
Submitted:
04 September 2023
Posted:
06 September 2023
You are already at the latest version
Abstract
Wheat dwarf disease (WDD) is an important disease of monocotyledonous species, including economically important cereals. The causative pathogen, the wheat dwarf virus (WDV), is a monopartite Mastrevirus that has a circular single-stranded (ss)DNA genome and is transmitted persistently and propagatively mainly by the leafhopper Psammotettix alienus. The disease can induce high yield losses in many European countries in a range of 20-100%. Due to climate change the periods of vector activity increased and the vectors spread to new habitats, leading to an increased importance of WDV in large parts of Europe. In the light of integrated pest management approaches cultivation practices and the use of resistant/tolerant host plants are currently the only effective methods for controlling WDV. To date, there is limited knowledge of the pathosystem and epidemiology of the disease. The few sources of genetic tolerance, indicate the need for further research. Considering the economic importance of the disease, and its likely increasing relevance in the coming decades, this review was prepared to compile and update knowledge on aspects of WDV.
Keywords:
wheat dwarf virus
; WDV
; resistance
; mastrevirus
; resistance genes
; geminiviridae
; resistance breeding
1. Introduction
As early as the 8th century a.d., the Japanese Anthology described the first observations of viroses on Eupatorium chinense L., which, according to current knowledge, were caused by geminiviruses [1]. As a consequence of climate change, insect-borne viruses, are gaining increased importance because vectors benefit e.g., from a temperature increase in different ways [2,3,4,5]. The damage caused by viruses in agriculture includes not only yield and biomass losses, but also the weakening of infected plants, making them more susceptible to abiotic and biotic stressors, so that quality losses may also occur [6]. Currently, there are no approved options for direct chemical control of viruses. So, appropriate measures in accordance with an integrated pest management include farm hygiene, quarantine programs for import and export of plant products, production of virus-free seeds and planting material, breeding of resistant varieties and the control of vector insects, as a last measure, by the use of chemical insecticides [7,8].
In Europe, more than 30 different viruses are known to occur in cereals [9]. These include the wheat dwarf virus (WDV, family Geminiviridae, genus Mastrevirus) as the causal agent of wheat dwarf disease (WDD). The virus is transmitted from plant to plant exclusively by leafhoppers [10,11,12]. The first occurrence was described in the former Czechoslovakia [10], followed by subsequent outbreaks in the 1990s [13,14,15,16]. Outbreaks vary from year to year and differ in the damage they cause with early infections in the fall leading to drastic yield losses [17,18]. Lindblad and Waern [17] put the average yield losses in winter wheat fields at 35-90 % for sites studied in Sweden, while a study in southern Finland found losses of 20-100% [18].
Due to the shift in seasons as a result of climate change and the resulting higher temperatures in late fall and February/March [19], a longer infection period can be expected due to a higher vector activity [3], possibly leading to increased disease incidences with higher infection rates in fields. Recent increases in the presence of WDV in European, African, and Asian cereal-growing regions are promoting the research activities with regard to plant resistance in wheat and barley. This article provides an overview of the virus, its vector, and ways of control, with particular emphasis on wheat as a crop.
2. Wheat dwarf virus (WDV)
2.1. Classification and Genomic Organization of WDV
WDV belongs to a group of viruses originally described as wheat dwarfing viruses within the family Geminiviridae, genus Mastrevirus [20,21,22,23].
Geminiviruses themselves are defined as plant pathogenic circular single-stranded DNA (ssDNA) viruses [24]. Their virion consists of twinned (geminate) icosahedra with a bipartite capsid [25,26] and a genome packaged in 11 subunits each [1,26,27]. Besides the nanoviruses (family Nanoviridae), they are the only phytopathogenic representatives with a genome consisting of a circular ssDNA [28]. Actual research on the family Geminiviridae began in the 1980s although they are already known since the beginning of the 20th century mainly as agents of yield loss in tomato, sugar beet, cassava, maize and cotton in tropical and subtropical countries [29,30,31,32]. Based on their genome structure, vector, host range, and phylogeny, geminiviruses are classified into 14 genera with 520 species (Figure 1) [21,22,23,33,34,35].
Currently, 45 different Mastreviruses are known, the type species of which is Maize streak virus (MSV) [34,36]. They predominantly infect monocotyledonous plants, with a few exceptions such as Tobacco yellow dwarf virus [37], Bean yellow dwarf virus [38], and Chickpea redleaf virus [39,40], which can infect susceptible dicotyledonous host plants. Transmission of these viruses to host plants is mainly persistent and non-propagative using leafhoppers as vectors [39,40]. The Mastrevirus genus has a monopartite circular ssDNA genome with a length of 2.6-2.8 kb [41,42]. The genome of WDV [20], which belongs to this group, is 2.73-2.75 kb in size [14,25,43,44].
The circular genome contains two open reading frames (ORFs) on the sense side and two ORFs on the antisense side, which are separated by two noncoding regions that encode four viral proteins. On the virion sense strand, ORFs V1 and V2 are responsible for encoding the viral movement protein (MP) and the coat protein (CP). On the complementary sense are C1 and C2, which encode the replication-associated proteins (Rep, RepA) and are expressed by script splicing [45,46,47,48,49,50]. The two strands are separated by a large (LIR) and a small (SIR) noncoding intergenic unit, whose sequences are substantially involved in viral replication and regulation of gene expression [51] and control bidirectional transcription based on promoter (transcription initiation step) and terminator (transcription termination step) sequences [52,53]. Between the 5’ ends of the Rep/RepA and MP genes is the LIR sequence [54].
The replication-associated proteins (Rep, RepA) are encoded by a gene and expressed by a complementary sense transcript. Both forms differ based upon an intron in the Rep gene [16,48,55,56,57,58] and are involved in the early stages of infection [27]. Rep is involved in viral replication, while RepA affects the control of the host cell cycle to support viral replication [27]. Translation of RepA occurs directly from the native RNA transcript, whereas production of Rep protein requires a splice cut of the RNA molecule. Therefore, the proteins have identical N-terminal sequences [59].
MP, as a product of V2, is a 10.9-kDa protein involved in systemic infection of the host by increasing the exclusion limit of plasmodesmata, allowing intercellular spread of viral DNA [60,61]. The functions of the coat protein (CP) have been studied most extensively for Mastreviruses [62]. In addition to encapsulating viral DNA with a capsid, it is involved in various functions in the infection cycle as well as virus-vector interaction during transmission [59]. Thus, it plays an important role in vector specificity [63], viral nuclear import [64], insect transmission, systemic viral movement, and symptom development [45,62]. For the establishment of systemic infection, both MP and CP (V1 and V2) have been found to be essential, although they do not contribute to virus replication. CP binds ssDNA and dsDNA in vitro in this process, so its presence is essential for the accumulation of viral ssDNA in infected host cells and protoplasts [65].
The geminiviral transcriptional activator protein (TrAP) plays a role in pathogenicity by inhibiting a plant’s transcriptional and posttranscriptional gene silencing [66,67,68,69,70,71,72,73,74]. Enhanced viral replication is initiated by replication enhancer protein (Ren), which interacts with host factors and Rep [63].
2.2. Life cycle of the virus
The life cycle processes of geminiviruses requires host proteins as well as viral ones. Infection of the host plant begins as soon as the virus-bearing insect vector feeds on the host. Deposition and unpacking of the viral genome occur in the phloem companion cells [78,79,80]. Replication of geminiviruses is taking place in the nucleus of companion cells because sieve elements are lacking a nucleus as a consequence of ontogenesis [81]. Entry of viral DNA into the nucleus is supported by the envelope protein (CP). This is thought to interact with host-specific transport receptors. Within the intergenic regions, there are signal motifs controlling the two phases of replication. The onset of DNA synthesis is initiated specifically for representatives of the genus by a primer (approximately 80 bp long) located in the SIR, which is complementary to the intergenic region [46,47]. In the first phase, ssDNA is converted into a double-stranded (ds) DNA intermediate [82], which serves as a template for the production of complementary and virus-sense transcripts [52,53]. Replication of the genomic (+) DNA strand is initiated (ori) by cleavage of the virion-sense strand at a specific highly conserved nona-nucleotide motif (5’ TAATATT ↓ AC 3’) by Rep (replication initiator protein) within the LIR sequence [54,79]. The motif is partially enclosed within the head of a stem-loop structure and contains the initiation point (↓) of the second replication phase to produce the (+) DNA strand using a rolling circle replication process [38,58,82,83,84,85].
For the amplification of viral ds-DNA and the production of ss-DNA genomes, the ds-DNA intermediate is used as a template. Starting from the LIR, passing through the (-) and (+) strand, and continuing to the SIR, the bidirectional transcription of the DNA occurs using host DNA polymerase [86]. Geminiviruses do not code for a DNA polymerase in this process, so production of dsDNA using complementary DNA synthesis depends exclusively on host factors recruited during early stages of replication [79]. Synthesis of the complementary minus (-) DNA strand begins at the 3’ end of a short complementary primer. This is packaged in viral particles and can hybridize to a sequence in the SIR region [82]. Transcription is bidirectional with coding regions diverging from the LIR in both strands. For gene expression, geminiviruses use multiple overlapping transcripts [79].
The movement of the virus depends on the outcome of the interaction with different parts of the cell (cytoskeleton), the type of plasmodesmata, and the ability of the virus to replicate in different cells [87]. In infected plants, electron microscopy has revealed altered nuclei in the phloem companion and in parenchyma cells of roots and leaves [88]. In these cells, an accumulation of virus particles, arranged in groups and rows, is found, filling almost the entire nucleoplasm. High particle concentrations have been detected especially in plants with wilted leaves in the stem region [89].
To spread the infection, the virus must overcome barriers such as the nuclear envelope and spread between adjacent cells [90]. The viral DNA is transported from the nucleus to the cell membrane as a V2-DNA complex with the help of the transport protein (MP), which binds to host receptors [41]. To spread the infection from one cell to another the virus has to pass plasmodesmata. This is exclusively possible between companion cells (CC) and sieve element (SE) of the CC/SE complex because they are isolated from the surrounding phloem parenchyma cells, indicated by a very low number of plasmodesmata in barley [91] and the absence of these in wheat [92]. Depending upon a leaves’ developmental stage the protein size that can pass through plasmodesmata varies, as has been shown for wheat [93]. The authors furthermore demonstrated that a viral movement protein is able to increase the open width of plasmodesmata, so that proteins with a higher molecular weight can pass through, independent of the leaves’ developmental stage. This would facilitate the systemic movement of a virus such as WDV. WDV is distributed together with photoassimilates and other nutrients along the sieve tube, where transport is based upon a turgor driven mass flow from source to sink [90]. For the maize streak virus in maize, it has been shown that younger leaves, formed after inoculation are more likely to be infected with the virus than older leaves, because the viral antigen is distributed according to the age of the tissue. The virus can therefore be detected in the basal meristem of young leaves as it reaches them through the phloem with the metabolites of older leaves. For long distance transport probably only the thin-walled SEs that form above mentioned CC/SE complexes are relevant while thick-walled SEs lack CCs and thus the basis for virus multiplication [94].
Regarding the molecular mechanisms of spread and the associated interaction with host components, many questions remain open in the relationship between geminiviruses and host. Cell-to-cell spread is ensured by phosphorilization of the transport protein (MP) by host kinases [95,96]. A study of begomoviruses (Geminiviridae) in tomato (Lycopersicon esculentum Miller) and soybean (Glycine max [L.] Merr) identified the cellular interaction partners that support transport of the viral genome from the nucleus to the cytoplasm. For both plant species, a membrane-associated plant species-specific kinase belonging to the LRR-RLK family of proteins (leucine rich-repeat receptor like kinase) was discovered. Within the highly specific interaction, short-term formation of a complex of nuclear shuttle protein (NSP) and NSP-interacting kinase (NIK) occurs, which provides targeted and active recognition of nuclear pores, plasma membrane and plasmodesmata modes. The complex presumably serves to regulate the biochemical activity of the viral protein in phosphorylating the transport protein. In this case, NSP would regulate the movement of viral DNA through the kinase activity of transmembrane receptors for this purpose. The host kinase as enzyme and viral NSP as substrate are related here [97]. Therefore, the non-host relationship between the wheat and barley strains of WDV could be due to the non-recognition of the viral protein by the plant receptor. In this case, the low incidence of winter barley infected with the wheat strain and winter wheat infected with the barley strain could be attributed to a sequence swap resulting from a mutation [98].
2.3. Phylogenetics
Based on phylogenetic analyses of WDV sequences from isolates of different host species, WDV has been shown to form a clade that is distinctly different from other mastreviruses and consists of multiple strains [99,100]. WDV sequence identity is below the delimitation criterion of <75% for mastrevirus species [36,101].
A further mastrevirus species was later identified in Avena fatua in Germany, based on sequences from isolates collected from plant samples from cereal fields. Oat dwarf virus (ODV) is closely related to WDV species but is distinct from wheat and barley strains and appears to be one of the causal agents of WDD in oats [101], with symptoms comparable to those of WDD (Figure 1a). Although some relationships do exist between WDV and ODV based upon a sequence analysis, the whole genome of ODV has only a nucleotide sequence similarity of approx. 70% compared to the wheat and barley strains of WDV. Based on a phylogenetic analysis, a revision of the classification of mastrevirus species into five phylogenetic groups (A-E) was proposed in 2013. In this context, WDV strains that preferentially infect wheat (WDV-W) or barley (WDV-B) should be assigned to groups A and C respectively [102]. A phylogenetic analysis of 230 isolates identified six strains (A-F) based on sequence similarity. Strains A- and F- were assigned to WDV-B (Figure 1, Clade A1, A1, WDV-Bar), and B-E strains were assigned mainly to WDV-W (Figure 1, Clade WDV-A, WDV-B) [103].
Macdowell et al. [14] and Matzeit [25] sequenced a 2,749 bp Swedish isolate (WDV-S), which was isolated from wheat in 1969 [75]. Two other wheat-adapted isolates from the Czech Republic (WDV-C, 43] and from France (WDV-F, 44] showed a genome size of 2,750 bp. Sequence analyses showed that barley WDV isolates have at least 94% similarity, whereas wheat isolates have at least 98.3 to 98.8% sequence similarity with the respective strains [43,44,75]. LIR and SIR represent the most variable parts of the WDV genome [101]. Within the genomes, nucleotide exchanges were observed in coding regions but these did not result in substitutions in the amino acid sequence, so this had no effect on the gene products [75].
Depending on the WDV isolate, differences in WDV virulence can be observed. Significantly increased symptoms of a WDV infection can be attributed to amino acid substitutions in the CP gene. This was reported within a Ukrainian study, in which the Ukrainian isolate Khm-K-Ukr caused a significantly greater reduction in seeds per ear and thousand grain weight compared to the isolate MIP-12-Ukr, which had fewer mutations in the CP gene than Khm-K-Ukr. The authors of the study suggested that the isolate MIP-K-Ukr has a higher potential for divergence, so that the CP sequence contains more non-synonymous changes that are subject to selection [104]. This has already been observed for Maize Streak virus, where even a few changes in nucleotide sequence have large effects on virus functionality [105].
Within a host, different WDV populations can occur [106], and a lack of antagonism between isolates may favor recombination between viral sequences during host infection. Such a case has already been described for the isolate WDV-Bar [TR]. The isolate is a variant of the barley WDV strain described in infected barley in Turkey [107]. Whole genomic sequence analysis showed that the WDV barley isolate partially corresponds to a novel WDV-like mastrevirus species [108]. In addition to the WDV bar [TR] isolate, sequence alignment analysis of field isolates revealed regions of the viral genome with short, few-nucleotide recombination patterns between wheat and barley strains. This suggests that sequences from barley strains were replaced by functionally homologous sequences from wheat strains [106]. Moreover, intra-specific recombinant genomes have been detected with two WDV wheat strains in China [109]. In this context, it should be noted that defective forms of wheat and barley strains containing at least part of the SIR and LIR sequences have also been detected in WDV-infected plants [15,106]. Putative recombinant isolates have also been identified for other members of the mastrevirus genus, such as maize streak virus [110].
3. Wheat dwarf disease (WDD)
3.1. History
The first dwarfing of wheat in Europe was observed in the early 20th century, with characteristic heavy tillering, dwarfing and deformation of the plants and subsequent death, while the first similar symptoms were described as early as 1863 in a region that is now part of Poland [111]. In Sweden, the leafhopper species Psammotettix alienus was made responsible for this by Tullgren in 1918 [143] (Table 1). At that time, it was assumed that other insects besides P. alienus were involved in the transmission of the so-called slidsjuka, or sheath disease, due to the partially stuck ears in the leaf sheaths. Overall, there were differing opinions about the cause, but it was consistently observed that the damage occurred particularly in dry and hot years [112]. Field prevalence in the 20th century was relatively low and thus there are only few descriptions of dwarfing symptoms in the scientific literature, but sometimes in context with severe outbreaks in wheat [113,114,115,116,117]. Slidsjuka, or WDD, declined around 1950 in Sweden and occurred only sporadically in the following 30-40 years until the 1980s/1990s [118,119,120]. This decline was attributed to changes in agricultural practices. The abandonment of undersowing in winter wheat, which was common in the first half of the century, or even the increased use of combine harvesters were considered positive effects for the control of the disease [20].
The direct relationship between virus, vector, and symptoms was first reported in 1961 using samples from wheat fields in western parts of the Czech Republic [10,122]. However, there was still confusion about the cause, as no clear virus particles or possible pathogens could be detected [13]. The identification and current taxonomic classification of the virus did not occur until 1980, when after three decades there was again an increased incidence of the disease in a number of European countries [20]. In the late 1980s, a new disease (pieds chétifs) occurred in central France, causing severe damage in wheat with yield losses of more than 50%, and associated with a high incidence of the leafhopper P. alienus [123]. Initially, only Mycoplasma-like organisms were diagnosed in this context [114]. In collaboration with a Swedish research group, the disease-causing pathogen was identified as WDV [124].
From this time on, the occurrence of vector and virus was studied, with WDV occurring mainly in central France and adjacent areas, but not in the coastal regions and in the south of the country [125,126]. The level of knowledge at that time was very low and was mainly based on studies from the Czech Republic [10], Sweden [20], and France [127]. In Germany, the first record probably occurred in 1990 near Dresden by Vacke [89] (Figure 2).
A concrete dispersal route cannot be deduced from the data. However, a natural spread over land seems most likely due to the biology of the animals and their activity. The virus was detected in the main Eurasian cereal-growing areas and in its region of origin in the Middle East. This can possibly be attributed to the fact that the climatic requirements for wheat cultivation, for example, match with those of P. alienus. Exceptions like India, as well as Canada and Australia underline these theories.
The reason for the increasing spread of WDV and the increased occurrence in areas where WDV was reported before is not clearly understood, but is probably caused by changes in agricultural practices. One of the main causes has been assumed to be the increased use of ploughless tillage. Also, the EU regulation on the use of a large part of stubble fields after winter wheat cultivation as set-aside areas was thought to be favorable for P. alienus reproduction and overwintering. Avoiding set-aside areas after the occurrence of WDV infected wheat and avoidance of undersown crops was therefore considered as a possible control measure in Sweden [119]. Furthermore, harvesting with short stubble, early tillage in autumn and avoiding early sowing had a positive effect on reducing the population of P. alienus [141]. Global climate change may also play a role in promoting the spread of vector-borne diseases. In this context, higher temperatures may favor the colonization of new habitats and hosts. Field monitoring is therefore essential, especially in cereal-growing regions, to identify additional regions where P. alienus together with WDV is potentially spreading [113,114,115,116,117], since the spread of WDV results from the migration of virulent vectors from wild or cultivated reservoirs into cereal fields [119,142]. Table 1 provides an overview of the history of WDD.

Table 1.
Overview of the historical development of WDV and its evidence in individual countries in relation to its reference in the literature. For some events, no direct data could be derived from the literature, so only a time span could be given.
Table 1.
