its expression offer therapeutic benefits? Further comprehensive studies might unravel the potential opportunities within miRNA regulation in T1DM.
Several pivotal antibodies can be detected in T1DM patients' years before the onset of the disease, notably antibodies against IA-2, IA-2b, and GAD. Intriguingly, research suggests that miRNAs may play a crucial role in the biosynthesis of specific auto-antibodies, as a cluster of 32 miRNAs has been found to modulate the expression pattern of these T1DM-related auto-antibodies [36]. However, our findings reveal no correlation between miRNAs and auto-antibodies.
Subsequently, we sought to understand the potential role of reduced CYP27B1 expression on the presence of autoantibodies in patients with T1DM. This investigation entailed correlating CYP27B1 mRNA levels with specific autoantibodies, notably IA2 and GAD65. We discovered a negative association between CYP27B1 mRNA levels and IA-2 autoantibodies, suggesting a compelling possibility: the CYP27B1 gene might play a pivotal role in the autoimmune cascade characteristic of T1DM. Moreover, previous studies have revealed associations between polymorphisms in codon 416 of the Vitamin D-binding protein gene and autoimmune markers of T1DM, particularly IA2 antibodies [37]. Given these findings, it raises the question: Could the intricate interplay of genetic factors, miRNAs, and Vitamin D metabolism be fundamental to the complex etiology of T1DM? Future research endeavors are poised to unravel this intricate network of interactions and illuminate the underlying mechanisms of the disease.
The clinical implications of this study underscore the profound significance of 1,25(OH)2D3 levels and related metabolic gene expressions such as CYP27B1 in the early detection and monitoring of the disease. Furthermore, altering miRNA expression in T1DM may provide promising targets for future pharmaceutical interventions, enhancing the overall management and treatment avenues. Additionally, as the study reveals correlations between CYP27B1 expressions and T1DM, it emphasizes the potential of personalized medicine, wherein treatments could be tailored to the unique genetic profiles of patients. This could revolutionize how T1DM is managed, shifting from generic treatments to more individual-centric ones. Clinicians might be encouraged to adopt a more holistic approach to managing T1DM, considering an array of factors beyond just the traditional metrics of glucose and insulin.
The study presented here offers several commendable strengths, making it a valuable contribution to the current scientific understanding. Firstly, its pioneering nature cannot be understated; this research appears to be the initial foray into connecting circulating miRNA expression with alterations in the Vitamin D metabolism genes, specifically in PBMCs of T1DM, thereby potentially bridging an existing knowledge gap. This comprehensive study approach stands out, as it delves into the surface-level investigation of serum 1,25(OH)2D3 levels and ventures deeper to uncover the underlying genetic and epigenetic mechanisms that might influence these levels in T1DM patients. Furthermore, the depth of analysis is worth noting. The research did not merely stop at identifying genes with altered expressions but extended its exploration to establish correlations with other physiological markers, such as autoantibodies. Considering the burgeoning interest in the role of Vitamin D in T1DM and the increasing significance attached to miRNAs in various disease processes, this study is timely and relevant.
However, like all scientific endeavors, this study is not devoid of limitations. A significant constraint is that the gene expression was gauged only at the mRNA level. While this offers a snapshot, it might not always mirror the protein levels or biological activity, which are critical for a complete understanding. The sample size, being limited, raises concerns about the robustness and generalizability of the findings. Moreover, while the study adeptly uncovers various correlations, the age-old causality conundrum remains. It is yet to be definitively ascertained whether Vitamin D deficiency is a consequence or a precursor to T1DM. Not to mention, there might be lurking variables.
In light of the above, it becomes evident that while this study paves the way, there is a long journey ahead. More extensive cohort studies, augmented with in vitro or animal model experiments, must validate and expand on these preliminary findings. Evaluations at the protein level, coupled with functional assays, might be the key to unlocking a more profound mechanistic understanding of the processes at play.
4. Materials and Methods
4.1. Ethics and Consent
This research project underwent a rigorous review and received approval from the Institutional Review Board at the General Directorate of Health Affairs (GDHA) in Madinah (IRB Number: 276). The GDHA, an official government ethics committee, issues ethical approvals for human research on behalf of the Ministry of Health (MOH). All studies involving human participants were conducted in compliance with the ethical guidelines set forth in the Declaration of Helsinki. Every participant provided written informed consent to participate in the study.
4.2. Study Design, Recruitment of T1DM Patients, Healthy Control, Inclusion and Exclusion Criteria
In this case-control study, patients with T1DM were evaluated for symptoms of diabetes and a casual plasma glucose concentration of 7.0 mmol/L or a 2-hour post-load glucose concentration of 11.1 mmol/L during an oral glucose tolerance test, based on the criteria set by the American Diabetes Society. Age-matched control children were selected from the Maternity and Children's Hospital.
Inclusion criteria for cases included an age group of 0 to 16 years and diagnosed cases of T1DM with classical symptoms (polyuria, polydipsia, and polyphagia) along with a random plasma glucose of ≥ 200 mg/dl. Patients with pancreatic disease, hepatic disease, renal disease, bone diseases, malignancy, and any history of drug use such as calcium and Vitamin D were excluded from the cases.
For healthy controls, individuals with blood sugar within normal limits, normal growth and puberty range, and no endocrine abnormalities or autoimmune conditions were included. Subjects with pancreatic disease, hepatic disease, renal disease, bone disease, malignancy, and any history of drug use such as calcium and Vitamin D were excluded from the control group.
Data, including demographic data, medical history, comorbidities, and medications, were collected for each patient from the medical records at the Maternity and Children's Hospital.
4.3. Isolation of Peripheral Blood Mononuclear Cells (PBMCs)
Blood samples (5 ml) were collected from both T1DM patients and healthy controls into tubes containing EDTA and were diluted 1:1 in PBS. Following this, density gradient separation of the diluted samples was carried out using Hypaque-Ficoll medium (Innotrain, Germany). PBMC samples were isolated through centrifugation of the Hypaque-Ficoll gradient, and fresh PBMCs were immediately utilized for RNA extraction.
4.4. Total RNA and miRNA Extraction from PBMCs
Total RNA, including miRNA, was extracted from the purified PBMCs using the Qiagen miRNeasy Mini Kit (Qiagen, Valencia, CA, USA), following the manufacturer's protocol. The purity and concentration of RNA were determined by obtaining OD260/280 readings using a dual-beam UV spectrophotometer (Eppendorf AG, Hamburg, Germany).
4.5. Reverse Transcription and Quantitative Real-Time PCR
Complementary DNA synthesis from miRNA samples was carried out using the miScript II RT Kit and HiSpec Buffer (Qiagen), following the manufacturer’s instructions. Quantitative Real-Time PCR (qRT-PCR) was conducted using the miScript SYBR Green PCR Kit (Qiagen). The 25 μl reaction included 1 μl of cDNA, forward and reverse primers at optimized concentrations, and RNase-free water to make up the volume. The PCR was set up in a 96-well plate with triplicates, under a UV-irradiated hood on an ABI 7700HT PCR machine (Applied Biosystems 7500 Fast Real-Time PCR System) (Applied Biosystems, USA). Minus RT and no-template controls were included for each assay. The cycling conditions were: 50ºC for 2 min, 95ºC for 10 min followed by 40 cycles of 95ºC for 15 sec and 60ºC for 60 sec, and a melt curve stage (95 °C for 15 s, 60 °C for 1 min, 95 °C for 15 s). All primer sequences for the selected miRNAs and Vitamin D-related genes for real-time RT-PCR analyses are listed in Table S2.
Results from the real-time PCR were computed using the equation [2ˆ(-∆Ct)]. The expression levels of CYP27B1 and CYP24A1 mRNA were normalized to the housekeeping gene, acidic ribosomal phosphoprotein P0 (RPLP0). For miRNAs, hsa-miR-U6B was used as a housekeeping gene to normalize all Ct values. Ct values were determined using the SDS software v.2.1, with manual baseline settings applied to fix the same threshold for both target miRNAs and reference genes
4.6. Serum Sample Collection and 1,25(OH)2D3 Level Measurement
A 3 ml blood sample was centrifuged at 1500g for 10 minutes to obtain serum samples from both patients and controls. These serum samples were then frozen at -80°C until required for analysis. Serum 1,25(OH)2D3 levels were measured using a Vitamin D Kit (IDS, UK) in a consistent laboratory setting. A concentration of 20 ng/ml or higher was considered a normal level of 1,25(OH)2D3. Concentrations between 10 and 20 ng/ml were classified as 1,25(OH)2D3 insufficiency, while levels below 10 ng/ml were categorized as 1,25(OH)2D3 deficiency.
4.7. Serological Analysis
Anti-GAD65 and anti-IA2 antibodies were assessed using enzyme immunoassay (ELISA) with Medizym commercial kits (Berlin, Germany). The detection of antibodies was performed semi-quantitatively, referencing a value of 5 IU/ml for GAD65 and 10 IU/ml for IA2. HbA1c levels were determined using a commercially available automatic system (DCA 2000, Bayer Diagnostics, Tarrytown, NY, USA).
4.8. Target Gene Identification and Bioinformatic Analysis
A comprehensive literature search was undertaken to identify a particular set of miRNAs whose expression has been previously linked with T1DM and are thought to influence the Vitamin D signaling pathway. Our study concentrated on three miRNAs, namely hsa-miR-125b-5p [31,32,38], has-miR-216b-5p [16,17],and has-miR-21-5p [39,40], which have been significantly dysregulated in serum, plasma, and PBMCs of T1DM patients, as identified using bioinformatics tools (TargetScan Human, available at http://www.targetscan.org). Based on the extant literature encompassing in vitro studies, we selected miRNAs potentially interacting with the 3′UTR sequences of CYP24A1 and CYP27B1 (refer to Supplementary table S1). Among several miRNAs that might interact with the CYP24A1 3′UTR sequence, we chose hsa-miR-125b-5p. For the CYP27B1 3′UTR, hsa-miR-216b-5p and hsa-miR-21 were selected. The involvement of these miRNAs in regulating CYP27B1 expression was substantiated by in vitro studies.
4.9. Statistical Analysis
Statistical analyses were conducted using GraphPad Prism 6.0 software (GraphPad Software, Inc. CA, USA). Clinical parameters are represented as mean ± standard deviation (SD). To compare statistical significance between the two groups (T1DM vs. healthy control), the unpaired t-test or Mann–Whitney U test was employed. Expression of miRNA or Vitamin D-related genes is denoted as mean ± standard error. To identify correlations between miRNA expression and mRNA of Vitamin D-related genes in all participants, Pearson or Spearman correlation testing was utilized. A two-tailed p-value (p) < 0.05 was deemed statistically significant for all statistical tests.
5. Conclusions
In conclusion, this study explored the role of Vitamin D and its metabolic genes in T1DM, focusing on the correlation between serum 1,25(OH)2D3 concentrations and Vitamin D related gene. A substantial association between Vitamin D deficiency and T1DM incidence was noted in the investigated patients. The study found a significant decrease in CYP27B1 mRNA levels in T1DM patients, which is crucial for Vitamin D metabolism, but no direct correlation with serum 1,25(OH)2D3 was observed. Further, the research delved into the impact of miRNAs on Vitamin D metabolism genes, revealing differential expression of certain miRNAs, yet without a direct correlation to CYP27B1 in T1DM patients. This highlights potential other mechanisms influencing gene expression.
The study also found no direct correlation between miRNAs and T1DM-related auto-antibodies but discovered a negative association between CYP27B1 mRNA levels and IA-2 autoantibodies, indicating a potential role of the CYP27B1 gene in T1DM autoimmunity. The research prompts inquiries about the intricate interactions between genetic factors, miRNAs, and Vitamin D metabolism in T1DM's complex etiology. Future research is needed to unravel this complexity and the potential therapeutic interventions based on miRNA regulation and Vitamin D metabolism in T1DM.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, H.A. and I.M.; methodology, H.A.; software, H.A; validation, H.A., I.M. and E.A.; formal analysis, H.A.; investigation.; resources, B.E.; data curation, A.A.; B.E and S.A. writing—original draft preparation, H.A.; writing—review and editing, H.A. I.M.; and J.F. visualization, H.A.; supervision,I.M.; project administration, H.A.; funding acquisition, S.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This research project was reviewed and approved by the Institutional Review Board, General Directorate of Health Affairs (GDHA) in Madinah (IRB Number: 276). This official government ethics committee issues ethical approvals on humans on behalf of the Ministry of Health (MOH). Ethical guidelines were followed in all human participant studies per the Declaration of Helsinki. Each participant provided written informed consent to participate in the study.
Informed Consent Statement
Each participant provided written informed consent to participate in the study.