Overview of the historical development of WDV and its evidence in individual countries in relation to its reference in the literature. For some events, no direct data could be derived from the literature, so only a time span could be given.
| Time | Event | Reference |
| Early 20th century | first observed dwarfing of wheat, called slidsjuka | 143, 112 |
| Early 20th century | relatively field prevalence of WDV, only few symptoms of dwarfing have been described in scientific literature | 113, 114, 115, 116, 117 |
| 1918 | leafhopper P. alienus was made responsible for WDV | 143 |
| Early 1950s | less undersowing in wheat, increased use of combine harvesters | 121 |
| Around 1950 | decline of slidsjuka due to changes in agricultural practices | 121, 118, 119, 120 |
| 1950-1980/1990 | Slidsjuka occured sporadically | 118, 119, 120 |
| 1961 | first report of direct relationship between virus, vector, symptoms, no virus particle detected | 10, 122 |
| 1980 | increased incidence of disease in european countries | 121 |
| 1980 | identification and taxonomic classification of WDV | 121 |
| Late 1980s | new disease (pieds chétifs) occured in France in association with P. alienus, disease was identified as WDV | 123, 124 |
3.2. Host range
The host range of WDV mainly includes monocotyledonous plants [102,144]. In addition to a variety of members of the Poaceae family, including important cereals such as wheat (Triticum aestivum L.), barley (Hordeum vulgare L.), rye (Secale cereale L.), oats (Avena sativa), and triticale [11,13,145], WDV also infects various wild and cultivated grasses, including Bromus secalinus L., Lolium multiflorum Lam. [13], Avena fatua L., B. inermis Leyss., B. tectorum L., H. murinum L., L. perenne L., L. temulentum L. [146], A. sterilis L., A. strigosa Schreb., Poa annua L. [99], L. remotum Schrk., Lagurus ovatus L. [147], and Apera spica-venti (L.) P. Beauv. [146], which are considered as virus reservoirs [13].
3.3. Symptoms of WDD
The name of the virus is derived from its main characteristics, the disruption of the shoot growth and the formation of numerous shoots in wheat, resulting in the typical dwarf and bushy growth (Figure 3).
Furthermore, symptoms of WDV infection in wheat also include chlorosis, reduced root size, intense yellow or red discoloration of leaves with or without a mosaic pattern, deformation of leaves, reduced growth hardiness, delayed ear emergence, and reduced number of ears and sterile flowers, resulting in significant yield losses and even complete plant death during early developmental stages of winter wheat and winter barley in winter and spring [13,119,148,149,150,151]. These are due in part to the side effects of infection, such as the effects of expression of viral suppressors of RNA silencing. In addition, symptoms may also affect plant defenses, leading to plant overreaction in the form of necrosis [152], chlorotic spots, and demarcated streaks on the leaves. The symptoms themselves first appear on the youngest and later on older leaves in association with small cracks and deformations on the youngest leaf, which are characteristic of the infection. This is followed by yellowing of the leaves from the leaf tips and margins with possible partial red coloration [13]. Symptomatic plants usually appear in patches on the field [11,13,150].
In addition to the described symptoms in wheat, the intensity of symptom expression varies among the other infested species. Symptoms in winter barley are similar to those in winter wheat, with no red coloration. Spring barley responds with a lower degree of dwarfing and yellowing of the leaf tips. Similar symptoms occur in winter rye, often associated with anthocyanin formation in leaves and culms. Spring rye shows only minor developmental depression, few leaf spots, and no disruption of generative plants. Oats show minor developmental depression, yellowing, and light red coloration [13]. Triticale shows no increased tillering after WDV infection compared to control plants, but spike-bearing culms shortened by half [153]. Growth reductions of 20%, severe tillering, yellowing, and chlorotic spots were observed for A. spica [13,99]. The wild grass Poa annua shows no symptoms after infection while Lolium perenne and Lolium multiflorum showed tolerance to WDV in studies with longer plant viability upon infection [11].
Furthermore, the extent of damage and development of symptoms depends on the time of infection. Early infections of winter cereals at the 2-3 leaf stage during fall result in reduced winter hardiness as well as severe developmental disorders with pronounced symptoms and negative effects on yield, as a result of ear formation that do often partially stuck in the leaf sheaths. Grain quality is reduced, as they are dried out, shriveled, and are partially unable to germinate [13,88]. The root system is also affected by WDV infection. As a result of infection, there is a reduced formation of secondary roots. Roots appear shorter and thinner overall [88].
Infections in spring result in shortening of internodes and, in some cases, ears. In spring wheat, no severe developmental disorders but shortening of shoots could be observed when infestation occurred during the period from the beginning of shooting to ear swelling (BBCH 31-45). Usually, the first signs of disease in winter wheat appear 18-25 days after infection. In general, symptoms are considered to appear 4-6 weeks after infection in early-sown wheat, while in late-sown wheat the corresponding symptoms do not become visible until spring, provided the plants are able to overwinter. If infection occurs in spring or early summer, the incubation period lasts 3-4 weeks. In spring wheat, under greenhouse conditions, the first symptoms are expected after 10-15 days, while infections in the field have an incubation period of 3 weeks [13].
Symptoms caused by Barley yellow dwarf virus (BYDV) infection, which belongs to the Luteoviridae family and is transmitted by aphids, are visually similar to those caused by WDV. When infected in early fall, it causes WDV-like growth depression. The two viruses can only be distinguished from each other by double antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) or by polymerase chain reaction (PCR), so prior to the discovery of WDV, plants were probably often assigned to BYDV on the basis of dwarfism [153,154].
4. WDV and its vector
4.1. Taxonomy and virus transmission of P. alienus
WDV is transmitted by the leafhopper species P. alienus, which belongs to the class Insecta, order Hemiptera, suborder Cicadomorpha in the family Cicadellidae. The vector itself is a holarctic species that is common in grass- and croplands [155]. Occurrence may be particularly high on fallow areas with many self-seeding plants of the Poaceae familiy. These may serve as reservoirs [12].
Many species of the family Cicadellidae are vectors of phytopathogenic viruses, including geminiviruses, phytorhabdoviruses, reoviruses, and marafiviruses [156,157,158,159]. In addition to WDV, P. alienus can persistently transmit a rhabdovirus, Wheat Yellow Striate Virus (WYSV, genus Nucleorhabdovirus) [160,161]. Furthermore, P. alienus appears to harbor entomopathogenic viruses that naturally infect insects and can only self-replicate in insect cells. In this context, filovirus-like particles were detected by Lundsgaard [162] in electron microscopic studies, which were confirmed as Taastrup virus (TV) and tentatively assigned to the Mononegavirales [162,163]. Using a next generation sequencing approach, additional insect-specific viruses including P. alienus iflavirus1 (PaIV1, genus Iflavirus, family Iflaviridae) [164], Tàiyuán leafhopper virus (TYLeV, genus Mivirus, family Chuviridae) [165], and Hancheng leafhopper Mivirus (HCLeV, genus Mivirus, family Chuviridae) have been detected [166]. Transmission electron microscopy (TEM) studies of WYSV-containing sites in salivary glands revealed the presence of reoviruses [167]. Reoviruses include insect-borne fijiviruses, which are the most common viral agents of a variety of diseases in gramineae, including Fiji virus (FDV) [168], garlic dwarf virus (GDV) [169], maize dwarf virus (MRDV) [170], Mal de Rio Cuarto virus (MRCV) [171], oat sterility dwarf virus (OSDV) [172], Pangola stunt virus (PaSV) [173], rice black-streaked dwarf virus (RBSDV) [26], and southern rice black-streaked dwarf virus (SRBSDV) [174]. Furthermore, the brown leafhopper Nilaparvata lugens was found to harbor Nilaparvata lugens reovirus (NLRV), a fijivirus that exclusively infects insects [175]. Most published data suggest that P. alienus is the sole vector of WDV. Some authors have also described a transmission by P. provincialis [176,137]. However, due to the complex taxonomy of species belonging to the genus, the leafhoppers used within the studies are often poorly characterized. This could lead to contradictory results, especially regarding the role of species in WDV transmission [177,178,179].
4.2. Morphology of P. alienus
To easily differentiate adult P. alienus from other leafhoppers, several criteria related to the morphological characteristics of the insects’ head, abdomen, and wings can be used [180,181]. Characteristic of adult P. alienus is their brown coloration with transparent wings, which are longer than the abdomen at a length of 2.7-3.7 mm [182,183]. Accurate species classification requires the morphological description of the male genitalia due to the high variability of the morphological characteristics of the aedeagus. The identification of nymphs and females based on morphological characteristics is currently not possible. This approach often turns out to be unreliable [184,185].
The accuracy of identification of individuals could be improved by using several criteria in parallel, such as morphometric parameters in combination with other approaches, such as emission of species- and sex-specific vibrational signals [186,187,188,189]. Only a few publications have described the vibrational signals emitted by leafhopper during their sexual communication [184,188,190,191] and a combination of body and aedeagus characteristics combined with the analysis of vibration signals revealed geographic differences between species related to these characteristics. However, this may allow not only the identification of this species but also of its origin. Therefore, future studies should include individuals from different countries to improve morphometric data [191]. A more straightforward approach that requires less expert knowledge is the use of DNA-Barcoding based upon sequencing of the cytochrome oxidase I (COI) [192,193]. To date, phylogenetic analysis using DNA barcoding were performed only for a limited number of species and individuals from Canada, Japan, and Korea [194,195]. Individual specimens of P. confinis and P. helvolus have already been found syntopic to P. alienus using this method [196].
4.3. Life cycle of P. alienus
The life cycle of P. alienus has been well studied (Figure 4). High population densities can occur in September, making this the most critical period for WDV infections on young winter cereal plants. Extensive primary infections could be observed until December [12].
Embryonic development is influenced by environmental conditions like temperature and day length. Low temperatures in winter are necessary for the abolition of dormancy (termination) [198]. P. alienus shows seven embryonic stages with an overall developmental duration of 16 to 24 days [12]. Depending on the temperature, the first larvae hatch in early May. In this context, protandry can be observed, where males hatch earlier than females [198]. The wingless nymphs develop into male and female adult leafhoppers in five stages with a developmental duration of 26-39 days until early summer. Development duration varies, again depending on temperature but also on host plant species, and sex of the leafhoppers. In winter barley, 31 days at a temperature of 20°C can be assumed. [197,12]. After hatching, the nymphs move through stocks exclusively by jumping, with older individuals being more mobile than the first two nymphal stages [199]. Newly hatched nymphs acquire the virus from host plants that were previously infected in the fall, which can lead to a secondary infection of plants. It has been observed that the first imagines appear at the end of May when temperature sum of all days above 9°C, measured from the 1st of January of a year, in general reaches 154°C [200,12]. Fertilization and oviposition occur after the tenth day of the adult stage, so that the first generation begins oviposition in June/July and the second-generation hatches about 18-20 days [197,12] later and lays its first eggs in early/mid-august. The duration of the entire life cycle from egg to egg is 58 days [197,12] where higher temperatures may reduce this time span as demonstrated for D. maidis [201]. Dormancy egg laying is induced with the onset of short day in mid/late August with a rate of 2-20%. From September onwards, up to 100% of eggs are laid as dormancy eggs [198]. Asexual reproduction, as observed in aphids, does not occur in leafhoppers [197,12].
In the temperate climate zone, two to four generations per year have been observed so far, depending on environmental conditions [98,183,179] with four complete generations from spring to fall in cereal-growing regions of France, whereas only two P. alienus generations per year occur in northern Europe and northwestern China [202]. Population dynamics studies showed that the density of individuals can reach 43 adults/m² [12]. The sex ratio in an adult population of P. alienus is close to 1 [202]. Adult numbers decrease above a temperature of 10°C [155]. Freezing temperatures of -5°C leads to induced death of the animals [200,12]. Temperatures above 35°C were associated with increased mortality rates [203]. In this context, the activity and population size of P. alienus increase significantly above a temperature of 15°C. A very mild autumn therefore leads to very active leafhoppers associated by increased WDV infection rates in the following summer [204].
4.4. Process of Virus Transmission
Indicated by taxonomic affiliation [205] and based upon electron microscopic observations [206], P. alienus belongs to the salivary sheath feeders (Auchenorrhyncha), which also includes most of the Sternorrhyncha (aphids, scale insects, psyllids). A salivary sheath is formed in the apoplast by secretions of gel saliva and surrounds the stylet as it moves through plant tissues towards the sieve elements, as shown for aphids [207]. When the stylet reaches the xylem or phloem, uptake of sap from the vascular cells occurs for the extraction of nutrients [205]. Direct damage by P. alienus caused by sucking activity is considered less important than indirect damage caused by transmission of phloem-restricted WDV [13]. WDV is persistently, circulatively, and non-propagatively transmitted from plant to plant [98,176]. Mechanical, soil- or seed-dependent transmission was not reported, so far [20].
A characteristic for persistent transmission is that a single virus uptake by the vector is sufficient to transmit the virus after a short latency period, i.e., the time between the uptake of virus particles and the subsequent release via the salivary glands, over months [20]. A latency period of one to several days is assumed [208,98,209]. Seventeen days after virus acquisition, transmission efficiency was found to be 90%. Transmission efficiency is influenced by environmental conditions, such as temperature, while transmission success depends on the virulence of the virus and the susceptibility of the host [210]. Vector studies on P. alienus are currently focusing on evaluating the transmission of WDV, determining the host plant range, and observing probing behavior on a variety of plants [211].
To date, two pathways of virus movement within the vector and transmission to healthy plants are known. Similar to persistent virus transmission of other insects, the virus can enter the salivary glands through the anterior midgut and hemocoel [212] or migrate into the lumen of the filtering chamber and on to the midgut lumen after entering the esophagus. Ten minutes after the first feeding, the virus is found throughout the insect’s midgut, and within the next ten minutes it accumulates throughout the entire filter chamber, midgut, hemocoel and salivary gland. Four hours after the first feeding, it is no longer be detectable in the filter chamber, while it has accumulated in the midgut, hemocoel and salivary glands, where it remains for the rest of the leafhopper’s life without replicating [209]. Transient direct transfer of particles to the salivary glands occurs within a few minutes, after which the normal circular, non-propagative pathway occurs with recruitment of the anterior and midgut organs of the leafhopper [213]. Here, the WDV CP not only has an encapsulation function but is also involved in retention and transmission of WDV in the leafhopper, virus propagation within the plant, and interaction with the Rep protein [103]. Once the vector has acquired the virus via ingestion [10,11,150], the virulent leafhopper can transmit the particles to new hosts each time it sucks. In this process, the virus particles are not lost during molting, so the virus remains in the vector for life. There, it interacts directly with the insect’s organs but does not replicate within the vector [150]. Although the WDV pathosystem is poorly documented in the literature, it has been clearly demonstrated that the virus is not transmitted vertically within virulent females to eggs. Vacke [10] assumed that after acquisition all developmental stages are capable of transmitting WDV. This was confirmed by Mehner et al. [11] using transmission tests with larval stages. Larval stages IV and V were more inefficient (22% and 9%, respectively) in terms of virus uptake compared to earlier larval stages and imagines (LI 43%, LII 50%, L3 45%, imago 41%) [11]. Larval stages appear to be more important than adult grasshoppers for WDV dispersal in this regard. Even at low densities, adults and larvae can cause significant yield losses by transmitting the virus to numerous host plants [200]. In the presence of the aphid species Rhopalosiphum padi, a negative effect on larval development, lifespan, and fertility of P. alienus has been observed. Studies of their interaction have ruled out food deprivation as a possible cause. It is hypothesized that the presence of aphids alters leafhopper behavior. This leads to an increase in the number of plants visited by individuals. Thus, this antagonistic interaction between aphids and leafhoppers, commonly found together in cereal fields, indirectly promotes efficient spread of WDV [200]. Within an experimental approach, the highest infection rates were observed at temperatures of 25°C. At higher temperatures, leafhoppers tended to settle on the ground, resulting in lower feeding rates and thus decrease in transmission rates [203].
4.5. Host Range and Wild Reservoirs
In particular, the presence of wild grasses in stubble fields as virus reservoirs can lead to an extension of the virus infection period in autumn and promote occurrence of the disease in spring [12]. The role of wild grasses as WDV virus reservoirs in cropland was demonstrated by Yazdkhasti et al. [214]. The results showed the potential role of ryegrass in the epidemiology of WDV [119] as a symptomless reservoir and underlined the wide host range of WDV [214]. In addition, removal of the overgrowth by ploughing immediately after harvest is strongly associated with leafhopper population reduction [12], probably reducing the spread of WDV from wheat and barley to wild grasses. The host range of the P. alienus as a first-degree oligophagous species [215] is restricted to known host plants of the Poaceae [216,182]. Therefore, in experimental studies, P. alienus has always been reared on grasses such as Hordeum vulgare L. [197,11], Triticum spp. [200], and Festuca gigantea (L.) Vill. [162]. Data from field studies also indicate feeding on other plant species including alfalfa, carrot [217,218] and ragwort [Ambrosia artemisiifolia L. (Asteraceae)] [219]. This indicates a possible diet of dicotyledonous plants and explains the detection of phytoplasma strains in the body of P. alienus [220,218,221]. These observations contradict the results of a previous study in which P. alienus was not able to survive longer than 2 days on the two non-grass plants A. artemisiifolia and the sedge Carex tomentosa L. (Cyperaceae). However, in this study the average survival time on the two species was longer than on the starvation control. This is due to the ability of the leafhoppers to possibly take up xylem cell sap from non-host plants [222], where the nutrient and water uptake may contribute to increased survival [211].
4.6. Studies of Insect-Plant Interactions
The behavioral sequence for host plant acceptance of hemipteran insects starts subsequent to landing with the exploration of the plant surface, where the plant surface is scanned with the tip of the labium, followed by probing including cell sap sampling [205,223]. For Cicadellidae, as observed for other hemipteran groups (e.g., aphids), probing seems to be critical in this process to distinguish between host and non-host plants [224]. As a result, not every plant is accepted as a suitable host, where rejection may occur during various stages of probing on the way to the phloem [225,226]. To better understand insect behavior and the mechanism of pathogen acquisition and transmission, electrical penetration graph (EPG) systems have been developed that provide real-time observation of feeding behavior of sap-sucking insects [227,228,229,230]. EPG is probably the most important and widely used technique to study insect-host-plant interactions, pathogen transmission and acquisition, insecticide mode of action and plant resistance [231,232,233,234,235,236]. Within an EPG measurement, insect and plant get integrated in an electrical circuit. The insect closes the electrical circuit by penetrating the plant with its stylet, acting like a switch. Insect and plant act as a variable resistor and different behavior patterns as well as the stylet-surrounding milieu affect the electrical resistance leading to voltage fluctuations resulting in different EPG waveforms, representing different feeding behavior patterns [230,237,238,239]. The EPG method has been used, for instance, in studies on aphids [228,230,240], leafhoppers [241,242,243,244], mealybugs [245], phylloxerids [246,247], thrips [248,249] and whiteflies [250]. Data on EPG studies of P. alienus are relatively limited in this regard [251,252,253,223]. Tholt et al. [1995] suggested that viruses like WDV are transmitted between insect and plant during the EPG phase Ps4, where the stylet of P. alienus is located in the phloem’s companion cells and sieve cells. In this context, phase Ps4 can be divided in phases 4a that is similar to waveforms E1 shown by aphids and is associated with the secretion of watery saliva into sieve elements, accompanied by virus transmission. Phase 4b appears to be homologue to waveform E2 shown by aphids, which is indicating the ingestion of sieve element sap [254,255,211], probably accompanied by virus inoculation [211]. In addition, Ps4a resembled the X-wave that occurs in other leafhoppers [256,257]. Thus, phase Ps4 is particularly important for WDV transmission [211] and could be used during WDV resistance research.
5. Possibilities of WDV Control
Knowledge on how to influence the population of P. alienus by appropriate countermeasures is currently insufficient. In field trials, parasitization has been detected only very rarely [198]. In Italy, parasitization of larvae and imagines by Gonatopus clavipes Thunberg, G. lunatus Klug (Heminoptera: Dryinidae (cicada wasps), representatives Pipunculidae (Diptera: eye flies), native to this country, have been observed more frequently [197]. Predominantly in the first generation in May to June, larvae of Gontopus sepoides Westwood were also found sitting on the abdomens of leafhoppers as exoparasites, while Alloneura nigritula Zetterstedt (Pipunculidae) occurs mainly in October to November on P. alienus [258]. In addition, experiments have shown that P. alienus is preyed by the spider Tibellus oblongus [259].