References
- Yoon, J.-W.; Jun, H.-S. Autoimmune destruction of pancreatic beta cells. Am. J. Ther. 2005, 12, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Pozzilli, P.; Manfrini, S.; Crino, A.; Picardi, A.; Leomanni, C.; Cherubini, V.; Valente, L.; Khazrai, M.; Visalli, N. Low levels of 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 in patients with newly diagnosed type 1 diabetes. Horm. Metab. Res. = Horm. und Stoffwechselforsch. = Horm. Metab. 2005, 37, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A. High Prevalence of Vitamin D Deficiency Among Saudi Children And Adolescents With Type 1 Diabetes In Albaha Region, Saudi Arabia; 2017; Vol. 12;
- Tashiro, K.; Abe, T.; Oue, N.; Yasui, W.; Ryoji, M. Characterization of vitamin D-mediated induction of the CYP 24 transcription. Mol. Cell. Endocrinol. 2004, 226, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Rak, K.; Bronkowska, M. Immunomodulatory Effect of Vitamin D and Its Potential Role in the Prevention and Treatment of Type 1 Diabetes Mellitus-A Narrative Review. Molecules 2018, 24. [Google Scholar] [CrossRef] [PubMed]
- Lin, R. Crosstalk between Vitamin D Metabolism, VDR Signalling, and Innate Immunity. Biomed Res. Int. 2016, 2016, 1375858. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xing, W.; Xie, L. Regulatory Roles of MicroRNAs in Diabetes. Int. J. Mol. Sci. 2016, 17, 1729. [Google Scholar] [CrossRef]
- Komagata, S.; Nakajima, M.; Takagi, S.; Mohri, T.; Taniya, T.; Yokoi, T. Human CYP24 Catalyzing the Inactivation of Calcitriol Is Post-Transcriptionally Regulated by miR-125b. Mol. Pharmacol. 2009. [Google Scholar] [CrossRef]
- Essa, S.; Denzer, N.; Mahlknecht, U.; Klein, R.; Collnot, E.M.; Tilgen, W.; Reichrath, J. VDR microRNA expression and epigenetic silencing of vitamin D signaling in melanoma cells. J. Steroid Biochem. Mol. Biol. 2010, 121, 110–113. [Google Scholar] [CrossRef]
- Mohri, T.; Nakajima, M.; Takagi, S.; Komagata, S.; Yokoi, T. MicroRNA regulates human vitamin D receptor. Int. J. cancer 2009, 125, 1328–1333. [Google Scholar] [CrossRef]
- Sebastiani, G.; Grieco, F.A.; Spagnuolo, I.; Galleri, L.; Cataldo, D.; Dotta, F. Increased expression of microRNA miR-326 in type 1 diabetic patients with ongoing islet autoimmunity. Diabetes. Metab. Res. Rev. 2011, 27, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Adorini, L.; Penna, G. Control of autoimmune diseases by the vitamin D endocrine system. Nat. Clin. Pract. Rheumatol. 2008, 4, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Gager, G.M.; Eyileten, C.; Postuła, M.; Nowak, A.; Gąsecka, A.; Jilma, B.; Siller-Matula, J.M. Expression Patterns of MiR-125a and MiR-223 and Their Association with Diabetes Mellitus and Survival in Patients with Non-ST-Segment Elevation Acute Coronary Syndrome. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.T.; Wheelwright, M.; Teles, R.; Komisopoulou, E.; Edfeldt, K.; Ferguson, B.; Mehta, M.D.; Vazirnia, A.; Rea, T.H.; Sarno, E.N.; et al. MicroRNA-21 targets the vitamin D-dependent antimicrobial pathway in leprosy. Nat. Med. 2012, 18, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, Q.; Zhao, H.; Bishop, J.O.; Zhou, G.; Olson, L.K.; Moore, A. miR-216a-targeting theranostic nanoparticles promote proliferation of insulin-secreting cells in type 1 diabetes animal model. Sci. Rep. 2020, 10, 5302. [Google Scholar] [CrossRef] [PubMed]
- El-Samahy, M.H.; Adly, A.A.; Elhenawy, Y.I.; Ismail, E.A.; Pessar, S.A.; Mowafy, M.E.-S.; Saad, M.S.; Mohammed, H.H. Urinary miRNA-377 and miRNA-216a as biomarkers of nephropathy and subclinical atherosclerotic risk in pediatric patients with type 1 diabetes. J. Diabetes Complications 2018, 32, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Margaritis, K.; Margioula-Siarkou, G.; Giza, S.; Kotanidou, E.P.; Tsinopoulou, V.R.; Christoforidis, A.; Galli-Tsinopoulou, A. Micro-RNA Implications in Type-1 Diabetes Mellitus: A Review of Literature. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Scherm, M.G.; Daniel, C. miRNA-Mediated Immune Regulation in Islet Autoimmunity and Type 1 Diabetes. Front. Endocrinol. (Lausanne). 2020, 11, 606322. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z.; Zhou, Z. miRNAs: novel regulators of autoimmunity-mediated pancreatic β-cell destruction in type 1 diabetes. Cell. &Amp; Mol. Immunol. 2017, 14, 488. [Google Scholar]
- Salas-Pérez, F.; Codner, E.; Valencia, E.; Pizarro, C.; Carrasco, E.; Pérez-Bravo, F. MicroRNAs miR-21a and miR-93 are down regulated in peripheral blood mononuclear cells (PBMCs) from patients with type 1 diabetes. Immunobiology 2013, 218, 733–737. [Google Scholar] [CrossRef]
- Sebastiani, G.; Grieco, F.A.; Spagnuolo, I.; Galleri, L.; Cataldo, D.; Dotta, F. Increased expression of microRNA miR-326 in type 1 diabetic patients with ongoing islet autoimmunity. Diabetes. Metab. Res. Rev. 2011, 27, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Azhir, Z.; Dehghanian, F.; Hojati, Z. Increased expression of microRNAs, miR-20a and miR-326 in PBMCs of patients with type 1 diabetes. Mol. Biol. Rep. 2018, 45, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Song, L.-J.; Qin, X.-Y. Advances in the cellular immunological pathogenesis of type 1 diabetes. J. Cell. Mol. Med. 2014, 18, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Ritterhouse, L.L.; Crowe, S.R.; Niewold, T.B.; Merrill, J.T.; Roberts, V.C.; Dedeke, A.B.; Neas, B.R.; Thompson, L.F.; Guthridge, J.M.; James, J.A. B lymphocyte stimulator levels in systemic lupus erythematosus: higher circulating levels in African American patients and increased production after influenza vaccination in patients with low baseline levels. Arthritis Rheum. 2011, 63, 3931–3941. [Google Scholar] [CrossRef]
- Piantoni, S.; Andreoli, L.; Scarsi, M.; Zanola, A.; Dall’Ara, F.; Pizzorni, C.; Cutolo, M.; Airò, P.; Tincani, A. Phenotype modifications of T-cells and their shift toward a Th2 response in patients with systemic lupus erythematosus supplemented with different monthly regimens of vitamin D. Lupus 2015, 24, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Chakhtoura, M.; Azar, S.T. The role of vitamin d deficiency in the incidence, progression, and complications of type 1 diabetes mellitus. Int. J. Endocrinol. 2013, 2013, 148673. [Google Scholar] [CrossRef]
- Bener, A.; Alsaied, A.; Al-Ali, M.; Al-Kubaisi, A.; Basha, B.; Abraham, A.; Guiter, G.; Mian, M. High prevalence of vitamin D deficiency in type 1 diabetes mellitus and healthy children. Acta Diabetol. 2009, 46, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, E.; Brück, P.; Jansen, T.; Pfeilschifter, J.M.; Radeke, H.H.; Badenhoop, K. CYP2R1-, CYP27B1- and CYP24-mRNA expression in German type 1 diabetes patients. J. Steroid Biochem. Mol. Biol. 2007, 103, 807–810. [Google Scholar] [CrossRef]
- Singh, R.; Yadav, V.; kumar, S.; Saini, N. MicroRNA-195 inhibits proliferation, invasion and metastasis in breast cancer cells by targeting FASN, HMGCR, ACACA and CYP27B1. Sci. Rep. 2015, 5, 17454. [Google Scholar] [CrossRef]
- Cheung, R.; Pizza, G.; Chabosseau, P.; Rolando, D.; Tomas, A.; Burgoyne, T.; Wu, Z.; Salowka, A.; Thapa, A.; Macklin, A.; et al. Glucose-Dependent miR-125b Is a Negative Regulator of β-Cell Function. Diabetes 2022, 71, 1525–1545. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, H.; Pan, X.; Wu, W.; Wang, H.; Yan, L.; Zhang, M.; Liu, X.; Xia, S.; Shao, Q. miR-34a and miR-125b are upregulated in peripheral blood mononuclear cells from patients with type 2 diabetes mellitus. Exp Ther Med 2017, 14, 5589–5596. [Google Scholar] [CrossRef] [PubMed]
- Garo, L.P.; Murugaiyan, G. Contribution of MicroRNAs to autoimmune diseases. Cell. Mol. Life Sci. 2016, 73, 2041–2051. [Google Scholar] [CrossRef] [PubMed]
- Assmann, T.S.; Recamonde-Mendoza, M.; De Souza, B.M.; Crispim, D. MicroRNA expression profiles and type 1 diabetes mellitus: systematic review and bioinformatic analysis. Endocr. Connect. 2017, 6, 773–790. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Martin, A.; Adams, B.D.; Lai, M.; Shepherd, J.; Salvador-Bernaldez, M.; Salvador, J.M.; Lu, J.; Nemazee, D.; Xiao, C. The microRNA miR-148a functions as a critical regulator of B cell tolerance and autoimmunity. Nat. Immunol. 2016, 17, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Abuhatzira, L.; Xu, H.; Tahhan, G.; Boulougoura, A.; Schäffer, A.A.; Notkins, A.L. Multiple microRNAs within the 14q32 cluster target the mRNAs of major type 1 diabetes autoantigens IA-2, IA-2β, and GAD65. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 4374–4383. [Google Scholar] [CrossRef] [PubMed]
- Ongagna, J.C.; Pinget, M.; Belcourt, A. Vitamin D-binding protein gene polymorphism association with IA-2 autoantibodies in type 1 diabetes. Clin. Biochem. 2005, 38, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Januszewski, A.S.; Cho, Y.H.; Joglekar, M.V.; Farr, R.J.; Scott, E.S.; Wong, W.K.M.; Carroll, L.M.; Loh, Y.W.; Benitez-Aguirre, P.Z.; Keech, A.C.; et al. Insulin micro-secretion in Type 1 diabetes and related microRNA profiles. Sci. Rep. 2021, 11, 11727. [Google Scholar] [CrossRef]
- Grieco, G.E.; Cataldo, D.; Ceccarelli, E.; Nigi, L.; Catalano, G.; Brusco, N.; Mancarella, F.; Ventriglia, G.; Fondelli, C.; Guarino, E.; et al. Serum Levels of miR-148a and miR-21-5p Are Increased in Type 1 Diabetic Patients and Correlated with Markers of Bone Strength and Metabolism. Non-coding RNA 2018, 4. [Google Scholar] [CrossRef]
- Morales-Sánchez, P.; Lambert, C.; Ares-Blanco, J.; Suárez-Gutiérrez, L.; Villa-Fernández, E.; Garcia, A.V.; García-Villarino, M.; Tejedor, J.R.; Fraga, M.F.; Torre, E.M.; et al. Circulating miRNA expression in long-standing type 1 diabetes mellitus. Sci. Rep. 2023, 13, 8611. [Google Scholar] [CrossRef]
Figure 1.
Distribution of Vitamin D status among study participants with T1DM and healthy controls.
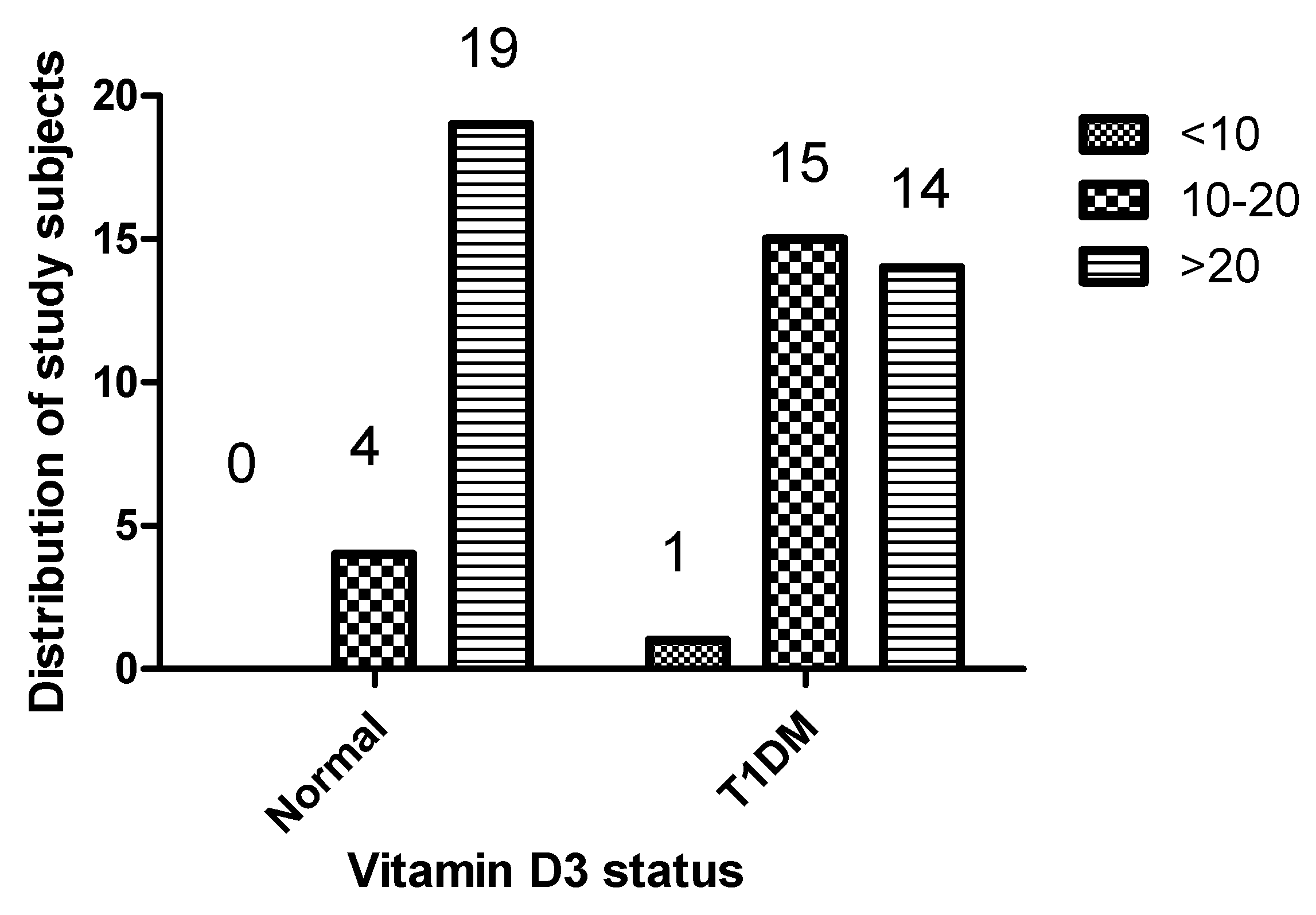
Figure 2.