The actual lack of systematically evaluated, commercially available WDV resistant and tolerant elite cultivars of wheat and barley means that protection of these cereals against WDV infection relies mainly on agronomic measures and the use of chemically synthesized control agents (insecticides) against P. alienus.
Prevailing cropping practices influence the presence and spread of plant virus diseases and are closely related to the fluctuating incidence of WDD and the extent of yield losses. The timing of sowing in coordination with the migration of vectors between fields is a critical element of an integrated pest management (IPM) strategy [260]. The presence of infected reservoirs, e.g., wild grasses, leads to an increase in the incidence of many viruses, including MSV and WDV [145,261], which in turn involves the field hygiene aspect to reduce WDV infection. Another risk is irregular germination of seedlings [179], as P. alienus is attracted to patchy stands [17]. In addition, feeding behavior, population density and activity, the latter influenced by weather conditions, affect the intensity and frequency of a WDV infestation [179,262]. A WDV infection is possible at different stages of development (Figure 2), with economic damage decreasing with later infection [17], as has been described for other viruses such as BYDV [263]. Furthermore, it has been shown in wheat that plant resistance can develop after the stage of pseudo stem break (Z30) at the time of first node (Z31) [204].
Although IPM has the aim to reduce the application of chemically synthesized insecticides and other pesticides it does not exclude the possibility for insecticide application. With regard to virus spread, the insecticide induced reduction of vector insects has been shown to reduce the spread of insect-borne viruses [261,179,264]. However, the application of insecticides is associated with negative environmental side effects [265,266] including harmful effects on beneficial insects [267,268,269]. Together, the consideration of these aspects, as well as the broad public request and political will to reduce the use of insecticides, means that the focus for controlling WDV is mainly on agronomic measures and the breeding of resistant/tolerant varieties.
6. Resistance Research
Abiotic and biotic factors exert a constant influence on plant populations. Naturally, plants have inherent defense mechanisms that make them resistant to virus invasion. [270]. One way is to combat the virus by induced mechanisms, such as RNA silencing with small interfering RNA (siRNA) in response to the virus’s double-stranded RNA (dsRNA), hypersensitive response (HR), or nucleic acid methylation, before infection occurs. [271]. To date, nothing is reported about direct protection against WDV [272].
In recent decades, various studies have attempted to identify WDV-resistant germplasm among the available genetic wheat and barley accessions. Disease resistance genes in wild relatives of wheat can be valuable sources for resistance breeding [273]. Differential resistance to Soil-borne wheat mosaic virus (SBWMV) has been demonstrated in Ae. tauschii and T. monococcum [274,275,276] and in Ae. geniculata to BYDV [277]. Furthermore, Ae. caudata, Ae. ovata and Ae. triuncialis have been shown to respond to WDV infection with milder forms of symptoms compared to spring wheat [13].
Transmission of the virus to the genotypes to be tested was carried out in previous studies with the natural vector P. alienus or agroinfections. Phenotyping of infected plants is possible under field [149,151,3], near-field conditions [278,279] or in the greenhouse [280,278,279]. For inoculation in the field with virus-bearing leafhoppers, both natural and artificial inoculation can be used. In order to protect the crop from natural insect infestation and bird-induced damage, it is possible to conduct the trials under semi-field conditions in a gauze house [278,279].
Within phenotyping for resistance, various agronomic parameters may be of interest. Virus infections with WDV affect the performance and yield of infected plants compared to healthy plants. Here, the traits plant height, number of ears per plant, grains per ear, grain yield per plant, thousand kernel weight (TKW) per plant can serve as suitable indirect parameters for characterizing resistance [281]. Between tillering and sprouting (BBCH 23-30) as well as after harvest (BBCH 92), a comparative symptom assessment from 1-9 can be performed according to Scheurer et al. [282].
Various serological and molecular techniques are available for the detection of WDV infection as well as for a precise assignment of isolates to the corresponding strain designations. For the verification of WDV infections in the field, direct virus detection, especially via ELISA [283] and PCR [284,285,98,286], has proven to be a reliable method [287,154]. A differentiation of the WDV strains in the host plants and vector samples can be made on the basis of the characteristics of viral compounds (capsid proteins, nucleic acids). Due to the high sequence similarity between the CP of the isolates, serological differentiation of these using polyclonal antisera is not possible [149], but the use of monoclonal antibodies has been reported [288]. Several established molecular methods are available for the identification of WDV strain-specific sequences, such as standard PCR [100,77], restriction fragment length polymorphism (RFLP) [289], rolling circle amplification restriction fragment length polymorphism [101], and isothermal recombinase polymerase amplification methods [290]. In addition, molecular-based quantification assays in form of real-time PCR assays targeting a conserved region of the CP gene sequence and using a Taq-Man probe have been added to the list of detection methods [176].
6.1. Status quo of the resistance in wheat
So far, no highly resistant WDV bread wheat variety is known. However, tendencies to favor different wheat varieties [291] and differences in susceptibility have been found. Based on yield reduction, studies were conducted on winter wheat to define susceptible groups [151]. These showed only minor quantitative differences between the tested host plants and reference genotypes [149,3]. Most genotypes were susceptible to WDV infection and only a few genotypes could be classified as moderately resistant. Within screenings, the Czech winter wheat cultivars ‘Banquet’ and Svitava’ showed reduced virus levels, with moderate susceptibility at a yield reduction of 87.3-93.1% [151]. Moderate yield reductions of 82.5-92.6% after WDV inoculation were shown by the Russian cultivars ‘Belocerkovskaya’, ‘Kharkovskaya’, ‘Mironovskaya 808’, ‘Yubileynaya’ and ‘Kawvale’ and the Slovak and Czech cultivars ‘Astella’, ‘Boka’, ‘Bruneta’, ‘Bruta’, ‘Ilona’, ‘Ina’, ‘Mona’, ‘Regina’, ‘Saskia’ and ‘Senta’ [149]. The winter wheat varieties ‘Mv Dalma’ and ‘Mv Vekni’ from Martonvásár (Hungary) were described by Benkovics et al. [292] as the first partially resistant varieties. In leafhopper Transmission tests, both cultivars were infected (50%), but showed milder symptoms and a 100-10,000 times lower virus titer than the susceptible reference host cultivars ‘Mv Emese’ and ‘Mv Regiment’ (100% infection) four weeks after infection. A difference in the survival rates of the leafhoppers could not be determined. It can therefore be assumed that the resistance mechanism of the cultivars is rather based on the movement or replication of the virus and not at insect feeding [292]. ‘Mv Dalma’ carries a homozygous 1AL.1RS, while ‘Mv Vekni’ carries a homozygous 1BL.1RS rye translocation and contains several stem, leaf and yellow rust resistance genes derived from Aegilops ventricose (VPM-1, SR38, Lr37, YR17) [293,292,294].
To clarify the genetic basis of partial resistance in ’MV Vekni’, in a recent work, F2 populations based on a cross between the susceptible cultivar Regiment were inoculated in greenhouse experiments and quantitative trait loci (QTL) analysis was performed. Significant QTL were found for the peak markers RFL_Contig6053_2072 and Kukri_rep_c95718_868 on chromosome 6A for virus extinction (LOD=22.6), explaining a phenotypic variance of 38.4%. The significant deviation from the expected segregation ratio of 3r:1s observed in this work indicated that the resistance is primarily inherited monogenetically due to the action of one major gene, eventually accompanied by additional minor QTL that could not be detected within the analysis. The hypothesis of coupling rye introgression with WDV resistance in Vekni could not be confirmed in this work. Within the main QTL interval, among others, a gene encoding protein kinase activity could be identified [295]. These are involved in various defense mechanisms against geminiviruses and lead to attenuation and reduction of infection [296]. Furthermore, genes associated with DNA-directed transcriptional regulation in Triticum aestivum have been found to act as viral defense modulators and influence the host-dependent DNA replication cycle [48,295].
In a recent study [297], the changes in transcriptome profiles of the resistant wheat genotypes ‘Svitava’ and ‘Fengyou 3’ compared to the susceptible cultivar ‘Akteur’ were investigated after WDV infection. The study provides insights into the specific transcriptome profiles and pathways associated with resistance and susceptibility to WDV in wheat genotypes. RNA-Seq analysis revealed significantly different expression of transcripts in response to WDV infection in ‘Akteur’, ‘Fengyou 3’, and ‘Svitava’ genotypes. Gene ontology (GO) analysis showed that different biological processes, cellular components, and molecular functions were activated in the tested genotypes. The resistant genotype showed significant activation of biological processes compared to the susceptible genotype. Certain classes of genes were affected by WDV infection. For example, transport activity was suppressed [297], which could prevent virus movement and accumulation [298]. On the other hand, oxidoreductase and lyase activities were activated [297], which are involved in defense responses and limit virus accumulation [299]. The ‘Svitava’ genotype suppressed reductase protein classes and chaperones. The latter include heat shock proteins (HSP), which play a role in viral DNA/protein aggregation and viral reduction [300,301,302]. Suppression of reductase activity is associated with a reduction in reactive oxygen species (ROS) accumulation, which is associated with better adaptation to viral infections [303]. Analyses of GO and KEGG metabolic pathways revealed reprogramming of several transcripts in response to WDV infection, particularly in the carbohydrate, energy, lipid, nucleotide, amino acid, glycan, and vitamin metabolism. Secondary metabolic and photosynthetic pathways were induced in ‘Svitava’. The susceptible genotype showed down-regulation of photosynthesis-related carbon fixation genes, which, in contrast, were induced in the resistant genotypes. Transcripts for biosynthesis of other secondary metabolites were upregulated in ‘Svitava’ and downregulated in ‘Fengyou 3’ and ‘Akteur’, possibly contributing to higher resistance through their antiviral properties [304,297]. Transcription factors (TFs), including AP2/ERF, bHLH, MYB, and WRKY families, were highly enriched under WDV infection [297]. These TFs are known to regulate plant responses to various biotic and abiotic stresses [305,306]. In particular, ERFs have been linked to plant immune responses and resistance to plant viruses [307].
In greenhouse experiments with 13 wild and 5 domesticated wheat taxa of different ploidy, accessions of the species Aeg. tauschii, Aeg. cylindrical, Aeg. Searsii and T. spelta showed WDV tolerance. The accessions were initially strongly affected by symptoms 28 days after infection (dpi). Thereafter, there was a decline in symptoms with a relative increase in leaves and shoots at 112 dpi. Within the study, the domesticated wheat cultivars did not always show more severe symptoms, but there was a differential impact of infection on growth traits and leaf chlorosis in wild and domesticated wheat cultivars [280]. This could be attributed to a slight RNA silencing suppressor activity of the WDV proteins Rep and RepA [308,59]. Both viral proteins, when expressed in infiltrated transgenic leaves of Nicotiana benthamiana with green fluorescent protein (GFP) reporter gene, resulted in inhibition of post-transcriptional gene silencing (PTGS) and RNA silencing of the GFP reporter gene [308].
Within another study, 500 wheat accessions were phenotyped for WDV resistance using artificial inoculation in gauze houses. The majority of accessions showed a strong impact of WDV infection with a wide range of reduction in plant height (3.6-100%), number of ears (0-100%) and yield (2.3-100%) [278]. In contrast to Nygren et al. [1995], domesticated wheat varieties within the panel did not show a generally higher infection rate than wild wheat varieties and relatives [278]. The authors concluded that the genetic bottleneck that arose during evolution and domestication did not necessarily lead to higher WDV susceptibility, but that these variations created by ancestral hybridization were compensated for. During the study, the partially resistant genotypes ’MV Dalma’ and ’MV Vekni’ were confirmed with an average infection rate of 34.5% and 21.5%, respectively, and weaker symptom expression compared to susceptible varieties. In addition, 19 other sources of WDV resistance with lower infection rates than ’MV Vekni’ were identified, including di-, tetra- and hexaploid genebank wheat accessions. Ten T. aestivum, two T. vavilovii, two T. sp. (genebank accessions with unknown subspecies), one T. boeoticum, one T. macha, one Ae. geniculata, one Ae. bicornis and one Ae. longissima accession had lower infection rates than ’MV Vekni’. The cultivar ’Fisht’ proved to be another resistant cultivar with a low average number of infected plants (5.7%) and less severe virus symptoms (average score 2.3) compared to the reference cultivars ‘Mv Dalma’ (34.5%, 5.9) and ‘Mv Vekni’ (21.5%, 4.6) and the susceptible ‘Mv Regiment’ (64.9%, 6.7) as well as ‘Mv Emese’ (68.1%, 6.9). Overall, the results indicated that there are natural sources of WDV resistance within the wheat gene pool. A subpanel was also used to identify QTL for WDV resistance in hexaploid wheat. The putative 35 QTL (FDR, α < 0.05) for partial WDV resistance for the traits relative plant height (relPH), relative yield (relYield), and relative thousand kernel weight (relTKW) are located on chromosomes 1B, 1D, 2B, 3A, 3B, 4A, 4B, 5A, 6A, 7A, and 7B. Among them, the most significant QTL were detected on chromosome 1B, especially six QTL explaining more than 10% of the phenotypic variance (LOD 5.0-8.7), and two highly significant yield-related QTL explaining 18.3% of the phylogenetic variance (LOD 5.0-8.7), which can be used to develop molecular markers in resistance breeding. The QTL identified here could be associated with genes encoding DNA template regulation of transcription, splicing mRNA by spliceosome, gene silencing by RNA, and protein kinase activity [278]. Genes responsible for regulation of DNA template transcription may serve as modulators of viral defense, particularly with respect to controlling the host-dependent DNA replication cycle of WDV [48]. Earlier research on RNA-mediated gene silencing has also demonstrated the ability of geminiviruses to trigger post-transcriptional gene silencing (PTGS) [309,310], such that viral dsRNA is degraded during the RNA splicing mechanism to small interfering RNAs (siRNAs) that align and degrade silencing complexes to sequence-specific mRNA [311]. Also involved in plant resistance to geminiviruses are protein kinase domains through phosphorylation of viral pathogenesis proteins. The viral protein ßC1 is phosphorylated by SNF1-related kinases, which has negative effects on RNA silencing suppressor function or labeling for degradation in the 26s proteosome. As a result, delayed/reduced viral infection may be observed [312]. Overall, the results suggest that other resistance genes are involved in defense against WDV.
Previous studies have shown that resistance to various viruses is localized to the D chromosome. For example, resistance to Soil-borne Wheat Mosaic Virus (SBWMV) is localized on chromosomes 4D and 5D, and the resistance gene encoding alleles on chromosome 5D is due to Aegilops tauschii [313,314]. Additional highly significant marker trait associations (MTA) were found on chromosome 2D for resistance to Wheat spindle streak mosaic virus (WSSMV) [315]. Of 35 QTL identified, 25 QTL, explaining between 7.4 and 18.3% of the phenotypic variance, were verified within in four biparental populations with the cultivar ’Fisht’ as parent [278]. Within the segregation analysis, two of the markers showed significant effects on relYield, 11 on relTKW and 10 on relative virus titers. The QTL on chromosome 1B consistently showed highly significant effects in all four populations [316].
A recent QTL study revealed two additional highly significant QTL associated with WDV resistance [317]. The primary QTL, Qwdv.ifa-6A, mapped to the long arm of chromosome 6A between markers Tdurum_contig75700_411 (at 601,412,152 bp) and AX-95197581 (at 605,868,853 bp). Qwdv.ifa-6A originated from the Dutch experimental line SVP-72017 and showed a strong effect in all populations, explaining a significant proportion (up to 73.9%) of the phenotypic variance. The second QTL, Qwdv.ifa-1B, was located on chromosome 1B and derived from the susceptible parental line P1314. The QTL is possibly linked to the 1RS.1BL translocation, which originated from the CIMMYT line CM-82036. Qwdv.ifa-1B was responsible for a substantial portion (up to 15.8%) of the phenotypic variance in WDV resistance [317]. Efficacy of rye chromatin segment 1RS.1BL against Wheat Streak Mosaic Virus (WSMV) has been reported previously [318], but there is no evidence to date that the same gene confers resistance to both WDV and WSMV. The QTL mapped on the short arm of chromosome 1B in the study by Pfrieme et al. [278], overlap with the Qwdv.ifa-1B QTL identified within the study by Buerstmayr & Buerstmayr [317]. Although Fisht has the preferable allele on chromosome 1B, the presence of the translocation 1RS.1BL remains unclear. Thus, it remains uncertain whether ‘Fisht’ and P1314 (the resistance donor for Qwdv.ifa-1B) have the same resistance gene. This study has shown that Qwdv.ifa-6A and Qwdv.ifa-1B are clearly additive, suggesting that pyramidization of resistance QTL could increase both the durability and extent of resistance [317].
The utility of the discovered QTL for wheat breeding depends on their ability to predict quantitative WDV resistance in a range of genetic backgrounds. For breeding, QTL associated with resistance should explain at least 10% of the phenotypic variance. Their pyramiding is an interesting approach to increase resistance to WDV [319,320,321,278], as already shown for BYDV in barley [322]. The use of the identified QTL in marker-assisted selection can be accomplished via the development of PCR-based markers from verified array-based markers. For example, the use of competitive allele-specific PCR marker (KASP) developed from flanking marker sequences offers an efficient approach in hexaploid wheat [323,324]. Introduction of WDV tolerance can be facilitated by the use of molecular markers, avoiding artificial inoculation with virus-bearing leafhoppers, which is difficult to integrate into applied breeding programs.
7. Perspective
WDV is a worldwide virus disease that affects most cereals and grasses. As a result of climate change, the importance of insect-transmitted viruses will inevitably increase in the coming years. Research conducted within the last decades allows a description of the biology of the putative vector, the virus and the plant hosts. In this context, the epidemiology of WDV is characterized by the presence of different strains, recombinants and virus species, as well as a complex taxonomy of vectors and a contradictory host range. Even though, WDV as a DNA virus is thought to have a lower mutation rate compared to RNA viruses, putative new variants and recombinants have already been detected in reservoirs and crop species in recent years. Since there are no approved chemical control agents in the European Union, agronomic measures are currently the only way for controlling WDV. The detection of the first WDV resistant genotypes and QTL in wheat indicates that resistance is present in the cereal pool. As can be seen from this review, further experimental studies of resistance to WDV and the epidemiology of the vector are needed and promising, especially given the economic importance of this viral disease. Development of resistant cereal varieties offer the prospect of minimizing the spread and losses from WDV infections.
Author Contributions
Conceptualization, AKP; writing—original draft preparation, AKP; writing—review and editing, AS, TW, KP; visualization, AKP; All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Buck, K. W. Geminiviruses (Geminiviridae). Encyclopedia of Virology 1999, 597–606. [CrossRef]
- Canto, T.; Aranda; M. A.; Fereres, A. Climate change effects on physiology and population processes of hosts and vectors that influence the spread of hemipteran-borne plant viruses. Global Change Biology 2009, 15. Jg., Nr. 8, 1884-1894. [CrossRef]
- Habekuß, A.; Riedel, C.; Schliephake, E.; Ordon, F. Breeding for resistance to insect-transmitted viruses in barley- an emerging challenge due to global warming. Journal für Kulturpflanzen 2009, 61. Jg., Nr. 2, 53-61. [CrossRef]
- Roos, J.; Hopkins, R.; Kvarnheden, A.; Dixelius, C. The impact of global warming on plant diseases and insect vectors in Sweden. European Journal of Plant Pathology 2011, 129. Jg., 9-19. [CrossRef]
- Ziesche, T. M.; Bell, J.; Ordon, F.; Schliephake, E.; Will, T. Long-term monitoring of insects in agricultural landscapes. Mitteilungen der DGaaE 2020, 22. Jg., 101-106.
- Barnett, O. W. & Main, C. E. Plant Virus Disease - Economic Aspects. Encyclopedia of Virology 1999, 1318–1326. [CrossRef]
- Waterworth, H. E.; Hadidi, A. Economic losses due to plant viruses. Plant virus disease control. APS, St. Paul 1998.
- Fraser, R. S. S. Plant Resistance to Viruses | Natural Resistance. In Encyclopedia of Virology, 2nd ed.; Granoff, A., Webster, R.G.; Publisher: Elsevier, 1999, 1300–1307. [CrossRef]
- van Regenmortel, M. H.; Fauquet, C. M.; Bishop, D. H.; Carstens, E. B.; Estes, M. K.; Lemon, S. M.; ... ; Wickner, R. B. Virus taxonomy: classification and nomenclature of viruses. Seventh report of the International Committee on Taxonomy of Viruses. Academic Press 2000.
- Vacke, J. Wheat dwarf virus disease. Biologia Plantarum 1961, 3. Jg., Nr. 3, 228-233. [CrossRef]
- Mehner, S.; Manurung, B.; Gruntzig, M.; Habekuss, A.; Witsack, W.; Fuchs, E. Investigations into the ecology of the Wheat dwarf virus (WDV) in Saxony-Anhalt, Germany. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 2003, 313-323.
- Manurung, B.; Witsack, W.; Mehner, S.; Gruntzig, M.; Fuchs, E. Studies on biology and population dynamics of the leafhopper Psammotettix alienus Dahlb. (Homoptera: Auchenorrhyncha) as vector of Wheat dwarf virus (WDV) in Saxony-Anhalt, Germany. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 2005, 497-507.
- Vacke, J. Host plants range and symptoms of wheat dwarf virus. Věd Pr Výz Ústavú Rostl Výroby Praha-Ruzyně 1972, 17. Jg., 151-162.
- MacDowell, S. W.; Macdonald, H.; Hamilton, W. D. O.; Coutts, R. H. A.; Buck, K. W. The nucleotide sequence of cloned wheat dwarf virus DNA. The EMBO journal 1985, 4. Jg., Nr. 9, 2173-2180. [CrossRef]
- Macdonald, H.; Coutts, R. H. A.; Buck, K. W. Characterization of a Subgenomic DNA Isolated from Triticum Aestivum Plants Infected with Wheat Dwarf. Journal of general virology 1988, 69. Jg., Nr. 6, 1339-1344. [CrossRef]
- Schalk H.J.; Matzeit V.; Schiller B.; Schell J.; Gronenborn B. Wheat dwarf virus, a geminivirus of graminaceous plants needs splicing for replication. The EMBO journal 1989, 8. Jg., Nr. 2, 359-364. [CrossRef]
- Lindblad, M.; Waern, P. Correlation of wheat dwarf incidence to winter wheat cultivation practices. Agriculture, ecosystems & environment 2002, 92. Jg., Nr. 2-3, 115-122. [CrossRef]
- Lemmetty, A.; Huusela-Veistola, E. First Report of Wheat dwarf virus in Winter Wheat in Finland. Plant disease 2005, 89. Jg., Nr. 8, 912-912. [CrossRef]
- Wang, J.; Guan, Y.; Wu, L.; Guan, X.; Cai, W.; Huang, J.; Dong, W.; Zhang, B. Changing Lengths of the Four Seasons by Global Warming. Geophysical Research Letters 2021, 48. Jg., Nr. 6. [CrossRef]
- Lindsten, K.; Lindsten, B.; Abdelmoeti, M.; Junti, N. Purification and some properties of wheat dwarf virus. Proceedings of the 3rd conference on virus diseases of Gramineae in Europe, Rothamsted. 1980. 27-31.
- Fauquet, C. M.; Briddon, R. W.; Brown, J. K.; Moriones, E.; Stanley, J.; Zerbini, M.; Zhou, X. Geminivirus strain demarcation and nomenclature. Archives of virology 2008, 153. Jg., 783-821. [CrossRef]
- Bernardo, P.; Golden, M.; Akram, M.; Naimuddin, Nadarajan, N.; Fernandez, E.; Granier, M.; Rebelo, A. G.; Peterschmitt, M.; Martin, D. P.; Roumagnac, P. Identification and characterisation of a highly divergent geminivirus: Evolutionary and taxonomic implications. Virus Research 2013, 177. Jg., Nr. 1, 35-45. [CrossRef]
- Varsani, A.; Navas-Castillo, J.; Moriones, E.; Hernández-Zepeda, C.; Idris, A.; Brown, J. K.; Murilo Zerbini, F.; Martin, D. P. Establishment of three new genera in the family Geminiviridae: Becurtovirus, Eragrovirus and Turncurtovirus. Archives of virology 2014, 159. Jg., 2193-2203. [CrossRef]
- Agrios, G. N. (1988). in Plant Pathology, 3rdEdition Edn, ed. G. N. Agrios (New York, NY: Academic Press), 3–39. [CrossRef]
- Matzeit, V. Wheat dwarf virus – Ein Geminivirus monokotyledoner Pflanzen-DNA-Sequenz, Replikation und Einsatz seines Genoms zur Amplifikation und Expression fremder Gene. Ph.D. Dissertation, Universität zu Köln 1988.
- Zhang, W.; Olson, N. H.; Baker, T. S.; Faulkner, L.; Agbandje-McKenna, M.; Boulton, M. I.; Davies, J. W.; McKenna, R. Structure of the Maize Streak Virus Geminate Particle. Virology 2001, 279. Jg., Nr. 2, 471-477. [CrossRef]
- Boulton, M. I. Functions and interactions of mastrevirus gene products. Physiological and molecular plant pathology 2002, 60. Jg., Nr. 5, 243-255. [CrossRef]
- Drews, G.; Adam, G.; Heinze, C. Molekulare Pflanzenvirologie. Springer-Verlag: Berlin Heidelberg, Germany, 2004. [CrossRef]
- Adejare, G. O.; Coutts, R. H. A. The Isolation and Characterisation of a Virus from Nigerian Cassava Plants Affected by the Cassava Mosaic Disease, and Attempted Transmission of the Disease. Journal of Phytopathology 1982, 103. Jg., Nr. 3, 198-210. [CrossRef]
- Harrison, B. D. Advances in Geminivirus Research. Annual Review of Phytopathology 1985, 23. Jg., Nr. 1, 55-82. [CrossRef]
- Damsteegt, V. D.; Igwegbe, E. C. K. Epidemiology and Control of Maize streak disease. Plant Virus Disease Control 1998, 484-494.
- Moffat, A. S. Geminiviruses Emerge as Serious Crop Threat. Science 1999, 286. Jg., Nr. 5446, 1835-1835. [CrossRef]
- Lefkowitz E.J.; Dempsey D.M.; Hendrickson R.C.; Orton R.J.; Siddell S.G.; Smith D.B. Virus taxonomy: the database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic acids research 2018, 46. Jg., Nr. D1, D708-D717. [CrossRef]
- Fiallo-Olivé, E.; Lett, J.-M.; Martin, D. P.; Roumagnac, P.; Varsani, A.; Zerbini, F. M.; Navas-Castillo, J. ICTV Virus Taxonomy Profile: Geminiviridae 2021. Journal of General Virology 2021, 102. Jg., Nr. 12, 001696. [CrossRef]
- Family: Geminiviridae. Available online: https://ictv.global/report/chapter/geminiviridae/geminiviridae (accessed on 12.11.2022).
- Fauquet, C. M.; Bisaro, D. M.; Briddon, R. W.; Brown, J. K.; Harrison, B. D.; Rybicki, E. P.; Stenger, D. C.; Stanley, J. Virology division news: Revision of taxonomic criteria for species demarcation in the family Geminiviridae, and an updated list of begomovirus species. Archives of virology 2003, 148. Jg., Nr. 2, 405-420. [CrossRef]
- Morris, B. A. M.; Richardson, K. A.; Haley, A.; Zhan, X.; Thomas, J. E. The nucleotide sequence of the infectious cloned dna component of tobacco yellow dwarf virus reveals features of geminiviruses infecting monocotyledonous plants. Virology 1992, 187. Jg., Nr. 2, 633-642. [CrossRef]
- Gutierrez, C. Geminivirus DNA replication. Molecular Life Sciences CMLS 1999, 56. Jg., 313-329. [CrossRef]
- Thomas, J. E.; Parry, J. N.; Schwinghamer, M. W.; Dann, E. K. Two novel mastreviruses from chickpea (Cicer arietinum) in Australia. Archives of virology 2010, 155. Jg., 1777-1788. [CrossRef]
- Zerbini, F. M.; Briddon, R. W.; Idris, A.; Martin, D. P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A. ICTV Virus Taxonomy Profile: Geminiviridae. Journal of general virology 2017, 98. Jg., Nr. 2, 131-133. [CrossRef]
- Gafni, Y.; Epel, B. L. The role of host and viral proteins in intra- and inter-cellular trafficking of geminiviruses. Physiological and Molecular Plant Pathology 2002, 60. Jg., Nr. 5, 231-241. [CrossRef]
- Ramsell, J.N.E. Genetic Variability of Wheat dwarf virus. Ph.D. dissertation, Swedish University of Agricultural Sciences, 2007.
- Woolston, C. J.; Barker, R.; Gunn, H.; Boulton, M. I.; Mullineaux, P. M. Agroinfection and nucleotide sequence of cloned wheat dwarf virus DNA. Plant Molecular Biology 1988, 11. Jg., 35-43. [CrossRef]
- Bendahmane, M.; Schalk, H. J.; Gronenborn, B. Identification and characterization of wheat dwarf virus from France using a rapid method for geminivirus DNA preparation. Phytopathology 1995, 85. Jg., Nr. 11, 1449-1455.
- Dickinson, V. J.; Halder, J.; Woolston, C. J. The Product of Maize Streak Virus ORF V1 Is Associated with Secondary Plasmodesmata and Is First Detected with the Onset of Viral Lesions. Virology 1996, 220. Jg., Nr. 1, 51-59. [CrossRef]
- Gutierrez, C. Geminiviruses and the plant cell cycle. Plant molecular biology 2000, 43. Jg., 763-772. [CrossRef]
- Gutierrez, C. DNA replication and cell cycle in plants: learning from geminiviruses. The EMBO journal 2000, 19. Jg., Nr. 5, 792-799. [CrossRef]
- Gutierrez, C.; Ramirez-Parra, E.; Mar Castellano, M.; Sanz-Burgos, A. P.; Luque, A.; Missich, R. Geminivirus DNA replication and cell cycle interactions. Veterinary microbiology 2004, 98. Jg., Nr. 2, 111-119. [CrossRef]
- Rojas, M.R.; Hagen, C.; Lucas, W.J.; Gilbertson, R.L. Exploiting chinks in the plant’s armor: Evolution and emergence of geminiviruses. Annu. Rev. Phytopathol. 2005, 43. Jg., 361-394. [CrossRef]
- Briddon, R. W.; Martin, D. P.; Owor, B. E.; Donaldson, L.; Markham, P. G.; Greber, R. S.; Varsani, A. A novel species of mastrevirus (family Geminiviridae) isolated from Digitaria didactyla grass from Australia. Archives of virology 2010, 155. Jg., 1529-1534. [CrossRef]
- Hofer, J. M. I.; Dekker, E. L.; Reynolds, H. V.; Woolston, C. J.; Cox, B. S.; Mullineaux, P. M. Coordinate Regulation of Replication and Virion Sense Gene Expression in Wheat Dwarf Virus. The plant cell 1992, 4. Jg., Nr. 2, 213-223. [CrossRef]
- Morris-Krsinich, B. A. M.; Mullineaux, P. M.; Donson, J.; Boulton, M. I.; Markham, P. G.; Short, M. N.; Davies, J. W. Bidirectional transcription of maize streak virus DNA and identification of the coat protein gene. Nucleic acids research 1985, 13. Jg., Nr. 20, 7237-7256. [CrossRef]
- Dekker, E. L.; Woolston, C. J.; Xue, Y.; Cox, B.; Mullineaux, P. M. Transcript mapping reveals different expression strategies for the bicistronic RNAs of the geminivirus wheat dwarf virus. Nucleic acids research 1991, 19. Jg., Nr. 15, 4075-4081. [CrossRef]
- Fenoll, C.; Black, D. M.; Howell, S. H. The intergenic region of maize streak virus contains promoter elements involved in rightward transcription of the viral genome. The EMBO Journal 1988, 7. Jg., Nr. 6, 1589-1596. [CrossRef]
- Accotto, G. P.; Donson, J.; Mullineaux, P. M. Mapping of Digitaria streak virus transcripts reveals different RNA species from the same transcription unit. The EMBO Journal 1989, 8. Jg., Nr. 4, 1033-1039. [CrossRef]
- Mullineaux, P. M.; Guerineau, F.; Accotto, G.-P. Processing of complementary sense RNAs of Digitariastreak virus in its host and in transgenic tobacco. Nucleic acids research 1990, 18. Jg., Nr. 24, 7259-7265. [CrossRef]
- Wright, E. A.; Heckel, T.; Groenendijk, J.; Davies, J. W.; Boulton, M. I. Splicing features in maize streak virus virion- and complementary-sense gene expression. The plant journal 1997, 12. Jg., Nr. 6, 1285-1297. [CrossRef]
- Palmer, K. E.; Rybicki, E. P. The Molecular Biology of Mastreviruses. Advances in virus research 1998, 50. Jg., 183-234. [CrossRef]
- Wang, Y.; Mao, Q.; Liu, W.; Mar, T.; Wei, T.; Liu, Y.; Wang, X. Localization and Distribution of Wheat dwarf virus in Its Vector Leafhopper, Psammotettix alienus. Phytopathology 2014, 104. Jg., Nr. 8, 897-904. [CrossRef]
- Noueiry, A. O.; Lucas, W. J.; Gilbertson, R. L. Two proteins of a plant DNA virus coordinate nuclear and plasmodesmal transport. Cell 1994, 76. Jg., Nr. 5, 925-932. [CrossRef]
- Liu, H.; Boulton, M. I.; Oparka, K. J.; Davies, J. W. Interaction of the movement and coat proteins of Maize streak virus: implications for the transport of viral DNA Journal of general virology 2001, 82. Jg., Nr. 1, 35-44. [CrossRef]
- Liu, H.; Andrew, Lucy, P.; Davies, J. W.; Boulton, M. I. A single amino acid change in the coat protein of Maize streak virus abolishes systemic infection, but not interaction with viral DNA or movement protein. Molecular plant pathology 2001, 2. Jg., Nr. 4, 223-228. [CrossRef]
- Noris, E.; Vaira, A. M.; Caciagli, P.; Masenga, V.; Gronenborn, B. & Accotto, G. P. Amino Acids in the Capsid Protein of Tomato Yellow Leaf Curl Virus That Are Crucial for Systemic Infection, Particle Formation, and Insect Transmission. Journal of virology 1998, 72. Jg., Nr. 12, 10050-10057. [CrossRef]
- Liu, H.; Boulton, M. I.; Thomas, C. L.; Prior, D. A. M.; Oparka, K. J.; Davies, J. W. Maize Streak Virus Coat Protein Is Karyophyllic and Facilitates Nuclear Transport of Viral DNA. Molecular plant-microbe interactions 1999, 12. Jg., Nr. 10, 894-900. [CrossRef]
- Kotlizky, G.; Boulton, M. I.; Pitaksutheepong, C.; Davies, J. W.; Epel, B. L. Intracellular and Intercellular Movement of Maize Streak Geminivirus V1 and V2 Proteins Transiently Expressed as Green Fluorescent Protein Fusions. Virology 2000, 274. Jg., Nr. 1, 32-38. [CrossRef]
- Sunter, G.; Bisaro, D. M. Transactivation of Geminivirus AR1 and BR1 Gene Expression by the Viral AL2 Gene Product Occurs at the Level of Transcription. The Plant Cell 1992, 4. Jg., Nr. 10, 1321-1331. [CrossRef]
- Hong, Y.; Saunders, K.; Hartley, M. R.; Stanley, J. Resistance to Geminivirus Infection by Virus-Induced Expression of Dianthin in Transgenic Plants. Virology 1996, 220. Jg., Nr. 1, 119-127. [CrossRef]
- Voinnet, O.; Pinto, Y. M.; Baulcombe, D. C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proceedings of the National Academy of Sciences 1999, 96. Jg., Nr. 24, 14147-14152. [CrossRef]
- Shivaprasad, P. V.; Akbergenov, R.; Trinks, D.; Rajeswaran, R.; Veluthambi, K.; Hohn, T.; Pooggin, M. M. Promoters, Transcripts, and Regulatory Proteins of Mungbean Yellow Mosaic Geminivirus. Journal of virology 2005, 79. Jg., Nr. 13, 8149-8163. [CrossRef]
- Trinks, D.; Rajeswaran, R.; Shivaprasad, P. V.; Akbergenov, R.; Oakeley, E. J.; Veluthambi, K.; Hohn, T.; Pooggin, M. M. Suppression of RNA Silencing by a Geminivirus Nuclear Protein, AC2, Correlates with Transactivation of Host Genes. Journal of virology 2005, 79. Jg., Nr. 4, 2517-2527. [CrossRef]
- Wang, H.; Buckley, K. J.; Yang, X.; Buchmann, R. C.; Bisaro, D. M. Adenosine Kinase Inhibition and Suppression of RNA Silencing by Geminivirus AL2 and L2 Proteins. Journal of virology 2005, 79. Jg., Nr. 12, 7410-7418. [CrossRef]
- Chowda-Reddy, R. V.; Dong, W.; Felton, C.; Ryman, D.; Ballard, K.; Fondong, V. N. Characterization of the cassava geminivirus transcription activation protein putative nuclear localization signal. Virus research 2009, 145. Jg., Nr. 2, 270-278. [CrossRef]
- Castillo-González, C.; Liu, X.; Huang, C.; Zhao, C.; Ma, Z.; Hu, T.; Sun, F.; Zhou, X.; Wang, X.J.; Zhang, X. Geminivirus-Encoded TrAP Suppressor Inhibits the Histone Methyltransferase SUVH4/KYP to Counter Host Defense. Elife 2015, 4. Jg., S. e06671. [CrossRef]
- Kumar, V.; Mishra, S. K.; Rahman, J.; Taneja, J.; Sundaresan, G.; Mishra, N. S.; Mukherjee, S. K. Mungbean yellow mosaic Indian virus encoded AC2 protein suppresses RNA silencing by inhibiting Arabidopsis RDR6 and AGO1 activities. Virology 2015, 486. Jg., 158-172. [CrossRef]
- Kvarnheden, A.; Lindblad, M.; Lindsten, K.; Valkonen, J. P. T. Genetic diversity of Wheat dwarf virus. Archives of virology 2002, 147. Jg., 205-216. [CrossRef]
- Koch, C. Die Bestimmung der DNA-Sequenz des Geminivirus WDV-ER Genoms und Versuche zur Übertragung des Virus auf Gerste mit Agrobacterium tumefaciens. Diploma thesis, Universität Köln, 1990.