Expression levels of CYP27B1(A) are decreased in the PBMCs of patients with T1DM, whereas no significant alteration is observed in the expression of CYP24A1(B) across n = 23, healthy control and n = 30, T1DM patients. Data are represented as mean ± SD of normalized 2−ΔCT values, relative to the expression of acidic ribosomal phosphoprotein P0 (RPLP0). The Mann–Whitney U test was employed for statistical analysis, with a significance threshold set at p < 0.05.
Figure 2.
Expression levels of CYP27B1(A) are decreased in the PBMCs of patients with T1DM, whereas no significant alteration is observed in the expression of CYP24A1(B) across n = 23, healthy control and n = 30, T1DM patients. Data are represented as mean ± SD of normalized 2−ΔCT values, relative to the expression of acidic ribosomal phosphoprotein P0 (RPLP0). The Mann–Whitney U test was employed for statistical analysis, with a significance threshold set at p < 0.05.
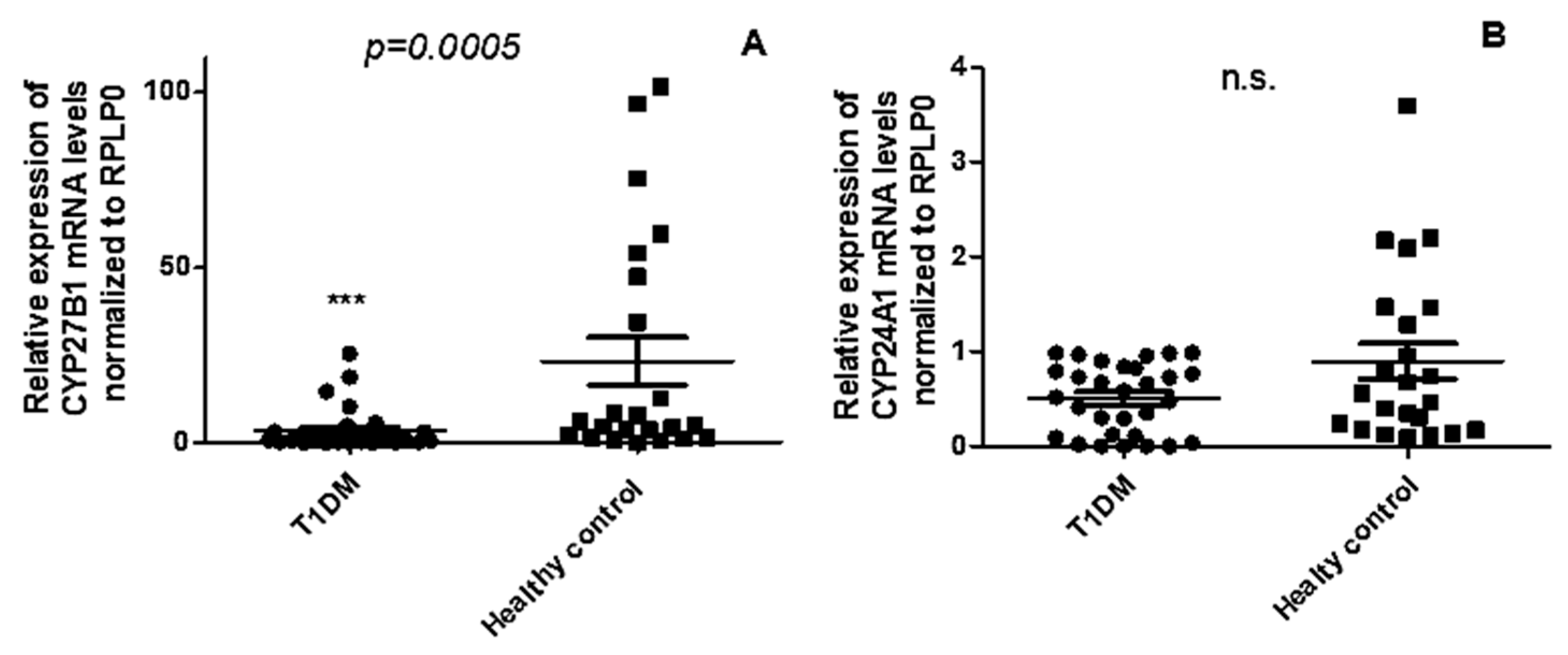
Figure 3.
The expression levels of CYP27B1 (A and B) and CYP24A1 (C and D) did not exhibit any correlation with serum 1,25(OH)2D3 in both T1DM patients and healthy control. The correlation analysis between the expression levels of CYP27B1 and CYP24A1 was represented by normalized 2−ΔCT values, while serum 1,25(OH)2D3 levels were reported in ng/ml. The Spearman R test was employed to determine the r-values and p-values, with significance set at p < 0.05.
Figure 3.
The expression levels of CYP27B1 (A and B) and CYP24A1 (C and D) did not exhibit any correlation with serum 1,25(OH)2D3 in both T1DM patients and healthy control. The correlation analysis between the expression levels of CYP27B1 and CYP24A1 was represented by normalized 2−ΔCT values, while serum 1,25(OH)2D3 levels were reported in ng/ml. The Spearman R test was employed to determine the r-values and p-values, with significance set at p < 0.05.