- Schubert, J.; Habekuß, A.; Rabenstein, F. Investigation of differences between wheat and barley forms of Wheat dwarf virus and their distribution in host plants. Plant Protection Science-Prague 2003, 38. Jg., 43-48. [CrossRef]
- Jeske, H. Geminiviruses. TT Viruses: The Still Elusive Human Pathogens 2009, 185-226. [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E. R.; Robertson, D.; Mansoor, S. Geminiviruses: masters at redirecting and reprogramming plant processes. Nature Reviews Microbiology 2013, 11. Jg., Nr. 11, 777-788. [CrossRef]
- Wu, B.; Shang, X.; Schubert, J.; Habekuß, A.; Elena, S. F.; Wang, X. Global-scale computational analysis of genomic sequences reveals the recombination pattern and coevolution dynamics of cereal-infecting geminiviruses. Scientific reports 2015, 5. Jg., Nr. 1, 8153. [CrossRef]
- Van Bel, A. J. E. The phloem, a miracle of ingenuity. Plant, Cell & Environment 2003, 26. Jg., Nr. 1, 125-149. [CrossRef]
- Kammann, M.; Schalk, H.-J.; Matzeit, V.; Schaefer, S.; Schell, J.; Gronenborn, B. DNA replication of wheat dwarf virus, a geminivirus, requires two cis-acting signals. Virology 1991, 184. Jg., Nr. 2, 786-790. [CrossRef]
- Heyraud, F.; Matzeit, V.; Schaefer, S.; Schell, J.; Gronenborn, B. The conserved nonanucleotide motif of the geminivirus stem-loop sequence promotes replicational release of virus molecules from redundant copies. Biochimie 1993, 75. Jg., Nr. 7, 605-615. [CrossRef]
- Laufs, J.; Jupin, I.; David, C.; Schumacher, S.; Heyraud-Nitschke, F.; Gronenborn, B. Geminivirus replication: Genetic and biochemical characterization of Rep protein function, a review. Biochimie 1995, 77. Jg., Nr. 10, 765-773. [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S. B.; Orozco, B. M.; Nagar, S.; Robertson, D. Geminiviruses: Models for Plant DNA Replication, Transcription, and Cell Cycle Regulation. Critical Reviews in Plant Sciences 1999, 18. Jg., Nr. 1, 71-106. [CrossRef]
- Bosque-Pérez, N. A. Eight decades of maize streak virus research. Virus research 2000, 71. Jg., Nr. 1-2, 107-121. [CrossRef]
- Astier, S.; Albouy J.; Maury Y.; Robaglia C.; Lecoq H. Principles of plant virology: genome, pathogenicity, virus ecology. Ecology. Paris: Science Publisher 2007.
- Tomenius, K.; Oxelfelt, P. Preliminary Observations of Viruslike Particles in Nuclei in Cells of Wheat Infected with the Wheat Dwarf Disease. Journal of Phytopathology 1981, 101. Jg., Nr. 2, 163-167. [CrossRef]
- Huth, W.; Lesemann, D.-E. Nachweis des wheat dwarf virus in Deutschland. Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 1994, 46. Jg., Nr. 5, 105-106.
- Hehnle, S.; Wege, C.; Jeske, H. Interaction of DNA with the Movement Proteins of Geminiviruses Revisited. Journal of virology 2004, 78. Jg., Nr. 14, 7698-7706. [CrossRef]
- Evert, R. F.; Russin, W. A.; Botha, C. E. J. Distribution and frequency of plasmodesmata in relation to photoassimilate pathways and phloem loading in the barley leaf. Planta 1996, 198. Jg., 572-579. [CrossRef]
- Aoki, N.; Scofield, G. N.; Wang, X.-D.; Patrick, J. W.; Offler, C. E.; Furbank, R. T. Expression and localisation analysis of the wheat sucrose transporter TaSUT1 in vegetative tissues. Planta 2004, 219. Jg., 176-184. [CrossRef]
- Crawford, K. M.; Zambryski, P. C. Non-Targeted and Targeted Protein Movement through Plasmodesmata in Leaves in Different Developmental and Physiological States. Plant Physiology 2001, 125. Jg., Nr. 4, 1802-1812. [CrossRef]
- Peterschmitt, M.; Quiot, J. B.; Reynaud, B.; Baudin, P. Detection of maize streak virus antigens over time in different parts of maize plants of a sensitive and a so-called tolerant cultivar by ELISA. Annals of applied biology 1992, 121. Jg., Nr. 3, 641-653. [CrossRef]
- Waigmann et al. 2000 cited in Mariano, A. C.; Andrade, M. O.; Santos, A. A.; Carolino, S. M. B.; Oliveira, M. L.; Baracat-Pereira, M. C.; Brommonshenkel, S. H.; Fontes, E. P. B. Identification of a novel receptor-like protein kinase that interacts with a geminivirus nuclear shuttle protein. Virology 2004, 318. Jg., Nr. 1, 24-31. [CrossRef]
- Maule, A.; Leh, V.; Lederer, C. The dialogue between viruses and hosts in compatible interactions. Current opinion in plant biology 2002, 5. Jg., Nr. 4, 279-284. [CrossRef]
- Mariano, A. C.; Andrade, M. O.; Santos, A. A.; Carolino, S. M. B.; Oliveira, M. L.; Baracat-Pereira, M. C.; Brommonshenkel, S. H. & Fontes, E. P. B. Identification of a novel receptor-like protein kinase that interacts with a geminivirus nuclear shuttle protein. Virology 2004, 318. Jg., Nr. 1, 24-31. [CrossRef]
- Mehner, S. Zur Ökologie des Wheat dwarf virus (WDV) in Sachsen-Anhalt. Ph.D. Dissertation, Martin-Luther-Universität Halle-Wittenberg, 2005.
- Lindsten, K.; Lindsten, B. Wheat dwarf - an old disease with new outbreaks in Sweden. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 1999, 325-332.
- Commandeur U.; Huth W. Differentiation of strains of Wheat dwarf virus in infected wheat and barley plants by means of polymerase chain reaction. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 1999, 550-552.
- Schubert, J.; Habekuß, A.; Kazmaier, K.; Jeske, H. Surveying cereal-infecting geminiviruses in Germany—Diagnostics and direct sequencing using rolling circle amplification. Virus research 2007, 127. Jg., Nr. 1, 61-70. [CrossRef]
- Muhire, B.; Martin, D. P.; Brown, J. K.; Navas-Castillo, J.; Moriones, E.; Zerbini, F. M.; Rivera-Bustamante, R.; Malathi, V. G.; Briddon, R. W.; Varsani, A. A genome-wide pairwise-identity-based proposal for the classification of viruses in the genus Mastrevirus (family Geminiviridae). Archives of virology 2013, 158. Jg., 1411-1424. [CrossRef]
- Wu, B.; Melcher, U.; Guo, X.; Wang, X.; Fan, L.; Zhou, G. Assessment of codivergence of Mastreviruses with their plant hosts. BMC evolutionary biology 2008, 8. Jg., Nr. 1, 1-13. [CrossRef]
- Mishchenko L.T.; Dunich A.A.; Mishchenko I.A.; Dashchenko A.V.; Kozub N.O.; Kyslykh T.M.; Molodchenkova O.O. Wheat dwarf virus in Ukraine: occurrence, molecular characterization and impact on the yield. Journal of Plant Diseases and Protection 2022, 129. Jg., Nr. 1, 107-116. [CrossRef]
- Shepherd, D. N.; Martin, D. P.; McGivern, D. R.; Boulton, M. I.; Thomson, J. A.; Rybicki, E. P. A three-nucleotide mutation altering the Maize streak virus Rep pRBR-interaction motif reduces symptom severity in maize and partially reverts at high frequency without restoring pRBR–Rep binding. Journal of general virology 2005, 86. Jg., Nr. 3, 803-813. [CrossRef]
- Schubert J.; Habekuß A.; Wu B.; Thieme T.; Wang X. Analysis of complete genomes of isolates of the Wheat dwarf virus from new geographical locations and descriptions of their defective forms. Virus Genes 2014, 48. Jg., 133-139. [CrossRef]
- Köklü, G.; Ramsell, J. N. E.; Kvarnheden, A. The complete genome sequence for a Turkish isolate of Wheat dwarf virus (WDV) from barley confirms the presence of two distinct WDV strains. Virus Genes 2007, 34. Jg., 359-366. [CrossRef]
- Ramsell, J. N. E.; Boulton, M. I.; Martin, D. P.; Valkonen, J. P. T.; Kvarnheden, A. Studies on the host range of the barley strain of Wheat dwarf virus using an agroinfectious viral clone. Plant pathology 2009, 58. Jg., Nr. 6, 1161-1169. [CrossRef]
- Wu, X.; Weigel, D.; Wigge, P. A. Signaling in plants by intercellular RNA and protein movement. Genes & development 2002, 16. Jg., Nr. 2, 151-158. [CrossRef]
- Owor, B. E.; Shepherd, D. N.; Taylor, N. J.; Edema, R.; Monjane, A. L.; Thomson, J. A.; Martin, D. P.; Varsani, A. Successful application of FTA® Classic Card technology and use of bacteriophage ϕ29 DNA polymerase for large-scale field sampling and cloning of complete maize streak virus genomes. Journal of virological methods 2007, 140. Jg., Nr. 1-2, 100-105. [CrossRef]
- Jungner J. Die Zwergzikade (Cicadula sexnotata Fall.) und ihre Bekämpfung. Berlin DLG 1906.
- Lindsten, K.; Vacke, J.; Gerhardson, B. A preliminary report on three cereal virus diseases new to Sweden spread by Macrosteles and Psammotettix leafhoppers. Meddelanden fran Statens Vaxtskyddsanstalt 1970, 14. Jg., Nr. 123, 285-297.
- Gaborjanyi, R.; Vacke, J. and Bisztray, G. Wheat dwarf virus: a new cereal pathogen in Hungary. Novenytermeles (Hungary) 1988.
- Lapierre, H.; Cousin, M.T.; Della Giustina, W.; Moreau, J.P.; Khogali, M.; et al. Nanisme blé: agent pathogéne et vecteur. Description, biologie, interaction. Phytoma 432 1991, 26–28.
- Conti, M. Leafhopper-borne plant viruses in Italy. Memorie della Societá Entomologica Italiana 72, 1993, 541–547.
- Jilaveanu, A.; Vacke, J. Isolation and identification of wheat dwarf virus (WDV) in Romania. Probleme de protectia plantelor 1995, 23. Jg., 51-62.
- Najar, A.; Makkouk, K.M.; Boudhir, H.; Kumari, S.G.; Zarouk, R.; Bessai, R.; Othman, F.B. Viral diseases of cultivated legume and cereal crops in Tunisia. Viral Diseases of Cultivated Legume and Cereal Crops in Tunisia 2000, 1000-1010.
- Sandgren, M.; Lindblad, M. Field studies of Wheat dwarf virus. In 7th International Congress of Plant Pathology, Edinburgh, UK, 9-16 August 1998.
- Lindsten, K.; Lindsten, B. Occurrence and transmission of Wheat dwarf virus (WDV) in France. In Proceedings of Third International Conference on Pest in Agriculture. Montpellier, France. 1993. 7-9.
- Lindblad, M. What happened to the wheat dwarf disease. Växtskyddsnotiser 2000, 64. Jg., Nr. 1, 11-13.
- Lindsten, K. Wheat dwarf – an old disease caused by a unique and earlier unknown virus. Vaextskyddsnotiser, 1980.
- Dlabola, J. Zur Schädlichkeit der Zikaden in Getreidefeldern. Nachrichtenblatt Deutscher Pflanzenschutzdienst (Berlin), 1961, 14. Jg., 120-122.
- Moreau, J.-P.; Lapierre, H.; Navarro, D.; Debray, P.; Fohrer, F.; Lebrun, I. Distinction des effets du nanisme et de la jaunisse sur le blé. Phytoma, la défense des végétaux 1992, Nr. 443, 21-25.
- Lindsten, K. and Vacke, J. A possible barley adapted strain of wheat dwarf virus (WDV). Acta phytopathologica et entomologica Hungarica 1991, 26. Jg., Nr. 1-2, 175-180.
- Giustina, W. D.; Lebrun, I.; Lapierre, H.; Lochon, S. et le Groupe de travail „Biologie et écologie de P. alienus “. Distribution géographique du vecteur et du virus. Phytoma – La défense des végétaux 1991, 432: 30-34.
- Anonym. New Knowledges about wheat dwarf virus. Phytoma – La défense des végétaux 1992, 443: 17-20.
- Vacher, C.; Felix, I.; Bonnand, E. Lutte contre Psammotettix alienus, Cicadelle vectrice de la maladie des „pieds chétifs “. Perspectives Agricoles 1991, 162: 86-89.
- Pridanceva 1965 cited in Bisztray, G.; Gaborjanyi, R.; Vacke, J. Isolation and characterization of wheat dwarf virus found for the first time in Hungary. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 1989, 449-454.
- Radulescu & Munteanu, 1970 cited in Vacke, J. Host plants range and symptoms of wheat dwarf virus. Věd Pr Výz Ústavú Rostl Výroby Praha-Ruzyně 1972, 17. Jg., 151-162.
- Stephanov & Dimov 1981 cited in Bisztray, G.; Gaborjanyi, R.; Vacke, J. Isolation and characterization of wheat dwarf virus found for the first time in Hungary. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 1989, 449-454.
- Bisztray, G.; Gaborjanyi, R.; Vacke, J. Isolation and characterization of wheat dwarf virus found for the first time in Hungary. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 1989, 449-454.
- Jezewska, J. First report of Wheat dwarf virus occurring in Poland. Phytopathologia Polonica 2001, Nr. 21, S. 93-100.
- Achon, M. A.; Serrano, L.; Ratti, C.; Rubies-Autonell, C. First Detection of Wheat dwarf virus in Barley in Spain Associated with an Outbreak of Barley Yellow Dwarf. Plant Disease 2006, 90. Jg., Nr. 7, 970-970. [CrossRef]
- Viršček Marn, M.; Mavrič Pleško, I. First Report of the Occurrence of Wheat dwarf virus Infecting Wheat in Slovenia. Plant Disease 2017, 101. Jg., Nr. 7, 1336. [CrossRef]
- Behjatnia, S. A. A.; Afsharifar, A. R.; Tahan, V.; Motlagh, M. H. A.; Gandomani, O. E.; Niazi, A.; Izadpanah, K. Widespread occurrence and molecular characterization of Wheat dwarf virus in Iran. Australasian Plant Pathology 2011, 40. Jg., 12-19. [CrossRef]
- Kapooria, R. G.; Ndunguru, J. Occurrence of viruses in irrigated wheat in Zambia. EPPO Bulletin 2004, 34. Jg., Nr. 3, 413-419. [CrossRef]
- Ekzayez, A. M.; Kumari, S. G.; Ismail, I. First Report of Wheat dwarf virus and Its Vector (Psammotettix provincialis) Affecting Wheat and Barley Crops in Syria. Plant Disease 2011, 95. Jg., Nr. 1, 76-76. [CrossRef]
- Xie, J.; Wang, X.; Liu, Y.; Peng, Y.; Zhou, G. First Report of the Occurrence of Wheat dwarf virus in Wheat in China. Plant disease 2007, 91. Jg., Nr. 1, 111-111. [CrossRef]
- Wang X.; Wu B.; Wang J.F.; First report of Wheat dwarf virus infecting barley in Yunnan, China. Journal of Plant Pathology 2008, 90. Jg., Nr. 2, 400-400. [CrossRef]
- Bivand, R.; Lewin-Koh, N.; Pebesma, E.; Archer, E.; Baddeley, A.; Bearman, N.; ... & Golicher, D. Package ‘maptools’.
- pers. Mitteilung Peder Waren in Lindsten, K.; Lindsten, B. (1999) Wheat dwarf - an old disease with new outbreaks in Sweden. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection 1999, 325-332.
- Felix, I.; Larcher, J.M.; Maraby, J.; Philippeau, G.; Vinatier, K. Risques d’attaques de cicadelles et conditions d’efficacité des insecticides. Perspectives agricoles (Paris) 1992, Nr. 173, 98-106.
- Tullgren, A. Zur Morphologie und Systematik der Hemipteren I. Entomologisk Tidskrift / Entomologiska Föreningen i Stockholm 1918, 113–133. [CrossRef]
- ICTV Report Virus taxonomy: classification and nomenclature of viruses: Ninth Report of the International Committee on Taxonomy of Viruses. King, A.M.Q.; Adams, M.J.; Carstens, E.B. and Lefkowitz, E.J (eds). Elsevier Academic Press 2012.
- Ramsell, J. N. E.; Lemmetty, A.; Jonasson, J.; Andersson, A.; Sigvald, R.; Kvarnheden, A. Sequence analyses of Wheat dwarf virus isolates from different hosts reveal low genetic diversity within the wheat strain. Plant pathology 2008, 57. Jg., Nr. 5, 834-841. [CrossRef]
- Vacke J.; Cibulka, R. Silky bent grass (Apera spica-venti [L.] Beauv.) – a new host and reservoir of wheat dwarf virus. Plant protection science 1999, 35. Jg., Nr. 2, 47-50. [CrossRef]
- Lindsten 1991 cited in Brunt, A.; Crabtree, K.; Dallwitz, M.; Gibbs, A.; Watson, L. Viruses of Plants 1996. [CrossRef]
- Fohrer, F.; Lebrun, I.; Lapierre E, H. Acquisitions recéntes sur le virus du nanisme du blé. Phytoma, la défense des végétaux 1992, Nr. 443, S. 18-20.
- Vacke, J.; Cibulka, R. Response of selected winter wheat varieties to wheat dwarf virus infection at an early growth stage. Czech Journal of Genetics and Plant Breeding 2000, 36. Jg., Nr. 1, 1-4.
- Manurung, B.; Witsack, W.; Mehner, S.; Gruntzig, M.; Fuchs, E. The epidemiology of Wheat dwarf virus in relation to occurrence of the leafhopper Psammotettix alienus in Middle-Germany. Virus Research 2004, 100. Jg., Nr. 1, 109-113. [CrossRef]
- Širlová, L.; Vacke, J.; Chaloupková, M. Reaction of selected winter wheat varieties to autumnal infection with Wheat dwarf virus. Plant Prot. Sci 2005, 41. Jg., 1-7. [CrossRef]
- Jones, R. A. C. Global Plant Virus Disease Pandemics and Epidemics. Plants 2021, 10. Jg., Nr. 2, 233. [CrossRef]
- Huth W. Weizenverzwergung - bisher übersehen? Pflanzenschutz-Praxis 1994, 4, 37-39.