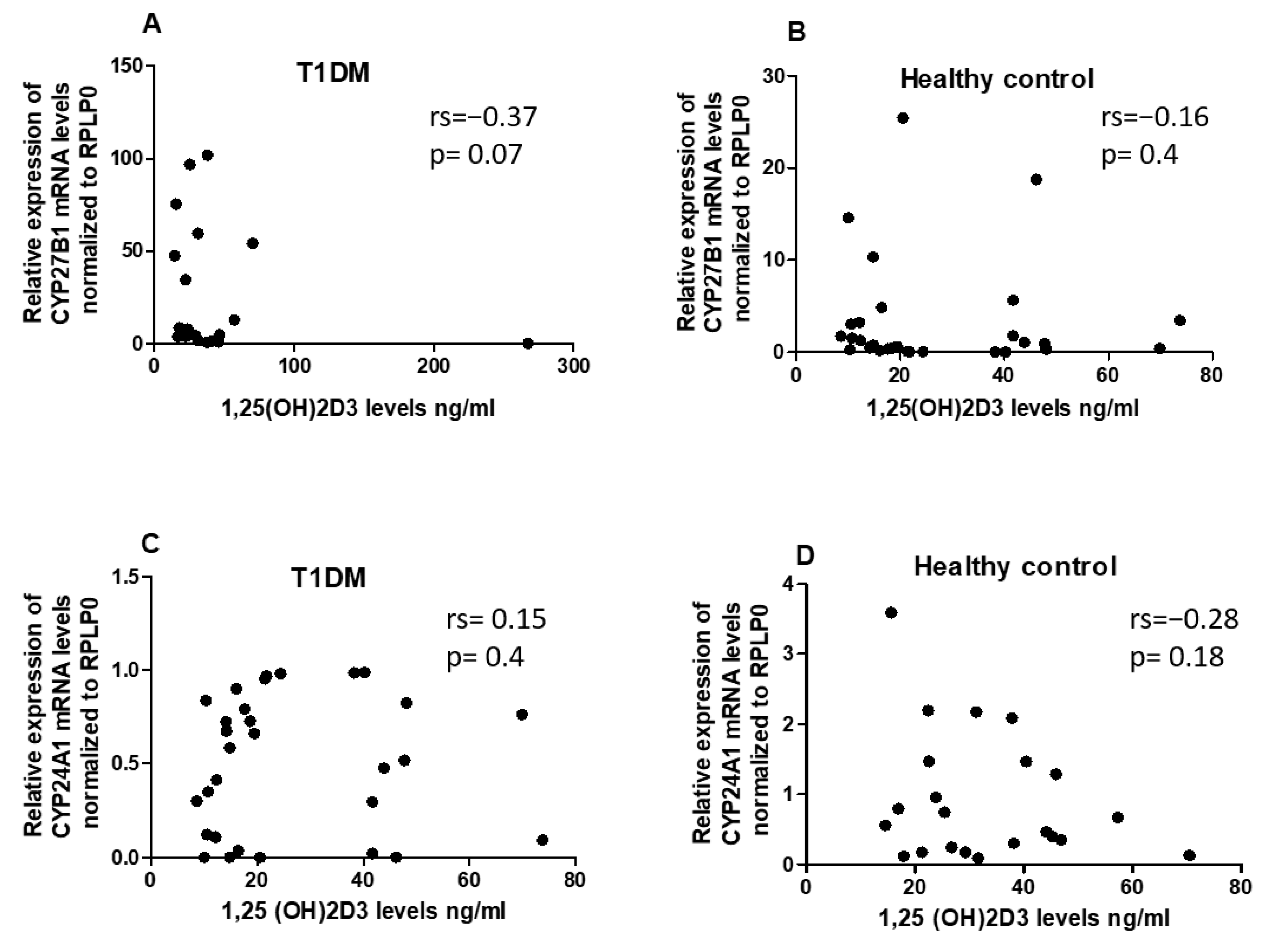
Figure 4.
Expression of miRNA in PBMCs from patients with T1DM and healthy controls. (A) The level of hsa-miR-21-5p (B) hsa-miR-216b-5p, and (C) hsa-miR-125b-5p levels in PBMCs. The error bars represent the standard deviation (SD). * p<0.05 vs controls.
Figure 4.
Expression of miRNA in PBMCs from patients with T1DM and healthy controls. (A) The level of hsa-miR-21-5p (B) hsa-miR-216b-5p, and (C) hsa-miR-125b-5p levels in PBMCs. The error bars represent the standard deviation (SD). * p<0.05 vs controls.
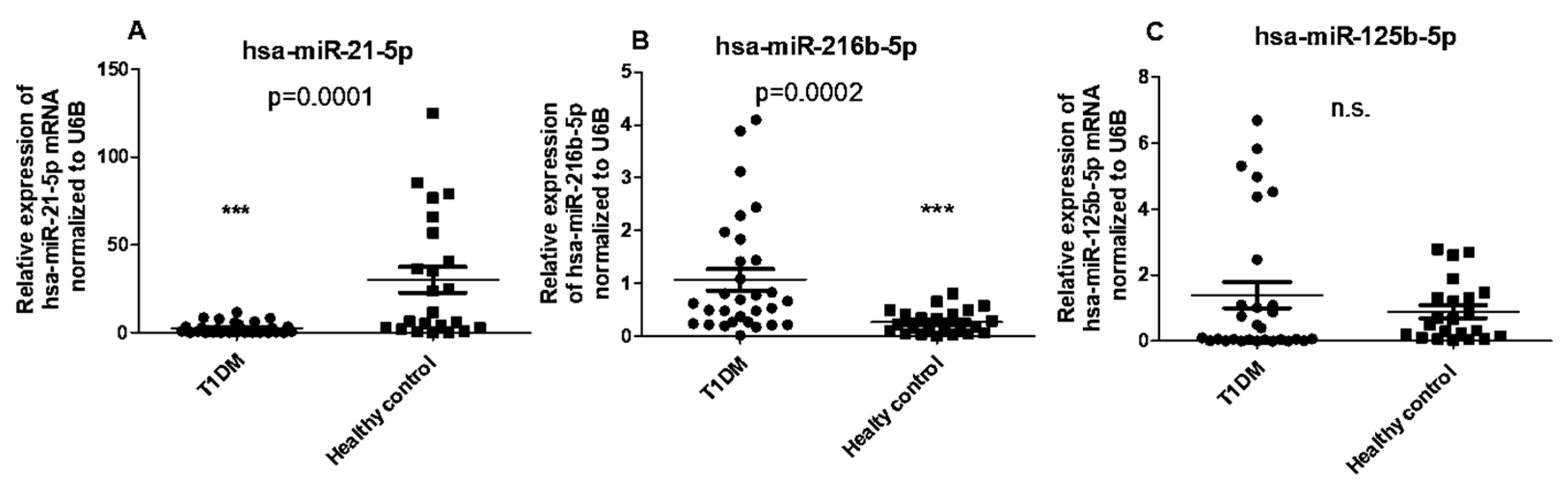
Figure 5.
The expression levels of hsa-miR-216b-5p (A) and hsa-miR-21-5p (B) are not correlated with CYP27B1 expression in T1DM patients. The correlation analysis is reported as normalized 2−ΔCT values. The Spearman R test was performed to evaluate R-values and p-values (p < 0.05).
Figure 5.
The expression levels of hsa-miR-216b-5p (A) and hsa-miR-21-5p (B) are not correlated with CYP27B1 expression in T1DM patients. The correlation analysis is reported as normalized 2−ΔCT values. The Spearman R test was performed to evaluate R-values and p-values (p < 0.05).
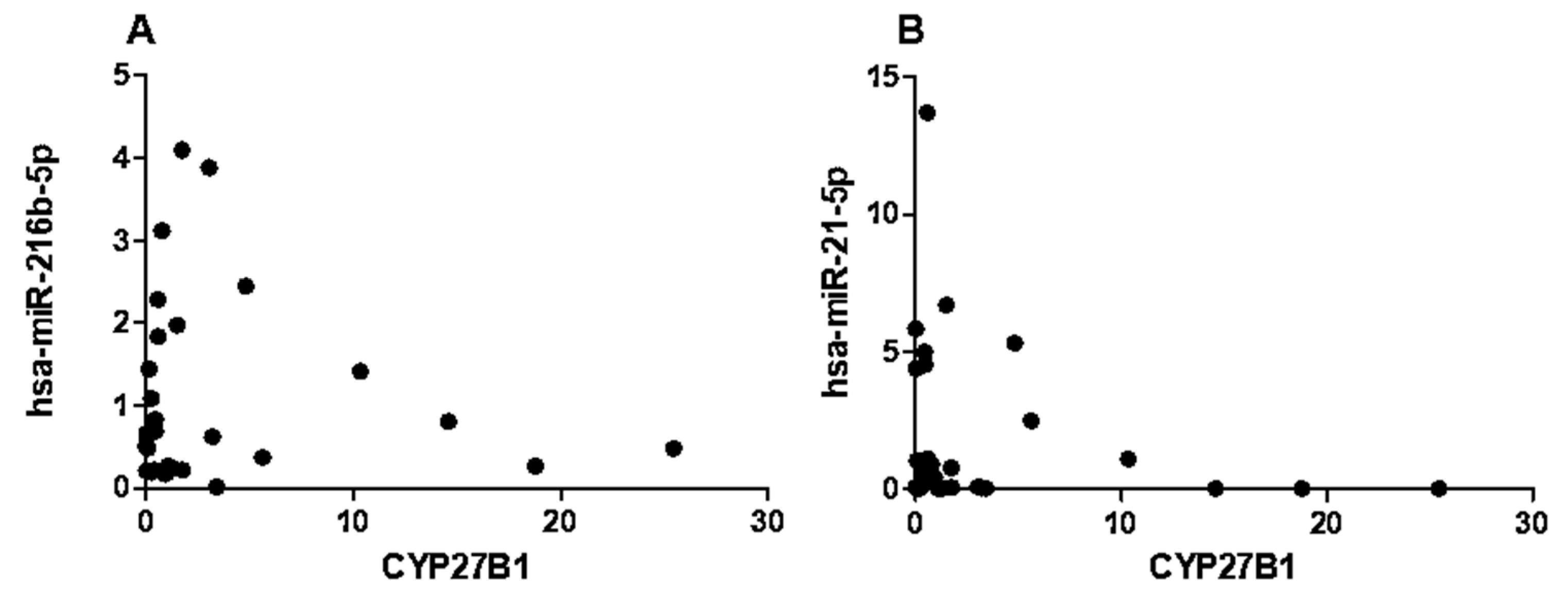
Figure 6.
Correlation of has-miR-216b-5p expression to Islet autoantibodies within the T1DM group. A) IA2, B) GADA65.
Figure 6.
Correlation of has-miR-216b-5p expression to Islet autoantibodies within the T1DM group. A) IA2, B) GADA65.
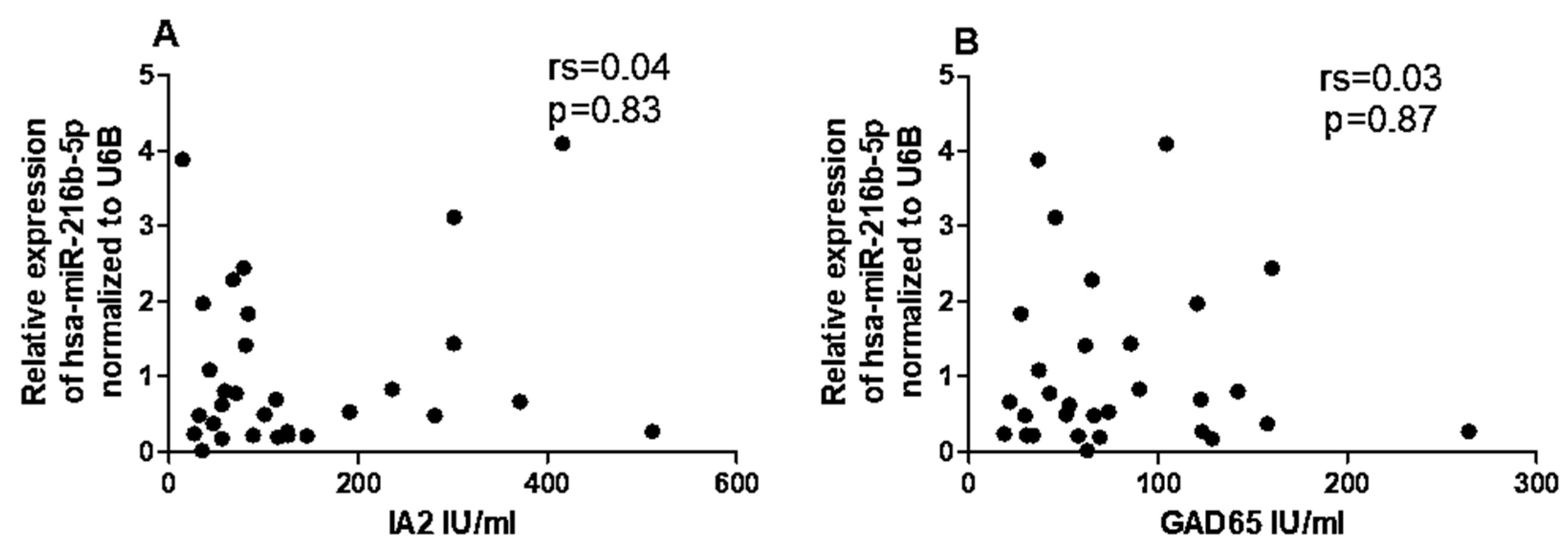
Figure 7.