- Áy, Z.; Kerényi, Z.; Takács, A.; Papp, M.; Petróczi, I.; Gáborjányi, R.; Silhavy, D.; Pauk, J.; Kertész, Z. Detection of cereal viruses in wheat (Triticum aestivumL.) by serological and molecular methods. Cereal Research Communications 2008, 36. Jg., Nr. 2, 215-224. [CrossRef]
- Lindblad, M.; Arenö, P. Temporal and spatial population dynamics of Psammotettix alienus, a vector of wheat dwarf virus. International journal of pest management 2002, 48. Jg., Nr. 3, 233-238. [CrossRef]
- Nault, L. R. & Ammar, E. D. Leafhopper and Planthopper Transmission of Plant Viruses. Annual review of entomology 1989, 34. Jg., Nr. 1, 503-529. [CrossRef]
- Whitfield, A. E.; Falk, B. W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479. Jg., 278-289. [CrossRef]
- Dietzgen, R. G.; Kondo, H.; Goodin, M. M.; Kurath, G.; Vasilakis, N. The family Rhabdoviridae: mono- and bipartite negative-sense RNA viruses with diverse genome organization and common evolutionary origins. Virus research 2017, 227. Jg., 158-170. [CrossRef]
- Liu, B.; Yuan, R.; Liang, Z.; Zhang, T.; Zhu, M.; Zhang, X.; Geng, W.; Fang, P.; Jiang, M.; Wang, Z.; Feng, Y.; Liu, X.; Zhou, Y.; Xue, R.; Cao, G.; Chen, H.; Hu, X.; Gong, C. Comprehensive analysis of circRNA expression pattern and circRNA–mRNA–miRNA network in Ctenopharyngodon idellus kidney (CIK) cells after grass carp reovirus (GCRV) infection. Aquaculture 2019, 512. Jg., 734349. [CrossRef]
- Du, Z.; Fu, Y.; Liu, Y.; Wang, X. Transmission Characteristics of Wheat Yellow Striate Virus by its Leafhopper Vector Psammotettix alienus. Plant disease 2020, 104. Jg., Nr. 1, 222-226. [CrossRef]
- Liu, Y.; Du, Z.; Wang, H.; Zhang, S.; Cao, M.; Wang, X. Identification and Characterization of Wheat Yellow Striate Virus, a Novel Leafhopper-Transmitted Nucleorhabdovirus Infecting Wheat. Frontiers in microbiology 2018, 9. Jg., 468. [CrossRef]
- Lundsgaard, T. Filovirus-like particles detected in the leafhopper Psammotettix alienus. Virus research 1997, 48. Jg., Nr. 1, 5-40. [CrossRef]
- Bock, J. O.; Lundsgaard, T.; Pedersen, P. A.; Christensen, L. S. Identification and partial characterization of Taastrup virus: a newly identified member species of the Mononegavirales. Virology 2004, 319. Jg., Nr. 1, 49-59. [CrossRef]
- Wang H.; Liu Y.; Liu W.; Cao M.; Wang X. Full genome sequence of a novel iflavirus from the leafhopper Psammotettix alienus. Archives of virology 2019, 164. Jg., 309-311. [CrossRef]
- Wang H.; Liu Y.; Liu W.; Cao M.; Wang X. Sequence analysis and genomic organization of a novel chuvirus, Tàiyuán leafhopper virus. Archives of virology 2019, 164. Jg., 617-620. [CrossRef]
- Han, X.; Wang, H.; Wu, N.; Liu, W.; Cao, M.; Wang, X. Leafhopper Psammotettix alienus hosts chuviruses with different genomic structures. Virus research 2020, 285. Jg., 197-992. [CrossRef]
- Fu, Y.; Cao, M.; Wang, H.; Du, Z.; Liu, Y.; Wang, X. Discovery and characterization of a novel insect-specific reovirus isolated from Psammotettix alienus. Journal of General Virology 2020, 101. Jg., Nr. 8, 884-892. [CrossRef]
- Harding, R. M.; Burns, P.; Geijskes, R. J.; McQualter, R. M.; Dale, J. L.; Smith, G. R. Molecular Analysis of Fiji Disease Virus Segments 2, 4 and 7 Completes the Genome Sequence. Virus Genes 2006, 32. Jg., 43-47. [CrossRef]
- Lot, H.; Delecolle, B.; Boccardo, G.; Marzachi, C.; Milne, R. G. Partial characterization of reovirus-like particles associated with garlic dwarf disease. Plant pathology 1994, 43. Jg., Nr. 3, 537-546. [CrossRef]
- Xie, L.; LV, M.-F.; Yang, J.; Chen, J.-P.; Zhang, H.-M. Genomic and phylogenetic evidence that Maize rough dwarf and Rice black-streaked dwarf fijiviruses should be classified as different geographic strains of a single species. Acta virologica 2017, 61. Jg., Nr. 4, 453-462. [CrossRef]
- Distéfano, A. J.; Conci, L. R.; Muñoz Hidalgo, M.; Guzmán, F. A.; Hopp, H. E. & del Vas, M. Sequence and phylogenetic analysis of genome segments S1, S2, S3 and S6 of Mal de Rı́o Cuarto virus, a newly accepted Fijivirus species. Virus research 2003, 92. Jg., Nr. 1, 113-121. [CrossRef]
- Isogai, M.; Lindsten, K.; Uyeda, I. Taxonomic characteristics of fijiviruses based on nucleotide sequences of the oat sterile dwarf virus genome. Journal of general virology 1998, 79. Jg., Nr. 6, 1479-1485. [CrossRef]
- Teakle, D.; Hicks, S.; Karan, M.; Hacker, J.; Greber, R.; Donaldson, J. Host range and geographic distribution of pangola stunt virus and its planthopper vectors in Australia. Australian Journal of Agricultural Research 1991, 42. Jg., Nr. 5, 819-826. [CrossRef]
- Zhou, G.; Wen, J.; Cai, D.; Li, P.; Xu, D.; Zhang, S. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chinese science bulletin 2008, 53. Jg., Nr. 23, 3677-3685. [CrossRef]
- Nakashima, N.; Koizumi, M.; Watanabe, H.; Noda, H. Complete nucleotide sequence of the Nilaparvata lugens reovirus: a putative member of the genus Fijivirus. Journal of General Virology 1996, 77. Jg., Nr. 1, 139-146. [CrossRef]
- Zhang, X.; Zhou, G.; Wang, X. Detection of wheat dwarf virus (WDV) in wheat and vector leafhopper (Psammotettix alienus Dahlb.) by real-time PCR. Journal of virological methods 2010, 169. Jg., Nr. 2, 416-419. [CrossRef]
- Le Roux, J. J.; Rubinoff, D. Molecular data reveals California as the potential source of an invasive leafhopper species, Macrostelessp. nr. severini, transmitting the aster yellows phytoplasma in Hawaii. Annals of Applied Biology, 154(3), 419–427. [CrossRef]
- Derlink, M.; Pavlovčič, P.; Stewart, A. J. A.; Virant-Doberlet, M. Mate recognition in duetting species: the role of male and female vibrational signals. Animal behaviour 2014, 90. Jg., 181-193. [CrossRef]
- Abt, I.; Jacquot, E. (2015). Wheat dwarf. In Virus Diseases of Tropical and Subtropical Crops, Tennant, P., Fermin, R., Eds.; Plant Protection Series; CAB International: Boston, MA, USA, 2015; pp. 27–41. [CrossRef]
- Vilbaste, J. Preliminary key for the identification of the nymphs of North European Homoptera, Cicadinea. II. Cicadelloidea. Annales Zoologici Fennici 19 1982, 1–20.
- Della Giustina, W. Homoptères Cicadellidae 3: compléments Faune de France 73. Fédération Française des Sociétés de Sciences Naturelles, INRA éditions 1989.
- Biedermann R.; Niedrighaus, R. Die Zikaden Deutschlands. In Bestimmungstafeln für alle Arten. Wissenschaftlich Akademischer Buchvertrieb-Fründ 2004.
- Kunz G., Nickel H.; Niedrighaus, R. In Photographic atlas of the planthoppers and leafhoppers of Germany. Wissenschaftlich Akademischer Buchvertrieb-Fründ 2011.
- Tishechkin, D. Yu. The variability of acoustic signals and some morphological characters in Psammotettix striatus (Homoptera, Cicadellidae) from Russia and adjacent countries. Зooлoгический журнал 1999, 78. Jg., Nr. 11.
- Biedermann, R.; Niedringhaus, R. The Plant- and Leafhoppers of Germany: Identification Key to all Species. Wabv Fründ, 2009.
- Heady, S. E.; Nault, L. R.; Shambaugh, G. F.; Fairchild, L. Acoustic and Mating Behavior of Dalbulus Leaf hoppers (Homoptera: Cicadellidae). Annals of the Entomological Society of America 1986, 79. Jg., Nr. 4, 727-736. [CrossRef]
- Gillham, M. C. Variation in acoustic signals within and among leafhopper species of the genus Alebra (Homoptera, Cicadellidae). Biological Journal of the Linnean Society 1992, 45. Jg., Nr. 1, 1-15. [CrossRef]
- Tishechkin, D. Yu. Vibrational communication in Aphrodinae leafhoppers (Deltocephalinae auct.; Homoptera: Cicadellidae) and related groups with notes on classification of higher taxa. Russian Entomological Journal 2000, 9. Jg., Nr. 1, 1-66.
- Percy, D. M.; Boyd, E. A.; Hoddle, M. S. Observations of acoustic signaling in three sharpshooters: Homalodisca vitripennis, Homalodisca liturata, and Graphocephala atropunctata (Hemiptera: Cicadellidae). Annals of the Entomological Society of America 2008, 101. Jg., Nr. 1, 253-259. [CrossRef]
- Tishechkin, D. Yu. The use of bioacoustic characters for distinguishing between cryptic species in insects: Potentials, restrictions, and prospects. Entomological Review 2014, 94. Jg., 289-309. [CrossRef]
- Derlink, M.; Abt, I.; Mabon, M.; Julian, C.; Virant-Doberlet, M.; Jacquot, E. Mating behaviour of Psammotettix alienus Dahlbom (Hemiptera: Cicadellidae). Insect Science 2018, 25. Jg., Nr. 1, 148-160. [CrossRef]
- Schlick-Steiner, B. C.; Steiner, F. M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R. H. Integrative Taxonomy: A Multisource Approach to Exploring Biodiversity. Annual review of entomology 2010, 55. Jg., 421-438. [CrossRef]
- Bluemel, J. K.; Derlink, M.; Pavlovcic, P.; Russio, I.-R. M.; Andrew King, R.; Corbett, E.; Sherrard-Smith, E.; Blejec, A.; Wilson, M. R.; Stewart, A. J. A.; Symondson, W. O. C.; Virant-Doberlet, M. Integrating vibrational signals, mitochondrial DNA and morphology for species determination in the genus Aphrodes (Hemiptera: Cicadellidae). Systematic Entomology 2014, 39. Jg., Nr. 2, 304-324. [CrossRef]
- Kamitani, S. (2011). DNA Barcodes of Japanese Leafhoppers. ESAKIA 2011, 50, 81–88. [CrossRef]
- Gwiazdowski, R. A.; Foottit, R. G.; Maw, H. E. L.; Hebert, P. D. N. The Hemiptera (Insecta) of Canada: Constructing a Reference Library of DNA Barcodes. PLoS One 2015, 10. Jg., Nr. 4, S. e0125635. [CrossRef]
- Abt, I.; Derlink, M.; Mabon, R.; Virant-Doberlet, M.; Jacquot, E. Integrating multiple criteria for the characterization of Psammotettix populations in European cereal fields. Bulletin of entomological research 2018, 108. Jg., Nr. 2, 185-202. [CrossRef]
- Guglielmino, A.; Virla, E.G. Postembryonic development and biology of Psammotettix alienus (Dahlbom) (Homoptera, Cicadellidae) under laboratory conditions. Bollettino di Zoologia, Agraria e di Bachicoltura 1997, 29. Jg., 65-80.
- Manurung, B. Untersuchungen zur Biologie und Ökologie der Zwergzikade Psammotettix alienus Dahlb. (Auchenorrhyncha) und zu ihrer Bedeutung als Vektor des Wheat dwarf virus (Weizenverzwergungs-Virus, WDV). Ph.D. Dissertation, Martin–Luther-Universität Halle-Wittenberg, 2002.
- Schiemenz, H.; Emmrich, R.; Witsack, W. Beiträge zur Insektenfauna Ost- deutschlands: Homoptera – Auchenorrhyncha (Cicadina) (Insecta) Teil IV: Unterfamilie Deltocephalinae. Faunistische Abhandlungen 1996, 20. Jg., 153-258.
- Alla, S.; Moreau, J. P.; Frerot, B. Effects of the aphid Rhopalosiphum padi on the leafhopper Psammotettix alienus under laboratory conditions. Entomologia Experimentalis et Applicata 2001, 98. Jg., Nr. 2, 203-209. [CrossRef]
- Van Nieuwenhove, G. A.; Frías, E. A.; Virla, E. G. Effects of temperature on the development, performance and fitness of the corn leafhopperDalbulus maidis (DeLong) (Hemiptera: Cicadellidae): implications on its distribution under climate change. Agricultural and Forest Entomology 2016, 18. Jg., Nr. 1, 1-10. [CrossRef]
- Schiemenz, H. Die Zikadenfauna mitteleuropäischer Trockenrasen (Homoptera, Auchenorrhyncha). Entomologische Abhandlungen Staatliches Museum für Tierkunde in Dresden 1969, 36. Jg., 201-280.
- Ghodoum Parizipour M.H.; Behjatnia S.A.A.; Afsharifar A.; Izadpanah K. Natural hosts and efficiency of leafhopper vector in transmission of Wheat dwarf virus. Journal of Plant Pathology 2016, 483-492. [CrossRef]
- Lindblad, M.; Sigvald, R. Temporal spread of wheat dwarf virus and mature plant resistance in winter wheat. Crop protection 2004, 23. Jg., Nr. 3, 229-234. [CrossRef]
- Backus, E. A. Sensory systems and behaviours which mediate hemipteran plant-feeding: A taxonomic overview. Journal of Insect Physiology 1988, 34. Jg., Nr. 3, 151-165. [CrossRef]
- Zhao, L.; Dai, W.; Zhang, C.; Zhang, Y. Morphological characterization of the mouthparts of the vector leafhopper Psammotettix striatus (L.) (Hemiptera: Cicadellidae). Micron 2010, 41. Jg., Nr. 7, 754-759. [CrossRef]
- Hewer A.; Becker A.; van Bel A.J.E. An aphid’s Odyssey –the cortical quest for the vascular bundle. Journal of Experimental Biology 2011, 214. Jg., Nr. 22, 3868-3879. [CrossRef]
- Storey, H. H. Transmission studies of maize streak disease. Annals of applied biology 1928, 15. Jg., Nr. 1, 1-25. [CrossRef]
- Wang, Y.; Dang, M.; Hou, H.; Mei, Y.; Qian, Y.; Zhou, X. Identification of an RNA silencing suppressor encoded by a mastrevirus. Journal of General Virology 2014, 95. Jg., Nr. 9, 2082-2088. [CrossRef]
- Reynaud, B.; Peterschmitt, M. A study of the mode of transmission of maize streak virus by Cicadulina mbila using an enzyme-linked immunosorbent assay. Annals of applied biology 1992, 121. Jg., Nr. 1, 85-94. [CrossRef]
- Tholt, G.; Samu, F.; Kiss, B. Feeding behaviour of a virus-vector leafhopper on host and non-host plants characterised by electrical penetration graphs. Entomologia Experimentalis et Applicata 2015, 155. Jg., Nr. 2, 123-136. [CrossRef]
- Hogenhout, S. A.; Ammar, E.-D.; Whitfield, A. E.; Redinbaugh, M. G. Insect Vector Interactions with Persistently Transmitted Viruses. Annu. Rev. Phytopathol. 2008, 46. Jg., 327-359. [CrossRef]
- Brault, V.; Uzest, M.; Monsion, B.; Jacquot, E.; Blanc, S. Aphids as transport devices for plant viruses. Comptes rendus biologies 2010, 333. Jg., Nr. 6-7, 524-538. [CrossRef]
- Yazdkhasti, E.; Hopkins, R. J.; Kvarnheden, A. Reservoirs of plant virus disease: Occurrence of wheat dwarf virus and barley/cereal yellow dwarf viruses in Sweden. Plant Pathology 2021, 70. Jg., Nr. 7, 1552-1561. [CrossRef]
- Nickel H.; Remane R. Artenliste der Zikaden Deutschands, (Checklist of the planthoppers and leafhoppers of Germany, with notes on food plants, diet width, life cycles, geographic range and conservation status). Beiträge zur Zikadenkunde 2002, 5. Jg., 27-64.
- Ossiannilsson, F. The Auchenorrhyncha (Homoptera) of Fennoscandia and Denmark. Part 2: The Families Cicadidae, Cercopidae, Membracidae, and Cicadellidae (excl. Deltocephalinae). Scandinavian Science Press 1981, 7. Jg., Nr. 2, 223-593. [CrossRef]
- Koppányi, T. A lucernásban kalakuló Cicadinea együttes évszakos és állományok korával járó változásának vizsgálata. A study into the seasonal and crop age related changes of Cicadinea assemblages in alfalfa.). Debreceni Agrártudományi Egyetem Tudományos Közleményei 1976, 18. Jg., 27-60.
- Drobnjakovic, T.; Peric, P.; Marcic, D.; Picciau, L.; Alma, A.; Mitrovic, J.; Duduk, B.; Bertaccini, A. Leafhoppers and cixiids in phytoplasma-infected carrot fields: Species composition and potential phytoplasma vectors. Pesticidi i Fitomedicina 2010, 25(4), 311–318. [CrossRef]
- Kiss B.; Redei D.; Koczor S. Occurrence and feeding of hemipterans on common ragweed (Ambrosia artemisiifolia) in Hungary. Bulletin of Insectology 2008, 61. Jg., Nr. 1, 195-196.
- Riedle-Bauer M.; Tiefenbrunner W.; Otreba J.; Hanak K.; Schildberger B.; Regner F. Epidemiological observations on Bois Noir in Austrian vineyards. Mitteilungen Klosterneuburg 2006, 56:177–181.
- Landi, L.; Isidoro, N.; Riolo, P. Natural Phytoplasma Infection of Four Phloem-Feeding Auchenorrhyncha Across Vineyard Agroecosystems in Central-Eastern Italy. Journal of Economic Entomology 2013, 106. Jg., Nr. 2, 604-613. [CrossRef]
- Tholt, G.; Kiss, B. Host range of Psammotettix alienus (Dahlbom). Növényvédelem 2011, 47. Jg., Nr. 6, 229-235.
- Stafford, C. A.; Walker, G. P. Characterization and correlation of DC electrical penetration graph waveforms with feeding behavior of beet leafhopper, Circulifer tenellus. Entomologia Experimentalis et Applicata 2009, 130. Jg., Nr. 2, 113-129. [CrossRef]
- Alvarez, A. E.; Tjallingii, W. F.; Garzo, E.; Vleeshouwers, V.; Dicke, M.; Vosman, B. Location of resistance factors in the leaves of potato and wild tuber-bearing Solanum species to the aphid Myzus persicae. Entomologia Experimentalis et Applicata 2006, 121. Jg., Nr. 2, 145-157. [CrossRef]
- Lei, H.; Lenteren, J. C.; Tjallingii, W. F. Analysis of resistance in tomato and sweet pepper against the greenhouse whitefly using electrically monitored and visually observed probing and feeding behaviour. Entomologia Experimentalis et Applicata 1999, 92. Jg., Nr. 3, 299-309. [CrossRef]
- Lei, H.; Van Lenteren, J. C.; Xu, R. M. Effects of plant tissue factors on the acceptance of four greenhouse vegetable host plants by the greenhouse whitefly: an Electrical Penetration Graph (EPG) study. European Journal of Entomology 2001, 98. Jg., Nr. 1, 31-36. [CrossRef]
- McLean, D. L.; Kinsey, M. G. A Technique for Electronically Recording Aphid Feeding and Salivation. Nature 1964, 202. Jg., Nr. 4939, 1358-1359. [CrossRef]
- Tjallingii, W. F. Electronic recording of penetration behaviour by aphids. Entomologia experimentalis et applicata 1978, 24. Jg., Nr. 3, 721-730. [CrossRef]
- Tjallingii, W. F. Electrical nature of recorded signals during stylet penetration by aphids. Entomologia experimentalis et applicata 1985, 38. Jg., Nr. 2, 177-186. [CrossRef]
- Tjallingii, W. F. Electrical recording of stylet penetration activities. In Aphids, their biology, natural enemies and control. Elsevier Science Publishers 1988. 95-108.
- Prado, E.; Tjallingii, W. F. Aphid activities during sieve element punctures. Entomologia experimentalis et applicata 1994, 72. Jg., Nr. 2, S. 157-165. [CrossRef]
- Harrewijn, P.; Kayser, H. Pymetrozine, a Fast-Acting and Selective Inhibitor of Aphid Feeding. In-situStudies with Electronic Monitoring of Feeding Behaviour. Pesticide Science 1997, 49. Jg., Nr. 2, S. 130-140. [CrossRef]
- Mesfin T.; Bosque-Pérez N.A. Feeding behavior of Cicadulina storeyi China (Homoptera: Cicadellidae) on maize varieties susceptible or resistant to maize streak virus. African entomology 1998, 6. Jg., Nr. 2, S. 185-191.