Correlation of has-miR-21-5p expression to Islet autoantibodies within the T1DM group. A) IA2, B) GADA65.
Figure 7.
Correlation of has-miR-21-5p expression to Islet autoantibodies within the T1DM group. A) IA2, B) GADA65.
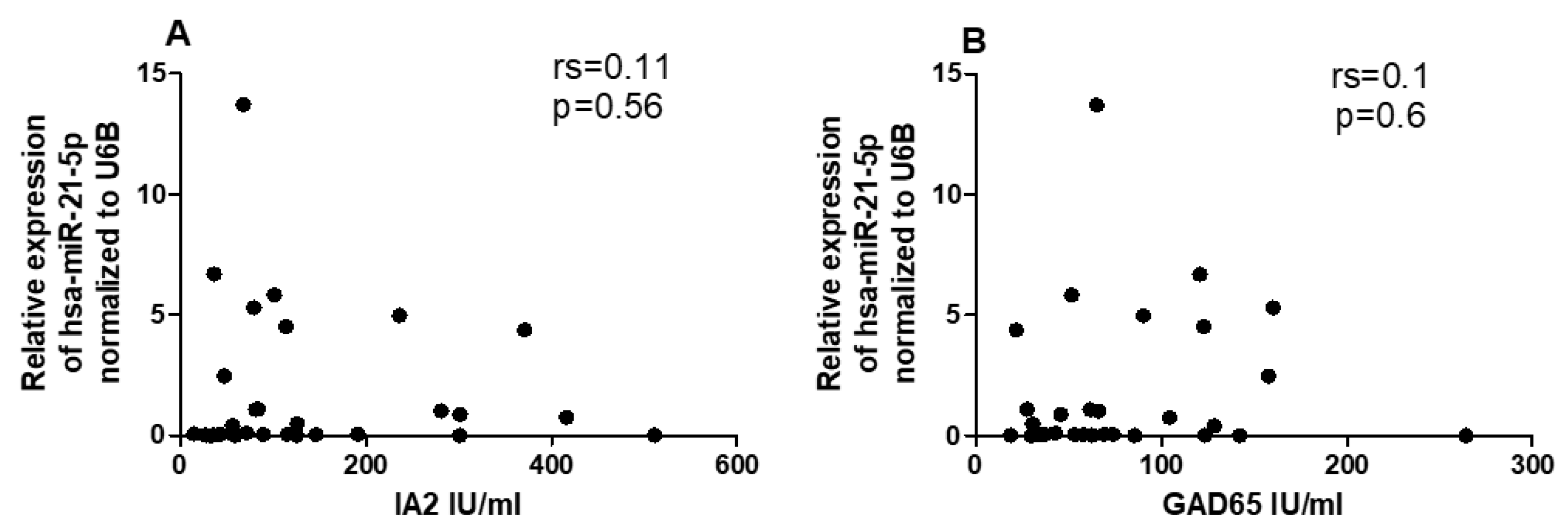
Figure 8.
Correlation of CYP27B1 expression to Islet autoantibodies within the T1DM group. A) IA2, B) GADA65.
Figure 8.
Correlation of CYP27B1 expression to Islet autoantibodies within the T1DM group. A) IA2, B) GADA65.
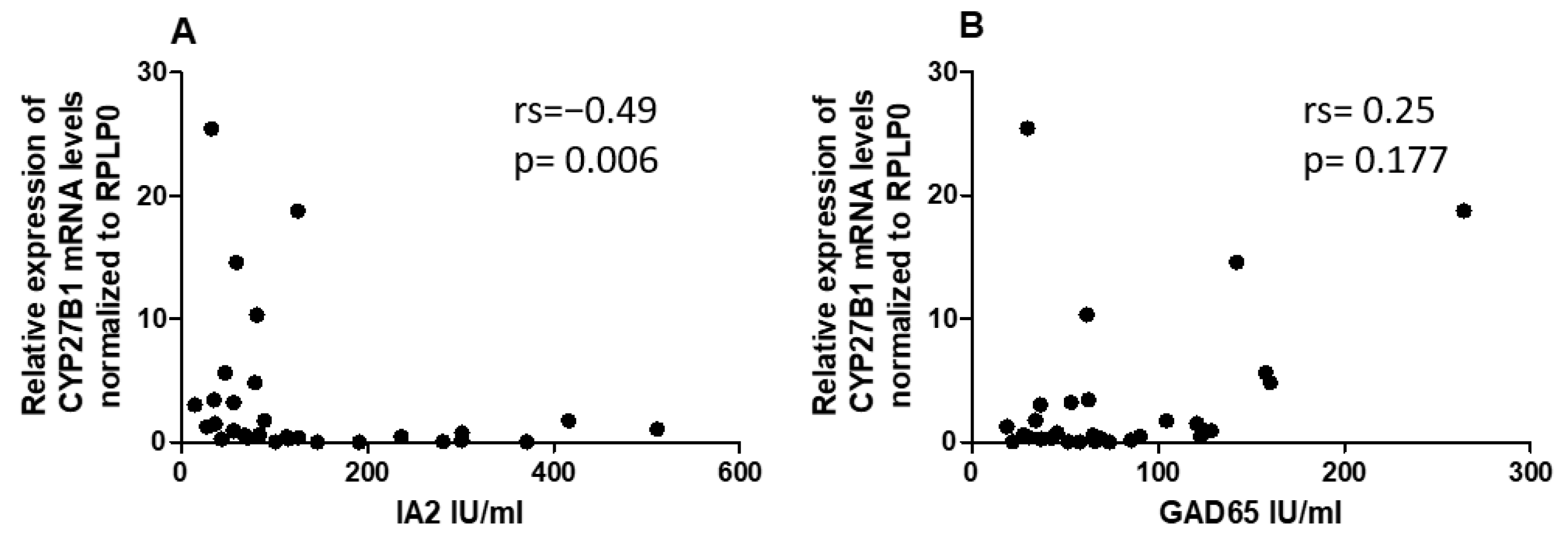

Table 1.
Demographic and clinical features of T1DM patients and healthy control groups.
| Variable | Control group, n = 23 | T1DM group, n = 30 | P-value |
|---|---|---|---|
| HbA1c (%) | 5.126± 0.4158 | 8.957 ±1.353 | <0.0001 |
| IA2 (IU/ml) | 0 | 140.4 (14.45–511.0) | - |
| GAD65 (IU/ml) | 2.62 (0.4–4.7) | 79.55 (18.77–264.3) | - |
| Age (years) | 9.296 ±3.006 | 10.71 ± 6.072 | 0.3098 |
| Sex: male/female | 11/12 | 15/15 | - |
| BMI | 21.86 ±3.250 | 22.31±4.248 | 0.6775 |
| 1,25(OH)2D3 (ng/ml) | 43.18±50.89 | 24.37±17.14 | 0.0045 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