- Helden, M. van; Tjallingii, W. F. Experimental Design and Analysis in EPG Experiments with Emphasis on Plant Resistance Research. In Principles and applications of electronic monitoring and other techniques in the study of homopteran feeding behavior. Entomological Society of America 2000. 144. [CrossRef]
- Liu, X. D.; Zhai, B. P.; Zhang, X. X.; Zong, J. M. Impact of transgenic cotton plants on a non-target pest, Aphis gossypii Glover. Ecological Entomology 2005, 30. Jg., Nr. 3, 307-315. [CrossRef]
- Prado, E.; Fred Tjallingii, W. Behavioral Evidence for Local Reduction of Aphid-Induced Resistance. Journal of Insect Science 2007, 7. Jg., Nr. 1, 48. [CrossRef]
- Martin, B.; Fereres, A.; Tjallingii, W. F.; Collar, J. L. Intracellular ingestion and salivation by aphids may cause the acquisition and inoculation of non-persistently transmitted plant viruses. Journal of General Virology 1997, 78. Jg., Nr. 10, 2701-2705. [CrossRef]
- Sauge M.H.; Lambert P.; Pascal T. Co-localisation of host plant resistance QTLs affecting the performance and feeding behaviour of the aphid Myzus persicae in the peach tree. Heredity 2012, 108. Jg., Nr. 3, 292-301. [CrossRef]
- Giordanengo, P. EPG-Calc: a PHP-based script to calculate electrical penetration graph (EPG) parameters. Arthropod-Plant Interactions 2014, 8. Jg., 163-169. [CrossRef]
- Tjallingii, W. F.; Esch, TH. H. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiological entomology 1993, 18. Jg., Nr. 3, 317-328. [CrossRef]
- Khan, Z. R.; Saxena, R. C. Probing Behavior of Three Biotypes of Nilaparvata lugens (Homoptera: Delphacidae) on Different Resistant and Susceptible Rice Varieties. Journal of Economic Entomology 1988, 81. Jg., Nr. 5, 1338-1345. [CrossRef]
- Kimmins, F. M. Electrical penetration graphs from Nilaparvata lugens on resistant and susceptible rice varieties. Entomologia experimentalis et applicata 1989, 50. Jg., Nr. 1, 69-79. [CrossRef]
- Powell, K. S.; Gatehouse, J. A. Mechanism of mannose-binding snowdrop lectin for use against brown planthopper in rice. Rice Genetics III: (In 2 Parts) 1996. 753-758. [CrossRef]
- Seo, B. Y.; Kwon, Y.-H.; Jung, J. K.; Kim, G.-H. Electrical penetration graphic waveforms in relation to the actual positions of the stylet tips of Nilaparvata lugens in rice tissue. Journal of Asia-Pacific Entomology 2009, 12. Jg., Nr. 2, 89-95. [CrossRef]
- Calatayud, P.-A.; Seligmann, C. D.; Polania, M. A.; Bellotti, A. C. Influence of parasitism by encyrtid parasitoids on the feeding behaviour of the cassava mealybug Phenacoccus herreni. Entomologia Experimentalis et Applicata 2001, 98. Jg., Nr. 3, 271-278. [CrossRef]
- Harrewijn P.; Piron P.G.M.; Ponsen M.B. Evolution of vascular feeding in aphids: an electrophysiological study. Entomologia Experimentalis et Applicata 1998, 9. Jg., 29-34.
- Kingston K.B. Digestive and feeding physiology of grape phylloxera (Daktulosphaira vitifoliae Fitch), Ph.D. thesis, Australian National University, Canberra, 2007.
- Joost P.H.; Riley D.G. Imidacloprid effects on probing and settling behavior of Frankliniella fusca and Frankliniella occidentalis (Thysanoptera: Thripidae) in tomato. Journal of economic entomology 2005, 98. Jg., Nr. 5, 1622-1629. [CrossRef]
- Kindt, F.; Joosten, N. N.; Tjallingii, W. F. Electrical penetration graphs of thrips revised: Combining DC- and AC-EPG signals. Journal of Insect Physiology 2006, 52. Jg., Nr. 1, 1-10. [CrossRef]
- Walker, G. P.; Janssen, J. A. M. Electronic Recording of Whitefly (Homoptera: Aleyrodidae) Feeding and Oviposition Behavior. Principles and Applications of Electronic Monitoring and Other Techniques in the Study of Homopteran Feeding Behavior. In Principles and applications of electronic monitoring and other techniques in the study of Homopteran feeding behavior. Entomological Society of America 2000. 172. [CrossRef]
- Kimmins, F. M.; Bosque-Perez, N. A. Electrical penetration graphs from Cicadulina spp. and the inoculation of a persistent virus into maize. Proceedings of the 9th International Symposium on Insect-Plant Relationships 1996. 46–49. [CrossRef]
- Lett, J.-M.; Granier, M.; Grondin, M.; Turpin, P.; Molinaro, F.; Chiroleu, F.; Peterschmitt, M.; Reynaud, B. Electrical penetration graphs from Cicadulina mbila on maize, the fine structure of its stylet pathways and consequences for virus transmission efficiency. Entomologia Experimentalis et Applicata 2001, 101. Jg., Nr. 2, 93-109. [CrossRef]
- Miranda, M. P.; Fereres, A.; Appezzato-da-Gloria, B.; Lopes, J. R. S. Characterization of electrical penetration graphs of Bucephalogonia xanthophis, a vector of Xylella fastidiosain citrus. Entomologia Experimentalis et Applicata 2010, 134. Jg., Nr. 1, 35-49. [CrossRef]
- Tjallingii, W. F.; Esch, TH. H. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiological entomology 1993, 18. Jg., Nr. 3, 317-328. [CrossRef]
- Tjallingii, W. F. Salivary secretions by aphids interacting with proteins of phloem wound responses. Journal of experimental botany 2006, 57. Jg., Nr. 4, 739-745. [CrossRef]
- Wayadande, A. C. Leafhopper Probing Behavior Associated with Maize Chlorotic Dwarf Virus Transmission to Maize. Phytopathology 1993, 83. Jg., Nr. 5, 522-526. [CrossRef]
- Backus, E. A.; Holmes, W. J.; Schreiber, F.; Reardon, B. J.; Walker, G. P. Sharpshooter X Wave: Correlation of an Electrical Penetration Graph Waveform with Xylem Penetration Supports a Hypothesized Mechanism for Xylella fastidiosa Inoculation. Annals of the Entomological Society of America 2009, 102. Jg., Nr. 5, 847-867. [CrossRef]
- Pillon, O. Importance en Champagne de deux parasitoides de la cicadelle vectrice du nanisme du ble. In ANPP – 3rd International Conference on Pests in Agriculture, France, 7-9 December 1993.
- Samu, F.; Beleznai, O.; Tholt, G. A potential spider natural enemy against virus vector leafhoppers in agricultural mosaic landscapes – Corroborating ecological and behavioral evidence. Biological Control 2013, 67. Jg., Nr. 3, 390-396. [CrossRef]
- Kogan, M. Integrated Pest Management: Historical Perspectives and Contemporary Developments. Annual review of entomology 1998, 43. Jg., Nr. 1, S. 243-270. [CrossRef]
- Shepherd, D. N.; Martin, D. P.; van der Walt, E.; Dent, K.; Varsani, A.; Rybicki, E. P. Maize streak virus: an old and complex ‘emerging’ 62 pathogen. Molecular plant pathology 2010, 11. Jg., Nr. 1, 1-12. [CrossRef]
- Yazdkhasti, E. (2022). Epidemiology of wheat dwarf virus. Ph.D. Dissertation, Acta Universitatis Agriculturae Sueciae, 2022.
- Eweida, M.; Oxelfelt, P.; Tomenius, K. Concentration of virus and ultrastructural changes in oats at various stages of barley yellow dwarf virus infection. Annals of applied biology 1988, 112. Jg., Nr. 2, 313-321. [CrossRef]
- Aranda, M. A.; Freitas-Astúa, J. Ecology and diversity of plant viruses, and epidemiology of plant virus-induced diseases. Annals of Applied Biology 2017, 171. Jg., Nr. 1, 1-4. [CrossRef]
- Weibull, A.-C.; Östman, Ö. Species composition in agroecosystems: The effect of landscape, habitat, and farm management. Basic and Applied Ecology 2003, 4. Jg., Nr. 4, 349-361. [CrossRef]
- Ansari, M. S.; Moraiet, M. A.; Ahmad, S. Insecticides: Impact on the Environment and Human Health. Environmental deterioration and human health: natural and anthropogenic determinants 2014, 99-123. [CrossRef]
- Biondi, A.; Desneux, N.; Siscaro, G.; Zappalà, L. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: Selectivity and side effects of 14 pesticides on the predator Orius laevigatus. Chemosphere 2012, 87. Jg., Nr. 7, 803-812. [CrossRef]
- Dudley, N.; Attwood, S. J.; Goulson, D.; Jarvis, D.; Bharucha, Z. P. Pretty, J. How should conservationists respond to pesticides as a driver of biodiversity loss in agroecosystems? Biological Conservation 2017, 209. Jg., 449-453. [CrossRef]
- Guimarães-Cestaro, L.; Martins, M. F.; Martínez, L. C.; Alves, M. L. T. M. F.; Guidugli-Lazzarini, K. R.; Nocelli, R. C. F.; Malaspina, O.; Serrão, J. E.; Teixeira, É. W. Occurrence of virus, microsporidia, and pesticide residues in three species of stingless bees (Apidae: Meliponini) in the field. The Science of Nature 2020, 107. Jg., 1-14. [CrossRef]
- Staskawicz, B. J. Genetics of Plant-Pathogen Interactions Specifying Plant Disease Resistance. Plant Physiology 2001, 125. Jg., Nr. 1, 73-76. [CrossRef]
- Wang, H.; Hao, L.; Shung, C.-Y.; Sunter, G.; Bisaro, D. M. Adenosine Kinase Is Inactivated by Geminivirus AL2 and L2 Proteins. The Plant Cell 2003, 15. Jg., Nr. 12, 3020-3032. [CrossRef]
- Leke, W.N. Molecular Epidemiology of Begomoviruses that Infect Vegetable Crops in Southwestern Cameroon. Ph.D. Thesis, Acta Universitatis Agriculturae Sueciae, Uppsala, Sweden, 2010.
- Wulff, B. B. H.; Moscou, M. J. Strategies for transferring resistance into wheat: from wide crosses to GM cassettes. Frontiers in Plant Science 2014, 5. Jg., 692. [CrossRef]
- Kanyuka, K.; Lovell, D. J.; Mitrofanova, O. P.; Hammond-Kosack, K.; Adams, M. J. A controlled environment test for resistance to Soil-borne cereal mosaic virus (SBCMV) and its use to determine the mode of inheritance of resistance in wheat cv. Cadenza and for screening Triticum monococcum genotypes for sources of SBCMV resistance. Plant Pathology 2004, 53. Jg., Nr. 2, 154-160. [CrossRef]
- Ward E.; Kanyuka K.; Motteram J.; Kornyukhin D.; Adams M.J. The use of conventional and quantitative real-time PCR assays for Polymyxa graminis to examine host plant resistance, inoculum levels and intraspecific variation. New Phytologist 2005, 875-885. [CrossRef]
- Hall, M. D.; Brown-Guedira, G.; Klatt, A.; Fritz, A. K. Genetic analysis of resistance to soil-borne wheat mosaic virus derived from Aegilops tauschii. Euphytica 2009, 169. Jg., 169-176. [CrossRef]
- Zaharieva, M.; Monneveux, P.; Henry, M.; Rivoal, R.; Valkoun, J.; Nachit, M. M. Evaluation of a Collection of Wild Wheat Relative Aegilops Geniculata Roth and Identification of Potential Sources for Useful Traits. Developments in Plant Breeding 2001. 739–746. [CrossRef]
- Pfrieme, A.-K.; Ruckwied, B.; Habekuß, A.; Will, T.; Stahl, A.; Pillen, K.; Ordon, F. Identification and Validation of Quantitative Trait Loci for Wheat Dwarf Virus Resistance in Wheat (Triticum spp.). Frontiers in Plant Science 2022, 13. Jg., 828639. [CrossRef]
- Pfrieme, A. K.; Stahl, A.; Pillen, K.; Will, T. Comparison of two different experimental environments for resistance screenings for leafhopper-transmitted wheat dwarf virus in wheat. Journal of Plant Diseases and Protection 2023, submitted.
- Nygren, J.; Shad, N.; Kvarnheden, A.; Westerbergh, A. Variation in Susceptibility to Wheat dwarf virus among Wild and Domesticated Wheat. PloS one 2015, 10. Jg., Nr. 4, e0121580. ttps://doi.org/10.1371/journal.pone.0121580.
- Riedel, C.; Habekuß, A.; Schliephake, E.; Niks, R.; Broer, I.; Ordon, F. Pyramiding of Ryd2 and Ryd3 conferring tolerance to a German isolate of Barley yellow dwarf virus-PAV (BYDV-PAV-ASL-1) leads to quantitative resistance against this isolate. Theoretical and applied genetics 2011, 123. Jg., Nr. 1, 69-76. [CrossRef]
- Scheurer, K. S.; Friedt, W.; Huth, W.; Waugh, R.; Ordon, F. QTL analysis of tolerance to a German strain of BYDV-PAV in barley (Hordeum vulgare L.). Theoretical and Applied Genetics 2001, 103. Jg., 1074-1083. [CrossRef]
- Clark, M. F.; Adams, A. N. Characteristics of the Microplate Method of Enzyme-Linked Immunosorbent Assay for the Detection of Plant Viruses. Journal of general virology 1977, 34. Jg., Nr. 3, 475-483. [CrossRef]
- Hayes, R. J.; Macdonald, H.; Coutts, R. H. A.; Buck, K. W. Agroinfection of Triticum aestivum with Cloned DNA of Wheat Dwarf Virus. Journal of general virology 1988, 69. Jg., Nr. 4, 891-896. [CrossRef]
- Boulton, M. I. Agrobacterium-Mediated Transfer of Geminiviruses to Plant Tissues. Plant Gene Transfer and Expression Protocols 1995, 77-93. [CrossRef]
- Boulton, M. I. Construction of Infectious Clones for DNA Viruses: Mastreviruses. Plant Virology Protocols: From Viral Sequence to Protein Function 2008, 503-523. [CrossRef]
- Bukvayová, N.; Henselová, M.; Vajcíková, V.; Kormanová, T. Occurrence of dwarf virus of winter wheat and barley in several regions of Slovakiaduring the growing seasons 2001-2004. Plant Soil and Environment 2006, 52. Jg., Nr. 9, 392. [CrossRef]
- Rabenstein, F.; Sukhacheva, E.; Habekuß, A.; Schubert, J. Differentiation of Wheat dwarf virus isolates from wheat, triticale, and barley by means of a monoclonal antibody. In Proceedings of the X Conference on Viral Diseases of Gramineae in Europe, Louvain-la-Neuve, Belgium, 2005. 60.
- Kundu, J. K.; Gadiou, S.; Červená, G. Discrimination and genetic diversity of Wheat dwarf virus in the Czech Republic. Virus genes 2009, 38. Jg., 468-474. [CrossRef]
- Glais, L.; Jacquot, E. Detection and Characterization of Viral Species/Subspecies Using Isothermal Recombinase Polymerase Amplification (RPA) Assays. Plant Pathology: Techniques and Protocols 2015, 207-225. [CrossRef]
- Thresh, J. M. Cropping Practices and Virus Spread. Annual Review of Phytopathology 1982, 20. Jg., Nr. 1, 193-216. [CrossRef]
- Benkovics, A. H.; Vida, G.; Nelson, D.; Veisz, O.; Bedford, I.; Silhavy, D.; Boulton, M. I. Partial resistance to Wheat dwarf virus in winter wheat cultivars. Plant pathology 2010, 59. Jg., Nr. 6, 1144-1151. [CrossRef]
- Schneider, A.; Molnár-Láng, M. Detection of the 1RS chromosome arm in Martonvásár wheat genotypes containing 1Bl.1Rs or 1Al.1Rs translocations using SSR and STS markers. Acta Agronomica Hungarica 2009, 57. Jg., Nr. 4, 409-416. [CrossRef]
- Schlegel R. Current list of wheats with rye and alien introgression. From http://www.ryegene-map.de/rye-introgression/html/entry_m.html. Retrieved 22/05/2020.
- Ruckwied, B. Sources of resistance and identification of QTLs for Wheat dwarf virus (WDV) resistance in wheat (Triticum spp.) and wild relatives. Julius Kühn-Institut, Bundesforschungsinstitut für Kulturpflanzen, 2022.
- Kumar, R. V. Plant Antiviral Immunity Against Geminiviruses and Viral Counter-Defense for Survival. Frontiers in Microbiology 2019, 10. Jg., 1460. [CrossRef]
- Sharaf, A.; Nuc, P.; Ripl, J.; Alquicer, G.; Ibrahim, E.; Wang, X.; Maruthi, M. N.; Kundu, J. K. Transcriptome Dynamics in Triticum aestivum Genotypes Associated with Resistance against the Wheat Dwarf Virus. Viruses 2023, 15. Jg., Nr. 3, 689. [CrossRef]
- Schoelz, J. E.; Harries, P. A.; Nelson, R. S. Intracellular Transport of Plant Viruses: Finding the Door out of the Cell. Molecular plant 2011, 4. Jg., Nr. 5, 813-831. [CrossRef]
- Frederickson Matika, D. E.; Loake, G. J. Redox Regulation in Plant Immune Function. Antioxidants & redox signaling 2014, 21. Jg., Nr. 9, 1373-1388. [CrossRef]
- Mayer, M. P. Recruitment of Hsp70 chaperones: a crucial part of viral survival strategies. Reviews of physiology, biochemistry and pharmacology 2005, 1-46. [CrossRef]
- Nagy, P. D.; Pogany, J. The dependence of viral RNA replication on co-opted host factors. Nature Reviews Microbiology 2012, 10. Jg., Nr. 2, 137-149. [CrossRef]
- Yuan, W.; Jiang, T.; Du, K.; Chen, H.; Cao, Y.; Xie, J.; Li, M.; Carr, J. P.; Wu, B.; Fan, Z.; Zhou, T. Maize phenylalanine ammonia-lyases contribute to resistance toSugarcane mosaic virusinfection, most likely through positive regulation of salicylic acid accumulation. Molecular plant pathology 2019, 20. Jg., Nr. 10, 1365-1378. [CrossRef]
- Zhu, F.; Yuan, S.; Wang, S.-D.; Xi, D.-H.; Lin, H.-H. The higher expression levels of dehydroascorbate reductase and glutathione reductase in salicylic acid-deficient plants may contribute to their alleviated symptom infected with RNA viruses. Plant signaling & behavior 2011, 6. Jg., Nr. 9, 1402-1404. [CrossRef]
- Selway, J.W. Antiviral activity of flavones and flavans. Progress in clinical and biological research 1986, 213. Jg., 521-536.
- Hrmova, M.; Hussain, S. S. Plant Transcription Factors Involved in Drought and Associated Stresses. International Journal of molecular sciences 2021, 22. Jg., Nr. 11, 5662. [CrossRef]
- Pandey, A.; Khan, Mohd. K.; Hamurcu, M.; Brestic, M.; Topal, A.; Gezgin, S. Insight into the Root Transcriptome of a Boron-Tolerant Triticum zhukovskyi Genotype Grown under Boron Toxicity. Agronomy 2022, 12. Jg., Nr. 10, S. 2421. [CrossRef]
- Li Y, Liu K, Tong G, Xi C, Liu J, Zhao H, Wang Y, Ren D, Han S. MPK3/MPK6-mediated phosphorylation of ERF72 positively regulates resistance to Botrytis cinerea through directly and indirectly activating the transcription of camalexin biosynthesis enzymes. Journal of Experimental Botany 2022, 73. Jg., Nr. 1, 413-428. [CrossRef]
- Liu, Y.; Jin, W.; Wang, L.; Wang, X. Replication-associated proteins encoded by Wheat dwarf virus act as RNA silencing suppressors. Virus research 2014, 190. Jg., 34-39. [CrossRef]
- Chellappan P.; Vanitharani R.; Pita J.; Fauquet C.M. Short interfering RNA accumulation correlates with host recovery in DNA virus-infected hosts, and gene silencing targets specific viral sequences. Journal of virology 2004, 78. Jg., Nr. 14, 7465-7477. [CrossRef]
- Li, F.; Wang, Y. & Zhou, X. SGS3 Cooperates with RDR6 in Triggering Geminivirus-Induced Gene Silencing and in Suppressing Geminivirus Infection in Nicotiana Benthamiana. Viruses 2017, 9. Jg., Nr. 9, 247. [CrossRef]
- Ding, S.-W.; Li, H.; Lu, R.; Li, F.; Li, W.-X. RNA silencing: a conserved antiviral immunity of plants and animals. Virus research 2004, 102. Jg., Nr. 1, 109-115. [CrossRef]
- Shen, Q.; Bao, M.; Zhou, X. A plant kinase plays roles in defense response against geminivirus by phosphorylation of a viral pathogenesis protein. Plant signaling & behavior 2012, 7. Jg., Nr. 7, S. 888-892. [CrossRef]
- Zhang, D.; Bai, G.; Hunger, R. M.; Bockus, W. W.; Yu, J.; Carver, B. F.; Brown-Guedira, G. Association Study of Resistance to Soilborne wheat mosaic virus in U.S. Winter Wheat. Phytopathology 2011, 101. Jg., Nr. 11, 1322-1329. [CrossRef]
- Liu, S.; Yang, X.; Zhang, D.; Bai, G.; Chao, S.; Bockus, W. Genome-wide association analysis identified SNPs closely linked to a gene resistant to Soil-borne wheat mosaic virus. Theoretical and Applied Genetics 2014, 127. Jg., 1039-1047. [CrossRef]
- Hourcade, D.; Bogard, M.; Bonnefoy, M.; Savignard, F.; Mohamadi, F.; Lafarge, S.; Du Cheyron, P.; Mangel, N.; Cohan, J. P. Genome-wide association analysis of resistance to wheat spindle streak mosaic virus in bread wheat. Plant Pathology 2019, 68. Jg., Nr. 3, 609-616. [CrossRef]
- Pfrieme, A. K.; Ruckwied, B; Habekuß, A.; Will, T.; Stahl, A.; Pillen, K.; Ordon, F. Corrigendum: Identification and validation of Quantitative Trait Loci for Wheat dwarf virus resistance in wheat (Triticum spp.). Frontiers in Plant Science, 2022, 13. Jg., 1088938. [CrossRef]
- Buerstmayr, M.; Buerstmayr, H. Two major quantitative trait loci control wheat dwarf virus resistance in four related winter wheat populations. Theoretical and Applied Genetics 2023, 136. Jg., Nr. 5, 103. [CrossRef]
- Li, H.; Conner, R. L.; Liu, Z.; Li, Y.; Chen, Y.; Zhou, Y.; Duan, X.; Shen, T.; Chen, Q.; Graf, R. J.; Jia, X. Characterization of Wheat-Triticale Lines Resistant to Powdery Mildew, Stem Rust, Stripe Rust, Wheat Curl Mite, and Limitation on Spread of WSMV. Plant disease 2007, 91. Jg., Nr. 4, 368-374. [CrossRef]
- Parlevliet, J.E. Durability of resistance against fungal, bacterial and viral pathogens; present situation. Euphytica 2002, 124. Jg., 147-156. [CrossRef]
- Palloix, A.; Ayme, V.; Moury, B. Durability of plant major resistance genes to pathogens depends on the genetic background, experimental evidence and consequences for breeding strategies. New Phytologist 2009, 183. Jg., Nr. 1, 190-199. [CrossRef]
- Brown J.K.; Idris A.M.; Ostrow K.M.; Goldberg N.; French R.; Stenger D.C. Genetic and phenotypic variation of the Pepper golden mosaic virus Complex. Phytopathology 2005, 95. Jg., Nr. 10, 1217-1224. [CrossRef]
- Riedel, C. Pyramidisierung von QTL im Hinblick auf eine Verbesserung der Barley yellow dwarf virus Toleranz der Gerste und genetische Analyse der Toleranz gegenüber Wheat dwarf virus. Ph.D. dissertation, university of Rostock, 2013.
- Ayalew, H.; Tsang, P.W.; Chu, C.; Wang, J.; Liu, S.; Chen, C.; et al. Comparison of TaqMan, KASP and rhAmp SNP genotyping platforms in hexaploid wheat. PloS one 2019, 14. Jg., Nr. 5, e0217222. [CrossRef]
- Karlstedt, F. (2020). Identification and mapping of QTL for resistance against Zymoseptoria tritici in the winter wheat accession HTRI1410 (Triticum aestivum L. subsp. Spelta) Julius Kühn-Institut, Bundesforschungsinstitut für Kulturpflanzen, 2020.
Figure 1.
Classification and genomic organization of Wheat dwarf virus (WDV), (a) Classification of the family Geminiviridae is based on their molecular and biological characteristics. The WDV species belong to the Mastreviruses and consists of the main strains of wheat and barley, to which the various isolates are subordinated in clades. The percentage of nucleotide similarity is given for the species, strains, and clades. WDV Bar [TR] refers to the recombinant isolate between a barley isolate and a yet unknown member of the Mastreviruses. (b) Genomic organization of the Mastreviruses, which include Wheat dwarf virus (WDV). These have a circular ssDNA genome (black circle) and four ORFs. Code of viral proteins: MP- movement protein, CP- capsid protein, RepA- replication-associated protein, Rep- replication initiation protein. Also shown are the non-coding regions of the large intergenic region (LIR) and small intergenic region (SIR).
Figure 1.
Classification and genomic organization of Wheat dwarf virus (WDV), (a) Classification of the family Geminiviridae is based on their molecular and biological characteristics. The WDV species belong to the Mastreviruses and consists of the main strains of wheat and barley, to which the various isolates are subordinated in clades. The percentage of nucleotide similarity is given for the species, strains, and clades. WDV Bar [TR] refers to the recombinant isolate between a barley isolate and a yet unknown member of the Mastreviruses. (b) Genomic organization of the Mastreviruses, which include Wheat dwarf virus (WDV). These have a circular ssDNA genome (black circle) and four ORFs. Code of viral proteins: MP- movement protein, CP- capsid protein, RepA- replication-associated protein, Rep- replication initiation protein. Also shown are the non-coding regions of the large intergenic region (LIR) and small intergenic region (SIR).
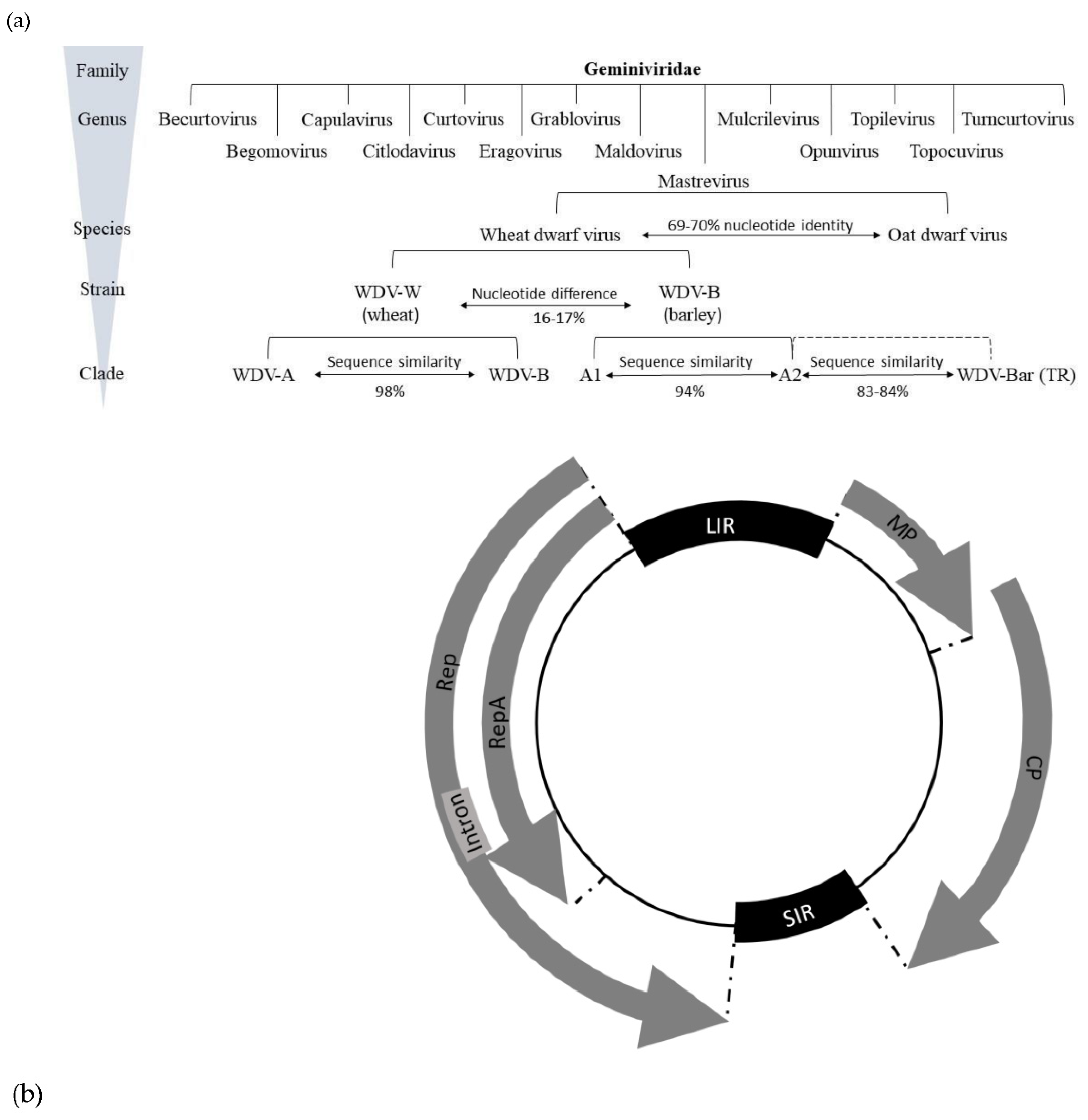
Figure 2.
World map with countries where WDV could be detected (marked in red). WDV was reported in Ukraine [128], Romania [129], Bulgaria [130], Hungary [131], Italy [115], France [44], Sweden [119], Poland [132], Finland [18], Spain [133], the United Kingdom [106], Austria [106] and Slovenia [134], as well as regions in Iran [135], the Middle East (Turkey [107], Africa (Tunisia [117] and Zambia [136]), West Asia (Syria [137], and China [138,139]). [140].
Figure 2.
World map with countries where WDV could be detected (marked in red). WDV was reported in Ukraine [128], Romania [129], Bulgaria [130], Hungary [131], Italy [115], France [44], Sweden [119], Poland [132], Finland [18], Spain [133], the United Kingdom [106], Austria [106] and Slovenia [134], as well as regions in Iran [135], the Middle East (Turkey [107], Africa (Tunisia [117] and Zambia [136]), West Asia (Syria [137], and China [138,139]). [140].
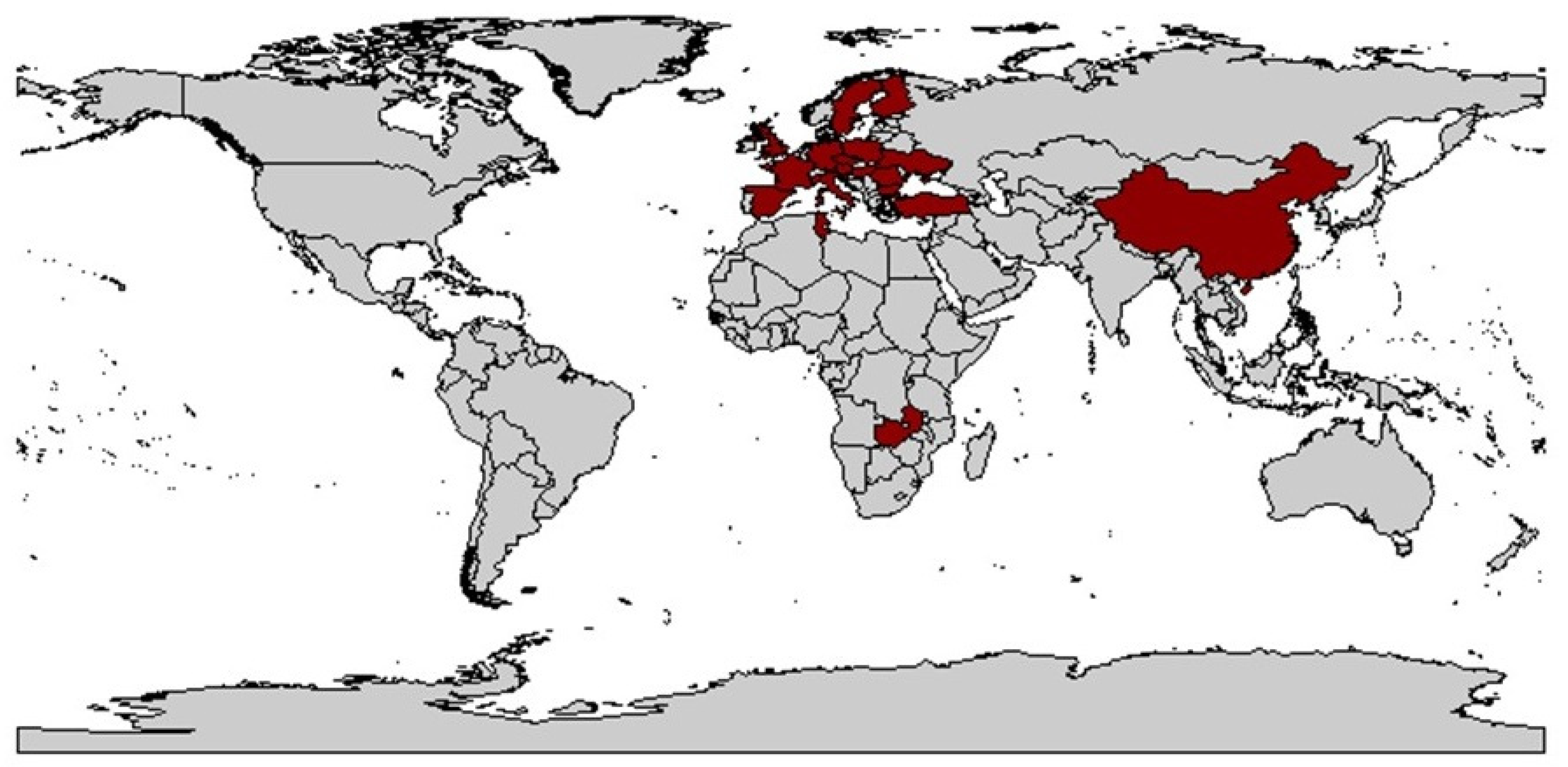
Figure 3.
Eight-week-old wheat plants with different degrees (symptom scoring 1,2,5,6,8) of dwarfing in the greenhouse depending on their genotype (a) and at BBCH stage 30-39 in Mai 2021 under field conditions (b) after artificial inoculation. (c) Leaves of WDV-infected plants (left) show a stripe-like lightening compared to healthy leaves (right), which later develops into yellowing.
Figure 3.
Eight-week-old wheat plants with different degrees (symptom scoring 1,2,5,6,8) of dwarfing in the greenhouse depending on their genotype (a) and at BBCH stage 30-39 in Mai 2021 under field conditions (b) after artificial inoculation. (c) Leaves of WDV-infected plants (left) show a stripe-like lightening compared to healthy leaves (right), which later develops into yellowing.

Figure 4.
Schematic representation of the life cycle of winter cereals and Psammotettix alienus. The major developmental stages of host cereal plants (from sowing to harvest) are represented by the outer circle. The successive and overlapping biological cycles of P. alienus are represented by arrows in the inner circle. Under optimal conditions (20 °C, 70-95% relative humidity, 18/6 light/dark hours), the length of the life cycle (from egg to adult death) is 71 days [197]. Eggs produced in the fall overwinter on cereals and hatch in the following growing season (the next spring). According to Manurung et al. [12], the duration of the five larval stages (L1 to L5) is 5.9, 5.1, 5.6, 3, and 9.4 days, respectively. The 7-day-old adults can mate to produce the next generation of insects [12].
Figure 4.
Schematic representation of the life cycle of winter cereals and Psammotettix alienus. The major developmental stages of host cereal plants (from sowing to harvest) are represented by the outer circle. The successive and overlapping biological cycles of P. alienus are represented by arrows in the inner circle. Under optimal conditions (20 °C, 70-95% relative humidity, 18/6 light/dark hours), the length of the life cycle (from egg to adult death) is 71 days [197]. Eggs produced in the fall overwinter on cereals and hatch in the following growing season (the next spring). According to Manurung et al. [12], the duration of the five larval stages (L1 to L5) is 5.9, 5.1, 5.6, 3, and 9.4 days, respectively. The 7-day-old adults can mate to produce the next generation of insects [12].
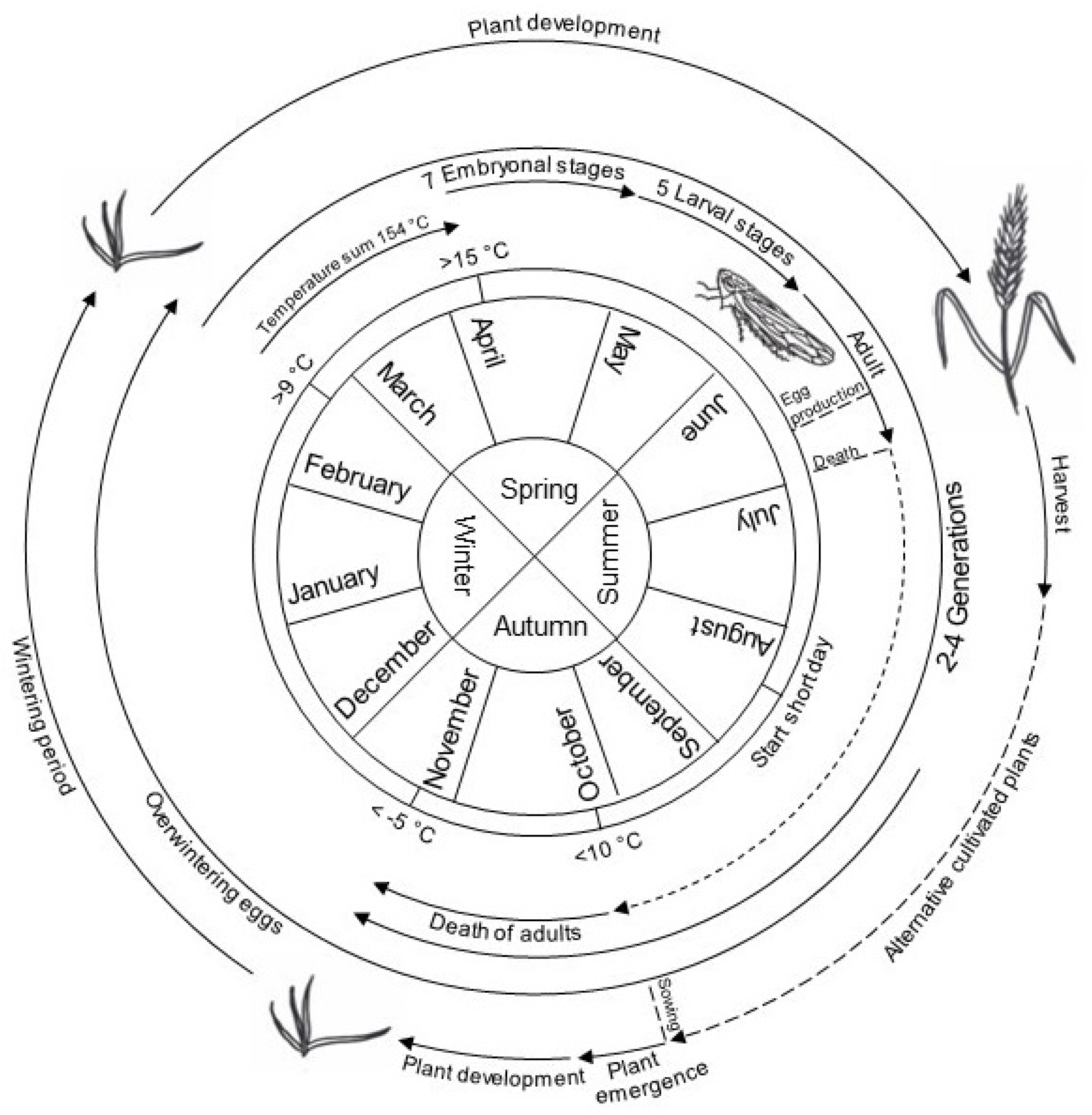
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



